Embed Size (px)
Citation preview

Freshwat. Biol 1972, Volume 2, pages 289-307
Studies on Gull Lake, MichiganL Seasonal and depth distribution of phytoplankton*
BRIAN MOSS Department of Botany and Plant Pathology, and W. K. KelloggBiological Station, Michigan State University
Manuscript accepted 13 July 1972
SummaryThe seasonal and depth distribution of phytoplankton in Gull Lake, a moderatelylarge, temperate, dimictic, recreational lake in southern Michigan, U.S.A., is describedand discussed. Diatoms (Fragilaria crotonensis, Cyclotella michiganiana, Asterionellaformosa, and Synedra acus) dominate the spring phytoplankton. Populations ofcryptomonads, green flagellates, and non-bloom-forming blue-green algae {Chroo-coccus and Synechococcus) grow mainly in summer. Diatoms grew under the winterice, and objections are lodged to the classic explanation of seasonal growth in temperatelakes, and to a recent hypothesis concerning the 'paradox of the plankton'. Tworaphe-bearing large diatom species were considered to be truly planktonic. A Synecho-coccus species was similar to that described previously only from Loch Leven.
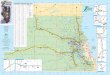
IntroductionGull Lake (Fig. I), one ofthe largest lakes in southern Michigan, is a popular recrea-tional lake. Its shoreline is now almost completely bordered by private houses, manyof them occupied all year, others in summer only.
Increased growth of littoral attached Cladopbora has been reported in recentyears, and minor blooms of blue-green algae have been noted in sheltered bays,particularly at the southern end ofthe lake. Evidence is presented in Moss (1972) thatthe lake is becoming increasingly eutrophic, and future expansion of the nearbycities of Battle Creek and Kalamazoo into the area may ultimately result in greaterfertilization of the lake. Extensive studies of the lake, based on the W. K. KelloggBiological Station, have now begun. This paper includes the earliest availablesubstantial data on the phytoplankton.
Gull LakeGull Lake lies in Kalamazoo and Barry Counties on the Gull outwash plain, backedby the Kalamazoo moraine. It was probably formed by glacial widening and moraine
* Publication No. 226 from ihe W. K. Kellogg Biological Station.
Correspondence: DrB. Moss, School of Environmental Sciences, University of East Anglia, Norwich,NOR 8SC, England.
19 289

290 Brian Moss
85" 25' W
GULL LAKE
Michigon, U.S.A.
I mile 42°25'N
Kellogg Biof Stn
Fig. 1. Map and location of Gu!l Lake, Michigan. Closed circles indicate places with more thanshoreline housing development.
damming of a pre-existing stream valley. The catchment area consists largely ofcalcium-rich glacial drift and comprises subdued rolling country, 268-305 m abovemean sea level. The lake's surface area is 822 ha, and much of it is over 15 m deep.The deepest portions have about 3! m of water. Inflow is provided by several smallstreams, but springs in the lake bed contribute an unknown amount of water. A singleoutflow, in the south, flows over a natural moraine dam. However, an artificial sluiceon the outflow permits lowering of the lake level by about 25-50 cm in autumn,before expansion during ice formation can damage the protective walls of the lakeshoregardens.
The climate of the area is typically continental for the latitude, though mollifiedby the nearby very large St Lawrence Great Lakes. The average January air temperatureis about — 3-3°C, and the average July air temperature is 22-5°C. Mean annualprecipitation is 86-3 cm (Senninger, 1963).
A partial major ion analysis of the water is shown in Table 1. Calcium andbicarbonate are predominant and may supersaturate the water. Carbonate deposits(marl) are found over much ofthe bottom, mixed, in shallow areas, with organic plantremains. Original fringing reedswamp vegetation has largely been cleared as themargins have been deepened to permit boat access. Typha latifolia L. and Scirpus spp.dominate a rich flora in some small bays and an area near the outlet. Sub-aquaticvegetation is present to depths of about 9 m and includes Oiara spp., Najas flexilis(Willd.) Rostk. & Schmidt, Myriophyllum heterophyllum Michx., Potamogeton spp.,Utricularia sp., Elodea canadensis (Michx.) and Vallisneria americana {Michx.).

Studies on Gull Lake, Michigan. I 291
Table 1. Chemical anjilysis of Gull Lake waters, November 1970(Analyses by Water Quality Laboratory, Michigan State University)
Analysed Result Analysed Result
K+Na+Ca++Mg++pH
O-9mg/I4-4 mg/1
40 0 mg/122-1 mg/17-8
ci-AlkalinityTot. diss'd PNH4-NNO3-N
9-7 mg/13-0 m-equiv./I
0 01 mg/i0 01 mg/i0 21 mg/1
Until the last century, the area was occupied only by the Potowatomi Indians(Senninger, 1963). European colonization began in 1833-34 with the building of gristand lumber mills on the outflow stream (Taube & Bacon, 1952). The pre-existingforest has now been almost completely felled and development has proceeded rapidly,particularly in the post-war period. Most ofthe open land in the area is farmed (dairy,livestock, maize). At points indicated in Fig. 1 house-building has taken place awayfrom the lake as well on the shoreline. At other points a single line of houses is present,many of them with lawns edging the lake. There is no central sewerage system, eachhouse having its own dry well, septic tank or drainage field, sometimes within 10 mof the lake (Henderson, 1970), and most of the lawns have been heavily fertilizedannually.
MethodsTemperature was measured at metre intervals with a thermistor (Yellow SpringsInstrument Co. model 43TB) over the station shown in Fig. 1, one ofthe two deepestparts of the lake. Measurements were taken on all dates when water samples weretaken (Table 3) and on a few additional occasions in winter.
Water samples were taken with a non-metallic 31 Van Dorn closing samplerusually at depths of 0, 5, 10, 15, 20, 25 and 30 m. During summer additional depths(variously 7, 8, 9, 11, 13 m) were sampled.
pH of each water sample was determined electrometrically on return from thefield, and weak acid salts (alkalinity) were measured by titration with dilute strongacid to pH 4-5. Glass stoppered bottles were completely filled, without bubbling, inthe field, where manganous sulphate and Winkler's reagent were added. Dissolvedoxygen content ofthe water was subsequently determined by the unmodified Winklermethod (American Public Health Association, I960).
A Wild inverted microscope was used for counting of phytoplankton preservedwith Lugol's iodine solution between December 1968 and July 1969. For the remainderof the period samples filtered through 'Millipore' HA filters (0 45 fixn pore size),subsequently cleared with cedar wood oil, were counted. Counts of ail but veryinfrequent species lie within ±10% ofthe true value with a 95% probability.
Results(a) Physico-chemical measurementsTemperature data are available from October 1968 until December 1969 (Fig. 2).Gull Lake is a dimictic deep lake typical of temperate continental regions (Hutchinson& Loffler, 1956), with two periods of mixing, and two of thermal stratification eachyear. It was isothermal from the beginning of November 1968 until late December

292 Brian Moss
cr>
o o o
Q
<• O

Studies on Gull Lake, Michigan. I 293
1968. Ice then covered the whole lake until late March 1969, with inverse temperaturestratification occurring beneath the ice. The water temperature rose to about 2°Cjust above the sediment surface. When the ice melted, mixing began at about 3°Cand the temperature rose for about a month before increasing insolation createddirect stratification in late April. Stratification persisted until late October, as it hadin 1968, when mixing occurred followed by the beginnings of inverse stratificationin late December. Ice was then beginning to form at the margins of the lake, but notover the sampling station. The summer epilimnion was about 9 m deep and thehypolimnion began at about 13 m. Hypolimnion temperature increased from S^C toICC during the summer, and surface temperatures of nearly 26°C were noted in July.
For the period June-August 1969, the rate of temperature increase {ddjdt), expressedlogarithmically, v^as linear with depth between 11 m and 16 m. For this clinolimnionstratum where, by definition (Hutchinson, 1941), the rate of heating falls exponentiallywith depth, the coefficients of turbulence (coefficients of eddy conductivity) {A) havebeen calculated using the procedure of Hutchinson (1941) applied to the equations ofMcEwan (1929). Log {B-C), where ^ = mean temperature, and C= virtual temperatureof isothermal circulation prior to development of stratification (for Gull Lake,C - 8 4X) . was also linear with depth from 11 m-I6 m, and parallel to the graph oflog ddjdt. This confirms that the application of McEwan's equations for the calculationof A was valid. Table 2 gives the calculated values of A.
Table 2. Coefficients of turbulence {A) for the clinolimnion of Gull Lake (June-Aug. 1969)
Depth (m)
111213141516
dBldt X/30 days
2-11-00-60-40-170-11
-4g.crm-i.s-iX 10^
0-520-490-500-580-430-47
A g.cm"*.s~^correctedfor molecular thermal
conductivityX102
0-400-370 380460-310 35
Depth-time distribution of dissolved oxygen is shown in Fig. 3. During theautumnal and vernal periods of mixing, dissolved oxygen levels were constant between9 and 13 mgOg/l throughout the water column. After direct stratification began inlate April 1969, surface concentrations decreased slightly, owing to the reducedcapacity of warm water to dissolve oxygen, and the hypolimnion concentrationsdecreased progressively. In late September and early October water below 25 mwas anoxic. Data are not avaiiabie for late October 1969, but in 1968 anoxia extendedup to 15 m in the water column for a short period just prior to turnover. Markeddepletion of oxygen was not observed during the winter stratification.
Summer oxygen depletion was undoubtedly due to heterotrophic respiration.Occasional light penetration measurements in summer 1969 showed that the euphoticzone extended to no more than 16 m, and below this depth net oxygen production byphotosynthesis would, by definition, be impossible.

294 Brian Moss
Seasonal distributions of alkalinity and pH are shown in Figs. 4 and 5. Changeswere relatively small, as would be expected from the buffering ability of the relativelylarge amounts of bicarbonate present (Hutchinson, 1957). During isothermal periodsthe alkalinity was about 3 m-equiv./l and the pH about 8-0. In the epilimnion pHincreased during summer to 8-4-8-5 whilst alkalinity decreased to about 2-7 m-equiv./l.Concomitantly, the hypolimnion alkalinity increased to about 3-3 m-equiv./l andthe pH decreased to 7-4. These changes are those expected from shifts in the
Gull Lake 1968-69 Dissolwed oxygen [mq/l
30
I Oci I Nov I Dec I Jon I Feb I Mor I Ac May I Jun I Jul I Agq i Sep.
Fig. 3. Depth-time distribution of dissolved oxygen. Sampling dates on which this figure andsubsequent figures are based are given in Table 3, and the legend to Fig. 1.
Gull Lake 1 9 6 8 - 6 9 Alkolmity /ll
10
n
Ocl I Nov I Dec Jan Feb War Apr May Jun Jul, AugH Sep
Fig. 4. Depth-time distribution of alkalinity.

Studies on Gull Lake, Michigan. I 295
Gull Lake 1968-69 pH
F ^ . 5. Depth-time distribution of pH.
bicarbonate-pH buffer system resulting from photosynthetic withdrawal and respira-tory production of carbon dioxide (Hutchinson, 1957). Although photosynthesis mayhave been most intense in April and May (see below), the turbulence of the watercolumn would permit equilibration with atmospheric CO2 and net changes inalkalinity and pH were consequently not observed.
(b) PhytoplanktonParticularly in spring, diatoms were predominant in the phytoplankton of Gull Lake.Figures 6, 7, 8, 9 and 10 show the depth-time distributions of the most abundantdiatom species, and Table 3 the seasonal occurrences of less numerous species.Table 4 gives all species, including several more diatoms, of sporadic occurrence inthe plankton. Fragilaria crotonensis Kitton (Fig. 6) was the most abundant diatomspecies during 1969. and was detected in the water column on all sampling dates.It was present in relatively large numbers (about 1000 cells/ml) under the winter ice,but the population increased about threefold during the isothermal April period.With direct thermal stratification it sank from the epiiimnion, but persisted in thehypolimnion until early August, when the region of the thermocline only still containedup to 100 cells/mi. Growth in the epilimnion began in late August and a populationof 500ceIls/mI had built up by overturn. Further growth during the mixing periodproduced the overwintering population of 1000 cells/ml.
Asterionella formosa Hass. (Fig. 7) underwent three periods of growth, though itsmaximum population was not large compared with those in other lakes. The firstperiod occurred entirely during the ice covered phase. A maximum of 600 cells/mldeveloped at about 15 m, but disappeared before the ice melted. A lesser growth peak,under directly stratified conditions, occurred at 12 m in July with up to 350 cells/mlbeing recorded. Thirdly, populations of up to 600 cells/ml were formed during theearly part of the autumnal mixing period. Between these growth periods there were

296 Brian Moss
Gull Lokei9fi8-69 Fragiiano cra'onens'S
Dec I Jon t Feb I Mor I fipr | Moy I Jun I M. I Aug I Sep I Ocl I Nov i Dec
Fig. 6. Dcplh-timc dislribution of Fragitaria crotonensts. In this and subsequent figures, horizontal
lines at {op of figure indicate periods of stratification, and vertical lines, periods of mi.xing.
Gull Lake iggg-gg Jisierionella formosa kells/ml)
Dec I Jon I Feb I War I Apr I May I Jjn I Jui
Fig. 7. Depth-time distribution of Asterionella formosa.
interludes when Asterionella was not detected. In practice, this meant that fewerthan 3-5 cells/ml were present.
A third major diatom species, Cyclotella michiganiana Skv. (Fig. 8) increased innumbers under the winter ice. though more slowly than it did after melting of the

Studies on Gull Lake., Michigan. I 297
ice and mixing of the water. Net population increase ceased when the lake stratifieddirectly. Some growth took place in !ate summer, but only 100 celis/ml were produced.In November growth began again, after a decrease in population during October.
A Synedra acus Kutz population began to grow towards the end of winterstratification, but accomplished most growth during the spring mixing (Fig. 9).The population then progressively decreased. A spurious growth maximum of 2000
GulILoke 1968 — 69 Cyc'ofeHa michiganiono [cells
30
I Dec I Jon I Feb I Mor I Apr I Moy I Ju i I Jiji I Aug ) Sep I Oci I Nov I Dec
Fig. 8. Depth-time distribution of Cyciotella michiganiana.
Gull LaHe 1966 —69 Syredraacus rx'ti Synecfiococcus
10
20
30
Synedra Synezhococcus
..-1000'
% (^2000 N.
V
100
j.,'m I [->& I Mof I Apr I Moy I Jun I Jul I Aijq I Sep I Ocl I Noi/ I Dec
Fig. 9. Deplh-time distribution of Synedra acus (solid lines) and Synechococcus sp. (dotted lines).

298 Brian Moss
Gull Loke 1968-69 Cydolello comta (cells/ml)
Aug I Sep > Ocl 1 Nov I OccDec I Jcjn I Feb I Ma' I Apt I May I Jun I J j l
Fig. 10. Depth-time distribution of Cyclotella comta.
GulILoke I 9 & e -
I >tar. I Fet3 I Mar I Ap' I May I Jun I Jul AUQ I Sep [ Ocl I Nov I Dec
Fig. II . Depth-time distnbution of Dinobryon divergens.
or more cells/ml in mid-May probably reflects delay in sinking at the thermoclinerather than continued net population increase after stratification began. No cellswere detected in the water column from then on.
Cyclotella comta (Ehr.) Kutz. underwent most of its population growth in summer,producing a maximum of 156 cells/ml at 13 m in mid-July (Fig. 10).

Studies on Gull Lake, Michigan. I 299
Table 3. Seasonal distribution of infrequent species in the Gull Lake phytopiankton, Dec. 1968-Dec.1969. Mean ceil or filament (Anahaemi) numbers per ml for the epilimnion during summer stratification,and of the whole water column at the oiher times
Dec. Jan. Feb. March April MayDate 12 9 23 6 21 8 27 11 25 1 8 16 23 31
Melosira italicaRhizosolenia eriensisAmphiprora ornataCymatopleura solea apic.Synedra sp.Ceratium hirundinellaAnabaena flos-aquae
3 . , _ _ _ _ ^ . ]8 -6 — 3 8 5 5 2 8 5 5 5 4 — —
_ _ _ _ _ _ _ _ 5 4 4 4 4 __ _ _ _ _ _ _ 3 5 5 4 _ _ _
— 8 13 5 11 — 25 30 11 34 4 — 7 7
DateJune July August
5 13 20 28 5 II 19 25 2 9 16 23 30
Melosira italicaRhizosolenia eriensisAmphiprora ornataCymatopleura solea apic.Synedra sp.Ceratium hirundinellaAnabaena flos-aquae
— 3 3 1 0-5 2 4 ]-5 2 2 0 7 1 11 — — — — — 1 _ _ 2 3 3 —
DateSeptember October November December
6 13 20 27 4 15 29 5 13 25 2 13 17 23
Melosira italicaRhizosolenia eriensisAmphiprora ornataCymatopleura solea apic.Synedra sp.Ceratium hiiumiinellaAnabaena flos-aquae
— — 25 25 13 4 3 41 — 1 1 1 0 7— —— 1 1 1 — 1 1
1 — 1 1 1- 2-5- 2 I
12 8 9 1 I I
Of the less abundant diatoms {Table 3), Melosira italica (Ehr.) Kiitz. was confinedto the autumn mixing period, and Amphiprora ornata Bailey, and Cymatopleura solea(Breb) W. Smith var. apiculata (W. Smith) Ralfs. occurred during the spring mixingperiod. Rhizosolenia eriensis H. L. Smith and a large Synedra sp. were present duringwinter and spring.
The only member of the Chrysophyta present abundantly in the open water wasDinobryon divergens Imhof. (Fig. 11), though D. bavaricum Imhof., Chrysosphaerellalongispina Lauterborn and four species of MaUomonas occurred very infrequently.Like those of some diatoms, populations of Dinobryon divergens grew slowly inwinter, undergoing about one cell division in 3 months, but then grew more rapidlyduring the month of spring mixing, after which they decreased. A minor increaseoccurred in June and an even smaller one in November.
Three cryptomonads were abundant members of the phytopJankton in 1969.Cryptomonas sp. (probably C. erosa Ehr.) (Fig. 12) was present at levels of about50 cells/ml during winter and spring, but increased during June and July to formmaximum populations around the thermocline. This vertical pattern was preserved.

300 Brian Moss
Gull Loke 1968 —59 Cryptomonris sp (cel ls /mi)
Mor I Apr I Moy I Jun I Jul I Aug I 5ep I Oct I Nov I Dec
Fig. 12. Depth-time distribution of Cryptomonas sp.
Gull Loke I9b8-b'3 Rhodomonas minulafrARhodomanas SD
30
Fig. 13,lines).
Dec 1 Jon I Feb I ^•D- I Apr I May I Jun I Jul I Auq I ^ep I Oc' I Nov I Dec
Depth-time distribution of Rhodomonos minula {solid lines) and Rhodomonas sp. (broken
more or less, until the autumn mixing, although numbers decreased. A further,equally large., growth and decline occurred after overturn. Cells were then evenlydistributed through the water column. Different varieties or physiological states ofthe same species may have constituted the summer and autumn populations, althoughmorphologically they appeared indistinct. The summer form was usually maroon

Studies on Gull Lake, Michigan. I 301
Gull Loke I 9 G 8 - 6 9 Green flagellates (cel ls/ml)
Dec. I Jan I Feb I Ma I Apr I May I Jan I Ju Auq I Sep I Oci I Nov I Dec
Fig. 14. Depth-time distribution of green flagellates.
Gull Lake 1968-59 Chroococcus di^ersus (cel!s/ml)
Dec I Jan I Feh I Mor I Api I Moy I Jun I Ju
30
Fig. 15. Depth-time distribution of Chroococcus dlspersus.
when preserved with iodine solution, whilst the autumn form was greenish. Rhodomonasminuta var. nannoplanctica Skuja (Fig. 13) was most abundant at 10 m and belowunder the winter ice, declined as temperature increased, and then increased in numbersduring summer stratification. Again its maximum populations were not found at thesurface, but at about 10 m. It disappeared from the water column in August, but

302 Brian Moss
produced a small population in October. In late August and September a secondRhodomonas sp., less than 4^m long, formed relatively large populations (Fig. 13)between 13 and 15 m, but then decreased in numbers.
Small (<5 ^m) chlamydomonad green flagellates were present in winter, summerand autumn (Fig. 14), but proved unidentifiable. Probably different species wereinvolved at different times. Under the ice they were most abundant towards thesurface, whilst in summer they were most frequent in the lower part of the epilimnion.A population present in the early part of the autumn mixing phase was evenly distri-buted with depth. Other than these flagellates. Chlorophyta were scarce. Somefourteen species were recorded (Table 4), usually as isolated individuals. Desmidswere notably absent, a single cell of Cosmarium sp. being recorded on one occasionin August. Euglenophyta were also extremely infrequent.
Table 4. Species occurring sporadically in Gull Lake phytoplankton 1969
CyanophytaAnabaena inequalis (Kiietz.) Bom. et Flah.Mkrocystis aeruginosa Kuetz., emend.
ElenkJnOscillatoria sp.
ChlorophytaTetraedron mimmum (A. Braun) Hansgirg.Svenedesmus sp.Crucigenia qiiadrciia MorrenOocyslis soltlaiia Wittr.Butryococcus biaunii Kuetz.Ankisirodesmus sp.Cosmarium sp.Micractiniuni pusillum Fres.Quadrigula sp.Eitdorina elegans Ehr.Scenedesnuts bijuga (Turp.) Lag.Dictyosphaerium ehrenbergianum Naeg.Pediasfrum sp.
EuglenophytaPhacus sp.Tracheloinonas volvochm Ehr.
ChrysophytaDinobryon bavaricum ImhofChrysosphaercUa longispina Laut.Mallomonas caudata IwanoffAf. dentata ConradM. ehngala ReverdinM. mirabilis Conrad
BacillariophytaStephanodiscus niagarae Ehr.Surirella angiista KiJtz
PyrrophytaPeridinitim (2 spp.)Cynmodmum sp.
Pyrrophyta were numerically scarce. Up to four cells/ml of Ceratium hirundinella(O. F. Muell.) Dujardin (Table 3) were generally detected in the epilimnion fromJune until August, and three other dinoflagellates were even less frequent (Table 4).
Blue-green algae were represented largely by Chroococcus dispersus (Keissl.) Lemm.(Fig. 15) and a species of Synechococcus (Fig. 9). C. dispersus grew in the epilimnionin late summer, and declined in numbers towards the end of the stratified period.The Synechococcus sp. population also grew in mid-summer, forming a well demar-cated maximum in the thermocline region in July. It became less abundant thereafter,and, following redistribution of the remaining population on mixing of the watercolumn, eventually disappeared.

Studies on Gull Lake, Michigan. I 303
Other blue-green algae were uncommon, despite observations of surface bloomsat the southern end of the lake. Anabaena fios-aquae (Lyngb.) De Brebisson, the formerof these blooms, was detected during most of the summer, though most abundantlyafter autumn overturn (Table 3). Two other potential bloom formers, Anabaenaitiequalis (Kuetz.) Born, et Flah., and Microcystis aeruginosa Kuetz. emend. Elenkinwere sporadic in occurrence.
DiscussionThe seasonal periodicity of the Guli Lake phytopiankton broadly eorresponded withthe classic pattern for large temperate lakes. A major growth of diatoms in springwas accompanied, in its later stages, by growth of Dinobryon. Green algae grew insummer, and were aceompanied by blue-green algae in late summer and autumn.A moderate growth of diatoms also occurred in autumn. With some similar, thoughmany different, species a similar pattern is reported by Lund (in Macan, 1970) forWindermere, by Birge & Juday (1922) for Lake Mendota, and by Rodhe, Vollenweider& Nauwerck (1958) for Lake Erken.
The usual explanation (Hutchinson, 1967) for this pattern is that increasing lightavailability stimulates growth of cold water diatoms in spring, and that Dinobryon.,being inhibited by all but very low phosphate concentrations, increases in numberswhen the growth of diatoms has lowered the phosphate content of the water. Greenand blue-green algae with, respectively, higher and yet higher temperature optimafor growth, grow as the water warms, and diatom numbers increase again as thewater cools towards the end of the year.
Although this explanation seems to fit much presently available data, aspects ofthese Gull Lake data emphasize that it is much over-simplified. Most diatom growthdid occur in spring, but a complete cycle of growth and decline o^ Asterionella formosatook place during January and February when both temperature and light were mini-mal. Growth of Cyclotella michiganiana also occurred at that time, four cell divisionsbeing undergone in 9 weeks.
Rhodomonas minuta var. nannoplanctica and a green flagellate also grew under theice, preceding the spring growth of diatoms often assumed to be the 'start' of theannual phytopiankton cycle.
Growth of Dinobryon dirergens was not subsequent to that of the diatoms, as inthe classic model, but was simultaneous during the mixing period. The explanationthat nutrient depleted conditions are necessary for Dinobryon growth is, therefore,probably not tenable for Gull Lake.
Both Wright (1964) and Pennak (1968) report considerable phytopiankton activityunder winter ice. Many flagellates, including Rhodomonas minuta var. nannoplanctica,are listed from Beaver Pond, Massachusetts, by Wright but diatoms, although present,were minor components of the under-ice phytopiankton. Turbulence under ice isminimal and motile flagellates seem well adapted to such conditions. Non-motile,relatively heavy diatoms, however, are not and the persistence of large populations ofFragilaria crotonensis, and the marked growth of populations o{ Asterionella formosaand of Cyclotella michiganiana under the ice is puzzling. Some source of turbulenceis necessary to maintain these organisms in suspension. Winter heating from solarradiation and from summer heat stored in the sediment are obvious sources, but thesedo not maintain large diatom populations under ice in other lakes. The inversestratifications in Wintergreen Lake and in Lawrence Lake (Manny, 1971), water

304 Brian Moss
bodies near Gull Lake, appear rather more stable than that in Gull Lake, since thehorizontal components of the winter isotherms in the former lakes are greater thanthose in Gull Lake.
Gull Lake is fed by springs in its bed. These were noted by Taube & Bacon (1952),and were observable over part of the lake's edge when a temporary coffer dam wasconstructed to aid building of a dock in 1971. Turbulence from springs was not clearlydetected when coefficients of turbulence were calculated for the summer heatingperiod (Table 2). These values are, in fact, relatively low for the area of the lake, beingonly just larger than those calculated by Hutchinson (1941) for the much smallerLinsley Pond (9 4 ha) and by Newcombe & Dwyer (1949) for the smaller Sodon Lake(94 ha). In general, values of/) increase with area of lake. In summer, wind inducedturbulence was thus much greater than any spring induced turbulence but it is possiblethat the latter, coupled with turbulence produced by inflow streams, may have beenresponsible for maintenance of the winter diatom populations.
The simultaneous growth of four species—Fragilaria crotonensis, Synedra acus,Cyclotella michiganiana and Dinobryon divergens—in the circulating water columnin spring, suggests that Hutchinson's fourth explanation (1967. p. 358) of his 'paradoxof the plankton' (Hutchinson, 1961) is the most likely of his four alternatives. Theparadox is that more than one species exists in the phytopiankton in what appearsto be, in a mixing water column, a single niche. The two most reasonable generalexplanations of this are that the water column consists of a three-dimensional mosaicof niches (Richerson, Armstrong & Goldman, 1970) or that competitive displacementand attainment of equilibrium do not occur before the single spatial niche changes intime. Under stratified conditions many niches exist in the water column, even in thesupposedly mixed epilimnion where microstratifications may persist for several daysduring calm weather (Lund, 1964). Development o^ Synechococcus sp., Cryptomonassp. and Rhodomonas spp. at intermediate depths in the summer water column ofGull Lake illustrate the existence of specialized niches. At the thermocline particulateorganic matter tends to collect and bacterial activity may be high resulting, perhaps,in production of exogenous vitamins necessary for growth of at least the cryptomonads(Pringsheim, 1968). Graphic examples of intricate vertical distribution may be foundin small fertile lakes (Moss, 1969; Baker & Brook, 1971). However, the many nichehypothesis is untenable for the spring mixing period. Richerson et al. (1970) postulatethat patches of water in a lake differ for long enough for different species to developbefore the patches eventually mix. This 'contemporaneous disequilibrium' model issupported by demonstration of horizontal patchiness in collections of suspendedalgae from the small (20 ha) Castle Lake, California. More than half of the specieslisted by Richerson et al. are either typically epipelic (mud-living) or epiphytic, andit seems not unlikely that much of the patchiness they observed may have dependedon proximity of some samples to weed beds or disturbed shores, rather than todifferential growth of species suspended in the water. Data from Gull Lake, wheresimultaneous growth occurred over a month's period during mixing conditions,suggest either that competitive exclusion has not time to occur, or, as suggested byFryer (1959) for species flocks of cichlid fishes in Lake Nyasa (L. Malawi), that theseveral species are not in competition. Fryer's fishes shared a common but veryabundant food supply (epilithic algae). When growing phytopiankton populationsare quite small and not self-shading, and when the nutrient supply is adequate, as isprobably the case for a time in spring as the reservoir of nutrients built up during the

Studies on Gull Lake, Michigan. I 305
winter is being used up, competition may not occur between the various algalspecies.
Pennate diatoms, other than those of the Araphidineae, are not normally com-ponents of the phytopiankton of large temperate lakes. Individuals of raphe-bearingspecies may be introduced into the plankton by wave disturbance of the weed bedsand sediments, where typically they are found (Round, 1964), but then they occuronly sporadically. In the open water of Guli Lake, where the samples were taken,typical benthic genera such as Nitzsehia and Navicula were only very rarely en-countered, and then only in net samples. Two Biraphidinean species, however, occurredwith such consistency, and in the absence of typically non-planktonic species, thatthey must be considered truly planktonic. These were Amphiprora ornata and Cymato-pleura solea var. apiculata, and were frequently present in net samples when notdetected in counts on whole water samples. A number of raphe-bearing species,including C. solea var. apiculata, are hsted as planktonic by Huber-Pestalozzi (1942),but definite evidence that they are truly planktonic, and not resuspended benthicforms, is not given. Cymatopleura solea and Amphiprora ornata are listed as planktonicfrom L. Michigan by Stoermer & Kopczynska (1967), though Stoermer & Yang(1969) consider A. ornata to be primarily a benthic species. Truly planktonic raphe-bearing species, including Nitzsehia spp. and Surirella spp., are abundant in tropicalAfrican lakes (Richardson, 1968; Tailing, 1957) and are also listed in Huber-Pestalozzi(1942). Like the Gull Lake Amphiprora (80-100/^m long) and Cymatopleura (about150 ̂ m) they tend to be large, and among planktonic members of the genus Surirella,the twisted form exhibited by A. ornata and the slipper shaped form of C. solea, arecommon. These shapes may confer resistance to sinking in these large, and presumablyheavy, planktonic species.
The blue-green algal populations in Gull Lake are not yet well developed, and itis clear that reports of blooms of Anabaena in the south-eastern arm of the lake nearBay View (Fig. 1) result from local hydromechanical concentration, during calmweather, of the gas vacuolate filaments, rather than from vigorous growth (Reynolds,1971). Even when the main body of the lake is quite rough, the south-eastern armremains very calm. Observations in 1971 showed that average concentrations ofAnabaena in the water column were no greater in this arm than in the main body.
The Synechococcus observed is remarkable since it seems closely similar to theSynechococcus first reported from Loch Leven by Bailey-Watts, Bindloss & Belcher(1968). The ceils compare in being almost colourless straight rods, generally about50 fim long by just under 1 fim wide. The Gull Lake cells tend to be slightly turnedto one side at one or both ends. Synechococcus sp. was predominant in the summerphytopiankton of the eutrophic Loch Leven, with several million cells per ml. Althoughnot nearly so abundant in Gull Lake, increasing fertilization may result in increasesof this and other blue-green algae. This possibility is discussed by Moss (1972).
AcknowledgmentsDr G. H. Lauff, Director of the W. K. Kellogg Biological Station, encouraged thecarrying out of this work and also made available necessary boats and samplingequipment, and the services of Mr Robert Keene, who carried out much of the fieldsampling. I am grateful also to Dr R. G. Wetzel for the use of an inverted microscope,to Mr W. Sedgefield White for his assistance in the field during inclement weather, andto my wife, Miss L. J. Simlett, for criticizing and typing the manuscript. Finance for
20

306 Brian Moss
laboratory supplies was provided by a grant from the U.S. Dept. of the Interior.I am grateful to the Science Research Council (U.K.) for support during preparationof the manuscript.
ReferencesAMERICAN PUBLIC HEALTH ASSOCIATION. (1960) Standard Methods for the Examination of Water and
Waste-water, Hth edn. New York.BAILEY-WATTS A.E., BINDLOSS M . E . & BELCHER J .H. (1968) Freshwater primary production by a
blue-green alga of bacterial size. Nature, Lond. 220,1344-1345.BAKER A.L . & BROOK A.J . (1971) Optical density profiles as an aid to the study of microstratified
phytopiankton populations in lakes. Arch. Hydrobiol. 69, 214-233.BiEGE E.A. & JUDAY C. (1922) The inland lakes of Wisconsin. The plankton. 1. Its quantity and
chemical composition. Bull. Wis. geol. nat. Hist. Surv. 64 (Sci. Ser. 13), 222 pp.FRYER G. (1959) The trophic interrelationships and ecology of some littoral communities of Lake
Nyasa with especial reference to the fishes and a discussion of the evolution of a group of rockfrequenting Cichlidae. Proc. zool. Soc. Lond. 132, 153-281.
HENDERSON J. (1970) Gull Lake Investigations. 1969 Studies, Part 1. Demographic Survey, pp. 1-17.Unpub'd report to Kalamazoo Nature Center.
HUBER-PESTALOZZI G. (1942) Das Phytopiankton des Siisswassers 2(2) Diatomeen. E. Schweizer-bart'sche Verlagsbuchhandlung, Stuttgart.
HUTCHINSON G.E . (1941) Limnological studies in Connecticut IV. The mechanism of intermediarymetabolism in stratified lakes. £col. Monogr. 11, 21-60.
HUTCHINSON G.E. (1957) A Treatise on Limnology 1. Wiley, New York.HUTCHINSON G.E. (1961) The paradox of the pianlcton. Am. Nat. 95, 137-146.HUTCHINSON G.E. (1967) A Treatise on Limnology 2. Wiley, New York.HUTCHINSON G.E. & LOFFLER H . (1956) The thermal classification of lakes. Proc. natn. Acad. Sci.
U.S.A. 42, 84-86.LUND J . W . G . (1%4) Primary production and periodicity of phytopiankton. Verh. int. Verein. theor.
angew. Limnol. 15, 37-56.MACAN T.T. (1970) Biological Studies of the English Lakes. American Elsevier, New York.MANNY B.A. (1971) Interactions of dissolved and particulate nitrogen in lake metabolism. Ph.D. thesis,
Michigan State University.McEwAN G.F. (1929) A mathematical theory of the vertical distribution of temperature and salinity
in water under the action of radiation, conduction, evaporation, and mixing due to the resultantconvection. Bull. Scripps Instn Oceanogr. tech. Ser. 2, 197-306.
Moss B. (1969) Vertical heterogeneity in the water column of Abbot's Pond. II. The influence ofphysical and chemical conditions on the spatial and temporal distribution of the phytopianktonand of a community of epipelic algae. J. Ecol. 57, 397-414.
Moss B. (1972) Studies on Gull Lake, Michigan. 11. Eutrophication—evidence and prognosis.Freshwat. Biol. 2, 309-320.
NEVt-coMBE C.L. & DWYER P.S. (1949) An analysis of the vertical distribution of temperature in adichothermic lake of southeastern Michigan. Ecology, 30, 443-449.
PENNAK R .W. (1968) Field and experimental winter limnology of three Colorado mountain lakes.Ecology, 49, 505-519.
PRINGSHEIM E.G. (1968) Zur Kenntnis der Cryptomonaden des Siisswassers. Nova Hedwigia, 16,367-401.
REYNOLDS C.S. (1971) The ecology of the planktonic blue-green algae in the North Shropshire meres.F/rf5/M£/. 3, 409-432.
RICHARDSON J .L. (1968) Diatoms and lake typology in East and Central Africa. Int. Revue ges.Hydrobiol. Hydrogr. 53, 299-338.
RICHERSON P., ARMSTRONG R. & GOLDMAN C.R. (1970) Contemporaneous disequilibrium, a newhypothesis to explain the'Paradox of the Plankton'. Prac. natn. Acad. Sci. U.S.A. 67,1710-1714.
RODHE W., VOLLENWEIDER R .A. & NAUWERCK A. (1958) The primary production and standingcrop of phytopiankton. In: Perspectives in Marine Biology (Ed. by A. Buzzati-Traverso), p. 299-322. University of California Press, Berkeley.

Studies on Gull Lake, Michigan. I 307
ROUND F .E . (1964)Theecology of benthic algae. In: Algae and Man (Ed. by D. F. Jackson), pp. 138-184. Plenum, New York.
SENN[NGER E.J. Jr. (1963) Atlas of Michigan. Flint Geographical Press, Flint, Michigan.STOERMER E .F . & KOPCZYNSKA E. (1967) Phytopiankton populations in the e.\treme southern basin
of Lake Michigan, 1962-1963. Ct. Lakes Res. Div'n Spec. Rep. 30, 19^6.STOERMER E.F. & YANG J.J. (1970) Distribution and relative abundance of dominant plankton diatoms
in Lake Michigan. Gt. Lakes Res. Div'n Pufn 16, 1-64.TALLING J . F . (1957) Some observations on the stratification of Lake Victoria. Limnol. Oceanogr.
2,213-221.TAUBE CM. &. BACON E .H . (1952) Gull Lake. Lake Inventory Summary No. 2. 4 pp. Michigan Depart-
ment of Conservation.WRIGHT R.T . (1964) Dynamics of a phytopiankton community in an ice-covered lake. Limnol.
Oceanogr. 9, 163-178.