Embed Size (px)
Citation preview

Synaptic Transmission
Jianhong Luo, Ph.D.
Department of Neurobiology
Zhejiang University School of Medicine
Main Reference: Neuroscience Exploring the Brain, 3rd Ed.
By M.F. Bear, B.W. Connors, and M.A. Paradiso

Introduction
Types of synapses
Electrical synapses
Chemical synapses
Principles of chemical synaptic transmission
- Neurotransmitters (NT)
- Synthesis and storage
- Release
- Receptors and effectors
- Recovery and degradation
- Neuropharmacology
Principles of synaptic integration
The integration of EPSPs
The contribution of dendritic properties
Inhibition
Modulation

Introduction
By the end of 19th century, it was recognized that this transfer of information
from one neuron to another occurs at specialized sites of contact.
(例:疼痛的反应)

Synapse
(1897 Charles Sherrington)
Synaptic transmission – two hypothesis
Argued for a century on its physical nature.
Electrical synapse
(Proven in 1959 by E. Furshpan and D. Potter in crayfish)
Chemical synapse
1. Solid evidence given in 1921 by Otto Loewi;
2. B. Katz et al. demonstrated fast transmission at NMJ
chemically mediated.
3. By 1951, John Eccles studied the synaptic transmission
in the mammalian CNS using the glass microelectrode
4. During the last decade, new methods of studying the
molecules revealed that synapses are far more complex.
A large and fascinating topic
Introduction

Introduction
Otto Loewi (1873-1961), a German born
pharmacologist, discoverer of acetylcholine,
nobel prize laureate in Physiology or Medicine
in 1936.
His most famous experiment came from his dream in the night
of Easter Sunday, 1921 and found ―vagusstoff‖, turned out to be
acetylcholine, showing that synaptic signaling used chemical
messengers.

Types of synapses
A synapse is the specialized junction where one part of a neuron
contacts and communicates with another neuron or cell type
(such as a muscle or glandular cell). Information tends to flow in
one direction, from a neuron to its target cell. The first is said to
be presynaptic and the target is said to be postsynaptic.
Electrical synapses (电突触)
Six connexins form a channel (connexon), and two connexons
(one from each cell) form a gap junction channel.
The pore of channels is relatively large with diameter 1–2 nm,
enough for all the major cellular ions, and many small organic
molecules, to pass through directly from the cytoplasm of one cell
to the other’s.
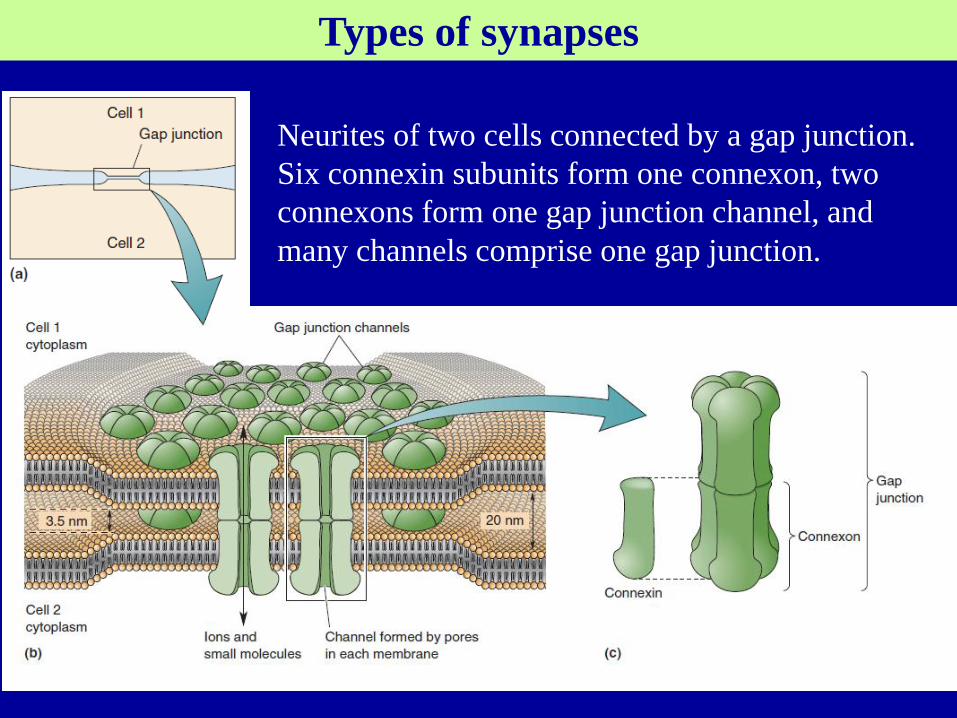
Types of synapses
Neurites of two cells connected by a gap junction.
Six connexin subunits form one connexon, two
connexons form one gap junction channel, and
many channels comprise one gap junction.

Types of synapses
Functional properties of electrical synapses:
Equally pass in both direction
Electrically coupled
Very fast, and if the synapse is large, fail-safe. Thus, an AP in
the presynaptic neuron can produce, almost instantaneously,
an AP in the postsynaptic neuron.
In invertebrate species, such as the crayfish, electrical synapses
are sometimes found between sensory and motor neurons in
neural pathways mediating escape reflexes.

Types of synapses
Electrical synapses also occur in the vertebrate brain.
Are common in every part of the mammalian CNS
Among electrically coupled neurons, AP in the presynaptic
neuron can cause a small amount of ionic current to flow
across the gap junction channels into the other neuron,
producing postsynaptic potential (PSP).
The PSP generated by a single electrical synapse in the
mammalian brain is usually small—about 1 mV or less at its
peak—and may not, by itself, be large enough to trigger an
AP in the postsynaptic cell.
The precise roles of electrical synapses vary from one brain
region to another (synchronize; developmental coordination;
in non-neuron cells).
Box 5-2 by Michael V. L. Bennett
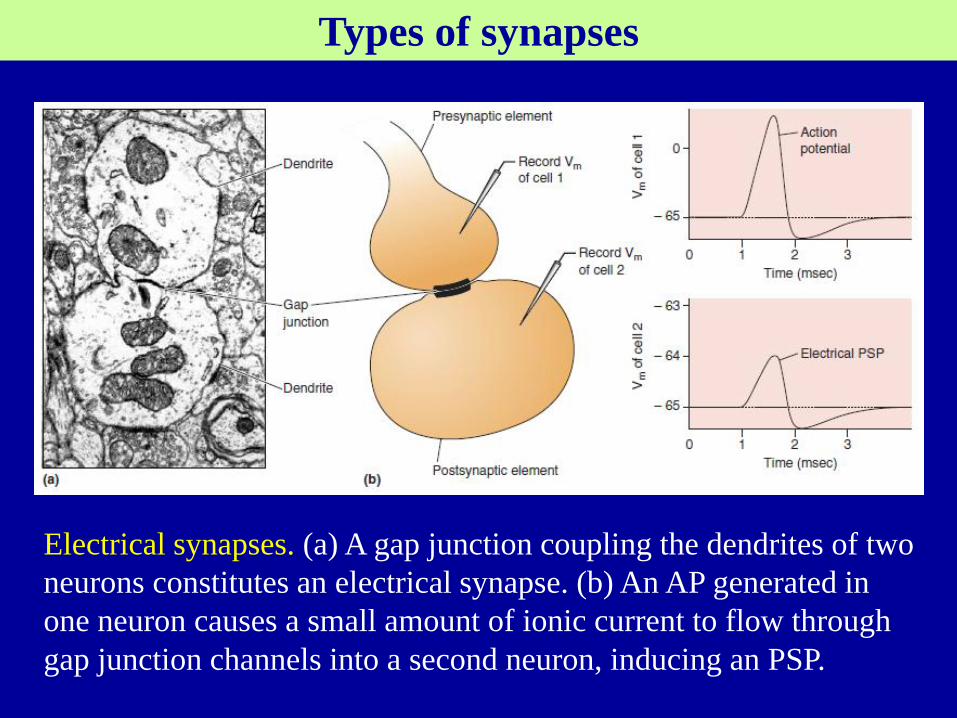
Types of synapses
Electrical synapses. (a) A gap junction coupling the dendrites of two
neurons constitutes an electrical synapse. (b) An AP generated in
one neuron causes a small amount of ionic current to flow through
gap junction channels into a second neuron, inducing an PSP.

Types of synapses
Chemical synapses
General description:
synaptic cleft (20–50 nm ), filled with a matrix of fibrous
extracellular protein. One function of this matrix is to make the pre-
and postsynaptic membranes adhere to each other.
presynaptic element, is usually an axon terminal.
synaptic vesicles (50 nm in diameter), store neurotransmitter
used to communicate with the postsynaptic neuron.
secretory granules (larger vesicles, about 100 nm diameter)
contain soluble protein (dark in EM, large dense-core vesicles)
Membrane differentiations on either side of the synaptic cleft
Active zone looks like pyramid, the sites of NT release
postsynaptic density contains receptors converting signal from
intercellular to intracellular

The components of a chemical synapse.
Types of synapses
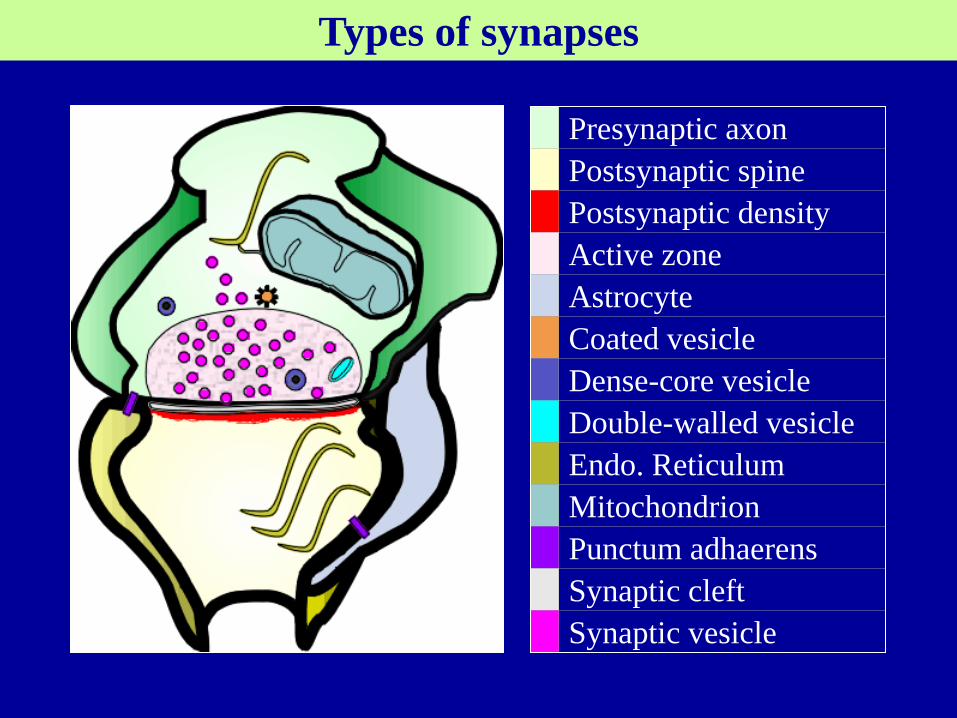
Presynaptic axon
Postsynaptic spine
Postsynaptic density
Active zone
Astrocyte
Coated vesicle
Dense-core vesicle
Double-walled vesicle
Endo. Reticulum
Mitochondrion
Punctum adhaerens
Synaptic cleft
Synaptic vesicle
Types of synapses

Different types in of synapse in the CNS (CNS synapses)
The sizes and shapes of CNS synapses also vary widely
Axodendritic, Axosomatic, axoaxonic, dendrodendritic synapses.
Types of synapses
Chemical synapses as seen with EM
(left) A fast excitatorysynapse in the CNS
(right) A synapse in the PNS, with numerous dense-core vesicles
DCV
AZ
Mt
Presynaptic
Postsynaptic
V
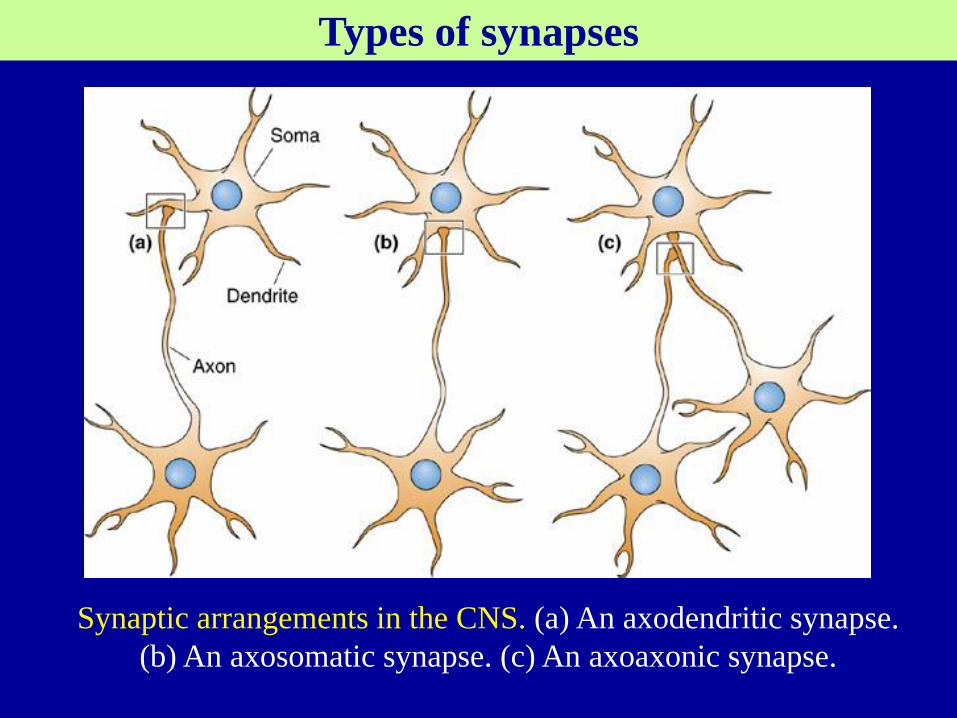
Synaptic arrangements in the CNS. (a) An axodendritic synapse.
(b) An axosomatic synapse. (c) An axoaxonic synapse.
Types of synapses

Various sizes of CNS synapses.
Notice that larger synapses
have more active zones.
Types of synapses
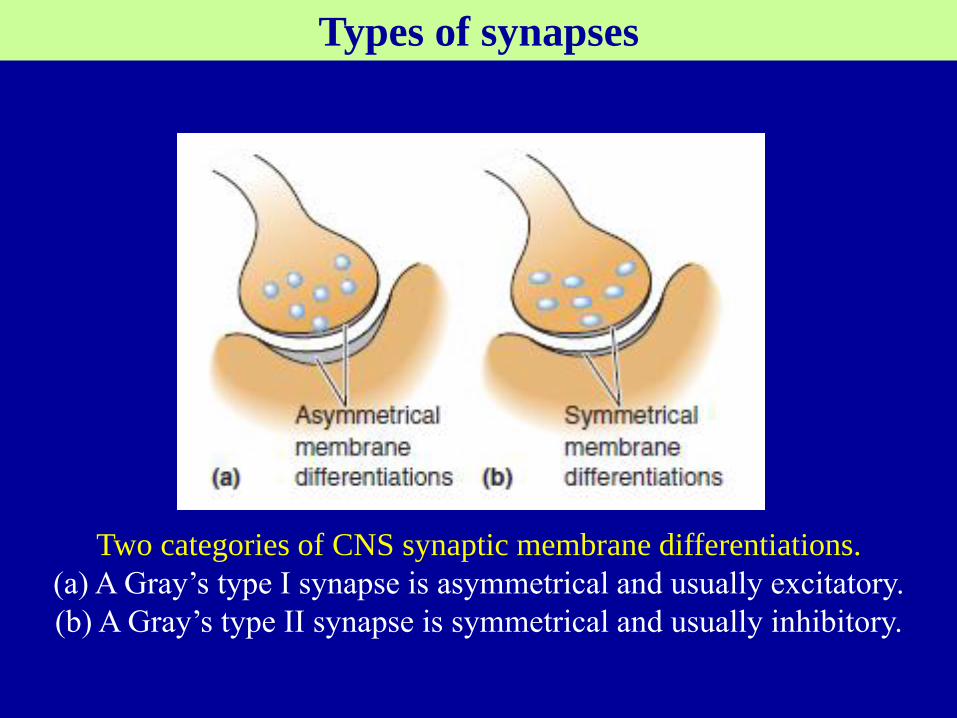
Two categories of CNS synaptic membrane differentiations.
(a) A Gray’s type I synapse is asymmetrical and usually excitatory.
(b) A Gray’s type II synapse is symmetrical and usually inhibitory.
Types of synapses

Synaptic junctions also exist outside the central nervous system.
Axons of the autonomic nervous system innervate glands,
smooth muscle, and the heart.
Neuromuscular junctions occur between the axons of motor
neurons of the spinal cord and skeletal muscle. NMJ has many
of the structural features of chemical synapses in the CNS.
Neuromuscular synaptic transmission is fast and reliable. An
AP in the motor axon always causes an AP in the muscle cell it
innervates (What structural features for this reliability?)
Most knowledge from the research on NMJ transmission.
Types of synapses

The neuromuscular junction. The postsynaptic membrane,
known as the motor endplate, contains junctional folds with
numerous neurotransmitter receptors.
Types of synapses

Principles of chemical synaptic transmission
There basic requirements for chemical synaptic transmission:
Synthesis and package into vesicles of neurotransmitter (NT);
Release of vesicle NT to cleft in response to a presynaptic AP;
Induction of an electrical or biochemical response to NT in the
postsynaptic neuron
Clearance of NT from the synaptic cleft
And, occur very rapidly to be useful for sensation, perception, and
the control of movement.

Principles of chemical synaptic transmission
Neurotransmitters (three chemical categories)
g-氨基丁酸 乙酰胆碱 胆囊收缩素
谷氨酸 多巴胺 强腓肽
甘氨酸 肾上腺素 脑啡肽
组胺 N-乙酰门冬氨酰谷氨酸
去甲肾上腺素 神经肽Y
5-羟色胺 生长抑素
P物质
促甲状腺素释放激素
血管活性肠肽

Neurotransmitters
Three chemical categories
Amine, amino acid, peptide
Secretory granules and synaptic vesicles
Often co-exist in the same axon terminals
amine + peptide
amino acid + peptide
Different neurons release different neurotransmitters
Fast transmission; NMJ; Slow transmission
Principles of chemical synaptic transmission

Representative neurotransmitters
(a) glutamate, GABA, and glycine.
(b) acetylcholine and norepinephrine.
(c) substance P.
Principles of chemical synaptic transmission

Principles of chemical synaptic transmission
Neurotransmitter Synthesis and Storage
Amine and amino acid neurotransmitters:
➀ Enzymes are transported to the axon terminal and convert
precursor molecules into neurotransmitter molecules in the cytosol.
➁ Transporter proteins load the neurotransmitter into synaptic
vesicles in the terminal, where they are stored.
Glu, Gly vs GABA, the amines
Peptides:
➀ A precursor peptide (a long peptide) synthesis in the rough
ER in cell body.
➁ Then split in the Golgi apparatus to yield the active one.
➂ Secretory vesicles with the peptide bud off from the Golgi
apparatus.
➃ The secretory granules are transported (axoplasmic) down
the axon to the terminal where the peptide is stored.

Principles of chemical synaptic transmission
Transporters, proteins in the vesicle membrane, take up and
concentrate the amino acid and amaine neurotransmitters inside
the vesicle.

Principles of chemical synaptic transmission
Neurotransmitter Release
An action potential in the axon terminal → depolarization of
the terminal membrane → voltage-gated calcium channels in
the active zones to open ([Ca2+]i 0.0002 mM → ˃0.1 mM)→ vesicles release(exocytosis)→ the contents to spill out
into the synaptic cleft
The exocytosis occurs very rapidly within 0.2 msec of the
Ca2+ influx into the terminal. Why?
The mechanism by which [Ca2+] i stimulates exocytosis:
Reserve pool of vesicles bound to the cytoskeleton
Docking of vesicles to active zone
SNARE protein complex, conformation altered by ↑[Ca2+]i
Endocytosis
Recycled vesicle refilled with neurotransmitter

Principles of chemical synaptic transmission
The Release of Neurotransmitter by Exocytosis

Principles of chemical synaptic transmission
(a) This is a view of the extracellular
surface of the active zone of a
neuromuscular junction in a frog.
The particles are believed to be
calcium channels.
(b) In this view, the presynaptic terminal
had been stimulated to release
neurotransmitter. The exocytotic
fusion pores are where synaptic
vesicles have fused with the
presynaptic membrane and released
their contents.
A “receptor’s eye” view of neurotransmitter release

Principles of chemical synaptic transmission
SNAREs and vesicle fusion
(Box 5.3)
SNARE: SNAP Receptor
SNAP: Soluble NSF Attach Protein
NSF: N-ethylmaleimide-sensitive factor
(N-乙基马来酰亚胺敏感的融合因子)

Principles of chemical synaptic transmission
Secretory granules also release peptide neurotransmitters by
exocytosis:
in a calcium-dependent fashion
typically not at the active zones
requires high-frequency trains of AP and more calcium influx.
a leisurely process to taking 50 msec or more.

Principles of chemical synaptic transmission
Neurotransmitter Receptors and Effectors
binding to specific receptor proteins in the postsynaptic
density.
key in a lock, induce conformational changes in the
receptor and lead to different functions.
More than 100 different receptors can be classified into
two types: transmitter-gated ion channels and G-protein-
coupled receptors.

Receptors Ion channels
Receptor
channels
or Ionotropic receptors,
or Ligand-gated ion channels
G-protein Coupled Receptors
Enzyme linked receptors
Nuclear receptors
Voltage-gated
Mechanically-gated
Non-gated
Principles of chemical synaptic transmission
Transmitter-gated ion channels

Principles of chemical synaptic transmission
Transmitter-Gated Ion Channels
Membrane-spanning proteins consisting of four or five
subunits to form a pore.
Closed to open, neurotransmitter, binds to specific sites,
induces a conformational change
The functional consequence depends on which ions.
The structure of an ACh-gated ion channel
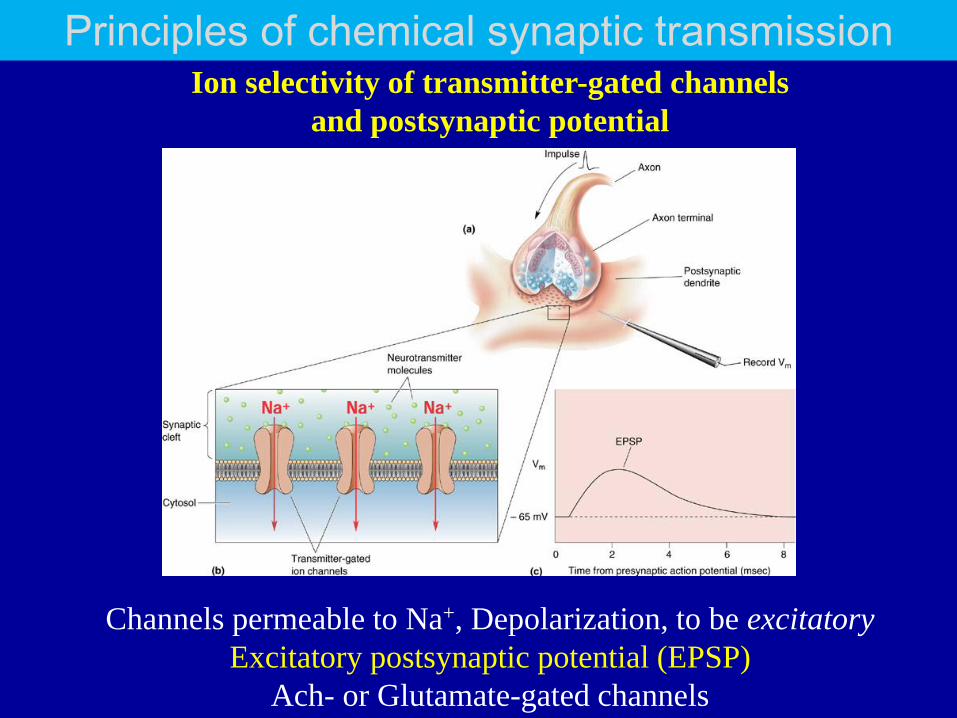
Principles of chemical synaptic transmission Ion selectivity of transmitter-gated channels
and postsynaptic potential
Channels permeable to Na+, Depolarization, to be excitatory
Excitatory postsynaptic potential (EPSP)
Ach- or Glutamate-gated channels
Principles of chemical synaptic transmission
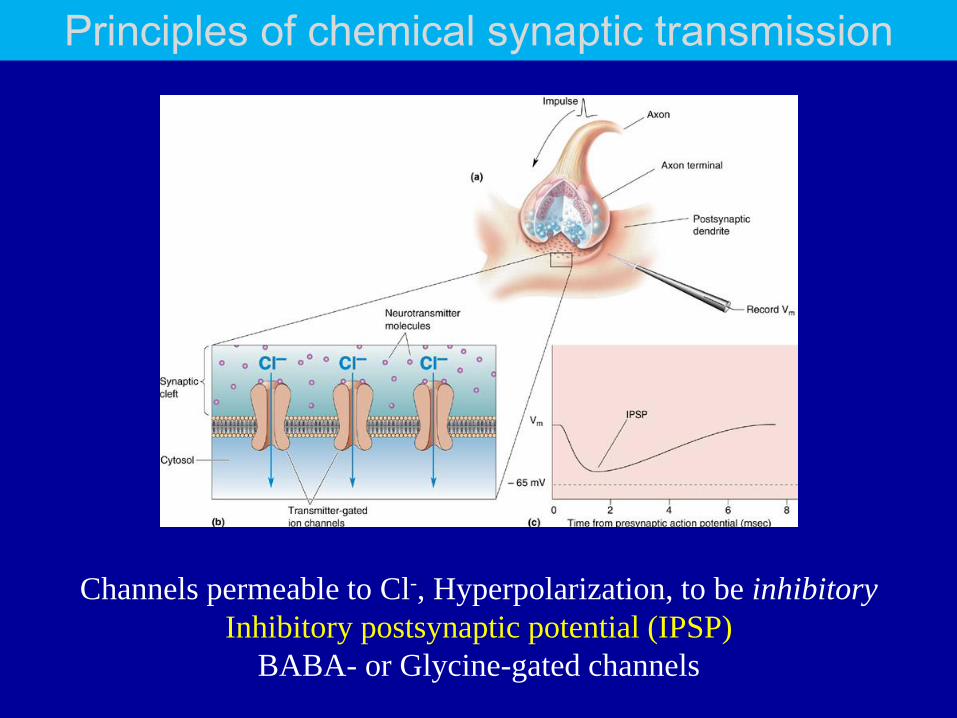
Channels permeable to Cl-, Hyperpolarization, to be inhibitory
Inhibitory postsynaptic potential (IPSP)
BABA- or Glycine-gated channels

Principles of chemical synaptic transmission
G-Protein-Coupled Receptors (GPCR)
Fast chemical synaptic transmission is mediated by amino acid
and amine neurotransmitters acting on transmitter-gated ion
channels. However, all three types of neurotransmitter, acting on
GPCR, can also have slower, longer-lasting, and much more
diverse postsynaptic actions.
This type of transmitter action involves three steps:
① Transmitters bind to receptors in the postsynaptic membrane.
② The receptors activate G-proteins, free to move along the
intracellular face of the postsynaptic membrane.
③ The activated G-proteins activate “effector” proteins.
Principles of chemical synaptic transmission
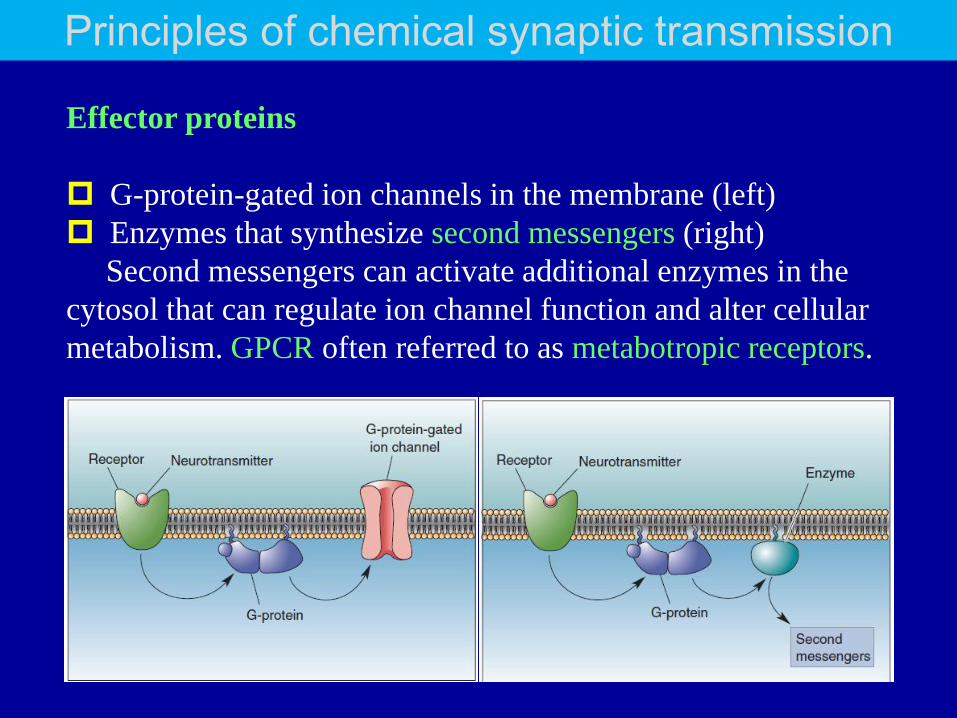
Effector proteins
G-protein-gated ion channels in the membrane (left)
Enzymes that synthesize second messengers (right)
Second messengers can activate additional enzymes in the
cytosol that can regulate ion channel function and alter cellular
metabolism. GPCR often referred to as metabotropic receptors.

Principles of chemical synaptic transmission
In the heart, a metabotropic ACh receptor is coupled by a
G-protein to a potassium channel. It slows the rhythmic
contractions of the heart by causing a slow
hyperpolarization of the cardiac muscle cells.
In skeletal muscle, the receptor is an ACh-gated ion channel,
permeable to Na+. ACh induces contraction by causing a
rapid depolarization of the muscle fibers.
The same neurotransmitter can have different postsynaptic
actions, depending on what receptors it binds to.
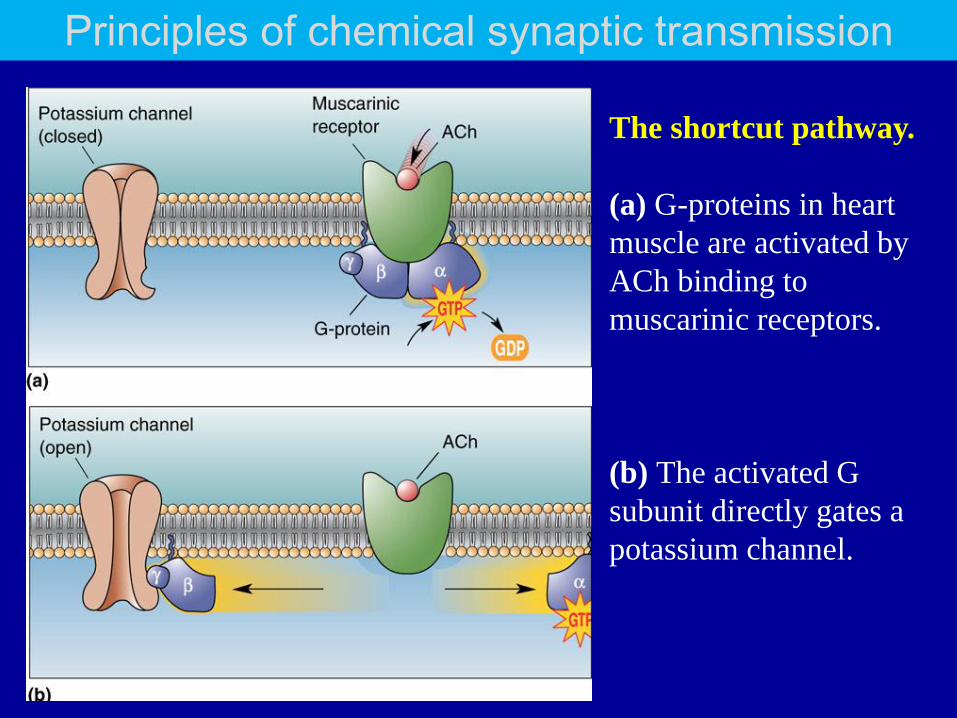
The shortcut pathway.
(a) G-proteins in heart
muscle are activated by
ACh binding to
muscarinic receptors.
(b) The activated G
subunit directly gates a
potassium channel.
Principles of chemical synaptic transmission

Principles of chemical synaptic transmission
Neurotransmitter receptors are also commonly found in the
membrane of the presynaptic axon terminal.
Sensitive to the neurotransmitter, called autoreceptors.
Typically, autoreceptors are GPCR
The common consequences of activating autoreceptors is
inhibition of neurotransmitter release. This allows a
presynaptic terminal to regulate itself
Autoreceptors

Neurotransmitter Recovery and Degradation
Principles of chemical synaptic transmission
Neurotransmitter in the synaptic cleft must be cleared to allow
another round of synaptic transmission.
Simple diffusion (For most of the amino acid and amine
neurotransmitters)
Reuptake occurs by the action of specific transporter proteins
located in the presynaptic membrane (once inside the cytosol,
enzymatically destroyed, or reloaded into synaptic vesicles)
Neurotransmitter transporters also exist in the membranes of
glia surrounding the synapse, which assist in such removal.
Enzymatic destruction in the cleft. Ach is removed at the NMJ
by enzyme acetylcholinesterase, deposited in the cleft.
Importance of removal: desensitization (脱敏); nerve gases

Neuropharmacology
Each of the steps of synaptic transmission is chemical, and
therefore can be affected by specific drugs and toxins.
Inhibitors: e.g. Nerve gases inhibite the enzyme AChE. Inhibitors
of neurotransmitter receptors, called receptor antagonists (e.g.
Curare, an arrow-tip poison, binds tightly to the ACh receptors)
Receptor agonists. e.g. nicotine, binds to, and activates, the ACh
receptors in skeletal muscle and CNS. nicotinic ACh receptors
(nAChR).
Wrong neurotransmission is the root cause of many
neurological and psychiatric disorders. Knowledge of
neuropharmacology of synaptic transmission will be helpful for
development of new and effective therapeutic drugs.
Principles of chemical synaptic transmission

Principles of synaptic integration
The integration of EPSPs
The contribution of dendritic properties
Inhibition
Modulation
Principles Of Synaptic Integration

Principles Of Synaptic Integration
The postsynaptic neuron integrates thousands of synaptic inputs
(complex ionic and chemical signals) and gives rise to a simple
form of output: AP
The transformation constitutes a neural computation. The brain
performs billions of neural computations every second.
Synaptic integration is the process by which multiple synaptic
potentials combine within one postsynaptic neuron.
Principles Of Synaptic Integration
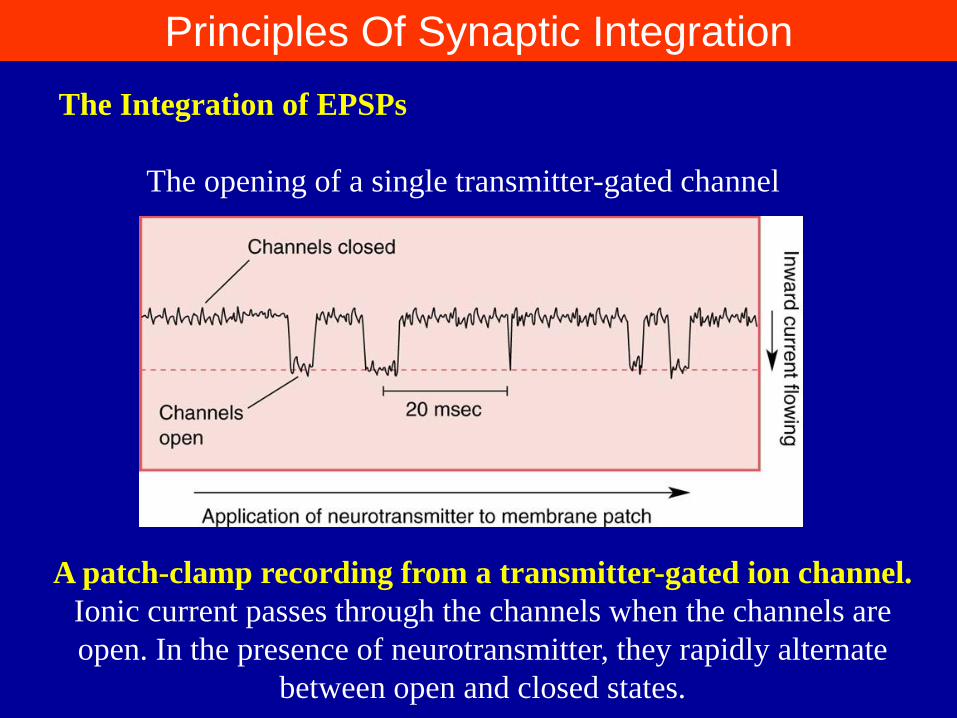
The Integration of EPSPs
The opening of a single transmitter-gated channel
A patch-clamp recording from a transmitter-gated ion channel.
Ionic current passes through the channels when the channels are
open. In the presence of neurotransmitter, they rapidly alternate
between open and closed states.

Patch Clamps Permit Measurement of
Ion Movements through Single Channel
(not only in a whole cell)
Patch Clamp (膜片钳)
Different configurations
Principles Of Synaptic Integration

Principles Of Synaptic Integration
Quantal Analysis (量子分析) of EPSPs: a method of
comparing the amplitudes of miniature and evoked
postsynaptic potentials.
The neurotransmitter content in a single synaptic vesicle.
Spontaneous release w/o AP, one vesicle → miniature EPSP
(miniEPSP, mEPSP)
Multiple vesicle release w AP (evoked) → EPSP (multiples of
mEPSP)
i.e. postsynaptic EPSPs at a given synapse are quantized; they
are multiples of an indivisible unit, the quantum, that reflects
the number of transmitter molecules in a single synaptic
vesicle and the number of postsynaptic receptors available at
the synapse.

Principles Of Synaptic Integration
There is a big difference between excitatory transmission at NMJ
and CNS synapses.
Most neurons in CNS perform more sophisticated computations,
requiring that many EPSPs add together to produce a significant
postsynaptic depolarization. This is what is meant by integration
of EPSPs.
EPSP summation is the simplest form of synaptic integration.
Spatial summation is the adding together of EPSPs
generated simultaneously at many different synapses on a dendrite.
Temporal summation is the adding together of EPSPs
generated at the same synapse if they occur in rapid succession,
within about 1–15 msec of one another.
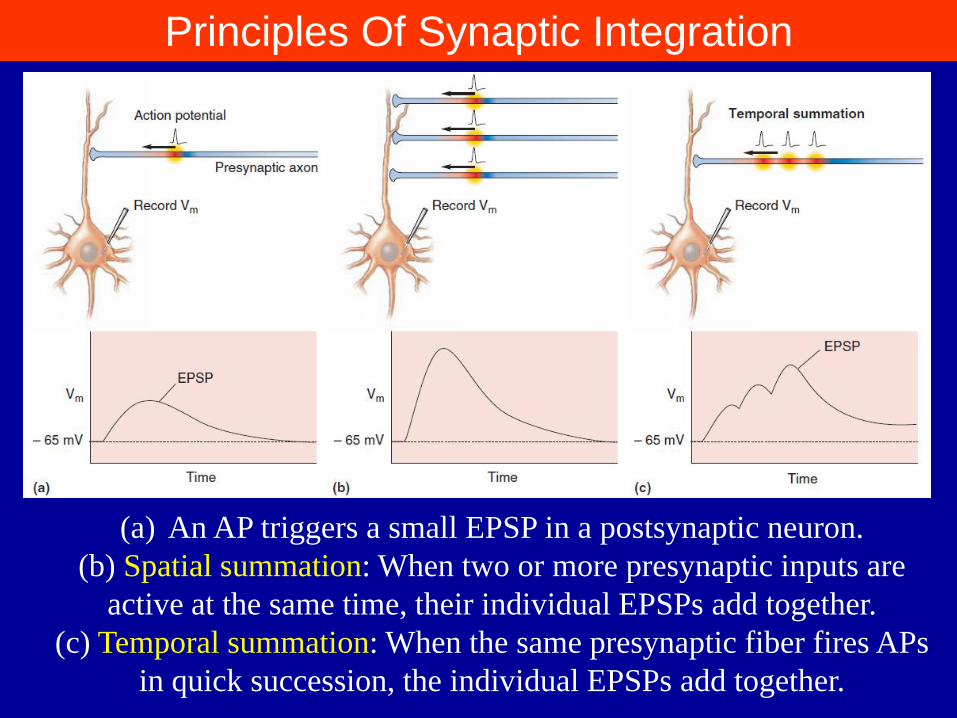
Principles Of Synaptic Integration
(a) An AP triggers a small EPSP in a postsynaptic neuron.
(b) Spatial summation: When two or more presynaptic inputs are
active at the same time, their individual EPSPs add together.
(c) Temporal summation: When the same presynaptic fiber fires APs
in quick succession, the individual EPSPs add together.

Principles Of Synaptic Integration
The Contribution of Dendritic Properties to Synaptic
Integration
The current of synaptic contact must spread down the dendrite and
the soma, and cause the membrane of the spike-initiation zone to be
depolarized beyond threshold, before an AP can be generated.
The effectiveness of an excitatory synapse in triggering an AP,
therefore, depends on how far the synapse is from the spike-
initiation zone and on the properties of the dendritic membrane.

Principles Of Synaptic Integration
Dendritic Cable Properties:
To simplify, let’s assume that dendrites function as cylindrical
cables that are electrically passive; that is, lacking voltage-gated ion
channels (in contrast, of course, with axons).
Imagine that the current at a synapse is like turning on the water
that will flow down a leaky garden hose (the dendrite).
Similarly, two paths that synaptic current can take: One is down the
inside of the dendrite; the other is across the dendritic membrane.
At some distance from the site of current
influx, the EPSP amplitude may approach
zero because of the dissipation of the current
across the membrane.

Principles Of Synaptic Integration
To simplify the mathematics, we assume the dendrite is infinitely
long, unbranched, and uniform in diameter.
The amount of depolarization falls off exponentially with increasing
distance:
Vx=V0/e x/λ
when x=λ, then Vx=V0/e. Put another way, Vλ=0.37 (V0).
This distance λ, where the depolarization is 37% of that at the
origin, is called the dendritic length constant.
(Remember that this analysis is an oversimplification) .
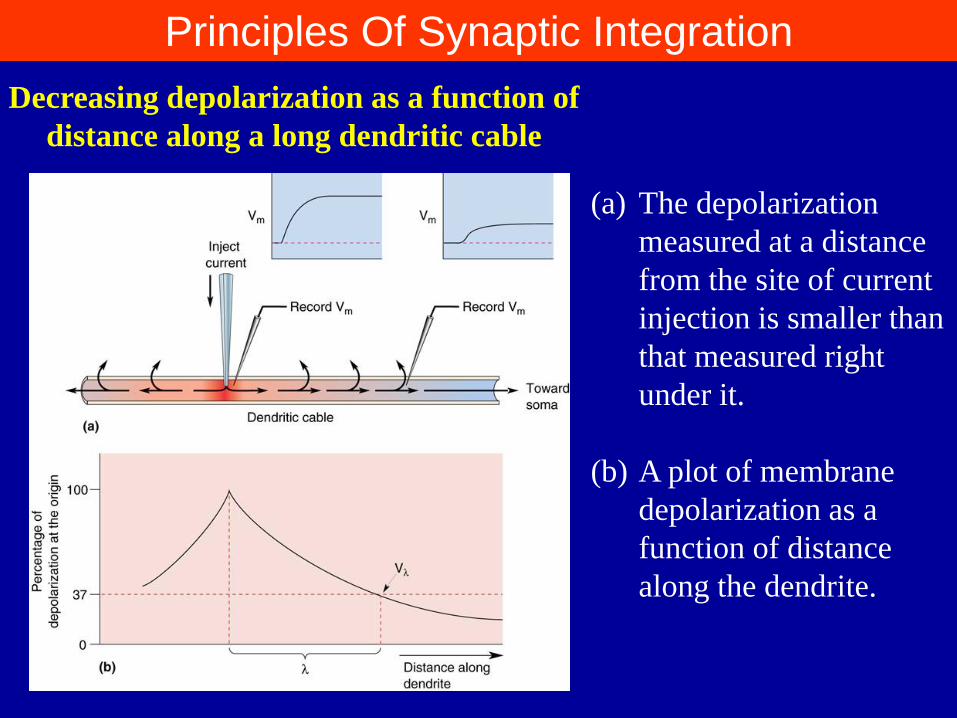
Principles Of Synaptic Integration
(a) The depolarization
measured at a distance
from the site of current
injection is smaller than
that measured right
under it.
(b) A plot of membrane
depolarization as a
function of distance
along the dendrite.
Decreasing depolarization as a function of
distance along a long dendritic cable

Principles Of Synaptic Integration
The length constant is an index of how far depolarization can
spread down a dendrite or axon. The longer the length constant, the
more likely it is that EPSPs generated at distant synapses will
depolarize the membrane at the axon hillock(轴丘).
λ depends on two factors:
(1) the internal resistance (ri); and (2) the membrane resistance (rm).
ri depends only on the diameter of the dendrite and the electrical
properties of the cytoplasm (relatively constant in a mature neuron)
rm, depends on the number of open ion channels, which changes
from moment to moment depending on what other synapses are
active.
The dendritic length constant, therefore, is not constant at all!

Principles Of Synaptic Integration
Excitable Dendrites.
Assumption: The dendrite’s membrane is electrically passive.
The dendrites of spinal motor neurons are very close to passive.
However, many other neuronal dendrites are decidedly not
passive.
The voltage-gated channels in dendrites can act as important
amplifiers of small EPSPs generated far out on dendrites.
Paradoxically, in some cells dendritic sodium channels may also
serve to carry electrical signals in the other direction—from the
soma outward along dendrites.

Principles Of Synaptic Integration
A cortical pyramidal neuron with a long apical dendrite
that has voltage-gated ion channels

Principles Of Synaptic Integration
Inhibition
EPSP → AP output depends on:
the number of coactive excitatory synapses
the distance the synapse is from the spike-initiation zone
the properties of the dendritic membrane
Plus:
inhibitory synapses that take the membrane potential away from
action potential threshold, and exert a powerful control over a
neuron’s output .

Principles Of Synaptic Integration
IPSPs and Shunting Inhibition (分流抑制) The postsynaptic inhibitory receptors are GABA or glycine-
gated ion channels that they only allow Cl- to pass through their
channels.
Opening of the chloride channel brings the membrane potential
toward the chloride equilibrium potential, ECl-, about - 65 mV.
So, whether its activation causes a hyperpolarizing IPSP or not
depend on the resting membrane potential.
If there is no visible IPSP, is the neuron really inhibited? The
answer is yes.
Shunting inhibition (分流抑制). The actual physical basis of
shunting inhibition is the inward movement of negatively
charged chloride ions, which is formally equivalent to outward
positive current flow.
Thus, inhibitory synapses also contribute to synaptic integration
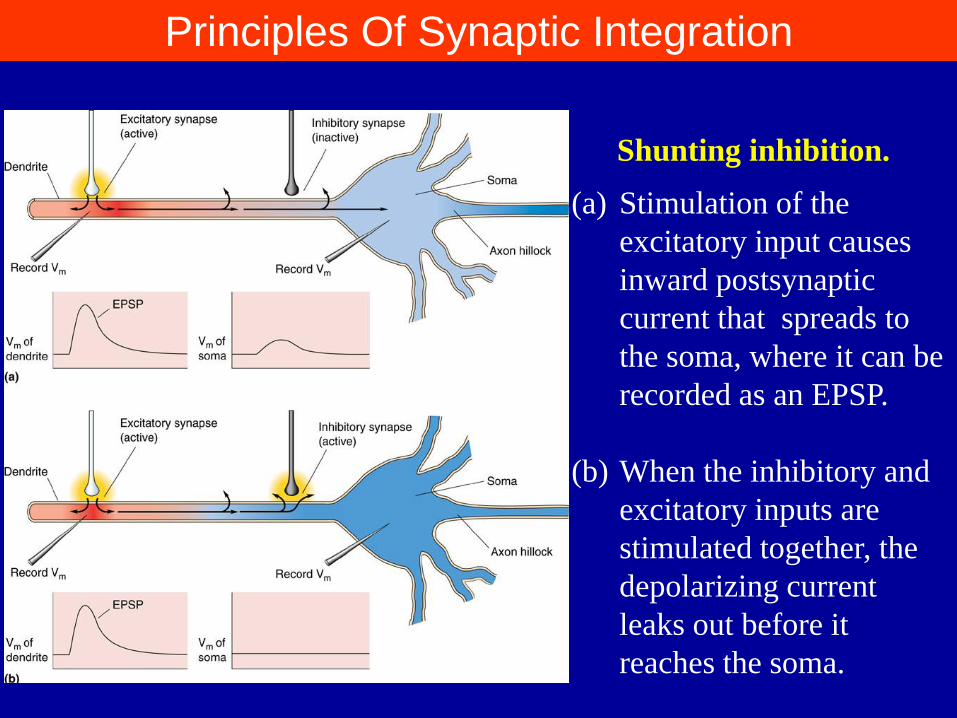
Principles Of Synaptic Integration
(a) Stimulation of the
excitatory input causes
inward postsynaptic
current that spreads to
the soma, where it can be
recorded as an EPSP.
(b) When the inhibitory and
excitatory inputs are
stimulated together, the
depolarizing current
leaks out before it
reaches the soma.
Shunting inhibition.

Principles Of Synaptic Integration
The Geometry of Excitatory and Inhibitory Synapses
Inhibitory synapses (GABA or glycine), Gray’s type II.
Excitatory synapses (glutamate), Gray’s type I
Inhibitory synapses on many neurons are found clustered on the
soma and near the axon hillock.

Principles Of Synaptic Integration
Modulation (调制)
In addition to synaptic transmitter-gated channels, there are many
synapses with G-protein-coupled neurotransmitter receptors that do
not directly evoke EPSPs and IPSPs, but instead modifies the
effectiveness of EPSPs generated by other synapses. This is called
modulation.
e.g. norepinephrine β receptor. The binding of norepinephrine (NE)
to the receptor triggers a cascade of biochemical events within the
cell to produce the second messager cAMP

Principles Of Synaptic Integration
Modulation by the NE receptor. ➀ The binding of NE to the receptor
activates a G-protein in the membrane. ➁ The G-protein activates the enzyme
adenylyl cyclase. ➂ Adenylyl cyclase converts ATP into the second messenger
cAMP. ➃ cAMP activates a protein kinase. ➄ The protein kinase causes a
potassium channel to close by attaching a phosphate group to it.

Principles Of Synaptic Integration
decreasing the K+ conductance increases the dendritic membrane
resistance and therefore increases the length constant λ.
Distant or weak excitatory synapses will become more effective in
depolarizing the spike-initiation zone beyond threshold. i.e. the
cell becomes more excitable.
It is why excitability of a neuron is increased when NE is released
presynaptically.
END

谢谢!