Embed Size (px)
Citation preview

Faculteit Wetenschappen
Departement Biologie
Study of intrapopulation variation in movement and habitat use in a stream fish (Cottus perifretum):
integrating behavioural, ecological and genetic data
Studie van individuele verschillen in verplaatsingsgedrag en habitatkeuze van een riviervis (Cottus perifretum):
integratie van gedrag, ecologische en genetische data
Dissertation for the degree of Doctor in Science: Biology
at the University of Antwerp to be defended by
ALEXANDER KOBLER
Promotor: Prof. Dr. Marcel Eens
Antwerpen, 2012

Doctoral Jury
Promotor
Prof. Dr. Marcel Eens
Chairman
Prof. Dr. Erik Matthysen
Jury members
Prof. Dr. Lieven Bervoets
Prof. Dr. Gudrun de Boeck
Prof. Dr. Filip Volckaert
Dr. Gregory Maes
Dr. Michael Ovidio
ISBN: 9789057283864
© Alexander Kobler, 2012.
Any unauthorized reprint or use of this material is prohibited. No part of this book may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system without express written permission from the author.

A naturalist’s life would be a happy one if he had only to observe and never to write.
Charles Darwin

4
Acknowledgments
My Ph.D. thesis was made possible through a FWO (Fonds Wetenschappelijk Onderzoek -‐ Vlaanderen) project-‐collaboration between the University of Antwerp and the Catholic University of Leuven.
First of all, I wish to thank Marcel Eens, the head of the Biology-‐Ethology research group in Antwerp, who supervised me during all phases of my thesis. Marcel, I am very grateful for your trust and patience. You gave me confidence and incentive during this difficult journey. Hartelijk bedankt!
In Leuven, I was guided by Filip Volckaert, the head of the Biodiversity and Evolutionary Genomics research group, and Gregory Maes. I truly appreciate all your time, patience, knowledge, spontaneous inputs and happy welcomes that you gave me. I learned a lot; also things that go beyond this thesis and are not included in it. Cheers!
I also wish to thank my other jury and committee members. Thanks Lieven Bervoets, Gudrun de Boeck, Erik Matthysen and Michael Ovidio for your contribution. I really appreciate it.
In the first two months of my work for this thesis, I had also a co-‐supervisor, Guy Knaepkens, who had initiated the FWO project together with Greg. Although our collaboration was only of short duration, I am happy to have met you. Thanks for believing in my abilities to achieve this high goal. Jammer dat we samen niet meer donderpadden konden vangen.
In Leuven, I met Alexandros Triantafyllidis, who was in Filip’s lab for a half year sabbatical. You invested a large part of your time into the “bullhead research”, I am grateful for all your interest and your help to guide me into the world of population genetics. I also met Bart Hellemans in Leuven, a very nice person and true connoisseur of genetic techniques and theory. I learned a lot and really enjoyed working together with you in the laboratory.
I got much support from the Ethology research group and I want to name the people that had their part in this thesis, motivated me, taught me Dutch and/or gave me friendship: Berber De Jong (Smakelijk!), Geert Eens, Igor Eulaers, Laszlo Garamszegi (Egészségedre!), Ann Geens (was good to have a nice fairy in the office!), Hector Rivera Gutierrez, Veerle Jaspers, Stefanie Lahaye (keep me updated about the villa), Carsten Lucass, Josie Meaney-‐Ward, Wendt Müller, Rianne Pinxten, Maud Poisbleau, Peter Scheys (Petri Heil!), Jeff Van Camp, Evi Van

5
den Steen (was wel een leuke tijd), Alain Van Hout, Jonas Vergauwen. Thanks a lot, I have lovely memories of all of you.
During my thesis I also shared great moments with the students that I supervised. Big cheers goes out to Dimitri Geelhand de Merxem, Katia Geudens, Yves Humblet and Sofie Vanwetswinkel.
I was always warm-‐hearted welcomed in the Leuven lab. I still see the smiles of Alessia Cariani, Conny Coeckelberghs, Auguste Chocha Manda, Eveline Diopere, Sarah Geldof, Pascal Habluetzel, Tine Huyse, Nellie Konijnendijk, Maarten Larmuseau, Joost Raeymaekers, Jo-‐Ann De Roos, Dirk Schaerlaekens, Sara Vandamme, Frederik Van den Broeck, Maarten Vanhove and Jeroen Van Houdt.
I contemplated about science and biology and lived together with Stuart Baird, Joelle Gouy de Bellocq and Loran Crespin. You were very important to me in my scientific but also personal development. Thanks for sharing life and living room. It was great!
Sabine Convent, thanks for having been such as nice landlady, you provided me best circumstances to start a new stage of my life here in Flanders.
I also received great support of many other friends and I want to name a few of them: Cordula Altendorf, Volker Huckstorf, Cecilia Iribarren, Karoline Kühnelt, Vito Martorana, Stefan, Susi and Simon Scharpf (the S-‐family), Solveig Schröder, the triathlon companions Jeroen Bartels, Bob Dejongh, Dieter Delbaere, Fritz Gerhart, Dave Lietaert, Xenia Luxem, Yeray Luxem, Joris Peeters, Bart Van de Velde and Andreas Vicic, and the fishing buddies Mathias Birkle, Lawrence De Geyseleer, Jan Hallerman, Daniel Hammer, Mattias Hempel, Gilles Lambert, Wulf Plickat, Bastian Reetz, Jan-‐Simon Saamen, Hendrik Schuster, Giovanni Vanhooren and Jens Verschaeren. Good to have friends like you!
Many thanks also to Anne’s family, Martine Gijsbrechts and Frederik Moonen, which always supported me and gave me a home in Turnhout. Many thanks!
My parents Christine and Georg Kobler, my sister Katja and her love Mark Dongus, my brother Matthias and my grandparents Hilde and Reinhold Rall encouraged and supported me during school, study and Ph.D. It was a long educational journey and I am endlessly grateful that you made this possible. I love you.
My girlfriend Anne shared with me the sweet as well as the bitter days during the writing of this thesis. Anne, je was een heel belangrijke (en zo mooie) steun tijdens moeilijke perioden. Zonder jou zal dit nooit zo leuk geweest zijn. Daarvoor ben ik jou oneindig dankbaar. Dikke kus, ik hou van jou.

6
Table of Contents
Summary .............................................................................................. 8
Samenvatting ..................................................................................... 10
General Introduction .......................................................................... 13 Intrapopulation heterogeneity in behaviour ............................................. 15 Individual differences in movement behaviour ......................................... 16 Intra-‐population heterogeneity in habitat use .......................................... 18 Temperament traits ................................................................................. 19 Study species ............................................................................................ 20 Passive integrated transponder telemetry ................................................ 21 Objectives and thesis outline .................................................................... 22
Comparison of laboratory and field behaviour .................................... 27 Abstract .................................................................................................... 29 Introduction ............................................................................................. 30 Materials and methods ............................................................................ 31 Results ..................................................................................................... 33 Discussion ................................................................................................ 35
Temperament traits and habitat use .................................................. 39 Abstract .................................................................................................... 41 Introduction ............................................................................................. 42 Materials and methods ............................................................................ 44 Results ..................................................................................................... 54 Discussion ................................................................................................ 59
Diel movement of bullhead ................................................................ 63 Abstract .................................................................................................... 65 Introduction ............................................................................................. 66 Materials and Methods ............................................................................ 68 Results ..................................................................................................... 71 Discussion ................................................................................................ 75
Period-‐dependent sex-‐biased movement ........................................... 79 Abstract .................................................................................................... 81 Introduction ............................................................................................. 82 Materials and methods ............................................................................ 84 Results ..................................................................................................... 88 Discussion ................................................................................................ 92

7
Movement range and temperament traits .......................................... 95 Abstract .................................................................................................... 97 Introduction ............................................................................................. 98 Materials and methods .......................................................................... 100 Results ................................................................................................... 110 Discussion .............................................................................................. 113
Movement range, fitness and heterozygosity ................................... 117 Abstract .................................................................................................. 119 Introduction ........................................................................................... 120 Materials and methods .......................................................................... 122 Results ................................................................................................... 127 Discussion .............................................................................................. 131
General discussion, conclusions and future research ........................ 135 Late preface: personal reflection ............................................................ 137 Laboratory activity of bullhead ............................................................... 138 Aggressiveness and the use of structured habitats ................................. 139 Movement range and dispersal distance ................................................ 141 Movement range and individual fitness .................................................. 142 Between-‐year consistency in individual behaviour ................................. 143 Candidate gene -‐ phenotype association ................................................ 144
References ....................................................................................... 147

Summary
8
SUMMARY There is growing interest in consistent individual differences in behaviour. This has led to an increasing number of studies that distinguish behavioural groups within single populations. For example, individual differences in movement distances, habitat selection or temperament traits have prompted scientists to differentiate populations into e.g. resident and mobile, specialist and generalist, or shy and bold individuals. Which individual characteristics are associated with such behavioural grouping is not completely understood. There are indications that environmental stimuli during early life-‐stage, life-‐history traits or resource limitations may be related. Furthermore, there may be a genetic basis for the expression of certain behavioural traits, which may also be heritable.
Whereas there are numerous studies examining consistent individual differences in movement patterns, resource specialisation (e.g. microhabitat use) or temperament traits, there are only few studies combining these research areas. This is one of the aims of this thesis. A non-‐migratory stream-‐fish with known intra-‐population heterogeneity in movement patterns (resident and mobile individuals) was used to study movement behaviour, microhabitat use and temperament traits. Attention was paid to the consistency of individual differences. Information on individual life-‐history traits such as sex and somatic growth rate, body size, body condition and genetic variation was used to explain individual differences in behaviours.
Individually tagged bullhead (Cottus perifretum) from a lowland stream population were studied in the field over ten months, a subsample of these individuals were recaptured and the expression of temperament traits (activity, boldness, aggressiveness, exploration of novel food and novel environment) was observed in subsequent laboratory observations. Furthermore, data collected before this thesis was used to compare the exploratory behaviour in the natural environment with this behaviour under laboratory conditions. The main findings of the study can be summarized as follows.
The exploratory behaviour in the laboratory predicted this behaviour under field conditions. There were indications that passive bullhead showing only little activity during the tests were more predictable than individuals with higher activity.
Two temperament trait tests, activity and novel environment activity, were repeated in the laboratory and consistent individual behaviour was observed.

Summary
9
There was no association with sex. Smaller individuals were more active in the novel environment.
Individual habitat use was consistent over the three-‐months observation period. Bullhead positively selected structured habitats and avoided open water. However, we also observed some individuals that were regularly tracked in stream patches with only little complexity. The individual habitat use was unrelated to sex or body size but an association with aggressiveness was found: bullhead that were more often associated with highly complex habitats were less aggressive. Other temperament traits were not associated with habitat use.
Movement of bullhead varied between diel and seasonal periods. Bullhead moved farther distances at night and dawn than during daytime. Some individuals expressed site-‐fidelity: after swimming several metres at night they returned to the same daytime location. Bullhead moved furthest in February and May. In these months, movement distances diversified between the genders and period-‐dependent sex-‐bias was observed: females moved furthest in February and males moved furthest in May.
Long-‐term movement range (the distance between the most up-‐ and downstream location during ten months) varied considerably between individuals and ranged from 1 to 1284 stream metres. A clear grouping of resident and mobile fish was, however, not found. Instead, movement distribution seemed to be leptokurtic. The inter-‐individual differences were not related to sex, body size, body condition and growth rate. The individual expression of temperament traits was also not associated with movement range. Furthermore, individual genetic diversity could not explain differences in movement range.
Bullhead with better body condition at the beginning of the study grew faster during the field observation period. No association between body condition and microsatellite heterozygosity as well as between growth rate and microsatellite heterozygosity was found.
The main conclusion of this thesis is that the studied population was heterogeneous in movement behaviour, habitat use and temperament traits. These behaviours varied consistently between individuals. Individual habitat use was related to aggressiveness. No individual characteristic was found that was associated with movement range. The thesis highlights the importance of structured habitat types for bullhead. The study suggests that inter-‐sexual differences in reproductive behaviour may cause (periodical) sex-‐bias in the movement of many fishes.

Samenvatting
10
SAMENVATTING Er bestaat een groeiende interesse voor consistente individuele gedragsverschillen. Dit heeft geleid tot een stijgend aantal studies die gedragsgroepen binnen een populatie wil onderscheiden. Individuele verschillen in verplaatsingsgedrag, habitatkeuze of gedragskenmerken hebben ertoe geleid dat wetenschappers populaties in bijvoorbeeld sedentaire en mobiele, specialisten en generalisten, of onverschrokken en verschrokken individuen onderscheiden. Welke individuele karakteristieken met de gedragsgroepering samenhangen, is niet volledig duidelijk.
Hoewel er talrijke studies zijn die consistente individuele verschillen in verplaatsingspatronen, habitatkeuze of gedragskenmerken onderzoeken, zijn er slechts weinig studies die deze verschillende aspecten combineren. Dit is één van de doelstellingen van deze thesis. Een niet-‐migrerende riviervis met gekende intra-‐populatie heterogeniteit in verplaatsingspatronen (sedentaire en mobiele individuen) werd gebruikt om verplaatsingsgedrag, habitatkeuze en gedragskenmerken te bestuderen. Hierbij werd bijzondere aandacht besteed aan de consistentie van individuele gedragsverschillen.
Individueel gemarkeerde rivierdonderpadden (Cottus perifretum) van een laagland beekpopulatie werden gedurende tien maanden in het veld bestudeerd. Een deel van deze populatie werd opnieuw gevangen en de expressie van gedragskenmerken (activiteit, onverschrokkenheid, agressiviteit en exploratie naar nieuw voedsel en een nieuwe omgeving) werd onderzocht in opeenvolgende laboratoriumobservaties. Bovendien werden eerder verzamelde data gebruikt om het exploratiegedrag in de natuurlijke omgeving te vergelijken met het gedrag onder laboratoriumomstandigheden. De belangrijkste bevindingen van deze studie kunnen als volgt worden samengevat.
Het exploratiegedrag in het laboratorium voorspelde het gedrag onder veld-‐ condities. Er waren indicaties dat passieve individuen, die slechts weinig activiteit vertoonden, voorspelbaarder waren dan individuen met een hogere activiteit.
Twee testen van gedragskenmerken, activiteit en activiteit in een nieuwe omgeving, werden herhaald tijdens de labostudie. Daaruit bleek dat de individuele gedragsverschillen consistent waren. Het geslacht verklaarde deze verschillen niet. Kleinere individuen waren actiever in de nieuwe omgeving.
De individuele habitatkeuze was consistent over de drie maanden durende observatieperiode. Rivierdonderpadden verkozen gestructureerde habitatten en

Samenvatting
11
vermeden open water. We observeerden echter ook enkele individuen die regelmatig voorkwamen in beekgebieden met maar weinig structuur. De individuele habitatkeuze was niet gerelateerd met geslacht of lichaamsgrootte. Wel werd er een relatie met agressiviteit gevonden: rivierdonderpadden die vaker met structuurrijke habitatten geassocieerd werden, waren minder agressief. Andere gedragskenmerken correleerden niet met de habitatkeuze.
Verplaatsingsafstanden waren afhankelijk van de tijden van de dag en de jaargetijden. Rivierdonderpadden verplaatsten zich ´s nachts en tijdens de ochtendschemering over een grotere afstand dan overdag. Sommige individuen vertoonden een sterke plaatstrouw: na enkele meters te hebben afgelegd in de nacht, keerden ze terug naar dezelfde locatie van overdag. Rivierdonderpadden verplaatsten zich het verst in februari en mei. In deze maanden werden uiteenlopende verplaatsingsafstanden tussen mannetjes en vrouwtjes waargenomen. Vrouwtjes zwommen het verst in februari, mannetjes in mei.
De verplaatsingsspanwijdte (de afstand tussen de meest stroomopwaartse en meest stroomafwaarste locatie gedurende tien maanden) varieerde onmiskenbaar tussen de individuen met waardes van 1 tot 1284 meters. Een duidelijke groepering van sedentaire en mobiele individuen werd echter niet gevonden; de verplaatsingsdistributie bleek leptokurtisch. Inter-‐individuele verschillen waren niet gerelateerd met geslacht, lichaamsgrootte, conditie of groeisnelheid. De individuele expressie van gedragskenmerken was ook niet gerelateerd met de verplaatsingsspanwijdte. Bovendien kon de individuele genetische diversiteit de verschillen in verplaatsingsspanwijdte niet verklaren.
Rivierdonderpadden met een betere lichaamsconditie aan het begin van de studie groeiden sneller tijdens de observatieperiode. Lichaamsconditie en microsatelliet heterozygositeit waren niet gecorreleerd, net zoals groeisnelheid en microsatelliet heterozygositeit.
De hoofdconclusie van deze thesis is dat de bestudeerde populatie heterogeen was wat betreft verplaatsingsgedrag, habitatkeuze en gedragskenmerken. De onderzochte gedragingen varieerden consistent tussen de individuen. Individuele habitatkeuze was gecorreleerd met agressiviteit. Er werden geen individuele karakteristieken gevonden die geassocieerd waren met de verplaatsingsspanwijdte. Deze thesis benadrukt het belang van gestructureerde habitattypen voor de rivierdonderpad. De studie suggereert dat interseksuele verschillen in voortplantingsgedrag (periodieke) seks-‐bias in de verplaatsing van vele vissoorten zouden kunnen veroorzaken.


CHAPTER 1
GENERAL INTRODUCTION


Introduction
15
Intrapopulation heterogeneity in behaviour
The variation in individual behaviour has often been ignored by ecologists. This is reflected in the common assumption that characterizing a species by the population’s average phenotype is sufficient to understand its functional role within an ecosystem (Bolnick et al. 2003; Sih et al. 2004; Réale et al. 2007). This view has been challenged through the identification of ecologically relevant variation in individual behavioural specialization across a variety of taxa ranging from gastropods to mammals (Robinson et al. 1996; Wilson 1998; Bolnick et al. 2003; Araujo et al. 2011). Inter-‐individual variation unrelated to sex, size or age has been observed e.g. in diet composition, foraging behaviour, oviposition preference, microhabitat use (reviewed by Bolnick et al. 2003), temperament traits (also referred to as personality traits, see further introduction; reviewed by Sih et al. 2004; Réale et al. 2007) or movement patterns (McLaughlin et al. 1992; Rodriguez 2000; Austin et al. 2004).
There is growing interest in individual differences in behaviour (Bell et al. 2009). Recent research suggests that an individual’s behaviour, once considered to be plastic, may be more predictable than previously assumed (Bell et al. 2009). For example, juvenile male great tits (Aves: Parus major) show consistent individual differences in how they respond to novel situations (exploratory behaviour, Verbeek et al. 1994). These individual differences can be consistent from juvenile phase to adulthood (Carere et al. 2005), correlate with other behavioural traits (forming behavioural types: Sih et al. 2004; Bell 2007) and may even be heritable (Dingemanse et al. 2002).
Recent theory suggests that the behavioural heterogeneity may be explained by individual differences in fitness strategies. For instance, trade-‐offs between current and future reproductive success may cause consistent individual differences in risk-‐taking behaviour; individuals with high future expectations should be more risk-‐averse (Wolf et al. 2007). Moreover, consistent individual differences in growth rates may encourage consistent individual differences in behavioural types that increase both growth and mortality (Stamps 2007). For example, more risk-‐prone behaviour while foraging (boldness) and higher aggressiveness during territory defence may result in higher food intake but also increases the risk of predation (Stamps 2007). In animals with indeterminate growth such as fish, amphibians or reptiles individuals with e.g. bolder behaviour may, therefore, benefit from higher growth rate but suffer from greater mortality risk.

Chapter 1
16
Whereas there are numerous studies examining consistent individual differences in resource specialization (e.g. diet composition or microhabitat use; reviewed by Bolnick et al. 2003), temperament traits (also referred to as personality traits; reviewed by Sih et al. 2004; Réale et al. 2007) or movement patterns (McLaughlin et al. 1992; Rodriguez 2000; Austin et al. 2004), there are surprisingly few studies combining these research areas. Studying relationships between microhabitat use, temperament traits and movement patterns is, therefore, part of this thesis and should help to increase knowledge about the existence of consistent behavioural types.
Individual differences in movement behaviour
Movement is a fundamental feature of animals (Nathan et al. 2008). Movement allows the exploitation of spatially and temporally variable resources and is essential for the persistence of a species (Bohonak 1999; Bowler & Benton 2005; Clobert et al. 2009). The importance of movement behaviour for the ecology and evolution of wild populations becomes increasingly apparent as populations face fragmentation of their habitat (Clobert et al. 2009). Furthermore, the linking of movement behaviour with a quantitative description of dispersal (the movement from a natal/breeding site to another breeding site, Clobert et al. 2009) has become increasingly important in dispersal ecology (Hawkes 2009). Successful management of a species, therefore, relies on an understanding of the patterns and dynamics of movement.
The variation of individual movement patterns within a population is a common phenomenon in animal ecology and has been found in many taxa ranging from crustaceans (e.g. Hays et al. 2001) to fishes (e.g. McLaughlin et al. 1992; Kobler et al. 2009a), reptiles (e.g. McLaughlin 1989), birds (e.g. McLaughlin 1989) and mammals (e.g. Austin et al. 2004). Intrapopulation variation in movement behaviour reflects the different strategies in e.g. exploration, foraging or mate search used by individuals or sexes within a population to meet the demands of survival (Austin et al. 2004). The examination of individual variation in movement patterns thus helps to increase the understanding of ecological and evolutionary processes within a population of a species.
An extreme example of individual differences in movement behaviour is partial migration. Populations that partially migrate consist of resident and migrant individuals. This phenomenon is widespread in animal kingdom and documented in migratory invertebrates, fish, birds and mammals (Chapman et al.

Introduction
17
2011). It is particularly expressed in migratory birds (reviewed by Lundberg 1988) and anadromous fish (reviewed by Jonsson & Jonsson 1993). Anadromous fish species such as salmonids (family: Salmonidae) migrate from freshwater to saltwater. Only a part of the juvenile population will transform to migrating salmonids. In comparison to residents, migrant salmonids benefit from faster growth but suffer from greater mortality risk during migration (reviewed by Jonsson & Jonsson 1993). Male salmonids tend to be less migratory than females (Jonsson & Jonsson 1993). This can be explained by the greater benefit of faster growth and larger body size for migrating females (Jonsson & Jonsson 1993) as female egg production increases with body size (Fleming & Gross 1991).
Less attention has been paid to elucidating the patterns of movement in non-‐migratory stream fish especially in small benthic species (Lucas & Baras 2001). This may be due to lower economic or recreational value (Lucas & Baras 2001). However, small benthic stream fish are important biological components of fish assemblages and stream ecosystems and often also target species for nature conservation and habitat recovery plans (e.g. species included in the European Habitat Directive) (Matthews 1998; Lucas & Baras 2001). Non-‐migratory stream fish exhibit different life-‐histories than for example salmonids (e.g. Atlantic salmon, Salmo salar; brown trout, Salmo trutta). Consequently, it is difficult to infer general patterns of movement behaviour between stream fishes.
Although movement distances of non-‐migratory stream fish are often very restricted (Gerking 1959), movement distribution within populations can also be heterogeneous (e.g. Skalski & Gilliam 2000; Fraser et al. 2001). Numerous studies highlighted the existence of a mobile fraction that moves over longer distances (e.g. Gerking 1959; Gowan et al. 1994; Rodriguez 2002). Owing to the inter-‐individual differences in movement behaviour, numerous studies differentiated non-‐migratory stream fish populations into resident and mobile individuals (e.g. Funk 1957; Gowan et al. 1994; Smithson and Johnston 1999; Rodriguez 2002; Knaepkens et al. 2004; Belica & Rahel 2008). The mobile fraction influences the level of inbreeding through the spread of new genes (Howard 1960) and the repopulation of depauperated areas (Gerking 1959). However, not much is known which individual characteristics distinguish resident and mobile fish.
There are indications that life-‐history traits such as body size (Gowan & Fausch 1996) or age (Hughes 2000; Petty & Grossman 2004) are associated to individual differences in movement distances of non-‐anadromous stream fish. Other studies could not find such relation although intra-‐population heterogeneity in movement was high (e.g. Knaepkens et al. 2005; Breen et al.

Chapter 1
18
2009; Hudy & Shiflet 2009; Ovidio et al. 2009). In the absence of a life-‐history trait association, a behavioural association to movement patterns or a behavioural grouping of resident and mobile stream fish may exist and remains to be determined (Howard, 1960; Rodriguez, 2002). Fitness traits such as body condition or growth rate may also be associated to movement differences in non-‐migratory stream fish, similar to what has been observed in anadromous salmonids. Furthermore, individual genetic diversity may be linked to distances moved. For instance, microsatellite heterozygosity in mountain goats (Oreamnos americanus) has been associated to a lower propensity to disperse (Shafer et al. 2011). Studies that relate genetic diversity to individual movement distances in fish are lacking and may help to further distinguish resident and mobile individuals.
Intra-‐population heterogeneity in habitat use
Habitat use is a universal activity among animals that affects nearly all of an individual’s subsequent choices (Orians & Wittenberger 1991). The study of habitat use, therefore, requires attention to individual differences in the use of the environment and how between-‐individual differences in behavioural traits influence success in habitats and distributions of individuals among habitats (Orians 1991). Similar to niche partitioning between species under competition (Schoener 1974) and the density-‐dependent ecological divergence between species (Rosenzweig 1991; Morris 2003), a few examples exist that habitat use specialization may also exist within a population of a species (Bolnick 2001; Swanson et al. 2003; Svanbäck & Bolnick 2007). Beyond critical density thresholds, single individuals of the population may expand to unused or underused habitats (Mayr 1926; Svärdson 1949; Svanbäck et al. 2008). For example, at high levels of intraspecific competition, Arctic charr (Salvelinus alpinus) form distinct subpopulations differing in habitat use, behaviour, morphology and life history (resource polymorphism, Knudsen et al. 2006).
Structurally complex habitats like reed, submerged macrophytes, tree roots, stones or woody debris provide more niches and are generally the preferred habitat of benthic fish species. However, individual habitat specialization between structure-‐rich, near-‐shore littoral and open water has been observed frequently, for example in bluegill sunfish (Lepomis macrochirus), European perch (Perca fluviatilis) and roach (Rutilus rutilus; Ehlinger & Wilson 1988; Svanbäck et al. 2008). In most of the studies, habitat divergence was attributable to intraspecific competition (Svanbäck et al. 2008). Little is known which individual

Introduction
19
characteristics correspond to the habitat use differences. The individual difference in environmental choice may cause a diversification of temperament traits. For example, it can be assumed that less aggressive individuals may be displaced to less structured and less favourable habitats (Fausch 1984; Weber & Fausch 2003). Boldness may also be related to habitat use; individuals in less structured habitats may be exposed to a higher predation risk and show riskier and bolder behaviour (Magnhagen & Borcherding 2008). The level of boldness is then positively adapted to the level of predation risk (Brown et al. 2007). The other causal pathway implicates the possibility that individuals with stronger expressed boldness, which can be heritable (Brown et al. 2007), are more risk-‐prone and therefore “freer” in habitat choice. As a consequence they might exploit more open habitats. Irrespective of the causal pathway, it seems plausible that temperament traits may be related to the individual use of differently structured microhabitats.
Temperament traits
Temperament traits are increasingly used to highlight consistent differences in individual behaviour between and within animal populations (Réale et al. 2007). Recent research suggests that individual behaviour, once considered to be plastic, may be more predictable than previously thought (Bell et al. 2009). Theoretically, consistency in behaviour is likely because of reduced future costs of ecological adaptation when individual behaviour is established and fine-‐tuned (Wolf et al. 2008). Future costs involve the time, energy, and mortality costs of sampling the environment, or the costs of building and maintaining the required sensory machinery (Wolf et al. 2008). Therefore, even small positive feedback between the established behaviour and fitness may induce behavioural consistency (Wolf et al. 2008).
Temperament traits are summarized in five main categories: 1) boldness-‐shyness, an individual’s reaction to any risky situation, but not a new situation, 2) exploration-‐avoidance, an individual’s reaction towards a new situation; this includes behaviour towards a new habitat, new food or novel objects (in this paper: activity in a novel environment, interest in a novel food item), 3) activity, the general level of activity of an individual in a non-‐risky and non-‐novel environment, 4) aggressiveness, an individual’s antagonistic reaction towards conspecifics and 5) sociability, an individual’s reaction to the presence or absence of conspecifics, excluding aggressive behaviour (for more details on these definitions see Réale et al. 2007).

Chapter 1
20
To date, temperament traits have mostly been measured in single, isolated individuals in a laboratory environment. Whether behaviour in captivity predicts this behaviour in the wild, however, is seldom tested (but see Fraser et al. 2001; Herborn et al. 2010; Marentette et al. 2011). Undoubtedly, the increased complexity of a natural ecosystem with greater habitat variety and social or other behavioural interactions can result in a different reaction of an individual towards a challenging situation than in a laboratory environment. For instance, individual differences in temperament traits of pumpkinseed sunfish (Lepomis gibbosus) are relatively stable in nature but seem to disappear when the fish are held in social and ecological isolation in the laboratory (Wilson et al. 1993). Furthermore, pumpkinseed sunfish show markedly increased aggression to conspecifics and heterospecifics under isolated laboratory conditions compared to under field conditions (Coleman & Wilson 1998). Therefore, studies that compare temperament traits measured in single individual experiments in the laboratory with a non-‐isolated situation in the field are necessary (Verbeek et al. 1994; Herborn et al. 2010) and will enable evaluation of the transferability of laboratory results to “real world” circumstances.
Study species
A large family of non-‐migratory stream fish are freshwater sculpins (Cottus ssp.). Cottus ssp. are small benthic fish (< 20 cm; Kottelat & Freyhof 2007), known for their benthic, cryptic (e.g. Smyly 1957) and territorial behaviour (e.g. Cottus pollux, Natsumeda 2001; C. bairdii, Petty & Grossman 2004; C. rhenanus, Ovidio et al. 2009). Some species of this family (e.g. Cottus gobio) are protected in several European countries under the European Habitat Directive. This may be one reason why this stream fish family is increasingly being studied in recent years. Another reason may be the development of small passive integrated transponder (PIT) tags that facilitate the individual observation in the field (see further introduction). Numerous studies found remarkable intra-‐population heterogeneity in movement distances of Cottus ssp. (Breen et al. 2009; Hudy & Shiflet 2009; Ovidio et al. 2009). Whereas some individuals stay over the year in a restricted area often not larger than a few square meters, other individuals move more than one kilometre (Hudy & Shiflet 2009).
Similar differences in individual movement behaviour have been found in Cottus perifretum (Knaepkens et al. 2004, 2005). The popular name of C. perifretum, the bullhead, is identical to that of C. gobio and it was only recently that these two very closely related species were differentiated (Freyhof et al.

Introduction
21
2005). The distribution of C. perifretum encompasses Atlantic drainages from Garonne in France to Scheldt in Belgium but also major Rhine tributaries in France and Germany.
Bullhead have a strong affinity for complex habitats (Greenberg & Holtzman 1987; Davey et al. 2005; Legalle et al. 2005), which they use for spawning (C. pollux and C. hangiongensis, Fujimoto et al. 2008), parental care (Morris 1954), foraging or as refuge (Smyly 1957). It is only at night that bullhead wander into open water (Kobler et al. 2012a; C. pollux, Natsumeda 1998). However, where bullhead occur at high population density, sometimes being the dominant fish species in the ecosystem (Prenda et al. 1997; C. bairdii, McCleave 1964), not all individuals may have access to complex habitats (C. pollux, Natsumeda 2001).
It is well-‐known that freshwater sculpins are mainly active at night (e.g. Smyly 1957; Andreasson 1969, 1973; Brandt 1986; Natsumeda 1998) and can forage in total darkness (Hoekstra & Janssen 1985). The diel movement behaviour has, however, not yet been quantified. Relevant quantitative information is only available from aquarium observations (Andreasson 1969; but see Greenberg & Holtzman 1987; Natsumeda 2007 for quantified home range field data).
The reproductive behaviour of bullhead has been extensively studied and is very similar to that of the Gasterosteidae family (e.g. three-‐spined stickleback). It is characterized by female mate choice (Bisazza & Marconato 1988) and male parental care (Morris 1954; for other Cottus spp. see also Goto 1990). Females prefer larger males (Bisazza & Marconato 1988; C. pollux, Natsumeda 2001) and males whose nests already contain eggs (Marconato & Bisazza 1986). The nest is situated in a cavity that the male digs under solid habitat types such as rocks (Morris 1954) but can also be in tree root systems or woody debris. Males remain at the nest site for approximately three weeks as they find a mate, fan eggs (approximately 2 weeks) and guard young-‐of-‐the-‐year (approximately 1 week; Morris 1954). Inter-‐sexual differences in reproductive behaviours such as mate choice or parental care may cause sex-‐bias in movement distances. While this relationship has been extensively studied in birds and mammals, little is known regarding fishes.
Passive integrated transponder telemetry
PIT tags are small (> 11 mm) glass cylinders comprised of a coil and an integrated circuit, programmed to transmit one of some billions of codes. An induction coil is

Chapter 1
22
used to energize each tag, causing them to transmit their corresponding 10-‐digit alphanumeric code to the digital display of the reader in which the coil is housed (Hutchings & Gerber 2001). Roussel et al. (2000) investigated the application of a portable antenna to track PIT-‐tagged fish in shallow streams. In comparison to the previously used radio-‐telemetry, the portable PIT-‐tag antenna enabled observing fine-‐scale movements of smaller-‐bodied fish (Roussel et al. 2000). The size of utilized PIT-‐tags (23 × 4 mm) was used for fishes > 84 mm fork length and had a detection range of up to 100 cm (Roussel et al. 2000). In order to study even smaller-‐bodied fishes, a similar antenna with a detection range of up to 36 cm was developed for 11 × 2 mm PIT-‐tags (Cucherousset et al. 2005). Owing to the low detection range, the detection efficiency of the 11 mm PIT-‐tag antenna is dependent on the studied species (Cucherousset et al. 2005); less mobile fish such as the members of the genus Cottus (> 60 species) are more efficiently detected. The detection efficiency is also related to the stream conditions and was determined for Cottus ssp. in several conditions to be greater than 80% (Cucherousset et al. 2005; Keeler 2006; Keeler et al. 2007). Field (C. cognatus, Keeler et al. 2007) and laboratory observations (C. cognatus, Keeler 2006; C. bairdii, Ruetz et al. 2006; C. gobio, Knaepkens et al. 2007) revealed that surgical implantation of 11 mm PIT-‐tags had negligible effects on the survival and growth of Cottus ssp. > 50 mm total length. Furthermore, no influence on swimming capacity was found (C. gobio, Knaepkens et al. 2007). Therefore, PIT-‐tags enable researchers to gather more information about behaviour of individuals of small fish species in the wild. The necessity of clear water or good light conditions for visual observations (e.g. Petty & Grossman 1996, 2004, 2007) are no longer a prerequisite and freshwater sculpins can be observed with precision in turbid water and/or at night.
Objectives and thesis outline
The general aim of this thesis was to investigate whether intra-‐population heterogeneity in movement and habitat use of the bullhead is associated with individual characteristics such as sex, body size, body condition, growth rate, temperament traits and genetic diversity. A further aim was to test whether the temperament trait test results were independent of the applied methods and appropriate to draw conclusions about bullhead behaviour in the wild. Therefore, the comparability of field and laboratory behaviour as well as the behaviour of PIT-‐tagged and untagged individuals was evaluated. We also aimed to address other gaps in the knowledge about C. perifretum behaviour such as the quantification of movement distances during diel periods. The thesis includes six

Introduction
23
first-‐author articles, four of which are already published (chapters 2 to 5) and two that are in preparation (chapters 6 and 7).
The field study was conducted in the Laarse Beek, a lowland stream in the region of Antwerp, Belgium. Most of the work was carried out in the stream reach situated in the beech-‐oak forest Peerdsbos, between Brasschaat and Schoten, an area that is protected as EU-‐habitatzone without manipulation of the stream such as removal of woody debris. PIT-‐tagged bullhead were tracked weekly from February until November 2008 in stream reaches up to 2500 m length. Additional data from previous tracking studies of the Biology-‐Ethology research-‐group of Antwerp between March and May 2007 was also included in the thesis. The laboratory observation of behaviour was conducted subsequent to the field study at the Campus Drie Eiken of the University of Antwerp. Data from previous studies in April 2007 was used and own observations were made between October and November 2008. The genetic work was done at the Laboratory of Biodiversity and Evolutionary Genomics of the University of Leuven and carried out between 2009 and 2010. It comprised population genetic analysis and the analysis of individual genetic variability by using neutral markers (microsatellites).
Chapter 2: Comparability of laboratory and field behaviour
Bullhead were tested in the laboratory for explorative behaviour. Subsequently, the same individuals were released into the stream and again tested for exploratory behaviour. Owing to these two measurements, in the aquarium and in the stream, the comparability of laboratory and field observation could be assessed. Individuals had similar exploratory tendency under both experimental conditions. Individuals that were rather non-‐exploring and showed passive behaviour in both trials seemed to be more predictable than other phenotypes. These findings were a prerequisite for the following studies including behavioural observations in the laboratory and indicated that laboratory observations can help to predict behaviour under natural circumstances.
Chapter 3: Temperament traits and habitat use
Microhabitat use of bullhead was determined during three months in summer. During this period, the stream had only low water levels and the temporal occurrence of microhabitat types was relatively stable. In autumn, after recapture of the field-‐observed bullhead, individual temperament traits were examined in the laboratory. The temperament traits comprised boldness, activity,

Chapter 1
24
aggressiveness and exploratory tendency (interest in novel food and activity in a novel environment). Both microhabitat use and repeated temperament traits were individually consistent. Aggressiveness was negatively related to the use of structurally complex habitats (which are commonly preferred): bullhead that were more often found in structured microhabitats were less aggressive during the laboratory assay. It was speculated that this relationship was caused by a higher level of aggressive defence of less structured habitats.
Chapter 4: Diel movement of bullhead
Movement behaviour of bullhead was differentiated between the times of the day. In two years, different tracking protocols with different labour intensity were used. Bullhead moved furthest at night and shortest during daytime in 2007 and 2008. The difference in 2008 was, however, not significant. It is shown that this may be due to site-‐fidelity of some individuals: after swimming several metres at night, they returned to the location they previously occupied during daytime. It is discussed that longer tracking intervals used in 2008 (three times per 24-‐h instead of every two hours) were not suitable to detect the sheer magnitude of distances covered during a diel period.
Chapter 5: Period-‐dependent sex-‐biased movement
In this chapter, the long-‐ and short-‐term movement distance (movement range, respectively minimum displacement) of bullhead was analysed and compared between the sexes. Bullhead moved furthest in February and May. In these months, minimum displacements diversified between the sexes. Females moved furthest in February. This may be related to female mate-‐choice at the beginning of the reproductive period. In May, at the end of the reproductive period, males moved longest distances. It is speculated that males shift to resource-‐richer habitats after the starvation during parental care. Long-‐term movement range did not differ between the sexes (and was also not related to body size) but varied considerably between individuals ranging from 1 to 1111 m. The importance of a long study period (e.g. annual) to draw conclusions about sex-‐biased movement is discussed.
Chapter 6: Movement range and the relation to temperament traits
After the previous chapter in which periodical inter-‐sexual differences in movement distances were observed, chapter 6 aimed to get more insights into between-‐individual differences in long-‐term movement range. Individual

Introduction
25
movement range information that was collected in the stream was related to temperament traits observations in the laboratory. We also tested whether the expression of temperament traits was different between tagged and untagged bullhead; this was done to test whether observed behaviours were independent of the used (tagging) method. There were no behavioural differences between tagged and untagged bullhead. Repeated temperament traits (activity and novel environment activity) indicated behavioural consistency. The comparison between individual movement range and temperament traits revealed weak positive but not significant correlations between movement range and boldness or interest in novel food. This might suggest that bullhead with longer movement range are more risk-‐prone. Other temperament traits were not related to movement range.
Chapter 7: Relating movement range to fitness and heterozygosity
This chapter aimed to identify which other individual characteristics that were not yet covered in chapter 5 and 6 are related to the movement range diversification in the studied bullhead population. It was examined whether body condition, growth rate or genetic diversity (individual microsatellite heterozygosity) would help to understand intra-‐population heterogeneity in movement range. The individual movement range varied from 1 to 1096 meters. These differences were not related to body condition, growth rate or heterozygosity (single-‐ or multilocus). Thus, there was no indication of fitness differences that were associated with individual movement range. There was also no indication that genetic diversity was associated to fitness because body condition and growth rate were not related to heterozygosity.
Chapter 8: General conclusions and future research
In the last section, the results of all chapters are integrated, conclusions are made and future research possibilities are being proposed.


CHAPTER 2
COMPARISON OF LABORATORY AND FIELD BEHAVIOUR


Comparison of laboratory and field behaviour
29
Temperament in bullheads: do laboratory and field explorative behaviour variables correlate?
Alexander Kobler, Brecht Engelen, Guy Knaepkens, Marcel Eens
Naturwissenschaften (2010) 96: 1229-‐1233
Abstract
The relevance of temperament traits for life history strategy or productivity is increasingly acknowledged. Temperament traits are often either observed in captivity or in the wild, but studies combining both observations are very rare. We examine whether exploratory behaviour in the bullhead (Cottus perifretum), assayed under laboratory conditions, predicts this behaviour under field conditions. Forty-‐three PIT-‐tagged individuals were first assayed for exploration of a novel environment in the aquarium and then released into an unfamiliar stream stretch, where they were later relocated using a mobile antenna. Explorative behaviour assayed in the laboratory was significantly positively related to the exploration in the field, thus predicting distance moved in the field release. Both in the laboratory and in the field, explorative behaviour was not related to individual body length. When bullheads that did not leave the refuge in the aquarium (laboratory assay) and, therefore, did not explore the new environment were excluded from the analysis, the correlation between laboratory and field explorative behaviour variables became weaker. However, overall, our results illustrate that exploration rate of bullheads in isolated single-‐individual experiments can be used to predict this behaviour in the natural ecosystem.

Chapter 2
30
Introduction
Behavioural differences within a species have long been treated as insignificant variation within the average behaviour of the species as a whole. However, due to its ecological and evolutionary relevance, inter-‐individual variation in behaviour and, more specifically, in temperament traits like exploration, aggressiveness, activity, boldness or sociability are of increasing interest. To date, temperament traits have mostly been measured in single, isolated individuals in a laboratory environment. Only a small number of studies have measured temperament traits in the field and studies combining both laboratory and field observations are rare (but see Wilson et al. 1993; Fraser et al. 2001; Dingemanse et al. 2003; Wilson & McLaughlin 2007).
Undoubtedly, the increased complexity of a natural ecosystem with greater habitat variety and social or other behavioural interactions can result in a different reaction of an individual towards a challenging situation as opposed to being in a laboratory environment. For instance, individual differences in the temperament of pumpkinseed sunfish (Lepomis gibbosus) are relatively stable in nature but seem to disappear when the fish are held in social and ecological isolation in the laboratory (Wilson et al. 1993). Furthermore, pumpkinseed sunfish show markedly increased aggression to conspecifics and heterospecifics under isolated laboratory conditions compared to under field conditions (Coleman & Wilson 1998). In birds, slow exploring great tits (Parus major) explore a new environment significantly faster when observed together with a companion (van Oers et al. 2005). Thus, studies comparing individual temperament measured in single-‐individual experiments in the laboratory with a non-‐isolated situation in the field are necessary (Verbeek et al. 1994) and will enable evaluation of the transferability of laboratory results to “real world” circumstances.
One of the main categories of temperament research and, from an ecological point of view, possibly the main target of selection is exploration (Smith & Blumstein 2008), an individual’s reaction to a new situation including the behavioural response towards a new habitat, new food or novel object (Réale et al. 2007). A species which is particularly suitable to study individual differences in exploration is the bullhead (Cottus perifretum), a small (up to 100 mm) bottom-‐dwelling freshwater fish species that exhibits considerable variability in movement behaviour (Knaepkens et al. 2004, 2005). Due to the hidden nature of bullhead, which seeks shelter for most of the time in structured habitats (Smyly 1957), swimming activity is restricted. Movement in an unfamiliar environment is

Comparison of laboratory and field behaviour
31
therefore a good indication of the exploration rate in bullhead (compare Verbeek et al. 1994).
The objective of the present study is to measure inter-‐individual differences in the exploration of C. perifretum in the laboratory and the field and to correlate these measures to value the predictability of natural ecosystem exploration by a laboratory assay.
Materials and methods
On 2 April 2007, 43 adult bullhead individuals ≥ 50 mm [total length (TL), minimum size to PIT tag, see further] were collected in the Laarse Beek, a small lowland stream of the Scheldt basin (Flanders, Belgium), using electric fishing gear (WFC7, 150 V DC, Electracatch Int. Ltd, UK). Fish were transported to the laboratory and a PIT tag (12 × 2.1 mm, 94 mg, ID100, EID Aalten, NL) was inserted into the abdominal cavity of the anaesthetised (using 0.25 ml L−1 2-‐phenoxy-‐ethanol) individual through a 2-‐ to 3-‐mm incision, a method which was shown to have negligible effects on bullhead survival, growth and swimming performances (Knaepkens et al. 2007). Tagging enabled individual recognition during the laboratory and field release experiment (see further). Fish were held in two 400-‐L tanks in oxygenated (using air stones) and filtered (bio-‐filter) tap water with PVC tubes (10 × 5 cm) and roof tiles as refuges to allow bullheads some cover. Fish were fed daily to saturation with frozen bloodworms.
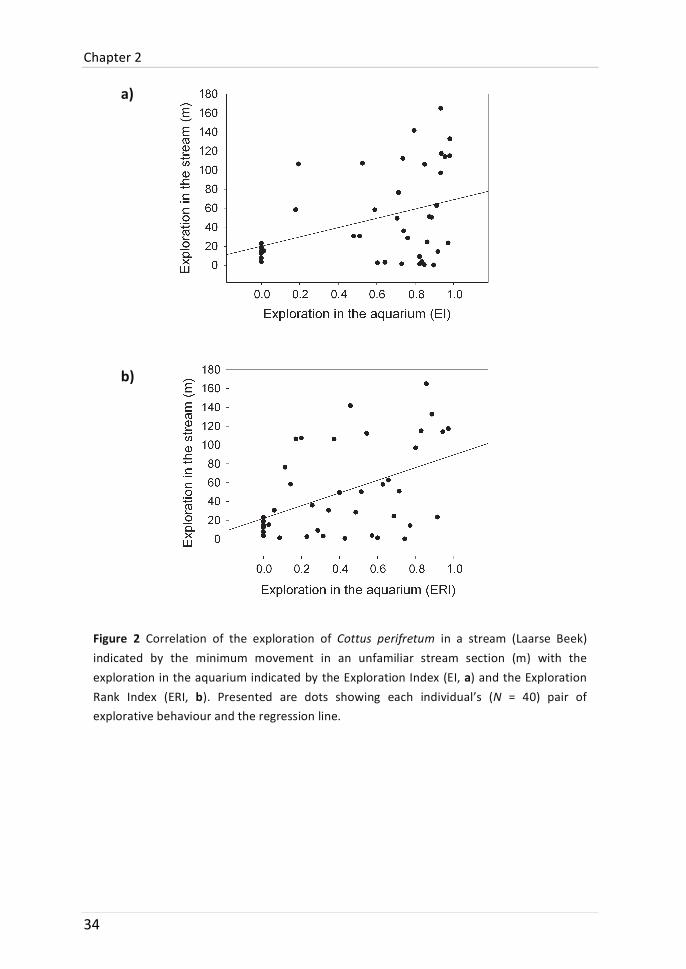
In the laboratory, exploration tests were recorded in the absence of an observer by a video camera (DCR-‐PC6E, Sony, Japan) in very low lighting, using the “nightshot” function, because the bullhead is known to be crepuscular and night active (Smyly 1957). Tests lasted 40 min and started 1 week after the initial capture. Individuals were placed in a familiar refuge [PVC tube, section 1 (S1); Figure 1] in a novel environment (aquarium, 120 × 40 × 40 cm). Similar to Fraser et al. (2001), two indices of activity in a novel environment were used, the Exploration Index (EI) and the Exploration Rank Index (ERI) (indices were renamed due to recent definitions of Réale et al. 2007). The EI is based on the time spent out of the refuge and calculated for the ith individual as EIi = (ti / T), where ti = time spent out of the refuge by the ith individual and T = total time in test (2,400 s).

Chapter 2
32
The ERI is based on the rank order of reaching the different sections of the aquarium (Figure 1). Rank 1 is assigned to the individual reaching S4 in the least amount of time, rank 2 to the next shortest time, until all individuals reaching S4 are ranked. The next ranks are assigned similarly first for individuals reaching S3 fastest (but never S4) followed by the fastest S2 reaching individuals (but never S3). Then, the individuals which never reached S2 are ranked by the shortest latency to leave the refuge. Finally, the last rank (R) is assigned to the remaining individuals that never left the refuge. The ERI for the ith individual is then calculated as ERIi = 1 -‐ (ri / R), where ri = rank of the ith individual and R = the last assigned rank in the sample. Bullheads with an ERI or EI value of 0 (individuals which never left the refuge) are from now on called non-‐exploring individuals.
After the laboratory observations and an additional week of holding in 400-‐L tanks, a release experiment in a novel stream section, as field counterpart to the aquarium observations, was conducted. The abundance of structured habitats in this stretch was low and previous electro-‐fishing confirmed that in this section ecosystem suitability for bullhead might be low because bullheads were extremely rare. We, therefore, expected long distance instead of fine-‐scale exploration in the released individuals. Furthermore, potential aquatic predators were not found there. The fish were released on 18 April 2007 at 9 A.M. together with a roof tile as refuge in a 20 m part of a novel stream section, which was situated 750 m upstream from the initial capture point. Release density was comparable to population density where they were originally caught (personal observation). Bullheads were tracked 24 h after release using a mobile PIT tag antenna (details in Cucherousset et al. 2005). This time interval was chosen to
Figure 1 The experimental aquarium was visually subdivided into four equal sections by putting tape on the observer side [section 1 (S1) to section 4 (S4)] with a familiar refuge (PVC tube) at both ends of the aquarium (S1 and S4). The intervening gap was unstructured.

Comparison of laboratory and field behaviour
33
ascertain possible movement because bullheads are known to be active mostly in the dark (Smyly 1957). Comparably to the aquarium observations, positions were used to calculate a minimum straight-‐line movement over time which served as the variable of the exploration for the field release experiment (here, m per 24 h).
In exploration studies, individuals who do not explore the experimental setup at all are sometimes excluded from analysis (e.g. Bell 2005). Correlations of the exploration variables (laboratory and field) were, therefore, run including (N = 40) as well as excluding (N = 34) the non-‐exploring (for definition, see previous) individuals. Additionally, we tested all three exploration variables on possible effects of body length (TL, mm). All variables but ERI without non-‐exploring individuals were non-‐normally distributed (Shapiro–Wilk, all p < 0.05). Spearman rank correlations were therefore used in all calculations. Statistical analyses were conducted with SPSS 15.0 (SPSS Inc., Chicago, IL, USA). Significance levels were adjusted (from originally α = 0.05) for the group of the exploration correlations (N = 4 correlations) as well as for the group of the correlations with body length (N = 3 correlations) using the false discovery rate (Benjamini & Hochberg 1995).
Results
As a general rule, bullheads remained motionless for a few minutes in the aquarium and explored the new environment in straightforward swimming. Exploration was inter-‐individually heterogeneous, ranging from individuals that did not leave the refuge (N = 6; non-‐exploring), swam half distance (N = 8), to individuals reaching the opposite side of the aquarium (N = 29) in time periods of between 73 and 2,323 s (mean ± SD, 618.1 ± 555.7 s).
We were able to relocate 40 of the 43 laboratory-‐assayed individuals 24 h after release in the novel area of their home stream. Straight-‐line minimum moved distance (indicating the exploration) varied considerably amongst individuals, ranging from 0 to 165 m (50.6 ± 47.9 m).
Chapter 2
34
a)
b)
Figure 2 Correlation of the exploration of Cottus perifretum in a stream (Laarse Beek) indicated by the minimum movement in an unfamiliar stream section (m) with the exploration in the aquarium indicated by the Exploration Index (EI, a) and the Exploration Rank Index (ERI, b). Presented are dots showing each individual’s (N = 40) pair of explorative behaviour and the regression line.

Comparison of laboratory and field behaviour
35
Exploration in the aquarium (EI and ERI) was significantly positively correlated to the exploration in the stream (N = 40; αadjusted = 0.025; EI: rs = 0.358, p = 0.023; ERI: rs = 0.407, p = 0.009; Figure 2a, b). However, when non-‐exploring individuals were removed from the analysis, correlation coefficients became lower and significance vanished (N = 34; αadj = 0.025; EI: rs = 0.242, p = 0.168; ERI: rs = 0.298, p = 0.087).
There was no significant correlation of both exploration indices in the aquarium (N = 40; EI: rs = −0.171, p = 0.291; ERI: rs = −0.124, p = 0.444) as well as the minimum movement in the novel stream section (N = 40; rs = −0.091, p = 0.575) to individual body size (63.9 ± 12.8 mm TL; range 50 to 98 mm).
Discussion
Studies which directly compare one particular temperament trait measured in the laboratory with observations in the field are rare although they show the transferability of isolated behaviour in a non-‐natural environment to the more complex natural ecosystem. Our study showed that explorative behaviour variables in bullhead are repeatable from an experimental situation in the aquarium to the circumstances in a natural environment. Comparable aquarium–stream repeatability with similar methods and variables was also observed in the freshwater killifish (Rivulus hartii, r = 0.41 to 0.48; Fraser et al. 2001). However, Wilson et al. (1993) observed that, although individual responses to novel situations of pumpkinseed sunfish (L. gibbosus) are inter-‐individually different and stable in nature, these inter-‐ individual differences vanished after pond-‐observed sunfish were caught and held in social and ecological isolation in aquaria over a longer time period. The social surrounding may, therefore, be an important determinant of individual fish’s exploratory behaviour (Magnhagen 2007) and holding time and group composition in captivity could be important variables on the original behaviour.
Although correlations of laboratory and field release of this study were significant, it is crucial to bear in mind that correlations were not high (EI, rs = 0.36; ERI, rs = 0.41). Particularly the individuals who did not leave the refuge in the aquarium (non-‐exploring) repeated their passive behaviour and showed only little variance in movement in the stream-‐release experiment (Figure 2a, b). Fish with higher levels of EI and ERI were more heterogenic and relative repeatability was lower. In this study, it seemed that individuals with the lowest exploration were more predict-‐ able than other phenotypes (compare with Sinn et al. 2008),

Chapter 2
36
although this should be examined in more detail.
Overall, our results imply slightly weaker short-‐term repeatability in comparison to other studies which examined explorative behaviour in one environmental setting only in fish (r = 0.68, Neolamprologus pulcher; Bergmüller & Taborsky 2007), insects (r = 0.51, Dolomedes triton; Johnson & Sih 2007), cephalopods (r = 0.39 to 0.54, Euprymna tasmanica; Sinn & Moltschaniwskyj 2005) and birds (r = 0.38 to 0.49, Fringilla coelebs; Quinn & Cresswell 2005). However, as we did not repeat our aquarium observations, it is not possible to distinguish between relative repeatability from laboratory to field and the repeatability per se. We are also well aware that the behavioural measures we used are only approximate values for the exploration avoidance. For instance, although the novel stream section of this study was of comparably lower quality than the original habitat and we therefore expected the fishes to move long distances during exploration, individuals who would have explored in finer scale but with similar intensity would have received a relatively lower field score.
The individual variation in exploration rate for the bullhead was not related to the total length of the fish. Other studies have speculated that a higher metabolic rate is responsible for faster exploration by smaller fishes (Brachyraphis episcopi; Brown et al. 2005). However, we did not find any evidence for this hypothesis despite the size range of the tested individuals being broad ranging from 50 to 98 mm. Although most previous studies have shown that the sex does not have an influence on the explorative behaviour of fish (Wilson et al. 1993; Fraser et al. 2001; Moretz et al. 2007; but see Brown et al. 2005), unfortunately we could not examine this here because, outside the spawning period, the bullhead cannot reliably be sexed based on external morphology (J. Freyhof, personal communication) and to date no genetic sex markers are available.
We assume that the exploration behaviour of bullhead, with correlating observations in the aquarium and in a stream, corresponds to further temperament traits and may be linked to the individual’s metabolism. Future studies should examine both the repeatability within a context and across contexts. A control for the social and ecological environment would elucidate to what degree the exploration of an individual is intrinsic (e.g. linked to the metabolism) and whether the individual rank in the group for the explorative behaviour remains the same despite environmental changes.

Comparison of laboratory and field behaviour
37
Acknowledgments
We wish to thank N. Adam, I. Boost, T. de Groote, S. Delen, J. Demeulemeester, D. Geelhand, K. Geudens, J. Hoefnagels, Y. Humblet, C. van de Sande and A. Vankerkhove, G. Eens, P. Scheys for technical assistance and J. Meaney-‐Ward for English proofreading. We also wish to thank four anonymous referees and T. Czeschlik for helpful comments and the FWO Flanders for funding (FWO postdoctoral grant to GK and FWO project to ME). This study was conducted under licenses of the relevant authorities that permitted us to capture and manipulate the bullheads.


CHAPTER 3
TEMPERAMENT TRAITS AND HABITAT USE


Temperament traits and habitat use
41
Temperament traits and microhabitat use in bullhead,
Cottus perifretum: fish associated with complex habitats are less aggressive
Alexander Kobler, Gregory E. Maes, Yves Humblet, Filip A.M. Volckaert, Marcel Eens
Behaviour (2011) 148: 603-‐625
Abstract
Temperament traits have been linked to fitness-‐related functional contexts such as dispersal or mating attractiveness, but few studies have linked inter-‐individual differences in habitat use to temperament traits. Therefore, we conducted a three-‐month field study with weekly tracking to define the individual microhabitat use of bullhead (Cottus perifretum). The species is known for its dependence on structured habitats. At the end of the field survey, bullhead were recaptured and tested in the laboratory for five temperament traits: boldness, interest in novel food, novel environment activity, aggressiveness and activity. Repeated trait tests (activity, r = 0.439; novel environment activity, r = 0.422) and habitat complexity use (r = 0.568) indicated behavioural consistency. Overall, bullhead significantly preferred complex habitats, such as branch jams, while avoiding open water. Individual frequency in the use of complex habitats could not be attributed to size or sex differences, but was significantly negatively correlated to the level of aggressiveness. We hypothesize that this relationship was caused by a higher level of aggressive defence of less structured territories. Other temperament traits were not significantly linked to individual habitat use. To our knowledge, this study is the first to show a relationship between aggressiveness measured under laboratory conditions and the use of complex habitats in situ.

Chapter 3
42
Introduction
One of the most important methods of reducing competition between species is niche partitioning by habitat use (Schoener 1974). This has also been shown to exist within single populations of one species (Bolnick et al. 2003). Some individuals of a population expand to unused or underused habitats when competition passes a threshold (Mayr 1926; Svanbäck et al. 2008). For instance, many lentic fish species prefer the near-‐shore littoral habitat; when faced with tighter competition, some individuals of a population use the less preferred open water habitat (e.g. Svanbäck et al. 2008). Such diversity in microhabitat use may occur in the absence of differences in sex, size or age, but could be related to other behavioural characteristics (Bolnick et al. 2003). However, little is known about the relation between the diversity in habitat use and other behavioural traits. Animal temperament (Réale et al. 2007) comprises a range of behavioural traits that might help to increase understanding of heterogeneous microhabitat use.
Temperament traits are increasingly used to highlight consistent differences in individual behaviour between and within animal populations (Réale et al. 2007). Populations have been subdivided into temperament types such as either bold and shy (Wilson et al. 1994; Réale et al. 2000; Sinn et al. 2008), fast and slow explorative (Verbeek et al. 1994) and responsive versus unresponsive (Wolf et al. 2008). Between-‐individual differences in temperament traits have been linked to fitness-‐related functional contexts (reviewed by Schuett et al. 2010) such as behaviour of bird singing (Garamszegi et al. 2008), brood care helping (Bergmüller & Taborsky 2007), mating attractiveness (Godin & Dugatkin 1996) and mating success (Reaney & Backwell 2007), dispersal or movement range (reviewed by Clobert et al. 2009) and growth (Millot et al. 2009). Surprisingly, relationships between individual microhabitat use in nature and temperament traits have hardly been studied (but see Boon et al. 2008).
An important question that arises is whether there is relation between temperament traits and individual use of structured habitats. The individual difference in environmental choice may cause a diversification of temperament traits. For example, individuals in the less structured habitat may be exposed to a higher predation risk and show riskier and bolder behaviour (Magnhagen & Borcherding 2008). The level of boldness is then positively adapted to the level of predation risk (Brown et al. 2007). The other causal pathway implicates the possibility that individuals with stronger expressed boldness, which can be heritable (Brown et al. 2007), are more risk prone and, therefore, ‘freer’ in habitat choice. As a consequence they might exploit more open habitats. Irrespective of

Temperament traits and habitat use
43
the causal pathway, it seems plausible that temperament traits may be related to the individual use of differently structured microhabitats.
Structurally complex habitats provide more niches, facilitate the exploitation of the environment and may increase the species richness of an environment (Tews et al. 2004). In aquatic ecosystems, the presence of complex habitats like reed, submerged macrophytes, tree roots, stones or woody debris can be directly linked to the reproductive success and abundance of individuals in a fish population (e.g. Grimm 1981).
Many fishes that are known for their strong dependence on complex habitats belong to the family of common sculpins (Cottidae, approximately 275 spp). Common sculpins consist mainly of small (< 15 cm) fishes, which are well camouflaged, have a reduced swim bladder and live in the demersal zones of lakes and streams. Similar to the Salmonidae, some species of Cottidae can exhibit non-‐breeding territoriality (Petty & Grossman 2007). They have a strong affinity for complex habitats (Greenberg & Holtzman 1987; Davey et al. 2005; Legalle et al. 2005), which they use for spawning (Fujimoto et al. 2008), parental care (Morris 1954), foraging or as refuge (Smyly 1957). It is only at night that adult common sculpins wander into open water (Natsumeda 1998; Kobler et al. in preparation). Where common sculpins occur at high population density, sometimes being the dominant fish species in the ecosystem (McCleave 1964; Prenda et al. 1997), not all individuals may have access to complex habitats (Natsumeda 2001).
A member of the Cottidae family, the bullhead (Cottus perifretum), was chosen to study possible relations between microhabitat use and temperament traits. Due to the importance of structured habitats as refuge shelter (Davey et al. 2005) and foraging habitat (Smyly 1957), it can be assumed that less aggressive individuals may be displaced to less structured and less favourable habitats (Fausch 1984; Weber & Fausch 2003). We further hypothesized that bullhead activity is negatively correlated to the inhabited level of habitat structure. Other studies found that fish inhabiting less structured habitats are more active (Sundbaum & Näslund 1998; Salvanes & Braithwaite 2005), which might be related to differences in foraging tactics (Kobler et al. 2009b). As mentioned earlier on, boldness can be negatively related to habitat complexity because of a higher predation risk in open habitats (Magnhagen & Borcherding 2008). However, we did not expect a strong relation between boldness and the individual use of complex habitats as predator abundance was low in the study system and predation risk presumably low.

Chapter 3
44
Materials and methods
Study area and habitat determination
The study was conducted in the Laarse Beek (51°16′28.40′′N, 4°29′4.63′′E), a lowland stream in northern Flanders, Belgium. The focal section of the stream was 324 m long, had a mean width of 2.87 m (range 2.09–3.53 m) and a mean depth of 0.3 m (range 0.1–0.5 m). It crossed a beech-‐oak forest with black alder (Alnus glutinosa) rooting partly in the stream. The stream bed was characterized by a sandy soil with woody debris patches. Bullhead were the most abundant fish species in the stream with a density of approximately 2.1 adults per m2 (see below). Aquatic predators, such as pike (Esox lucius), eel (Anguilla anguilla) and perch (Perca fluviatilis), were rare (personal observation). Similarly, the predation risk through avian predators such as grey heron (Ardea cinerea) and kingfisher (Alcedo atthis) was probably low, as they appeared only a few times.
We characterized the habitat types and the structure of the habitat types in two days (26 and 27 June 2008). The stream section was divided into 972 quadrats of approximately 1 m2 (mean 0.96 m2), three quadrats covered the stream width and 324 the stream length. The quadrats were assigned to one of the six main habitat types according to the dominant habitat type (> 50% abundance): open water which consisted either of a bare sand bottom or a limited abundance of twigs (approximately 553 m2), tree branches (woody debris, approximately 279 m2), tree roots (approximately 48 m2), naturally formed tree branch jams (approximately 34 m2), a stone pile at the foot of a bridge (approximately 34 m2) and submerged water plants (approximately 24 m2). The quadrats were categorized according to the degree of habitat complexity, because bullhead have a strong affinity for complex habitats (see introduction). Habitat complexity was defined at three levels based on its occurrence in the field. It was done to have a methodological way to clearly distinguish habitat complexity per quadrat. Quadrats with < 33% structure and mainly consisting of open water, received a habitat complexity value of 0 (approximately 616 m2). A habitat complexity value of 1 was given when ≥ 33% and ≤ 66% structure was provided by intermediate abundances of branches, tree roots, naturally formed branch jams, stones and/or water plants (approximately 252 m2). If the stream quadrat was highly structured with > 66% of complex habitats, it received a complexity value of 2 (approximately 104 m2).
A second habitat characterization was conducted on 17 October 2008 to evaluate the temporal consistency of the spatial occurrence of the habitat types

Temperament traits and habitat use
45
and the habitat complexity levels. The second characterization was carried out every 10 m by defining three quadrats in stream width resulting in 96 re-‐evaluated quadrats.
Capture and tagging
The focal section of the stream was marked every other 2 m with consecutively numbered wooden poles along the right stream bank. Bullhead were caught in two sessions, on 4, 5 and 7 February and on 13 and 14 February 2008 by electro-‐fishing (WFC7-‐10, Electracatch, Wolverhampton, UK) using a 40 cm ring anode. During the two electro-‐fishing sessions 348 bullhead > 50 mm [minimum size to tag with passive integrated transponders (PIT), Knaepkens et al. 2007] were caught including 14 recaptures during the second session. Bullhead (N = 334) had a mean total length (TL) of 62.1 ± 6.3 mm (mean ± SD; range 51–88 mm) and mean wet weight of 3.3 ± 1.2 g. Fish were anaesthetized (0.25 ml L-‐1 2-‐phenoxy-‐ethanol) and individually coded 12 mm PIT-‐tags (12 × 2.1 mm, 0.094 g; ID100, EID, Aalten, The Netherlands) were inserted horizontally through a 2-‐mm incision into the body captivity (by using a scalpel). Due to the small size of the fish and the opening, the incision was not stitched (Bruyndoncx et al. 2002). After implantation, bullhead were allowed to recover in buckets with fresh stream water and released at their capture position. Due to tagging shortly before spawning, the most bullhead could be reliably sexed (138 females and 139 males). Gonadal development did not allow to use weight as a variable as it was strongly gender-‐dependent (females were proportionally much heavier) and also related to the stage of gonadal development. Using the capture-‐ recapture data of the two electro-‐fishing sessions, an abundance estimate of 2006 bullhead > 50 mm (confidence interval 1228–3459) for the 324 m section was calculated (Petersen single census method, Chapman modified; Ricker 1975).
Tracking and recapture
Tracking took place between 12 June and 19 September 2008. This period was chosen as spawning at the Laarse Beek generally ends in May (Knaepkens, personal communication). Male egg guarding (nesting) behaviour should have ended also (see Morris 1954) and males are no longer restricted to the nest site and hence less constrained in habitat choice. The 324 m electro-‐fishing section was scanned by a mobile PIT-‐tag antenna with a detection precision of < 30 cm (Cucherousset et al. 2005). Tracking was conducted during the day, which is a time when bullhead prefer structured habitats (see Introduction), by carefully walking through the stream and moving the antenna like a vacuum cleaner. The

Chapter 3
46
magnetic field of the antenna was always moved with a distance of < 30 cm to the ground (bullhead are bottom-‐dwelling). To increase the probability of detection in stream-‐bank habitats, such as tree roots, the antenna was turned by 90 degrees and moved directly along the shoreline. Once a bullhead was located, the square in which it was found was noted. After its detection and during recording of the stream position (e.g. 1400 stream m, left side), the antenna was kept above the detected bullhead. An escape response was never observed and the tagged individuals relied on their camouflage, a typical behaviour for Cottus ssp. The immobile behaviour was a prerequisite of the methodological independence of temperament traits and habitat use. It excludes for example that bolder individuals were more likely detected in open water while more shy individuals preferred the more structured habitats following a flee attempt.
Tracking sessions took place three times per month (with at least 1-‐week intervals). During each session the stream section was scanned once on two consecutive days (two scans per session), to increase the probability that each individual was located once. To equalize the disequilibrium of the individual localizations (not all individuals were found twice) we only used the first localization of each tracking. This led to a total maximum of 15 individual tracking locations during the tracking period (12 June to 19 September 2008).
From 22 September to 19 November 2008 we attempted to recapture tagged bullhead in the 324 m section. The fish were localized in two screenings with the mobile PIT-‐tag antenna. The spotted individuals were then caught with a big landing net (one person) and two small aquarium nets (another person), which was more efficient than electro-‐fishing. In total 20 tagged individuals were caught with a TL (mean ± SD) of 64.8 ± 6.31 mm (range 55–77 mm) and a mean wet weight of 3.72 ± 0.99 g (range 2.2–6.7 g). The small number of recaptures was probably due to the large dispersal ranges of the bullhead (up to 1284 m range; Kobler et al., data not shown) but also due to natural mortalities; bullhead are short-‐lived. Every week slightly smaller, untagged bullhead (TL 48.41 ± 5.14 mm; range 40–62 mm) were caught; they served as stimuli for the subsequent aggressiveness test in the laboratory (see below). Sex could not be determined of the stimulus fish, as they were not in breeding condition. On the day of capture, the fish were transferred to a 400-‐L tank with oxygenated and bio-‐filtered tap water at the university wet lab. The tank contained PVC tubes (10 × 5 cm) as structuring refuges; the mean ± SD water temperature of 9.8 ± 2.7 °C (range 5.3–14.1 °C) and the mean oxygen concentration of 9.1 ± 1.2 mg L-‐1 (range 7.7–11.1 mg L-‐1) were constantly monitored. Bullhead were fed daily to satiation with frozen bloodworms.
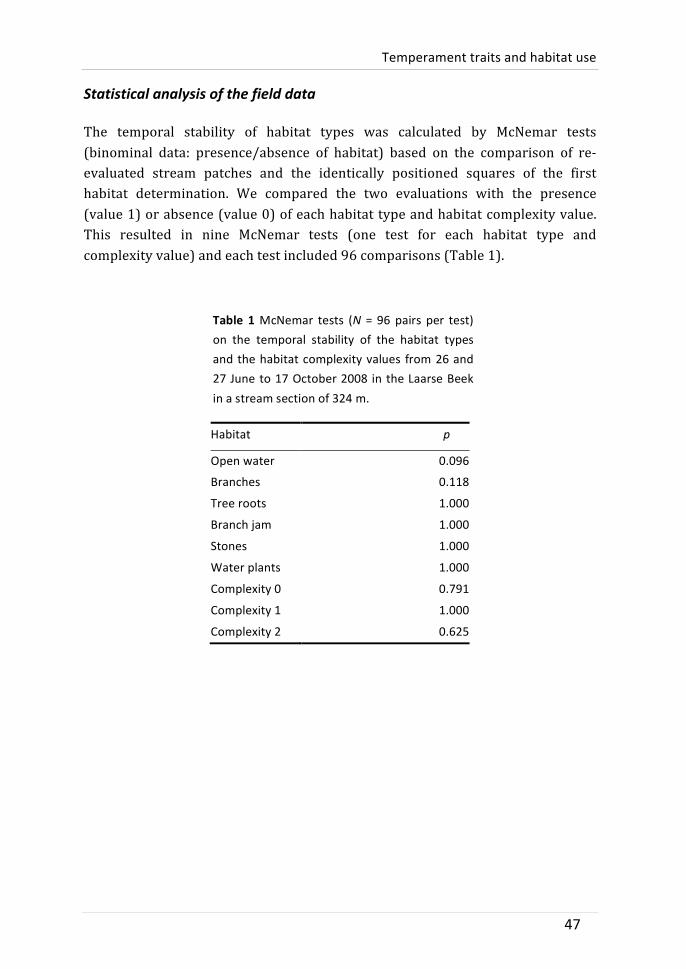
Temperament traits and habitat use
47
Statistical analysis of the field data
The temporal stability of habitat types was calculated by McNemar tests (binominal data: presence/absence of habitat) based on the comparison of re-‐evaluated stream patches and the identically positioned squares of the first habitat determination. We compared the two evaluations with the presence (value 1) or absence (value 0) of each habitat type and habitat complexity value. This resulted in nine McNemar tests (one test for each habitat type and complexity value) and each test included 96 comparisons (Table 1).
Habitat p
Open water 0.096
Branches 0.118
Tree roots 1.000
Branch jam 1.000
Stones 1.000
Water plants 1.000
Complexity 0 0.791
Complexity 1 1.000
Complexity 2 0.625
Table 1 McNemar tests (N = 96 pairs per test) on the temporal stability of the habitat types and the habitat complexity values from 26 and 27 June to 17 October 2008 in the Laarse Beek in a stream section of 324 m.

Chapter 3
48
To examine whether selection is occurring for individual habitat types or habitat complexity levels by some of the PIT tagged individuals, we used an individual-‐based chi square test for repeated measures by Manly et al. (1993) (see Rogers & White 2007). Following the notation by Manly et al. (1993), uij is the amount of habitat type i used by fish j; ui+ is the amount of habitat type i used by all fish; u+i is the total amount of habitat units used by the fish j; and u++ is the total number of habitat units used by all fish.
X 2 = 2 uij loge uij / E uij( )!" #$i=1
I
%j=1
n
% ,
where E(uij ) = ! iu+ j , and πi is the proportion of available resource units in category i (Rogers & White 2007).
Which habitat types or complexities the bullhead selected was calculated by selection ratios and their associated Bonferroni-‐adjusted 95% confidence intervals (individual-‐based log-‐likelihood test for repeated measures; Manly et al. 1993; Rogers & White 2007). Following the notation by Manly et al. (1993), the selection by the population for the i-‐th habitat type is estimated by
wij = uij / (! i +u+ j ) .
The selection ratio estimates are generated by pooling observations from all tagged fish, but the equation takes variation in selection from individual fish into account (Manly et al. 1993). Habitat selection was considered significant if the lower confidence interval (CI) of the selection ratio was greater (preference) or the upper CI was smaller (avoidance) than 1. For CI generating for the selection ratios see Rogers & White (2007).
The consistency (repeatability, r) of individual habitat complexity use was calculated by using mean squares of among-‐groups and within-‐groups variance of a one-‐way ANOVA with ‘individual’ as a factor on the dependent variables (Lessells & Boag 1987).
r = s2^ / s2 + s2^( ) ,
where s2^ is the among-‐groups variance component and s2 is the within-‐group variance component.

Temperament traits and habitat use
49
Due to the unbalanced number of individual habitat localizations, the co-‐ efficient (n0) related to the sample size had to be calculated (here n0 = 10.439; Lessells & Boag 1987). Individual habitat complexity use was calculated by averaging the habitat complexity values of each individual’s localizations. These individual values were related to TL (mm) and sex by an ANCOVA (variance homogeneity of sex, Levene-‐test, p > 0.05).
Significance of all calculations was assessed at α < 0.05. Statistical analyses were conducted with SPSS 15.0 (SPSS, Chicago, IL, USA). Habitat selection ratio was calculated by the software package Fishtel 1.4 (Rogers 2002).
Laboratory analyses of temperament traits
We used for the experiments eleven plastic tanks (79 × 42 × 57 cm), lined with blue plastic foil. The tanks were half-‐filled and contained approximately 80 L of oxygenated tap water with a mean temperature of 14.9 ± 1.5 °C (range 12.7–17.2 °C) and a mean oxygen concentration of 7.8 ± 0.8 mg L-‐1 (range 6.5– 9.6 mg L-‐1). We adjusted the light/dark cycle weekly to the seasonal natural conditions. Light intensity was dimmed to mimic the stream conditions in the forest.
The tanks were set up such that all tests for each individual could be con-‐ ducted in just one tank (Figure 1), although two transfers into novel environments and the introduction of a predator were done (see later). Therefore, the tanks were temporally visually (white plastic plate) and constantly physically (metal mesh) divided into two parts. Ten percent of the bottom of both tank sides (A and B) was structured; different objects created two different environments (Figure 1). Under the transparent bottom of the tank a grid of 7.5 × 7.5 cm squares was drawn; it was chosen in view of the maximum size of the bullhead (70 mm TL). Above the tanks we positioned a digital camcorder (DCR-‐PC6E or DCR-‐DVD310E, Sony, Tokyo, Japan).
Under the artificial tank conditions bullhead were mostly hidden in a refuge and showed no activity at all during the simulated daylight. To record behavioural activity, the tests were recorded at the beginning of the night period by using the night-‐shot function of the camcorders. Therefore, our study compares bullhead daytime shelter use in the field with temperament traits measured in the laboratory at the beginning of the night period. An observer was only present to turn the camcorders on in complete darkness.

Chapter 3
50
The tests lasted for 10 min starting from 2 min after the recordings started (2 min acclimatization after turning on the camcorders). After the first six tanks were filmed, the positions of the cameras were switched to the last five tanks. After this disturbance a 30 min acclimatization period before the succeeding test was provided. Four temperament categories were examined: (1) boldness-‐shyness, an individual’s reaction to any risky situation, but not a new situation, (2) exploration-‐avoidance, an individual’s reaction towards a new situation; this includes behaviour towards a new habitat, new food or novel objects (in this paper: activity in a novel environment, interest in a novel food item), (3) activity, the general level of activity of an individual in a non-‐risky and non-‐novel environment and (4) aggressiveness, an individual’s antagonistic reaction towards conspecifics (for more details on these definitions see Réale et al. 2007).
Figure 1 Laboratory set-‐up of the 80-‐L experimental tanks for the temperament trait tests. Both tank sections were similar in the amount of structure they provided. A vertical white plastic plate was fixed 0.5 cm below the surface, water was allowed to flow between the sides and the same water was used for each test. The white plastic plate guaranteed that the fish, that was introduced into tank section A, did not experience the environment in tank section B. Removal of the plastic plate before the boldness test enabled the perception of visual and chemical cues of the predator on the opposite tank section.

Temperament traits and habitat use
51
The general procedure of the behavioural laboratory tests was based on the study of Dingemanse et al. (2007) and is summarized in Table 2. The temperament trait tests (3 October to 28 November 2008) started with the introduction of the bullhead into the unfamiliar 80-‐L laboratory tanks through a vertically positioned plastic tube (30 × 10 cm) that was removed directly before the test. Novel environment activity (exploration-‐avoidance category) was quantified by the number of squares crossed per minute (squares min-‐1) regardless of the direction or position in the tank and was measured twice (Table 2). The transfer from outdoors to the wet laboratory tanks included the slowly acclimation of the bullhead by adding water to the bucket.
Interest in a novel food item (exploration-‐avoidance category) was measured in a familiar environment (Table 2) by the bullhead’s reaction towards a small maggot of terrestrial Diptera, which we anticipated to be an unknown food item. The reaction was measured in terms of (1) the approach to the maggot
Day Action and temperament test
0 -‐ 2 -‐(Re)-‐capture of tagged and untagged bullhead -‐Transport to a 400-‐L outdoors tank at the university
3
-‐Renewal of water of the 80-‐L laboratory tank (9 am) -‐Translocation from outside tank to laboratory tanks -‐Novel environment activity (NEA) 1 in tank section A
4 -‐ 5 -‐Familiarization to tank section A
6 -‐Activity-‐test (ACT) 1 in tank section A -‐Aggressiveness-‐test: introduction of stimulus fish
7 -‐NEA 2 in tank section B
8 -‐Removal of visual barrier between tank sides (9 am) -‐ACT 2 in tank section B -‐Interest in novel food test in tank section B
9
-‐ACT 3 in tank section B -‐Boldness-‐test: predator (pike) into tank section A -‐End of tests; remove fishes to second outdoors tank
Table 2 Schedule of the temperament trait analysis of bullhead in the laboratory from 3 October to 28 November 2008 (N = 53 tagged, N = 21 untagged individuals). When two tests were conducted on a day, they followed each other immediately.

Chapter 3
52
(bullhead was situated in the square next to the square of the maggot), (2) the interest in the maggot (both in the same square) and (3) the ingestion of the maggot.
Activity was measured three times after allowing familiarization to the environment for at least 12 h (Table 2) by quantifying the distance moved (regardless of direction) with squares min-‐1 as variable.
Aggressiveness was tested in a familiar environment (Table 2) with the introduction of a smaller conspecific into the tank. Stimulus fish were distributed across the tanks by size (mean ± SD 46.8 ± 4.4 mm; range 35–62 mm) to guarantee a similar size ratio between the individuals assayed. Stimulus fish were introduced into the tanks through a vertically positioned plastic tube (30 × 10 cm) that was removed directly before the test. The 10 min test started with the removal of the tube (without 2 min acclimatization as the reaction to the stimulus was often very sudden). The number of bites was counted and the distance moved was recorded (regardless of direction; squares min-‐1). The individual’s distance moved was later compared to the distance moved directly before the test (see statistics).
Boldness-‐shyness was tested in a tank section B with the introduction of a pike (approximately 30 cm TL) into tank section A (Figure 1). Because the tank sections were only separated by a metal mesh (Table 2), visibility was provided and the odour of the predator diffused between the sections. We measured (1) the mean time (s), as well as the proportion of time (%) spent in the proximity (distance < 1 square) to structuring tank objects (Figure 1), (2) the proportion of time spent in the tank section half close to the pike (%) and (3) the distance moved in the presence of the predator (squares min-‐1). These variables were quantified directly before the test (control), as well as during the presence of the predator, and statistically compared to measure the reaction towards the predator (see statistics).
After the test series bullhead were brought to a second 400-‐L outdoor tank, where they were stored until their release into the stream (Table 2). The water of the laboratory tanks was renewed before the next test series.
Statistics of laboratory analyses and correlation to field data
Two temperament traits (aggressiveness and boldness) measured the difference between the distance moved from just before to during a stimulation test (see

Temperament traits and habitat use
53
laboratory analyses). Each individual tested represented its own control and only the change from ‘normal’ to stimulated behaviour was used as behavioural variable. The change in movement was defined by the individual residual value of the regression of all tested individuals’ stimulus activity [LN (squares min-‐1 + 1), y-‐axis] on the control activity [LN (squares min-‐1 + 1), x-‐axis; see also Garamszegi et al. 2009]. Negative values meant that the individual decreased movement during the stimulus test more than expected based on the reaction of all individuals tested. Similar calculations of the behavioural difference of control and stimulus were also made for the boldness variables refuge use (%), mean refuge use (s) and tank side positioning (%; see laboratory analyses of temperament traits).
For temperament traits that had more than one behavioural measure by definition (interest in novel food, aggressiveness and boldness), we merged several measures to one variable. By principal component analysis (PCA) values were z-‐transformed, Eigenvalues above 1 were extracted, multiple components rotated by varimax method and the new variables of the principal components were saved by regression method (Dingemanse et al. 2007). Due to the use of only two variables for aggressiveness, the variables were tested for correlation before running the PCA (Spearman, N = 18, rs = 0.379, p = 0.120).
Temperament traits, which had only one behavioural measure but were replicated (activity and novel environment activity), were represented by the mean value of the multiple measurements (squares min-‐1). Repeatability (r) of activity (three replicates) and novel environment activity (two replicates) were calculated by using the mean squares of among-‐groups and within-‐ groups variance of a one-‐way ANOVA with ‘individual’ as a factor on the dependent variables (Lessells & Boag 1987; see field statistics).
Temperament traits were correlated to habitat complexity by Pearson correlations (every trait instead of ‘interest in novel food’) and Spearman rank correlation (only interest in novel food; Kolmogorov–Smirnov, p < 0.05). Whether the temperament traits were correlated to body size (TL) and sex was tested by ANCOVAs (sex factor: variance homogeneity, Levene test, p > 0.05).
Significance of all calculations was assessed at α < 0.05. Significance of sets of multiple pair-‐wise comparisons was assessed at a false discovery rate (FDR) adjusted α (Benjamini & Hochberg 1995). Statistical analyses were conducted with SPSS 15.0.

Chapter 3
54
Results
Habitat use
The temporal occurrence of habitat types and their degree of complexity was relatively stable. We did not find any significant difference between stream quadrats (approximately 1 m2, N = 972) from June 2008 and October 2008 (N = 96) for any of the parameters considered (Table 1). From the originally tagged bullhead, we were able to recapture 20 individuals, which stayed over the whole study period in the habitat-‐determined stretch. These 20 individuals had a mean ± SD TL of 58.9 ± 6.0 mm; the gender of 19 individuals could be determined (7 males and 12 females). The tracking sessions and the individual recapture resulted in a mean ±SD of 10.45 ± 1.50 habitat localizations per individual (N = 20).
Bullhead (N = 20) showed a non-‐random selection between habitat types (likelihood 2 χ2 = 529.02, df = 100, p < 0.0001) and habitat complexity (likelihood 2 χ2 = 451.92, df = 40, p < 0.0001). Fish significantly preferred stream patches with natural branch jams (Figure 2a); open water was significantly avoided. The result is accompanied by a significant preference for stream patches with high habitat complexity and the significant avoidance of patches, which provided < 33% structure (Figure 2b). Bullhead showed no significant habitat choice for patches with intermediate complexity (≥ 33% ≤ 66% structure). Individual habitat complexity use was consistent, indicated by a high repeatability value of r = 0.568 (ANOVA, mean square, between groups = 4.264, within group = 0.289; see also Figure 3). The individual preferences of habitat complexity could not be explained (ANCOVA, N = 20, corrected model, sum of squares = 0.494, df = 2, F = 0.624, p = 0.549) by body size (TL, ss = 0.194, df = 1, F = 0.490, p = 0.494) or sex (ss = 0.163, df = 1, F = 0.413, p = 0.530).
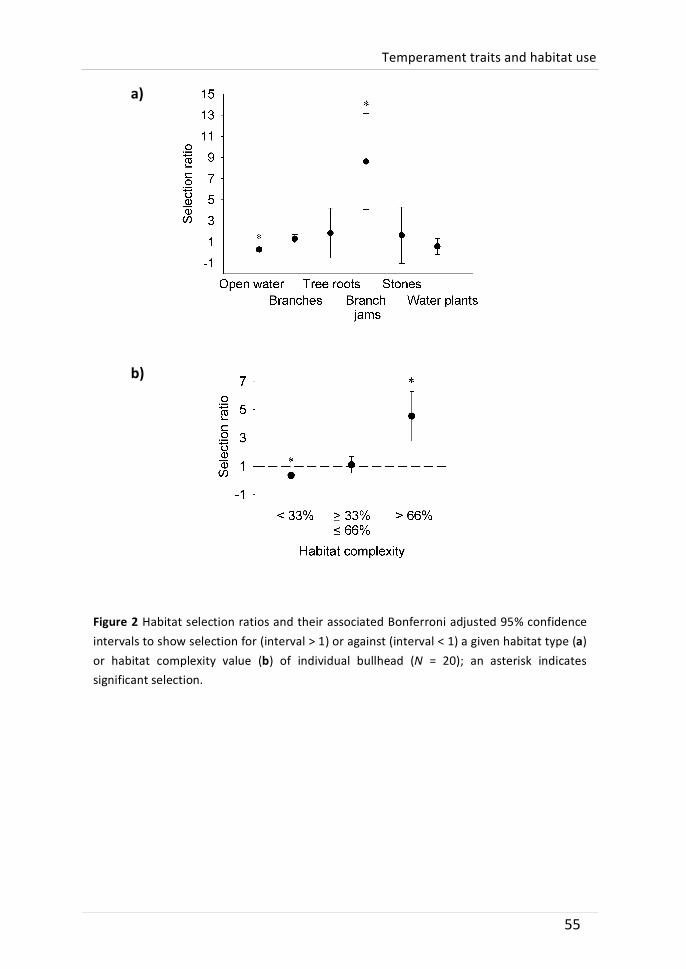
Temperament traits and habitat use
55
a)
b)
Figure 2 Habitat selection ratios and their associated Bonferroni adjusted 95% confidence intervals to show selection for (interval > 1) or against (interval < 1) a given habitat type (a) or habitat complexity value (b) of individual bullhead (N = 20); an asterisk indicates significant selection.

Chapter 3
56
Temperament traits and the relation to habitat use
Of the 20 field-‐observed and recaptured individuals, 18 were successfully tested in the laboratory for five temperament traits. The tests were behaviourally consistent (N = 18 individuals) during the laboratory test period with r = 0.439 for activity (ANOVA, ms, between groups = 35.645, within group = 10.649) and r = 0.422 for novel environment activity (ANOVA, ms, between groups = 63.741, within group = 25.937). Interest in novel food was represented by a principal component (PC1) in which the approach, the interest and the ingestion of the maggot were positively correlated (Table 3). In the aggressiveness PC1 the individual’s activity change and the number of bites were positively correlated, suggesting that fish, which bite more often became also more active when the stimulus fish was introduced. A positive value of boldness PC1 meant that after the introduction of a predator (pike) individuals spent relatively more time close to a refuge (%), had on average longer stays at a refuge (s), spent relatively less time in the tank half close to the predator (%) and were less active (squares min-‐1) compared to the control test (Table 3). Boldness PC2 was mainly dominated by the activity change from control to predator test.
Of the five temperament traits tested only aggressiveness was significantly related to habitat use (Table 4). Aggressiveness was strongly negatively correlated to habitat complexity (Pearson, N = 18, rp = −0.527, p = 0.025; Figure 3): individuals who were less aggressive towards the stimulus fish under laboratory conditions were more often found in complex habitat patches under field conditions. This correlation was, however, not significant after α adjustment for multiple comparisons (FDR αadjusted = 0.0083). After exclusion of two exceptional individuals who were almost exclusively found in open water (Figure 3), the negative correlation between aggressiveness and habitat complexity became even stronger and also significant (Pearson, N = 16, rp = −0.921, p < 0.001). Aggressiveness was not significantly related (ANCOVA, N = 18; corrected model, ss = 1.255, df = 2, F = 0.598, p = 0.563) to body size (TL, ss = 0.892, df = 1, F = 0.850, p = 0.371) or sex (ss = 0.835, df = 1, F = 0.795, p = 0.387).
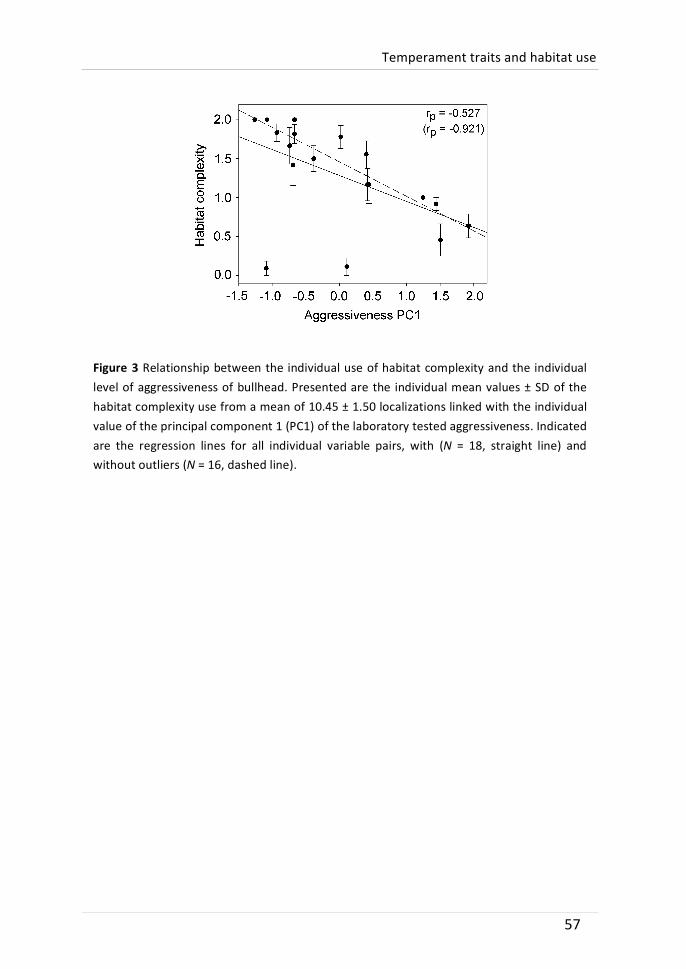
Temperament traits and habitat use
57
Figure 3 Relationship between the individual use of habitat complexity and the individual level of aggressiveness of bullhead. Presented are the individual mean values ± SD of the habitat complexity use from a mean of 10.45 ± 1.50 localizations linked with the individual value of the principal component 1 (PC1) of the laboratory tested aggressiveness. Indicated are the regression lines for all individual variable pairs, with (N = 18, straight line) and without outliers (N = 16, dashed line).
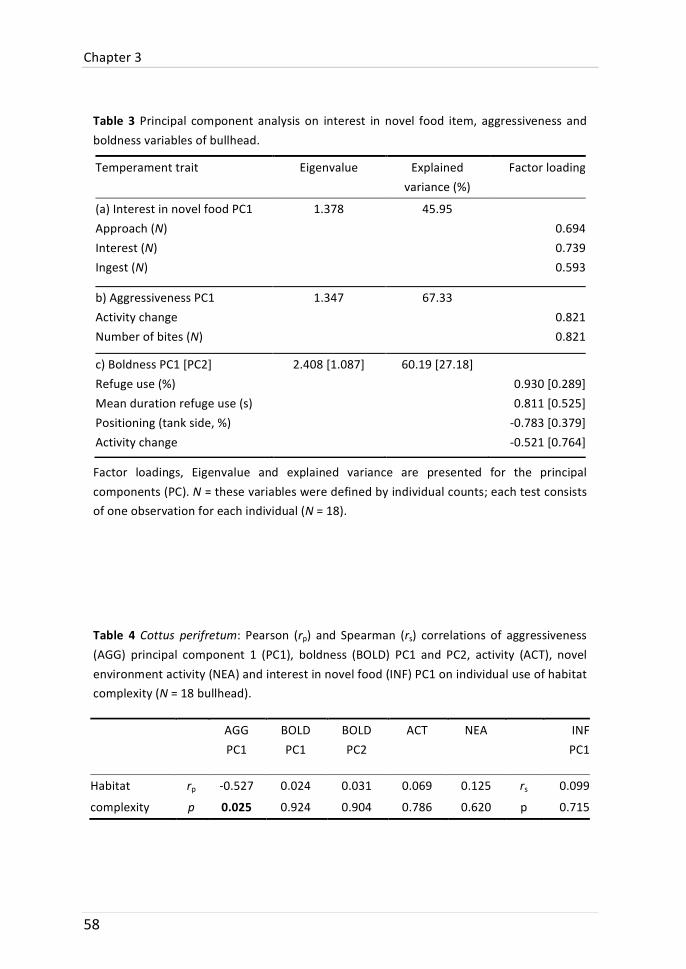
Chapter 3
58
Temperament trait Eigenvalue Explained variance (%)
Factor loading
(a) Interest in novel food PC1 Approach (N) Interest (N) Ingest (N)
1.378
45.95
0.694 0.739 0.593
b) Aggressiveness PC1 Activity change Number of bites (N)
1.347
67.33
0.821 0.821
c) Boldness PC1 [PC2] Refuge use (%) Mean duration refuge use (s) Positioning (tank side, %) Activity change
2.408 [1.087]
60.19 [27.18]
0.930 [0.289] 0.811 [0.525] -‐0.783 [0.379] -‐0.521 [0.764]
AGG PC1
BOLD PC1
BOLD PC2
ACT
NEA
INF PC1
Habitat rp -‐0.527 0.024 0.031 0.069 0.125 rs 0.099
complexity p 0.025 0.924 0.904 0.786 0.620 p 0.715
Table 3 Principal component analysis on interest in novel food item, aggressiveness and boldness variables of bullhead.
Factor loadings, Eigenvalue and explained variance are presented for the principal components (PC). N = these variables were defined by individual counts; each test consists of one observation for each individual (N = 18).
Table 4 Cottus perifretum: Pearson (rp) and Spearman (rs) correlations of aggressiveness (AGG) principal component 1 (PC1), boldness (BOLD) PC1 and PC2, activity (ACT), novel environment activity (NEA) and interest in novel food (INF) PC1 on individual use of habitat complexity (N = 18 bullhead).

Temperament traits and habitat use
59
Discussion
This is, to the best of our knowledge, the first study demonstrating a link between field-‐observed habitat use of various complexity levels and individual aggressiveness measured under standardized laboratory conditions. Bullhead that were found more often in highly structured habitats were less aggressive towards a smaller conspecific. Such a pattern has previously been shown in laboratory-‐based no-‐choice habitat structure studies for finfish (Danio rerio, Basquill & Grant 1998; Carfagnini et al. 2009; Salmo trutta, Sundbaum & Näslund, 1998) and crayfish (Orconectes propinquus, Corkum & Cronin 2004; Cherax destructor, Baird et al. 2006). Here the habitat structure–aggressiveness relation at the individual level was confirmed in a habitat choice survey under natural conditions.
We speculate that the defense of a territory was more difficult in less structured habitats. This hypothesis is supported by a study in house mouse (Mus domesticus) where aggressiveness per encounter of a conspecific tended to be higher in a territory located in an open habitat than in a habitat with a complex structure (Jensen et al. 2005). Furthermore, Baird et al. (2006) showed for a crayfish that the number of and the time spent in agonistic interactions is higher in less complex habitats. Therefore, bullhead occupying less structured habitats should be more aggressive. Our study design can, however, not resolve the causal pathway of the negative relation between habitat structure and aggressiveness. Do bullhead become more aggressive when defending their territory in less structured habitats or do more aggressive individuals choose to live in less structured habitats to benefit from underused resources (compare Svanbäck et al. 2008; Kobler et al. 2009a)?
Regardless of the causal pathway, we assume that the level of aggressiveness in the laboratory may still have been related to previous habitat use on site. This indicates that the habitat structure-‐aggressiveness relation of this study may not be phenotypically plastic in the short-‐term. Each individual was held for a period of approximately one week under habitat-‐standardized conditions in the laboratory tanks.
The negative relation between habitat complexity use and aggressiveness contrasts with the hypothesis that more aggressive stream fish have higher competitive abilities and, therefore, the ability to exclude less aggressive individuals from preferred habitats (cf., Fausch 1984; Weber & Fausch 2003). Because bullhead prefer highly complex habitats (Greenberg & Holtzman 1987;

Chapter 3
60
Davey et al. 2005; Legalle et al. 2005), similar to this study, complex habitats should be occupied by more aggressive individuals. We speculate that competition had a minor effect on the choice of habitat structure. The focal stretch of the Laarse Beek is situated in a EU-‐habitat zone, has not been artificially manipulated, has meandering parts and contains plenty of woody debris from a dense beech-‐oak forest surrounding the study area. It is plausible that structured habitats were not limiting and, hence, the influence of competition on microhabitat use was relatively low. This would explain why more aggressive individuals were found in less structured habitats, similarly to the laboratory studies mentioned earlier on that were conducted in the absence of competition for structure.
When two outlier fish were excluded from the aggressiveness-‐habitat use calculation, the correlation became almost 100%. The constant daytime open water use of these two individuals is rather uncommon, which was shown by the significant avoidance found in this study (see also references in the introduction). Further, it is surprising that these two individuals had rather low levels of aggressiveness, based on what should be expected from the behaviour of the population (Figure 3). In line with the previous discussion of higher aggression for the defense of less structured territories (see before) it can be speculated that these two individuals were less territorial and expressed a spatial behaviour similar to juveniles that use less stable, depositional habitats (see Cottus bairdii; Petty & Grossman 2007).
Against our hypothesis, we did not find a negative link between habitat complexity use in the field and the laboratory tested activity. The significant and repeatable between-‐individual differences of bullhead activity in the laboratory (Kobler et al., data not shown) are, thus, not related to the observed habitat use differences in the field. It may be that the habitat-‐uniform holding of bullhead in the outside tank and in the aquaria during approximately a one-‐week period may have led to a uniformity of activity. This is, however, rather surprising as Salvanes & Braithwaite (2005) observed that juvenile cod (Gadus morhua) reared in different habitat structures showed activity differences even after keeping them together for eight days under similar conditions. The latter study was conducted with cod reared from the larval to juvenile stage under either structured or non-‐structured tank conditions (Salvanes & Braithwaite 2005). Large variance in habitat use of early juvenile phases of bullhead has not been reported previously (Van Liefferinge et al. 2005). It seems that a possible habitat-‐activity adaption of bullhead is either only of short term and, thus, could not be detected with our experimental protocol, or did not exist in the studied population due to

Temperament traits and habitat use
61
homogeneous habitat use in the juvenile phase. However, our dataset only includes daytime shelter habitat use and does not cover possible habitat switches during the night when sculpins tend to be more variable in habitat choice (Greenberg & Holtzman 1987; Natsumeda 2007). Future studies should include diel changes in habitat use and examine how this affects inter-‐ individual differences in activity levels (see Kobler et al. 2009a).
We assume behavioural consistency during the aggressiveness test since we repeated only two of the five temperament tests. There was, however, a repeatability of r = 0.439 for activity and of r = 0.422 for novel environment activity. These values indicate an average degree of individual consistency (Bell 2009; Kobler et al. 2009b). By comparing repeatability between temperament traits, Bell et al. (2009) showed, in a meta-‐analysis, that aggressiveness is more repeatable than activity. We speculate that the individual responses of bullhead to the stimulus fish are repeatable in time. Moreover, Kobler et al. (2009b) have shown that a temperament trait, the exploration rate, observed in the laboratory reflects this behaviour in the field. The probable repeatability and the transferability of laboratory observation to field behaviour confirm the significance of the aggressiveness-‐habitat correlation of this study.
The present study used 20 out of 334 originally PIT-‐tagged individuals that were recaptured in the stream section in which they were initially captured. In an accompanying study, Kobler et al. (data not shown) detected large movement ranges in the same bullhead population (up to 1284 m). As a result, the stream stretch of the present study contained only a subset of the originally tagged individuals and a biased sample towards more sedentary individuals. However, aggressiveness, boldness, interest in novel food and basic activity were not related to the movement range values (N = 53 field-‐ and laboratory-‐observed individuals; Kobler et al., data not shown). Thus, we do not expect any bias of these temperament traits in the current bullhead sample.
Acknowledgments
We thank G. Eens, P. Scheys, K. Geudens, D. Geelhand and S. Vanwetswinkel for technical assistance and G. Knaepkens for advice. Research was funded by FWO-‐Flanders and the University of Antwerp. G.E.M. is a post-‐doctoral researcher funded by the FWO-‐Flanders. This study was conducted under permits for capture, manipulation and telemetry issued by Flemish authorities.


CHAPTER 4
DIEL MOVEMENT OF BULLHEAD


Diel movement of bullhead
65
Diel movement of bullhead (Cottus perifretum) in a lowland stream
Alexander Kobler, Yves Humblet, Guy Knaepkens, Brecht Engelen, Marcel Eens
Ecology of Freshwater Fish (2012) 21: 453-‐460
Abstract
Passive integrated transponder tags have been successfully applied in Cottus spp. and have enabled researchers to gather more information about the movement patterns of individual fish in the wild. In two succeeding years during springtime, a portable antenna was used to determine diel movements of bullhead (Cottus perifretum). In 2007, bullhead (N = 26) moved significantly farther distances at night (mean, 0.42 m h-‐1) and dawn (mean, 0.35 m h-‐1) than during daytime (mean, 0.11 m h-‐1; mixed model, p < 0.001, respectively; p = 0.001), which may be due to foraging activities for Gammarus spp. Irrespective of diel period, smaller fish covered significantly longer distances (p = 0.001). In 2008, similar diel movement patterns were observed, but the differentiation between daytime (mean, 0.12 m h-‐1) and night periods (mean, 0.18 m h-‐1) was not significant (mixed model, p = 0.087; N = 49 bullhead). It is discussed that longer tracking intervals used in 2008 (three times per 24-‐h instead of every two hours) were not suitable to detect the sheer magnitude of distances covered during a diel period. It is shown that this may be due to ‘site fidelity’ of some individuals: after swimming several metres at night, they returned to the exact location they previously occupied during daytime. In 2008, sex and body size were not related to diel movement. The present study is the first to present a quantitative differentiation between diel distances covered in a Cottus spp.

Chapter 4
66
Introduction
Knowledge about the behaviour of small stream fish species, particularly small benthic species, is limited (Lucas & Baras 2001; Petty & Grossman 2004). This is not only due to the lack of commercial interest (Lucas & Baras 2001), but also due to the difficulties in tracking individual fish of small body size in lotic environments. Whereas several recent studies focused on the long-‐term movement of small stream fishes (e.g. ide, Leuciscus idus, Winter & Fredrich 2003; gilt darters, Percina evides: Skyfield & Grossman 2008; creek chubs, Semotilus atromaculatus: Belica & Rahel 2008), there are only few studies that investigated diel differences in distances covered (e.g. brown trout, Salmo trutta: Bunnell et al. 1998; Young 1999; cutthroat trout, Oncoryhnchus clarki pleuriticus: Young et al. 1997).
Diel movements are characteristic for many freshwater stream fishes, particularly for juveniles or small-‐bodied species (Lucas & Baras 2001). In lotic environments, diel movements can be longitudinal and occur between distinct daytime and night-‐time sites (Lucas & Baras 2001). For instance, it has been observed that common dace (Leuciscus leuciscus, Clough & Ladle 1997) and brown trout (Clapp et al. 1990; Young 1999) have specific diurnal home sites, which they leave during dusk and to which they return during dawn. Possible behaviours that influence these movement patterns include anti-‐predator behaviour and foraging activity (Clough & Ladle 1997). Neither stream site, on its own, meets all requirements (shelter, temperature, oxygen and food availability) for a fish species during an entire diel cycle (Clough & Ladle 1997). Therefore, fish often move between different sites during their active and inactive phases (Helfman et al. 1997).
The stream fish species of the present study, the bullhead (Cottus perifretum), is a small benthic fish species living in the demersal zones of streams and rivers where they express a strong preference for highly structured habitat types (Kobler et al. 2011). Bullhead belong to the freshwater sculpins [genus Cottus, species rarely exceeding 15 cm in total length (TL)] that are found in small streams, large rivers or lakes of the northern hemisphere (Nelsen 1994). The extremely reduced swim-‐bladder (Freyhof et al. 2005) and the dorso-‐ventrally flattened tapering body (Tomlinson & Perrow 2003) of bullhead is an adaptation to lotic environments in which they swim with a hopping motion over substrates by using their large pectoral fins. While several recent long-‐term studies have focused on dispersal distances of freshwater sculpins (Breen et al. 2009; Hudy & Shiflet 2009; Ovidio et al. 2009), the diel movement behaviour has not yet been

Diel movement of bullhead
67
quantified. It is well-‐known that freshwater sculpins are mainly active at night (e.g. Smyly 1957; Andreasson 1969, 1973; Brandt 1986; Natsumeda 1998) and can forage in total darkness (Hoekstra & Janssen 1985) but relevant quantitative information is only available from aquarium observations (Andreasson 1969; but see Greenberg & Holtzman 1987; Natsumeda 2007 for quantified home range field data).
Because of the larger nocturnal home range of the banded sculpin (Cottus carolinae, Greenberg & Holtzman 1987) and the Japanese fluvial sculpin (Cottus pollux, Natsumeda 2007) and the nocturnal activity of bullhead (e.g. Smyly 1957; Andreasson 1969) we hypothesized that bullhead cover longer distances at night. Further, foraging success may also be higher during night as one of the common food resources, freshwater shrimps (amphipods, Gammarus spp.) are nocturnally active (e.g. Holomuzki & Hoyle 1990). We further expected that bullhead swim only very short distances during daytime as bullhead seek shelter in structured habitats during daylight (Davey et al. 2005). Similar to diurnal home site fidelity of common dace and brown trout (Clapp et al. 1990; Young 1999; Clough & Ladle 1997), it is possible that bullhead show site-‐fidelity to a daytime shelter to which they return after nocturnal movements.
The present study examined the distances that bullhead cover during diel periods in a small lowland stream in Flanders (Belgium) by using passive integrated transponder (PIT) tag telemetry. PIT tags have been successfully applied in freshwater sculpins [e.g. C. perifretum (formerly determined as Cottus gobio), Knaepkens et al. 2007; Cottus bairdii, Ruetz et al. 2006; Cottus cognatus, Keeler 2006, Keeler et al. 2007] enabling researchers to locate them with less than 30 cm precision (Cucherousset et al. 2005) in even very turbid water and/or at night. It has been shown that PIT tag insertion has no significant influence on survival, growth and swimming capacity (Ruetz et al 2006; Knaepkens et al 2007). The tracking was conducted in two succeeding years with two different tracking protocols: tracking every two hours and the assignment of movement to all diel periods (twelve times tracking during 24-‐h, 2007) and only three times tracking during 24-‐h (synchronized with sunrise and sunset) and assignment of movement to daytime and night-‐time (twilights and night) period (2008). It was compared whether the two protocols (with different labour-‐intensity) would result in a similar difference between diurnal and nocturnal movements. In addition to the comparison of the protocols, the potential relations between sex, body size and movement distances were also assessed.

Chapter 4
68
Materials and Methods
Study area and habitat determination
The study was conducted in the Laarse Beek (51°16'28.40"N; 4°29'4.63"E), a lowland stream in northern Flanders, Belgium. The focal stretches of the stream have an average width of ∼3 m and a depth of ∼0.3 m. The stream crosses a beech-‐oak forest with black alder (Alnus glutinosa) rooting partly in the stream. The stream bed is characterized by a sandy soil with woody debris patches. Bullhead is the most abundant fish species in the stream with a density of approximately 2.1 adults per m2 (Kobler et al. 2011). Aquatic predators such as northern pike (Esox lucius), European eel (Anguilla anguilla) and European perch (Perca fluviatilis) were rare (Kobler et al. 2011). Similarly, the predation risk through avian predators such as grey heron (Ardea cinerea) and kingfisher (Alcedo atthis) was probably low, as they were rarely observed.
Capture and tagging
The focal stretch of the stream was marked every 2 m with consecutively numbered wooden poles along the right stream bank. Adult bullhead (> 50 mm total length, TL) were caught by electro-‐fishing (WFC7-‐10, Electracatch Int., Wolverhampton, UK). Bullhead were then anaesthetized (0.25 ml L-‐1 2-‐phenoxyethanol) and individually coded PIT tags (12 x 2.1 mm, 0.094 g; ID100, EID Aalten, NL) were inserted horizontally into the body captivity, through a 2 mm incision, close to the genital papillae (by using a scalpel). Due to the small size of the fish and the opening, the incision was not stitched (Bruyndoncx et al. 2002). After implantation, bullheads were allowed to recover in buckets with fresh stream water and released at their capture position.
The electro-‐fishing and tagging for the 2007 tracking took place in a 250 m stream stretch of the Laarse Beek on 13 November 2006. The 66 PIT-‐tagged individuals had a mean TL (± SD) of 63.8 ± 7.0 mm and mean wet weight of 3.3 ± 1.4 g. In 2008, we electro-‐fished from 4 to 14 February. This period was chosen because of the possibility of sex-‐determination due to the start of the annual gonadal maturation (see also further). A 324 m stream stretch that overlapped large parts of the electro-‐fishing stretch from 2006 was fished twice and more intensively than in 2006 (to receive a good abundance estimate for another study, Kobler et al. 2011). 334 PIT-‐tagged individuals had a mean TL of 62.1 ± 6.3 mm and mean weight of 3.3 ± 1.2 g. In 2008, every tagged individual was photographed.

Diel movement of bullhead
69
Sex determination in 2008
Because an external sex determination of bullhead is not reliable outside the reproductive period (J. Freyhof, personal communication), bullhead caught in November 2006 were not sexed. For 2008, the gender of bullhead was determined retrospectively by evaluating the pictures of the individuals. The gender could be determined by morphological characteristics. Individuals with a very round belly (looking tadpole-‐alike), that clearly suggested the content of eggs, were determined as females. Males could be determined by the triangular shape of the head (Freyhof et al. 2005) and the proportionally smaller body cavity. The sex of every individual was determined three times on three different days, with the researcher being blind to the identity of the individual. Ambiguity about the sex of an individual led to a non-‐defined gender of the individual. The majority of individuals (83%) were sexed including 138 females and 139 males.
Tracking
Tracking was conducted manually by carefully walking upstream in the stream centre (Kobler et al. 2011) using a portable PIT antenna (Trovan components, EID Aalten, NL, custom made see Cucherousset et al. 2005). Once a bullhead was located, the time and stream position were recorded. While recording, the antenna was kept above the bullhead and an escape response following localization was never observed (Kobler et al. 2011).
In 2007, 24-‐h tracking sessions were conducted on 13 March and on 14 May. This period was chosen as bullhead activity is highest during reproductive period (Kobler A., Humblet Y., Geudens K., Eens M., in preparation) and the night is long enough (in May in Antwerp approx. 5.5 h) for a good assignment of movements to the night period. The stream stretch was scanned every two hours (10 a.m. to 10 a.m.) to estimate real distances covered (Natsumeda 2007) and good assignment to diel periods (Kobler et al. 2008). The 2-‐h interval allowed precise screening of a stream stretch of 165 m, centrally located in the electro-‐fishing stretch. Therefore, the observed individuals were only a subsample of the originally tagged bullhead. One researcher scanned the stream while another researcher recorded notes at the streamside. The locations of detected bullhead were recorded using stream length (e.g. 125 m) and stream width (left, left-‐middle, middle, right-‐middle and right) information. Each tracking day resulted in a maximum of twelve localizations per individual.

Chapter 4
70
In 2008, the 24-‐h tracking sessions were conducted on 14 March, 11 April and 9 May (one tracking session more than in 2007 due to a less labour-‐intensive tracking protocol; see further). The tracking stretch was 110 m long and centrally located in the electro-‐fishing stretch. This stretch length and screening duration (∼80 minutes for this stretch) was a prerequisite as the twilight lasted only between 72 and 93 minutes and we aimed to locate every individual during the dawn period. The start of each tracking interval was adapted to the time of sunset and sunrise. The first interval started 1.5 h before sunset (daytime), the second 1.5 h before sunrise (dawn), and the third interval again 1.5 h before sunset. This tracking protocol enabled the assignment of the movement to night (night and both twilights: after sunset to before sunrise; from first to second tracking) and daytime period (sunrise to sunset; from second to third tracking). This was done to test whether this protocol (only three times tracking per 24-‐h: less labour intensive and less disturbance in the stream) would reveal a similar difference between diel periods than in 2007 (twelve times tracking per 24-‐h). As a further change compared to 2007, the stream width information was reduced to “left, middle and right” as this differentiation allowed a more distinct and faster assignment to a location than in the previous year.
Statistics
Movement was represented by the variable “minimum displacement” (m h-‐1), the distance that an individual at the minimum moved between consecutive tracking locations divided through the time (minutes) between the locations (Rogers & White 2007). This standardized measure accounted for the different time intervals between consecutive tracking locations.
In 2007, minimum displacements were assigned to daytime, dusk, night and dawn. The beginning and end of each diel period were calculated with nautical definition by using http://www.cgi.stadtklima-‐stuttgart.de/mirror/sonne.exe (with Düsseldorf, Germany as reference city; Kobler et al. 2008). Movement values were excluded when a missing individual location resulted in a minimum displacement that could not be assigned to a single diel period. For instance, a missing dawn location would result in a minimum displacement from night location to daytime location and it would be unclear when the distance was covered, during dawn or daytime. However, if a fish was not localized during a twilight period (twilight was shorter than the tracking interval) but shortly after that period (e.g. some minutes afterwards) the delayed location was used to estimate twilight movement (Kobler et al. 2008). In general, individuals with less than nine out of twelve tracking localizations per 24-‐h were excluded from the

Diel movement of bullhead
71
tracking session. In total 394 movements from 26 bullhead were analysed. In 2008, individuals were excluded from a tracking session if one out of three possible locations was missing. This resulted in 1 daytime and 1 night period (night and twilight) movement per individual per tracking session (see before); 148 movements from 49 individuals.
The relationship between minimum displacement (dependent variable), diel period, body size (TL), sex (only available for 2008) and diel period × sex was calculated by a linear mixed model (one for each year). The “diagonal” repeated covariance type was used. Diel period and sex (the factors) were treated as fixed effects. Because the tagged individuals were a defined selection of the population (only individuals > 50 mm TL were studied) body size was also treated as a fixed effect. The individuals were treated as random subjects. The significance of the differences between factors with more than two levels (diel period in 2007) was compared by the mixed model using Bonferroni confidence interval adjustment. Variances of minimum displacement between the factor levels (diel period and sex) were heterogeneous (Levene-‐test, p > 0.05) and minimum displacement was, therefore, transformed [Ln (x + 1)]. Insignificant variables were step-‐wise excluded from the mixed models. Significance of all calculations was assessed at α < 0.05 and conducted with SPSS 15.0 (SPSS Inc., Chicago, IL, USA).
Results
In 2007, the distances covered (minimum displacements, m h-‐1) differed significantly between diel periods (Table 1). The minimum displacements during night (Figure 1, range 0 to 5.4 m h-‐1) were significantly higher than those during daytime (p < 0.001, daytime: 0 to 5.6 m h-‐1) and dusk (p = 0.005, dusk: 0 to 6.5 m h-‐1;). Minimum displacements during dawn (0 to 3.8 m h-‐1) were also significantly higher than those during daytime (p = 0.001; Figure 1). Irrespective of diel period, smaller fish covered significantly longer distances (Table 1).
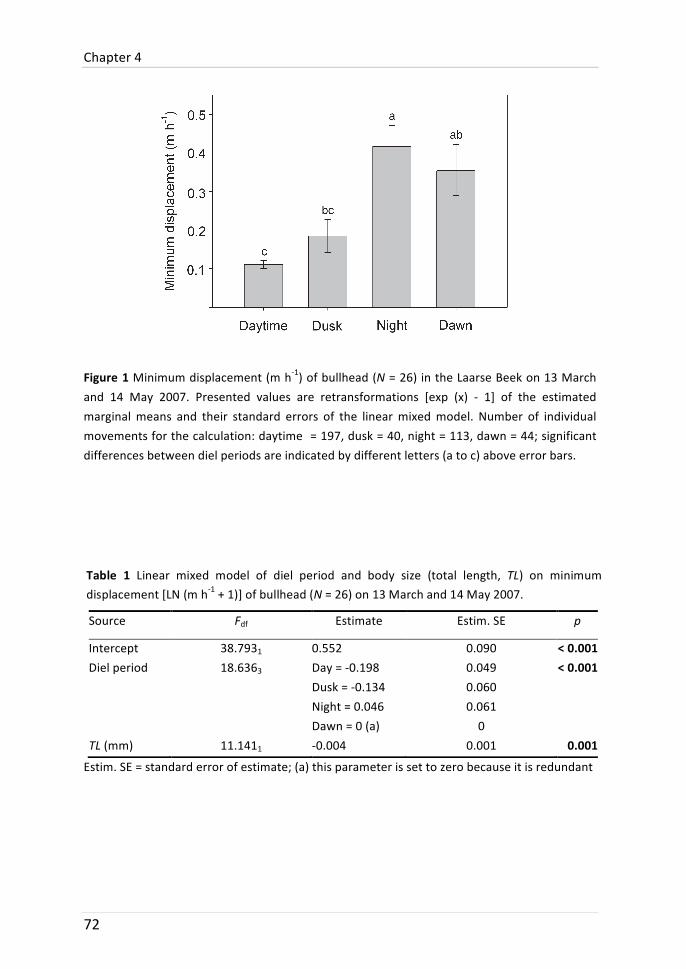
Chapter 4
72
Figure 1 Minimum displacement (m h-‐1) of bullhead (N = 26) in the Laarse Beek on 13 March and 14 May 2007. Presented values are retransformations [exp (x) -‐ 1] of the estimated marginal means and their standard errors of the linear mixed model. Number of individual movements for the calculation: daytime = 197, dusk = 40, night = 113, dawn = 44; significant differences between diel periods are indicated by different letters (a to c) above error bars.
Estim. SE = standard error of estimate; (a) this parameter is set to zero because it is redundant
Source Fdf Estimate Estim. SE p
Intercept 38.7931 0.552 0.090 < 0.001 Diel period 18.6363 Day = -‐0.198
Dusk = -‐0.134 Night = 0.046 Dawn = 0 (a)
0.049 0.060 0.061 0
< 0.001
TL (mm) 11.1411 -‐0.004 0.001 0.001
Table 1 Linear mixed model of diel period and body size (total length, TL) on minimum displacement [LN (m h-‐1 + 1)] of bullhead (N = 26) on 13 March and 14 May 2007.

Diel movement of bullhead
73
Forty-‐one 24-‐h cycles from 26 individuals were recorded during 13 March and 14 May 2007. Twelve 24-‐h cycles from eleven individuals (29.3% of all 24-‐h cycles in 2007) were characterized by activity during twilight and night while being stationary during daytime. These individuals left their diurnal location during dusk, swam some meters during night and dawn and were back at their previous diurnal location on the following daytime localization (Figure 2). Of these eleven “homing” individuals, eight were recorded twice but only one individual showed a similar diel habitat shift pattern with fidelity to a diurnal location during the second tracking session. Further, only three of the eleven homing individuals were found at the same diurnal location (within 1 m2) during the second tracking session. Other 24-‐h cycles were either characterized by a completely stationary behaviour (recorded within 1 m2; 13 cycles, 31.7%), upstream movement (1 cycle, 2.4%), downstream movement (7 cycles, 17.1%) or a more random direction of diel movements (8 cycles, 19.5%).
Similarly to the previous year, in 2008 the mean minimum displacement was higher during night period (night and twilight combined; mean of individual means ± SD, N = 49; 0.18 ± 0.19 m h-‐1; range 0 to 0.9 m h-‐1) than during daytime (0.12 ± 0.10 m h-‐1; range 0 to 0.6 m h-‐1). However, even after stepwise exclusion of the insignificant variables the difference between diel periods did not reach significance (p = 0.087, Table 2). The result of the linear mixed model (intercept, p = 0.931) that included all independent variables was: diel period (p = 0.137), body size (p = 0.348, estimate 0.002), sex (p = 0.635) and sex × diel period (p = 0.445).
Of 76 24-‐h cycles from 49 individuals observed from 14 March to 9 May 2008, 13 24-‐h cycles from 13 individuals (17.1% of all 24-‐h cycles in 2008) were characterized by a diel location shift with fidelity to a diurnal location, similar to what was observed in 2007 (Figure 3). None of the 13 homing individuals repeated this behaviour during a second tracking session (ten of these 13 individuals were recorded twice, one was recorded three times) nor were they found at exactly the same diurnal location at the second tracking session (in most cases they shifted the diurnal location by one or two metres). The other 63 24-‐h cycles were characterized by stationary behaviour (14 cycles, 18.4%), upstream movement (13 cycles, 17.1%), downstream movement (17 cycles, 22.4%) or a more random movement direction (19 cycles, 25%).
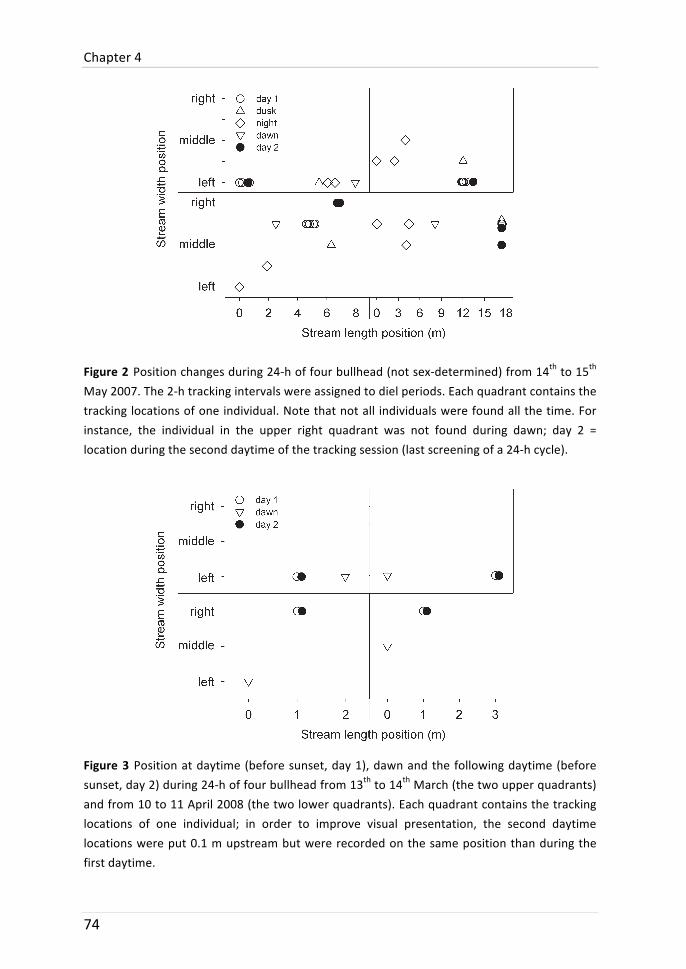
Chapter 4
74
Figure 2 Position changes during 24-‐h of four bullhead (not sex-‐determined) from 14th to 15th May 2007. The 2-‐h tracking intervals were assigned to diel periods. Each quadrant contains the tracking locations of one individual. Note that not all individuals were found all the time. For instance, the individual in the upper right quadrant was not found during dawn; day 2 = location during the second daytime of the tracking session (last screening of a 24-‐h cycle).
Figure 3 Position at daytime (before sunset, day 1), dawn and the following daytime (before sunset, day 2) during 24-‐h of four bullhead from 13th to 14th March (the two upper quadrants) and from 10 to 11 April 2008 (the two lower quadrants). Each quadrant contains the tracking locations of one individual; in order to improve visual presentation, the second daytime locations were put 0.1 m upstream but were recorded on the same position than during the first daytime.
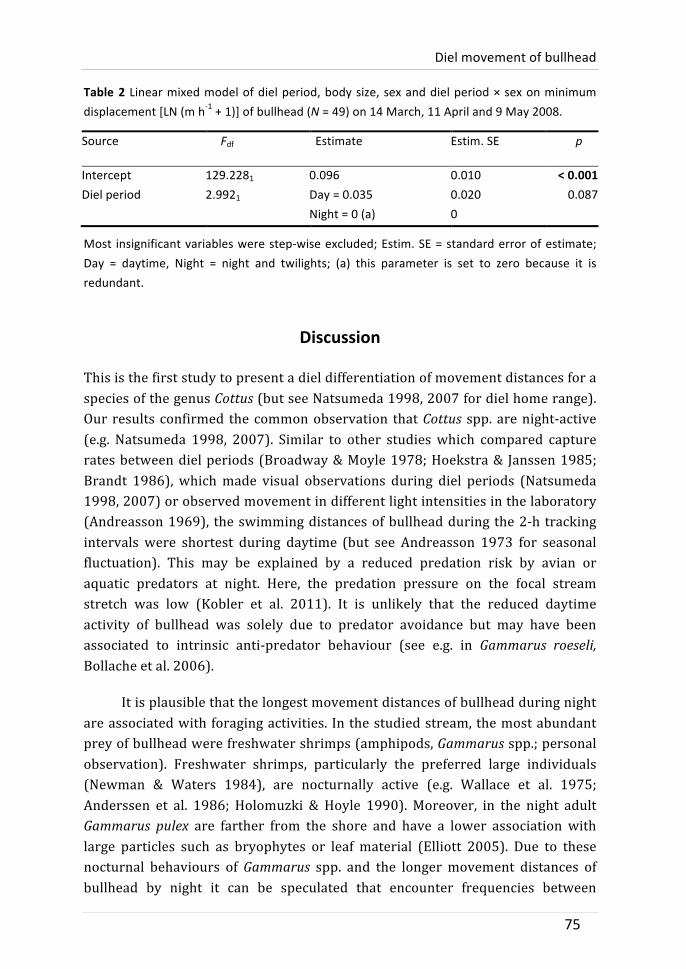
Diel movement of bullhead
75
Table 2 Linear mixed model of diel period, body size, sex and diel period × sex on minimum displacement [LN (m h-‐1 + 1)] of bullhead (N = 49) on 14 March, 11 April and 9 May 2008.
Source Fdf Estimate Estim. SE p
Intercept 129.2281 0.096 0.010 < 0.001 Diel period 2.9921 Day = 0.035
Night = 0 (a) 0.020 0
0.087
Most insignificant variables were step-‐wise excluded; Estim. SE = standard error of estimate; Day = daytime, Night = night and twilights; (a) this parameter is set to zero because it is redundant.
Discussion
This is the first study to present a diel differentiation of movement distances for a species of the genus Cottus (but see Natsumeda 1998, 2007 for diel home range). Our results confirmed the common observation that Cottus spp. are night-‐active (e.g. Natsumeda 1998, 2007). Similar to other studies which compared capture rates between diel periods (Broadway & Moyle 1978; Hoekstra & Janssen 1985; Brandt 1986), which made visual observations during diel periods (Natsumeda 1998, 2007) or observed movement in different light intensities in the laboratory (Andreasson 1969), the swimming distances of bullhead during the 2-‐h tracking intervals were shortest during daytime (but see Andreasson 1973 for seasonal fluctuation). This may be explained by a reduced predation risk by avian or aquatic predators at night. Here, the predation pressure on the focal stream stretch was low (Kobler et al. 2011). It is unlikely that the reduced daytime activity of bullhead was solely due to predator avoidance but may have been associated to intrinsic anti-‐predator behaviour (see e.g. in Gammarus roeseli, Bollache et al. 2006).
It is plausible that the longest movement distances of bullhead during night are associated with foraging activities. In the studied stream, the most abundant prey of bullhead were freshwater shrimps (amphipods, Gammarus spp.; personal observation). Freshwater shrimps, particularly the preferred large individuals (Newman & Waters 1984), are nocturnally active (e.g. Wallace et al. 1975; Anderssen et al. 1986; Holomuzki & Hoyle 1990). Moreover, in the night adult Gammarus pulex are farther from the shore and have a lower association with large particles such as bryophytes or leaf material (Elliott 2005). Due to these nocturnal behaviours of Gammarus spp. and the longer movement distances of bullhead by night it can be speculated that encounter frequencies between

Chapter 4
76
bullhead and Gammarus spp. as well as prey susceptibility are highest during night. Because of higher encounter frequencies with prey and the ability of freshwater sculpins to forage non-‐visually (e.g. Cottus bairdii, Hoekstra & Janssen 1985) nocturnal foraging success of bullhead might be high. It can, therefore, be assumed that the longer movement distances of bullhead at night (and twilight) are associated with foraging activities.
Some individuals were stationary during daytime and showed site-‐fidelity to a diurnal location to which they returned after movement during night and twilight. It can be assumed that the diurnal location was used as a daytime refuge shelter, similar to what has been observed in Cottus gobio (Davey et al. 2005). The benefit of returning to the same diurnal location after movement during night might include spatial segregation from conspecifics to avoid competition, aggressive attacks and predation (Nilsson 2006). However, bullhead individuals did not consistently use the same location as daytime shelter. Further, none of the bullhead with diel habitat shift and site-‐fidelity to a diurnal location showed this behaviour during a preceding or a following 24-‐h cycle. During late winter and spring, the studied stream commonly has the highest annual water levels with increased stream current (personal observations). Because the shelter structures largely consist of woody debris (twigs, branches and naturally formed branch jams, Kobler et al. 2011), we assume that the spatial occurrence of shelter habitats in spring is not stable over time (in contrast to conditions in summer, Kobler et al. 2011). This may have forced bullhead to be flexible in choosing a shelter habitat as the previously occupied diurnal shelter might have vanished or shifted its location by the return after nocturnal activity. More consistent diel habitat shifts between more stable habitats had been observed in common dace. Common dace repeatedly returned to a small location in a tributary during daytime after nocturnal movement to a pool in the main river (consistent diel habitat shift during 34 days, Clough & Ladle 1997).
The 2-‐h tracking intervals used in 2007 (as suggested by Natsumeda 2007 for Cottus pollux) were crucial to detect minimum displacements that were close to the real distances that were covered during a diel cycle. Clearly, if an individual with “site-‐fidelity” to a diurnal location would have been tracked only during daytime, the distances that it covered during the entire diel cycle could not have been detected. Based on 2007 findings, we tried to cover such diel movement patterns with a less labour-‐intensive tracking protocol in 2008. Three times tracking, shortly before sunset, during dawn and again shortly before sunset, should enable the separation of nocturnal and crepuscular from diurnal movement distances. Although we found in 2008 a clear trend towards longer

Diel movement of bullhead
77
distances covered during the night period, the difference to daytime period was not significant. It is possible that an individual was close to or even had already returned to its diurnal location at the end of the night or at the beginning of dawn. Consequently, the dawn screening might have detected the fish at its diurnal location and the distances covered during night would have been underestimated; this might be indicated by the clearly shorter nocturnal movement distances recorded in the second tracking year. Although Horton et al. (2004) even proposed shorter tracking intervals and reported that 2-‐h tracking intervals poorly estimate the diel movement in spotted bass (Micropterus punctulatus), we believe that the 2-‐h interval is sufficient to detect diel movement patterns in the less active freshwater sculpins. Furthermore, by using a portable PIT tag antenna and 2-‐h tracking intervals it is still feasible to track a sufficient number of individuals in a representative length of the studied stream. We, therefore, agree with Natsumeda (2007) and recommend using 2-‐h tracking intervals.
Sex as well as the interaction between sex and diel period had no significant influence on the minimum displacements in 2008. Bullhead usually spawn in the Laarse Beek between March and April and the last young-‐of-‐the-‐year bullhead commonly hatch during May (G. Knaepkens, unpublished data). Therefore, the tracking period fell in both years into the reproductive period. The parental care of males, which construct nests and rear the larvae (Morris 1954), may restrict their movements (day and night) as the risk of egg predation by conspecifics is very high once a nest is left alone (Marconato & Bisazza 1988). And indeed, in an accompanying study, we found that the mating system of bullhead can cause a temporal sex-‐bias in the movement distances during the reproductive period (Kobler A., Humblet Y., Geudens K., Eens M., in preparation). These sex-‐biased movement distance peaks occured at the beginning (February, female-‐biased) and the end of the reproductive period (end of May, male-‐biased; same study year and site than in 2008, Kobler A., Humblet Y., Geudens K., Eens M., in preparation). The present study was conducted exactly between these phases of sex-‐biased movement (Kobler A., Humblet Y., Geudens K., Eens M., in preparation) which explains a lack of sex-‐bias and potentially the short movement distances recorded in the present study. Therefore, it would be important that future studies examine whether there is a sex-‐bias in diel movement distances during the reproductive period.
Irrespective of diel period, smaller fish covered longer distances. This may be explained by the body-‐length related refuge use under predation risk. Smaller fish (three-‐spined stickleback, Gasterosteus aculeatus) spend under predation risk

Chapter 4
78
less time in a refuge then larger fish, which can be explained by higher metabolic requirements of smaller individuals (Krause et al. 1998).
Acknowledgments
We thank G. Eens, P. Scheys and L. Damen for technical assistance. We also thank two anonymous referees and the associated editor, Bernard Hugueny, for very helpful reviews, Alain van Hout for comments and the University of Antwerp and the FWO-‐Flanders for funding (FWO-‐project to ME). The experimental work was performed in agreement with the Belgian and Flemish laws and was approved by the ethical committee of the University of Antwerp (ID number 2007/42).

CHAPTER 5
PERIOD-‐DEPENDENT SEX-‐BIASED MOVEMENT


Period-‐dependent sex-‐biased movement
81
Period-‐dependent sex-‐biased movement in a polygamous stream fish (Cottus perifretum Freyhof, Kottelat & Nolte, 2005 -‐
Actinopterygii, Cottidae) with male parental care
Alexander Kobler, Yves Humblet, Katia Geudens, Marcel Eens
Hydrobiologia (2012) 693: 195-‐204
Abstract
Inter-‐sexual differences in reproductive behaviours such as mate choice or parental care may cause sex-‐bias in movement distances. While this relationship has been extensively studied in birds and mammals, little is known regarding fishes. Fifty-‐four bullhead (Cottus perifretum), polygamous stream fish with male nest holding, were tracked by means of a portable antenna in a 2500 m stream reach. Movement was measured at two time scales: monthly movement distance and long-‐term movement range. Bullhead moved furthest in February and May. In these months, movement distances diversified between the sexes. Females moved significantly furthest in February. This may be related to female mate-‐choice at the beginning of the reproductive period. In May, at the end of the reproductive period, males moved significantly longest distances. It is speculated that males shift to resource-‐richer habitats after the starvation during parental care. In general, smaller individuals moved longer distances per month. Long-‐term movement range did not differ between the sexes but varied considerably between individuals ranging from 1 to 1111 m. It is concluded that movement studies should encompass an annual time scale as well as a more precise monthly time scale to present an accurate description of sex-‐biased movement in (annual spawning) fish.

Chapter 5
82
Introduction
Intraspecific variation in movement patterns is a common phenomenon in animal ecology and has been found in many taxa ranging from crustaceans (e.g. Hays et al. 2001) to fishes (e.g. McLaughlin et al. 1992; Kobler et al. 2009a), reptiles (e.g. McLaughlin 1989), birds (e.g. McLaughlin 1989) and mammals (e.g. Austin et al. 2004). The identification of ecological factors that trigger individuals to move is necessary to understand why movement patterns can be heterogeneous within a species or a population of a species. For instance, resource competition can force less competitive mottled sculpin (Pisces: Cottus bairdii) individuals, such as juveniles, to move to areas where intra-‐population competition is lower (Petty & Grossman 2004). The individual’s decision to move is then dependent on competitive abilities and/or its ontogenetic stage. Depending on the reproductive behaviour of the species, the triggers to move can also differ between the sexes (Greenwood 1980; Croft et al. 2003). In mammals, males typically move among female-‐defended breeding sites to maximize their reproductive success (Greenwood 1980). On the contrary, female-‐biased movement is expected when females can benefit by selecting the best males while philopatric males benefit by defending territories (Greenwood 1980; Croft et al. 2003). This is true for many bird species, in which females search for males that occupy a breeding territory (Greenwood 1980; Clarke et al. 1997). Much less is known regarding fishes and the number of studies that have examined sex-‐biased movement (here defined as the stream distance covered per time unit) in fish are still very limited (Hutchings & Gerber 2002; Croft et al. 2003; Marentette et al. 2011).
Similar to mammals and birds, the reproductive behaviour of fish can have influence on movement or dispersal (attempt to move from a natal/breeding site to another breeding site, Clobert et al. 2009). For instance, in the non-‐territorial viviparous guppy (Poecilia reticulata), male individuals do not invest in parental care and move extensively while searching for mating opportunities (Croft et al. 2003). Reproductive success of female guppies is strongly dependent on their investment in offspring while the success of male guppies is closely related to increased movement (Croft et al. 2003). These inter-‐sexual differences cause male-‐biased movement during the reproductive period (Croft et al. 2003). Female-‐biased movement during the reproductive period, on the other hand, can be expected in fish with philopatric males that invest in the offspring. These characteristics are applicable to fish in which males construct nests and rear their offspring. Prominent examples of this reproductive behaviour are three-‐spined and ten-‐spined stickleback (Gasterosteus aculateus and Pungitius pungitius). And

Period-‐dependent sex-‐biased movement
83
indeed, in both species males are less likely to move between reproduction sites than females (Whoriskey et al. 1986). Thus, the reproductive behaviour of fishes may have direct influence on sex-‐biased movement, particularly during reproductive period.
After the reproductive period and the behavioural diversification between the genders, a sex-‐bias in movement may be less probable. However, most studies that examined sex-‐biased movement in fishes have only encompassed the reproductive period (e.g. Gasterosteidae, Whoriskey et al. 1986; Apogon niger, Okuda 1999; P. reticulata, Croft et al. 2003). Other studies have examined dispersal by using genetic markers (Pseudotropheus spp., Knight et al. 1999; Carcharodon carcharias, Pardini et al. 2001; Salvelinus fontinalis, Hutchings & Gerber 2002) without the possibility of being able to differentiate periodical sex-‐bias in movement. Studies that cover both short-‐ and long-‐term movement of a fish species and differentiate between the sexes are rare (but see e.g. Natsumeda 1999; Marentette et al. 2011), but may help to detect periodical changes in sex-‐biased movement.
The present paper aims to contribute to the paucity of studies examining sex-‐biased movement in fishes under consideration of the reproductive behaviour (Hutchings & Gerber 2002; Croft et al. 2003; Marentette et al. 2011) and also takes plausible periodical variations into account. A fish species that is particularly suitable to study sex-‐biased movement is the bullhead (Cottus perifretum), a small fish that belongs to the genus Cottus (freshwater sculpins) and is found in streams and rivers in Middle Europe. The reproductive behaviour of bullhead has been extensively studied and is very similar to that of the Gasterosteidae family; polygamous males construct nests and rear their young (Morris 1954; for other Cottus spp. see also Goto 1990). Due to female mate choice in bullhead (Bisazza & Marconato 1988; Marconato & Bisazza 1988) and the bonding of a large fraction of males to a nest, we hypothesized that the movement during the reproductive period should be female-‐biased. Male courtship and nest holding behaviour (e.g. Marconato & Bisazza 1988) should result in only short distance movements in males. After the reproductive period, movement may not differ between the sexes. The results of the present study provide important data on sex-‐biased movement of a polygamous stream fish with female mate choice and nest holding males.

Chapter 5
84
Materials and methods
Study species
The popular name of C. perifretum, the bullhead, is identical to that of Cottus gobio and it was only recently that these two very closely related species were differentiated (Freyhof et al. 2005). The distribution of C. perifretum encompasses Atlantic drainages from Garonne in France to Scheldt in Belgium but also major Rhine tributaries in France and Germany. Bullhead belong to the genus Cottus that consists of small bottom-‐dwelling fishes (< 20 cm) with territorial behaviour (e.g. Japanese fluvial sculpin, Cottus pollux, Natsumeda 2001; C. bairdii, Petty & Grossman 2004; Rhine sculpin, Cottus rhenanus, Ovidio et al. 2009). They have an extremely reduced swim-‐bladder and are known for their benthic and cryptic behaviour (e.g. C. gobio, Smyly 1957; C. pollux and river sculpin, Cottus hangiongensis, Fujimoto et al. 2008). The reproductive behaviour of bullhead is characterized by female mate choice (Bisazza & Marconato 1988) and male parental care (Morris 1954). Females prefer larger males (Bisazza & Marconato 1988; C. pollux, Natsumeda 2001) and males whose nests already contain eggs (Marconato & Bisazza 1986). The nest is situated in a cavity that the male digs under solid habitat types such as rocks (Morris 1954) but can also be in tree root systems or woody debris (as in the present study). Males remain at the nest site for approximately three weeks as they find a mate, fan eggs (∼2 weeks) and guard young-‐of-‐the-‐year (∼1 week, Morris 1954).
Several studies have found remarkable intra-‐population heterogeneity in Cottus movement behaviour (C. perifretum, Knaepkens et al. 2004, 2005; other Cottus spp., Breen et al. 2009; Hudy & Shiflet 2009; Ovidio et al. 2009). Between-‐individual differences in the range of movements can be extreme. Some individuals stay during an annual period in a restricted area often not larger than a few square meters; other individuals move in the same period over distances of more than one kilometre (Hudy & Shiflet 2009; see also Knaepkens et al. 2004; Breen et al. 2009).
Study site
The study was conducted in the Laarse Beek (51°16'28.40"N; 4°29'4.63"E), a lowland stream in the north of Belgium with a mean width of 3 m (range 2 -‐ 4 m) and a mean depth of 0.3 m (0.1 -‐ 0.5 m). The focal stream reach was 2500 m long and marked every 2 m with consecutively numbered wooden poles along one side of the stream. Bullhead were the most abundant fish species in the stream, and

Period-‐dependent sex-‐biased movement
85
aquatic as well as avian predators were rare (more detailed description of the study area in Kobler et al. 2011). Bullhead usually spawn in the Laarse Beek between March and April and the last young-‐of-‐the-‐year bullhead commonly hatch during May (G. Knaepkens, unpublished data). During the reproductive period, late winter and spring, the studied stream commonly has the highest annual water levels (personal observations).
Capture, tagging and sexing
In the centre of the 2500 m observation reach, bullhead were initially caught within 324 m in two sessions, on 4, 5 and 7 February and on 13 and 14 February 2008 by electro-‐fishing (WFC7-‐10, Electracatch Int., Wolverhampton, UK) using a power generator, a 100 m cable and a 40 cm ring anode. During the two electro-‐fishing sessions 334 bullhead > 50 mm total length (TL) were caught (14 recaptures during the second session, for abundance estimate see Kobler et al. 2011). Bullhead (N = 334) had a mean (± SD) TL of 62.1 ± 6.3 mm (range 50 to 88 mm) and a mean wet weight of 3.3 ± 1.2 g (range 1.7 to 9.9 g). Shortly after capture (less than 1 h) bullhead were anaesthetized (0.25 ml L-‐1 2-‐phenoxy-‐ethanol) and individually coded 12 mm PIT-‐tags (12 x 2.1 mm, 0.094 g; ID100, EID Aalten, NL) were inserted horizontally through a 2 mm incision into the body cavity close to the genital papillae (by using a scalpel). Due to the small size of the fish and the opening, the incision was not stitched (Bruyndoncx et al. 2002). Every tagged individual was photographed (from above lying on a board coated with scale paper). After implantation and recovery in buckets (∼1 h) with stream water, bullhead were released at their capture position.
The gender of bullhead was determined retrospectively by evaluating the photographs of the individuals. At the time of first capture (and photographing), the annual gonadal maturation had already started and the gender could be determined by morphological characteristics. Individuals with a very round belly (looking tadpole-‐like), that clearly suggested the content of eggs, were determined as females. The prickles on the skin of these individuals were also stronger expressed. Males could be determined by the triangular shape of the head (Freyhof et al. 2005) and the proportionally smaller body cavity. Ambiguity about the sex led to a non-‐defined gender of the individual. This was mainly the case for larger individuals with intermediately thick belly that could be either from few eggs (e.g. older females) or extensive foraging before capture (e.g. males). The sex of every individual was determined by evaluating the photograph of an individual three times on three different days; the researcher was blind to the identity of the individual. Only when a consensus between three sex

Chapter 5
86
determinations was reached was the gender used in this study. With this method, the majority of bullhead (83%) was sexed including 138 females and 139 males.
Tracking and recapture
The study reach was scanned monthly over a distance of 2500 m: 1250 m up-‐ and downstream of the middle of the electro-‐fishing reach. This tracking distance enabled us to cover the longest movements of bullhead (observed maximum distance of a Cottus spp.: Potomac sculpin, Cottus girardi, 1711 m in 75 d, Hudy & Shiflet 2009). Tracking started two weeks after the second electro-‐fishing session and was conducted on 28 February, 27 March, 24 April, 21 May, 25 June, 16 July, 13 August and 9 September 2008. Further tracking was conducted during the individual recapture (see further). The duration of a 2500 m tracking was three days with ∼8 h d-‐1. A portable PIT-‐tag antenna with a detection precision of < 30 cm was used (Cucherousset et al. 2005). While carefully walking through the stream the antenna was moved like a vacuum cleaner. The detection probability is high with this method. Keeler et al. (2007) measured a mean detection probability of PIT tagged slimy sculpins (Cottus cognatus) in five river tributaries of 80%. Once a bullhead was located, the individual PIT-‐tag code, the stream position (e.g. stream metre 1400; marked with poles, see before) and the tracking date were noted (for more details see Kobler et al. 2011). This positioning led to an overall detection precision of less than one metre (Kobler et al. 2011). During making notes, the antenna was kept above the detected bullhead. An escape response was never observed and the tagged individuals relied on their camouflage, a typical behaviour for Cottus spp. (for more details see Kobler et al. 2011).
From 22 September to 19 November 2008 we attempted to recapture the PIT-‐tagged bullhead within the tracking reach (334 initially tagged individuals). This period was chosen as the annual gonadal maturation had not started yet and bullhead were in the following tested for animal personality differences for an accompanying study (without influence of reproductive behaviours; Kobler et al., in preparation). The fish were detected in two complete screenings of the 2500 m reach (in total 8 d) by using the portable PIT-‐tag antenna. Fifty-‐nine bullhead were caught with a large landing net (one person) and two small aquarium nets (another person). The relatively low recapture rate (18%) was most likely not due to emigration of tagged bullhead. During two additional screenings some hundred meters outside the study reach we did not find a tagged bullhead. However, we detected loose PIT tags on top of the bottom or within the stream sediment during the recapture session. PIT tag loss shortly after the tagging event is negligible (Keeler et al. 2007; Knaepkens et al. 2007), survival after PIT-‐tagging

Period-‐dependent sex-‐biased movement
87
is high (> 90%, Knaepkens et al. 2007) and predation in the stream reach was relatively low (see Study site). However, bullhead are short-‐lived often not getting older than four years (Marconato et al. 1993; Kottelat & Freyhof 2007) and the youngest tagged individuals were almost two years old (in the Laarse Beek, 0+ aged bullhead have maximum 45 mm TL, personal observations). Furthermore, while bullhead were initially tagged with up to 88 mm TL, the largest individuals that were recaptured had initially only up to 70 mm TL. We, therefore, assume that natural mortality was the main cause for the relatively low recapture rate (besides the probability to have missed some individuals during the recapture screening, see before).
Definitions and data selection
Movement was measured by a shorter-‐ (monthly movement distance) and a longer-‐term variable (movement range). Monthly movement distance was represented by the variable “minimum displacement” (m d-‐1), the distance (measured along one stream-‐side) that an individual at the minimum moved between consecutive tracking locations divided by the number of days between the locations (Rogers & White, 2007). This standardized measure accounted for the slightly varying time intervals between consecutive tracking locations (a tracking session lasted three days). The movement range (m) was calculated by the absolute distance of the most up-‐ and downstream location of an individual during the entire study period. This variable should indicate bullhead dispersal distance (for definition see Introduction).
Minimum displacements (m d-‐1) were assigned to months. A month lasted from one tracking session to the following tracking session, a period of four weeks. The last monthly period started at the September tracking session and lasted until the individual recapture date; the length of this period was very variable between individuals ranging from 12 to 82 days (mean ± SD: 35.2 ± 19.4 d). In cases where an individual was not found during a tracking session and the monthly location was missing, the minimum displacement (m d-‐1) could not be assigned to a specific month; this individual value was then excluded.
Only individuals that were recaptured (N = 59) were used in the present study. Due to this procedure, the survival throughout the study was guaranteed. A further prerequisite was the successful sex-‐determination of the individual. With this procedure, in total 54 individuals were included in the calculations.

Chapter 5
88
Statistical analysis
The distances between tracking locations (absolute values, only > 1 m) were tested for stream direction bias (one-‐way ANOVA, factor: stream direction, Levene-‐test, p > 0.05 after Ln transformation). This was done for males and females.
The effects of month, sex, body size (TL) and month × sex on minimum displacements were calculated by a linear mixed model (due to repeated measures of individuals). After transformation [due to 0 values: Ln (x + 1)] the variance of sex was homogeneous (Levene-‐test, p > 0.05). All independent variables were treated as fixed effects. The individual fish was treated as random subject in the linear mixed model. Due to the repeated measures of moving subjects (here: fish), the “diagonal” repeated covariance type was used in the linear mixed model. Pairwise comparisons between multiple factor levels were Bonferroni corrected. In case the interaction between month and sex was significant, two additional linear mixed models were run for each sex including month and body size and the dependent variable minimum displacement [transformation: Ln (x + 1)].
The effects of sex, body size and sex × body size on the individual movement range were calculated with an ANCOVA (variance homogeneity of sex, Levene-‐test, p > 0.05). The plausible influence of the individually different duration of the observation (mean ± SD 248.6 ± 18.5 d, range 221 to 289 d) on the movement range was tested by a Spearman correlation (movement range was non-‐normally distributed, Kolmogorov-‐Smirnov, p < 0.05). Significance of all calculations was assessed at α < 0.05. Statistical analyses were conducted with SPSS 15.0 (SPSS Inc., Chicago, IL, USA).
Results
Fifty-‐four individuals were sexed (including 32 females and 22 males), tracked and recaptured at the end of the study period. Their mean (± SD) body size was 65.2 ± 5.9 mm TL, (range 53 to 77 mm TL) and mean wet weight was 3.1 ± 0.9 g (range 1.7 to 5.4 g). During the observation period from 4 February to 19 November 2008, bullhead (N = 54) were on average 9.2 ± 1.1 located (range 6 to 10). 464 monthly movements consisted of 39.7% upstream movements, 33.4% downstream movements and 26.9% stationary records (< 1 m). There was no indication of up-‐ or downstream bias of the movement distances of female (N =

Period-‐dependent sex-‐biased movement
89
32, ANOVA, df = 1, ss = 0.798, F = 0.184, p = 0.668) or male bullhead (N = 22, df = 1, ss = 1.335, F = 0.298, p = 0.586). The furthest upstream movement (1096 m in ≤ 99 d; female individual) was also similar in distance to the furthest downstream movement (1000 m in ≤ 21 d; male individual).
Bullhead movement distances (minimum displacement, m d-‐1) differed significantly between the months (Table 1). Longer distances were covered in February and May (Figure 1). On average, bullhead moved 5.3 ± 11.8 m d-‐1 in February and 5.8 ± 11.6 m d-‐1 in May while for instance in August movement distances averaged only 0.3 ± 1.0 m d-‐1. Movement in February was significantly further than in all other months except March and May (all p < 0.05, pairwise comparisons from the linear mixed model of Table 1). The movement in May was significantly further than in all months from June until recapture (all months p < 0.05). Irrespective of the differentiation between months, smaller individuals had significantly higher minimum displacement values (m d-‐1; Table 1 mixed model estimate ± SE, 0.018 ± 0.005).
Monthly movement distances did not differ between males and females (Table 1). The significant interaction between sex and month on minimum displacements (p = 0.023), however, indicated temporal differences in male and female movement. Females moved longest distances in February (Table 2 estimate of fixed effect, p = 0.003). Movement distances by females in February (7.8 ± 15.1 m d-‐1) were significantly longer than in April (p = 0.009), June (p = 0.001), July (p = 0.01), August (p = 0.006) and September (p = 0.007, pairwise comparisons from the linear mixed model of Table 2; Figure 1). Shortest movement distances by females were recorded in June with only 0.1 ± 0.2 m d-‐1. During the month of their movement peak, February, females moved around four times longer distances per day than males (males, 1.8 ± 3.3 m d-‐1; Figure 1). Monthly movement of males was furthest in May (Table 2 estimate of fixed effect, p = 0.001). During May, males moved around four times longer distances per day (8.3 ± 12.2 m d-‐1) than females (females: 2.2 ± 6.8 m d-‐1). The movement by males in May was significantly longer than in August (p = 0.033) when male movement averaged only 0.1 ± 0.1 m d-‐1.
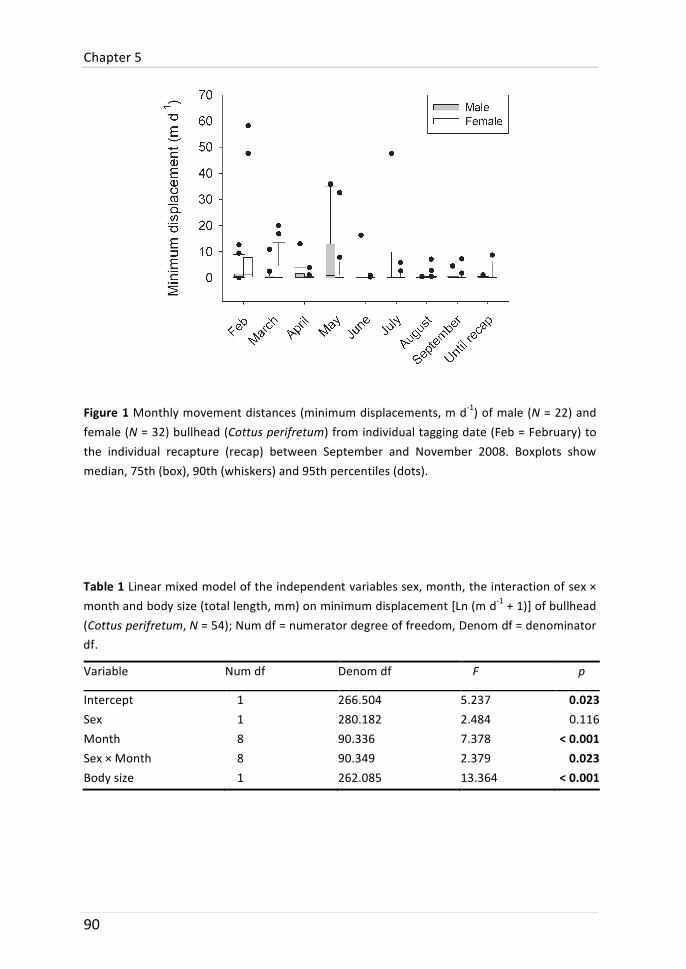
Chapter 5
90
Figure 1 Monthly movement distances (minimum displacements, m d-‐1) of male (N = 22) and female (N = 32) bullhead (Cottus perifretum) from individual tagging date (Feb = February) to the individual recapture (recap) between September and November 2008. Boxplots show median, 75th (box), 90th (whiskers) and 95th percentiles (dots).
Variable Num df Denom df F p
Intercept 1 266.504 5.237 0.023 Sex 1 280.182 2.484 0.116 Month 8 90.336 7.378 < 0.001 Sex × Month Body size
8 1
90.349 262.085
2.379 13.364
0.023 < 0.001
Table 1 Linear mixed model of the independent variables sex, month, the interaction of sex × month and body size (total length, mm) on minimum displacement [Ln (m d-‐1 + 1)] of bullhead (Cottus perifretum, N = 54); Num df = numerator degree of freedom, Denom df = denominator df.
Period-‐dependent sex-‐biased movement
91
Table 2 Two linear mixed models of month on minimum displacement [Ln (m d-‐1 + 1)] of female (N = 32) and male (N = 22) bullhead (Cottus perifretum); the intercept of both models was not significant when body size (TL) was included; Num df = numerator degree of freedom, Denom df = denominator df.
Gender Variable Num df Denom df F p
Female Intercept 1 123.900 86.712 < 0.001 Month 8 46.986 6.727 < 0.001
Male Intercept Month
1 8
81.110 30.732
50.747 5.570
< 0.001 < 0.001
The movement range (m) over the entire study period differed considerably among individuals (N = 54) ranging from 1 to 1111 m. For instance, during the ten-‐month period 18 individuals were recorded in stream ranges of ≤ 33 m while 13 individuals had a movement range ≥ 665 m. These between-‐individual differences in movement range were neither significantly related to body size nor sex (Table 3). On average (± SD), the movement range of females was 287.2 ± 332.6 m (N = 32, range 5 to 1096 m) while males had a movement range of 348.4 ± 381.6 m (N = 22, range 1 to 1111 m). The differences between individual observation duration from initial capture until recapture were not significantly correlated to the movement range (Spearman, N = 54, rs = -‐0.123, p = 0.375).
Source Type III ss Fdf p
Corrected model 638500.00a 1.8043 0.158 Intercept 98632.176 0.8361 0.365 Sex Body size
403460.695 235174.768
3.4201 1.9941
0.070 0.164
Sex × body size 407630.042 3.4561 0.069
R2 = 0.098 (adjusted R2 = 0.044)
Table 3 ANCOVA of sex and body size (total length, mm) on movement range of bullhead (Cottus perifretum, N = 54). No other variable combination gave a lower model p-‐value of the model.

Chapter 5
92
Discussion
At the beginning of the reproductive period (between February and March), females moved markedly further than males. In May, during the time period when spawning generally ends (last egg clutches in previous years were observed in the middle of May, G. Knaepkens, unpublished data), movement distances of females were clearly reduced and males moved further. The seasonal difference in movement distances of males and females indicates synchronized movement patterns within each sex. The distances covered by males and females were significantly shorter after May and bullhead were restricted to small stream areas between June and November confirming the sedentary nature of bullhead (e.g. Smyly 1957).
The longest movements distances of female bullhead were observed during the reproductive period, while afterwards they were relatively sedentary. An association with reproductive activities seems probable; spawning-‐associated movements are a common phenomenon in stream fishes (Lucas & Baras 2001) and are also reported for Cottus spp. (e.g. C. pollux, Natsumeda 1999). For instance, Japanese fluvial sculpins move long distances while shifting from marsh habitats to stream areas with abundance of stones that are more suitable for spawning (Fujimoto et al. 2008). In the present study, however, spawning habitat types (here: tree roots and woody debris) occurred in high abundance and were homogeneously distributed in the stream (Kobler et al. 2011). Extreme habitat type differences that were separated by long stream distances did also not occur (Kobler et al. 2011). Therefore, it seems unlikely that the long distance movements of females were associated to habitat shifts. Another explanation for those movements may be mate-‐choice processes. It is well known that female bullhead actively search for a mate to guard the eggs (Bisazza & Marconato 1988). Larger males (Bisazza & Marconato 1988; Natsumeda 2001) and males whose nests already contain eggs (Marconato & Bisazza 1986) are generally preferred. During this period males occupy a potential nest site where they attract females for spawning (Morris 1954). After successful spawning, males may move only very little as the risk of egg predation by conspecifics is very high once a nest is left unattended (Marconato & Bisazza 1988; see also further discussion). These sex-‐typical reproductive behaviours imply that bullhead movement should be female-‐biased during the reproductive period, movement observations in the present study are in accordance with this hypothesis. It may be concluded that females cover longer distances during mate-‐search and that this behaviour has a pronounced influence on sex-‐biased movements during reproductive period.

Period-‐dependent sex-‐biased movement
93
There was no relation between female movement (monthly movement distance and movement range) and body size. Similarly, Natsumeda (2001) could not relate the individual spawning movement differences in female Japanese fluvial sculpins to their ontogenetic stage or body size. Approximately half of the observed female Japanese fluvial sculpins moved longer distances and left their home range for spawning (Natsumeda 2001). Natsumeda (2001) speculated that these females separated spawning from feeding areas, a phenomenon also common in other fish species (for references see Natsumeda 2001). Natsumeda (2001) further speculated that the temporal limitation in mate availability may force some females to move over greater distances .In this case, females with faster gonadal development could choose for mates nearby while females with slower gonadal development and a later spawning time may have to swim over greater distances during mate search. The between-‐individual differences in female movement may thus be related to gonadal development. Based on this speculation, a future study with shorter observation intervals and the determination of gonadal development may find a negative relation between the distance covered by females and the stage of gonadal development.
Movement in May was male-‐biased. At the beginning of the reproductive period, males stay at a potential nest-‐site where they attract females for spawning (Morris 1954). Males remain at the nest site for approximately three weeks as they find a mate, fan eggs and guard young-‐of-‐the-‐year (Morris 1954). It can be assumed that the increased movement between the end of April and the end of May occurred when males finished with parental care. Accordingly, last egg clutches were observed in previous years in May (G. Knaepkens, unpublished data). During the entire period of parental care the food intake is very low and the condition of the male bullhead strongly decreases (Marconato et al. 1993; C. pollux, Natsumeda 2001). Starvation can even trigger males to cannibalise the eggs in their own nest (Marconato & Bisazza 1988). It can be assumed that male bullhead forage extensively after the period of starvation. Due to spatial and temporal fluctuations of prey abundance in lotic bodies of water (Schlosser & Angermeier 1995) and the active choice of Cottus spp. for stream patches with higher abundance of macroinvertebrates (C. bairdii, Petty & Grossman 1996), it seems plausible that some males will have moved from their breeding site to stream areas with higher food abundance. Therefore, the longer movement distances after reproduction might be related to resource-‐associated home range shifts (see Crook 2004). After recovery of the energy deficiency of nest holding, males may then switch again to their typical sedentary behaviour (e.g. Smyly 1957) as observed in the present study.

Chapter 5
94
Inconsistent with the hypothesis and results of the present study, Natsumeda (1999) observed male-‐biased movement of C. pollux during the reproductive period; males moved from pools or riffles to raceways while females stayed in their occupied channel-‐unit habitat. This contrary finding is surprising (based on the hypothesis of the present paper) and may be related to the distribution of suitable spawning habitats (see beginning of discussion). Future studies need to take the distribution of suitable spawning habitats into account as this may affect or even change the sex-‐bias of spawning movements in stream fishes with female mate-‐choice and male parental care.
In conclusion, the movement of C. perifretum, a stream fish species with male nest holding and parental care, is periodically sex-‐biased. As predicted based on the reproductive behaviour of bullhead, movement was female-‐biased at the beginning of the reproductive period. After reproduction, movement was male-‐biased resulting in equal movement ranges between the sexes during the ten-‐month observation period. The male-‐biased movement may be associated with home-‐range shifts to resource-‐rich stream areas after the starvation during nest holding. The short periods in which the movement diversified between the sexes emphasize the importance of short observation intervals (e.g. weekly or monthly) and a sufficiently long study period (preferably one year) to make conclusions about sex-‐biased movement patterns. Sex-‐biased movement may play a role in many stream fishes and its consideration may add extra complexity to the conservation of a species (Croft et al. 2003). Furthermore, the consideration of reproductive behaviours may help to better understand intra-‐population heterogeneity in movement distances, for instance the occurrence of resident and mobile individuals within single stream fish populations (e.g. Gowan et al. 1994).
Acknowledgments
Many thanks to Geert Eens, Peter Scheys, Sofie Vanwetswinkel, Brecht Engelen and Lennart Damen for technical assistance, Guy Knaepkens, Gregory E. Maes and Filip A.M. Volckaert for support. Many thanks also to the University of Antwerp and the Fonds Wetenschappelijk Onderzoek -‐ Vlaanderen for funding (FWO-‐project to Marcel Eens). The experimental work was performed in agreement with the Belgian and Flemish laws and was approved by the ethical committee of the University of Antwerp (ID number 2007/42).

CHAPTER 6
MOVEMENT RANGE AND TEMPERAMENT TRAITS


Movement range and temperament traits
97
Dispersal and behaviour: is the movement range of the bullhead
Cottus perifretum related to temperament traits?
Alexander Kobler, Gregory E. Maes, Sofie Vanwetswinkel, Dimitri Geelhand de Merxem, Marcel Eens
In preparation
Abstract
Populations of non-‐migratory stream fish may consist of resident and mobile individuals. The propensity to disperse may be related to body size or age but a behavioural association may also exist. This study aims to explain individual differences in movement range (individual distance between the most upstream and downstream location) by testing temperament traits. Tagged bullhead (Cottus perifretum) were tracked during ten months. Movement range, which ranged from 1 to 1284 m, was independent of either body size or sex. A clear distinction between resident and mobile individuals was not found. Fifty-‐three tagged and 21 untagged bullhead were caught and tested for temperament traits. There was no behavioural difference between tagged and untagged fish; repeated measurements of behavioural traits (activity and novel environment activity) indicated consistency. Smaller bullhead were significantly more active in a novel environment. Boldness (rs = 0.227, p = 0.106) and interest in novel food (rs = 0.222, p = 0.109) correlated weakly with movement range. These results may indicate that bullhead that expanded further in the stream may be more risk-‐prone. It was concluded that bullhead movement range is only weakly related to some temperament traits suggesting no different “personality” as cause or consequence of movement range diversification.

Chapter 6
98
Introduction
Although the majority of non-‐migratory stream fish prefer to stay in a very restricted area, there are also fish individuals that move farther away (Gerking 1959). Consequently, numerous studies have differentiated non-‐migratory stream fish populations into resident and mobile individuals (e.g. Smithson & Johnston 1999; Nakamura et al. 2002; Rodriguez 2002; Knaepkens et al. 2004). The existence of this more mobile fraction is not surprising due to its importance for the reduction of inbreeding, the spread of new genes (Howard 1960), adaptation to changing conditions and the repopulation of depauperate areas (Gerking 1959). Which individual traits distinguish resident and mobile individuals is, however, not well understood. Gerking (1959) doubted whether resident and mobile stream fish can be phenotypically distinguished and grouped.
Recently, there are indications that some individual traits are associated to long-‐term movement distances (hereafter called movement range) such as body size (Gowan & Fausch 1996) and age (Hughes 2000; Petty & Grossman 2004). Other studies could not find a relation between life-‐history traits and movement range of stream fish although intra-‐population heterogeneity was high (e.g. Knaepkens et al. 2005; Breen et al. 2009; Hudy & Shiflet 2009; Ovidio et al. 2009). In the absence of a life-‐history trait association, a behavioural relation to movement range or a behavioural grouping of resident and mobile stream fish may exist and remains to be determined (Howard 1960; Rodriguez 2002).
Animal temperament is classified in five main temperament categories (boldness, activity, exploration, aggressiveness and sociability (Réale et al. 2007). It may represent a key for an improved understanding of movement range diversification in stream fish (Fraser et al. 2001). And indeed, temperament traits have been linked to behaviours that are similar or closely related to the movement range of stream fish, such as dispersal (reviewed by Clobert et al. 2009) and movement in an unfamiliar environment (Fraser et al. 2001; Cote et al. 2010, 2011). Why, for example, temperament traits may be related to dispersal can be explained by the costs that are involved in the process of dispersal especially during transience and at settlement (Clobert et al. 2009; Bonte et al. 2012). Such costs can be reduced due to enhanced expression of a temperament trait (Duckworth & Badyaev 2007; Clobert et al. 2009). For instance, western bluebirds (Sialia mexicana) adaptively express a higher level of aggressiveness after dispersal and during the colonization process suggesting that behavioural changes may play an important role in range expansion processes (Duckworth & Badyaev 2007). Similar adaptations may also be involved after stream fish moved

Movement range and temperament traits
99
over long distances. Individual movement range may thus be related to a different expression of temperament traits.
To test whether the movement range of a non-‐migratory stream fish is associated with individual temperament traits, we used the bullhead (Cottus perifretum) as model species. Its vernacular name (bullhead) is identical to that of C. gobio and it was only recently that these two very closely related species were differentiated (Freyhof et al. 2005). The distribution of C. perifretum encompasses Atlantic drainages from the Garonne in France to the Scheldt in Belgium but also major Rhine tributaries in France and Germany. The genus Cottus (freshwater sculpins) consists of small bottom-‐dwelling fishes (< 20 cm) with territorial behaviour (e.g., Japanese fluvial sculpin, C. pollux, Natsumeda 2001; mottled sculpin, C. bairdii, Petty & Grossman 2004, 2007; Rhine sculpin, C. rhenanus, Ovidio et al. 2009). They have an extremely reduced swim-‐bladder (Kottelat & Freyhof 2007) and are known for their benthic and cryptic behaviour (e.g. C. gobio, Smyly 1957; C. pollux and river sculpin, C. hangiongensis, Fujimoto et al. 2008). Nevertheless, numerous studies reported remarkable intra-‐population heterogeneity in movement range (C. perifretum, Knaepkens et al. 2004, 2005; other Cottus ssp., Breen et al. 2009; Hudy & Shiflet 2009; Ovidio et al. 2009). Whereas some individuals stay over time in a restricted area often not more than a few square meters, other individuals may move more than one kilometre (Hudy & Shiflet 2009; see also Knaepkens et al. 2004; Breen et al. 2009; Ovidio et al. 2009).
We hypothesized that bullhead with longer movement range (“mobile” individuals) are more explorative. A similar relation has been found in juvenile great tits (Aves: Parus major) in which dispersal correlates positively with exploratory tendency (Dingemanse et al. 2003). Furthermore, a positive relation between movement range and aggressiveness seemed plausible similar to what has been observed in Western bluebirds (see previous introduction, Duckworth & Badyaev 2007). These hypotheses as well as the plausible relations between movement range and other temperament traits were tested by using passive integrated transponder (PIT) telemetry during a ten-‐month observation period, recapture of the tagged bullhead and subsequent measurements of boldness, novel environment activity, interest in novel food, activity and aggressiveness in the laboratory. The results of this study should give insight into between-‐individual differences in non-‐migratory stream fish movement range by including information on body size, sex and temperament traits. Furthermore, the study should also give insight into the individual repeatability of temperament traits

Chapter 6
100
and whether PIT tagged and untagged bullhead show differences in the expression of temperament traits.
Materials and methods
Study site
The study was conducted in the Laarse Beek (51°16'28.40"N; 4°29'4.63"E), a lowland stream belonging to the Scheldt basin (Flanders, Belgium). The focal stream stretch was 2500 m long, had a mean width of 3 m (range 2 -‐ 4 m) and a mean depth of 0.3 m (0.1 -‐ 0.5 m). The focal stream reach was 2500 m long and marked every 2 m with consecutively numbered wooden poles along one side of the stream. Bullhead were the most abundant fish species in the stream, and aquatic as well as avian predators were rare (for a more detailed description of the study area see Kobler et al. 2011).
Capture and tagging
In the centre of the 2500 m observation reach, bullhead were caught within 324 m in two sessions, on 4, 5 and 7 February and on 13 and 14 February 2008 by electro-‐fishing (WFC7-‐10, Electracatch Int., Wolverhampton, UK) using a power generator, a 100 m cable and a 40 cm ring anode. During the two electro-‐fishing sessions 334 bullhead > 50 mm total length [TL; minimum size to PIT-‐tag, Knaepkens et al. 2007] were caught (14 recaptures during the second session, for abundance estimate see Kobler et al. 2011). Bullhead (N = 334) had a mean (± SD) TL of 62.1 ± 6.3 mm (range 50 to 88 mm) and a mean wet weight of 3.3 ± 1.2 g (range 1.7 to 9.9 g). Thanks to tagging shortly before spawning, the most bullhead were sexed reliably (138 females and 139 males; Kobler et al. 2011, 2012b). Shortly after capture (less than 1 h) bullhead were anaesthetized (0.25 ml L-‐1 2-‐phenoxy-‐ethanol) and individually coded 12 mm PIT-‐tags (12 x 2.1 mm, 0.094 g; ID100, EID Aalten, NL) were introduced horizontally through a 2 to 3 mm incision into the body cavity close to the genital papillae (by using a scalpel). Due to the small size of the fish and the opening, the incision was not stitched (Bruyndoncx et al. 2002). After implantation and recovery in buckets (∼1 h) with stream water, bullhead were released at their capture position.

Movement range and temperament traits
101
Tracking and recapture
The study reach was scanned monthly over a distance of 2500 m: 1250 m up-‐ and downstream of the middle of the electro-‐fishing reach. This tracking distance enabled us to cover the longest known movements of bullhead (observed maximum distance of a Cottus spp.: Potomac sculpin, C. girardi, 1711 m in 75 d, Hudy & Shiflet 2009). Tracking started two weeks after the second electro-‐fishing session and was conducted monthly on 28 February, 27 March, 24 April, 21 May, 25 June, 16 July, 13 August and 9 September 2008. Further tracking was conducted during the individual recapture (see further). A portable PIT-‐tag antenna with a detection precision of < 30 cm was used (for more details see Kobler et al. 2011, 2012b). The detection probability of Cottus ssp. is high with this method (∼80%, Keeler et al. 2007). Once a bullhead was located, the individual PIT-‐tag code, the stream position (e.g. stream meter 1400; marked with poles, see before) and the tracking date were noted (for more details see Kobler et al. 2011). This positioning led to an overall detection precision of less than one meter (Kobler et al. 2011). During making notes, which took approximately half a minute, the antenna was kept above the detected bullhead. An escape response was never observed and the tagged individuals relied on their camouflage, a typical behaviour for Cottus spp. (for details see Kobler et al. 2011).
From 22 September to 19 November 2008 we attempted to recapture the PIT-‐tagged bullhead within the 2500 m tracking stretch. This period was chosen as the annual gonadal maturation had not started yet and reproductive behaviours would not influence the following temperament traits tests (see further). The fish were detected in two complete screenings of the 2500 m reach (in total 8 d) by using the portable PIT-‐tag antenna. Fifty-‐nine bullhead were caught with a large landing net (one person) and two small aquarium nets (another person). The relatively low recapture rate (18%) was most likely not due to emigration of tagged bullhead. During two additional screenings some hundred meters outside the study reach we did not find a tagged bullhead. However, we detected loose PIT tags on top of the bottom or within the stream sediment during the recapture session. PIT tag loss shortly after tagging is negligible, the survival after PIT-‐tagging is high (∼90%; Keeler et al. 2007; Knaepkens et al. 2007) and the predation in the stream reach was relatively low (see Study site). However, bullhead are short-‐lived often not reaching the age of four years (Marconato et al. 1993; Kottelat & Freyhof 2007); the youngest tagged individuals were almost two years old (in the Laarse Beek, 0+ aged bullhead have maximum 45 mm TL, personal observation). Furthermore, while bullhead were initially tagged up to a size of 88 mm TL, the largest individuals that were

Chapter 6
102
recaptured had initially only up to a size of 70 mm TL. We, therefore, assume that natural mortality was the main cause for the relatively low recapture rate (besides the probability to have missed some individuals during the recapture screening).
The monthly localisations from initial tagging to recapture were used to calculate the individual movement range. It is the absolute distance between the most up-‐ and downstream location of an individual during the entire ten-‐month study period (Kobler et al. 2012b). The study period included the reproductive period of bullhead, during which the longest distances are covered and an association to reproductive activities is likely (Kobler et al. 2012b). Therefore, movement range served as an indicator for the individual breeding dispersal distance, which is the attempt to move from a breeding site to another breeding site (Clobert et al. 2009).
Additionally, untagged individuals (> 50 mm TL, N = 21) were collected and marked with individually coded stripes of visible implant elastomer (VIE; Northwest Marine Technology Inc., Salisbury, UK) along the anal fin. These individuals were later used to test the eventual influence of the PIT-‐tag on the temperament traits of the bullhead and therefore defined “untagged” (see further). In addition, every week bullhead of all size classes were caught to serve as stimuli for the aggressiveness test that would be carried out later in the laboratory (see further). Due to the capture date and insufficient body size, sex could not be determined for the untagged and the stimulus fish (Freyhof et al. 2005). The fish were measured (TL) and weighed (wet weight) and transferred on the day of capture to an outdoor 400-‐L tank with oxygenated and (bio-‐) filtered tap water at the animalium of the university. The tank contained PVC tubes (10 x 5 cm) as structuring refuges (more tubes than fish). The mean (± SD) water temperature was 9.8 ± 2.7 °C (range 5.3 – 14.1 °C) and mean oxygen concentration was 9.1 ± 1.2 mg L-‐1 (range 7.7 – 11.1 mg L-‐1). Bullhead were fed daily to satiation with frozen bloodworms.
Temperament traits
We used eleven plastic tanks (79 x 42 x 57 cm), lined with blue plastic foil for the experiments. The tanks were half-‐filled and contained approximately 80 L of oxygenated tap water with a mean of 14.9 ± 1.5 °C (range 12.7 – 17.2 °C) and a mean oxygen concentration of 7.8 ± 0.8 mg L-‐1 (range 6.5 – 9.6 mg L-‐1). We adjusted the light/dark cycle weekly to the seasonal natural conditions. Light intensity was dimmed to mimic the stream conditions in the forest.

Movement range and temperament traits
103
The tanks were set up such that all tests for each individual could be conducted in just one tank (Figure 1) although two transfers into novel environments and a predator introduction were done (see later). Therefore, the tanks were temporally visually (white plastic plate) and constantly physically (metal mesh) divided into two parts. Ten percent of the bottom of both tank sides (A and B) was structured; different objects created two different environments (Figure 1). Under the transparent bottom of the tank a grid of 7.5 x 7.5 cm quadrats was drawn; it was chosen in view of the maximum size of the recaptured bullhead (77 mm TL). Above the tanks we positioned a digital camcorder (DCR-‐PC6E or DCR-‐DVD310E, Sony, Tokyo, Japan).
Figure 1 Laboratory set-‐up of the 80-‐L experimental tanks for the temperament trait tests. Both tank sections were similar in the amount of structure they provided. A vertical white plastic plate was fixed 0.5 cm below the surface, water was allowed to flow between the sides and the same water was used for each test. The white plastic plate guaranteed that the fish, that was introduced into tank section A, did not experience the environment in tank section B. Removal of the plastic plate before the boldness test enabled the perception of visual and chemical cues of the predator on the opposite tank section.
Under the artificial tank conditions, bullhead were mostly hidden in a refuge and showed no activity at all during the simulated daylight. To record behavioural activity, the tests were recorded at the beginning of the night period
Chapter 6
104
by using the night-‐shot function of the camcorders. An observer was only present to turn the camcorders on in complete darkness. The tests lasted for 10 min starting from 2 min after the recordings started (more details in Kobler et al. 2011). Four temperament categories were examined: 1) Boldness-‐shyness, an individual’s reaction to any risky situation, but not a new situation, 2) Exploration-‐avoidance, an individual’s reaction towards a new situation; this includes behaviour towards a new habitat, new food or novel objects, 3) Activity, the general level of activity of an individual in non-‐risky and non-‐novel environment and 4) Aggressiveness, an individual’s agonistic reaction towards conspecifics (for more details on these definitions see Réale et al. 2007).
Day Action and temperament test
0 -‐ 2 -‐(Re)-‐capture of tagged and untagged bullhead -‐Transport to a 400-‐L outdoors tank at the university
3
-‐Renewal of water of the 80-‐L laboratory tank (9 am) -‐Translocation from outside tank to laboratory tanks -‐Novel environment activity (NEA) 1 in tank section A
4 -‐ 5 -‐Familiarization to tank section A
6 -‐Activity-‐test (ACT) 1 in tank section A -‐Aggressiveness-‐test: introduction of stimulus fish
7 -‐NEA 2 in tank section B
8 -‐Removal of visual barrier between tank sides (9 am) -‐ACT 2 in tank section B -‐Interest in novel food test in tank section B
9
-‐ACT 3 in tank section B -‐Boldness-‐test: predator (pike) into tank section A -‐End of tests; remove fishes to second outdoors tank
Table 1 Schedule of the temperament trait analysis of bullhead in the laboratory from 3 October to 28 November 2008 (N = 53 tagged, N = 21 untagged individuals). When two tests were conducted on a day, they followed each other immediately.

Movement range and temperament traits
105
The general procedure of the behavioural laboratory tests was based on the study of Dingemanse et al. (2007) and is summarized in Table 1. The temperament trait tests (3 October to 28 November 2008) started with the introduction of a bullhead into the unfamiliar 80-‐L laboratory tanks through a vertically positioned plastic tube (30 x 10 cm) that was removed directly before the test (Kobler et al. 2011). Novel environment activity (exploration-‐avoidance category) was quantified by the number of quadrats crossed per minute (quadrats min-‐1) regardless of the direction or position in the tank and was measured twice (Table 1). Novel environment activity is a good indicator for the exploratory tendency of an individual (e.g. Cote et al. 2010).
Interest in a novel food item (exploration-‐avoidance category) was measured in a familiar environment (Table 1) by the bullhead’s reaction towards a small maggot of terrestrial Diptera, that we anticipated being an unknown food item (Kobler et al. 2011). The reaction was measured in terms of 1) the approach to the maggot (bullhead was situated in the quadrat next to the square of the maggot), 2) the interest in the maggot (both in the same quadrat) and 3) the ingestion of the maggot.
Activity was measured three times after allowing familiarization to the environment for at least 12 h (Table 1) by quantifying the distance moved (regardless of direction) with quadrats min-‐1 as variable (Kobler et al. 2011).
Aggressiveness was tested in a familiar environment (Table 1) with the introduction of a smaller conspecific into the tank (Kobler et al. 2011). Stimulus fish were distributed across the tanks by size (mean ± SD, 46.8 ± 4.4 mm TL; range 35 -‐ 62 mm) to guarantee a similar size ratio between the individuals assayed. The number of bites was counted and the distance moved was recorded (regardless of direction; quadrats min-‐1). The individual’s distance was later compared to the distance moved directly before the test (see statistics).
Boldness-‐shyness was tested in tank section B with the introduction of a pike (∼30 cm TL) into tank section A (Figure 1). Because the tank sections were only separated by a metal mesh (Figure 1), visibility was provided and the odour of the predator diffused between the sections. We measured 1) the mean time (s) as well as the proportion of time (%) spent in the proximity to structuring tank objects (distance < 1 quadrat, Figure 1), 2) the proportion of time spent in the tank section half that was close to the pike (%), and 3) the distance moved in the presence of the predator (quadrats min-‐1). These variables were quantified directly before the test (control) as well as during the presence of the predator,

Chapter 6
106
and statistically compared to measure the reaction towards the predator (see statistics).
Although PIT tagging has been shown to have negligible effects on bullhead survival, growth and swimming performances (Knaepkens et al. 2007), all tests were conducted with field-‐observed PIT-‐tagged (N = 53 of 59 recaptured bullhead; the temperament tests failed for six individuals due to different reasons) and untagged bullhead (N = 21). This allowed us to examine whether tagging had any effects on the temperament traits under study (see statistics).
After the test series (day 10, Table 1) bullhead were brought to a second 400-‐L outdoors tank, where they were stored until release back into the stream. The water of the laboratory tanks was renewed before the next test series.
Statistics
Only individuals that were recaptured and successfully tested for temperament traits (N = 53) were used for the movement range analyses. Whether body size (TL), sex and body size × sex had an influence on the individual movement range (m) was calculated with an ANCOVA (variance homogeneity of sex, Levene-‐test, p > 0.05).
Additionally, we also tested whether there would be similar results when only the extreme 33% of the movement range values are included in the calculations [(lowest and highest 33% movement range values (N = 36) of all bullhead studied (N = 53)]. This was done to exclude individuals with intermediate movement range distances and put emphasis on distinct individuals, similar to a categorization into resident and mobile individuals. The resulting groups were tested for differences in sex ratio (Chi-‐square test) and body size (independent t-‐test, equal variances, Levene-‐test, p > 0.05).
Two temperament traits (aggressiveness and boldness) measured the difference between the distances moved from just before to during a stimulation test (see Temperament traits). Each individual tested represented its own control and only the change from “normal” to stimulated behaviour was used as behavioural variable. The change in distance moved was defined by the individual residual value of the regression of all tested individuals’ stimulus activity [LN (quadrats min-‐1 + 1), y-‐axis] on the control activity of all tested individuals [LN (quadrats min-‐1 + 1), x-‐axis; see also Garamszegi et al. 2009]. Negative values meant that the individual decreased movement during the stimulus test more

Movement range and temperament traits
107
than expected based on the reaction of all individuals tested. Similar calculations of the behavioural difference of control and stimulus were also made for the boldness variables refuge use (%), mean refuge use (s) and tank side positioning (%).
For temperament traits that had more than one behavioural measure by definition (interest in novel food, aggressiveness and boldness), we merged several measures to one variable (Table 2). In the principal component analysis (PCA) values were z-‐transformed, Eigenvalues above one were extracted, multiple components rotated by varimax method and the new variables of the principal components (PC) were saved by regression method (Dingemanse et al. 2007; see also Kobler et al. 2011). Interest in novel food was represented by a PC in which the approach, the interest and the ingestion of the maggot were positively correlated (Table 2). In the aggressiveness PC the individual’s activity change and the number of bites were positively correlated, suggesting that fish, which bite more often became relatively more active when the stimulus fish was introduced. A positive value of the boldness PC meant that after the introduction of a predator (pike) individuals spent relatively more time close to a refuge (%), had on average longer stays at a refuge (s) and spent relatively less time in the tank half close to the predator (%) compared to what would be expected from the reaction of all tested individuals (see previous paragraph). The boldness PC was included in further calculations as additive inverse [x = boldness * (-‐1)] so that positive values meant a bolder (and not a shyer) behaviour.
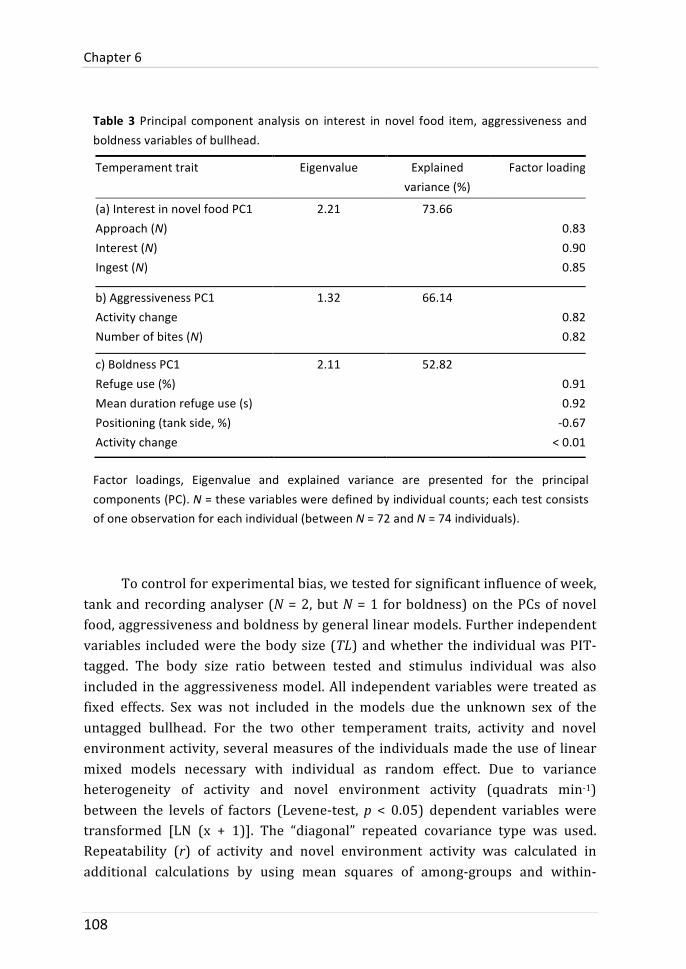
Chapter 6
108
To control for experimental bias, we tested for significant influence of week, tank and recording analyser (N = 2, but N = 1 for boldness) on the PCs of novel food, aggressiveness and boldness by general linear models. Further independent variables included were the body size (TL) and whether the individual was PIT-‐tagged. The body size ratio between tested and stimulus individual was also included in the aggressiveness model. All independent variables were treated as fixed effects. Sex was not included in the models due the unknown sex of the untagged bullhead. For the two other temperament traits, activity and novel environment activity, several measures of the individuals made the use of linear mixed models necessary with individual as random effect. Due to variance heterogeneity of activity and novel environment activity (quadrats min-‐1) between the levels of factors (Levene-‐test, p < 0.05) dependent variables were transformed [LN (x + 1)]. The “diagonal” repeated covariance type was used. Repeatability (r) of activity and novel environment activity was calculated in additional calculations by using mean squares of among-‐groups and within-‐
Temperament trait Eigenvalue Explained variance (%)
Factor loading
(a) Interest in novel food PC1 Approach (N) Interest (N) Ingest (N)
2.21
73.66
0.83 0.90 0.85
b) Aggressiveness PC1 Activity change Number of bites (N)
1.32
66.14
0.82 0.82
c) Boldness PC1 Refuge use (%) Mean duration refuge use (s) Positioning (tank side, %) Activity change
2.11
52.82
0.91 0.92 -‐0.67 < 0.01
Table 3 Principal component analysis on interest in novel food item, aggressiveness and boldness variables of bullhead.
Factor loadings, Eigenvalue and explained variance are presented for the principal components (PC). N = these variables were defined by individual counts; each test consists of one observation for each individual (between N = 72 and N = 74 individuals).

Movement range and temperament traits
109
groups variance of a one-‐way ANOVA with “individual” as a factor on the dependent variables (Lessells & Boag 1987).
,
where s2^ is the among-‐groups variance component and s2 is the within-‐group variance component.
Due to the unbalanced number of individual habitat localizations, the coefficient (n0) related to the sample size had to be calculated (Lessells & Boag, 1987).
The relation between the temperament traits and the gender of an individual was calculated for the PIT-‐tagged individuals by one-‐way ANOVAs.
Temperament traits were tested for correlation by Spearman rank correlation due to non-‐normality of novel environment activity, interest in novel food, activity, and aggressiveness (no transformation normalized, Kolmogorov-‐Smirnov, p < 0.05). Novel environment activity and activity were represented by the mean value of the multiple measurements while the other temperament traits were represented by the PCs.
Whether the movement range of bullhead was correlated to temperament traits was tested by Spearman rank correlations (non-‐normal distribution of movement range, Kolmogorov-‐Smirnov, p < 0.05).
Whether resident and mobile groups of bullhead are different in their expression of temperament traits was tested by independent t-‐tests. For the temperament traits novel environment activity and interest in novel food equal variances were not assumed (Levene-‐test, p < 0.05).
Significance of sets of multiple pair-‐wise comparisons was assessed at a false discovery rate (FDR) adjusted α (Benjamini & Hochberg 1995). Significance of all other calculations was assessed at α < 0.05. Statistical analyses were conducted with IBM SPSS Statistics 20.0 for Mac (IBM Corporation, New York, USA).
r = s2^ / s2 + s2^( )
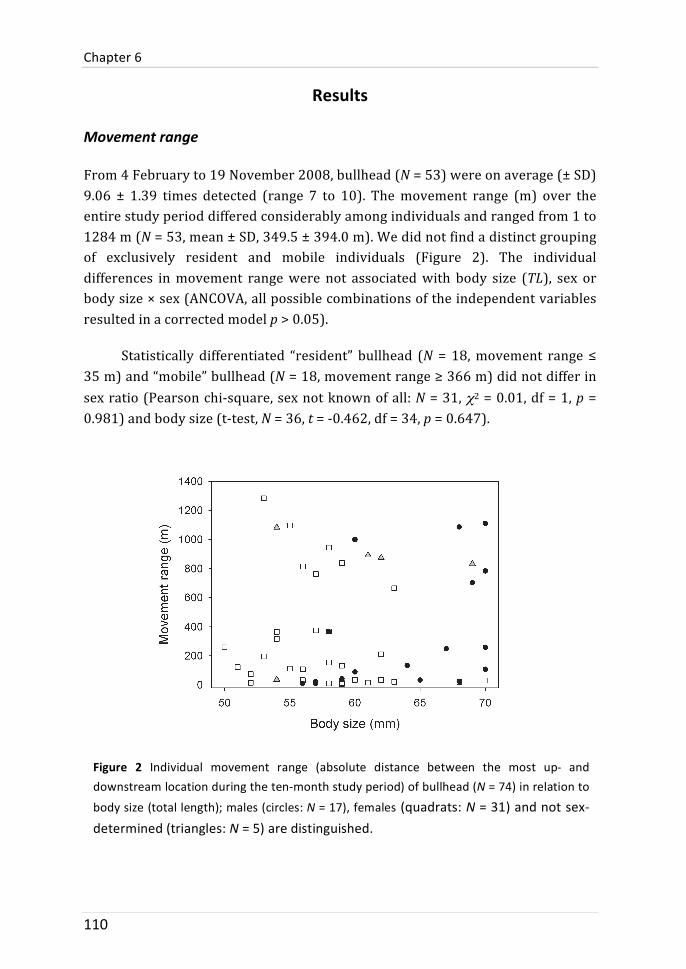
Chapter 6
110
Results
Movement range
From 4 February to 19 November 2008, bullhead (N = 53) were on average (± SD) 9.06 ± 1.39 times detected (range 7 to 10). The movement range (m) over the entire study period differed considerably among individuals and ranged from 1 to 1284 m (N = 53, mean ± SD, 349.5 ± 394.0 m). We did not find a distinct grouping of exclusively resident and mobile individuals (Figure 2). The individual differences in movement range were not associated with body size (TL), sex or body size × sex (ANCOVA, all possible combinations of the independent variables resulted in a corrected model p > 0.05).
Statistically differentiated “resident” bullhead (N = 18, movement range ≤ 35 m) and “mobile” bullhead (N = 18, movement range ≥ 366 m) did not differ in sex ratio (Pearson chi-‐square, sex not known of all: N = 31, χ2 = 0.01, df = 1, p = 0.981) and body size (t-‐test, N = 36, t = -‐0.462, df = 34, p = 0.647).
Figure 2 Individual movement range (absolute distance between the most up-‐ and downstream location during the ten-‐month study period) of bullhead (N = 74) in relation to body size (total length); males (circles: N = 17), females (quadrats: N = 31) and not sex-‐determined (triangles: N = 5) are distinguished.

Movement range and temperament traits
111
Temperament traits
There was no influence of PIT-‐tagging on the expression of temperament traits: PIT-‐tagged and untagged (solely VIE-‐marked) individuals behaved similarly (Table 3). The individual expression of the temperament traits was not related to the gender (sex not known of all 53 bullhead: N = 47, ANOVAs, all p > 0.05). Novel environment activity was found to be significantly higher in smaller individuals (Table 3). Unintentional potential influences on experiments (e.g. observation week difference) showed no experimental bias. Replicated temperament trait tests indicated behavioural consistency during the laboratory test period with r = 0.479 for activity (N = 73 bullhead) and r = 0.529 for novel environment activity (N = 74). Novel environment activity was significantly correlated to all other temperament traits and activity was significantly related to boldness (Table 4).
Ta
ble 3 Tw
o linear m
ixed
mod
els (with
rand
om effe
ct) a
nd th
ree gene
ral linear m
odels
to con
trol for significan
t influ
ences of experim
ental varia
bles on the tempe
ramen
t traits. C
ontrolled were diffe
rences between expe
rimen
tal w
eek (N = 8), tank (N
= 11),
the pe
rson
who
analysed the vide
os (N = 2; except for boldn
ess), tagged/un
tagged
status and
for b
ody siz
e (to
tal len
gth, m
m). Ad
ditio
nally, the
eventua
l effe
ct of the
size
ratio
of the tested
bullhead an
d the stim
ulus fish
was tested in the
aggressiven
ess
mod
el. F
or rep
eated measures the individu
al differen
ces as w
ell a
s the repe
atab
ility
are presen
ted; significant re
sults are in bold.
!"#$%&!
%'$()#'*
%'+!
,-+($
(+.!
/'+%)%0+!('!!
'#$%&!1##
2!!
! 3-+($
(+.!
! 344)%00($
%'%00!
! 5#&2'%
00!!
6,)(,
7&%!
! 21!!
"#! 2
1!"#
! 21!
"#! 2
1!"#
! 21!
"#
!"#$%&$''$()*&
/'+%)-%8
+!
9%%:!
;,':!
3',&.<%)!
;,44%2
!!
5#2.!0(<%!
=(<%!),
+(#!
+,-%
./&$''$
()*&
/'2($(2>
,&!
?$@!
! ABCDEF
A!
ACGFH I!
ACAAF A
F!
DCGHD A!
FCFJK A!
HCBGE A
!
L ! 9,&2!%!
DCBIA!
?FCBDK
@!
! MFCFFA
!
FCAFE!
FCJID!
FCFKJ!
FCGEB!
01023&
L! & 01020&
L!
! FCAIB A
!
FCJJB I
!
FCGBG A
F!
FCDBJ A
!
FCFEI A
!
FCAGE A
!
L ! ! L! L!
! FCHIG!
FCKJE!
FCBII!
FCHAI!
FCGDK!
FCHIF!
L! ! L! L!
! ECFKG A
!
FCEAF I!
FCDEB A
F!
DCHGH A!
FCDHF A!
ACJAJ A!
L! 9,&2!%!
ECAFG!
?FCEIK
@!
! FCFEK!
FCKAH!
FCKKJ!
FCAFB!
FCBII!
FCDBI!
L! & 401002
&
L!
! FCFFJ A
!
FCEDB I!
ACAEJ A
F!
ACEEF A!
FCFAA A!
FCJEG A!
FCJDA A
!
! L! L!
! FCKBH!
FCGGD!
FCJBF!
FCDJH!
FCKAH!
FCBBG!
FCBIJ!
! L! L!
! FCHIA A
!
FCKFF I
!
FCHJJ A
F!
L! FCGEJ A
!
FCIEA A!
L ! ! L! L!
! FCEAI!
FCBAJ!
FCIIK!
L! FCJHJ!
FCJKJ!
L! ! L! L!
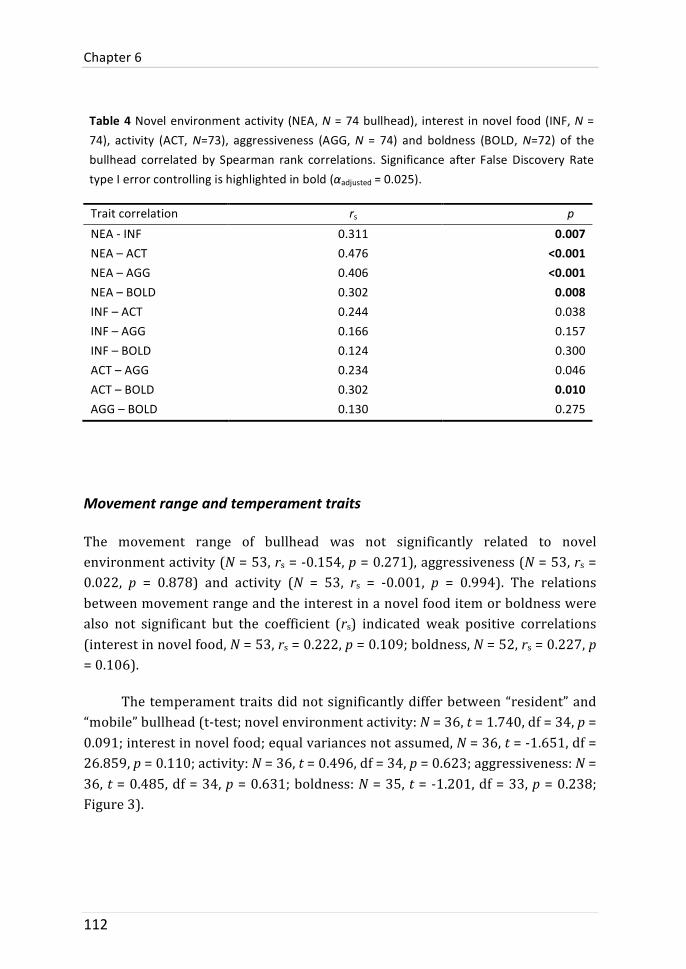
Chapter 6
112
Movement range and temperament traits
The movement range of bullhead was not significantly related to novel environment activity (N = 53, rs = -‐0.154, p = 0.271), aggressiveness (N = 53, rs = 0.022, p = 0.878) and activity (N = 53, rs = -‐0.001, p = 0.994). The relations between movement range and the interest in a novel food item or boldness were also not significant but the coefficient (rs) indicated weak positive correlations (interest in novel food, N = 53, rs = 0.222, p = 0.109; boldness, N = 52, rs = 0.227, p = 0.106).
The temperament traits did not significantly differ between “resident” and “mobile” bullhead (t-‐test; novel environment activity: N = 36, t = 1.740, df = 34, p = 0.091; interest in novel food; equal variances not assumed, N = 36, t = -‐1.651, df = 26.859, p = 0.110; activity: N = 36, t = 0.496, df = 34, p = 0.623; aggressiveness: N = 36, t = 0.485, df = 34, p = 0.631; boldness: N = 35, t = -‐1.201, df = 33, p = 0.238; Figure 3).
Trait correlation rs p NEA -‐ INF 0.311 0.007 NEA – ACT 0.476 <0.001 NEA – AGG 0.406 <0.001 NEA – BOLD 0.302 0.008 INF – ACT 0.244 0.038 INF – AGG 0.166 0.157 INF – BOLD 0.124 0.300 ACT – AGG 0.234 0.046 ACT – BOLD 0.302 0.010 AGG – BOLD 0.130 0.275
Table 4 Novel environment activity (NEA, N = 74 bullhead), interest in novel food (INF, N = 74), activity (ACT, N=73), aggressiveness (AGG, N = 74) and boldness (BOLD, N=72) of the bullhead correlated by Spearman rank correlations. Significance after False Discovery Rate type I error controlling is highlighted in bold (αadjusted = 0.025).
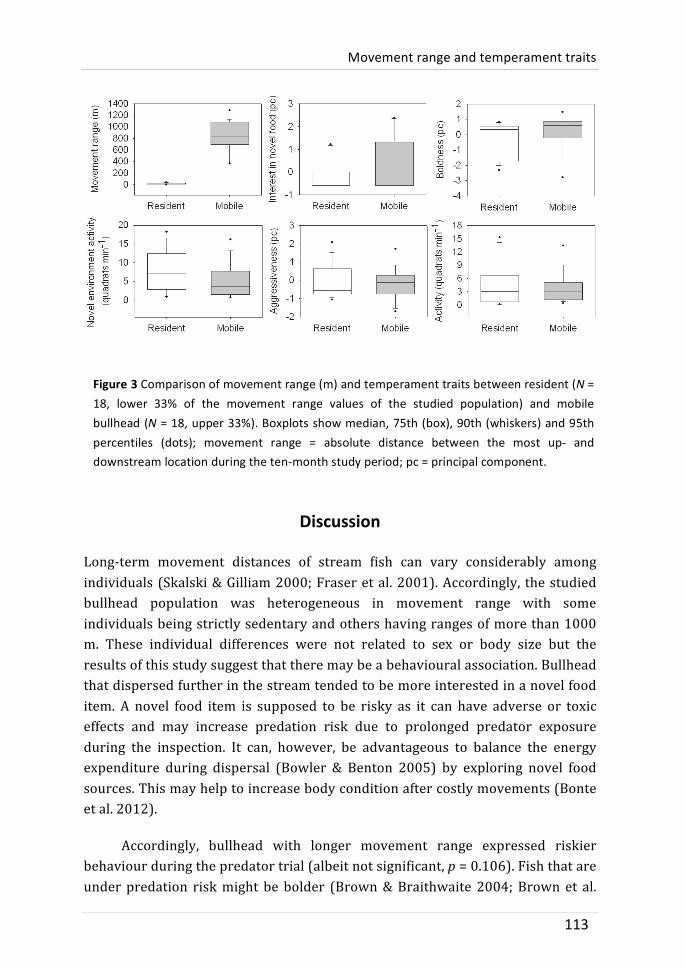
Movement range and temperament traits
113
Discussion
Long-‐term movement distances of stream fish can vary considerably among individuals (Skalski & Gilliam 2000; Fraser et al. 2001). Accordingly, the studied bullhead population was heterogeneous in movement range with some individuals being strictly sedentary and others having ranges of more than 1000 m. These individual differences were not related to sex or body size but the results of this study suggest that there may be a behavioural association. Bullhead that dispersed further in the stream tended to be more interested in a novel food item. A novel food item is supposed to be risky as it can have adverse or toxic effects and may increase predation risk due to prolonged predator exposure during the inspection. It can, however, be advantageous to balance the energy expenditure during dispersal (Bowler & Benton 2005) by exploring novel food sources. This may help to increase body condition after costly movements (Bonte et al. 2012).
Accordingly, bullhead with longer movement range expressed riskier behaviour during the predator trial (albeit not significant, p = 0.106). Fish that are under predation risk might be bolder (Brown & Braithwaite 2004; Brown et al.
Figure 3 Comparison of movement range (m) and temperament traits between resident (N = 18, lower 33% of the movement range values of the studied population) and mobile bullhead (N = 18, upper 33%). Boxplots show median, 75th (box), 90th (whiskers) and 95th percentiles (dots); movement range = absolute distance between the most up-‐ and downstream location during the ten-‐month study period; pc = principal component.

Chapter 6
114
2007). For instance, predator-‐sympatric poeciliids (Brachyraphis episcopi) are bolder than their predator-‐allopatric conspecifics (Brown & Braithwaite 2004; Brown et al. 2007). Therefore, an association of movement range with boldness may be explained by higher predation risk for bullhead with longer movement range (see Howard 1960). The low correlation coefficient (rs = 0.227) may be due to the relatively low predation risk in the studied stream. Aquatic predators were rare and avian predators appeared only a few times. The weak correlations between movement range and interest in novel food or boldness seem to indicate that bullhead with longer movement range may express a more risk-‐prone behaviour. Future studies on the relation between movement range and risk-‐prone behaviour may also consider the level of predation risk in the studied population(s).
We did not find a significant relation between the movement range and activity in a novel environment. Based on the findings of Dingemanse et al. (2003), who showed that juvenile great tit immigrants were faster explorers than locals we had hypothesized that bullhead with longer movement range are more explorative. On the opposite, mobile bullhead tended to be less active in a novel environment than resident bullhead (p = 0.091). And indeed, the relation between dispersal and explorative behaviour may be negative. Myers & Krebs (1971) observed dispersing field voles (Microtus pennsylvanicus and M. ochrogaster) to be less explorative in a maze experiment than residents. This may be explained by individual differences in search strategies. Philopatric or resident individuals may be more intensive and systematic searchers while dispersing or mobile individuals may search less intensively on a small scale. Comparably, Doerr & Doerr (2005) showed that individual treecreeper birds (Climacteris picumnus and Cormobates leucophaeus) that dispersed over greater areas searched less thoroughly suggesting a trade-‐off in search tactic. During reproductive period when the individual movement distances diversified in the bullhead population (Kobler et al. 2012b) “mobile” bullhead may have searched for mates or habitats with a straight-‐line approach (see Zollner & Lima 1999), which resulted in a longer movement range. It is possible that the movement range diversification in bullhead may be related to the individual degree of thoroughness in search tactics.
The movement range of bullhead was not related to individual levels of aggressiveness. Due to territorial behaviour of Cottus spp. even outside breeding-‐period (e.g. C. bairdii, Petty & Grossman 2007) it is plausible that mobile bullhead are more involved in aggressive interactions with conspecifics after movement range expansion and settlement. Duckworth & Badyaev (2007) observed

Movement range and temperament traits
115
philopatric western bluebirds to be less aggressive than their colonizing conspecifics. The highly aggressive individuals appeared to be better suited for colonization because of their ability to exclude less aggressive conspecifics from new territories (Duckworth & Badyaev 2007). We assumed that despite a possibly long time interval between the movement range expansion and the temperament traits tests in the present study (up to several months) a relation of movement range and aggressiveness would have been detectable. Meylan et al. (2009) showed that ten months after the dispersal phase common lizards (Lacerta vivipara) still showed behavioural differences between dispersing and resident individuals. However, it is possible that the length of movement range is not a good indication of the number of territory establishments. Individuals with short and long movement range may establish similar amounts of territories but on a different spatial scale. Future studies may examine whether the movement range (dispersal distance) and territory establishments of non-‐migratory stream fish are related. Furthermore, it may be tested whether the number of territory establishments is related to aggressiveness. The degree of habitat complexity of the territory may also be related to aggressiveness (see Kobler et al. 2011).
Bullhead that were more active in the novel environment in the laboratory were significantly smaller. This was already shown in a previous independent study (albeit not significant, Kobler et al. 2009b). Why smaller individuals explore faster can be explained by the hypothesis that smaller fish have higher metabolic requirements and locomotion is more costly than for larger fish (Krause et al. 1998). Similarly, Brown & Braithwaite (2004) showed a positive correlation of body size and time to emerge from a shelter. It may be concluded that an individual’s exploration of a novel environment is linked to its metabolism (see Careau et al. 2008).
Although the individual differences in movement range of bullhead were remarkable, we did not find a clear grouping of resident and mobile individuals as has been suggested in other studies in non-‐migratory stream fish (e.g. Belica & Rahel 2008) and also in the bullhead (Knaepkens et al. 2004). This may be due to biological but also methodological differences. Distinct groupings of mobility are often an artefact of data collection and the methodological definition when an individual is mobile (Porter & Dooley Jr. 1993; Gowan et al. 1994). Many studies define resident fish by those not leaving a stream area of a given length (e.g. 20 m stream length, sensu Gowan et al. 1994) or an estimated home range (Gerking 1959). Our approach of following individual fish over large stream areas with multiple localizations and quantified movement distances affirmed that the distribution of movement distances is rather leptokurtic than bimodal (Skalski &

Chapter 6
116
Gilliam 2000; Fraser et al. 2001). The conceptual grouping of non-‐migratory stream fish into resident and mobile individuals thus may be reconsidered.
To conclude, we found considerable intra-‐population heterogeneity in the movement range of bullhead. The individual movement range was not related to body size or sex. We found weak positive correlations between movement range and interest in novel food or boldness. Bullhead that expanded further in the stream seemed to be more risk-‐prone. Smaller bullhead were more active in a novel environment. This may be due to higher metabolic requirements of smaller fish. Future studies may also address the question whether the individual movement range or the individual dispersal distance of non-‐migratory stream fish is related to behavioural characteristics. It may also be tested whether individual differences in temperament traits are the cause or the consequence of heterogeneity in movement ranges.
Acknowledgments
Many thanks to F. Volckaert, S. Lahaye and A. Geens for helpful comments, P. Scheys, G. Eens, Y. Humblet and K. Geudens for technical assistance and G. Knaepkens for support. Many thanks also to the University of Antwerp and the Fund for Scientific Research-‐Flanders for funding (FWO-‐project nr G.0119.08N). The experimental work was performed in agreement with the Belgian and Flemish laws and was approved by the ethical committee of the University of Antwerp (ID number 2007/42).

CHAPTER 7
MOVEMENT RANGE, FITNESS AND HETEROZYGOSITY


Movement range, fitness and heterozygosity
119
Dispersal in a heterogeneous stream population: is movement
range of bullhead related to fitness and heterozygosity?
Alexander Kobler, Filip A.M. Volckaert, Alexander Triantafyllidis, Marcel Eens, Gregory E. Maes
In preparation
Abstract
Variation in individual dispersal propensity can depend on differences in life-‐history traits such as developmental state, size and sex but may also be associated to fitness. We examined whether the movement range, that is the distance between the most up-‐ and downstream location, of a non-‐migratory stream fish is associated to fitness-‐related traits and individual genetic diversity. Fifty bullhead (Cottus perifretum) were tagged with passive integrated transponders, genotyped, sexed, field-‐observed during ten months and recaptured. The individual movement range varied from 1 to 1096 meters. These differences were not related to body condition, growth rate or microsatellite heterozygosity (single-‐ or multilocus) and also not to sex or body size. No association between heterozygosity and body condition or growth rate was found. A possible reason why body condition was not associated with movement range may be counterbalancing effects of strong intra-‐specific competition and low predation risk during dispersal. Growth rate measurements over the entire study period were not suitable to differentiate a growth-‐association before dispersal from a possible growth benefit after dispersal. Instead of using neutral genetic variation a candidate-‐gene approach with genes associated to dispersal or temperament traits might help to detect the evolutionary background of between-‐individual differences in the movement range (dispersal) of non-‐migratory stream fish.

Chapter 7
120
Introduction
Dispersal is the active or passive attempt to move from one natal / breeding site to another breeding site (Clobert et al. 2001; Clobert et al. 2009). Dispersal allows the exploitation of spatially and temporally variable resources and is essential for the persistence of a species (Bohonak 1999; Bowler & Benton 2005; Clobert et al. 2009). The importance of dispersal behaviour for the ecology and evolution of wild populations becomes increasingly apparent as populations face fragmentation of their habitat (Clobert et al. 2009). Successful management of a species, therefore, relies on an understanding of the patterns and dynamics of dispersal.
Animal dispersal might be linked to competition with conspecifics and kin, and competition for mates and quality habitats (Clobert et al. 2001; Clobert et al. 2009). Individuals that are less successful during competitive interactions should preferentially disperse and tend to settle farther away, a mechanism that is recognized as “fitness-‐associated dispersal” (Hadany et al. 2004 and references therein). The pressures to disperse are, therefore, unequally distributed among individuals (Bowler & Benton 2005) and dispersing individuals rarely constitute a random sample of the population (Hadany et al. 2004). The variation in individual dispersal propensity might depend on differences in life-‐history traits such as developmental state, size and sex (Bowler & Benton 2005 and references therein). For instance, in many birds, territory acquisition by a young adult may only be possible through dispersal (Bowler & Benton 2005). The trait dispersal may have also a genetic component because fitness-‐associated traits can be related to individual genetic diversity (David 1998; Chapman et al. 2009; Szulkin et al. 2010). For instance, heterozygosity in mountain goats (Oreamnos americanus) is associated to increased survival (Mainguy et al. 2009) and a lower propensity to disperse (Shafer et al. 2011).
Individual propensity to disperse is hard to assess due to the difficulties in following individuals over a long period and in gathering information on their breeding locations. Non-‐migrating stream fish are no exception. Although movement distances are often very restricted (Gerking 1959), movement distribution within populations may be heterogeneous (e.g. Skalski & Gilliam 2000; Fraser et al. 2001) and numerous studies differentiated non-‐migratory stream fish populations into resident and mobile individuals (e.g. Smithson & Johnston 1999; Nakamura et al. 2002; Rodriguez 2002; Knaepkens et al. 2004). The mobile fraction influences the level of inbreeding through the spread of new genes (Howard 1960) and the repopulation of depauperated areas (Gerking

Movement range, fitness and heterozygosity
121
1959). However, not much is known which individual characteristics distinguish resident and mobile fish.
The studied species, the bullhead (Cottus perifreum), belongs to the genus Cottus that consists of small benthic fishes (< 20 cm, Kottelat & Freyhof 2007), known for their benthic and cryptic behaviour (e.g. Smyly 1957). Cottus ssp. are also known for their non-‐breeding territoriality (Cottus bairdii, Petty & Grossman 2007). Nevertheless, numerous studies found remarkable intra-‐population heterogeneity in movement distances (C. perifretum, Knaepkens et al. 2004, 2005; Cottus ssp., Breen et al. 2009; Hudy & Shiflet 2009; Ovidio et al. 2009). Whereas some individuals stay over the year in a restricted area often not larger than a few square meters, other individuals move more than one kilometre (Hudy & Shiflet 2009; see also Knaepkens et al. 2004; Breen et al. 2009; Kobler et al. 2012). The studied population density was high (Kobler et al. 2011) and intra-‐specific competition presumably strong.
The aim of the study is to examine movement range of a non-‐migratory stream fish, the bullhead, in the context of the phenotype and genetic diversity using a single population in a small lowland stream in Flanders, Belgium. The breeding sites of stream fish are hard to identify; therefore the existing literature commonly uses long-‐term movement range (the distance between the most up-‐ and downstream location during several months) to indicate dispersal distance. Consequently, rather than using the terms philopatric/dispersing individuals non-‐migratory stream fish are differentiated into resident and mobile individuals.
The present study examined whether individual movement range is positively associated to fitness, similar to what has been observed for the dispersal distances of other vertebrates (Gaines & McClenaghan Jr. 1980; Greenwood 1980; Belichon et al. 1996; Alonso et al. 1998; Hadany et al. 2004). For instance, better-‐conditioned, juvenile great bustards (Aves: Otis tarda) are competitively superior and stay closer to their natal sites (Alonso et al. 1998). Such dispersal-‐fitness-‐association might have a genetic cause; individual heterozygosity can be positively associated with fitness components in a broad range of taxa (David 1998; Chapman et al. 2009; Szulkin et al. 2010). To test our hypothesis, two common indicators of fitness in fish, body condition (e.g. Bolger & Connolly 1989; Jakob et al. 1996) and specific growth rate (e.g. Werner & Hall 1976), as well as microsatellite heterozygosity were related to individual movement range of bullhead.

Chapter 7
122
Materials and methods
Sampling
The study was conducted in the Laarse Beek (51°16'28.40"N; 4°29'4.63"E), a lowland stream in northern Belgium with a mean width of 3 m (range 2 -‐ 4 m) and a mean depth of 0.3 m (0.1 -‐ 0.5 m). Bullhead were the most abundant fish species in the stream. Aquatic and avian predators were rare. A more detailed description of the study area is given in (Kobler et al. 2011). The focal stream reach was 2500 m long and marked every 2 m with consecutively numbered wooden poles along one side of the stream.
In the middle of the 2500 m observation reach bullhead were initially caught within 324 m in two sessions, on 4, 5 and 7 February and on 13 and 14 February 2008 by electro-‐fishing (WFC7-‐10, Electracatch Int., Wolverhampton, UK) using a 40 cm ring anode. Only adult bullhead > 50 mm total length (TL) were collected and kept individually in buckets. Fifty mm is the minimum size for tagging C. perifretum (see further; Knaepkens et al. 2007). Bullhead (N = 334) had a mean (± SD) TL of 62.1 ± 6.3 mm (range 50 to 88 mm) and a mean wet weight of 3.3 ± 1.2 g (range 1.7 to 9.9 g). Fourteen recaptures during the second session resulted in an abundance estimate of 2006 bullhead > 50 mm TL (confidence interval 1228–3459) in the 324 m electro-‐fishing reach (Kobler et al. 2011). Thus, the captured bullhead represented ~17% of the mature population in the electro-‐fishing reach. In comparison to other bodies of waters in Flanders the abundance of C. perifretum was very high.
Bullhead were anaesthetized (0.25 ml L-‐1 2-‐phenoxy-‐ethanol) and individually coded 12 mm PIT-‐tags (12 x 2.1 mm, 0.094 g; ID100, EID Aalten, NL) were inserted horizontally into the body captivity through a 2 mm incision close to the genital papillae (Kobler et al. 2011). Every tagged individual was photographed dorsally with a scale. A small piece of tissue from the anal fin of each tagged bullhead was sampled for genetic analyses and preserved in 95% ethanol. After implantation and recovery in buckets with stream water, bullhead were released at their capture site.
The gender of bullhead was determined morphologically by evaluating the individual photographs. At the time of first capture (and photographing), seasonal gonadal maturation had already started. Individuals with a very round belly (looking tadpole-‐like), that clearly suggested the content of eggs, were determined as females. Males could be determined by the triangular shape of the

Movement range, fitness and heterozygosity
123
head (Freyhof et al. 2005) and the proportionally smaller body cavity. Ambiguity about the sex of an individual led to a non-‐defined gender of the individual. The sex of every individual was determined “blindly” three times on three different days by evaluating the photograph of an individual. Only when a consensus between three sex determinations was reached was the gender used in this study. The majority of bullhead (83%) was sexed and included 138 females and 139 males.
Tracking and recapture
The stream was scanned monthly over a distance of 2500 m: 1250 m up-‐ and downstream of the middle of the electro-‐fishing reach. This tracking distance should allow to cover the longest movements of bullhead (observed maximum distance of a Cottus ssp.: Potomac sculpin, Cottus girardi, 1711 m in 75 d, Hudy & Shiflet 2009). Tracking started two weeks after the second electro-‐fishing session and was conducted monthly on 28 February, 27 March, 24 April, 21 May, 25 June, 16 July, 13 August and 9 September 2008. Tracking after 9 September 2008 was conducted during the individual recapture (see further). The period covered the reproductive period (March to May), when farthest annual movement distances were recorded in 2008 in the Laarse Beek (Kobler et al. 2012).
The duration of a 2500 m tracking was three days with an effort of ~8 h d-‐1. A portable PIT-‐tag antenna with a detection precision of < 30 cm was used (Kobler et al. 2011, 2012). Once a bullhead was located, the individual PIT-‐tag code, the stream length position (m) and the tracking date were noted (for more details see Kobler et al. 2011, 2012). During reporting, the antenna was kept above the detected bullhead. An escape response was never observed and the tagged individuals relied on their camouflage, a typical behaviour for Cottus spp. (for more details see Kobler et al. 2011).
From 22 September to 19 November 2008 we attempted to recapture the PIT-‐tagged bullhead within the tracking reach. This period was chosen as bullhead were in the following tested for animal personality differences in an accompanying study (Kobler et al. in preparation, chapter 6) and this had to be done before the annual gonadal maturation started. The fish were detected in two complete screenings of the 2500 m stretch (in total 8 d) by using the portable PIT-‐tag antenna. Fifty-‐nine bullhead were caught with a large landing net (one person) and two small aquarium nets (another person). The relatively low recapture rate (18%) may be explained by natural mortality and not emigration (Kobler et al. 2012). Bullhead are short-‐lived often not getting older than four

Chapter 7
124
years (Marconato et al. 1993; Kottelat & Freyhof 2007) and the youngest tagged individuals of the present study were almost two years old (in the Laarse Beek, 0+ aged bullhead are maximum 45 mm TL long, personal observations).
DNA isolation and genotyping
Genomic DNA was extracted using a commercial kit (NucleoSpin, Macherey-‐Nagel, Düren, Germany). Nine microsatellite loci were amplified in a GeneAmp PCR System 2700 thermocycler (Applied Biosystems, Foster City, CA, USA) in two multiplex PCRs using the QIAGEN Multiplex PCR Kit (Qiagen N.V., Venlo, NL). Annealing temperatures were 55° C for Cgo56, Cgo91, Cgo1016, Cgo1033, Cgo1114 (Englbrecht et al. 1999) and 60° C for CottE6, CottE23, CottES21, LCE22, LCE59 (Nolte et al. 2005). PCR products were visualised on an ABI 3130 Genetic Analyser (Applied Biosystems). Allele size was determined by means of an internal Genescan 500-‐LIZ size standard (Applied Biosystems) and genotypes were obtained using GENEMAPPER version 3.7 (Applied Biosystems). To identify potential genotyping errors in the microsatellite data (i.e. stuttering, large allele dropout or null alleles), the software Microchecker version 2.2.3 (Van Oosterhout et al. 2006) was used. One locus (CottE23) was not in Hardy-‐Weinberg equilibrium and therefore excluded from further analysis (calculated by FSTAT version 2.9.3.2, Goudet 1995). For all genetic analyses we used only individuals for which at least seven of the nine loci were successfully amplified (N = 310 bullhead).
The number of alleles, the allele frequency per locus, the expected heterozygosity (HE) and the observed heterozygosity (HO) were calculated using the microsatellite toolkit for Excel 3.1.1 (Park 2001). Calculation of the inbreeding coefficient (FIS) for each locus was performed in the program FSTAT version 2.9.3.2 (Goudet 1995).
Individual diversity
Two common measures of diversity were used (Chapman et al. 2009): individual heterozygosity (HI) and internal relatedness (IR). HI measures the proportion of heterozygous typed loci for an individual. IR is centered around zero for individuals born to “unrelated” parents, with negative values suggesting relatively “outbred” individuals and high positive values being suggestive of inbreeding (Amos et al. 2001).

Movement range, fitness and heterozygosity
125
IR was calculated in Storm 1.1 (Frasier 2008).
2! − !!2! − !!
where H is the number of loci that are homozygous, N is the number of loci and fi is the frequency of the ith allele in the genotype.
HI (non-‐normally distributed, Kolmogorov-‐Smirnov, p < 0.05) and IR were strongly correlated (Spearman correlation, N = 310, rs = -‐0.971, p < 0.001).
Fitness indicators
Body condition was calculated separately for males and females because of seasonal gonadal development before capture and the associated inter-‐sexual differences in body shape (see Sampling). As proposed by (Bolger & Connolly 1989) we related the actual weight of an individual to its expected weight, which was calculated as a function of its length. Therefore, (unstandardized) residuals of the linear regression of wet weight (y-‐axis) on body size (TL, x-‐axis) for males and females were calculated as proposed by (Jakob et al. 1996; Schulte-‐Hostedde et al. 2005). A residual value was assigned to every individual; positive values meant that the individual was heavier than what was expected from the length-‐weight function of all sampled males or females. Residual values were normally distributed (Kolmogorov-‐Smirnov, N = 277, p = 0.200).
Since body growth in fish is an indicator of individual fitness at low predation risk (sensu Werner & Hall 1976; Mangel & Stamps 2001), specific growth rate (µ, Fausch 1984) was also used as fitness indicator.
where TL1 is the final total length (mm), TL0 is the initial total length (mm) and t is the growth period (d).
Movement range
As quantitative measure of dispersal distance we used the individual movement range (m) of each bullhead. Movement range was calculated with the absolute distance of the most up-‐ and downstream location of an individual during the
tTLTL 01 lnln −
=µ

Chapter 7
126
entire ten-‐month study period (Kobler et al. 2012). An accompanying study showed that significantly longer movement distances were recorded during the annual spawning period and an association to reproductive activities was likely (Kobler et al. 2012). After long distance movements, bullhead showed again their typical sedentary behaviour (Kobler et al. 2012). Therefore, movement range was used as an indicator of individual breeding dispersal distance (breeding dispersal: attempt to move from a breeding site to another breeding site, (Clobert et al. 2009).
Statistics
Only individuals that were genotyped, sexed and recaptured (N = 50) were used for movement range analyses. The relation between movement range (dependent variable), individual genetic diversity (HI or IR) and body condition was assessed with an ANCOVA (one model including HI and another including IR). Sex (variance homogeneity, Levene-‐test, p > 0.05) and body size (TL, mm) were also included in the model. The interactions of heteroyzgosity × sex, heterozygosity × length and sex × length were also tested. Most non-‐significant variables or interactions were excluded stepwise. Whether HI and IR of the “movement range individuals” (non-‐normally distribution, Kolmogorov-‐Smirnov, p < 0.05) were related to body condition was assessed with Spearman rank correlations.
We also tested whether single locus heterozygosity was associated with movement range and body condition. Eight polymorphic loci were tested individually in eight independent t-‐tests for movement range and eight tests for body condition. The groups were defined as homozygote or heterozygote. For 15 tests equal variances were assumed (Levene-‐tests, all p > 0.05). The variance of the locus CottES21 were heterogeneous for movement range and equal variances not assumed in the test.
Whether specific growth rate (µ, dependent variable) was related to movement range, individual genetic diversity (HI or IR) or body condition was tested with an ANCOVA (one model including HI and another including IR). This calculation was chosen as fish length-‐growth differs at different body sizes and the model was run with TL (mm) as controlling covariate. Because the variable µ consisted of values < 0.0001 we multiplied it by 1000. This facilitated the presentation of the Type III sum of squares and the parameter estimates (± SE) but did not change the F-‐ and p-‐values of the model. Sex (variance homogeneity, Levene-‐test, p > 0.05) and the interactions individual diversity × sex and

Movement range, fitness and heterozygosity
127
movement range × body condition were included in the model. Most non-‐significant variables or interactions between them were stepwise excluded.
The effect of single locus heterozygosity on specific growth rate was tested with eight polymorphic loci. Each locus was included separately as a factor substituting the covariate heterozygosity (HI or IR) in the previously mentioned growth rate ANCOVA. This resulted in eight ANCOVAs (variance homogeneity of all loci, Levene-‐tests, all p > 0.05).
Additionally, only the lower and higher 33% the of movement range values were included in the calculations. This was done to exclude individuals with intermediate movement range distances and put emphasis on the individuals with distinct distances, similar to a categorization into resident and mobile individuals. One-‐way ANOVAs with body condition, HI or IR as dependent variables and an ANCOVA with growth rate as dependent variable were run with the factor “mobility” (“resident” and “mobile”). For the body condition ANOVA, the variance between the factor levels was heterogeneous (Levene-‐test, p < 0.05); a Welch one-‐way ANOVA was conducted in this case.
All calculations (unless indicated differently) were conducted in PASW Statistics 18.0 for Mac (SPSS Inc., Chicago, IL, USA). Significance was assessed at α < 0.05.
Results
Genetic characteristics of the population
A total of 310 individuals were successfully genotyped from the originally 334 tagged bullhead. The mean (± SD) number of alleles per locus was 2.44 ± 0.73 (Table 1). Allele (N = 22) frequency ranged from 0.16% to 100% (40.91 ± 25.54%). Ten pairs of individuals had an identical multilocus genotype (based on eight polymorphic loci). The low number of alleles and the identical multilocus genotypes indicate only little genetic variability in the population. However, a mean HE over all loci of 0.44 ± 0.06 and a mean HO was 0.44 ± 0.01 did not indicate inbreeding (mean FIS = -‐0.007, Table 1). Also the internal relatedness (IR) = -‐0.0026 did not indicate inbreeding nor outbreeding.
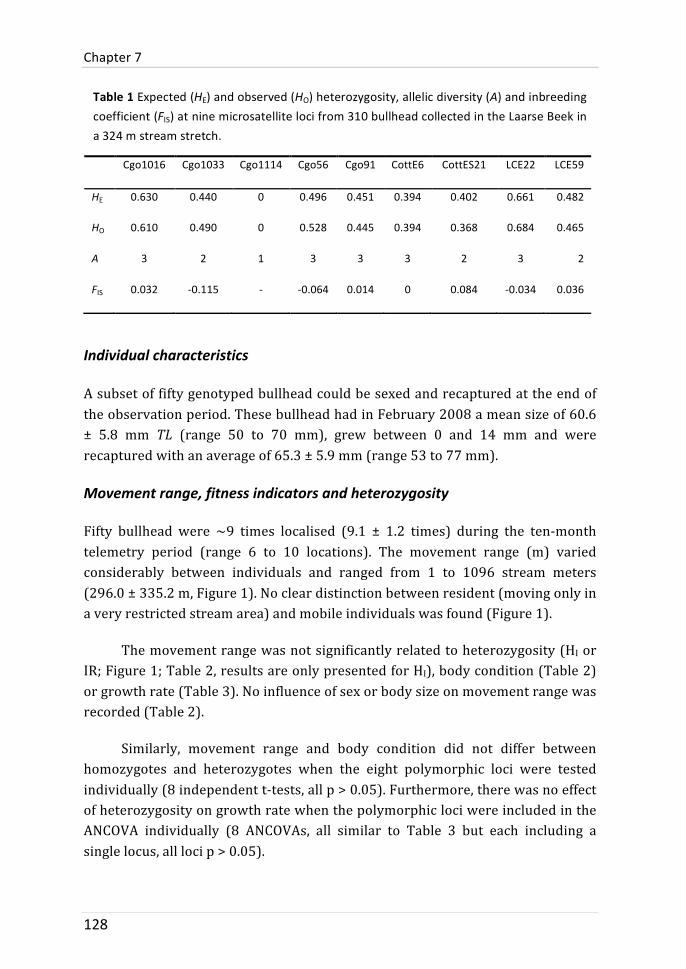
Chapter 7
128
Individual characteristics
A subset of fifty genotyped bullhead could be sexed and recaptured at the end of the observation period. These bullhead had in February 2008 a mean size of 60.6 ± 5.8 mm TL (range 50 to 70 mm), grew between 0 and 14 mm and were recaptured with an average of 65.3 ± 5.9 mm (range 53 to 77 mm).
Movement range, fitness indicators and heterozygosity
Fifty bullhead were ∼9 times localised (9.1 ± 1.2 times) during the ten-‐month telemetry period (range 6 to 10 locations). The movement range (m) varied considerably between individuals and ranged from 1 to 1096 stream meters (296.0 ± 335.2 m, Figure 1). No clear distinction between resident (moving only in a very restricted stream area) and mobile individuals was found (Figure 1).
The movement range was not significantly related to heterozygosity (HI or IR; Figure 1; Table 2, results are only presented for HI), body condition (Table 2) or growth rate (Table 3). No influence of sex or body size on movement range was recorded (Table 2).
Similarly, movement range and body condition did not differ between homozygotes and heterozygotes when the eight polymorphic loci were tested individually (8 independent t-‐tests, all p > 0.05). Furthermore, there was no effect of heterozygosity on growth rate when the polymorphic loci were included in the ANCOVA individually (8 ANCOVAs, all similar to Table 3 but each including a single locus, all loci p > 0.05).
Cgo1016 Cgo1033 Cgo1114 Cgo56 Cgo91 CottE6 CottES21 LCE22 LCE59
HE 0.630 0.440 0 0.496 0.451 0.394 0.402 0.661 0.482
HO 0.610 0.490 0 0.528 0.445 0.394 0.368 0.684 0.465
A 3 2 1 3 3 3 2 3 2
FIS 0.032 -‐0.115 -‐ -‐0.064 0.014 0 0.084 -‐0.034 0.036
Table 1 Expected (HE) and observed (HO) heterozygosity, allelic diversity (A) and inbreeding coefficient (FIS) at nine microsatellite loci from 310 bullhead collected in the Laarse Beek in a 324 m stream stretch.
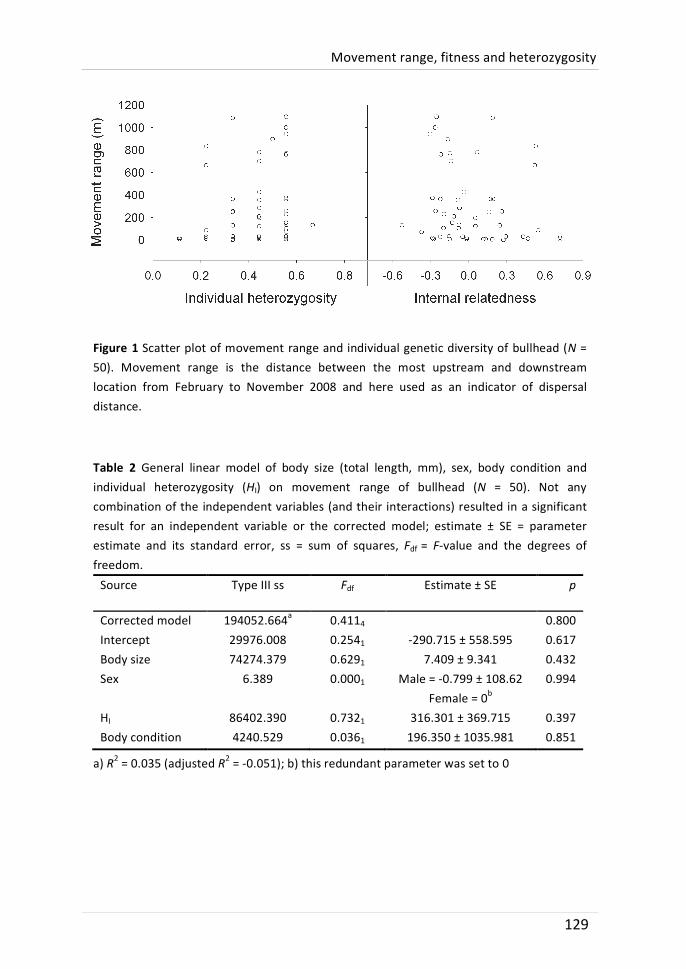
Movement range, fitness and heterozygosity
129
Source Type III ss Fdf Estimate ± SE p
Corrected model Intercept
194052.664a
29976.008 0.4114 0.2541
-‐290.715 ± 558.595
0.800 0.617
Body size Sex HI
Body condition
74274.379 6.389
86402.390 4240.529
0.6291 0.0001
0.7321 0.0361
7.409 ± 9.341 Male = -‐0.799 ± 108.62
Female = 0b
316.301 ± 369.715 196.350 ± 1035.981
0.432 0.994
0.397 0.851
a) R2 = 0.035 (adjusted R2 = -‐0.051); b) this redundant parameter was set to 0
Figure 1 Scatter plot of movement range and individual genetic diversity of bullhead (N = 50). Movement range is the distance between the most upstream and downstream location from February to November 2008 and here used as an indicator of dispersal distance.
Table 2 General linear model of body size (total length, mm), sex, body condition and individual heterozygosity (HI) on movement range of bullhead (N = 50). Not any combination of the independent variables (and their interactions) resulted in a significant result for an independent variable or the corrected model; estimate ± SE = parameter estimate and its standard error, ss = sum of squares, Fdf = F-‐value and the degrees of freedom.
Chapter 7
130
Statistically differentiated “resident” (lower 33% of movement range values, range 1 to 33 m, 16.6 ± 10.4 m) and “mobile” individuals (upper 33%, range 364 to 1096 m, 714.8 ± 263.7 m) were also compared. There was no significant difference in HI (ANOVA, between groups ss = 0.032, F = 1.657, p = 0.207), IR (ANOVA, ss = 0.186, F = 2.083, p = 0.159) and body condition (Welch ANOVA, Stat = 0.006, df2 = 26.898, p = 0.941) between “resident” and “mobile” bullhead (see also Figure 1). Similarly, this categorized mobility was also not a significant factor on specific growth rate (in a similar ANCOVA than in Table 3; corrected model: Fdf = 5.3684, p = 0.002, mobility: Fdf = 0.4981, p = 0.486).
Fitness indicators and heterozygosity
Growth rate was positively related to body condition (p = 0.003) in males and females (Table 3, no significant interaction of body condition and sex). No relation was found between growth rate and heterozygostiy (HI or IR; Table 3; results are only presented for HI). As expected, growth rate was negatively related to body size (p < 0.001). Male bullhead grew faster than females (p = 0.005).
Body condition was not significantly correlated to heterozygosity in males (Spearman, N = 21; HI: rs = -‐0.218, p = 0.343; IR: rs = 0.156, p = 0.499) and females (N = 29; HI: rs = 0.081, p = 0.677; IR: rs = -‐0.082, p = 0.672).
Source Type III ss Fdf Estimate ± SE p Corrected model 0.905a 9.0153 < 0.001 Intercept 0.822 24.5581 1.416 ± 0.292 < 0.001 Body size Condition
0.525 0.324
15.6751 9.6651
-‐0.020 ± 0.005 1.715 ± 0.552
< 0.001 0.003
Sex 0.296 8.8521 Male = 0.171 ± 0.058 Female = 0b
0.005
a) R2 = 0.370 (adjusted R2 = 0.329); b) this redundant parameter was set to 0
Table 3 General linear model of body size (total length, mm; controlling variable), body condition and sex on specific growth rate (µ) of bullhead (N = 50) during the study period (4 February to 19 November 2008). The movement range, the heterozygosity variables and their interactions were not significantly related to the specific growth rate and stepwise excluded; estimate ± SE = parameter estimate and its standard error, ss = sum of squares, Fdf = F-‐value and the degrees of freedom.

Movement range, fitness and heterozygosity
131
Discussion
The movement range during the ten-‐month observation period varied between individuals; individuals with restricted movement ranges and individuals with movement ranges > 1 km were observed. We did not detect a distinct grouping of resident and mobile individuals (Figure 1) as has been observed in non-‐migratory stream fish including bullhead (e.g. Smithson & Johnston 1999; Nakamura et al. 2002; Rodriguez 2002; Knaepkens et al. 2004). This may be due to biological but also methodological differences as many studies define resident fish by those not leaving a stream area of a given length (e.g. 20 m stream length, sensu Gowan et al. 1994). Our approach of following individual fish over large stream areas with multiple localizations and quantified movement distances affirmed that the distribution of movement distances is rather leptokurtic than bimodal (Skalski & Gilliam 2000; Fraser et al. 2001). Nevertheless, the between-‐individual differences in bullhead movement range were remarkable and a relation to individual characteristics or fitness differences seemed plausible.
The hypothesis that resident individuals can be distinguished from mobile individuals by a higher level of “fitness” was not confirmed. First, there was no relation between movement range and body condition. The measurement of fish body condition is used as an appropriate indicator for “well-‐being or fitness” (Bolger & Connolly 1989). Under strong intra-‐specific competition, body condition may be negatively associated with dispersal propensity (Hadany et al. 2004; Bonte & De La Pena 2009). Bullhead abundance was high and intra-‐specific competition presumably strong (Kobler et al. 2011). However, when the risk of predation during dispersal is low and fluctuations in environmental conditions are spatiotemporally unpredictable body condition can be positively associated to dispersal propensity (Bonte & De La Pena 2009). Under these conditions, individuals with better body condition are more likely to disperse and favoured during the potentially energetically expensive dispersal (Meylan et al. 2002; Barbraud et al. 2003; Cote & Clobert 2007). For bullhead, the risk of mortality during dispersal was presumably low as aquatic and avian predators were rare (Kobler et al. 2011) and distances relatively short. The conditions in the studied ecosystem thus favour both negative and positive associations of body condition and dispersal propensity. Several study systems with different levels of intra-‐specific competition and mortality risk during dispersal may help to disentangle their probably opposite effects on the relationship between body condition and movement range.

Chapter 7
132
Second, we hypothesized that fish with slower growth rate and, therefore, a lower level of fitness would preferentially disperse. Our results do not support this hypothesis. However, we did not measure growth rate at the moment of most dispersal activity (which mostly occurred during springtime, Kobler et al. 2012). Instead, our measurements include growth before, during and after dispersal. Therefore, we cannot disentangle whether dispersal was negatively associated to growth before the dispersal (sensu Belichon et al. 1996; Hadany et al. 2004) or, on the other hand, individuals might have benefitted from dispersal by a higher growth rate after the dispersal event (Belichon et al. 1996; Lowe 2010). For example, in anadromous salmonids (Pisces: Salmonidae) migrating individuals benefit from faster growth but suffer from greater mortality risk (reviewed by Jonsson & Jonsson 1993). A similar growth-‐mortality trade-‐off may also exist in non-‐migratory stream fish and may be tested in future research.
Individual genetic diversity was neither related to body condition, growth rate, movement range nor different between (statistically distinguished) “resident” and “mobile” bullhead. The relationship between individual genetic diversity and fitness-‐related traits has become known as heterozygosity-‐fitness correlation (HFC) and has been reported in populations of many species (David 1998; Chapman et al. 2009; Szulkin et al. 2010). Two main hypotheses prevail for multilocus HFCs, the functional overdominance and the associative overdominace (local and general effect) hypothesis (reviewed by Hansson & Westerberg 2002). There was no indication of inbreeding in the bullhead population and functional overdominance (wide-‐scale genomic heterozygosity that may be linked to inbreeding, Hansson & Westerberg 2002) less likely. Associative overdominance (heterozygote advantage as a result of genetic associations between the neutral marker loci and the loci under selection, reviewed by Hansson & Westerberg 2002) seemed more probable. Low allelic diversity, ten pairs of individuals with identical multilocus genotype but a relatively high abundance of bullhead may indicate a recently bottlenecked and expanded population. The local effect hypothesis is expected under such circumstances and predicts a heterozygote advantage at the markers as a result of effects of homozygosity at closely linked fitness loci (reviewed by Hansson & Westerberg 2002). Positive and negative HFCs of single microsatellite loci may be blurred in analyses of the multilocus effect (Lieutenant-‐Gosselin & Bernatchez 2006). However, our analysis with single loci did not reveal a significant difference of body condition and growth rate between homozygotes and heterozygotes. The markers used were randomly distributed and the proximity to genes that are coupled with growth rather unlikely; allozyme loci involved in metabolic energy pathways can be more

Movement range, fitness and heterozygosity
133
suitable (Pujolar et al. 2009). But clearly, HFCs can also vary across species, populations, temporal samples and sexes (Pujolar et al. 2006). For instance, it has been shown that growth rate of 12-‐month old European eel (Anguilla anguilla) is positively linked to individual genetic diversity (allozyme loci heterozygosity) but this relationship vanishes in 22-‐month old eel (Pujolar et al. 2006).
While HFC studies that measure fitness with body condition or growth rate are numerous (Chapman et al. 2009), only very few studies related dispersal to heterozygosity (Selonen & Hanski 2010; Shafer et al. 2011). This relation can be positive or negative. For instance, Shafer et al. (2011) showed that resident mountain goats have a higher multi-‐locus heterozygosity. They hypothesize that this is based on a higher level of competitive abilities of more heterozygote individuals. Individuals with lower heterozygosity should, therefore, preferentially disperse (Shafer et al. 2011). On the other hand, inbreeding may shorten dispersal distances (Szulkin & Sheldon 2008) and individual genetic variability may be linked to dispersal distances. For example, the dispersal distance of Siberian flying squirrels (Pteromys volans) is positively correlated with heterozygosity (Selonen & Hanski 2010). This correlation was mainly driven by one out of seven microsatellites, which indicates an associative overdominance effect (Selonen & Hanski 2010). The markers used in the present study did not indicate multi-‐ or single-‐locus heterozygosity-‐dispersal correlations. Genetic maps based on microsatellites that are linked to a physical map of a model species my help to initiate future quantitative trait loci (QTL) studies in Cottus ssp. (Stemshorn et al. 2005) and help to find local effects of individual genetic diversity on the individual propensity to disperse.
The variation in movement range was not body size-‐dependent probably because exclusively adult bullhead in a narrow body size range (50 to 70 mm TL) were observed. It is known that individual differences in developmental state or size are related to competitive abilities which can have an impact on dispersal distances (Bowler & Benton 2005). In some stream fish smaller individuals tend to move further away, which might confirm the hypothesis that movement range can be driven by competitive interactions (Gowan & Fausch 1996; Hughes 2000; Petty & Grossman 2004).
The movement range of bullhead was not sex-‐biased. Inter-‐sexual differences in reproductive behaviour are a possible source for sex-‐biased dispersal. This is well-‐known in birds and mammals (e.g. Greenwood 1980) but only very few studies examined sex-‐biased dispersal in fish (Pardini et al. 2001; Hutchings & Gerber 2002). Because of female mate choice and male parental care

Chapter 7
134
in bullhead (Bisazza & Marconato 1988) it is plausible that females disperse farther than males. This differentiation seems particularly plausible during reproductive period (Kobler et al. 2012). Indeed, movement distances may diversify between the genders during breeding period (Croft et al. 2003) but equalize during the remainder of the annual period due to movements that are not directly linked to the reproduction (Kobler et al. 2012). A shorter time scale (e.g. monthly) as well as an annual time scale may help to detect period-‐dependent sex-‐biased movements in annual spawning fish (Kobler et al. 2012).
In summary, we did not find evidence that between-‐individual differences in bullhead movement range were related to body condition, growth rate and heterozygosity. Furthermore, no association to sex or body size was found. Future studies may use a candidate-‐gene approach with genes associated to dispersal or temperament traits (e.g. Amstutz et al. 2006; Fidler et al. 2007; Korsten et al. 2010; Mueller et al. 2011). This might help to detect the evolutionary background of between-‐individual differences in the movement range (dispersal) of non-‐migratory stream fish (see Van Oers & Mueller 2010). Alternatively, gene expression studies can detect direct responses of an individual to changes in the environment and help to define the ecological circumstances when individual dispersal is initiated (see Bell & Aubin-‐Horth 2010; Van Oers & Mueller 2010; Aubin-‐Horth et al. 2012).
Acknowledgments
Many thanks to B. Hellemans, J.-‐A. De Roos, P. Scheys, G. Eens, K. Geudens and Y. Humblet for technical assistance and G. Knaepkens, Maarten Larmuseau, Jeroen Van Houdt and Joost Raeymaekers for support and helpful comments. Many thanks also to the University of Antwerp, the Catholic University of Leuven and the Fund for Scientific Research-‐Flanders for funding (FWO-‐project nr G.0119.08N). The experimental work was performed in agreement with the Belgian and Flemish laws and was approved by the ethical committee of the University of Antwerp (ID number 2007/42).

CHAPTER 8
GENERAL DISCUSSION, CONCLUSIONS AND FUTURE RESEARCH


Discussion, conclusions and future research
137
Late preface: personal reflection
In the beginning, my intention was to find individual characteristics that would help to explain the heterogeneity in movement distances within a bullhead population. To put it in a nutshell, we did not find a single characteristic that was related to the individual range of movements in the stream. Of course, this disappointed me very much. In the end, however, I realized that we discovered several other ecological relationships that increased the understanding of bullhead movement behaviour and habitat use. So all in all, I am satisfied with this thesis as it puts bullhead and more generally small benthic stream fish into the picture, fish of which most people don’t even know about their existence let alone they have an image of their appearance and yet benthic stream fish are so important for healthy ecosystems (e.g. Gelwick & Matthews 1992; Holmlund & Hammer 1999; Covich et al. 2004; Moore 2006).
It occurred uncountable times that people were passing us, stopped and asked what for heaven’s sake we were searching with our equipment in the narrow and shallow stream. Well, they basically believed us every humbug as they could not imagine anything else. The lies ranged from the search for bombs of the second world war, gold nuggets or the cleaning of the stream with our special vacuum cleaner. Everything seemed more plausible than tracking fish called rivierdonderpad (directly translated: river-‐thunder-‐toad or in German Flussdonnerkröte J). Most often the walkers thought that my Dutch was so bad that I got something wrong when I mentioned that I searched fish that are called rivierdonderpad. “You search frogs?” was often the question. “No, fish” I answered. On a lucky day, they then walked on and I had again peace for another half hour when the next walker stopped. They just could not believe that fish could live in this sometimes very “dirty” (turbid) stream. This showed me the necessity of communication between a biologist and people that seem to be less and less connected to mother nature. By knowing that something special, a toad that is actually a fish J, is living in the stream, people are made aware of the ecosystem and the animals therein, animals that might need protection.
In the following paragraphs I will focus on results and findings that have not yet been discussed in the chapters 2 to 7 as well as on topics that overreach the results of this PhD thesis.
The behavioural observations of bullhead in the laboratory are part of chapter 2, 3 and 6. The observation technique, the swimming behaviour and the tank set-‐up are discussed in the following.

Chapter 8
138
Chapter 3 is one of the rare studies linking individual habitat use with temperament traits. A negative relation between the use of structured habitats and aggressiveness was found. The study design could, however, not resolve the causal pathway of this relation. This topic will be taken up in the following.
I enlarge upon the characteristics of the movement range variable, why we used it and discuss the similarities and differences of movement range and dispersal distance.
This thesis examined whether the individual level of fitness is related to the movement range. I discuss the difficulty of the relation between a longer-‐term fitness measure, the individual growth rate, and movement range, which diversified mainly during only a short period. Another possibility to test for a movement range-‐fitness relation is proposed.
I did not test for individual consistency in behaviour across years. One reason was the short lifespan of the bullhead and the increasing number of natural mortalities towards the end of the study. Nevertheless, this topic is discussed in the following and future study possibilities are proposed.
Finally, some information about a candidate gene study that was initiated during the period of my Ph.D. thesis is given. A short overview of previous studies that analysed the association between dispersal or temperament traits and candidate gene variation is also presented.
Laboratory activity of bullhead
Bullhead are mostly active at night (for results and references see chapter 4). In the rather “sterile” environment in the laboratory (e.g. no sediment, clearer water and less structure than in the natural environment) the nocturnal behaviour was strongly expressed and we did not observe any activity during daytime when bullhead were mostly hidden in a refuge. This hidden behaviour during daytime is typical for Cottus spp. (e.g. Greenberg & Holtzman 1987; Greenberg 1991; Natsumeda 1998) and behavioural observations had to be performed at night by means of infra-‐red light recording. The recordings were of poorer quality than during daytime. Nevertheless, the night recordings revealed a swimming behaviour of very active fish that was unexpected: they swam in a continuous motion just below the water surface, mostly along the tank sides. This was rather surprising as bullhead have a very reduced swim-‐bladder (Freyhof et al. 2005;

Discussion, conclusions and future research
139
Kottelat & Freyhof 2007) and I expected them to swim with a hopping motion over the tank bottom by using their large pectoral fins. This motion is to be expected in the natural environment, particularly in stronger current, and is an adaptation to the benthic habitat (Tomlinson & Perrow 2003). And indeed, bullhead with a “normal” level of activity moved through the tank in a hopping motion over the bottom.
The maximum distances that bullhead covered during ten minutes of the laboratory observations were up to 24 quadrats of the measurement grid (one quadrat was 7,5 × 7,5 cm; see chapter 3). This corresponds to approximately 10 m h-‐1 and is higher than the maximum distance covered in short-‐term observations in the field (6.5 m h-‐1, 2-‐h tracking interval, chapter 4). Even with relatively short tracking intervals of two hours it is possible to miss some individual movement information and the distances covered can be underestimated (Horton et al. 2004). Nevertheless, it seems probable that the higher activity in the laboratory and the untypical swimming behaviour of some individuals indicates a higher stress level than in the natural environment. However, the exploration rate of bullhead in the laboratory and in the field correlated positively and the individual exploration rate was repeatable under both conditions (chapter 2). This indicates that a potential influence of stress on behavioural traits still resulted in similar differences between individuals and comparability between the individual rank order of laboratory and field behaviour. It thus seems that the laboratory observations of the present study may be suitable to reflect individual behaviour in the wild. Nevertheless, tank circumstances may be adapted in future studies to maximize the similarity to the field conditions. It is a balancing act between providing species-‐specific circumstances and keeping relatively empty tank spaces that ensure good circumstances for behavioural observations of bottom-‐dwelling fish in complete darkness.
Aggressiveness and the use of structured habitats
Structured habitats like tree roots, rocks or branch jams are preferred habitats of bullhead and are used as refuge shelter (Davey et al. 2005) and foraging habitat (Smyly 1953). Before we studied the relation between habitat use and temperament traits (chapter 3), we had hypothesized that less aggressive individuals should be displaced to less structured and less favourable habitats. This hypothesis was based on a study of Fausch (1984; reviewed by Weber & Fausch 2003) that observed in an experimental stream that more aggressive salmonids (Onconrhynchus kisutch, Salvelinus fontinalis and Salmo trutta) displace

Chapter 8
140
subordinate conspecifics to less profitable stream patches in which the cost of swimming is higher than the energy gain from drifting invertebrates. In bullhead, we assumed that a profitable stream position would be directly related to a preferred (selected) habitat type and more aggressive bullhead should, therefore, be in habitats that provide more complexity.
On the contrary, we found that bullhead that were associated with stream patches with lower habitat complexity were more aggressive. We discussed that the defence of a territory was more difficult in less structured habitats (see discussion in chapter 3). Our study design could, however, not resolve the causal pathway of the negative relation between habitat structure and aggressiveness. Do bullhead become more aggressive when defending their territory in less structured habitats or do more aggressive individuals choose to live in less structured habitats to benefit from underused resources (compare Svanbäck et al. 2008; Kobler et al. 2009a)? Similar difficulties in resolving the causal pathway are to be expected in single correlations between other temperament traits and microhabitat use. For example, individuals in less structured habitats may be exposed to a higher predation risk (Magnhagen & Borcherding 2008) and adapt to their environment by expressing a more risk-‐prone (bolder) behaviour (Brown et al. 2007). The other causal pathway implicates that bolder individuals (boldness might have a genetic component, Brown et al. 2007), are more risk prone and therefore “freer” in habitat choice. As a consequence they might exploit more open habitats.
The question whether the individual expression of aggressiveness (or another temperament trait) is the cause or consequence of habitat use diversification may be disentangled by a habitat choice experiment with limited amount of complex habitats. Several individuals that are individually held under standardized conditions should be tested for aggressiveness before and after the introduction into a habitat choice set-‐up in an experimental tank or stream:
If aggressiveness is the (main) origin of habitat choice, the intra-‐population rank order of the individual expression of aggressiveness should be repeatable between both temperament trait tests. Two scenarios seem possible: 1) Under the assumption that aggressiveness is positively related to dominance (sensu Fausch 1984), more aggressive individuals should occupy the more complex and preferred habitats. 2) The choice of less structured habitats of more aggressive individuals may indicate the usage of an underused resource.

Discussion, conclusions and future research
141
If aggressiveness is not the origin of individual habitat use but the consequence, the rank order of the individual level of aggressiveness should change between both temperament tests. The degree of selected habitat structure should be correlated to the individual level of aggressiveness during the second temperament test but not to aggressiveness during the first test. For example, the individual distribution among different habitat types should be random according to the rank order of the first aggressiveness test but individuals that scored higher during the second test might have been more often found in less structured habitats. This would indicate that the individual level of aggressiveness is a consequence of more difficulties in the defence of less structured habitats (Jensen et al. 2005; Baird et al. 2006; see discussion chapter 3).
Movement range and dispersal distance
During the writing process I encountered continuously the problem that we did not observe movement distances during an entire year period but recaptured the field-‐observed bullhead after ten months and transported them to the university for additional behavioural observations. The duration of the field observations did not suffice to measure dispersal distance; dispersal studies need at least twelve months duration because dispersal is defined as the distance from a natal/breeding site to another breeding site (Clobert et al. 2009). Therefore, we could only assume that the movement distances during ten months indicate individual dispersal distances. This made some part of the writing rather speculative when I aimed to implement our results into the existing animal dispersal literature (that was commonly higher in impact than movement studies in fish). The reason why we recaptured the fish before the end of the annual circle was the necessary independence from reproductive behaviours during the succeeding temperament tests in the laboratory assays (more details can be found in the discussion of chapter 7). Nevertheless, I would do this differently in the future and observe fish over an entire annual cycle. This would give certainty about dispersal distances and may increase the potential impact of the subsequent publications. Furthermore, movements in early winter and, therefore, preceding to the spawning period would enable drawing a more complete picture of sex-‐biased movements as discussed in chapter 5.
To be able to implement our study into the existing dispersal literature we defined the variable “movement range”, the distance between the most up-‐ and downstream location during the ten-‐month observation period (that included one reproductive period). This variable should be a good proximate for the individual

Chapter 8
142
dispersal distance and may be suitable for studies in which the determination of individual breeding sites is rather difficult such as in fish. The correlation between movement range and dispersal distance should be tested in future research to verify whether movement range suits as an indicator of dispersal in fish. A suitable indicator may facilitate the implementation of long-‐term movement studies in fish into the dispersal literature of other vertebrates. For example, while there are numerous studies about sex-‐biased dispersal in birds or mammals (reviewed by Greenwood 1980; Pusey 1987) there are only very few studies conducted in fish (but see Knight et al. 1999; Pardini et al. 2001; Hutchings & Gerber 2002). However, there are indications that sex-‐biased dispersal may be also a common phenomenon in fish (see chapter 5). In addition to movement studies, the study of dispersal in non-‐migratory (stream) fish may thus reveal new insights and facilitate the comparison with dispersal in other vertebrates.
Movement range and individual fitness
In chapter 7 we did not find a relation between movement range of bullhead and body condition, growth rate or heterozygosity. Thus, there was no indication that the individual differences in movement range were associated with fitness. One of the examined fitness indicators, the individual growth rate, might not have been suitable to measure the benefit of the individual expansion in the stream. Longest movement distances were covered during reproductive period in springtime (Kobler et al. 2012). The growth measurements, however, included growth before, during and after that period. Therefore, we could not disentangle whether movement range was negatively associated to growth before the dispersal (sensu Belichon et al. 1996; Hadany et al. 2004) or if individuals might have benefitted from higher growth rate after the dispersal event (Belichon et al. 1996; Lowe 2010; see chapter 7). Another approach would be to consider the individual benefit of dispersal through the reduction of the inbreeding risk.
Breeding between close relatives may result in a fitness decrease of the offspring (reviewed by Pusey & Wolf 1996). This phenomenon is referred to as inbreeding depression and its avoidance may play a major role in the evolution of dispersal and breeding behaviours (Amos et al. 2001 and references therein). Theoretically, the probability of a mating with a closely related conspecific is reduced when the distance of dispersal is longer. Therefore, dispersal distances might be directly linked to fitness (sensu Howard 1960; Pusey & Wolf 1996). For example, meadow voles (Microtus pennsylvanicus) that were released into experimental plots together with siblings were more likely to disperse than voles

Discussion, conclusions and future research
143
released with non-‐siblings (Bollinger et al. 1993). It thus appears that the avoidance of inbreeding can influence dispersal movements (Bollinger et al. 1993).
Future studies may test whether dispersal movements in fish reduce the probability of a mating with a closely related conspecific. Kin recognition may be involved in this process. Three-‐spined sticklebacks (Gasterosteus aculeatus) make decisions about preferred shoaling partners dependent on kinship (Frommen et al. 2007). Similar decisions may also be involved in mate choice. Regarding bullhead, movement distances of females during reproductive period (chapter 5) might be associated with the search for unrelated males. This might be tested in future research. Useful for such a study could be the measure “Identity” that calculates the expected proportion of loci that are homozygous in the offspring of a chosen pair of individuals (software Identix 1.1, Belkhir et al. 2002).
Between-‐year consistency in individual behaviour
This thesis involves movement behaviour, habitat use and temperament trait observations over a period of up to ten months. There is no data to validate behavioural consistency across years. During the study period, however, individual habitat use (r = 0.57), activity (r = 0.44 and r = 0.48) and activity in a novel environment (r = 0.42 and r = 0.53) was relatively consistent (chapter 3 and 6) when compared with correlation coefficients of other studies (reviewed by Bell et al. 2009). A between-‐year consistency of individually distinct habitat use has been demonstrated in e.g. adult lake trout (Salvelinus namaycush, Morbey et al. 2006) and in Icelandic cod (Gadus morhua, Pàlsson & Thorsteinsson 2003). Also temperament traits can be consistent between years. Male great tits can be consistent in their exploratory behaviour from juvenile phase to adulthood (Carere et al. 2005). Theoretically, consistency in behaviour between years is likely because of reduced future costs of ecological adaptation when individual behaviour is established and fine-‐tuned (Wolf et al. 2008). Future costs involve the time, energy, and mortality costs of sampling the environment, or the costs of building and maintaining the required sensory machinery (Wolf et al. 2008). Therefore, even small positive feedback between the established behaviour and fitness may induce behavioural consistency (Wolf et al. 2008).
Nevertheless, the repeatability of individual behaviour declines over time (e.g. Bell et al. 2009; Chervet et al. 2011) as individuals adapt to environmental circumstances. For example, although boldness has a heritable component in

Chapter 8
144
Brachyraphis episcopi fish, its expression in young-‐of-‐the-‐year adapts to the exposure to predation risk (Brown et al. 2007). Other influences on the expression of behaviour can be for example the individual development (changes in ontogenetic stage, Stamps & Groothuis 2010ab), stress in early life-‐stages (Sih 2011), changes in the social context (Webster et al. 2007) or the composition of the group of conspecifics (Sih & Watters 2005; Moretz et al. 2007). Furthermore, different classes of behaviour differ in their repeatability; mating behaviour, habitat selection and aggressiveness are most repeatable (Bell et al. 2009). Based on these findings there is no room for speculation about between-‐year consistency in movement behaviour, habitat use or temperament traits in bullhead. Future studies might compare individual repeatability in behaviour under consideration of species-‐specific lifespan. The positive feedbacks of an established type of behaviour (Wolf et al. 2008) may drive individuals of short-‐lived species to be relatively consistent (even under drastically changing environmental conditions, Dingemanse et al. 2004) while it might be more advantageous for individuals of long-‐lived species to be more adaptive (compare Wolf et al. 2008). Study species that allow relatively easy tracking of individual life histories in the wild provide best opportunities to study the consequences of individual behavioural strategies for major life history traits, including survival, dispersal and reproduction (Dingemanse et al. 2002).
Candidate gene -‐ phenotype association
Variation in individual behaviour (e.g. the degree of explorative or bold behaviour) may have a heritable component (e.g. Verbeek et al. 1994; Dingemanse et al. 2002, 2004; Drent et al. 2003; Bell 2005; Brown et al. 2007). This may indicate a genetic basis of some behavioural traits. To study the genetic basis of dispersal and temperament traits in bullhead, we aim to perform an association analysis with candidate gene variation. Gene polymorphisms may help to explain consistent individual variation in behaviour. For example, a single nucleotide polymorphism (SNP) in a neurotransmitter-‐associated gene (Drd4) differs significantly between slow-‐ and fast-‐exploring great tits (Fidler et al. 2007; this may, however, vary between populations, Korsten et al. 2010). A SNP in the candidate gene ADCYAP1 explains approximately 3% of the migratory tendency of blackcaps (Aves: Sylvia atricapilla; Mueller et al. 2011). The identification of genes or genome regions that underlie behavioural traits will open exciting possibilities to study natural selection at the molecular level (Van Oers & Mueller 2010).

Discussion, conclusions and future research
145
A candidate gene – phenotype association in bullhead was already initiated during the period of my Ph.D. thesis (study by M. Eens, A. Kobler, G. E. Maes, A. Triantafyllidis, F. A. M. Volckaert). Information from public databases (GenBank and ENSEMBL) on behavioural genes in other species was searched and sequence information extracted. Contigs (sets of overlapping DNA segments) of a pyrosequenced Cottus ssp. transcriptome (62,696 recently sequenced contigs provided by Arne Nolte, Max-‐Planck-‐Institute for Evolutionary Biology, Plön, Germany) could then be blasted against the extracted sequences and putative homologues were found for a total of 22 genes. Primers were designed that enabled the successful amplification of 16 genes. Sanger sequencing was used to detect SNPs and 15 candidate SNPs are now available that can be associated to the behavioural heterogeneity in the studied bullhead population. Preliminary analysis revealed an association of a SNP in the transaldolase 1 gene with movement range of bullhead. This needs to be verified but there are indications that this association may be significant (G. Maes, personal communication). Previously it had been shown that sedentary and migratory brown trout populations differently express transaldolase 1 (in the liver) during the period before the onset of migration (Amstutz et al. 2006). It thus seems possible that this gene is also involved in the movement diversification in bullhead. This research area is the follow-‐up of this thesis and it may help to understand the observed intra-‐population heterogeneity in movement range or temperament traits.


References
147
REFERENCES
Alonso J. C., Martin E., Alonso J. A. & Morales M. B. (1998) Proximate and ultimate causes of natal dispersal in the great bustard Otis trada. Behavioral Ecology 9: 245-‐252.
Amos W., Worthington Wilmer J., Fullard K., Burg T. M., Croxall J. P., Bloch D. & Coulson T. (2001) The influence of parental relatedness on reproductive success. Proceedings of the Royal Society of London B 268: 2021-‐2027.
Amstutz U., Giger T., Champigneulle A., Day P. J. R. & Largiadèr C. R. (2006) Distinct temporal patterns of Transaldolase 1 gene expression in future migratory and sedentary brown trout (Salmo trutta). Aquaculture 260: 326-‐336.
Andersson K. G., Brönmark C., Herrmann J., Malmqvist B., Otto C. & Sjörström P. (1986) Presence of sculpins (Cottus gobio) reduces drift and activity of Gammarus pulex (Amphipoda). Hydrobiologia 133: 209-‐215.
Andreasson S. (1969) Locomotory activity patterns of Cottus poecilopus Heckel and C. gobio L. (Pisces). Oikos 20: 78-‐94.
Andreasson S. (1973) Seasonal changes in diel activity of Cottus poecilopus and C. gobio (Pisces) at Arctic circle. Oikos 24: 16-‐23.
Araujo M. S., Bolnick D. I. & Layman C. A. (2011) The ecological causes of individual specialisation. Ecology Letters 14: 948-‐958.
Aubin-‐Horth N., Deschenes M. & Cloutier S. (2012) Natural variation in the molecular stress network correlates with a behavioural syndrome. Hormones and Behaviour 61: 140-‐146.
Austin D., Bowen W. D. & McMillan J. I. (2004) Intra-‐specific variation in movement patterns: modeling individual behaviour in a large marine predator. Oikos 105: 15-‐30.
Baird H. P., Patullo B. & MacMillan D. L. (2006) Reducing aggression between freshwater crayfish (Cherax destructor Clark: Decapoda, Parastacidae) by increasing habitat complexity. Aquaculture Research 37: 1419-‐1428.
Barbraud C., Johnsson A. R. & Bertrault G. (2003) Phenotypic correlates of post-‐fledging dispersal in a population of greater flamingos: the importance of body condition. Journal of Animal Ecology 72: 246-‐257.

References
148
Basquill S. P. & Grant J. W. A. (1998) An increase in habitat complexity reduces aggression and monopolization of food by zebra fish (Danio rerio). Canadian Journal of Zoology 76: 770-‐772.
Belica L. A. T. & Rahel F. J. (2008) Movements of creek chubs, Semotilus atromaculatus, among habitat patches in a plains stream. Ecology of Freshwater Fish 17: 258-‐272.
Belichon S., Clobert J. & Massot M. (1996) Are there differences in fitness components between philopatric and dispersing individuals? Acta Oecologica 17: 503-‐517.
Belkhir K., Castric V. & Bonhomme F. (2002) IDENTIX, a software to test for relatedness in a population using permutation methods. Molecular Ecology Notes 2: 611-‐614.
Bell A. M. (2005) Behavioural differences between individuals and two populations of stickleback (Gasterosteus aculeatus). Journal of Evolutionary Biology 18: 464-‐473.
Bell A. M. (2007) Future direction in behavioural syndromes research. Proceedings of the Royal Society of London B 274: 755-‐761.
Bell A. M. & Aubin-‐Horth N. (2010) What can whole genome expression data tell us about the ecology and evolution of personality? Philosophical Transactions of the Royal Society B 365: 4001-‐4012.
Bell A. M., Hankison S. J. & Laskowski K. L. (2009) The repeatability of behaviour: a meta-‐analysis. Animal Behaviour 77: 771-‐783.
Benjamini Y. & Hochberg Y. (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society B 57: 289-‐300.
Bergmüller R. & Taborsky M. (2007) Adaptive behavioural syndromes due to strategic niche specialization. BMC Ecology 7: doi:10.1186/1472-‐6785-‐7-‐12.
Bisazza A. & Marconato A. (1988) Female mate choice, male-‐male competition and parental care in the river bullhead, Cottus gobio L. (Pisces, Cottidae). Animal Behaviour 36: 1352-‐1360.
Bohonak A. J. (1999) Dispersal, gene flow and population structure. The Quarterly Review of Biology 74: 21-‐45.
Bolger T. & Connolly P. L. (1989) The selection of suitable indexes for the measurement and analysis of fish condition. Journal of Fish Biology 34: 171-‐182.

References
149
Bollache L., Kaldonski N., Troussard J.-‐P., Lagrue C. & Rigaud T. (2006) Spines and behaviour as defences against fish predators in an invasive freshwater amphipod. Animal Behaviour 72: 627-‐633.
Bollinger E. K., Harper S. J. & Barrett G. W. (1993) Inbreeding avoidance increases dispersal movements of the meadow vole. Ecology 74: 1153-‐1156.
Bolnick D. I. (2001) Intraspecific competition favours niche width expansion in Drosophila melanogaster. Nature 410: 463-‐466.
Bolnick D. I., Svanbäck R., Fordyce J. A., Yang L. H., Davis J. M., Husley C. D. & Forister M. L. (2003) The ecology of individuals: incidence and implications of individual specialization. The American Naturalist 161: 1-‐28.
Bonte D. & De la Pena E. (2009) Evolution of body condition-‐dependent dispersal in metapopulations. Journal of Evolutionary Biology 22: 1242-‐1251.
Bonte D., Van Dyck H., Bullock J. M., Coulon A., Delgado M., Gibbs M., Lehouck V., Matthysen E., Mustin K., Saastamoinen M., Schtickzelle N., Stevens V. M., Vandewoestijne S., Baguette M., Barton K., Benton T. M., Chaput-‐Bardy A., Clobert J., Dytham C., Hovestadt T., Meier C. M., Palmer S. C. F., Turlure C. & J. M. J. Travis J. M. J. (2012) Costs of dispersal. Biological Reviews 87: 290-‐312.
Boon A. K., Réale D., Boutin S. (2008) Personality, habitat use, and their consequences for survival in North American red squirrels Tamiasciurus hudsonicus. Oikos 117: 1321-‐1328.
Bowler D. E. & Benton T. G. (2005) Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamics. Biological Reviews 80: 205-‐225.
Brandt S. B. (1986) Ontogenic shifts in habitat, diet, and diel-‐feeding periodicity of slimy sculpin in Lake Ontario. Transactions of the American Fisheries Society 115: 711-‐715.
Breen M. J., Ruetz C. R., Thompson K. J. & Kohler S. L. (2009) Movements of mottled sculpins (Cottus bairdii) in a Michigan stream: how restricted are they? Canadian Journal of Fisheries and Aquatic Sciences 66: 31-‐41.
Broadway J. E. & Moyle P. B. (1978) Aspects of the ecology of the prickly sculpin, Cottus asper Richardson, a persistent native species in Clear Lake, Lake County, California. Environmental Biology of Fishes 3: 337-‐343.
Brown C. & Braithwaite V. A. (2004) Size matters: a test of boldness in eight populations of the poeciliid Brachyraphis episcopi. Animal Behaviour 68: 1325-‐1329.

References
150
Brown C., Burgess F. & Braithwaite V. A. (2007) Heritable and experiential effects on boldness in a tropical poeciliid. Behavioral Ecology and Sociobiology 62: 237-‐243.
Brown C., Jones F. & Braithwaite V. A. (2005) In situ examination of boldness-‐shyness traits in the tropical poeciliid, Brachyraphis episcopi. Animal Behaviour 70: 1003-‐1009.
Bruyndoncx L., Knaepkens G., Meeus W., Bervoets L. & Eens M. (2002) The evaluation of passive integrated transponder (PIT) tags and visible implant elastomer (VIE) marks as new marking techniques for the bullhead. Journal of Fish Biology 60: 260-‐262.
Bunnell Jr. D. B. & Isely J. J. (1998) Diel movement of brown trout in a southern Appalachian river. Transactions of the American Fisheries Society 127: 630-‐636.
Careau V., Thomas D., Humphries M. M. & Réale D. (2008) Energy metabolism and animal personality. Oikos 117: 641-‐653.
Carere C., Drent P. J., Privitera L., Koolhaas J. M. & Groothuis T. G. G. (2005) Personalities in great tits, Parus major: stability and consistency. Animal Behaviour 70: 795-‐805.
Carfagnini A. G., Rodd F. H., Jeffers K. B. & Bruce A. E. E. (2009) The effects of habitat complexity on aggression and fecundity in zebrafish (Danio rerio). Environmental Biology of Fishes 86: 403-‐409.
Chapman B. B., Brönmark C., Nilsson J.-‐A. & Hansson L.-‐A. (2011) The ecology and evolution of partial migration. Oikos 120: 1764-‐1775.
Chapman B. B., Hulthén K., Blomqvist D. R., Hansson L.-‐A., Nilsson J.-‐A., Nilsson P. A., Skov C. & Brönmark C. (2011) To boldly go: individual differences in boldness influence migratory tendency. Ecology Letters 14: 871-‐876.
Chapman J. R., Nakagawa S., Coltman D. W., Slate J. & Sheldon B. C. (2009) A quantitative review of heterozygosity-‐fitness correlations in animal populations. Molecular Ecology 18: 2746-‐2765.
Chervet N., Zöttl M., Schürch R., Taborsky M. & Heg D. (2011) Repeatability and heritability of behavioural types in a social cichlid. International Journal of Evolutionary Biology 2011: ID 321729, doi: 10.4061/2011/321729.
Clapp D. F. & Clark Jr. R. D. (1990) Range, activity, and habitat of large, free-‐ranging brown trout in a Michigan stream. Transactions of the American Fisheries Society 119: 1022-‐1034.

References
151
Clarke A. L., Saether B.-‐E. & Roskaft E. (1997) Sex biases in avian dispersal: a reappraisal. Oikos 79: 429-‐438.
Clobert J., Danchin E., Dhondt A. A. & Nichols J. D. (2001) Dispersal. Oxford, United Kingdom: Oxford University Press.
Clobert J., Le Galliard J. F., Cote J., Meylan S. & Massot M. (2009) Informed dispersal, heterogeneity in animal dispersal syndromes and the dynamics of spatially structured populations. Ecology Letters 12: 197-‐209.
Clough S. & Ladle M. (1997) Diel migration and site fidelity in a stream-‐dwelling cyprinid, Leuciscus leuciscus. Journal of Fish Biology 20: 1117-‐1119.
Coleman K. & Wilson D. S. (1998) Shyness and boldness in pumpkinseed sunfish: individual differences are context-‐specific. Animal Behaviour 56: 927-‐936.
Cote J. & Clobert J. (2007) Social personalities influence natal dispersal in a lizard. Proceedings of the Royal Society B 274: 383-‐390.
Cote J., Fogarty S., Brodin T., Weinersmith K. & Sih A. (2011) Personality-‐dependent dispersal in the invasive mosquitofish: group composition matters. Proceedings of the Royal Society B 278: 1670-‐1678.
Cote J., Fogarty S., Brodin T., Weinersmith K., Brodin T. & Sih A. (2010) Personality traits and dispersal tendency in the invasive mosquitofish (Gambusia affinis). Proceedings of the Royal Society B 277: 1571-‐1579.
Corkum L. D. & Cronin D. J. (2004) Habitat complexity reduces aggression and enhances consumption in crayfish. Journal of Ethology 22: 23-‐27.
Covich A. P., Austen M. C., Bärlocher F., Chauvet E., Cardinale B. J., Biles C. L., Inchausti P., Dangles O., Solan M., Gessner M. O., Statzner B. & Moss B. (2004) The role of biodiversity in the functioning of freshwater and marine benthic ecosystems. Bioscience 54: 767-‐775.
Croft D. P., Albanese B., Arrowsmith B. J., Botham M., Webster M. & Krause J. (2003) Sex-‐biased movement in the guppy (Poecilia reticulata). Oecologia 137: 62-‐68.
Crook D. A. (2004) Is the home range concept compatible with the movements of two species of lowland river fish? Journal of Animal Ecology 73: 353-‐366.
Cucherousset J., Roussel J. M., Keeler R., Cunjak R. A. & Stump R. (2005) The use of two new portable 12-‐mm PIT tag detectors to track small fish in shallow streams. North American Journal of Fisheries Management 25: 270-‐274.

References
152
Davey A. J. H., Hawkins S. J., Turner G. F. & Doncaster C. P. (2005) Size-‐dependent microhabitat use and intraspecific competition in Cottus gobio. Journal of Fish Biology 67: 428-‐423.
David P. (1998) Heterozygosity-‐fitness correlations: new perspectives on old problems. Heredity 80: 531-‐537.
Dingemanse N. J., Both C., Drent P. J., Van Oers K. & Van Noordwijk A. J. (2002) Repeatability and heritability of exploratory behaviour in great tits from the wild. Animal Behaviour 64: 929-‐938.
Dingemanse N. J., Both C., Van Noordwijk A. J., Rutten A. L. & Drent P. J. (2003) Natal dispersal and personalities in great tits (Parus major). Proceedings of the Royal Society B 270: 741-‐747.
Dingemanse N. J., Both C., Drent P. J. & Tinbergen J. M. (2004) Fitness consequences of avian personalities in a fluctuating environment. Proceedings of the Royal Society B 271: 847-‐852.
Dingemanse N. J., Wright J., Kazem A. J. N., Thomas D. K., Hickling R. & Dawnay N. (2007) Behavioural syndromes differ predictably between 12 populations of three-‐spined stickleback. Journal of Animal Ecology 76: 1128-‐1138.
Doerr E. D. & Doerr V. A. J. (2005) Dispersal range analysis: quantifying individual variation in dispersal behaviour. Oecologia 142: 1-‐10.
Drent P. J., Van Oers & Van Noordwijk A. J. (2003) Realized heritability of personalities in the great tit (Parus major). Proceedings of the Royal Society B 270: 45-‐51.
Duckworth R. A. & Badyaev A. V. (2007) Coupling of dispersal and aggression facilitates the rapid range expansion of a passerine bird. Proceedings of the National Academy of Science USA 104: 15017-‐15022.
Ehlinger T. M. & Wilson D. S. (1988) Complex foraging polymorphism in bluegill sunfish. Proceedings of the National Academy of Science USA 85: 1878-‐1882.
Elliott J. M. (2005) Day-‐night changes in the spatial distribution and habitat preferences of freshwater shrimps, Gammarus pulex, in a stony stream. Freshwater Biology 50: 552-‐566.
Englbrecht C. C., Largiadèr C. R., Hänfling B. & Tautz D. (1999) Isolation and characterization of polymorphic microsatellite loci in the European bullhead Cottus gobio L. (Osteichthyes) and their applicability to related taxa. Molecular Ecology 8: 1957-‐1969.
Fausch K. D. (1984) Profitable stream positions for salmonids: relating specific growth rate to net energy gain. Canadian Journal of Zoology 62: 441-‐451.

References
153
Fidler A. E., Van Oers K., Drent P. J., Kuhn S., Mueller J. C. & Kempenaers B. (2007) Drd4 gene polymorphisms are associated with personality variation in a passerine bird. Proceedings of the Royal Society B 274: 1685-‐1691.
Fraser D. F., Gilliam J. F., Daley M. J., Le A. N. & Skalski G. T. (2001) Explaining leptokurtic movement distributions: intrapopulation variation in boldness and exploration. The American Naturalist 158: 124-‐135.
Frasier T. R. (2008) STORM: software for testing hypotheses of relatedness and mating patterns. Molecular Ecology Resources 8: 1263-‐1266.
Freyhof J., Kottelat M. & Nolte A. (2005) Taxonomic diversity of European Cottus with description of eight new species (Teleostei: Cottidae). Ichthyological Exploration of Freshwaters 16: 107-‐172.
Frommen J. G., Mehlis M., Brendler C. & Bakker T. C. M. (2007) Shoaling decisions in three-‐spined sticklebacks (Gasterosteus aculeatus) – familiarity, kinship and inbreeding. Behavioral Ecology and Sociobiology 61: 633-‐539.
Fujimoto Y., Kurosaka K., Ojima D. & Iwata M. (2008) Habitat use and shift of two sympatric freshwater sculpins (Cottus pollux and Cottus hangiongensis) during the spawning and non-‐spawning seasons. Journal of Freshwater Ecology 23: 341-‐346.
Gaines M. S. & McClenaghan Jr. L. R. (1980) Dispersal in small mammals. Annual Review of Ecology and Systematics 11: 163-‐196.
Garamszegi L. Z., Eens M. & Török J. (2008) Birds reveal their personality when singing. PLoS ONE 3: e2647, doi:10.1371/journal.pone.0002647.
Garamszegi L. Z., Eens M. & Török J. (2009) Behavioural syndromes and trappability in free-‐living collared flycatchers, Ficedula albicollis. Animal Behaviour 77: 803-‐812.
Gelwick F. P. & Matthews W. J. (1992) Effects of an algivorous minnow on temperate stream ecosystem porperties. Ecology 73: 1630-‐1645.
Gerking S. D. (1959) The restricted movement of fish populations. Biological Reviews of the Cambridge Philosophical Society 34: 221-‐242.
Godin J.-‐G. & Dugatkin L. A. (1996) Female mating preference for bold males in the guppy, Poecilia reticulata. Proceedings of the National Academy of Science USA 93: 10262-‐10267.
Goto A. (1990) Alternative life-‐history styles of Japanese freshwater sculpins revisited. Environmental Biology of Fishes 28: 101-‐112.

References
154
Goudet J. (1995) FSTAT (Version 1.2): a computer program to calculate F-‐statistics. Journal of Heredity 86: 485-‐486.
Gowan C. & Fausch K. D. (1996) Mobile brook trout in two high-‐elevation Colorado streams: re-‐evaluating the concept of restricted movement. Canadian Journal of Fisheries and Aquatic Sciences 53: 1370-‐1381.
Gowan C., Young M. K., Fausch K. D. & Riley S. C. (1994) Restricted movement in resident stream salmonids: a paradigm lost? Canadian Journal of Fisheries and Aquatic Sciences 51: 2626-‐2637.
Greenberg L. A. (1991) Habitat use and feeding behaviour of thirteen species of benthic stream fishes. Environmental Biology of Fishes 31: 389-‐401.
Greenberg L. A. & Holtzman D. A. (1987) Microhabitat utilization, feeding periodicity, home range and population-‐size of the banded sculpin, Cottus carolinae. Copeia 1: 19-‐25.
Greenwood P. J. (1980) Mating systems, philopatry and dispersal in birds and mammals. Animal Behaviour 28: 1140-‐1162.
Grimm M. P. (1981) The composition of northern pike (Esox lucius L.) populations in four shallow waters in the Netherlands, with special reference to factors influencing 0+ pike biomass. Fisheries Management 12: 61-‐76.
Hadany L., Eshel I. & Motro U. (2004) No place like home: competition, dispersal and complex adaptation. Journal of Evolutionary Biology 17: 1328-‐1335.
Hansson B. & Westerberg L. (2002) On the correlation between heterozygosity and fitness in natural populations. Molecular Ecology 11: 2467-‐2474.
Hawkes C. (2009) Linking movement behaviour, dispersal and population processes: is individual variation a key? Journal of Animal Ecology 78: 894-‐906.
Hays G. C., Kennedy H. & Frost B. W. (2001) Individual variability in diel vertical migration of a marine copepod: why some individuals remain at depth when others migrate. Limnology and Oceanography 46: 2050-‐2054.
Helfman G. S., Collette B. B. & Facey D. E. (1997) The diversity of fishes. Malden, Massachusetts: Blackwell Science. pp. 384-‐405.
Herborn K. A., Macleod R., Miles W. T. S., Schofield A. N. B., Alexander L. & Arnold K. E. (2010) Personality in captivity reflects personality in the wild. Animal Behaviour 79: 835-‐843.

References
155
Hoekstra D. & Janssen J. (1985) Non-‐visual feeding behavior of the mottled sculpin, Cottus bairdi, in Lake Michigan. Environmental Biology of Fishes 12: 111-‐117.
Holmlund C. A. & Hammer M. (1999) Ecosystem services generated by fish populations. Ecological Economics 29 : 253-‐268.
Holomuzki J. R. & Hoyle J. D. (1990) Effect of predatory fish presence on habitat use and diel movement of the stream amphipod, Gammarus minus. Freshwater Biology 24: 509-‐517.
Horton T. B., Guy C. S. & Pontius J. S. (2004) Influence of time interval on estimations of movement and habitat use. North American Journal of Fisheries Management 24: 690-‐696.
Howard W. E. (1960) Innate and environmental dispersal of individual vertebrates. American Midland Naturalist 63: 152-‐161.
Hudy M. & Shiflet J. (2009) Movement and recolonization of Potomac sculpin in a Virginia stream. North American Journal of Fisheries Management 29: 196-‐204.
Hughes N. F. (2000) Testing the ability of habitat selection theory to predict interannual movement patterns of a drift-‐feeding salmonid. Ecology of Freshwater Fish 9: 4-‐8.
Hutchings J. A. & Gerber L. (2002) Sex-‐biased dispersal in a salmonid fish. Proceedings of the Royal Society B 269: 2487-‐2493.
Jakob E. M., Marshall S. D. & Uetz G. W. (1996) Estimating fitness: a comparison of body condition indices. Oikos 77: 61-‐67.
Jensen S. P., Gray S. J. & Hurst J. L. (2005) Excluding neighbours from territories: effects of habitat structure and resource distribution. Animal Behaviour 69: 785-‐795.
Johnson J. C. & Sih A. (2007) Fear, food, sex and parental care: a syndrome of boldness in the fishing spider, Dolomedes triton. Animal Behaviour 74: 1131-‐1138.
Jonsson B. & Jonsson N. (1993) Partial migration: nice shift versual sexual maturation in fishes. Reviews in Fish Biology and Fisheries 3: 348-‐365.
Keeler R. A. (2006) Development and application of passive integrated transponder technology to investigate the movement and reproductive ecology of adult slimy sculpin (Cottus cognatus) in small New Brunswick streams. M.Sc. thesis. New Brunswick, Canada: The University of New Brunswick.

References
156
Keeler R. A., Breton A. R., Peterson D. P. & Cunjak R. A. (2007) Apparent survival and detection estimates for PIT-‐tagged slimy sculpin in five small New Brunswick streams. Transactions of the American Fisheries Society 136: 281-‐292.
Kemp P. S., Armstrong J. D. & Gilvear D. J. (2005) Behavioural responses of juvenile Atlantic salmon (Salmo salar) to presence of boulders. River Research Applications 21: 1053-‐1060.
Knaepkens G., Baekelandt K. & Eens M. (2005) Assessment of the movement behaviour of the bullhead (Cottus gobio), an endangered European freshwater fish. Animal Biology 55: 219-‐226.
Knaepkens G., Bruyndoncx L. & Eens M. (2004) Assessment of residency and movement of the endangered bullhead (Cottus gobio) in two Flemish rivers. Ecology of Freshwater Fish 13: 317-‐322.
Knaepkens G., Maerten E., Tudorache C., De Boeck G. & Eens M. (2007) Evaluation of passive integrated transponder tags for marking the bullhead (Cottus gobio), a small benthic freshwater fish: effects on survival, growth and swimming capacity. Ecology of Freshwater Fish 16: 404-‐409.
Knight M. E., Van Oppen M. J. H., Smith H. L., Rico C., Hewitt G. M. & Turner G. F. (1999) Evidence for male-‐biased dispersal in Lake Malawi cichlids from microsatellites. Molecular Ecology 8: 1521-‐1527.
Knudsen R., Klemetsen, A., Amundsen P.-‐A. & Hermansen B. (2006) Incipient speciation through niche expansion: an example from the Arctic charr in a subarctic lake. Proceedings of the Royal Society B 273: 2291-‐2298.
Kobler A., Engelen B., Knaepkens G. & Eens M. (2009b) Temperament in bullheads: do laboratory and field explorative behaviour variables correlate? Naturwissenschaften 96: 1229-‐1233.
Kobler A., Humblet Y., Knaepkens G., Engelen B. & Eens M. (2012a) Diel movement of bullhead (Cottus perifretum) in a lowland stream. Ecology of Freshwater Fish 21: 453-‐460.
Kobler A., Humblet Y., Geudens K. & Eens M. (2012b) Period-‐dependent sex-‐biased movement in a polygamous stream fish (Cottus perifretum Freyhof, Kottelat & Nolte, 2005 -‐ Actinopterygii, Cottidae) with male parental care. Hydrobiologia 693: 195-‐204.
Kobler A., Klefoth T., Mehner T. & Arlinghaus R. (2009a) Coexistence of behavioural types in an aquatic top predator: a response to resource limitation? Oecologia 161: 837-‐847.

References
157
Kobler A., Klefoth T., Wolter C., Fredrich F. & Arlinghaus R. (2008) Contrasting pike (Esox lucius L.) movement and habitat choice between summer and winter in a small lake. Hydrobiologia 601: 17-‐27.
Kobler A., Volckaert F. A. M., Humblet Y., Maes G. E. & Eens M. (2011) Temperament traits and microhabitat use in bullhead, Cottus perifretum; fish associated with complex habitats are less aggressive. Behaviour 148: 603-‐625.
Korsten P., Mueller J. C., Hermannstädter C., Bouwman K. M., Dingemanse N. J., Drent P. J., Liedvogel M., Matthysen E., Van Oers K., Van Overveld T., Patrick S. C., Quinn J. L., Sheldon B. C., Tinbergen J. M. & Kempenaers B. (2010) Association between DRD4 gene polymorphism and personality variation in great tits: a test across four wild populations. Molecular Ecology 19: 832-‐843.
Kottelat M. & Freyhof J. (2007) Handbook of European Freshwater Fishes. Cornol, Switzerland: Publications Kottelat.
Krause J., Loader S. P., McDermott J. & Ruxton G. D. (1998) Refuge use by fish as a function of body length-‐related metabolic expenditure and predation risks. Proceedings of the Royal Society B 265: 2373-‐2379.
Legalle M., Mastrorillo S., Santoul F. & Céréghino R. (2005) Ontogenetic microhabitat shifts in the bullhead, Cottus gobio L., in a fast flowing stream. International Review of Hydrobiology 90: 310-‐321.
Lessells C. M. & Boag P. T. (1987) Unrepeatable repeatabilities: a common mistake. The Auk 104: 116-‐121.
Lieutenant-‐Gosselin M. & Bernatchez L. (2006) Local heterozygosity-‐fitness correlations with global positive effects on fitness in threespine stickleback. Evolution 60: 1658-‐1668.
Lowe W. H. (2010) Explaining long-‐distance dispersal: effects of dispersal distance on survival and growth in a stream salamander. Ecology 91: 3008-‐3015.
Lucas M. C. & Baras E. (2001) Migration of freshwater fishes. Oxford, United Kingdom: Blackwell Science.
Lundberg P. (1988) The evolution of partial migration in birds. Trends in Ecology & Evolution 3: 172-‐175.
Magnhagen C. (2007) Social influence on the correlation between behaviours in young-‐of-‐the-‐year perch. Behavioral Ecology and Sociobiology 61: 525-‐531.
Magnhagen C. & Borcherding J. (2008) Risk-‐taking behaviour in foraging perch: does predation pressure influence age-‐specific boldness? Animal Behaviour 75: 509-‐517.

References
158
Mainguy J., Côté S. D. & Coltman D. W. (2009) Multilocus heterozygosity, parental relatedness and individual fitness components in a wild mountain goat, Oreamnos americanus population. Molecular Ecology 18: 2297-‐2306.
Mangel M. & Stamps J. A. (2001) Trade-‐offs between growth and mortality and the maintenance of individual variation in growth. Evolutionary Ecology Research 3: 583-‐593.
Manly B. F. J., McDonald L. & Thomas D. (1993) Resource selection by animals: statistical design and analysis for field studies. London, United Kingdom: Chapman & Hall.
Marconato A. & Bisazza A. (1986) Males whose nests contain eggs are preferred by female Cottus gobio L. (Pisces, Cottidae). Animal Behaviour 34: 1580-‐1582.
Marconato A. & Bisazza A. (1988) Mate choice, egg cannibalism and reproductive success in the river bullhead, Cottus gobio L. Journal of Fish Biology 33: 905-‐916.
Marconato A., Bisazza A. & Fabris M. (1993) The cost of parental care and egg cannibalism in the river bullhead, Cottus gobio L. (Pisces, Cottidae). Behavioral Ecology and Sociobiology 32: 229-‐237.
Marentette J. R., Wang G., Tong S., Sopinka N. M., Taves M. D., Koops M. A. & Balshine S. (2011) Laboratory and field evidence of sex-‐biased movement in the invasive round goby. Behavioral Ecology and Sociobiology 65: 2239-‐2249.
Matthews W. J. (1998) Patterns in freshwater fish ecology. New York, United States of America: Chapman & Hall.
Mayr E. (1926) Die Ausbreitung des Girlitz (Serinus canaria serinus L.). Journal für Ornithologie 74: 571-‐671.
McCleave J. D. (1964) Movement and population of the mottled sculpin (Cottus bairdi Girard) in a small Montana stream. Copeia 1964: 506-‐513.
McLaughlin R. L., Grant J. W. A. & Kramer D. L. (1992) Individual variation and alternative patterns of foraging movements in recently-‐emerged brook charr (Salvelinus fontinalis). Behaviour 120: 286-‐301.
McLaughlin R. L. (1989) Search modes of birds and lizards: evidence for alternative movement patterns. The American Naturalist 133: 654-‐670.
Meylan S., De Fraipont M., Aragon P., Vercken E. & Clobert J. (2009) Are dispersal-‐dependent behavioral traits produced by phenotypic plasticity? Journal of Experimental Zoology 311A: 377-‐388.

References
159
Millot S., Bégout M. L. & Chatain B. (2009) Risk-‐taking behaviour variation over time in sea bass Dicentrarchus labrax: effects of day-‐night alternation, fish phenotypic characteristics and selection for growth. Journal of Fish Biology 75: 1733-‐1749.
Moore J. W. (2006) Animal ecosystem engineers in streams. Bioscience 56: 237-‐246.
Morbey Y. E., Addison P., Shuter B. J. & Vascotto K. (2006) Within-‐population heterogeneity of habitat use by lake trout Salvelinus namaycush. Journal of Fish Biology 69: 1675-‐1696.
Moretz J. A., Martins E. P. & Robison B. D. (2007) The effects of early and adult social environment on zebrafish (Danio rerio) behaviour. Environmental Biology of Fishes 80: 91-‐101.
Moretz J. A., Martins E. P. & Robison B. D. (2007) Behavioral syndromes and the evolution of correlated behavior in zebrafish. Behavioral Ecology 18: 556-‐562.
Morris D. (1954) The reproductive behaviour of the river bullhead (Cottus gobio L.) with special reference to the fanning activity. Behaviour 7: 1-‐32.
Morris D. W. (2003) Toward an ecological synthesis: a case for habitat selection. Oecologia 136: 1-‐13.
Mueller J. C., Pulido F. & Kempenaers B. (2011) Identification of a gene associated with avian migratory behaviour. Proceedings of the Royal Society B 278: 2848-‐2856.
Myers J. H. & Krebs C. J. (1971) Genetic, behavioral, and reproductive attributes of dispersing field voles Microtus pennsylvanicus and Microtus ochrogaster. Ecological Monographs 41: 53-‐78.
Nakamura T., Maruyama T. & Watanabe S. (2002) Residency and movement of stream-‐dwelling Japanese charr, Salvelinus leucomaenis, in a central Japanese mountain stream. Ecology of Freshwater Fish 11: 150-‐157.
Natsumeda T. (1998) Home range of the Japanese fluvial sculpin, Cottus pollux, in relation to nocturnal activity patterns. Environmental Biology of Fishes 53: 295-‐301.
Natsumeda T. (1999) Year-‐round local movements of the Japanese fluvial sculpin, Cottus pollux (large egg type), with special reference to the distribution of spawning nests. Ichtyological Research 46: 43-‐48.
Natsumeda T. (2001) Space use by the Japanese fluvial sculpin, Cottus pollux, related to spatio-‐temporal limitations in nest resources. Environmental Biology of Fishes 62: 393-‐400.

References
160
Natsumeda T. (2007) Estimates of nocturnal home range size of the adult Japanese fluvial sculpin, Cottus pollux (Pisces: Cottidae) in relation to bottom topography and sampling intervals. Journal of Ethology 25: 87-‐93.
Nelsen J. S. (1994) Fishes of the World. New York, United States of America: John Wiley & Sons.
Newman R. M. & Waters T. F. (1984) Size-‐selective predation on Gammarus pseudolimnaeus by trout and sculpins. Ecology 65: 1535-‐1545.
Nilsson P. A. (2006) Avoid your neighbours: size-‐determined spatial distribution patterns among northern pike individuals. Oikos 113: 251-‐258.
Nolte A., Stemshorn K. C. & Tautz D. (2005) Direct cloning of microsatellite loci from Cottus gobio through a simplified enrichment procedure. Molecular Ecology Notes 5: 628-‐636.
Okuda N. (1999) Female mating strategy and male brood cannibalism in sand-‐dwelling cardinalfish. Animal Behaviour 58: 273-‐279.
Orians G. H. (1991) Preface. The American Naturalist 137: Supplement Habitat Selection S1-‐S4.
Orians G. H. & Wittenberger J. F. (1991) Spatial and temporal scales in habitat selection. The American Naturalist 137: Supplement Habitat Selection S29-‐S49.
Ovidio, M., Detaille, A., Botninck, C. & J.-‐C. Phillippart, 2009. Movement behaviour of the small benthic Rhine sculpin Cottus rhenanus (Freyhof, Kottelet & Nolte, 2005) as revealed by radio-‐telemetry and pit-‐tagging. Hydrobiologia 636: 119-‐128.
Pàlsson O. K. & Thorsteinsson V. (2003) Migration patterns, ambient temperature, and growth of Icelandic cod (Gadus morhua): evidence from storage tag data. Canadian Journal of Fisheries and Aquatic Sciences 60: 1409-‐1423.
Pardini A. T., Jones C. S., Noble L. R., Kreiser B., Malcolm H., Bruce B. D., Stevens J. D., Cliff G., Scholl M. C., Francis M., Duffy C. A. J. & Martin A. P. (2001) Sex-‐biased dispersal of great white sharks. Nature 412: 139-‐140.
Park S. D. E. (2001) Trypanotolerance in West African cattle and the population genetic effects of selection. Ph.D. thesis. Dublin, Ireland: University of Dublin.
Petty J. T. & Grossman G. D. (1996) Patch selection by mottled sculpin (Pisces: Cottidae) in a southern Appalachian stream. Freshwater Biology 35: 261-‐276.
Petty J. T. & Grossman G. D. (2004) Restricted movement by mottled sculpin (Pisces: Cottidae) in a southern Appalachian stream. Freshwater Biology 49: 631-‐645.

References
161
Petty J. T. & Grossman G. D. (2007) Size-‐dependent territoriality of mottled sculpin in a southern Appalachian stream. Transactions of the American Fisheries Society 136: 1750-‐1761.
Porter J. H. & Dooley Jr. J. L. (1993) Animal dispersal patterns: a reassessment of simple mathematical models. Ecology 74: 2436-‐2443.
Prenda J., Armitage P. D. & Grayston A. (1997) Habitat use by the fish assemblages of two chalk streams. Journal of Fish Biology 51: 64-‐79.
Pujolar J. M., Bevacqua D., Capoccioni F., Ciccotti E., De Leo G. A. & Zane L. (2009) Genetic variability is unrelated to growth and parasite infestation in natural populations of the European eel (Anguilla anguilla). Molecular Ecology 18: 4604-‐4616.
Pujolar J. M., Maes G. E., Vancoillie C. & Volckaert F. A. M. (2006) Environmental stress and life-‐stage dependence on the detection of heterozygosity-‐fitness correlations in the European eel, Anguilla anguilla. Genome 49: 1428-‐1437.
Pusey A. E. (1987) Sex-‐biased dispersal and inbreeding avoidance in birds and mammals. Trends in Ecology & Evolution 10: 295-‐299.
Pusey A. E. & Wolf M. (1996) Inbreeding avoidance in animals. Trends in Ecology & Evolution 11: 201-‐206.
Quinn J. L. & Cresswell W. (2005) Personality, anti-‐predation behaviour and behavioural plasticity in the chaffinch Fringilla coelebs. Behaviour 142: 1377-‐1402.2
Réale D, Gallant BY, Leblanc M, Festa-‐Bianchet M. 2000. Consistency of temperament in bighorn ewes and correlates with behaviour and life history. Anim Behav. 60:589-‐597.
Réale D., Reader S. M., Sol D., McDougall P. T. & Dingemanse N. J. (2007) Integrating animal temperament within ecology and evolution. Biological Reviews 82: 291-‐318.
Reaney L. T. & Backwell P. R. Y. (2007) Risk-‐taking behavior predicts aggression and mating success in a fiddler crab. Behavioral Ecology 18: 521-‐525.
Ricker W. E. (1975) Computation and interpretation of biological statistics of fish populations. Bulletin of the Fisheries Research Board of Canada 191: 1-‐382.
Robinson B. W., Wilson D. S. & Shea G. O. (1996) Trade-‐offs of ecological specialization: an intraspecific comparison of pumpkinseed sunfish phenotypes. Ecology 77: 170-‐178.

References
162
Rodriguez M. A. (2002) Restricted movement in stream fish: the paradigm is incomplete, not lost. Ecology 83: 1-‐13.
Rogers K. B. (2002) FishTel: telemetry analysis package. Version 1.4. Colorado, United States of America: Colorado Division of Wildlife.
Rogers K. B. & White G. C. (2007) Analysis of movement and habitat use from telemetry data. In: Guy C. S. & Brown M. L., eds. Analysis and interpretation of freshwater fisheries data. Bethesda (Maryland), United States of America: American Fisheries Society, pp. 625-‐676.
Rosenzweig M. L. (1991) Habitat selection and population interactions: the search for mechanism. The American Naturalist 137: Supplement Habitat Selection S5-‐S28.
Roussel J.-‐M., Haro A. & Cunjak R. A. (2000) Field test of a new method for tracking small fishes in shallow rivers using passive integrated transponder (PIT) technology. Canadian Journal of Fisheries and Aquatic Sciences 57: 1326-‐1329.
Ruetz C. R., Earl B. M. & Kohler S. L. (2006) Evaluating passive integrated transponder tags for marking mottled sculpins: effects on growth and mortality. Transactions of the American Fisheries Society 135: 1456-‐1461.
Salvanes A. G. V. & Braithwaite V. A. (2005) Exposure to variable spatial information in the early rearing environment generates asymmetries in social interactions in cod (Gadus morhua). Behavioral Ecology and Sociobiology 59: 250-‐257.
Schlosser I. J. & Angermeier P. L. (1995) Spatial variation in demographic processes of lotic fishes: conceptual models, empirical evidence, and implications for conservation. American Fisheries Society Symposium 17: 392-‐401.
Schoener T. W. (1974) Resource partitioning in ecological communities. Science 185: 27-‐39.
Schuett W., Tregenza T. & Dall S. R. X. (2010) Sexual selection and animal personality. Biological Reviews 85: 217-‐246.
Schulte-‐Hostedde A. I., Zinner B., Millar J. S. & Hickling G. J. (2005) Restitution of mass-‐size residuals: validating body condition indices. Ecology 86: 155-‐163.
Selonen V. & Hanski I. K. (2010) Condition-‐dependent, phenotype-‐dependent and genetic-‐dependent factors in the natal dispersal of a solitary rodent. Journal of Animal Ecology 79: 1093-‐1100.

References
163
Shafer A. B. A., Poissant J., Côté S. D. & Coltman D. W. (2011) Does reduced heterozygosity influence dispersal? A test using spatially structured populations in an alpine ungulate. Biology Letters 7: 433-‐435.
Sih A. (2011) Effects of early stress on behavioural syndromes: an integrated adaptive perspective. Neuroscience and Biobehavioral Reviews 35: 1452-‐1465.
Sih A. & Watters J. V. (2005) The mix matters: behavioural types and group dynamics in water striders. Behaviour 142: 1417-‐1431.
Sih A., Bell A. & Johnson J. C. (2004) Behavioral syndromes: an ecological and evolutionary overview. Trends in Ecology & Evolution 19: 372-‐378.
Sinn D. L., Gosling S. D. & Moltschaniwskyj N. A. (2008) Development of shy/bold behaviour in squid: context-‐specific phenotypes associated with developmental plasticity. Animal Behaviour 75: 433-‐442.
Sinn D. L. & Moltschaniwskyj N. A. (2005) Personality traits in dumpling squid (Euprymna tasmanica): context-‐specific traits and their correlation with biological characteristics. Journal of Comparative Psychology 119: 99-‐110.
Skalski G. T. & Gilliam J. F. (2000) Modeling diffusive spread in a heterogeneous population: a movement study with stream fish. Ecology 81: 1685-‐1700.
Skyfield J. P. & Grossman G. D. (2008) Microhabitat use, movements and abundance of gilt darters (Percina evides) in southern Appalachian (USA) streams. Ecology of Freshwater Fish 17: 219-‐230.
Smith B. R. & Blumstein D. T. (2008) Fitness consequences of personality: a meta-‐analysis. Behavioral Ecology 19: 448-‐455.
Smithson E. B. & Johnston C. E. (1999) Movement patterns of stream fishes in a Ouachita Highlands stream: an examination of the restricted movement paradigm. Transactions of the American Fisheries Society 128: 847-‐853.
Smyly W. J. P. (1957) The life-‐history of the bullhead or Miller's thumb (Cottus gobio L.). Proceedings of the Zoological Society of London 128: 431-‐453.
Stamps J. A. (2007) Growth-‐mortality tradeoffs and “personality traits” in animals. Ecology Letters 10: 355-‐363.
Stamps J. A. & Groothuis T. G. G. (2010a) Developmental perspectives on personality: implications for ecological and evolutionary studies of individual differences. Philosophical Transactions of the Royal Society B 365: 4029-‐4041.
Stamps J. A. & Groothuis T. G. G. (2010b) The development of animal personality: relevance, concepts and perspectives. Biological Reviews 85: 301-‐325.

References
164
Stemshorn K. C., Nolte A. W. & Tautz D. (2005) A genetic map of Cottus gobio (Pisces, Teleostei) based on microsatellites can be linked to the physical map of Tetraodon nigroviridis. Journal of Evolutionary Biology 18: 1619-‐1624.
Sundbaum K. & Näslund I. (1998) Effects of woody debris on the growth and behaviour of brown trout in experimental stream channels. Canadian Journal of Zoology 76: 56-‐61.
Svanbäck R. & Bolnick D. I. (2007) Intraspecific competition drives increased resource use diversity within a natural population. Proceedings of the Royal Society B 274: 839-‐844.
Svanbäck R., Eklöv P., Fransson R. & Holmgren K. (2008) Intraspecific competition drives multiple species resource polymorphism in fish communities. Oikos 117: 114-‐124.
Svärdson G. (1949) Competition and habitat selection in birds. Oikos 1: 157-‐174.
Swanson B. O., Gibb A. C., Marks J. C. & Hendrickson D. A. (2003) Trophic polymorphism and behavioral differences decrease intraspecific competition in a cichlid, Herichtys minckleyi. Ecology 84: 1441-‐1446.
Szulkin M., Bierne N. & David P. (2010) Heterozygosity-‐fitness correlations: a time for reappraisal. Evolution 64: 1202-‐1217.
Szulkin M. & Sheldon B. C. (2008) Dispersal as a means of inbreeding avoidance in a wild bird population. Proceedings of the Royal Society of London B 275: 703-‐711.
Tews J., Brose U., Grimm V., Tielbörger K., Wichmann M. C., Schwager M. & Jeltsch F. (2004) Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. Journal of Biogeography 31: 79-‐92.
Tomlinson M. L. & Perrow M. R. (2003) Ecology of the bullhead. Conserving Natura 2000 Rivers Ecology Series No. 4. Peterborough, United Kingdom: English Nature.
Van Oers K. & Mueller J. C. (2010) Evolutionary genomics of animal personality. Philosophical Transactions of the Royal Society B 365: 3991-‐4000.
Van Oers K., Klunder M. & Drent P. J. (2005) Context dependence of personalities: risk-‐taking behavior in a social and a nonsocial situation. Behavioral Ecology 16: 716-‐723.
Van Liefferinge C., Seeuws P., Meire P. & Verheyen R. F. (2005) Microhabitat use and preferences of the endangered Cottus gobio in the River Voer, Belgium. Journal of Fish Biology 67: 897-‐909.

References
165
Van Oosterhout C., Weetman D. & Hutchinson W. F. (2006) Estimation and adjustment of microsatellite null alleles in nonequilibrium populations. Molecular Ecology Notes 6: 255-‐256.
Verbeek M. E. M., Drent P. J. & Wiepkema P. R. (1994) Consistent individual differences in early exploratory behaviour of male great tits. Animal Behaviour 48: 1113-‐1121.
Wallace R. R., Hynes H. B. N. & Kaushik N. K. (1975) Laboratory experiments on factors affecting the activity of Gammarus pseudolimnaeus Bousfield. Freshwater Biology 5: 533-‐546.
Weber E. D. & Fausch K. D. (2003) Interactions between hatchery and wild salmons in streams: differences in biology and evidence for competition. Canadian Journal of Fisheries and Aquatic Sciences 60: 1018-‐1036.
Webster M. M., Ward A. J. W. & Hart P. J. B. (2007) Boldness is influenced by social context in threespine sticklebacks (Gasterosteus aculeatus). Behaviour 144: 351-‐371.
Werner E. E. & Hall D. J. (1976) Niche shifts in sunfishes: experimental evidence and significance. Science 191: 404-‐406.
Whoriskey F. G., Fitzgerald G. J. & Reebs S. G. (1986) The breeding-‐season population structure of three sympatric, territorial sticklebacks (Pisces: Gasterosteidae). Journal of Fish Biology 29: 635-‐648.
Wilson A. D. W. & McLaughlin R. L. (2007) Behavioural syndromes in brook charr, Salvelinus fontinalis: prey-‐search in the field corresponds with space use in novel laboratory situations. Animal Behaviour 74: 689-‐698.
Wilson D. S. (1998) Adaptive individual differences within single populations. Philosophical Transactions of the Royal Society of London B 353: 199-‐205.
Wilson D. S., Clark A. B., Coleman K. & Dearstyne T. (1994) Shyness and boldness in humans and other animals. Trends in Ecology & Evolution 9: 442-‐446.
Wilson D. S., Coleman K., Clark A. B. & Biederman L. (1993) Shy-‐bold continuum in pumpkinseed sunfish (Lepomis gibbosus): an ecological study of a psychological trait. Journal of Comparative Psychology 107: 250-‐260.
Winter H. V. & Fredrich F. (2003) Migratory behaviour of ide: a comparison between the lowland rivers Elbe, Germany, and Vecht, The Netherlands. Journal of Fish Biology 63: 871-‐880.
Wolf M., Van Doorn G. S., Leimar O. & Weissing F. J. (2007) Life-‐history trade-‐offs favour the evolution of personalities. Nature 447: 581-‐584.

References
166
Wolf M., Van Doorn G. S. & Weissing F. J. (2008) Evolutionary emergence of responsive and unresponsive personalities. Proceedings of the National Academy of Science USA 14: 15825-‐15830.
Young M. K. (1999) Summer diel activity and movement of adult brown trout in high-‐elevation streams in Wyoming, U.S.A. Journal of Fish Biology 54: 181-‐189.
Young M. K., Rader R. B. & Belish T. A. (1997) Influence of macroinvertebrate drift and light on the activity and movement of Colorado river cutthroat trout. Transactions of the American Fisheries Society 126: 428-‐437.
Zollner P. A. & Lima S. L. (1999) Search strategies for landscape-‐level interpatch movements. Ecology 80: 1019-‐1030.




![Research Article Morphobiochemical Variability and Selection …downloads.hindawi.com/journals/jb/2014/869167.pdf · 2018. 11. 12. · intrapopulation variations [ ]. 5. Conclusions](https://img.pdfslide.us/doc/110x75/5ff655f2fbf2652f532d87ce/research-article-morphobiochemical-variability-and-selection-2018-11-12-intrapopulation.jpg)

























