Embed Size (px)
Citation preview

Strigolactone signaling is required for auxin-dependentstimulation of secondary growth in plantsJavier Agustia, Silvia Herolda,1, Martina Schwarza, Pablo Sancheza, Karin Ljungb, Elizabeth A. Dunc, Philip B. Brewerc,Christine A. Beveridgec, Tobias Siebererd,2, Eva M. Sehra,3, and Thomas Greba,4
aGregor Mendel Institute of Molecular Plant Biology, Austrian Academy of Sciences, 1030 Vienna, Austria; bUmeå Plant Science Center, Department of ForestGenetics and Plant Physiology, Swedish University of Agricultural Sciences, SE-901 83 Umeå, Sweden; cSchool of Biological Sciences, University of Queensland,St. Lucia QLD 4072, Australia; and dMax F. Perutz Laboratories, Department of Microbiology, Immunobiology, and Genetics, University of Vienna, 1030Vienna, Austria
Edited by Athanasios Theologis, Plant Gene Expression Center, Albany, CA, and accepted by the Editorial Board November 8, 2011 (received for reviewJuly 21, 2011)
Long distance cell-to-cell communication is critical for the de-velopment of multicellular organisms. In this respect, plants areespecially demanding as they constantly integrate environmentalinputs to adjust growth processes to different conditions. Oneexample is thickening of shoots and roots, also designated assecondary growth. Secondary growth is mediated by the vascularcambium, a stem cell-like tissue whose cell-proliferating activity isregulated over a long distance by the plant hormone auxin. Howauxin signaling is integrated at the level of cambium cells and howcambium activity is coordinated with other growth processes arelargely unknown. Here, we provide physiological, genetic, andpharmacological evidence that strigolactones (SLs), a group of planthormones recently described to be involved in the repression ofshoot branching, positively regulate cambial activity and that thisfunction is conserved among species. We show that SL signaling inthe vascular cambium itself is sufficient for cambium stimulationand that it interacts strongly with the auxin signaling pathway. Ourresults provide a model of how auxin-based long-distance signalingis translated into cambium activity and suggest that SLs act asgeneral modulators of plant growth forms linking the control ofshoot branching with the thickening of stems and roots.
meristem | wood production | MORE AXILLARY BRANCHES
Hormone-based long-distance signaling is essential for co-ordinating the growth and activity of different organs and
tissues during the life cycle of multicellular organisms. In par-ticular, plants are demanding in this respect because, due to theirsessile and indeterminate lifestyle, their reproductive successdepends on the competence to adjust their developmental pro-grams to changing environmental conditions and requirements.Auxin is one of the best-characterized hormones involved inlong-distance signaling, regulating a tremendous number of de-velopmental processes. One of these is apical dominance, which,indeed, represents a classical example for long-distance signalingin plants (1). In this case, apex-derived auxin is transported ba-sipetally along the main shoot and this is required for sup-pressing the outgrowth of axillary buds. Another process thatdepends on the basipetal transport of auxin is secondary growth(2). Secondary or lateral growth of growth axes is based on theactivity of the vascular cambium, a meristematic tissue organizedas a cylinder encompassing the center of growth axes. Cambiumactivity leads to the production of secondary vascular tissue,which results in an increase of shoot or root diameter. Theinterruptibility of the process by decapitation of the shoot tip andthe subsequent reversibility by artificial auxin application isshared between apical dominance and secondary growth (1, 2).Even though both processes thus share upstream processes and,presumably, regulators, how basipetal transport of auxin istranslated into two different outputs and to what degree they areinterconnected is unknown.Significant progress has been made in the understanding of the
molecular control of apical dominance by the characterization of
the strigolactone (SL) biosynthesis and signaling pathway. SLs, acarotenoid-derived group of molecules, show all attributes ofplant hormones, meaning that they travel through the plant (3)and are effective at low concentrations (4, 5). In addition toshoot branching, SLs have been identified as germinationstimulants of parasitic plants and as triggers for the establish-ment of interactions with mycorrhizal fungi, in which contextthey were originally identified (reviewed in ref. 6). The SL sig-naling pathway has, so far, been defined by a series of four genesfor which homologs have been characterized in Arabidopsis (7–10), pea (11), petunia (12), tomato (13), and rice (14). Also inrice, two additional genes associated with the SL pathway havebeen recently described, indicating that the discovery of SL-re-lated genes is not yet completed (15, 16). The identification ofSL signaling in distantly related species, among them mono- anddicotyledonous species, suggests that this communication toolrepresents an ancient invention and that it is widely distributedwithin the plant kingdom.The function of SL-related genes is characterized best in
Arabidopsis where three of four MORE AXILLARY BRANCHES(MAX) proteins (MAX1, MAX3, and MAX4) have been sug-gested to be involved in the biosynthesis of SLs starting fromcarotenoids. This suggestion is based on their resemblance tocarotenoid dioxygenases (MAX3 and -4) and to cytochrome P450family members (MAX1), on corresponding biochemical activities(7–9) and on SL deficiency of knockout mutants (4, 5, 17). Incontrast to max1, -3, and -4 mutants, max2 mutants are SL in-sensitive with regard to an effect on apical dominance (4, 5, 9),consistent with the idea that MAX2 functions as a receptor for SLsor an unknown downstream product. Moreover, its similarity to F-box leucin-rich repeat proteins and its physical interaction withcompounds of Skp/Cullin/F-box (SCF) E3 ligase complexes sug-gests that MAX2 functions as an SL-dependent recruiting factor forproteins destined for proteasomal degradation (10).max2 mutants (which were also isolated as ore9) (18) display
various growth alterations in addition to a decrease of apicaldominance, like delayed leaf senescence (18, 19), altered leaf
Author contributions: J.A. and T.G. designed research; J.A., S.H., M.S., P.S., K.L., E.A.D., P.B.B.,C.A.B., T.S., and E.M.S. performed research; J.A. and T.G. analyzed data; and J.A. and T.G.wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission. A.T. is a guest editor invited by the EditorialBoard.1Present address: Institute of Chemical Engineering, Vienna University of Technology,Gumpendorferstraße 1a, 1060 Vienna, Austria.
2Present address: Department of Plant Sciences, Technische Universität München–Wei-henstephan, Liesel Beckmann Str. 1, 85354 Freising, Germany.
3Present address: Roche Diagnostics, Engelhorngasse 3, 1211 Vienna, Austria.4To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1111902108/-/DCSupplemental.
20242–20247 | PNAS | December 13, 2011 | vol. 108 | no. 50 www.pnas.org/cgi/doi/10.1073/pnas.1111902108
Dow
nloa
ded
by g
uest
on
June
18,
202
0 D
ownl
oade
d by
gue
st o
n Ju
ne 1
8, 2
020
Dow
nloa
ded
by g
uest
on
June
18,
202
0 D
ownl
oade
d by
gue
st o
n Ju
ne 1
8, 2
020
Dow
nloa
ded
by g
uest
on
June
18,
202
0

morphology (19), and hypocotyl elongation (20). In addition, theMAX2 gene is required for karrikin-induced seed germination inpostfire environments (21) and a role of the SL signaling path-way in the regulation of root system architecture was also de-scribed recently (17, 22, 23). Intriguingly, in contrast to pea andrice (4, 24), SLs are only reduced by 50% in Arabidopsis max1 andmax4 mutants (17). Together, these observations suggest that thebroadness of SL-dependent processes is currently underestimated.One explanation for a broader role of the SL signaling path-
way in various developmental processes is an interactive mode ofaction with the auxin signaling pathway. The interaction betweenboth pathways seems to exist on several levels. On the one hand,the influence of SL signaling on auxin transport has beenreported. max mutants in Arabidopsis display an enhanced ex-pression of PIN1 and PIN3, two members of the auxin effluxtransporter family and show an increase in auxin transport ca-pacity (25–29). These observations indicate that SLs act in-directly on apical dominance by repressing auxin transport,which impedes auxin synthesized in lateral buds to be trans-ported into the main stem (25, 29, 30). On the other hand, SL-dependent inhibition of the outgrowth of side shoots functionsalso in auxin-depleted plants and the application of SLs andauxin transport inhibitors, like N-1-naphthylphthalamic acid (NPA),results in different physiological responses (26). This suggests SL’sfunction to also be downstream of auxin signaling. In fact, MAX4and RMS1, its homolog in pea, are inducible by auxin application(7, 31), and the same holds true for RMS5, the pea homolog ofMAX3 (32). Even though the significance of MAX4 inducibilityhas been questioned (33), these observations indicate that SLbiosynthesis is stimulated by auxin.In this study, we reveal a role of SL signaling in the regulation
of secondary growth and show that the regulation of cambiumactivity is an SL response, which is independent from the regu-lation of apical dominance and conserved among species. On thebasis of genetic and pharmacological data, we propose that SLsfulfill their roles directly in the cambium mainly downstream ofauxin signaling.
ResultsStrigolactone-Deficient Mutants Display a Reduction in SecondaryGrowth. To characterize long-distance regulation of the cam-bium, we studied the effect of SLs on cambium activity in wild-type and max mutant Arabidopsis plants. Cambium activity wasdefined by the lateral extension of the tissue produced by theinterfascicular cambium (IC) immediately above the uppermostrosette leaf (IC-derived tissue; ICD) and by the acropetal pro-gression of IC initiation along the inflorescence stem (34, 35).Quantifying these parameters, we observed that the IC-basedtissue production was decreased by 30% in all max mutants (Fig.1 A–E) and that the progression of IC initiation was reduced onaverage by 40% (Fig. 1F). Consistent with a reduced cambiumactivity in SL-defective plants, qRT-PCR analyses demonstratedlower transcript levels of cambium-specific (35) and cell cycle-re-lated (36, 37) genes in max1-1 stems in comparison with wild type(Fig. S1). Together, these observations demonstrate that plantswith reduced SL signaling or biosynthesis show reduced cambiumactivity.
Local GR24 Treatments Stimulate Cambium Activity. To test whetherthe reduction in cambium activity in max mutants is based ona direct regulation of the vascular cambium by SLs, we treatedwild-type,max1-1, andmax2-1 stems locally with the synthetic SLanalog GR24. Stems were treated and analyzed in the first in-ternode of the main inflorescence stem in a region where, with-out treatment, no IC is established in any of the genotypes usedin this study (Fig. 1G) (34). In contrast to mock treatments, lo-cally applied GR24 induced cell divisions in interfascicularregions of plants of all three genetic backgrounds at the treat-ment site (Fig. 2 A–C). The pattern of cell divisions was similarto that observed during secondary growth initiation at the base ofthe inflorescence stem of untreated plants (Fig. 1 A and B) (34),and the effect was dose dependent (Fig. S2A). max1-1 mutantswere more sensitive than wild type and, consistent with theemployment of the canonical SL signaling pathway, the effectwas significantly weaker in max2-1 (Fig. 2C). Remarkably, max2-1
Fig. 1. Genetic analysis of the role of SL signaling and biosynthesis in cambium regulation. (A–D) Cross-sections from immediately above the uppermostrosette leaf of wild-type (A and B) and max1-1 (C and D) stems. Note that B and D are higher magnification images for areas labeled in A and C, respectively.The extension of the IC-derived tissue (ICD) is indicated by braces (}). [Scale bar in C (100 μm) also applies to A, and scale bar in D (50 μm) also applies to B.]Asterisks indicate the position of primary vascular bundles. (E) Quantification of lateral ICD extension immediately above the uppermost rosette leaf asindicated in A and C. (F) Quantification of the longitudinal progression of IC initiation, as illustrated in D. (G) Scheme of longitudinal IC extension in theArabidopsis stem (red) and the relative position of the treatment zone described in Fig. 2 and Fig. 4.
Agusti et al. PNAS | December 13, 2011 | vol. 108 | no. 50 | 20243
PLANTBIOLO
GY
Dow
nloa
ded
by g
uest
on
June
18,
202
0

plants were not fully GR24 insensitive with respect to the inductionof cambium-like cell divisions, which is in contrast to their in-sensitivity with respect to other processes (5, 23) and suggests thatthere are factors acting in parallel to MAX2 in this case. Collec-tively, these observations show that GR24 can locally stimulatecambium-like cell divisions in the Arabidopsis inflorescence stem.Next, we tested whether GR24-induced cell divisions represent
secondary growth as observed under natural conditions by ana-lyzing the dynamics of cambium- and phloem-specific markers inresponse to GR24 treatments. PHLOEM INTERCALATEDWITH XYLEM (PXY, also known as TDIF RECEPTOR, TDR)is a receptor-like kinase expressed in cambium cells throughoutthe Arabidopsis plant body (38, 39). A reporter line expressing thecyan fluorescent protein (CFP) under the control of the PXYpromoter (PXY:CFP) visualized the fascicular cambium in pri-mary stems and, after GR24 treatments, was detected in inter-fascicular regions indicating GR24-induced formation of theinterfascicular cambium (Fig. 2 D and E). To visualize the for-mation of secondary vascular tissue, we used the promoter of theAPL gene encoding for a MYELOBLASTOSIS (MYB) tran-scription factor specifically expressed in phloem tissues (40).
Initially only active in the phloem of primary bundles, an APL:CFP reporter was also active in clusters of cells in interfascicularregions after stems were treated with GR24 (Fig. 2 F and G),indicating GR24-dependent formation of secondary vasculartissue. To characterize the spatial extension of the GR24-de-pendent effect, we treated two reporter lines expressing theβ-glucuronidase (GUS) gene under the control of the PXY andAPL promoters, respectively. GR24 treatments resulted in theinduction of both reporters in the treatment zone and notextending outwards by more than 0.5 cm (Fig. 2 H and I). Im-portantly, our treatments did not repress the outgrowth of ro-sette branches (Fig. S2B). Together, these results demonstrate alocal induction of the cambium-specific stem cell niche and ofvascular tissue formation by GR24 applications and suggest thata local increase in SL levels is capable of stimulating secondarygrowth in Arabidopsis inflorescence stems independently from aneffect on shoot branching.
Increased Auxin Content and Signaling in SL Mutant InflorescenceStems. Previous reports showed that SL signaling can reduceauxin transport in the Arabidopsis stem (25–27, 29), presumablyby suppressing members of the PIN family of auxin transporters(25, 28, 29). Furthermore, the analysis of auxin-sensitivereporters and measurements of auxin exported out of isolatedstem fragments suggest that auxin levels are increased in maxmutant backgrounds (25, 28). Because auxin is a positive keyregulator of cambium activity (35, 41, 42), we thus analyzedwhether SLs stimulate cambium activity indirectly by repressingauxin transport, which might result in enhanced auxin accumu-lation in certain cell types. Initially, we analyzed pin1-613 andpin3-5 mutants to see how changes in auxin transport capacitiesinfluence cambium activity. Both mutants were chosen becausePIN1 and PIN3 are strongly expressed in stems (Fig. S2C) and, atleast for pin1 mutants, it has been shown that auxin transportcapacity along the stem is strongly impaired (43). Histologicalanalysis of the acropetal progression of IC initiation and ICDextension revealed a decrease in both pin mutants (Fig. 3A andFig. S3), implying that basipetal auxin transport along the stem ispositively correlated with cambium activity and that an enhancedauxin transport rate in max mutants is not causal for decreasedcambium activity.Next, we measured the concentration of free indole-3-acetic
acid (IAA) in the stems of wild-type and max1-1 mutants atdifferent positions including the segment immediately above theuppermost rosette leaf displaying pronounced cambium activity(34). Our analyses revealed enhanced IAA concentration inmax1-1 stems at all positions with an increasing difference to-ward the shoot base, where max1-1 contained ∼5.5 times moreIAA in comparison with wild type (Fig. 3B). To determinewhether enhanced IAA concentration also leads to enhancedauxin signaling, especially in segments with prominent secondarygrowth, we analyzed DR5:GUS reporter activity visualizing auxinsignaling (44) in stems of both wild-type and max1-1 mutants.Enhanced reporter gene activity was detected in max1-1 alongthe whole stem including the base, where secondary growth ismost prominent (34) (Fig. 3 C and D). We also analyzed thedistribution of auxin signaling at tissue level resolution in wild-type and max1-1 backgrounds taking advantage of the DR5rev:GFP reporter (45). The analysis of stem cross-sections revealed acomparable pattern of reporter gene activity with more intenseactivity in max1-1 than in wild type. Increased reporter geneactivity was observed in tissues including primary bundles alongthe whole stem and in interfascicular regions at the stem basewhere IC formation takes place (Fig. 3 E–H).Collectively, these results show that, even though secondary
growth is reduced, auxin concentration and signaling is enhancedin the vasculature of plants with reduced SL biosynthesis. Thissuggests that the decrease in secondary growth observed in max
Fig. 2. GR24 treatments stimulate secondary growth in Arabidopsis in-florescence stems. (A and B) Sections from GR24- (A) and mock-treated (B)wild-type plants. Extension of tissue produced in interfascicular regions isindicated by the brace (}) in A. (C) Quantification of GR24-induced tissueproduction in interfascicular regions of wild-type, max1-1, and max2-1. Notethat in mock-treated plants no cell divisions were observed in any of thegenetic backgrounds (shown here for wild type). (D and E) PXY:CFP activityin mock- (D) and GR24-treated plants (E). Reporter gene-derived signal inthe fascicular cambium is indicated by arrows in D and in interfascicularregions in E. (F and G) APL:CFP activity in mock- (F) and GR24-treated plants(G). Reporter gene-derived signal in the phloem of vascular bundles is in-dicated by arrows in F and in interfascicular regions in G. (H) APL:GUS activityin GR24- (Left) and mock-treated (Right) plants. (G) PXY:GUS detection inGR24- (Left) and mock-treated (Right) plants. Asterisks indicate the positionof vascular bundles. [Scale bar in B (100 μm) also applies to A; scale bar in D(100 μm) also applies to E–G; scale bar in I (5 mm) also applies to H.] Notethat sections in D–G were counterstained by propidium iodide (red), whichhighlights cell walls.
20244 | www.pnas.org/cgi/doi/10.1073/pnas.1111902108 Agusti et al.
Dow
nloa
ded
by g
uest
on
June
18,
202
0

mutants is not a consequence of decreased auxin levels, sup-porting a direct role of SL signaling in the regulation of cambiumactivity independently or downstream of auxin accumulation.
Single Signaling Pathway for Auxin and SL in Secondary GrowthRegulation. Local treatments of stems by NPA are able to blockbasipetal auxin transport leading to auxin accumulation abovethe treatment zone and local initiation of secondary growth(36, 41, 46). We took advantage of this effect to decipher theinteraction of auxin and SL signaling pathways. We speculatedthat if SL signaling is important for a positive effect of auxin oncambium activity, then max mutants should exhibit either noresponse or, at least, a weaker response than wild type. To testthis hypothesis, we performed local NPA treatments of wild-type, max1-1, and max2-1 stems. We observed that interfa-scicular tissue production in NPA-treated max1-1 and max2-1mutants was reduced by 75% (Fig. 4A), indicating that SL
biosynthesis and signaling is important for auxin to stimulatevascular cambium activity. The fact that max mutants are notcompletely devoid of cambium activity might be explained by anSL-independent effect of auxin on the cambium or alternatively,by redundancy in the MAX pathway or a role of as yet uniden-tified factors.To determine whether SL and auxin signaling might act se-
quentially or in parallel, we tested the genetic interaction betweenthe axr1-3 and max mutants. axr1-3 mutants are impaired in auxinsignaling (47) and we reasoned that mutant phenotypes should notbe additive if both act through a common signaling pathway.Consistent with a role of AXR1 in cambium regulation, IC initia-tion and activity was reduced in axr1-3 mutants (Fig. S4). Asexpected, axr1-3 max double mutants displayed the same reductionin the NPA response as the corresponding single mutants (Fig.4A). These findings support a sequential order of the two signalingpathways and, together with the observation of elevated auxinlevels in max1-1 mutants and their reduced NPA responsiveness,suggest that SLs act downstream of auxin in a common signalingcascade. If this is the case, the axr1-3 mutation should not reduceGR24 responsiveness. Indeed, histological analyses showed thataxr1-3, axr1-3 max1-1, and axr1-3 max2-1 plants displayed the sameGR24-induced response as wild type, max1-1, and max2-1, re-spectively (Fig. 4B). Taken together, these results indicate that SLsfunction predominantly downstream of auxin in a signaling cas-cade that positively regulates secondary growth.
Tissue-Specific SL Signaling.MAX2 activity can be found in a broadspectrum of tissues (10) and the other MAX genes can influenceshoot branching over long distances (3). To dissect whether SLsignaling in the vascular cambium is sufficient for stimulatingcambium activity, we expressed MAX2 under the control of the(pro)cambium-specific WOX4 promoter (48) in max2-1 mutants.As controls, we also established lines expressing MAX2 in thestarch sheath, phloem, and xylem, with the help of the SCR (49),APL (40), and NST3 (50) promoters, respectively. RT-PCR withprimers specific for the respective constructs demonstrated thatall transgenes were actively transcribed (Fig. S2D). We analyzedthe lateral ICD extension at the base of the main inflorescencestem from two independent lines for each construct. This anal-ysis demonstrated that only the lines expressing MAX2 in thevascular cambium displayed secondary growth at wild-type-likelevels (Fig. 4C). Lines expressingMAX2 within the phloem or thestarch sheath displayed no difference compared with max2-1,whereas only a minor increase could be observed in linesexpressing MAX2 in the xylem (Fig. 4C). All in all, our resultsshow that SL signaling within the vascular cambium is sufficientto promote cambium activity arguing that MAX2-dependent SLsignaling regulates the process in a cell-autonomous manner.
SL-Dependent Cambium Regulation Is Conserved Between Species.To elucidate whether SLs fulfill a positive role in cambiumregulation in woody species displaying prominent secondarygrowth, we performed local GR24 treatments on stems of Eu-calyptus globulus. As a result, we observed a significant increase inthe production of secondary vascular tissues and in particular, thelateral extension of the cambium zone was increased (Fig. S5). Tosee whether the endogenous SL pathway itself is involved incambium regulation in species other than Arabidopsis, we quan-tified cambium activity in the pea mutant rms1-1, impaired in theactivity of the MAX4 ortholog (7, 32). Our analysis revealed asignificant reduction in cambium activity in comparison with wildtype (Fig. S6). Collectively, these results suggest a conserved rolefor SLs in the regulation of cambium activity across species, in-cluding species with prominent wood production.
Fig. 3. Auxin levels and signaling are enhanced in max1 mutants. (A) Inpin1-613 and pin3-5 mutants, the acropetal progression of IC initiation isdiminished. Plants were analyzed when shoots were 2, 5, 15, and 30 cm tall.(B) Comparison of levels of free IAA in wild type and max1-1 at differentpositions along the inflorescence stem. The first elongated internode abovethe rosette was counted as the first internode. IN, internode. (C and D)Analysis of DR5:GUS activity in wild-type (C) and max1-1 inflorescence stems(D). Rosette leaves have been removed for clarity. (E–H) Analysis of DR5rev:GFP activity at different positions of the inflorescence stem. (E and F)DR5rev:GFP detection 1 cm above the rosette in wild type (E) andmax1-1 (F).(G and H) DR5rev:GFP detection immediately above the uppermost rosetteleaf of wild type (G) and max1-1 (H). [Scale bar in D (5 mm) also applies to C;scale bar in E (100 μm) also applies to F–H.]
Agusti et al. PNAS | December 13, 2011 | vol. 108 | no. 50 | 20245
PLANTBIOLO
GY
Dow
nloa
ded
by g
uest
on
June
18,
202
0

DiscussionIn this work, we present the role of SLs and their interaction withthe auxin signaling pathway in the control of secondary growth.We report a significant reduction in cambium activity in SL-de-ficient mutants (Fig. 1) and show induction of secondary growthupon local treatments with the SL analog GR24 (Figs. 1 and 2).Collectively, these results demonstrate a positive role of SLsignaling in the regulation of cambium activity.Previous studies have shown that PIN protein levels and auxin
transport capacity are enhanced inmax1-1mutant stems and thatGR24 applications reduce PIN1 protein levels and dampen auxintransport (25, 29). It could, hence, be possible that an SL-in-duced change in auxin transport is the reason for an effect of SLsignaling on cambium activity. However, several observationssupport an influence of SLs on cambium activity, which is mainlyindependent from regulating auxin transport. First, cambiumactivity is reduced in pin1 and pin3 mutants (Fig. 3), indicatingthat, in general, basipetal auxin transport along the stem ispositively correlated with cambium activity. Second, even thoughcambium activity is decreased, auxin concentration and signalingis increased in the stem of max1-1 mutants. Third, geneticanalyses of the interaction between SL and auxin signaling in thecontext of cambium regulation are in line with a requirement ofSL signaling for auxin becoming fully active on cambium cellsand show that SL responsiveness is not impaired in auxin sig-naling mutants (Fig. 4). Moreover, the axr1-3 max1-1 and axr1-3max2-1 double mutants did not display additive phenotypes incomparison with single mutants arguing for a linear relationshipof both signaling pathways.Recently, it was shown that auxin induces the expression of the SL
biosynthesis genes MAX3 and MAX4 in the stem of Arabidopsis inan AXR1-dependent manner (51). In agreement with that obser-vation, we propose that the reduced responsiveness of axr1-3 andmax1-1 toward NPA is, at least partly, due to a reduced SL bio-synthesis. In the case of axr1-3, this model would imply that
exogenous SL applications can bypass auxin signaling in the in-duction of secondary growth. In the case of max1-1, GR24treatments would stimulate cambium activity largely indepen-dently from an effect on the auxin signaling pathway. Togetherwith the idea that there is SL production in the shoot (3) and theobservation that the expression of MAX2 in the cambium is suf-ficient to restore wild-type–like levels of cambium activity (Fig. 4),these results argue for an important role of auxin-dependentstimulation of SL biosynthesis in the cambium for secondarygrowth. As the evidence presented in this study is rather indirect,we want to point out, however, that our interpretation does notexclude the possibility that SLs influence cambium activity alsoby regulating auxin transport.Importantly, the reduction of cambium activity in the pea
mutant rms1-1 (impaired in SL biosynthesis) as well as the pos-itive effect of GR24 treatments on the production of secondaryvascular tissue in Eucalyptus, demonstrate that the role of SL incambium activity is conserved among species and that it repre-sents a common signaling mechanism for coordinating growth ofdifferent plant organs.
ConclusionsThe converse effects of SLs on branching and secondary growthsuggest a central role for SL in regulating plant architecture.Shoot branching and secondary growth are both influenced byenvironmental stimuli such as shading, temperature, or daylength (52–55). On the basis of our finding that SLs are involvedin the regulation of both these processes, we hypothesize thatSLs function as modulators used by plants to switch between twogrowth forms: a bushier form displaying a strong outgrowth ofmany side shoots and a weak main stem, and a form in which themain shoot dominates and displays enhanced secondary growth.The ability to oscillate to a certain extent between these growthforms could be regarded as being beneficial if, for example,differences in the necessity to compete for light, or in the total
Fig. 4. Genetic interaction between axr1 and max mutants and the effect of tissue-specific SL signaling. (A) Quantification of NPA-induced tissue productionin interfascicular regions of wild type, max1-1, max2-1, axr1-3, axr1-3 max1-1, and axr1-3 max2-1. (B) Quantification of GR24-induced tissue production ininterfascicular regions of wild type, max1-1, max2-1, axr1-3, axr1-3 max1-1, and axr1-3 max2-1. Note that in mock-treated plants no interfascicular celldivisions were observed in any of the genetic backgrounds (shown here for wild type). (C) Quantification of lateral ICD extension immediately above theuppermost rosette leaf in wild-type, max2-1, and max2-1 plants carrying APL:MAX2, NST3:MAX2, SCR:MAX2, or WOX4:MAX2 transgenes. For each construct,two independent transgenic lines were analyzed. In all cases, n = 10.
20246 | www.pnas.org/cgi/doi/10.1073/pnas.1111902108 Agusti et al.
Dow
nloa
ded
by g
uest
on
June
18,
202
0

number of flowers produced by the shoot system under changingenvironmental conditions, are considered. This feature couldrepresent a strategy in which the plant can choose the optimalway to grow to cope with the challenges imposed by a sessilelifestyle.
Materials and MethodsDetails about growth conditions, cloning and plant lines, histological anal-yses, pharmacological treatments, RT-PCR, auxin measurements, and othertechniques are in SI Materials and Methods. Arabidopsis thaliana (L.) Heynh.
plants of the accession Columbia were used with the exception of the PXY:GUS reporter line, which is in the Ler background (38).
ACKNOWLEDGMENTS. We are grateful to Ottoline Leyser for providing themax1-1, max2-1, max3-9, max4-1, axr1-3 max1-1, and axr1-3 max2-1 mutantlines; Christian Luschnig for providing the pin3-5 mutant; and Simon Turnerfor providing the PXY:GUS marker line. This work was supported by AustrianScience Fund (FWF) Grants P21258-B03 and P23781-B16 (toM.S., P.S., and J.A.)the Wiener Wissenschafts-, Forschungs-, und Technologiefonds (T.S.), theAustralian Research Council (E.A.D., P.B.B., and C.A.B.), and the SwedishGovernmental Agency for Innovation Systems and the Swedish ResearchCouncil (K.L.).
1. Thimann KV, Skoog F (1933) Studies on the growth hormone of plants: III. The in-hibiting action of the growth substance on bud development. Proc Natl Acad Sci USA19:714–716.
2. Snow R (1935) Activation of cambial growth by pure hormones. New Phytol 34:347–360.
3. Turnbull CG, Booker JP, Leyser HM (2002) Micrografting techniques for testing long-distance signalling in Arabidopsis. Plant J 32:255–262.
4. Umehara M, et al. (2008) Inhibition of shoot branching by new terpenoid planthormones. Nature 455:195–200.
5. Gomez-Roldan V, et al. (2008) Strigolactone inhibition of shoot branching. Nature455:189–194.
6. Bouwmeester HJ, Roux C, Lopez-Raez JA, Bécard G (2007) Rhizosphere communica-tion of plants, parasitic plants and AM fungi. Trends Plant Sci 12:224–230.
7. Sorefan K, et al. (2003) MAX4 and RMS1 are orthologous dioxygenase-like genes thatregulate shoot branching in Arabidopsis and pea. Genes Dev 17:1469–1474.
8. Booker J, et al. (2004) MAX3/CCD7 is a carotenoid cleavage dioxygenase required forthe synthesis of a novel plant signaling molecule. Curr Biol 14:1232–1238.
9. Booker J, et al. (2005) MAX1 encodes a cytochrome P450 family member that actsdownstream of MAX3/4 to produce a carotenoid-derived branch-inhibiting hormone.Dev Cell 8:443–449.
10. Stirnberg P, Furner IJ, Ottoline Leyser HM (2007) MAX2 participates in an SCF complexwhich acts locally at the node to suppress shoot branching. Plant J 50:80–94.
11. Beveridge CA, Ross JJ, Murfet IC (1994) Branching mutant rms-2 in Pisum sativum(grafting studies and endogenous indole-3-acetic acid levels). Plant Physiol 104:953–959.
12. Napoli C (1996) Highly branched phenotype of the petunia dad1-1 mutant is reversedby grafting. Plant Physiol 111:27–37.
13. Vogel JT, et al. (2009) SlCCD7 controls strigolactone biosynthesis, shoot branching andmycorrhiza-induced apocarotenoid formation in tomato. Plant J 61:300–311.
14. Ishikawa S, et al. (2005) Suppression of tiller bud activity in tillering dwarf mutants ofrice. Plant Cell Physiol 46:79–86.
15. Lin H, et al. (2009) DWARF27, an iron-containing protein required for the biosynthesisof strigolactones, regulates rice tiller bud outgrowth. Plant Cell 21:1512–1525.
16. Arite T, et al. (2009) d14, a strigolactone-insensitive mutant of rice, shows an accel-erated outgrowth of tillers. Plant Cell Physiol 50:1416–1424.
17. Kohlen W, et al. (2011) Strigolactones are transported through the xylem and playa key role in shoot architectural response to phosphate deficiency in nonarbuscularmycorrhizal host Arabidopsis. Plant Physiol 155:974–987.
18. Woo HR, et al. (2001) ORE9, an F-box protein that regulates leaf senescence in Ara-bidopsis. Plant Cell 13:1779–1790.
19. Woo HR, Kim JH, Nam HG, Lim PO (2004) The delayed leaf senescence mutants ofArabidopsis, ore1, ore3, and ore9 are tolerant to oxidative stress. Plant Cell Physiol 45:923–932.
20. Shen H, Luong P, Huq E (2007) The F-box protein MAX2 functions as a positive reg-ulator of photomorphogenesis in Arabidopsis. Plant Physiol 145:1471–1483.
21. Nelson DC, et al. (2011) F-box protein MAX2 has dual roles in karrikin and strigo-lactone signaling in Arabidopsis thaliana. Proc Natl Acad Sci USA 108:8897–8902.
22. Ruyter-Spira C, et al. (2011) Physiological effects of the synthetic strigolactone analogGR24 on root system architecture in Arabidopsis: Another belowground role forstrigolactones? Plant Physiol 155:721–734.
23. Kapulnik Y, et al. (2011) Strigolactones affect lateral root formation and root-hairelongation in Arabidopsis. Planta 233:209–216.
24. Alonso-Blanco C, et al. (2005) Genetic and molecular analyses of natural variationindicate CBF2 as a candidate gene for underlying a freezing tolerance quantitativetrait locus in Arabidopsis. Plant Physiol 139:1304–1312.
25. Bennett T, et al. (2006) The Arabidopsis MAX pathway controls shoot branching byregulating auxin transport. Curr Biol 16:553–563.
26. Brewer PB, Dun EA, Ferguson BJ, Rameau C, Beveridge CA (2009) Strigolactone actsdownstream of auxin to regulate bud outgrowth in pea and Arabidopsis. PlantPhysiol 150:482–493.
27. Lazar G, Goodman HM (2006) MAX1, a regulator of the flavonoid pathway, controlsvegetative axillary bud outgrowth in Arabidopsis. Proc Natl Acad Sci USA 103:472–476.
28. Prusinkiewicz P, et al. (2009) Control of bud activation by an auxin transport switch.Proc Natl Acad Sci USA 106:17431–17436.
29. Crawford S, et al. (2010) Strigolactones enhance competition between shoot branchesby dampening auxin transport. Development 137:2905–2913.
30. Balla J, Kalousek P, Reinöhl V, Friml J, Procházka S (2011) Competitive canalization ofPIN-dependent auxin flow from axillary buds controls pea bud outgrowth. Plant J 65:571–577.
31. Foo E, et al. (2005) The branching gene RAMOSUS1 mediates interactions among twonovel signals and auxin in pea. Plant Cell 17:464–474.
32. Johnson X, et al. (2006) Branching genes are conserved across species. Genes con-trolling a novel signal in pea are coregulated by other long-distance signals. PlantPhysiol 142:1014–1026.
33. Bainbridge K, Sorefan K, Ward S, Leyser O (2005) Hormonally controlled expression ofthe Arabidopsis MAX4 shoot branching regulatory gene. Plant J 44:569–580.
34. Sehr EM, et al. (2010) Analysis of secondary growth in the Arabidopsis shoot revealsa positive role of jasmonate signalling in cambium formation. Plant J 63:811–822.
35. Agusti J, Lichtenberger R, Schwarz M, Nehlin L, Greb T (2011) Characterization oftranscriptome remodeling during cambium formation identifies MOL1 and RUL1 asopposing regulators of secondary growth. PLoS Genet 7:e1001312.
36. Suer S, Agusti J, Sanchez P, Schwarz M, Greb T (2011) WOX4 imparts auxin re-sponsiveness to cambium cells in Arabidopsis. Plant Cell 23:3247–3259.
37. Menges M, Hennig L, GruissemW, Murray JA (2003) Genome-wide gene expression inan Arabidopsis cell suspension. Plant Mol Biol 53:423–442.
38. Fisher K, Turner S (2007) PXY, a receptor-like kinase essential for maintaining polarityduring plant vascular-tissue development. Curr Biol 17:1061–1066.
39. Hirakawa Y, et al. (2008) Non-cell-autonomous control of vascular stem cell fate bya CLE peptide/receptor system. Proc Natl Acad Sci USA 105:15208–15213.
40. Bonke M, Thitamadee S, Mähönen AP, Hauser MT, Helariutta Y (2003) APL regulatesvascular tissue identity in Arabidopsis. Nature 426:181–186.
41. Little CHA, MacDonald JE, Olsson O (2002) Involvement of indole-3-acetic acid infascicular and interfascicular cambial growth and interfascicular extraxylary fiberdifferentiation in Arabidopsis thaliana inflorescence stems. Int J Plant Sci 163:519–529.
42. Nilsson J, et al. (2008) Dissecting the molecular basis of the regulation of wood for-mation by auxin in hybrid aspen. Plant Cell 20:843–855.
43. Okada K, Ueda J, Komaki MK, Bell CJ, Shimura Y (1991) Requirement of the auxinpolar transport system in early stages of Arabidopsis floral bud formation. Plant Cell3:677–684.
44. Ulmasov T, Murfett J, Hagen G, Guilfoyle TJ (1997) Aux/IAA proteins repress expres-sion of reporter genes containing natural and highly active synthetic auxin responseelements. Plant Cell 9:1963–1971.
45. Friml J, et al. (2003) Efflux-dependent auxin gradients establish the apical-basal axisof Arabidopsis. Nature 426:147–153.
46. Sundberg B, Tuominen H, Little C (1994) Effects of the Indole-3-Acetic Acid (IAA)transport inhibitors N-1-Naphthylphthalamic Acid and Morphactin on endogenousIAA dynamics in relation to compression wood formation in 1-year-old Pinus sylvestris(L.) Shoots. Plant Physiol 106:469–476.
47. Leyser HM, et al. (1993) Arabidopsis auxin-resistance gene AXR1 encodes a proteinrelated to ubiquitin-activating enzyme E1. Nature 364:161–164.
48. Hirakawa Y, Kondo Y, Fukuda H (2010) TDIF peptide signaling regulates vascular stemcell proliferation via the WOX4 homeobox gene in Arabidopsis. Plant Cell 22:2618–2629.
49. Wysocka-Diller JW, Helariutta Y, Fukaki H, Malamy JE, Benfey PN (2000) Molecularanalysis of SCARECROW function reveals a radial patterning mechanism common toroot and shoot. Development 127:595–603.
50. Mitsuda N, et al. (2007) NAC transcription factors, NST1 and NST3, are key regulatorsof the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell 19:270–280.
51. Hayward A, Stirnberg P, Beveridge C, Leyser O (2009) Interactions between auxin andstrigolactone in shoot branching control. Plant Physiol 151:400–412.
52. Aguilar-Martínez JA, Poza-Carrión C, Cubas P (2007) Arabidopsis BRANCHED1 acts asan integrator of branching signals within axillary buds. Plant Cell 19:458–472.
53. Doust AN (2007) Grass architecture: Genetic and environmental control of branching.Curr Opin Plant Biol 10:21–25.
54. Oribe Y, Kubo T (1997) Effect of heat on cambial reactivation during winter dormancyin evergreen and deciduous conifers. Tree Physiol 17:81–87.
55. Schrader J, et al. (2003) Polar auxin transport in the wood-forming tissues of hybridaspen is under simultaneous control of developmental and environmental signals.Proc Natl Acad Sci USA 100:10096–10101.
Agusti et al. PNAS | December 13, 2011 | vol. 108 | no. 50 | 20247
PLANTBIOLO
GY
Dow
nloa
ded
by g
uest
on
June
18,
202
0

Corrections
PLANT BIOLOGYCorrection for “Strigolactone signaling is required for auxin-de-pendent stimulation of secondary growth in plants,” by Javier Agusti,Silvia Herold, Martina Schwarz, Pablo Sanchez, Karin Ljung,Elizabeth A. Dun, Philip B. Brewer, Christine A. Beveridge, TobiasSieberer, Eva M. Sehr, and Thomas Greb, which appeared in issue
50,December 13, 2011, ofProcNatlAcadSciUSA (108:20242–20247;first published November 28, 2011; 10.1073/pnas.1111902108).The authors note that Fig. 3 appeared incorrectly. The cor-
rected figure and its legend appear below. This error does notaffect the conclusions of the article.
www.pnas.org/cgi/doi/10.1073/pnas.1211779109
Fig. 3. Auxin levels and signaling are enhanced inmax1 mutants. (A) In pin1-613 and pin3-5 mutants, the acropetal progression of IC initiation is diminished.Plants were analyzed when shoots were 2, 5, 15, and 30 cm tall. (B) Comparison of levels of free IAA in wild type and max1-1 at different positions along theinflorescence stem. The first elongated internode above the rosette was counted as the first internode. IN, internode. (C and D) Analysis of DR5:GUS activity inwild-type (C) and max1-1 inflorescence stems (D). Rosette leaves have been removed for clarity. (E–H) Analysis of DR5rev:GFP activity at different positions ofthe inflorescence stem. (E and F) DR5rev:GFP detection 1 cm above the rosette in wild type (E) and max1-1 (F). (G and H) DR5rev:GFP detection immediatelyabove the uppermost rosette leaf of wild type (G) and max1-1 (H). [Scale bar in D (5 mm) also applies to C; scale bar in E (100 μm) also applies to F–H.]
www.pnas.org PNAS | August 28, 2012 | vol. 109 | no. 35 | 14277–14279
CORR
ECTIONS

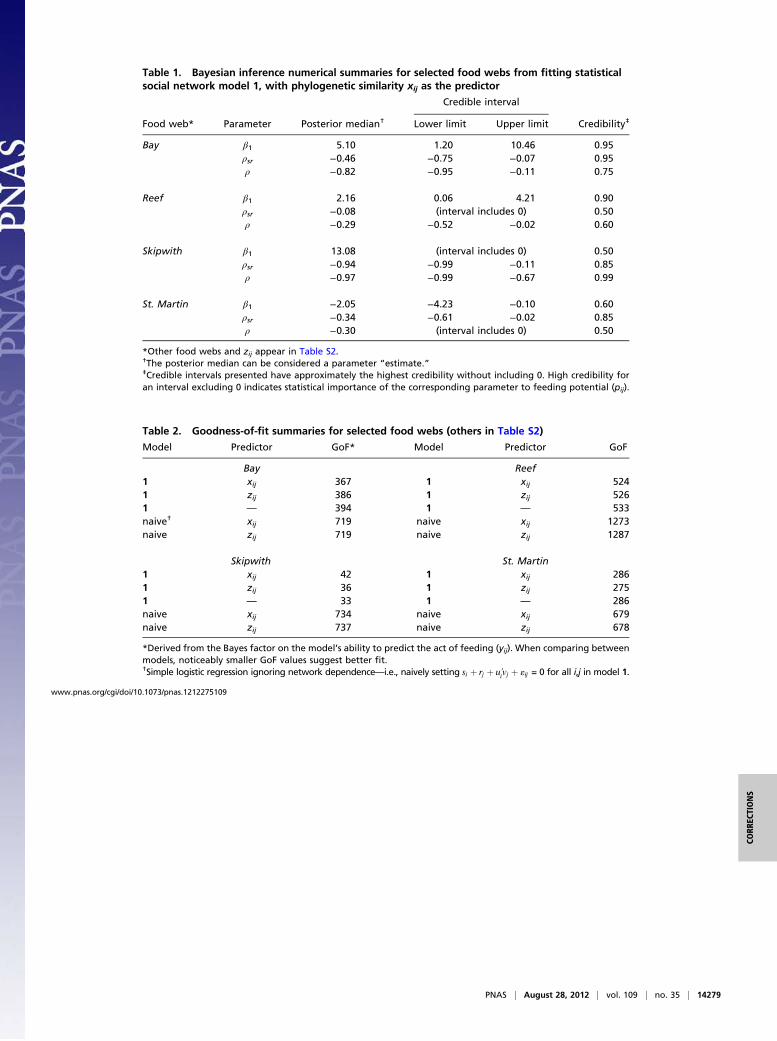
ECOLOGY, STATISTICSCorrection for “A unifying approach for food webs, phylog-eny, social networks, and statistics,” by Grace S. Chiu andAnton H. Westveld, which appeared in issue 38, September 20,2011, of Proc Natl Acad Sci USA (108:15881–15886; first pub-lished September 6, 2011; 10.1073/pnas.1015359108).
The authors note that the data for three of the eight datasetsanalyzed in the article (Skipwith, St. Martin, and Ythan) wereincorrectly processed. As a result, Fig. 2, Table 1, and Table 2appeared incorrectly. The corrected figure, its corrected corre-sponding legend, and the corrected tables appear below. Theseerrors do not affect the conclusions of the article.
Fig. 2. Food web graphs displaying trophic structure from fitting model 1 with phylogeny measure xij as the predictor. (SR) Feeding activity in Bay. The s axisrefers to activity as prey, and the r axis, as predator. Label of node i is located at the mean of the bivariate posterior distribution (an “estimate”) of [si, r i]; thisdistribution describes the probability of the position of [si, r i] on the sr plane, given the knowledge of the observed food web. Distribution density appears asheat map for benthos-eating birds (“37”) and detritus (“48”). Legend for node labels appears in SI Text. (U, V) Preference of being consumed/consuming inSkipwith. They are similarly interpreted as (SR), but referring to ui vectors for small oligochaetes (“2”), C. praeusta (“14”), L. marmoratus (“30”), and detritus(“37”), and vi vectors for A. juncea (“11”), A. germari (“19”), great diving beetle (“27”), and P. sagittalis (“31”). Nodes far apart in the latent u or v space differsubstantially with respect to feeding preference.
14278 | www.pnas.org
www.pnas.org/cgi/doi/10.1073/pnas.1212275109
Table 1. Bayesian inference numerical summaries for selected food webs from fitting statisticalsocial network model 1, with phylogenetic similarity xij as the predictor
Credible interval
Food web* Parameter Posterior median† Lower limit Upper limit Credibility‡
Bay β1 5.10 1.20 10.46 0.95ρsr −0.46 −0.75 −0.07 0.95ρ −0.82 −0.95 −0.11 0.75
Reef β1 2.16 0.06 4.21 0.90ρsr −0.08 (interval includes 0) 0.50ρ −0.29 −0.52 −0.02 0.60
Skipwith β1 13.08 (interval includes 0) 0.50ρsr −0.94 −0.99 −0.11 0.85ρ −0.97 −0.99 −0.67 0.99
St. Martin β1 −2.05 −4.23 −0.10 0.60ρsr −0.34 −0.61 −0.02 0.85ρ −0.30 (interval includes 0) 0.50
*Other food webs and zij appear in Table S2.†The posterior median can be considered a parameter “estimate.”‡Credible intervals presented have approximately the highest credibility without including 0. High credibility foran interval excluding 0 indicates statistical importance of the corresponding parameter to feeding potential (pij).
Table 2. Goodness-of-fit summaries for selected food webs (others in Table S2)
Model Predictor GoF* Model Predictor GoF
Bay Reef1 xij 367 1 xij 5241 zij 386 1 zij 5261 — 394 1 — 533naive† xij 719 naive xij 1273naive zij 719 naive zij 1287
Skipwith St. Martin1 xij 42 1 xij 2861 zij 36 1 zij 2751 — 33 1 — 286naive xij 734 naive xij 679naive zij 737 naive zij 678
*Derived from the Bayes factor on the model’s ability to predict the act of feeding (yij). When comparing betweenmodels, noticeably smaller GoF values suggest better fit.†Simple logistic regression ignoring network dependence—i.e., naively setting si þ rj þ u′ivj þ «ij = 0 for all i,j in model 1.
PNAS | August 28, 2012 | vol. 109 | no. 35 | 14279
CORR
ECTIONS













![Unraveling the Evolution of Auxin Signaling - Plant Physiology · Unraveling the Evolution of Auxin Signaling1[C][W] ... PIN-FORMED orthologs calls for a deeper investigation of their](https://img.pdfslide.us/doc/110x75/5c79a60109d3f200208caa28/unraveling-the-evolution-of-auxin-signaling-plant-unraveling-the-evolution.jpg)







![LBD29-Involved Auxin Signaling Represses NAC Master · LBD29-Involved Auxin Signaling Represses NAC Master Regulators and Fiber Wall Biosynthesis1[OPEN] Kwang-Hee Lee,a Qian Du,a](https://img.pdfslide.us/doc/110x75/5ec8d6b920162707222fa0c8/lbd29-involved-auxin-signaling-represses-nac-lbd29-involved-auxin-signaling-represses.jpg)







