Embed Size (px)
Citation preview

VOL. VIII, No. 2 APRIL, 193I
STUDIES ON THE NUTRITION OFBLOW-FLY LARVAE
I. STRUCTURE AND FUNCTION OF THEALIMENTARY TRACT
B Y R. P . HOBSON, B . S C , P H . D .
(From the Department of Entomology, London School ofHygiene and Tropical Medicine.)
(Received August 12th, 1930.)
(With Four Text-figures.)
CONTENTS.PAGE
Introduction 109
Observations and experiments:(i) The anatomy and histology of the mid-gut . . . . 1 1 0(ii) The digestive enzymes in the gut and excreta . . . 113
(iii) The hydrogen-ion concentration in the alimentary tract . 115(iv) The process of digestion and assimilation . . . . 1 1 8
Discussion:(i) Structure and function of the cells of the mid-gut . . 120
(ii) Digestive enzymes and hydrogen-ion concentration in thealimentary tract . . . . . . . . 1 2 0
(iii) The function of the middle segment of the mid-gut . . 121(iv) Synthesis of fat from protein 121
Summary 122
References 123
INTRODUCTION.
THE economic importance of insect nutrition requires no further emphasis asUvarov (1928) has dealt adequately with this subject. An outstanding physiologicalproblem is the role of micro-organisms in the nutrition of insects, and especially ofthose insects which feed habitually on decaying matter. It has been variouslysuggested in such cases that micro-organisms either form the actual food, aid indigestion by their activities, or supply essential accessory substances (vitamins).A classical example is the digestion of meat by blow-fly larvae, and this forms thesubject of the present work.
The larvae can only take in liquid food as the mouth-parts are not adapted formastication and Guyenot (1907), by feeding a paste of animal charcoal, has shown
JEB-VIII ii 8

no R. P. HOBSON
that larvae are unable to ingest any but microscopic particles of solid. Fabre, im-pressed by the rapid liquefaction of meat by maggots, suggested the emission of apowerful pepsin; he did not, however, confirm this experimentally. Guyenot (1906,1907) concluded that Lucilia larvae were entirely dependent upon bacteria fordigestion and incapable of aseptic growth. Bogdanow (1906, 1908) confirmed thistheory by showing that Calliphora larvae could only develope on sterile meat in thepresence of trypsin. However, using an improved technique, Wollman (1911, 1921,1922) claimed to have grown Calliphora larvae under aseptic conditions. Frew(1928), who confirmed the possibility of rearing blow-fly larvae aseptically, did notconsider that growth under these conditions was entirely normal.
As the observations on other lines have been either incidental or confined toparticular aspects, the information available is scattered and incomplete. The presentwork was conceived as a comprehensive study of the general problem of nutritionin blow-fly larvae.
This paper gives an account of the histology of the mid-gut and the physiologyof digestion. The species used was Lucilia sericata Mg., the sheep maggot fly.Although this larva is an important pest on sheep, its normal habitat is decayingmeat.
The larvae have been bred from a pure strain of adults, for the supply of whichI am indebted to Dr G. Salt, of the Farnham House Laboratory.
I take this opportunity of thanking the Empire Marketing Board, for providinga grant which has entirely financed this work, and Dr P. A. Buxton, for his assist-ance and hospitality in extending to me the facilities of his department. I wish alsoto express my appreciations to Dr V. B. Wigglesworth for his numerous suggestionsand active interest throughout this investigation.
OBSERVATIONS AND EXPERIMENTS.
(i) THE ANATOMY AND HISTOLOGY OF THE MID-GUT.
Perez (1910) described the cells of the mid-gut in blow-fly larvae as epithelialcells containing fat globules. Weinland (1908) recognised three regions in the mid-gut; in the first were large epithelial cells containing fat; in the second, cells almostdevoid of fat; in the third, large polygonal cells loaded with fat globules. Theseobservations have been confirmed in various ways.
Larvae were fixed in Carnoy's fluid and serial sections were cut, stained witheosin and counter-stained with Ehrlich's haemotoxylin. The presence of fat wasshown by staining the mid-gut with Sudan III after dissecting out and fixing in10 per cent, formol.
The mid-gut in Lucilia larvae is long and convoluted; the general arrangement,after dissecting and untwisting, is shown in Fig. 1 A. A more exact idea of thedisposition of the coils would be realised by imagining this representation to bespirally twisted. The hind-gut is coiled outside the mid-gut and passes forward asfar as the proventriculus before returning to the hind end of the body. Guyenot

Studies on the nutrition of blow-fly larvae m(1906) figures the undissected gut in Lvdlia larvae, but the course of the mid-gut isdifficult to follow in his illustration.
The different regions of the mid-gut (termed respectively anterior, middle andposterior segments) can be readily recognised under a dissecting microscope. The
Fig. i . A. Mid-gut of Lucilia larva after partial dissection. B. Enlarged view of the distended partof the middle segment. 1, oesophagus; z, proventriculus; 3, gastric diverticula; 4, junction ofanterior and middle segments; 5, distended part of the middle segment; 6, junction of middle andposterior segments; 7, Malpighian tubes; 8, hind-gut; 9, peritrophic membrane coiling withinthe distended mid-gut.
anterior and posterior segments are comparatively thick and contain liquid food, theposterior segment becoming narrow and convoluted at the hind end. The middlesegment is constricted at either end and in between (in a fed larva) is distendedinto a sac in which the peritrophic membrane, filled with nearly solid food, can beseen folding over within the gut (Fig. 1 B). In a starved larva the middle segmentis uniformly narrow throughout.
8-2
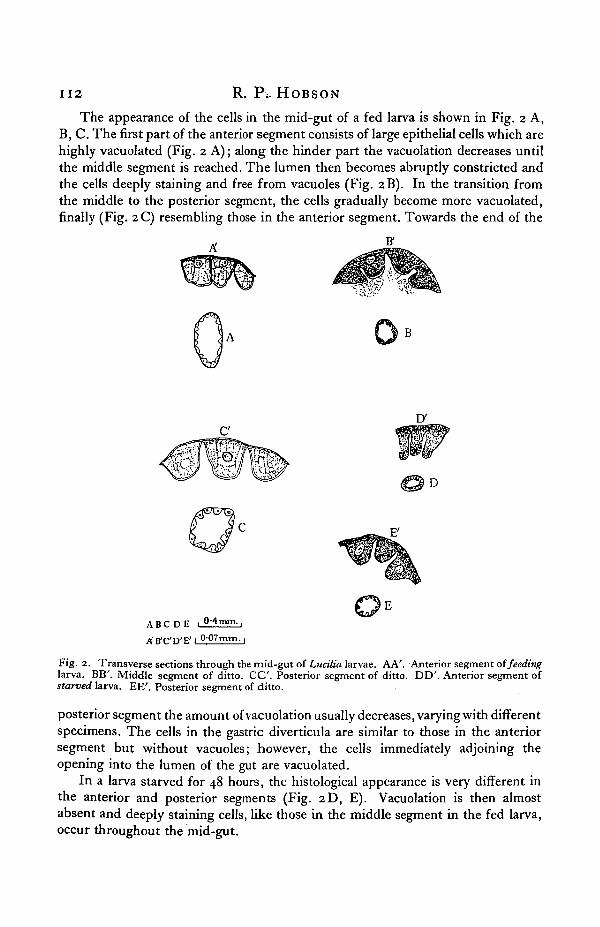
1 1 2 R. Pi- HOBSON
The appearance of the cells in the mid-gut of a fed larva is shown in Fig. 2 A,B, C. The first part of the anterior segment consists of large epithelial cells which arehighly vacuolated (Fig. 2 A); along the hinder part the vacuolation decreases untilthe middle segment is reached. The lumen then becomes abruptly constricted andthe cells deeply staining and free from vacuoles (Fig. 2B). In the transition fromthe middle to the posterior segment, the cells gradually become more vacuolated,finally (Fig. 2 C) resembling those in the anterior segment. Towards the end of the
D
ABC D E 1 0-4mm.,
A'ffC'D'E' 1 007mm. |
Fig. 2. Transverse sections through the mid-gut of Lucilia larvae. AA'. Anterior segment of feedinglarva. BB'. Middle segment of ditto. C C . Posterior segment of ditto. DD'. Anterior segment ofstarved larva. EE'. Posterior segment of ditto.
posterior segment the amount of vacuolation usually decreases, varying with differentspecimens. The cells in the gastric diverticula are similar to those in the anteriorsegment but without vacuoles; however, the cells immediately adjoining theopening into the lumen of the gut are vacuolated.
In a larva starved for 48 hours, the histological appearance is very different inthe anterior and posterior segments (Fig. 2D, E). Vacuolation is then almostabsent and deeply staining cells, like those in the middle segment in the fed larva,occur throughout the mid-gut.

Studies on the nutrition of blow-fly larvae 113
The presence of fat has been shown to coincide with vacuolation, and onlyoccurs in the cells of the anterior and posterior segments of fed larvae. As this fatdisappears after 48 hours' starvation, it is probably a temporary storage product.
(ii) THE DIGESTIVE ENZYMES IN THE GUT AND EXCRETA.
The previous evidence on the digestive enzymes secreted by blow-fly larvae isconflicting. Guyenot (1906, 1907) found no enzymes in extracts made from salivaryglands, diverticula and whole larvae (Lucilid), but he did not test the mid-gutseparately. Wollman (1922) claimed that the liquid residues from aseptic culturesof Calliphora larvae contain tryptase, amylase and peptidase; the methods used,however, would not serve to differentiate between the action of peptidase, as definedby Waldschmidt-Leitz and Harteneck (1925), and tryptase. Weinland (1906) showedthe presence of tryptase in the excreta of Calliphora larvae.
As the previous work supplies only indirect evidence on the digestive enzymesin the gut, this subject has been re-investigated. The methods used were essentiallythose devised by Wigglesworth (1927a, 1928, 1929); his technique is described indetail in the earlier publications and summarised in the last-named. The procedureadopted will be described briefly.
Enzyme preparations.
For the dissections half-grown larvae were starved until the gut was empty(about 48 hours). To keep the larvae moist during starvation and prevent ingestionof the excreta, they were placed over a sintered glass filter covered with gauze anda slow stream of water passed through the filter with the aid of suction.
The following preparations were made:(i) "Glycerol extracts": the region in question dissected out from 50 larvae,
ground up with thymol and stored in 5 c.c. of 50 per cent, glycerol.(ii) " Dry preparations ": a similar quantity of material ground up to a uniform
paste with about \ gm. of moist clean sand and thymol and dried at room temperature.Preparations from the excreta were made as follows: 400 larvae of all sizes were
removed from the meat, well washed, placed on a glass filter, and 10 c.c. of waterallowed to drip through thelarvae and filter in 4 hours. 5 c.c. of the filtrate weremixed with thymol and 5 c.c. of glycerol; the remaining 5 c.c. were mixed with15 gm. of sand and thymol and dried at room temperature.
Methods for the detection of the various enzymes.
Amylase. Glycerol extracts acting upon f per cent, starch solutions. Testedwith iodine at intervals.
Invertase. Glycerol extracts acting upon 3 per cent, sucrose. Fehling's test atintervals.
Maltase. Dry preparations extracted shortly before use with phosphate buffer/>H 6*8. This extract acting upon 3 per cent, maltose. Osazone test at intervals.
Lactase. As for maltase, using 3 per cent, lactose as substrate.

R. P. HOBSON
Tryptase. Glycerol extracts acting upon 5 per cent, gelatine at />H 8*o. Form-aldehyde titration at intervals.
Peptase. As for tryptase but at pH 2-2.Peptidase. Glycerol extracts acting upon 1 per cent, glycylglycine. Formalde-
hyde titration at intervals.Lipase. Stable preparations could not be obtained. Fresh preparations shaken
with 10 per cent, ethyl butyrate at pH 7-8. Titration at intervals to pH 9-0.The results are given in Table I, in which the + signs indicate roughly the
relative activity of the various enzymes, the sign ± representing definite but veryweak activity.
Table I. Distribution of digestive enzymes in larvae of Lucilia sericata.
Enzyme
AmylaseInvertaseMaltaseLactaseTryptasePeptasePeptidaseLipase
Salivaryglands
+1 1 1 1 1 1 1 ~-
Mid-gut*
+ ~ +
Hind-gut Excreta
±
* The gastric diverticula and proventriculus were included with the mid-gut.
These results with Lucilia larvae are such as might be expected in a flesh-feedinginsect and agree with the findings of Weinland and Wollman: enzymes acting uponcarbohydrates absent, except for a weak amylase in the salivary glands; proteolyticand lipolytic enzymes active in the mid-gut and the former present also in theexcreta. The lipase from Lucilia larvae is unstable in glycerol extracts and resemblesin this respect the lipase found by Wigglesworth (1928) in the cockroach.
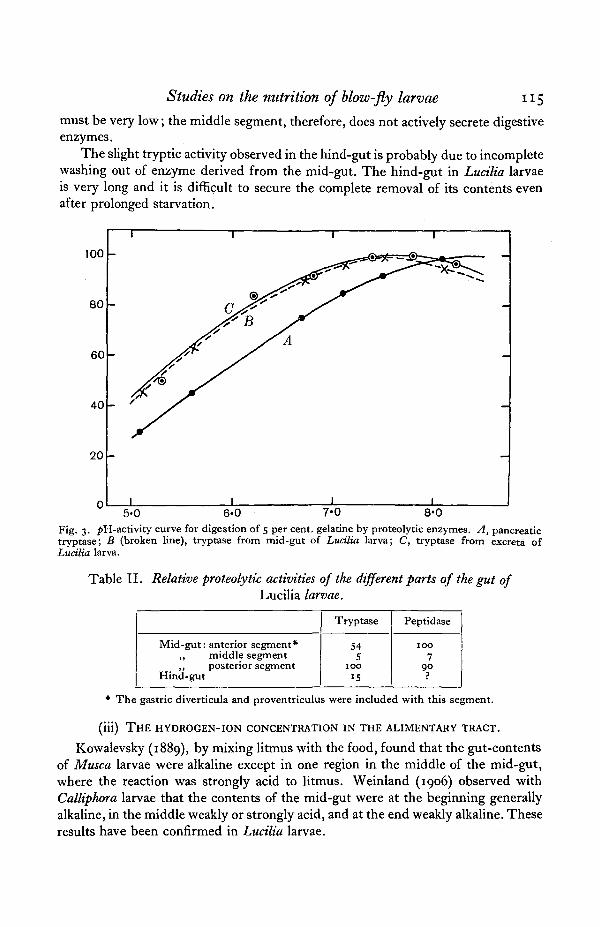
In order to compare the tryptase from the mid-gut and excreta, pii activitydeterminations were made with these enzymes and pancreatic tryptase; the resultsare shown in Fig. 3. The similarity of the curves for the tryptase from the mid-gutand excreta leaves little doubt that the enzyme in the excreta is derived from themid-gut. The tryptase from Lucilia larvae is more active in the acid region thanpancreatic trypsin, as was found to be the case also by Wigglesworth (1928, 1929)with the tryptase of the cockroach and Glossina.
The distribution of the proteolytic enzymes in the gut is given in Table II, inwhich the relative activities of the different regions (from the same number oflarvae) are expressed as percentages of the most active region.
This table shows that the proteolytic enzymes are concentrated mainly in theanterior and posterior segments of the mid-gut. The distribution of the lipase,which was not determined, is presumably the same, since in insects the secretionof different enzymes usually occurs in the same region of the gut.
The results for the middle segment are barely outside the experimental errorand might be due to a slight contamination in dissection. The amount of tissue inthis segment is admittedly small in bulk, but even so the concentration of enzyme
Studies on the nutrition of blow-fly larvae 115
must be very low; the middle segment, therefore, does not actively secrete digestiveenzymes.
The slight tryptic activity observed in the hind-gut is probably due to incompletewashing out of enzyme derived from the mid-gut. The hind-gut in Luctlia larvaeis very long and it is difficult to secure the complete removal of its contents evenafter prolonged starvation.
100
80
60
40
20
n
I
-
„
- ' y
i
i i i
'A—
-
-
i i
5-0 6-0 7 '0 8*0
Fig. 3. />H-activity curve for digestion of 5 per cent, gelatine by proteolytic enzymes. A, pancreatictryptase; B (broken line), tryptase from mid-gut of Luctlia larva; C, tryptase from excreta ofLucilia larva.
Table II. Relative proteolytic activities of the different parts of the gut ofLucilia larvae.
Mid-gut: anterior segment*„ middle segment„ posterior segment
Hind-gut
Tryptase
545
100IS
Peptidase
1007
90
* The gastric diverticula and proventriculus were included with this segment.
(iii) THE HYDROGEN-ION CONCENTRATION IN THE ALIMENTARY TRACT.
Kowalevsky (1889), by mixing litmus with the food, found that the gut-contentsof Musca larvae were alkaline except in one region in the middle of the mid-gut,where the reaction was strongly acid to litmus. Weinland (1906) observed withCalliphora larvae that the contents of the mid-gut were at the beginning generallyalkaline, in the middle weakly or strongly acid, and at the end weakly alkaline. Theseresults have been confirmed in Lucilia larvae.

u 6 R. P. HOBSON
Methods.
Owing to the small size of the lumen it was found impossible to determine thereaction except by adding indicators to the food and observing the colour changesalong the gut. This method can only be regarded as approximate, since the tintscannot be compared accurately and the presence of protein may cause errors; how-ever, it is convenient for showing changes of reaction in a long narrow gut such asthat of Lucilia larvae.
The indicators were usually fed in neutralised 5 per cent, gelatine or peptonesolutions to starved larvae, the gut being dissected out after suitable intervals andthe/»H in the different parts noted. To check this method under more natural con-ditions, the indicators were also fed mixed with liquefied meat but, owing to thedark colour of the food, the pH could only be estimated very approximately. Exceptin the crop and the anterior segment of the mid-gut, the reaction in the gut wasindependent of the nature of the food.
Results.
The reaction of the crop contents was found to be the same as that of the foodfed; when the food was decomposing meat, the reaction in the crop was alkaline,the pH ranging from 7-5 to 8-o. In the anterior segment of the mid-gut, the pHvaried from 6-5, with fresh neutral gelatine, to 7*5, with decomposing meat; thelatter value probably represents the reaction in this region when the larvae are ontheir normal diet. The indicators used were chlorphenol red, neutral red, cresol redand phenol red.
The contents of the middle segment reacted acid to methyl red, congo red,bromcresol green and bromphenol blue. Gunzberg's test proved negative, but,owing to the minute amount of material available, the failure of this reaction is notentirely conclusive; however, it probably indicates the absence of free hydro-chloric acid and a reaction more alkaline than^>H 3-0. As the results with indicatorsshow that the pH is not more alkaline than 3-5, the most probable value for thisregion is pU 3-0-3-5.
Table III. Average pH values of gut contents of Lucilia larvae.
Region of gut
CropMid-gut: anterior segment
,, middle segment,, posterior segment
Hind-gut
Food
Freshgelatine(/>H yo)
706-S
Liquefiedmeat
(pU 77)
. 777-5
J
3'2
8082
In the posterior segment the reaction was always alkaline but varied with theposition of the food in the segment, the alkalinity generally increasing towards the

Studies on the nutrition of blow-fly larvae 117
hind-gut. The outside values obtained, using phenol red and cresol red, werepH 7-3 and 8-4; generally the pH was about 8-o. No gradual change could beobserved between this alkaline region and the preceding acid region, as little foodwas usually present in the intermediate zone. The contents of the hind-gut wereabout pH 80-8-5; alkaline to phenol red and cresol red, acid to phenolphthalein.
These results are summarised in Table III. The range of reaction found in thegut of Ludlia larvae is so unusually wide for an insect that the method of itsproduction requires consideration.
Causes of the changes in reaction.
Wigglesworth (1927) explained the acidity in the crop of the cockroach as theresult of micro-organisms acting on sugars, the pH. being 4-8 with a carbohydratediet and 6-3 with a protein diet. As the food of blow-fly larvae is free from carbo-hydrate, it is unlikely that micro-organisms produce the acid reaction noted in themiddle segment; in addition, the degree of acidity suggests an inorganic acid. Thepresence of free hydrochloric acid has not been recorded in the gut of insects and isrendered improbable in this case by the failure of Gunzberg's test. Biedermann(1898) has suggested that a monobasic salt of phosphoric acid is responsible for themild acidity noted in the gut of meal-worms.
Larvae of Ludlia were starved until the gut was empty and then fed on gelatine,a food deficient in phosphorus. The mid-guts were dissected out from 10 larvae,and the contents, opened into 1 c.c. of water and tested with proportionate amountsof Briggs' (1922) phosphate reagents, gave a positive result. In order to examine theamounts of phosphate in the tissues of the gut in starved larvae, the "dry prepara-tions " (cf. p. 113) were extracted and tested in the same way. Phosphate was foundto be present, the amount for an equal bulk of tissue being approximately the samein the anterior and middle segments and definitely less in the posterior segment.
The reaction in the acid region (pH 3-0-3-5) corresponds to the partial neutrali-sation of the first hydrogen-ion of phosphoric acid, and this condition could easilyresult if the free acid were secreted and partly neutralised by the bases in the food.Moreover, the production of an acid secretion in the middle segment of the mid-gutis not unlikely, as the cells in this region have the appearance of secreting cells andyet contain little or no digestive enzymes. Although the secretion of free phosphoricacid cannot be regarded as proven and the evidence is largely circumstantial, never-theless this hypothesis seems to afford the most plausible explanation of the facts.
The vigorous production of ammonia, which accompanies the liquefaction ofmeat by larvae, explains the alkaline reaction in the crop and anterior segment ofthe mid-gut. This alkalinity is absent after a meal of fresh food and is, therefore,derived from the food and not from bacterial action in the crop. The alkaline re-action in the posterior segment is presumably due also to ammonia, which ispresent in large amounts in the excreta. This view has been confirmed by showingthe presence of ammonia in the mid-gut. The mid-guts were dissected out from10 larvae, the contents opened into 1 c.c. of water, and proteins precipitated withtrichloracetic acid; the filtrate gave a positive reaction with Nessler's solution.

u8 R. P. HOBSON
The source of the ammonia present in the excreta was held by Bogdanow (1908)to be the action of micro-organisms; on the other hand, Weinland (1908) insistedthat deamination of amino-acids occurs in the tissues of the larva, either in the mid-gut or elsewhere. Tests were, therefore, made for the presence of a deaminase in themid-gut.
The mid-guts from 10 starved larvae were divided into the separate segmentsand each lot ground up to a fine emulsion with water containing thymol. The freshpreparations were incubated with 2 per cent, glycine solutions in the presence ofphosphate bufferpH 6-8. Aliquots were removed at intervals, diluted fifty times andtested with suitable amounts of Nessler's solution. No evidence of deamination wasfound with any of the preparations. This does not, however, preclude the possi-bility of deamination occurring in vivo; for, although deamination is a commonoccurrence in the animal body and in bacteria, no well-defined deaminase has beendescribed except those acting upon special amino-acids. The origin of the ammonia,therefore, remains an open question.
(iv) THE PROCESS OF DIGESTION AND ASSIMILATION.
Concentration of the food.
When larvae were fed with coloured foods, the concentration of dye was seento be higher in the middle segment of the mid-gut than in the anterior segment. Thesemi-solid state of the food in the middle segment is due, therefore, to the absorp-tion of water, possibly also to a partial coagulation of the protein in the food by theacid reaction (pH 3-0-3-5).
The middle segment appears to be the main seat of water absorption, as the firstfood taken up by a starved larva passes rapidly to this region which soon becomesdistended with food. Moreover, the peculiar structure of the middle part of thissegment, shown macroscopically in Fig. 1B and in section in Fig. 4, suggests amechanism for expressing water from the food. The gut in the region is distendedand the peritrophic membrane coils freely within it.
This accumulation of the membrane can only be produced by peristalsis at theentrance, resistance at the end and relaxation of the muscles in the intermediateregion. The increased hydrodynamic pressure of the liquid will tend to force waterout of the gut and the cells, being flattened, will offer less resistance. Vigorousperistalsis can be seen to occur in the anterior segment, whose function seems to beto drive the food into the constricted portion at the beginning of the middle segment.
Digestion.
When larvae were fed with fresh blood, blackening occurred first in the middlesegment of the mid-gut, the anterior segment and crop containing red blood. Thisobservation completely refutes Perez's suggestion that digestion within the larva islimited to bacterial action in the crop. Actually, the crop appears to function onlyas a storage organ since in a starved larva the first food passes directly into themid-gut.

Studies on the nutrition of blow-fly larvae 119
The blackening of blood in the middle segment is probably due to hydrolysis ofhaemoglobin to haematin and globin and not to digestion as, at the pH found inthis region (3"O—3-5), blood has been shown to blacken in the cold and the tryptasewould be inactive. Protein digestion, therefore, occurs mainly in the posteriorsegment.
D
Fig. 4. Longitudinal sections through the distended region in the middle segment ofLucilia larva. A, coils of the peritrophic membrane enclosing food; B, cells of the gutdistended; C, cells of the gut not distended ;•£>, haemocoel fluid.
The absence of digestion in the anterior segment, despite the presence ofenzyme and a favourable reaction, may be due to several causes. In this segmentthe dilution of the food is greater, and its passage through the gut more rapid, thanin the posterior segment. In addition, the enzyme secreted may tend to passforwards between the peritrophic membrane and the gut, so escaping diffusioninto the food until the constriction at the beginning of the middle segment isreached.

I2O R. P . HOBSON
Absorption.
The presence of fat in the anterior and posterior segments of the mid-gut in-dicates that absorption of food occurs in both these regions. Weinland (1906)suggested that the fat in the former is derived from fat in the food, and in the latterby synthesis from protein. The following experiments show that fat can result ineach region from either source in Lucttia larvae.
When larvae were starved until the gut was free from fat and then fed on oliveoil emulsified in 10 per cent, peptone, fat was found after 8 hours' feeding in theanterior and posterior segments; controls fed on 10 per cent, peptone alone for thesame period showed no fat in the mid-gut. Fat was also found in both segments inlarvae reared entirely on a protein diet (fish). This confirms Weinland's claim thatblow-fly larvae can synthesise fat from protein.
DISCUSSION,
(i) Structure and function of the cells of the mid-gut.
The mid-gut in Lucilia larvae can be separated on various grounds into threesegments. The histology of the anterior and posterior segments is similar, and varieswith the nutrition. In a feeding larva, the cells appear distended with vacuoles andcontain fat; after starvation the cells are deeply staining and free from vacuolationand have been shown chemically to contain digestive enzymes. In these segments,therefore, the cells possess the dual function of absorption and secretion.
Buchmann (1929, 1930) arrived at similar conclusions with larvae of Stomoxysand Musca. From histological studies and feeding experiments with colloidal ironand intravital stains, he suggested that the cells are monomorphous throughout themid-gut and pass through alternate stages of absorption and secretion. Buchmanndid not find any differentiation of the mid-gut; this is not the case in Lucilia larvaesince the cells in the middle segment always appear deeply staining and do notabsorb fat.
Probably the supply of food determines whether absorption or secretion pre-dominates in the mid-gut of Lucilia larvae. Under favourable conditions theactivities of bacteria will supply the larva with partly digested liquid food, absorptionbecoming the main function of the mid-gut. In the absence of liquid food, the cellswill become free from fat and secrete more actively, thus securing an increasedliquefaction of the meat since a part of the tryptase is excreted.
(ii) Digestive enzymes and hydrogen-ion concentration in the alimentary tract.
The enzymes present in the mid-gut cover the requirements of the larva andinclude tryptase, peptidase and lipase. The liquefaction of the meat outside thebody is rendered possible by the persistence of tryptase in the excreta.
It is of interest to make a comparison between the blow-fly larva and adult. Inthe larva (Lucilia), proteolytic enzymes predominate ahd enzymes acting uponcarbohydrate are deficient. In the imago (Calliphora), Wigglesworth (1929) found

Studies on the nutrition of blow-fly larvae 121
the reverse; enzymes acting upon carbohydrate being active, upon protein relativelyinactive. Thus in blow-flies the type of enzymes secreted changes with the diet inthe passage from larva to imago.
The tryptase from Lucilia larvae is well adapted for acting in the mid-gut, sinceits optimum lies at about pH 7-6 and the reaction is approximately pH 8-o in theposterior segment where digestion mainly occurs.
That the enzymes described are derived from the tissues of the mid-gut, andnot from micro-organisms, I have tacitly assumed in the belief that there are notenough organisms in the empty gut to interfere appreciably. The only alternativeis to suppose that digestion is due to an obligative symbiont, which is unable togrow on the usual nutrient media and so escaped detection in the alleged asepticexperiments of Wollman and Frew. Since this interpretation involves considerableassumption, it seems more reasonable to suppose that the enzymes are secreted bythe mid-gut. The subject, however, requires critical bacteriological examinationunder septic and aseptic conditions and will be further investigated.
(iii) The function of the middle segment of the mid-gut.
Since a peptase is not present in the mid-gut, the acid reaction in the middlesegment does not assist directly in the digestive process. The acidity may serve tocheck the excessive multiplication of bacteria, which might otherwise occur on adiet of decomposing meat; also, the bacteria killed would become available fordigestion. This would be an important function if micro-organisms are of any dieteticvalue in nutrition; Baumberger (1919) regarded them as the actual food of the blow-fly larvae, Portier (1919) as an important source of vitamins for insects.
The structure of the middle segment suggests that it acts as mechanism forexpressing water from the food. The peculiar appearance of the peritrophic mem-brane as a tube coiling within the mid-gut, noted in this region in Lucilia larvae, hasalso been described by Wagner (1863) and Mecznikow (1866) in the larva of Miastor(Diptera, Nematocera).
Wigglesworth, discussing the mechanics of water absorption in the gut ofGlossina, suggests the following as effective factors: (a) active work on the part ofthe epithelial cells, (b) mechanical pressure, (c) the action of the Malpighian tubes inmaintaining the osmotic pressure of the haemocoel fluid and so opposing the risingosmotic pressure within the gut. In Lucilia larvae another factor may operate, theacidity in the middle segment decreasing the osmotic pressure within the gut bypartly coagulating the food. Although the exact nature of the gut contents in thisregion is uncertain, a partial precipitation of protein is likely.
(iv) Synthesis of fat from protein.
The presence of fat in larvae reared on a protein diet proves the synthesis of fatfrom protein; it is not possible to decide whether the fat found in the mid-gut hasbeen synthesised in situ or deposited in the gut after synthesis elsewhere. Weinlandclaimed that this synthesis occurs in the tissues of blow-fly larvae, but he did notexclude the possibility of bacterial co-operation; however, as the chemical side of

122 R. P. HOBSON
the process presents no difficulties, his claim appears reasonable. The formation ofsugar from protein and of fat from sugar is generally admitted to occur in mam-malian tissue, and there is no reason to suppose that insect tissues possess inferiorsynthetic powers. Frew reared blow-flies aseptically from egg to imago on a fat-free diet (peptone agar) and observed that the fat metabolism was abnormal in thelarva; nevertheless, fat must presumably have been synthesised in the larvae, assome pupated successfully.
SUMMARY.
1. The mid-gut in Lucilia larvae can be divided into three distinct regions(termed anterior, middle and posterior segments).
2. Histologically the anterior and posterior segments are similar. In a feedinglarva the cells are highly vacuolated and contain fat; in a starved larva the cells aredeeply staining and non-vacuolated. In the middle segment the cells are alwaysdeeply staining and free from vacuoles and fat, whatever the state of nutrition.
3. The hydrogen-ion concentration varies along the gut and with the nature ofthe food. With liquefied meat as food, the^>H is 7-5-8-0 in the crop, 7-5 in the an-terior segment, 3-0-3-5 in the middle segment, 7-5-8-3 in the posterior segment,and 8-0-8-5 in the hind-gut. With fresh gelatine (pH 7-0) as food, the values are thesame except in the crop and anterior segment, for which the figures are respectivelypH 7-0 and 6-5.
4. It has been suggested that the acidity in the middle segment may be due toan acid secretion, the most likely component being phosphoric acid. The alkalinereaction in other parts of the gut is probably caused by ammonia, which is presentin the gut-contents and excreta.
5. Tryptase, peptidase and lipase are present in the mid-gut, the enzymes beingconcentrated in the anterior and posterior segments. The proteolytic enzymes per-sist in the excreta and some extra-intestinal digestion, therefore, can occur withoutthe aid of micro-organisms. Carbohydrate-splitting enzymes are absent except fora feeble secretion of amylase in the salivary gland.
6. By combining the evidence from various sources, I have attempted to obtaina complete picture of the process of digestion, which I suggest is as follows:
The food is stored unchanged in the crop and, passing into the mid-gut, israpidly forced into the middle segment. Some absorption of water and simple pro-ducts of bacterial action occurs along the anterior segment, the concentration of thefood being completed in the middle segment where the acidity prevents digestion.The food, by now of a pasty constituency, passes into the posterior segment, dis-solves in the alkaline fluid and is digested and absorbed. The digestive enzymes aresecreted in the anterior and posterior segments, but digestion does not progress farin the anterior segment owing to the rapid passage of the food.

Studies on the nutrition of blow-fly larvae 123
REFERENCES.
BAUMBERGER, J. P. (1919). A nutritional study of insects, with special reference to micro-organismsand their substrata. Journ. Exp. Zool. 28, i.
BIEDERMANN, W. (1898). Beitrage zur vergleichenden Physiologie der Verdauung. I. Die Verdauungder Larve von Tenebrio molitor. Arch. ges. Physiol. 72, 105.
BOGDANOW, E. A. (1906). Ueber das Zuchten der gewohnlichen Fleischfliege (Calliphora vomitoria)in sterilisierten Nahrmitteln. Arch. ges. Physiol. 113, 97.(1908). Ueber die Abhangigkeit des Wachstums der Fliegenlarven von Bakterien und Fermen-
ten und iiber Variabilitat und Vererbung bei den Fliegenlarven. Arch. Anat. Physiol. 1908,Suppl. 173.
BRIGGS, A. P. (1922). A modification of the Bell-Doisy phosphate method. Journ. Biol. Chem. 53, 13.BUCHMANN, W. (1929). Untersuchungen iiber die Ernahrungsphysiologie der Dipterenlarven.
Teil I. Histophysiologie der Sekretion. Z. Desinf. 21, 237.(1930). Untersuchungen iiber die Ernahrungsphysiologie der Dipterenlarven. Teil II. Die
Histophysiologie der Resorption. Z. Desinf. 22, 6.FABRE, J. H. La mouche bleue de la viande. Le ver. Souvenirs Entomol. 10, 259.FREW, J. G. H. (1928). A technique for the cultivation of insect tissues. Brit. Journ. Exp. Biol. 6, 1.GUYENOT, E. (1906). Sur la mode de nutrition de quelques larves de mouches. C.R. Soc. Biol. 61,634.
(1907). L'appareil digestif et la digestion de quelques larves de mouches. Bull. Sci. Fr. Belg.41, 353-
KOWALEVSKY, A. (1889). Ein Beitrag zur Kenntnis der Exkretionsorgane. Biol. Zentralbl. 9, 33.MECZNIKOW, E. (1866). Embryologische Studien an Insecten. Z. Wiss. Zool. 16, 389.PEREZ, C. (1910). Recherches histologiques sur la metamorphose des Muscides: Calliphora erythro-
cephala, Mg. Arch. Zool. Exp. Gener. (5), 4, 1.PORTIER, P. (1919). DeVeloppement complet des larves de Tenebrio molitor, obtenu au moyen d'une
nourriture st6rilis£e a haute temperature (130°). C.R. Soc. Biol. 62, 59.UVAROV, B. P. (1928). Insect nutrition and metabolism. A summary of the literature. Trans. Ent.
Soc. Lond. (1928), 255.WAGNER, N. (1863). Beitrag zur Lehre von der Fortpflanzung der Insectenlarven. Z. Wiss. Zool.
13, 513-WALDSCHMIDT-LEITZ, E. and HARTENECK, A. (1925). Ueber die tryptische und ereptische Wirkung
der Pankreasdriise. Z. physiol. Chem. 149, 203.WEINLAND, E. (1906). Ueber die Ausscheidung von Ammoniak durch die Larven von Calliphora und
iiber eineBeziehungdieserTatsache zudemEntwickelungs-stadiumdieser Tiere. Z.Biol. 47,232.(1908). Ueber die Bildung von Fett aus eiweissartiger Substanz im Brei der Calliphoralarven.
Z. Biol. 51, 197.WIGGLESWORTH, V. B. (1927). Digestion in the cockroach. I. The hydrogen-ion concentration in the
alimentary canal. Biochem. Journ. 21, 791.(1927a). Digestion in the cockroach. II. The digestion of carbohydrates. Biochem. Journ.
21, 797-(1928). Digestion in the cockroach. III. The digestion of proteins and fats. Biochem. Journ.
.22, 150.(1929). Digestion in the tsetse-fly; a study of structure and function. Parasitol. 21, 288.
WOLLMAN, E. (1911). Sur l'elevage de mouches st£riles. Contribution a la connaissance du role desmicrobes dans les voies digestives. Ann. Inst. Pasteur, 25, 79.(1921). La methode des elevages aseptiques en physiologie. Arch. Internat. Physiol. 18, 194.(1922). Biologie de la mouche domestique et des larves de la mouche a viande, en elevages
aseptiques. Ann. Inst. Pasteur, 36, 784.





























