Embed Size (px)
Citation preview

Research article
Received: 11 November 2013, Revised: 28 January 2014, Accepted: 2 February 2014 Published online in Wiley Online Library
(wileyonlinelibrary.com) DOI 10.1002/bio.2656
Stimulation of luminescence of mycelium ofluminous fungus Neonothopanus nambi byionizing radiationTatiana V. Kobzeva,a Anatoly R. Melnikov,a,b Tatiana Y. Karogodina,a
Samat B. Zikirin,a,b Dmitri V. Stass,a,b* Yuri N. Molin,a Emma K. Rodicheva,c,d
Svetlana E. Medvedeva,c,d Alexey P. Puzyr,c,d Andrey A. Burov,d,e
Vladimir S. Bondarc,d and Joseph I. Gitelsonc,d
ABSTRACT: The luminescent system of higher luminous fungi is not fully understood and the enzyme/substrate pair of thelight emission reaction has not been isolated. It was suggested that luminescence of fungi involves oxidase-type enzymes,and reactive oxygen species are important for fungal light production. Generation of reactive oxygen species can be stimulatedby ionizing irradiation, which has not been studied for luminous fungi. We report the effect of X-irradiation on the luminescenceof fungusNeonothopanus nambi. Experiments were performedwithmycelium on a home-built setup based on an X-ray tube andmonochromator/photomultiplier tube. Application of X-rays does not change the emission spectrum, but after approximately20min of continuous irradiation, light production fromunsupportedmycelium starts growing and increases up to approximatelyfive times. After peaking, its level decreases irrespective of the presence of X-irradiation. After staying at a certain level, lightproduction collapses to zero, which is not related to the drying of the mycelium or thermal impact of radiation. Theobserved shape of kinetics is characteristic of a multistage and/or chain reaction. The time profile of light production mustreflect the current levels of radicals present in the system and/or the activity of enzyme complexes involved in light production.Copyright © 2014 John Wiley & Sons, Ltd.
Keywords: Higher luminous fungi; Neonothopanus nambi; ionizing irradiation; reactive oxygen species; lipid peroxidation
* Correspondence to: Dmitri Stass, Institute of Chemical Kinetics and Com-bustion SB RAS, 630090 Novosibirsk, Russia. E-mail: [email protected]
a Institute of Chemical Kinetics and Combustion SB RAS, 630090 Novosibirsk,Russia
b Novosibirsk State University, 630090 Novosibirsk, Russia
c Institute of Biophysics SB RAS, 660036 Krasnoyarsk, Russia
d Siberian Federal University, 660041 Krasnoyarsk, Russia
e Special Design-Technology Bureau “Nauka” SB RAS, 660049 Krasnoyarsk,Russia
IntroductionFor many biological species the luminescent systems and mech-anisms of luminescence are rather well understood, and boththe enzymes that catalyze the reactions leading to light emission(luciferases) and their substrates (luciferins) have been isolatedand characterized (1). For higher luminous fungi, this problemis not yet solved. The molecular organization of the luminescentsystem of higher fungi has not yet been fully determined. It isstill not clear which enzyme (or enzyme complex) functions asluciferase in fungi, or what the structure of the substrate of thelight emission reaction is in this system, i.e., luciferin.
Several decades ago in their in vitro experiments with fungiextracts Airth et al. demonstrated (2-4) that the emission reactioninvolves two enzyme systems that separated upon centrifuga-tion, a “heat labile, nondialyzable” NAD(P)H-dependent reduc-tase found in supernatant and a luciferase found in the pellet,as well as a thermostable luciferin and NAD(P)H. Later this resultwas reproduced by two independent groups (5-8). However,the structure and properties of the fungal luciferase, NAD(P)H-dependent reductase and luciferin are yet unknown, as thesecomponents have not yet been isolated in pure form.
Earlier it has been suggested that themechanismof luminescenceof higher fungi involves reactive oxygen species and oxidase-typeenzymes (9,10). This is supported by results for the luminous fungusNeonothopanus nambi obtained by the Krasnoyarsk team (11–13).The obtained data also imply a connection between the fungal
Luminescence 2014 Copyright © 2014 John
luminescent system and membrane structures (11–14), which isin line with other reports demonstrating that fungal luciferaseis indeed a membrane-bound system (2–8). The available dataindicate that light production in fungi can be mediated by thefollowing enzyme systems associated with membrane structures:oxidases (in particular, peroxidases) of the ligninolytic complex,the system of cytochrome Р-450 and mitochondrial breathingchain enzymes (12,13). All these enzyme systems are capable ofproducing reactive oxygen species, and two of them (ligninolyticcomplex peroxidases and cytochrome Р-450 system) can cata-lyze oxidation of organic substrates, including luciferin, with theparticipation of reactive oxygen species. This was also mentionedin earlier studies, e.g., Kamzolkina et al. (5) suggested that the
Wiley & Sons, Ltd.

T. V. Kobzeva et al.
insoluble particles of the fungal luciferase are similar to mito-chondria, while Oliveira and Stevani (7) argued that the processof fungal light production might involve the system of cyto-chrome Р-450.
It is thus clear that investigation of connections between theluminescence of higher fungi and the processes of generationand transformation of reactive oxygen species in them is impor-tant for understanding the mechanisms of fungal light produc-tion. It is well known (15,16) that generation of reactive oxygenspecies in biological systems can be stimulated by various phys-ical, chemical and biological factors. Chemical stimulation isexperimentally convenient, but, once initiated, it cannot be can-celed, as after adding the reagent to the system the agent notyet consumed cannot be removed. On the other hand, stimulationby irradiationmakes it possible to remove the stimulus at any timesimply by blocking the beam of radiation. The effect of ionizingirradiation on chemiluminescence of plants and bacteria was dem-onstrated previously (17-20). The authors link the observed effectof irradiation with activation of protecting functions of the livingsystems (18), as well as with the generation of radicals, includingreactive oxygen species (17). Furthermore, some authors (21) sug-gest that the luminescent function helps neutralize the effect ofradicals, as it was observed that bioluminescent strains could copewith irradiation more effectively. It has also been noted thatthe more complex and organized the biological system and itsdefense system, the more pronounced is the effect of ionizingirradiation (18). The activation of radical processes in bacteria andplant cells under irradiation and the role of reactive oxygen specieshave previously been discussed (22,23).
A search of literature showed no published accounts of stud-ies on the effect of ionizing radiation on light production in lumi-nous fungi. In this paper, we report and discuss the observedeffect of X-irradiation on the luminescence of fungus N. nambi.
ExperimentalAll experiments were performed with mycelium of luminoushigher fungus N. nambi originally coming from tropic forests ofsouthern Vietnam (24). The fungal culture was generously pro-vided by Vietnamese researcher Dao Thi Van (private strain col-lection BIO-LUMI Co., Ltd., Ho Chi Minh City, Vietnam). The aerialand globular mycelium samples with a prolonged luminescencelevel were produced using the techniques of stationary and sub-merged cultivation of fungus. The biomass of aerial myceliumwas cultivated by the stationary method in Petri dishes on liquidor solid nutrient potato–sucrose media (200 g/L potato brothand 20 g/L sucrose in distilled water or in 2% agar) at 27°С for8-10 days. The submerged cultivation of fungus was carried outin 100-500mL conical vessels containing liquid potato–sucrosenutrient medium (50-250mL). The fungal mycelium grown for8-10 days on Petri dishes in liquid nutrient medium wasfragmented and used as the inoculum for the submergedculture. The volume of the seeding material was 2-5% of the vol-ume of nutrient medium. Cultivation was carried out for 3-4 daysat 27-28°С with continuous stirring at 180-200 rpm (Environmen-tal Shaker-Incubator ES-20; Biosan, Riga, Latvia).
The samples of grown mycelium were washed with deionized(DI) water (Milli-Q system; Millipore, Billerica, MA, USA) overnightbefore the experiments. A significant enhancement of the levelof luminescence from themycelium ofN. nambi after washingwithDI water was demonstrated earlier (12,13). The appearance andintensity of luminescence of mycelium before and after washing
Copyright © 2014 Johnwileyonlinelibrary.com/journal/luminescence
were monitored with a Universal Hood II system (Bio-Rad, Hercules,CA, USA), which makes possible to have the object image in visiblelight and record its luminescence in signal accumulation mode.
The effect of ionizing radiation on the intensity and spectra ofemission from samples of mycelium of N. nambi were studied ona home-built setup described in detail elsewhere (25) compris-ing a CW X-ray tube (BSV-27, Мо, 40 kV × 20mA) and aphotomultiplier tube (PMT; FEU-100, 2200 V) with a system oflenses and a grating monochromator (MDR-206, objective focuslength 180mm, grating 1200 lines/mm, inverse linear dispersion4.3 nm/mm, input and output slits set at 2.2–2.2mm) for regis-tration of the spectra and kinetics of luminescence before, dur-ing and after irradiation. The samples were stimulated with full40 kV X-ray Bremsstrahlung at an estimated dose rate 85 krad/hfor specified periods of time, luminescence was detected in adirection normal to the incident X-ray beam. All presented spec-tra and kinetics were recorded in cylindrical polystyrene cellswith outer diameter 8mm and height 8mm, into which eitherpieces of dense mycelium with dimensions approximately5 × 5mm cut from a film of aerial mycelium, or several globulesfrom the submerged cultivation, were placed. Before the experi-ment, samples were activated by overnight exposition in DIwater with air bubbling. No correction for the PMT spectral sensi-tivity curve was performed when recording the luminescence ki-netics, as it was found that in the conditions of the describedexperiments the apparent maximum in the spectrum of lumines-cence shifts from 530 nm to 525 nm, while the spectral resolutionof the monochromator at used slit widths is about 10 nm. Allpresented kinetics were taken at 525 nm, in the apparent maxi-mum of the emission band. The kinetics consisted of 50,000 timepoints with time step 1 s and were smoothed with a slidingwindow averaging over 20 adjacent points. All Y-axes in thefigures are labeled in “a.u.”, which are PMT voltages in mV andare consistent throughout all figures so that different traces canbe directly compared for signal intensity.
ResultsThe samples of aerial and globular mycelium of N. nambi pro-duce detectable bioluminescence (Fig. 1). The conditions ofsubmerged cultivation (vide supra) produce round globules ofmycelium reaching 2-7mm in diameter after 3-4 days of cultiva-tion. Washing the mycelium samples with DI water leads to asubstantial (10-fold or more) enhancement of their lumines-cence intensity level, which is retained for several days, withoutchanges in the luminescence spectrum (Fig. 2) that agrees withearlier results for this species of fungus (12,13).
First, the response of the produced samples of mycelium toexogenous chemical stimulus was tested using hydrogen perox-ide, which stimulates light production by mycelium of N. nambi(11,13). Fig. 3 shows a typical kinetics of luminescence upon ad-dition of hydrogen peroxide, which agrees with published data.
Figure 4 shows that application of X-rays does not change thefungal emission spectrum: the double-band spectrum was takenunder X-irradiation, the other spectrum was taken from thesame sample immediately after the first spectrum after closingthe X-ray beam shutter. The shorter wavelength emission lineappearing under X-irradiation comes from intrinsic radiation-generated luminescence of the polystyrene cuvette, which wasverified directly by taking a spectrum from the empty cuvette(not shown). It was further verified that the remnants of the cul-ture medium used in the cultivation of dense mycelium produce
Luminescence 2014Wiley & Sons, Ltd.

Figure 1. Appearance (left) and luminescence (right) of samples of mycelium of N. nambi grown in Petri dishes (А) and by submerged cultivation and washed with DIwater (В). Registered using a Universal Hood II system (Bio-Rad, Hercules, CA, USA).
300 400 500 600 700-2
0
2
4
6
8
10
12
14
16
18
20
Sig
nal,
a.u.
Wavelength, nm
Figure 2. Typical spectrum of bioluminescence from a globular sample. The spec-trum for dense mycelium (not shown) is identical to the spectrum for globules.
0 5 10 15 20 25 30 35 40 45 50 55 60 6510
20
30
40
50
60
Sig
nal,
a. u
.
Time, min
Figure 3. Typical kinetics of light production for a globule of mycelium ofN. nambi placed in 40μL of water. At 30min, 10μL of 0.6% hydrogen peroxide wasadded to the sample.
Stimulation of luminescence of N. nambi by ionizing radiation
no luminescence either per se or under X-irradiation. Finally, itwas found that a sample of fungus with intrinsic biolumines-cence is required to obtain X-ray stimulated emission, and thedegree of stimulation is roughly proportional to the initial levelof the intrinsic bioluminescence.
Figure 5 shows typical bioluminescence kinetics for a sampleof dense mycelium taken in the same conditions as the spectrashown above. This type of sample has a higher initial level oflight production, as indicated by higher Y-axis scale values. Theinitial increase of light production followed by relaxation isalways observed after placing a sample into the experimentalsetup and is the known reaction for mechanical stimulus upon
Luminescence 2014 Copyright © 2014 John
sample manipulation (26). The level of light production graduallyrelaxes to a certain background level and stays there forprolonged times (in this case overnight), with fungus visuallyremaining healthy and moist. This behavior is reproducible forall types of samples used in this study, with varying absolutelevels but similar overall shapes, and does not depend onwhether the kinetics are recorded through the day or overnight.Figure 6 shows the range of responses that radiation stimula-
tion elicits from different preparations of mycelium. The figureconsists of three pairs of panels showing a stimulation experi-ment and a matching control experiment for two pieces of thesame mycelium (Fig. 6a + b, c + d and e + f), and two additional
Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/luminescence

200 300 400 500 600 700-2
0
2
4
6
8
10
12
14
16
X-ray off
Sig
nal,
a.u.
Wavelength, nm
X-ray on
Figure 4. Spectrum of luminescence for a globular sample in a polystyrenecuvette taken under X-irradiation (solid line) and from the same sample with X-raysturned off by closing the beam shutter (dashed line).
0 60 120 180 240 300 360 420 480 540 600 660 720 780
0
10
20
30
40
50
60
70
80
Sig
nal,
a. u
.
Time, min
Figure 5. Kinetics of light production from a piece of dense mycelium. The sam-ple was healthy at the end of measurement.
T. V. Kobzeva et al.
stimulation experiments. Figure 6(a–d) were obtained for densemycelium on agar, Fig. 6(e–g) for unsupported dense myceliumgrown on liquid medium and Fig. 6(h) for globular mycelium.Each experimental cycle took approximately 20 h and was pre-ceded by overnight washing of globules or cut pieces of densemycelium in DI water to activate the intrinsic bioluminescence.The sample was loaded into the experimental setup as describedin the Experimental section at approximately 16.00 h local time.An optional 1 h waiting period was allowed to relax after the me-chanical shock and enter the falling part of the light productionkinetics (cf. Fig. 5), and then the 50,000 s kinetics shown in thepanels was launched that ended by early morning the followingday. When required, X-ray stimulation was applied and removedas shown by the corresponding markers in the panels. The sam-ple was left in the setup until midday, when short kinetics wasrecorded to determine the resulting level of light production.After that, the sample was taken out of the system and visuallyinspected. Because the X-ray irradiation/luminescence detection
Copyright © 2014 Johnwileyonlinelibrary.com/journal/luminescence
system has provisions only for one sample at a time, the controlexperiments were performed after the corresponding stimula-tion experiment using a piece of mycelium from the same dish,with the entire experimental cycle, including cutting the sampleand initial washing shifted by 1 day. For stimulation experimentsshown in Fig. 6, irradiation was applied continuously betweenthe indicated time points. While inside the experimental setup,the sample stayed in the dark closed compartment and wasnot moved in any way. Initially, 40μL of water was added toeach sample upon loading, and part of it always remained inthe cuvette when the sample was taken out irrespective of theobserved state of the sample. For samples on agar, it waschecked that neither agar itself, nor a piece of dried myceliumon agar produced any background emission under X-irradiation.Full 2-day exposition/control cycles were replicated at leasttwice for each sample.
Figure 6(a) shows the kinetics of light production for a piece ofdense mycelium on agar. When X-rays are applied as describedin the Experimental section (the first arrow marker in Fig. 6(a),1 h after the start of kinetics), growth in the level of light produc-tion begins after approximately 20min. For sample light peakedat approximately twofold the intensity of bioluminescence rela-tive to the non-stimulated control. After peaking, the level oflight production decreases, and the decrease is independent ofthe presence of X-irradiation. The second arrow marker in Fig. 6(a) shows the time when the irradiation was stopped, at 4 h30min after the start of the kinetics. When checked for lightproduction at next noon the next day, the sample producedabout 80% of the light intensity, consistent with the gradualdecaying slope that the kinetics shows. When the sample wastaken out of the cuvette, it was noted that brownish patcheshave developed at the surface of the mycelium although itremained moist. When left for a longer time, the sample driednaturally into an off-white film without further developing thepatches. Figure 6(b) shows the matching control experiment.The kinetics broadly reproduce the curve shown in Fig. 5; afterthe experimental cycle the sample produced the consistent levelof light, and visually remained in the state at which it was loadedinto the experimental setup. When left for a longer time, thesample dried naturally into a homogeneous off-white film with-out any coloration.
Figure 6(c,d) shows the kinetics of light production for anothersample of dense mycelium on agar from a different batch, whichdemonstrates the lower end of the observed spectrum of re-sponse intensity. The control experiment of Fig. 6(d) producedresults virtually identical to the sample in Fig. 6(b). However,the response of X-irradiation in this case is less prominent,although and with approximately the same timings. The appar-ently longer delay of onset of increase in light production isthe result of a much lower absolute increase superimposed ona rather steep decaying slope. This sample that respondedpoorly to stimulation produced again about 80% of the lightintensity at the end of kinetics next day, visually remained inits initial state without signs of patching and dried naturally intoa homogeneous off-white film.
Figure 6(e) shows the kinetics of light production for a pieceof unsupported dense mycelium grown on liquid nutrientmedium. This type of sample preparation turned out to bemore responsive to X-ray stimulation. Although starting withapproximately the same initial level of light production, theunsupported mycelium produces five times its intrinsic biolumi-nescence upon stimulation (note the different scale of the Y-axis
Luminescence 2014Wiley & Sons, Ltd.

0 60 120 180 240 300 360 420 480 540 600 660 720 7800
40
80
120
160
Sig
nal,
a. u
.S
igna
l, a.
u.
Sig
nal,
a. u
.S
igna
l, a.
u.
Time, min
0 60 120 180 240 300 360 420 480 540 600 660 720 7800
40
80
120
160
Sig
nal,
a. u
.
Time, min
0 60 120 180 240 300 360 420 480 540 600 660 720 780
Time, min
0 60 120 180 240 300 360 420 480 540 600 660 720 780
Time, min
0 60 120 180 240 300 360 420 480 540 600 660 720 780
Time, min
0 60 120 180 240 300 360 420 480 540 600 660 720 780
Time, min
0 60 120 180 240 300 360 420 480 540 600 660 720 780
Time, min
0 60 120 180 240 300 360 420 480 540 600 660 720 780
Time, min
turn on
turn off
a b
0
40
80
120
turn on
turn off
c d
0
20
40
60
80
turn on
turn off
g
turn on
turn off
h
f
0
100
200
300
400
Sig
nal,
a. u
.S
igna
l, a.
u.
Sig
nal,
a. u
.0
40
80
120
0
10
20
30
0
100
200
300
400
turn on turn off
e
Figure 6. Range of responses to radiation stimulation: (a) mycelium on agar, stimulated as shown; (b) matching control without stimulation; (c) mycelium on agar,stimulated; (d) matching control, without stimulation; (e) unsupported mycelium, stimulated; (f) matching control without stimulation; (g) aged unsupported mycelium,stimulated; (h) globular mycelium, stimulated. One hour waiting time used for (a). See text for details.
Stimulation of luminescence of N. nambi by ionizing radiation
and better signal-to-noise ratio) with the same overall shape andtiming of the main response. However, the further course of thekinetics is strikingly different. After a period at a level exceedingthe initial intensity level, the light production abruptly drops andfalls to zero – the horizontal tail in the kinetics of Fig. 6(e) is themeasured intensity of luminescence, which is identical to the
Luminescence 2014 Copyright © 2014 John
background level of the experimental setup. When removedfrom the cuvette, the sample was completely covered withbrown patches mentioned for the sample of Fig. 6(a), driedand shrunk, although some water did remain in the cuvette.Figure 6(f) shows the matching control experiment, which issimilar to two already described controls of Fig. 6(b,d). The
Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/luminescence
T. V. Kobzeva et al.
kinetics again broadly reproduces the curve shown in Fig. 5, af-ter the experimental cycle the sample produced a consistentlevel of light and visually remained in its original state, and whenleft for an extended time dried into a homogeneous off-whitefilm.
Figure 6(g) shows the kinetics of light production for anothersample of unsupported dense mycelium, this time from an olderfungus already starting to senesce. The overall level of light pro-duction in this case is lower, and the initial stress relaxation periodis longer, so stimulation was applied later than in the precedingpanels. This sample again develops a bell-shaped response to radi-ation stimulation with the same lag period, although at a lower in-tensity, and shows the abrupt disappearance of light productionafter the stimulation, producing a kinetic trace similar to Fig. 6(e).This sample was also completely brown and dried after the endof experiment, and its light production could not be recovered.
An obvious candidate for the observed terminal effect of irra-diation on the unsupported mycelium is direct physical heatingby energy deposition via irradiation. However, simple estimatesshow this is not the case. The used dose rates are too low: at awater heat capacity of 75.35 J/mol × K and the quoted dose rateof 85 krad/h, even neglecting heat losses, the estimated rate oftemperature rise does not exceed 0.2ºС/h.
Finally, Fig. 6(h) shows the kinetics of stimulated light produc-tion for a sample of globular mycelium. The overall level of lightproduction in this case is much lower than for dense mycelium,but stimulation does produce an increase in light productionwith the same lag period. The globular samples have a shorterlife span of about 1week and thus are not very convenient forthe extended kinetics experiments of the type described withthe sequential control experiments. The initial increasing re-sponse with a 20-min lag period and following decrease of thetype shown in Fig. 6(h) was seen reproducibly.
2
1
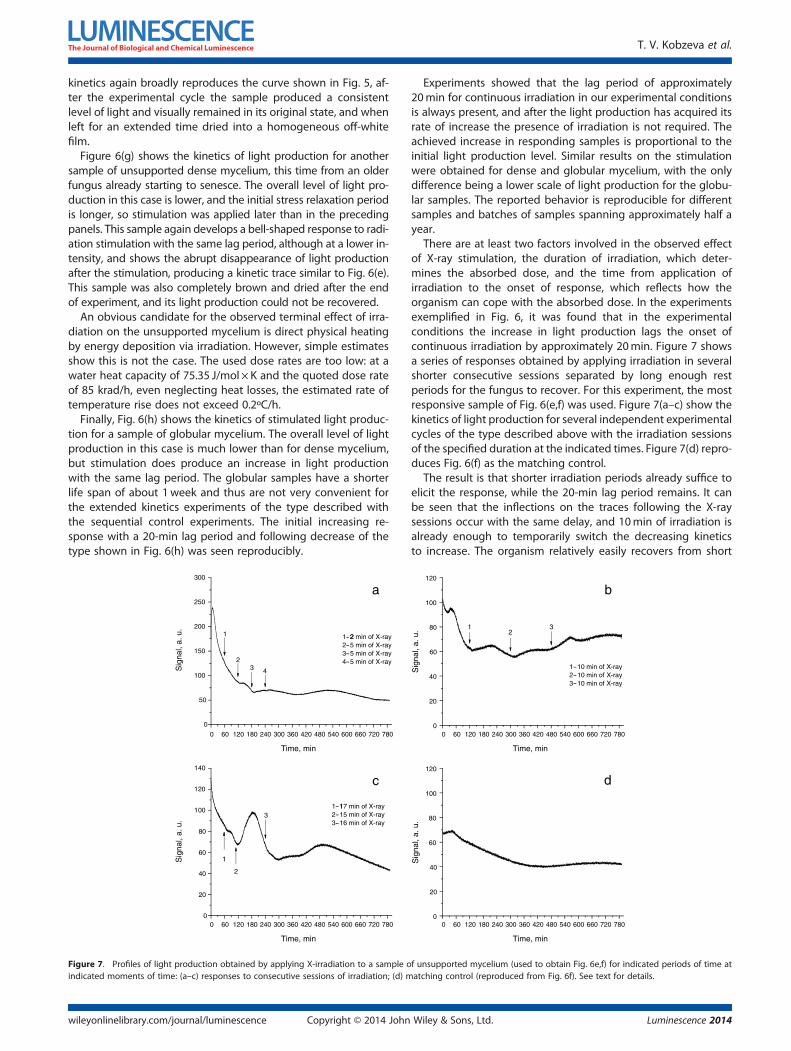
Figure 7. Profiles of light production obtained by applying X-irradiation to a sample oindicated moments of time: (a–c) responses to consecutive sessions of irradiation; (d) m
Copyright © 2014 Johnwileyonlinelibrary.com/journal/luminescence
Experiments showed that the lag period of approximately20min for continuous irradiation in our experimental conditionsis always present, and after the light production has acquired itsrate of increase the presence of irradiation is not required. Theachieved increase in responding samples is proportional to theinitial light production level. Similar results on the stimulationwere obtained for dense and globular mycelium, with the onlydifference being a lower scale of light production for the globu-lar samples. The reported behavior is reproducible for differentsamples and batches of samples spanning approximately half ayear.
There are at least two factors involved in the observed effectof X-ray stimulation, the duration of irradiation, which deter-mines the absorbed dose, and the time from application ofirradiation to the onset of response, which reflects how theorganism can cope with the absorbed dose. In the experimentsexemplified in Fig. 6, it was found that in the experimentalconditions the increase in light production lags the onset ofcontinuous irradiation by approximately 20min. Figure 7 showsa series of responses obtained by applying irradiation in severalshorter consecutive sessions separated by long enough restperiods for the fungus to recover. For this experiment, the mostresponsive sample of Fig. 6(e,f) was used. Figure 7(a–c) show thekinetics of light production for several independent experimentalcycles of the type described above with the irradiation sessionsof the specified duration at the indicated times. Figure 7(d) repro-duces Fig. 6(f) as the matching control.
The result is that shorter irradiation periods already suffice toelicit the response, while the 20-min lag period remains. It canbe seen that the inflections on the traces following the X-raysessions occur with the same delay, and 10min of irradiation isalready enough to temporarily switch the decreasing kineticsto increase. The organism relatively easily recovers from short
f unsupported mycelium (used to obtain Fig. 6e,f) for indicated periods of time atatching control (reproduced from Fig. 6f). See text for details.
Luminescence 2014Wiley & Sons, Ltd.

Stimulation of luminescence of N. nambi by ionizing radiation
irradiation periods, but as the duration of each period increases,the changes in light production profile become irreversible.Furthermore, the effect of such an intermittent irradiation iscumulative, as the kinetics does not return to its initial courseafter the irradiation.
DiscussionIt follows that X-irradiation stimulates the light production sys-tem of the fungus with a certain lag period, 20min in the condi-tions of the described experiment, the minimum exposure timerequired to elicit the response is shorter than the lag period, theeffect of irradiation is cumulative, and after the light productionhas acquired its rate of increase the presence of irradiation is notrequired to sustain it. There is a range of responses to stimula-tion, from moderate to quite high (up to fivefold increase wasobserved), and the samples that respond to irradiation developa brown coloration and shrink, the more so the higher theresponse was. Samples of mycelium on agar seem to be lessresponsive to stimulation, while samples of unsupported densemycelium may produce a very pronounced response leadingto complete coloration of mycelium and loss of light production.The ultimate drop of signal to zero for the unsupported samplesis not related to the simple drying of mycelium (the control sam-ples of the same mycelium persist quite well) and to the plainthermal impact of radiation (the temperature rise is negligible).The observed increase in light production clearly accompaniesan acceleration of fungal catabolism that, for unsupportedmycelium, leads to the terminal self-destruction of the organism.
It is reasonable to suggest that, similar to chemical stimulationupon addition of hydrogen peroxide, radiation stimulation ofluminescence is also mediated by radiation generation ofradicals in aqueous medium that surrounds and fills mycelium.The yields of radicals in water are known and are equal to about5/100 eV, and thus the amount of radicals produced in the sam-ple of mycelium can be estimated from the used dose rate at thesample and the typical duration of irradiation (85 krad/h and 2 h,respectively) (27) as about 1.5 × 1016. The amount of hydroxylradicals formed upon complete decomposition of 10μL of0.6% hydrogen peroxide is about 1.2 × 1015. Finally, the totalamount of oxygen dissolved in the fragment of mycelium usedfor the experiment is estimated at 1.5 × 1016. The close numbersof about the same order of magnitude support the involvementof reactive oxygen species in the processes induced by ionizingirradiation of fungus. The observed shape of kinetics with a lagperiod and the acceleration phase is characteristic of a multi-stage and/or chain reaction. The initial increase and the follow-ing decreаse in light production must reflect the current levelsof radicals present in the system and/or the activity of enzymecomplexes involved in light production.
A possible candidate for the underlying chain process is thecycle of lipid peroxidation (28). Initiation of lipid peroxidationby ionizing irradiation with varied characteristics was indeedreported in literature. Khalil and Fülöp (29) used a cobalt gunwith a dose rate 0.12 Gy/s (which corresponds to 43 krad/h at1Gy = 100 rad), exposed to doses of 0–400Gy (0–40 krad, whichcorresponds to about 30min of irradiation in the conditions ofthe described experiment) and observed an increase in the levelof products of peroxidation for low-density lipoproteins with alag period of 10-20min. Although the energies of the quantaof radiation in the cited work (60Co, 1.17MeV and 1.33MeV)and in this work (full-spectrum X-ray Bremsstrahlung with
Luminescence 2014 Copyright © 2014 John
maximum energy 40 keV) are different, this is of no consequencefor radiolysis of water having molecules composed of lightatoms, and thus the conditions of experiments on the modelsystem in Khalil and Fülöp (29) and the experiments describedhere can be considered similar.It is clear that the reported observation of stimulated light
production per se is not sufficient to establish the mechanismof the enhancement, and further study is required. It is alsopossible that ionizing irradiation initiates several concurrentprocesses that have no immediate relationship to each other. Stillthe reported effect provides an indirect support to the involve-ment of reactive oxygen species in the luminescence of higherfungi, and introduces a useful tool to study fungal luminescence.
Acknowledgments
The work was supported by the Program of Siberian Branch ofRussian Academy of Sciences (project no. 71), Council for Grantsof the President of the Russian Federation for Support of Lead-ing Scientific Schools (project no. NSh 2272.2012.3), the RussianFoundation for Basic Research (project no. 12-03-33082), and theProgram of Government of Russian Federation “On the Effortsfor Attracting Leading Researchers to Educational Institutionsof Russia” (grant no. 11.G34.31.0058).
References1. Shimomura O. Bioluminescence: chemical principles and methods.
Singapore: World Scientific Publishing Co. Pte. Ltd., 2006.2. Airth RL, McElroy WD. Light emission from extracts of luminous
fungi. J Bacteriol 1959;77:249–50.3. Airth RL, Foerster GE. The isolation of catalytic components required
for cell-free fungal bioluminescence. Arch Biochem Biophys1962;97:567–73.
4. Airth RL, Foerster GE. Enzymes associated with bioluminescence ofPanus stipticus luminescens and Panus stipticus nonluminescens.J Bacteriol 1964;88:1372–9.
5. Kamzolkina OV, Danilov VS, Egorov NS. Nature of luciferase fromthe bioluminescent fungus Armillariella mellea. Dokl Biochem1983;271:274–6.
6. Kamzolkina OV, Bekker ZE, Egorov NS. Extraction of the luciferin-luciferase system from the fungus Armillariella mellea. Biol Nauki1984;1:73–7.
7. Oliveira AG, Stevani CV. The enzymatic nature of fungal biolumines-cence. Photochem Photobiol Sci 2009;8:1416–21.
8. Oliveira AG, Desjardin DE, Perry BA, Stevani CV. Evidence that a sin-gle bioluminescent system is shared by all known bioluminescentfungal lineages. Photochem Photobiol Sci 2012;11:848–52.
9. Shimomura O. Superoxide-triggered chemiluminescence of theextract of luminous mushroom Panellus stipticus after treatment withmethylamine. J Exp Botany 1991;42:555–60.
10. Shimomura O. The role of superoxide dismutase in regulating thelight emission of luminescent fungi. Exp Botany 1992;43:1519–25.
11. Bondar VS, Puzyr AP, Purtov KV, Medvedeva SE, Rodicheva EK,Gitelson JI. The luminescent system of the luminous fungusNeonothopanus nambi. Dokl Biochem Biophys 2011;438:138–40.
12. Bondar VS, Shimomura O, Gitelson JI. Luminescence of higher mush-rooms. J Siberian Fed Univ Biol 2012;5:331–51.
13. Bondar VS, Rodicheva EK, Medvedeva SE, Tyulkova NA, Tyaglik AB,Shpak BA, Gitelson JI. On the mechanism of luminescence of thefungus Neonothopanus nambi. Dokl Biochem Biophys 2013;449:80–3.
14. Puzyr AP, Burov AE, Bondar VS. Source of light emission in a luminousmycelium of the fungus Panellus stipticus. J Res Biol 2013;3:900–5.
15. Vladimirov YA. Intrinsic chemiluminescence of living tissues. In: Nohl H,Esterbauer H, Rice-Evans C, editors. Free Radicals in the Environment,Medicine and Toxicology. London: Richelieu Press, 1994:345–73.
16. Halliwell B, Gutteridge JMC. Free Radicals in Biology and Medicine.Oxford: University Press, 1999.
17. Radotic K, Radenovic C, Jeremic M. Spontaneous ultraweak biolumi-nescence in plants: origin, mechanisms and properties. Gen PhysiolBiophys 1998;17:289–308.
Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/luminescence

T. V. Kobzeva et al.
18. Rozhko TV, Kudryasheva NS, Kuznetsov AM, Vydryakova GA, BondarevaLG, Bolsunovsky AY. Effect of low-level α-radiation on bioluminescent as-say systemsof various complexity. PhotochemPhotobiol Sci 2007;6:67–70.
19. Mantel J, Freidin M, Bulich AA, Perry H. The effect of radiation onbioluminescent bacteria: possible use of luminescent bacteria as abiological dosemeter. Phys Med Biol 1983;28:599–602.
20. Asada K. The water-water cycle in chloroplasts: scavenging of activeoxygens and dissipation of excess photons. Annu Rev Plant PhysiolPlant Mol Biol 1999;50:601–39.
21. Cryz A, Wrobel B, Wegrzyn G. Vibrio harveyi bioluminescence plays arole in stimulation of DNA repair. Microbiology 2000;146:283–8.
22. Mackemess SA-H, John CF, Jordan В, Thomas B. Early signaling com-ponents in ultraviolet-B responses: distinct role for different reactiveoxygen species and nitric oxide. FEBS Lett 2001;489:237–42.
23. Mika A, Minibayeva F, Beckett R, Lüthje S. Possible functions of extra-cellular peroxidases in stress-induced generation and detoxificationof active oxygen species. Phytochem Rev 2004;3:173–93.
Copyright © 2014 Johnwileyonlinelibrary.com/journal/luminescence
24. Vydryakova GA, Van DT, Shoukouhi P, Psurtseva NV, Bissett J.Intergenomic and intragenomic ITS sequence heterogeneity inNeonothopanus nambi (Agaricales) from Vietnam. Mycology 2011;3(2):89–99.
25. Kalneus EV, Melnikov AR, Korolev VV, Ivannikov VI, Stass DV. A low-field magnetically affected reaction yield (MARY) spectrometer withspectral fluorescence resolution. Appl Magn Reson 2013;44:81–96.
26. Mihail JD, Bruhn JN. Dynamics of bioluminescence by Armillariagallica, A. mellea and A. tabescens. Mycologia 2007;99:341-50.
27. Spinks JWT, Woods RJ. An Introduction to Radiation Chemistry.New York: John Wiley & Sons, 1976.
28. Yin H, Xu L, Porter NA. Free radical lipid peroxidation: mechanismsand analysis. Chem Rev 2011;111:5944–72.
29. Khalil A, Fülöp T. A comparison of the kinetics of low-density lipopro-tein oxidation induced by copper or by γ-rays: Influence of radiationdose-rate and copper concentration. Can J Physiol Pharmacol2001;79:114–21.
Luminescence 2014Wiley & Sons, Ltd.





![DLF&x, A...DLF Land India Limited [w.e.f. 26 April 2019] ee Nambi Buildwell Limited (formerly Nambi Buildwell Private Limited ) [w.e.f. 30 September 20 19 . d) Key ma Dr. K.P. po Name](https://img.pdfslide.us/doc/110x75/5e7b3c953ebfbf5cec7b1214/dlfx-a-dlf-land-india-limited-wef-26-april-2019-ee-nambi-buildwell.jpg)























