Embed Size (px)
Citation preview

177
Advanced Concepts of Strength & Conditioning
SportMetabolism
NCSF
CertifiedStrengthCoach
Chapter
6

Sport MetabolismWinning or losing in competitive sports has as much to do
with metabolic factors as it does with the physical attributes and
technical skills of the athletes. The loss of energy from the body
leads to deleterious effects on performance, and is often a
difference maker in the last stages of a race or second half of
a game. For this reason, strength and conditioning programs
should be constructed to exploit the metabolic systems for com-
petition, while considering proper nutritional support to ensure
adequate provisions are available for work and recovery.
The term bioenergetics essentially describes the body’s use of
energy following the process of nutrient consumption and absorp-
tion. The metabolic activity supporting both rest and active
conditions occur through chemical reactions that free energy from
nutrient sources into bodily systems. Unlike mechanical objects,
which have an on and off switch, living organisms need a constant
flow of energy to support the requirements of tissue. To further
complicate this matter, humans also require a variety of nutrients
for different purposes. Therefore, to manage the constant need
for energy and the variability of a system’s affinity for a particular
type of fuel, the body functions using an ongoing process of
energy transfer. The laws of thermodynamics encompass this
notion, as the first law states “energy cannot be created or
destroyed”. Assuming the universe is a closed-system, all energy
in it is reusable, either existing in a state of potential or being
transferred kinetically. In a closed-system, energy is constant
because it is locked in and infinitely recycled; while the total quan-
tity of energy in an open system, such as the human body, is
variable because energy can be added or removed. Due to the fact that all living organisms func-
tion as open systems, energy is constantly needed to replenish that which is being lost.
The second law of thermodynamics relates to the concept that during any energy exchange
there is a loss in order, or entropy, as it is scientifically referred. Whenever energy is transferred
in the body from one form to another, some of the energy is lost due to inefficiency (loss of
order). This makes it impossible for the body to harness all the energy in a given reaction. The
aerobic use of a six-carbon glucose molecule for instance, is roughly 35%; similar to the efficiency
of a light bulb. From a practical perspective, an athlete running a race is losing energy through
all biological mechanisms. Energy is lost as it is transferred to manage ongoing mechanical work,
heat dissipation, and metabolic homeostasis. Due to the fact that energy is constantly being lost
from the body, an athlete in a long race will need to add new energy to sustain their workload, or
they will be forced to slow down.
Energy-yielding nutrients are formed from the elements carbon, hydrogen, and oxygen, with
the addition of nitrogen to form protein. These chemical structures can be separated or recon-
figured to take on other forms of energy or become byproducts and substrates as elements are
removed during metabolism. Interestingly, the byproducts themselves reflect configurations of
the foundational elements so they too can be restructured with other elements to form new
178
DEFINITIONS
Bioenergetics –
Describes the processes of energy usewithin the body
Entropy –
Energy disorder associated withchemical reactions within the body, thesystem loses order and efficiency isconsequently reduced
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism

179
energy. Forms of energy in the body may be mechanical, biochemical, or thermal depending on
the role it is fulfilling. For example, a fully-fueled athlete who participates in a competition will
lose a notable proportion of his or her stored energy by the end of the game, which is made evi-
dent by fatigue. But the body does not simply become drained from the mechanical activity used
to perform the sport’s movements; rather it loses energy through the metabolic mechanisms
necessary to keep it alive.
The sweat dripping off the athlete represents energy lost through heat-related mechanisms;
the increased respiration from high-intensity actions release gases, while the constant movement
during competition derives mechanical energy from inside the tissues; mobilizing it into a free
form. By the end of a two hour event an athlete may have lost over a thousand calories of energy
from the body. Energy that participates in any body function is always reduced by the amount
of free energy it creates. Therefore, energy in storage represents a whole value, and metabolic
processes reduce that value by the quantity freed into the system. Once free, the energy may
follow any number of paths. Carbons freed through glucose metabolism may connect with
oxygen (O2) and leave the body as carbon dioxide (CO2), or may connect with the nitrogen and
hydrogen (H+) liberated from amino acids to form urea; which will leave the body as urine.
Energy released into the environment can no longer be re-used to form energy within the body,
and is therefore lost, but as the laws of thermodynamics state, it is not destroyed. The CO2
breathed into the air will likely be recaptured by plants to form more usable energy such as
carbohydrates (CHO). Humans must constantly replace or regenerate energy due to continuous
loss. Performance outcomes are ultimately determined by the body’s ability to manage available
energy; without a constant energy supply the systems cannot function properly and performance
declines.
Free Energy and the Laws of Thermodynamics
The idea of free energy referenced in the prior text can be better understood when appliedthrough the laws of thermodynamics:
It is important to recognize that this equation attempts to represent what happens to free energy
when energy is transferred. Change in free energy (∆G) is related to three other factors: 1) total
internal energy within the system, or enthalpy (H), 2) absolute temperature (T) and 3) entropy
(S) or the loss in order. In essence, this formula aims to calculate available energy by taking the
initial amount of energy within the system and subtracting the energy that is lost due to disrup-
tion and inefficiency.
The change in free energy (∆G) during any reaction is derived from the difference between
the free energy released from the reaction (byproduct) and the amount of free energy used to
cause the reaction. From a training perspective, biochemical reactions either release energy for
direct use during exercise (ATP ↔ ADP + Pi + energy) or require energy in the process of
providing energy for exercise (PCr + ADP + energy↔ ATP + C). For a reaction to provide
energy, the free energy contained in the final molecules must be lower than the free energy in the
initial molecules – the difference represents the cost of the reaction (second law of thermodynam-
ics). This is where the concept of energy breakdown occurs, and in its process, how free energy
is released. It also explains why different energy sources may provide more or less energy at a
given time and provide different limits in total energy availability during a physical event.
DEFINITIONS
Enthalpy –
The sum of the internal energy of thesystem plus the product of the pressureof the gas in the system and its volume
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
The human body is not a closed system andtherefore lacks efficiency, leading to a constantdepletion of energy.

Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
The concept of free energy is important toward understanding the reactions that occur
within sports metabolism because different reactions cause different outcomes. It is also impor-
tant to understand that the higher the metabolic rate associated with any given sport action, the
faster energy is depleted during its execution. In some cases, the metabolic requirements are so
demanding that only certain sources of energy can support the activity. This causes a rapid and
notable drain on local stores leading to fatigue. Maximal-speed sprints for instance, are too fast
to be supported by the oxidation of fat or proteins; therefore, only CHO can contribute to the
supportive metabolism. To put this into a sport example, pre-game glycogen storage can be
reduced by as much as 40-70% by the end of a soccer match due to the quantity of high-speed
activity that occurs over the 90-minute competition. Without adequate training and nutrition
to support the energy demands an athlete will slow to non-competitive speeds, potentially
contributing to a losing outcome.
Food and drink provides a variety of energy-yielding nutrients for the body, but when it
comes to using the energy for mechanical purposes there is only one exchange medium; the
breakdown of adenosine triphosphate (ATP). ATP is unique in that it is the only energy source
utilized by muscles within the contractile units (sarcomere). Therefore, ATP availability is
requisite to muscle activity. Energy-yielding nutrients hold the elements necessary to form ATP,
but they must be refined through reactions to provide the ATP to working tissue. This is con-
ceptually similar to providing fuel for a car. Crude oil must go through significant processing
before it becomes gasoline, which is the only usable form of fuel to drive the pistons in a gasoline
engine. Pouring crude or even refined oil in the gas tank would not work, even though the key
elements exist. This is the same reason proteins and lipids cannot be used for immediate energy
as they are still in a “crude” form.
ATP consists of an adenine molecule (a nitrogenous base), a sugar (ribose) and three
phosphate groups. The actual “energy” is held in the bonds that form its molecular structure.
When the phosphate bonds break (as a result of their negative charge and their tendency to repel
each other), energy is released.
ATP reaction:
The splitting of the ATP molecule frees energy, resulting in one molecule of adenosine
diphosphate (ADP), an inorganic phosphate (Pi), and roughly 7.3 kcal of energy. This does not
mean the body losses energy at this rate with ATP metabolism, but rather it releases energy into
the system during each reaction. The arrows in the formula above imply this reaction is
reversible, allowing for ATP to be metabolized and resynthesized. This is important because
ATP storage in muscle represents a relatively small quantity of energy at any given time. Relative
to the full spectrum of the metabolic pathways, it is important to understand that while ATP
will serve as the medium for reactions that drive sports, immediately accessible stores of ATP in
the muscle provide only enough energy for a single maximal action. Therefore, any energy
expenditure used to perform additional work will require the resynthesis of ATP via rephospho-
rylation – being the remanufacturing of ATP from energy-yielding compounds through various
metabolic pathways.
Sport Metabolism
The breaking of an ATP molecule frees energyresulting in one molecule of adenosinediphosphate (ADP), an inorganic phosphate (Pi)and roughly 7.3 kcal of energy released.
180

181
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
The Metabolic Pathways
The small quantities of ATP stored in muscle tissue necessitate an ongoing process of
remanufacturing to support static or dynamic muscle function. The more immediate the avail-
ability of ATP, the higher the potential force output from muscle contractions. This presents
relevant consideration for programming because the type of energy used indicates the absolute
force potential of the work. It also determines the length of necessary recovery between repeat
actions requiring the same force. The more reactions needed to produce ATP, the lower the force
associated with the contraction. Stored ATP requires a single reaction and therefore provides
maximal force, whereas the multiple reactions required for lipid metabolism to form ATP can
only support the force needed for an activity such as jogging. This is partially related to the
interactions between the energy pathways and select muscle fiber affinity for a particular path-
way. Fast-twitch fibers are optimized with phosphagen-derived pathways, whereas slow-twitch
fibers adapt more efficiently to the aerobic production of ATP from the breakdown of sugar and
fats – a much longer process requiring more reactions.
Each energy pathway is characterized by the specific metabolic substrate used to produce
ATP as well as its explicit purpose. The type of substrate and the dynamics of how ATP is derived
through its reactions provide for three metabolically-controlled performance factors. The path-
way determines 1) the associated availability of energy at a given time, 2) the quantity of potential
force produced, and 3) the duration of time needed for rest between repeat actions.
The immediate energy system is normally referred to as the phosphagen system due to the
interaction of phosphagen-driven fuel to support short duration, intense work. The system
reflects energy derived from stored ATP and phosphocreatine (PCr). Once stored ATP is
exhausted (≤3 seconds), new ATP is formed from stored or end-product elements within the cell
to support high-force actions lasting ≤15 seconds. Phosphagen stores in the muscle provide rapid
and powerful energy but have very limited support times. The body
accounts for this limitation using a second anaerobic system to maintain
ongoing work called the glycolytic pathway or glycolysis if referring to the
reaction. Sugar stored in muscle as glycogen provides anaerobic energy
for intermediate-length work demands lasting roughly 90 seconds. As a
consequence of the increased number of reactions needed to produce ATP
in this pathway, total force production declines. Sugar metabolism how-
ever, better accounts for sustaining force when compared to the phosphagen
system due to the higher provisional storage in muscle tissue. The higher
storage of sugar-based fuel provides for greater total work capacity at
intensities reaching moderate-high levels. This energy can sustain the work
necessary to perform an 800-m run.
Sport Metabolism
Figure 6.1 Practical Applications of the Metabolic Systems

182
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
If the body is forced to continue activity for a prolonged period of time, energy support
shifts towards a preference for aerobic metabolism. Aerobic metabolism provides energy for very
long periods of sustained work but at an even greater reduction in force potential. This signifi-
cant drop in force explains why most athletes do not train using the aerobic pathway as it does
not align with actual sports demands.
For the purposes of learning, these energy systems are typically presented in a manner that
seems arbitrary and absolute in terms of energy contribution related to time. In reality, they co -
exist in a much more cooperative and fluid state. It is important to comprehend that these
“metabolic” time frames are not concrete, but instead reflect a continuum (or overlap) of energy
production to support various physiological actions occurring at the same time. This concept
will be detailed later in the chapter. The reason each energy system is normally described as an
independent variable, often expressed by time and intensity, is the characteristics of each pathway
have particular nuances. Each system demonstrates unique aspects related to the efficiency of
energy-availability and the consequent contribution to work, expressed by force, during a
particular activity.
Realistically, all energy pathways work synergistically to drive human movement. A soccer
player, for instance, jogging down the field may break into a sprint toward the ball then perform
a maximal leap to a header resulting in a shot on goal. In this case the athlete is using all meta-
bolic pathways together. Dissecting each part by contribution is inconsequential; having the
support of each part though is paramount to success. Additionally, each metabolic pathway
represents provisional storage, which may vary based on the daily consumption of energy-yielding
nutrients and physical condition of the athlete. These facts underscore the relevance of both
specific conditioning by sport and adequate nutritional support for physical work and recovery.
Immediate (ATP Stores)
Textbooks commonly cite that immediate stores of ATP provide provisions for a 1RM effort,
or roughly three seconds of maximal work. The extremely short duration of tension is not
enough to provide significant adaptations, so most programs do not use single-repetition lifts.
Normally, 1RM efforts are only used during assessments, such as when testing Olympic cleans or
measuring vertical jump height. If a maximal effort is performed, recovery of depleted ATP stores
requires about 90 seconds for rephosphorylation. Some coaches challenge this notion suggesting
90 seconds is not enough time to recover from a single-repetition, maximal lift. In this instance,
the thought process is correct but out of factual context. Repeated maximal vertical jump meas-
Sport Metabolism
Figure 6.2

183
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
ures can be obtained accurately with a 90 second recovery, but one must consider the time-
under-tension and total mass employed for the movement. When a strength assessment such as
a maximal squat is used the conceptual factors change. A 1RM back squat requires a lifter to
remove the bar from the rack position, step to an appropriate stance, and eccentrically lower the
bar to the transition point before reversing the process. For a maximal effort, this takes more
than three seconds and uses many muscle groups. Therefore it is not reflective of the point that
stored ATP serves a single maximal effort, but rather the whole phosphagen system supports the
act – explaining why the recovery may be as long as five minutes before the effort can be repeated
successfully.
During the process of recovery from one of the aforementioned actions, ADP and an inor-
ganic phosphate use energy to re-bond into a new ATP molecule through rephosphorylation.
Rephosphorylation is used to manage energy because ATP is a very heavy molecule; prompting
the body to forgo storage in exchange for remanufacturing the product. Conceptually, it would
seem that rephosphorylating ATP and PCr back to initial stores would provide for long periods
of high-force work since a recovery period allows for renewed energy. The problem though, is
the process requires energy for reformation due to the negatively-charged bonds. This causes an
ongoing depletion of energy during system use, and ultimately fatigue over time. If muscle was
a closed energy system high-intensity work could continue for long periods, but since it is an open
system, phosphagen-based work usually totals 5-10 minutes of time-under-tension.
Immediate (Creatine Phosphate)
When the phosphagen system is considered, PCr more often provides the primary energy
to support high-intensity movements consistent with sports training. Creatine is an amino acid
produced by the human body which can be synthesized into PCr. The system synthesizes ATP
when PCr donates its phosphate group to ADP (PCr + ADP + energy ↔ ATP + Cr) to become
simply creatine. The depletion of PCr associated with muscle work requires between 2-5 minutes
for the process of rephosphorylation to replenish stores. The reason there is a range rather than
a single value is several factors affect recovery including 1) an athlete’s efficiency (based on train-
ing in the system), 2) total storage, 3) the duration and intensity of the action, and 4) available
oxygen. Since the phosphagen system supports most relevant sport outcomes, an athlete’s
exposure to the system often predicts efficiency in the recovery process.
A detailed examination of the phosphagen system through literary review indicates that
there are three reactions that comprise the resultant energy outcome. However, only the creatine
kinase and adenylate kinase reactions actually produce ATP. These reactions are named according
to the enzyme that catalyzes them for energy. The two reactions serve different purposes: the
creatine kinase reaction is important as it creates greater quantities of ATP, whereas the adenylate
kinase reaction serves as a signaling mechanism when PCr becomes depleted. During ongoing
intense work the body recognizes energy loss from the phosphagen system due to an increase in
adenylate kinase reactions. This signals the body to increase ATP production from the glycolytic
system. During this process some ADP is split to produce AMP which is used by enzymes to
initiate glycogenolysis (the breakdown of glycogen) and the production of an activated form of
glucose called glucose-6-phosphate (G6P). G6P provides the energy for continued exercise via
glycolysis.
In many texts, the phosphagen and glycolytic systems are described independently, but in
reality they are cooperative and interactive. This explains why time-under-tension, workloads
Sport Metabolism

184
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
and recovery periods are codependent for adaptations. When the phosphagen system is signifi-
cantly depleted using heavy, low-repetition actions (3RM), a longer-duration recovery is needed.
But, when that same exercise is performed at 6RM intensity the recovery shifts to a shorter-
duration requirement. This is due to 1) less PCr contribution and 2) the interaction of PCr and
glucose to fuel the muscle tissue. Ultimately, recovery time is based on the total requirements for
PCr rephosphorylation and the environmental levels of byproduct in the cell at the end of an
exercise set. The more an athlete is exposed to this system interaction, using the correct amount
of tension and metabolic stress, the more he or she will improve. In a study evaluating elite
sprinters at 40-, 60-, 80- and 100-m distances, those with better capacity to breakdown PCr
(marked by higher enzyme concentrations) achieved a higher maximal speed and were able to
maintain that speed for a longer period of time [1].
Once resting measures of PCr are depleted, they must be regenerated by creatine kinase (ATP
+ Cr → PCr + ADP + H+). Therefore, the ability of an athlete to recuperate PCr stores following
bursts of maximal efforts is central to their ability to repeat performance over an event’s entirety.
This ability is a crucial element in performance outcomes, particularly in explosive sports such
as American football, rugby, basketball, tennis, hockey and soccer. Research concerning how to
most effectively expedite PCr system recovery is not easy to conduct; resulting in limited appli-
cable findings in the literature. The available information indicates that heavy loading with high
velocity seems to be the most effective technique for increasing creatine kinase and adenylate
kinase activity. This makes sense if one again looks at the relationship between sport metabolism
and motor unit physiology. Heavier loading and ballistic actions recruit mostly larger and faster
motor units. This recruitment pattern preferentially recruits fast-twitch fibers, which have a par-
ticular affinity for the immediate energy system. Thus, it is no surprise that the average PCr
content at rest in type II fibers is about 20% higher than in type I fibers and these values can
increase with training [2].
The text has thus far supported an emphasis on the immediate system, but simply focusing
on improved phosphagen efficiency would not be optimal for overall sport performance. There
have been estimates that during a 10-second maximal sprint, the energy provided by PCr is lower
than previously thought [3]. While somewhat controversial, it is clear that athletes can improve
their PCr system through fast, heavily-loaded movements, but also by improving their oxidative
capacity through sprint-based training. This was made evident from a clinical trial employing
intense, 30-second repeat sprints on a Wingate. The resistance used for the high-speed sprints
was 7.5% body weight for males, and 6.5% bodyweight for females, respectively. Results indicated
an average 14% improvement in PCr recovery time following the six training sessions [4]. The
important note here is that the rest periods used were relatively long at 4 minutes. This duration
is in line with energy system specificity of the immediate system, explaining its impact on effi-
ciency. It also explains why some forms of high-intensity training (HIT) have failed to replicate
these results in the PCr system due to insufficient recovery. Conceptually though, maximal
improvements lie in the interactions of the system due to the dynamics of recovery. This suggests
a relational load-velocity–recovery matrix is most effective for programming compared to any
independent focus on the training. The use of heavy loading combined with high-speed ballistic
actions and sprint-based training, using appropriate rest intervals, will positively influence the
recovery-rate of the phosphagen system and improve energy availability.
Sport Metabolism
DEFINITIONS
High Intensity Training –
A style of training which uses minimalrest periods to greatly challenge thelactate tolerance system
185
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Intermediate (Glycolysis)
When exercise or sport-related tension begins to encroach upon the 10-second mark, the
adenylate kinase reaction described previously signals the need for energy support via the
glycolytic system. In response, enzymatic activity increases the rate of ATP production from
glucose and glycogen. Glycogen is simply the storage form of sugar; attaching glucose to water
to retain energy in muscles and the liver. Glycolysis can be generally thought of as the process
by which one molecule of glucose is broken down into two molecules of pyruvate, while
simultaneously synthesizing two molecules of ATP. An important molecule for energy transport
called nicotinamide adenine dinucleotide (NAD) is also activated, and helps in supporting the
energy continuum. NAD accepts H+ to form NADH, which can later facilitate ongoing reactions.
Compared to the immediate system, glycolysis is a relatively slower process in terms of ATP
synthesis as it requires 10 reactions; however it is still much faster than aerobic metabolism. See
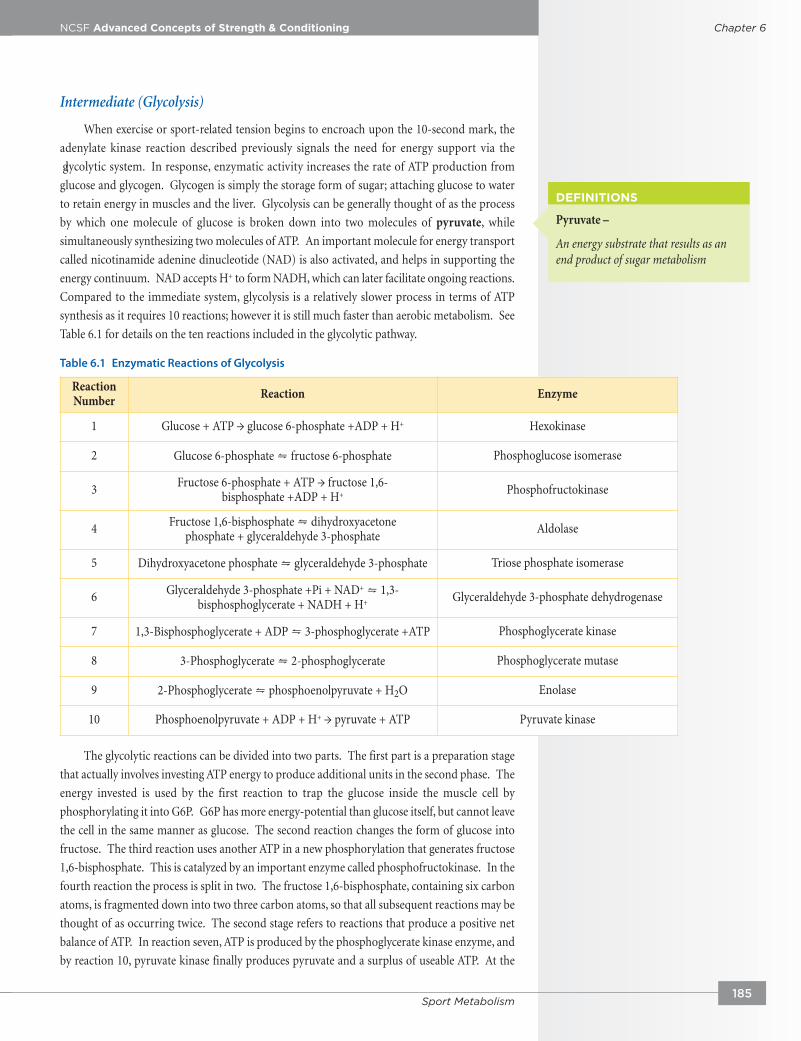
Table 6.1 for details on the ten reactions included in the glycolytic pathway.
The glycolytic reactions can be divided into two parts. The first part is a preparation stage
that actually involves investing ATP energy to produce additional units in the second phase. The
energy invested is used by the first reaction to trap the glucose inside the muscle cell by
phosphorylating it into G6P. G6P has more energy-potential than glucose itself, but cannot leave
the cell in the same manner as glucose. The second reaction changes the form of glucose into
fructose. The third reaction uses another ATP in a new phosphorylation that generates fructose
1,6-bisphosphate. This is catalyzed by an important enzyme called phosphofructokinase. In the
fourth reaction the process is split in two. The fructose 1,6-bisphosphate, containing six carbon
atoms, is fragmented down into two three carbon atoms, so that all subsequent reactions may be
thought of as occurring twice. The second stage refers to reactions that produce a positive net
balance of ATP. In reaction seven, ATP is produced by the phosphoglycerate kinase enzyme, and
by reaction 10, pyruvate kinase finally produces pyruvate and a surplus of useable ATP. At the
Sport Metabolism
DEFINITIONS
Pyruvate –
An energy substrate that results as anend product of sugar metabolism
ReactionNumber Reaction Enzyme
1 Glucose + ATP → glucose 6-phosphate +ADP + H+ Hexokinase
2 Glucose 6-phosphate ⇋ fructose 6-phosphate Phosphoglucose isomerase
3 Fructose 6-phosphate + ATP → fructose 1,6-bisphosphate +ADP + H+ Phosphofructokinase
4 Fructose 1,6-bisphosphate ⇋ dihydroxyacetone phosphate + glyceraldehyde 3-phosphate Aldolase
5 Dihydroxyacetone phosphate ⇋ glyceraldehyde 3-phosphate Triose phosphate isomerase
6 Glyceraldehyde 3-phosphate +Pi + NAD+ ⇋ 1,3-bisphosphoglycerate + NADH + H+ Glyceraldehyde 3-phosphate dehydrogenase
7 1,3-Bisphosphoglycerate + ADP ⇋ 3-phosphoglycerate +ATP Phosphoglycerate kinase
8 3-Phosphoglycerate ⇋ 2-phosphoglycerate Phosphoglycerate mutase
9 2-Phosphoglycerate ⇋ phosphoenolpyruvate + H2O Enolase
10 Phosphoenolpyruvate + ADP + H+ → pyruvate + ATP Pyruvate kinase
Table 6.1 Enzymatic Reactions of Glycolysis
186
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
end there is a balance of two ATP molecules gained since 4 were produced but two were invested
in the preparation phase.
The efficiency of the system is limited by the amount of reactions involved in the manu -
facturing process, as well as the number of compounds involved. As mentioned earlier, the
glycolytic system’s advantage lies in its storage potential. Whereas phosphagen storage is limited,
difficult to manipulate, and requires replenishment; glycogen storage is much more significant
and does not require replenishment to repeat force production. This allows for shorter recovery
periods. Another added benefit with storing energy prior to competition, rather than attempting
to fuel it during the event, is glycogen converts directly into G6P. Therefore, the body does not
need to invest extra ATP in the process, as seen with free glucose. The use of glycogen over glucose
represents a 50% increase in total energy production because the body saves on the initial
investment. ATP production from glycogen increases from two (2) to three (3) ATP. Due to this
efficiency differential, the body will prefer glycogen as its primary fuel for glycolysis; underscoring
the need for coaches to monitor pre- and post-activity nutrition.
The human body normally stores around 400-450 grams of glycogen, of which about 25%
is maintained in the liver to preserve adequate blood glucose levels. While still a viable option to
support the demands of energy for exercise and sports, it is particularly sensitive to the needs of
the central nervous system (CNS). This fact provides both positive and negative implications.
On the positive side, higher hepatic (liver) glycogen levels aid in the maintenance of blood glucose
during an event and help prevent fatigue. On the negative side, the liver is sensitive to the needs
of the CNS; therefore, when liver glycogen content drops too low (<55% capacity), central fatigue
occurs at a rapid rate. This negative consequence is often a result of poor pre-activity glycogen
levels, performance of high-intensity actions and prolonged exercise duration.
Many athletes associate the glycolytic energy system with higher levels of discomfort, and
many blame lactic acid as the instigator. Lactic acid is actually not the problem; it is a lack of
oxygen and an accumulation of hydrogen protons that create the metabolic issue. During
anaerobic efforts, the enzyme lactate dehydrogenase allows the body to produce lactate from the
lactic acid or pyruvate released during glycolysis. This actually improves efficiency, as lactate is
a metabolic transport vehicle that removes pyruvate from the system, therefore limiting the
saturation rate of the product that would normally slow down the process. Additionally, lactate
regenerates an inactivated version of the NAD carrier, which allows it to receive energy from
glycolysis. If an inactivated version of NAD is not present to receive energy in the form of
electrons, glycolysis will not be efficient. Lactate is also a hydrogen proton acceptor and lowers
acidity, buffering the internal system. It can be converted into glucose through the hepatic process
of gluconeogenesis to increase glucose availability, and can also be readily used by metabolic
tissues and the CNS for energy. During exercise, lactate spares valuable glucose and glycogen as
muscle and organ cells have the capacity to use its energy without the need to convert it into
glucose first. Lactate is an important player in protein-sparing, as the increased efficiency in
glucose management makes it unnecessary for the liver to use protein products to form glucose.
This suggests that training in the glycolytic system is also relevant to improving energy during
competition while limiting fatigue.
Longer Duration (Aerobic)
It has been illustrated that anaerobic metabolism is not compartmentalized, but actually an
interaction of several pathways working together to supply energy to working tissues. Aerobic
metabolism is often assumed to function independently as its own system, but it too works as
Sport Metabolism

187
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
part of the continuum. Aerobic metabolism simply reflects another shift in energy support; and
in the same way the immediate system signals the intermediate system to increase contribution,
the intermediate system signals the aerobic system to help support ongoing work. All cells need
oxygen, and without adequate supply they become dysfunctional. During anaerobic metabolism
no oxygen is needed to make fuel, but the cells still require oxygen to manage ongoing metabolism.
In aerobic metabolism, oxygen directly functions to form energy within the mitochondria of the
cell. The associated processes are referred to as mitochondrial respiration, where energy substrates
are metabolized in the presence of oxygen.
The anaerobic system produces pyruvate as a byproduct of glycolysis – which is now used
by the aerobic system to form acetyl CoA in the Krebs cycle. Also referred as the citric acid cycle,
the Krebs cycle represents a process of numerous molecular exchanges involving carbons,
hydrogen, and oxygen to produce energy. For example, glucose’s molecular chain represents
C6H12O6. When the glucose molecule is metabolized via mitochondrial respiration, the main
constituents are removed and 36 ATP remain as an end product. Since components of the glucose
molecule were removed during this process some energy is lost, while other components are
migrated to a different process to produce more energy. Due to the fact that aerobic metabolism
requires oxygen, it functions as a “pay as you go” system which enables the reactions to create a
stable environment during steady-state work. Unlike anaerobic glycolysis, there exist no
extraneous metabolites, such as hydrogen protons, that inhibit the process by affecting the cell’s
environment. Therefore, the act of producing force is ongoing and does not require a rest period.
It could be argued that while at steady state, low tension training could continue as long as energy
is available to keep the body moving. This explains the ultra-marathon, covering distances of
50-100 miles, as well as the need for refueling during distance race events.
Earlier text has stated that anaerobic athletes should not use aerobic training for improve -
ments in oxygen efficiency. The reason for this recommendation is oxygen is not just a fuel, but
a fuel-mediator. During anaerobic actions oxygen helps maintain proficiency in the anaerobic
systems. For instance, even well-trained anaerobic athletes experience decreases in tissue pH
during training, as lactate levels increase in working muscle and spill into the blood. This
chemical overflow triggers an increase in respiration through a signaling effect in attempts to
augment oxygen supply to working muscle cells. If an athlete has a higher oxygen capacity, they
will more efficiently buffer the anaerobic byproducts. The increased oxygen levels also promote
pyruvate clearance as it can be converted into Acetyl CoA, instead of lactate, and be used
aerobically. This substrate removal allows for improved glycolysis and increases the rate of ATP
production via mitochondrial respiration. This explains why a soccer player can tolerate higher
intensities for longer periods of time (aerobic) during the match, but also explains why they can
repeat high speed (anaerobic) sprints due to energy system interaction.
To this point sugar metabolism has rightfully been the focus of athletic-driven fuel; but lipids
and proteins can also be used in the aerobic system. Although they offer limited contribution
during team sports, endurance athletes must remain cognizant that both protein and fat represent
viable energy sources during prolonged, continuous work. In fact, an endurance athlete may use
as much as 15% of total energy from protein contribution via the breakdown of amino acids.
Lipids are even more important for endurance athletes, as higher fat oxidation rates support
glycogen-sparing. This will be explored in chapter 17. The detailed functions of the system
essentially involve the citric acid cycle and the electron transport chain. It is relevant to under -
stand though that lipid and protein products do not support higher-intensity work. Lipid use
for instance, is optimized below 70% VO2max (~62-65%).
DEFINITIONS
Mitochondria –
An intracellular organelle responsible for generating most of the ATP requiredduring cellular operations
Krebs cycle –
A series of enzymatic reactions thatoccur in the mitochondria, involvingoxidative metabolism of acetyl com -pounds, which produce high-energyphosphate compounds for cellularenergy

188
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism
Adaptations stemming from aerobic training are represented in both the cardiopulmonary
and muscular systems. An increase in stroke volume and cardiac output, both of which are critical
in improving maximal oxygen use, occur as the heart improves in conduction signaling and
metabolic efficiency. Training stress will also trigger architectural changes in the left ventricle
leading to desirable hypertrophy and increased contractile force capacity. Often overlooked is
the fact that training to improve cardiovascular conditioning reduces systolic blood pressure and
relative heart rate responses; consequently lowering rate-pressure product (RPP). By reducing
the RPP, the muscles of the cardiopulmonary system do less work, which spares energy over the
duration of the competition. When an athlete can exert the same force at lower cardiovascular
demands, the energy saved from cardiopulmonary function can be used by skeletal tissues for
improved endurance. This benefit is added at the same time the skeletal muscles increase
in capillary density and fiber efficiency for better oxygen management. A major structural
adaptation is an increase in the expression of mitochondria within muscle fibers.
These adaptations demonstrate relevant consideration for sports training, but the method
by which the adaptations are reached is a sensitive issue. Due to the permanent inverse relation -
ship between motor unit power capacity and aerobic efficiency, it is important not to improve
one at the expense of the other. More specifically, if a higher aerobic training time is employed
using low-to-moderate intensities while also training for strength and power development, any
improvements in the anaerobic aspects will be proportionally negated by improvements in the
aerobic system. This is common of aerobic training used for baseball pitchers, boxers and tennis
players. Steady-state training as a means for conditioning anaerobic athletes is generally
counterintuitive to maximal performance benefit. Conditioning drills need to be performed at
sport-speed, and the rest intervals need to be short enough to allow for adequate metabolic stress.
The emphasis is placed on system interaction, not simply aerobic efficiency. Combined or
concurrent system interaction aimed at strength, power and anaerobic metabolic endurance
(relative to the sport) is the best option for most team sport athletes.
A recently published example of this concept employed professional soccer players from
two categories; a conditioning group and a control. Both groups engaged in regular soccer
participation, but the treatment group also performed an 8-week resistance training program.
The program consisted of two concurrent anaerobic training sessions per week; which included
4 sets of 6 maximal-effort repetitions of high pulls, jump squats, bench press, back squat and
pull-up exercises with interval training using 16 maximum-intensity sprints; lasting 15 seconds
with a 1:1 work to rest ratio. Those that participated in the concurrent sessions not only experi -
enced greater performance improvements in vertical jump height as well as 10- and 30-m sprints
speeds, but also in the Yo-Yo intermittent recovery test. Most importantly, these athletes increased
their maximal aerobic speed, which is a major factor for success in soccer [5].
The Continuum Concept
Energy systems interact more than they function in a segregated fashion. Therefore, it is
more practical to view them by duration and intensity versus simply segments of time, particularly
when considering the overlap of metabolic process between them as intensities increase
(continuum concept). The first step to using the energy systems for programming for specific
adaptations is to realize that during any given activity the respective energy systems interact in
attempts to create metabolic homeostasis. Therefore the interactive relationship should be
emphasized when conditioning athletes.
189
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
To accomplish this goal there are three major factors to take into consideration:
1) The training tenure and current condition of the athlete or team – An untrained or
novice athlete will not have foundational system aspects necessary to support high
quantities of work, particularly when high-tension is employed. Detriments in training
performance will be associated with both neuromuscular and metabolic issues;
essentially a coach cannot bypass pre-requisite adaptations by training harder or longer.
2) The volume of the work performed and total recovery – Athletes maintain limited
provisions of energy at a given time. Therefore, activities must be balanced across
phosphagen-driven acts and those supported by glycolysis. Adding volume with or
without changes in frequency requires consideration for proper nutrition and recovery
to account for residual fatigue. If the body is pushed beyond its energy support it will
suffer from fatigue; and consequently, muscle damage.
3) The end-product and rest interval relationship –When creating work-to-rest relation -
ships a coach must consider the athlete’s efficiency in the specific energy system as well
as the total energy system interaction. This is based on the intensity, each action’s
duration, and the rest interval used between sets. Following every sprint there are
residual metabolic byproducts (residuals) in the system. If partial recovery is used, the
athlete’s condition for the next set = residuals + new byproduct. By the fifth set of
sprints (with partial recovery); the athlete’s condition would be = 4x residuals + new
byproduct. This suggests an athlete cannot go from 0-100%, or maximally, over the
course of a workout because the system does not start at zero each time. Rest intervals
should be programmed by residual tolerance with sport-specific purpose and be
adjusted to maintain desired velocities.
Most programmatic difficulties are associated
with controlling all the factors at play. For coaches
to determine proper balance (intensity/duration =
recovery schematics) they must account for each
factor. Specifically, what affect each factor plays
on the recovery demand as well as how long the
recovery must be to 1) drive the energy-specific
adaptation, 2) allow for optimal movement tech -
nique, and 3) permit work at the desired velocity.
High speed, low load, short duration: 20 m shuttle
(5 sec) – byproduct: low ADP/H+; recovery required
is 1:4-6 due to the low percentage of energy used
High speed, moderate load, short duration: 7 tuck
jumps, loaded 10% BW (6 sec) – byproduct: low-
moderate ADP/H+; recovery required is 1:5-7
Moderate-high speed, high load, short duration: sled drive loaded 30% BW (6 sec) – byproduct:
moderate ADP/H+; recovery required is 1:7-9
High speed, high load, short duration: Olympic cleans 3RM (10 sec) – byproduct: high ADP and
moderate H+; recovery required is 1:12-15
In the prior examples all of the activities were performed using the phosphagen system, but
Sport Metabolism
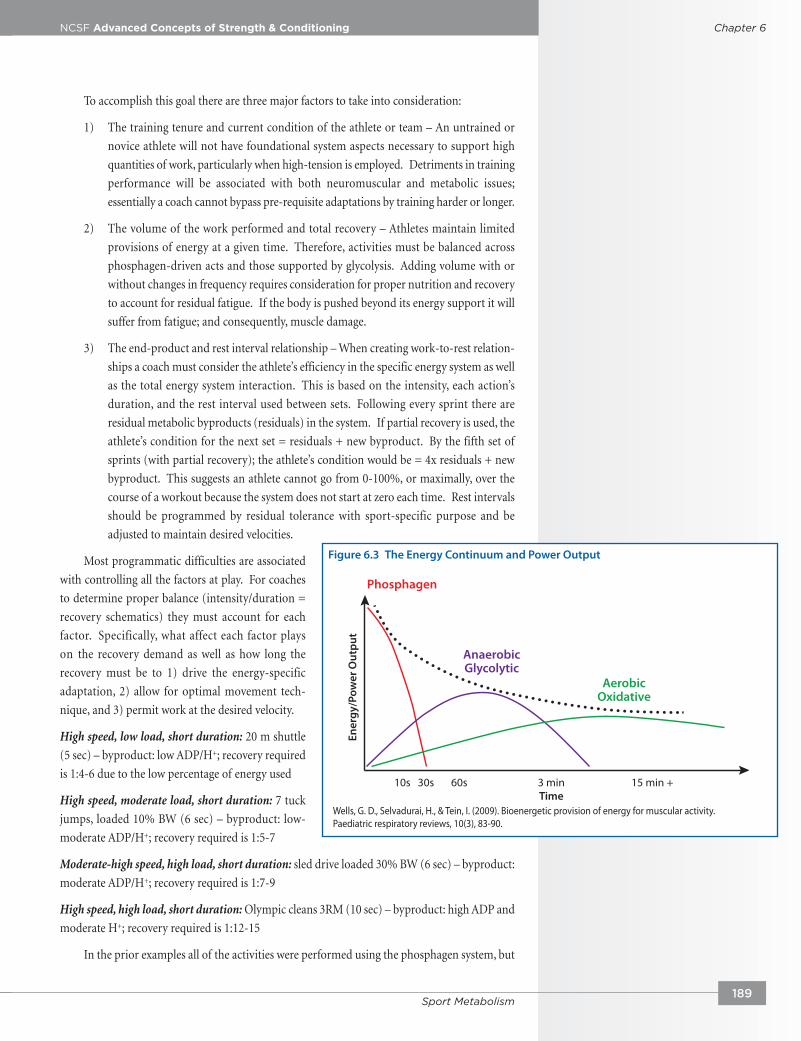
Figure 6.3 The Energy Continuum and Power Output
Wells, G. D., Selvadurai, H., & Tein, I. (2009). Bioenergetic provision of energy for muscular activity. Paediatric respiratory reviews, 10(3), 83-90.

190
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
the necessary recovery varied for each. Factors specific to the load, velocity, time-under-tension,
total muscle mass employed and the resultant rest interval all affect relative performance.
Therefore, assigning a rest interval simply based on the duration of the activity is flawed. A 20
m shuttle lasting five (5) seconds would not justify a full recovery of 120 seconds because the
action will not fully drain the system. The Olympic cleans involved resistance of a much higher
load, and therefore used much more energy; thereby warranting a recovery 4-6 times longer than
the shuttle run even though both were maximal-effort, phosphagen-driven activities.
This concept becomes even more complicated when systems overlap. Consider the follow -
ing: a high-speed activity, such as eight (8) repeat plyometric tuck jumps, will be supported mainly
by phosphagen system because the duration of time needed to complete the act will be less than
10 seconds. A back squat, on the other hand, would likely take about 25 seconds to complete the
equivalent eight (8) repetitions using 80%1RM. Here, PCr stores overlap with glycolysis to
support the activity because it takes a longer period of time to complete. Interestingly, the tuck
jumps will warrant 30 seconds recovery to repeat the performance, whereas the back squat will
require a rest interval of about 90 seconds. To some, this may not make sense because the tuck
jumps were performed using the phosphagen system, which normally is associated with at least
120-300 second rest periods; while the squats were performed using the glycolytic system, which
usually warrants 30-90 seconds for recovery. Again it is the relative intensity, time-under-tension,
total energy demands and byproduct residuals that determine the rest interval – not simply the
amount of time it took to complete the act. Many practitioners fail to realize the practicality of
the energy systems. The duration of the rest must always be consistent with the quantified use
of available fuel, the time it takes to clear limiting by-products from the environment associated
with metabolic disruption, and the relative condition of the physiological systems.
There is a distinct difference in the need for recovery when comparing actions driven by the
glycolytic and PCr systems because the resultant fatigue is based on two different effects. Anaerobic
glycolysis represents the breakdown of sugar from circulating glucose and stored glycogen in
muscle, so the byproduct includes H+ protons and limited ADP residuals. The phosphagen system
on the other hand results in ADP and limited H+ protons residuals. Additionally, sugar is stored
in higher concentrations than phosphagen in the muscle tissue, and therefore is not depleted to a
point of inhibition. Due to the fact that sugar metabolism produces ADP at a slower rate when
compared to the PCr system, the body does not have to wait for
rephosphorylation during recovery between glycolytic-driven actions
as it does with phosphogen-driven work. Instead, it must remove the
H+ and related byproducts from the environment to continue to
produce force. With ongoing work, H+ accumulates in both the cell
and blood, promoting a progressively acidic environment. This acid
accumulation lowers cellular and blood pH levels which inhibits
enzyme activity and “turns off” the energy system – but not due to a
lack of available energy.
Athletes with the most experience working in the phosphagen
and glycolytic pathways with recovery matched for proper adap -
tations will benefit from two perspectives: 1) a prolonged higher-
intensity work capacity and 2) they will require shorter periods
of recovery between sets or activities on the field. This identifies
why training experience and specific stress exposure are so impor -
tant in athletic conditioning for sport. A body unaccustomed to
Sport Metabolism

191
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
work will not manage relative byproducts or heat efficiently, and will therefore shut down
prematurely. Likewise, rest intervals must be aligned in accordance with the energy demands or
the metabolic system will not properly acclimate to the situation. Too long of a rest interval will
reduce the interaction of systems while too short of a rest interval will compromise the
phosphagen system’s contributions to force and velocity. In order to achieve maximal athletic
performance, one must understand the fundamentals of each metabolic pathway and how a
continuum exists between them.
Sport-Specific MetabolismRegardless of a sport’s metabolic preference, all team sports require energy
from each of the aforementioned systems. This is a relevant concept as different
sports are better suited for different energy systems, but no team sport depends
exclusively on a single system. Coaches must therefore attempt to determine the
balance or distribution of energy requirements and the respective relationship
between systems for each sport using evidence-based criteria. From a sport-specificity
perspective, an analysis of the energy system’s contributions should be reflected
accordingly in the training. A common error is to emphasize the aerobic or glycolytic
pathway during conditioning, but analysis of most team sports suggest that event-
determining outcomes mainly occur in less than 10 seconds and at the highest
velocities. For example, the usual duration of a rally in volleyball lasts 6-10 seconds,
during which there is a need for quick lateral movements and explosive jumps [6].
Following the energy continuum, approximately 50% of the energy would be
provided by the PCr system, 35% by glycolysis, with only 15% support from the
aerobic system. Tennis players face a similar situation; while matches can go on for hours, the
sport actions per point last only seconds. A specialist in the 200-m distance swim (efforts up to
two minutes) on the other hand, could expect 4% of the energy to come from the PCr system,
46% from glycolytic contribution, and 50% from aerobic support. Extending the swim effort to
10 minutes causes the relative contribution of the systems to adjust to 1% PCr, 9% glycolytic
and 90% aerobic [7].
Figure 6.4

192
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Depending on the nature of the activity, some sports may require a varied combination of
system support as different demands are applied throughout an event. For example, a soccer
player will experience moments of low-intensity activity (1/3 of game time is spent walking) as
well as several series of vigorous sprints. In this situation, the system predominance during a
game is aerobic, but the game-changing events all occur at the highest speeds; reflecting very high
force demands. Therefore, the PCr system will be the most relevant system for success in soccer,
but acknowledgement and attention must still be paid to the other areas of the game to prevent
fatigue-related performance decline. At this point, the interaction of conditioning becomes most
relevant. In most team sports, the phosphogen system should be exploited using ballistic training,
sprints and interplay; with rest intervals determined based on the sport’s aerobic system demands.
Fundamental Applications of the Neuromuscular SystemMotor units serve as the functional component of muscle. They are comprised of a nerve
and a homogenous group of attached muscle fibers. When innervated by the nerve all of the
attached muscle fibers contract simultaneously to create force. Interestingly, while all motor units
need energy to produce force, all motor units do not use energy in a uniform manner. Rather,
they demonstrate preferences to particular energy pathways based on structural and enzymatic
distinctions found in muscle cells. These inherent differences are seen through a motor units’
expression of force, velocity, and endurance. Due to the fact that all the fibers of a single motor
unit have consistent characteristics, when innervated they all function in the same manner. From
this perspective the structural and metabolic properties of the motor unit is commonly referred
to as the fiber type.
Fiber types can be differentiated by several characteristics, but mainly by their ability
to use energy. The enzyme concentration within the myofilament structures, more
specifically myosin, heavily determine whether the contraction speed is fast or slow. The
more ATPase found in the myosin, the faster the ability of the muscle fiber to contract.
Since there are many isoforms and hybrids of myosin, most classifications differentiate
two types of fibers; fast twitch (type II) and slow twitch (type I). Fast-twitch fibers may
be further divided into Types IIA and IIX. The reason for the secondary classification of
fast-twitch fibers is the force-energy system specificity associated with each. Analyzing
fibers by these distinctions identifies an important relationship between the fiber
characteristics and its alignment with a particular sport. This is partly where genetics
connect to performance measures as certain fibers are better suited for certain sports
situations. Essentially, elite athletes in a particular sport have the best composition of fibers
for the relative demands of the competition. This identifies why a 100-m sprinter does
not also compete in the 800-m event, and explains the very limited crossover of athletes
between professional sports.
Athletes at the highest level of competition have a genetic make-up that aligns with the
specific demands of a sport. If the demands require the fastest movements for short periods, as
seen in the Olympic 100-m race, the athlete will need a preferential distribution of fast-twitch
fibers. If the race is extended to 5000 m the athlete would need a better distribution of slow-
twitch fibers to complement the endurance requirements. It would be difficult for an athlete to
successfully compete in both race distances because an inverse relationship exists between fibers;
Sport Metabolism
The more ATPase found in the myosin, the faster the ability of the muscle fiber to contract.

193
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
when fibers produce high force they fatigue rapidly, whereas fibers with the best endurance
produce limited force.
Fast-twitch, type IIX fibers are highly excitable and fire rapidly due to a well-developed
sarcoplasmic reticulum (SR) and a high concentration of ATPase. They produce roughly 50
grams of force per motor unit and demonstrate the largest diameter by cross sectional measures.
The performance limitation to type IIX fibers lies in their fatigue rate. This identifies why a
percentage of these fibers migrate toward type IIA characteristic as an adaptation in response to
training. Initial and ongoing training promotes force endurance within fast-twitch fibers;
demonstrating that fiber characteristics change to accommodate applied stresses. When the
fastest-twitch fibers are recruited for high-force activation they fatigue in roughly five (5) minutes
of accumulated time-under-tension – to a level that no longer warrants training in that system.
This explains why phosphagen-supported training intensities cannot be sustained for long
durations. Type IIA fibers serve an intermediate role for both high force and endurance. At peak
activation, each motor unit produces approximately 30 grams of force. The firing rate is slightly
lower due to a reduced SR complexity and myosin efficiency, with absolute force production
limited by differences in total circumference size (compared to IIX fibers). The uniqueness of
the type IIA fiber is its ability to share responsibilities. The high-force contribution from the
fiber fatigues at approximately six (6) minutes under significant tension. However, unlike type
IIX fibers, IIA fibers do not fully exhaust to a compromised condition. Instead, they shift to a
lower force capacity. Motor units fatigued from high-force contribution will drop output to
about four (4) grams of force. Type IIA motor units can maintain force at this rate for extended
periods of time to aid Type I fibers in endurance activities. For this reason, IIA fibers are referred
to as a fast-twitch, fatigue-resistant or fast-twitch oxidative fibers, due to their capacity to function
aerobically. Type I fibers represent the slow twitch category. The slow twitch is a result of a less
complex SR and reduced myosin capacity. Interestingly, slow-twitch fibers are the first fiber
recruited due to a low threshold for excitation, but contribute the lowest levels of force; about
five (5) grams per motor unit. They are the smallest of the fibers (by circumference) and do not
demonstrate a significant capacity for hypertrophy or force with resistance training, but they
are resilient. Slow-twitch fibers rely mostly on mitochondrial respiration, whereas Type II fibers
are more capable of utilizing the PCr and glycolytic systems.
It is important to note a few concepts related to
generalizing in this classification system. When
examining an entire muscle, a continuum of muscle
fiber types can be identified. Therefore, no muscle
is characterized by only type I or type II fibers.
Likewise, individual muscle cells may be hybrids
(e.g., I/IIA, IIA/IIX, I/IIA/IIX); adding further
diversity to recruitment dynamics. Finally, even if a
particular fiber was identified as type I or type II, or
a specific hybrid (i.e., I/IIA), it should be understood that differences exist among muscle types
in the myosin heavy chains from molecule to molecule. This supports the role of adaptations;
as all skeletal muscle tissue has the capacity to change to accommodate new stresses applied to it.
The genetic factor lies in the initial distribution of muscle fiber types, the proficiency of an
individual’s nervous systems, how quickly and to what magnitude the athlete can adapt, and the
inherent potential for change within the athlete.
The ability to change an athlete’s distribution of slow or fast twitch fibers does not seem
Sport Metabolism
DEFINITIONS
Sarcoplasmic reticulum –
A tubular network that surrounds each individual myofibril and acts as a storage site for calcium within skeletal muscle
Table 6.2 Muscle Fiber Force Contribution
Fiber type Force per unit Fatigue rate
Type II X 50 grams of force per motor unit 5 minute fatigue rate
Type II A 30 grams of force per motor unit 6 minute fatigue rate4 grams of force per motor unit Fatigue resistant
Type I 5 grams of force per motor unit Fatigue resistant

194
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
possible as findings do not support a transition from slow twitch to fast twitch as an adaptation
response. Rather, research suggests the most common adaptation involves characteristic changes
in the muscle fiber architecture and cross-sectional area [8]. This adaptation provides for greater
production of force via parallel fibers within the muscle. Essentially, when a muscle fiber grows
in size it produces more force. Therefore, since genetic manipulation is limited, it is better to
understand these factors as useful in gauging performance potential(s) only. A coach can be
most impactful by adjusting training efforts to account for an athlete’s natural limitations. As
mentioned earlier, there exist specific fiber compositions and efficiencies that best serve different
sports based on their characteristics, but those concerns are better addressed by recruiting the
right athlete for the right position. For example, long-distance runners have shown a higher
percentage of type I fibers in the gastrocnemius (71.1%) compared to middle-distance and
recreational runners at 56.3% and 59.8%, respectively [9]. Essentially at the elite level, the sport
will pick its athletes, not the other way around.
Metabolic Implications for Training
The characteristics that define the performance of skeletal muscle fibers also delineate the
specific training necessary to exploit the fibers’ adaptation potential. With this in mind, the
volume within exercise programs can be tailored to specific adaptation responses. The first step
is matching the energy system and fiber recruitment preferences with the volume of work. Fast-
twitch fibers thrive in the phosphagen system but fatigue within minutes of accumulated
near-maximal tension. This suggests efforts aimed at adaptations in the immediate system require
high intensities with limited volume and appropriate rest intervals. Emphasis should be placed
on maximal loading (or speed) and technique for movement proficiency. Attempting to prolong
periods of heavy work (>85%1RM or maximal power) is counterintuitive to the adaptations.
This is where many coaches make programming mistakes. Based on the fatigue rates of fast-
twitch fibers, total work attainable through the phosphagen energy system will equate to less than
5-10 minutes of total time-under-tension. This should be measured by time of contraction.
Longer durations of tension at this intensity are not beneficial, and commonly lead to uninten -
tional overtraining syndrome.
When the intensity is reduced to 70-85%1RM (or velocity is reduced), time-under-tension
may double as metabolic support shifts to the glycolytic pathway. As a consequence of reduced
Sport Metabolism
Table 6.3 The Biochemical Characteristics of Muscle Fiber Types
Muscle Fiber Characteristic Type I Type IIA Type IIX
Capillary density 1.0 0.8 0.6
Mitochondrial density 1.0 0.7 0.4
Myoglobin content 1.0 0.6 0.3
Phosphorylase content 1.0 2.1 3.1
Glycogen content 1.0 1.3 1.5
Triglyceride content 1.0 0.4 0.2
Phosphocreatine content 1.0 1.2 1.2
Myosin ATPase activity 1.0 >2.0 >2.0
Phosphofructokinase activity 1.0 1.8 2.3
Adapted from: Sport Nutrition: An Introduction to Energy Production and Performance, 2nd edition;Jeukendrup & Gleeson

195
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
loading during work sets, training regimens aimed at moderate to moderately-high intensity
work increase to as much as 20-30 minutes of time-under-tension. The actual amount of time
is determined by the load used to create the volume, particularly if combined with phosphagen-
based activities. A traditional hypertrophy workout for instance, may use 10-12 exercises with
3-4 sets of 8-12 repetitions. At approximately 30-40 seconds per set, the time-under-tension will
range from 15-30 minutes (depending on the schematic). Conditioning (tension) periods will
be even less. Attempting to increase or maximize volume for the purpose of simply doing more
work will most often lead to residual muscle fatigue, poor training technique and compromised
recovery. Adaptations that promote sport performance from anaerobic training are derived from
the details of the program, not on maximal training volume. It is important to realize fitness
training is different than sports training as it attempts to prolong activity to burn calories, whereas
sports performance enhancement is specific to adaptations in an attempt to spare calories through
improved efficiency.
As acknowledged earlier, aerobic training is not commonly used during team sport condi -
tioning. Endurance sports however, rely on aerobic metabolism for success. Here adaptations
continue to reflect the metabolic- and tension-specificity for motor unit recruitment. The
distances covered by most endurance sport athletes require prolonged durations of continuous
activity. Interestingly though, training for endurance events should also be based on the inter -
action of the anaerobic and aerobic systems. Among elite endurance athletes, VO2max is not the
primary factor separating the winners from the losers; it is normally the anaerobic system that
makes the difference. Those individuals with both an efficient aerobic and anaerobic system
usually dominate the race. Anaerobic training emphasizes high-intensity intervals for up to 20-
30 minutes to increase demands placed upon fast-twitch fibers; whereas aerobic adaptations such
as slow-twitch fiber hypertrophy and improved lipid oxidation are derived from long, slow
distance training for 60-90 minutes (or more). Endurance athletes must use the metabolic
pathways to adapt both slow- and fast-twitch fibers to promote the desired performance benefits.
Gradation of Contractile Force
It was stated earlier that within a motor unit a distinct number of muscle fibers are con-
nected to each nerve which signals information to each fiber using what are referred to as action
potentials. An action potential starts as an electrical impulse that runs along the nerve. When
DEFINITIONS
Action potential –
A wave-like change in the electricalproperties of a cell membrane, electricalsignaling causes a chemical message thatforces the muscle cell to contract
Figure 6.5

Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism
it reaches the neuromuscular junction the electrical impulse is converted and relayed via
chemical signaling to all of the fibers innervated by that motor neuron. This process of
events is known as the all-or-none principle. When considering force gradation, it
is important to understand that contrary to the individual fibers within a motor unit
which all fire at the same time; the individual motor units throughout an entire muscle
are not innately synchronized. Since gradation of force, or tension, has the capacity to
vary at different points, sports specificity becomes more relevant for improvements in key
areas. In order to execute sport movements successfully, the athlete must precisely adjust
the tension within the musculature to regulate specific actions. If the athlete activates too
many fibers or not enough; the action may falter or fail. Furthermore, if the recruitment
pattern is not organized, the movements of associated bodily segments lose force effi-
ciency. This includes syncing motor unit recruitment so slow-twitch fibers, which
stabilize motion segments, precede fast-twitch fibers to maximize force transfer across the
segment. Essentially, if the motor units fire at different times, or they lack proper
sequencing or synchronization, the precision needed within an action will not occur.
What often separates the good athletes from the great athletes lies in the measures of intel-
ligence and coordination within nerves.
The muscular system has the capacity for incredible force output, but that capacity lies
mainly in potential. Untrained females for instance, have been observed lifting extremely heavy
objects in efforts to save their child. Certainly, adrenaline is the driving force behind this remark-
able feat, but the fact remains that the individual’s musculature was capable of producing forces
well beyond the tissues measurable 1RM.
This observational data suggests if the nerv-
ous system is able to recruit the correct
quantity of motor units with precise timing
– it can far exceed its normal performance.
The ability of a strength coach to extract as
much of this potential as possible is the key
to success. A caveat to this concept is the
training must be aimed at applicable force
(sports proficiency), not just weightlifting
force.
Motor Unit Firing Rate, Recruitment and Synchronicity
Muscle force (and consequently movement control) is based on motor unit firing rates,
recruitment and synchronicity; all of which are trainable components of the nervous system.
However, there seems to be differences in adaptation responses when comparing small and large
muscle groups. Adaptations to firing frequency are more easily attained in smaller muscles,
whereas improved motor unit recruitment seems to be more pronounced in larger muscles.
This is likely due to the ratio of muscle fibers recruited per motor neuron in the respective areas
as well as the specificity of the tasks they perform. The thigh musculature for instance, may
have a ratio of 1:3000, whereas the calf may have a ratio 1:1800; very small muscles within the
hand are further refined to a ratio of 1:100. Where recruitment and firing rate seem to have
some adaptation specificity, factors related to improved synchronicity seem to be based on motor
rehearsal and can be improved to an equal extent in most small and large muscles, respectively.
In regards to motor unit firing, research has indicated that a change in firing rate occurs in
An action potential starts as an electrical impulse that runsalong the nerve to the neuromuscular junction; at whichtime the electrical impulse is converted and relayed viachemical signaling to all of the fibers innervated by thatmotor neuron.
All-or-None Principle
DEFINITIONS
Neuromuscular junction –
Physiological structure that allows anaction potential from a nerve toinnervate a muscle via an electrical tochemical conversion
All-or-none principle –
A muscle cell either fully contracts, ornot at all; if a nerve impulse is at leaststrong enough to depolarize the cell, itwill contract regardless of the absolutestrength of the impulse
196

197
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
response to adjustments in the threshold stimulus. The reduction in electrical threshold allows
for an earlier release of signaling, and consequently a more rapid firing rate. Additionally, firing
rate has been identified as a key component in the development of important performance
mechanisms such as the rate of force development (RFD) or “explosiveness”, which represents
one of the best predictors of athletic ability [10,11,12,13]. The most effective means for achieving a
reduced firing threshold is the performance of ballistic training. Researchers have found an
increase in the percentage of motor unit firing doublets (two motor unit signals that occur at
very short intervals) following 12 weeks of movement-specific power training. Specifically, firing
doublets increased from 5.2% to 32.7% [13]. The best results in terms of firing frequencies are
usually detected when the athlete is beginning his or her training process. This explains, in part,
the improvement rates noted in the initial stages of training compared to adaptation responses
that occur later on in the athlete’s tenure. An untrained athlete will see significant improvements
in strength and power during the first 4-6 weeks of training as motor units improve their firing
efficiency. In addition, the firing patterns learned can be environmentally specific. For example,
elite sprinters demonstrate maximum motor unit signaling frequency at the start of a 100-meter
race, as a fast start is paramount to success [14]. This supports the rationale
of sport-specificity in training to develop neural signaling in a manner con-
sistent with the sport.
The size principle has been used to describe the activation pattern
of motor units. Smaller motor units that innervate type I fibers will be
activated before the larger, more powerful motor units that innervate and
excite the type IIA and IIX fibers. The size principle comes into play as a
result of the different thresholds of each unit. The smaller units have lower
thresholds, thus requiring lower electrical levels of activation than those
required by larger motor units. This is likely due to the need to stabilize a
segment before loading it and to spare energy by using smaller units when-
ever possible such as to support postural requirements. Attempts have been
made to promote recruitment adjustments, but studies suggest that the size
principle remains unaltered even after periods of power training [13]. How-
ever, thresholds for recruitment of high-power motor units can be reduced with ballistic and
plyometric training; demonstrating the important role of power exercises in explosive movement
development. Training programs which properly emphasize velocities and loading can optimize
recruitment-rate specificity. A combination of ballistic training and compound exercises using
heavy loads seems to best exploit this adaptation response.
Fast-Twitch Recruitment Patterns
When analyzing recruitment factors it is important to understand that specific motor units
need to be trained using speeds, power levels, and patterns that mimic the dynamics of the sport.
For instance, if a given sport demands quick explosive power, training should involve similarly
characteristic movements that require recruitment of the fastest-twitch fibers in order to reduce
motor unit thresholds. For concurrent improvements in firing synchronicity, it is important that
both the movement patterns and training speeds remain consistent with the speed and type of
sport actions. When training employs actions that resemble the sport requirements in whole or
in part, at the proper velocities, improved firing synchronicity has been noted. Analysis has
shown that improvements occur in the connections between the motor neurons, which results
in enhanced coordination and an increased rate of force production [15].
DEFINITIONS
Size principle –
Tension on muscle promotes progressiverecruitment of fibers based on motorunit size and activation threshold;smaller motor units that innervate Type I fibers are activated before larger,more powerful motor units thatinnervate Type II fibers
If a given sport demands quick explosive power,training should involve movements that requirerecruitment of the fastest-twitch fibers so thattheir motor unit thresholds can be reduced.

198
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism
Considering these concepts, it is important to provide appropriate balance between the
resistance and speed used during training for sport. There are two developmental considerations
here; (1) ensuring adequate prime mover strength, strength-balance across movement segments
and stability across the skeleton, and (2) ensuring load applications properly serve the force-
velocity curve. These suggest faster speed is more important than heavier loading. Expectedly,
the two most common errors in promoting motor unit adaptations for sport are the performance
of ballistic training without musculoskeletal balance, and excess loading at the expense of move-
ment velocity. Therefore, phasically-appropriate resistance assigned to compound and ballistic
movements is extremely relevant. A key element in the program is to understand that when too
much weight is employed during ballistic and plyometric exercises – the benefits of the motor
unit adaptations will be relatively reduced. Common examples include weighted jumps using
heavy loaded vests or excessive dumbbell loads, overloading drag or sled training, and using high
heights for depth jumps. When these types of activities are properly employed, jump resistance
should be less than 12% of total bodyweight, loads used for drag training should not have any
notable effects on running biomechanics (3-8% BW) and depth jumps should occur at heights
that allow for the shortest amortization phase possible (maximally 30 inches – less than 18 inches
is recommended). The force-velocity curve should be referenced for appropriate load and
velocity by training phase. A progressive model in training loads and speeds should logically
reflect a neuromuscular path towards consistent game speed. Additionally, when deciding on
traditional weight training volumes, a strength coach must be sure to consider adequate quantities
of athletic-based ballistic training. This is exemplified by the fact that greater improvements in
vertical jump height were found among NCAA Division I volleyball players following ballistic
jump squat training using loads at 30, 60, and 80% 1RM, when compared to traditional heavy
(squat) training [16]. Later chapters will proportionate concurrent strength training and intensity-
specific power training in a progressive model.
Increased Motor Unit RecruitmentThe execution speed of exercises may also be reduced to promote specific adaptations.
During the earlier phases of a training cycle a coach may emphasize muscle tension via slow-
velocity training (tempos) for certain benefits. A recent study showed that the performance of
leg extensions using a relatively light load (30% of 1RM), executed at a slow tempo of six (6)
seconds of concentric work and six (6) seconds of eccentric work, produced a greater increase in
muscle protein production than when executed at a fast tempo of one (1) second up and one (1)
second down [17]. While not surprising for its “bodybuilding effects” on muscle growth, the
stimulus also increased the number of motor units recruited. Bodybuilders have long identified
that as time-under-tension increases, motor unit fatigue triggers greater motor unit recruitment
and anabolic responses. This has application for both hypertrophy and strength as a result of
fiber cross-sectional improvements.
Activities aimed at improving sport performance will not involve twelve-second repetitions
that activate isolated muscle groups, but evidence related to the time-tension relationship on
motor unit recruitment holds merit. Hypertrophy-endurance and hypertrophy-strength phases
can both exploit this condition by utilizing athletic lifting techniques such as squats, deadlifts,
presses and related compound movements to increase motor unit recruitment in muscles used
for sport training and performance. Sport differences will determine the specific emphasis on
muscle growth, but it is important to recognize that improvements in muscle force production
DEFINITIONS
Amortization phase –
Also known as the transition or contactphase; constitutes the period of timebetween the concentric and eccentricphases of a plyometric exercise whereinthe stretch-shortening cycle is exploitedto maximize power production
Bodybuilders have long identified that as timeunder tension increases, motor unit fatiguetriggers greater fiber recruitment.

199
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
are associated with 1) an increased cross-sectional area of associated muscle fibers and 2) the
recruitment of more motor units as a result of increased time-under-tension. Both are beneficial
to performance because these newly-recruited motor units will become more active;
consequently increasing force production as more fibers activate during work. A
caveat to the aforementioned is when programming hypertrophy-based training, as
the strength coach must be cognizant of morphological considerations for the sport.
Athletes carrying additional mass tend to be slower and require more oxygen for the
same task when compared with a lighter counterpart. Training for muscle mass
should not negate ballistic and/or athletic-based training nor affect metabolic
conditioning.
It is evident that phases of training should be balanced and present varied
methods of stress for desired results. It should be understood however that multiple
variables can be applied simultaneously. This suggests the use of appropriate velocities
throughout all stages of a periodized training program; including the hypertrophy
phase when more emphasis is usually placed on motor unit recruitment. When
constructing training programs using multiple adaptation goals per exercise bout,
operations should be ordered based on the energy system-tension relationship.
Since most programs are aimed at multiple objectives the order will reflect the energy
system first, and then the speed of the movement second. Therefore phosphagen-
driven power exercises (<5 reps) precede phosphagen-based strength exercises (<5
reps), which precede phosphagen/glycolytic strength exercises (<10 reps). Essentially,
nerves are trained before muscle, both on the field and in the weight room; neural
adaptation training will precede metabolic conditioning.
Potential of Motor Unit Force Output – Program AttentionThere are two neurophysiological aspects of muscle fibers that may help promote the desired
training effect when attempting to develop power. The first is post- activation potentiation
(PAP). PAP describes the improvement in muscle performance (e.g., power and force) resulting
from a previous, high-intensity stimuli. Pre-activation elicits a PAP response when performed
within 12 minutes of the prior exercise. Essentially, the acute contractile history of motor units
can have a positive effect on future actions due to changes in neural excitation. These effects
are commonly targeted during training via the use of complex sets. A complex set describes a
set couple where a heavy resistance exercise (>85% 1RM) is followed by an explosive activity.
Numerous studies have concluded that complex sets, (sometimes referred to as contrast sets),
such as heavy squats followed by rapid jump squats, are effective for improving performance vari-
ables such as jump height through the benefits of PAP during the execution of the explosive
component. It is important to note that the load of the initial exercise is the key aspect in pro-
moting an optimal PAP response. The second consideration lies in the lengthening of muscle
tissue and its affect on motor unit activation. When muscle moves through a full ROM, twitch
force increases; consequently reducing the fatigability of the motor unit by lowering localized
force production. This occurs through an increased number of contractions [18]. In addition to
total muscle activation, full ROM contractions promote cross-fascial excitation and improve force
coupling potential. This identifies that the use of very heavy loads moved over a short duration
is not as beneficial as heavy loading across a full ROM. Unilateral exercises can optimally exploit
this benefit.
Figure 6.6 Proper Training Stress Order
DEFINITIONS
Post-activation potentiation –
Post-movement excitation in thenervous system which enhances powerand force output during performance ofa subsequent movement using the sametissue
Complex set –
Set couple where a heavy resistanceexercise (>85% 1RM) is followed by anexplosive activity (e.g., heavy squats andbox jumps)

200
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism
Practical Implications of Motor Unit Metabolism and PhysiologyProper exercise prescription for elite athletes should address three key elements connected
with the motor unit and metabolic adaptations including the total force produced, the maximal
velocity of movement, and the time-tension relationship. Due to the fact most sports require
repeat bursts of high-force and high-velocities; phosphagen-derived energy becomes the priority
system. Research has demonstrated that PCr recovery is slowed when the system is placed under
hypoxic conditions, and improved when oxygen is available during recovery. These findings
suggest two things: 1) optimal benefits within the PCr system occur with adequate recovery, and
2) athletes who develop efficient cardiovascular proficiency will experience higher neuromuscular
performance in the PCr system due to greater oxygen availability [19]. This concept was tested
with hockey, tennis, basketball, handball and football players using work intervals with varying
periods of recovery. Researchers found that interval training with short rest periods compromised
phosphagen-recovery and the resultant adaptations, whereas intervals with longer rest periods
demonstrated higher PCr efficiency and contributed to improved PCr synthesis rates after five
weeks of HIT[20]. These findings are relevant because these adaptations are necessary for repeating
high force in sports situations.
The evidence suggests if an athlete wants to effectively produce, reproduce, or sustain high
force they must comply with proper tension-time/recovery relationships during training. An
important concept to understand is recovery is dictated by the level of resistance and volume of
muscle employed to move it and not simply the time to complete the activity. For example, an
experienced athlete performing four Olympic cleans using 87.5%1RM will require two or more
minutes of recovery before repeating the set, whereas an athlete performing the same exercise
and repetition schematic using 65%1RM will need only 90 seconds of recovery to duplicate the
effort. The difference in recovery is associated with the depletion of PCr during the set. In the
case of competitive weightlifting, recovery from maximal capacity lifts are sometimes five minutes.
But in sports training recovery of this magnitude is unrealistic, and of little consequence, as
strength and conditioning focuses on improving sports performance not weightlifting.
HIT has become a popular technique to train multiple systems simultaneously. The issue
with HIT as an all-encompassing model is the short rest intervals cause hypoxia – which impedes
phosphagen system function. Therefore, even though HIT circuits may include lifts normally
associated with PCr adaptations (such as the Olympic lifts), the training does not provide the
athlete with the same benefits. Repeat power output associated with the PCr system is improved
among athletes who perform phosphagen-supported training using the correct work-rest
relationships. Olympic lifts and related PCr-driven activities should be employed independently
and not be used in circuit-based training. HIT training is designed for metabolic conditioning
using moderate-force applications. It is an ideal system of training to enhance the body’s ability
to deal with high-intensity, intermittent work associated with glycogen-supported actions.
Unlike the phosphagen system, training for improvements in glycolytic capacity does not
warrant long recovery periods for efficient adaptations. The recovery should reflect the metabolic
byproducts, which in the case of glycogen-supported activities is H+. Coaches should always
base the rest interval on the amount of ADP and H+ in the system after each training set. When
intensities drop below 80% of load capacity, ADP rephosphorylation becomes reduced; and
subsequently, so do the rest periods. Training the glycolytic system does not require exhausting
the system at terminal levels of time to be affective either. Typically, intervals of 10, 15, 30, 45 or

201
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
60 seconds are used with the shortest, tolerable rest period. The term tolerable reflects repeat -
ability; in the glycolytic system, recovery can be minimized to the point that allows for repeat
performance with proper movement efficiency at the desired velocity. If the technique or velocity
is compromised due to residual byproducts, the rest interval should be lengthened to accom -
modate improved movement quality. The glycolytic system can also be pushed for longer periods
of time (anaerobic capacity is generally 90 seconds), but it would have to reflect a defined purpose
in conditioning. The primary decisions around load, time, and recovery will be dependent on
either 1) improving the system’s efficiency with particular applications or 2) increasing the
athlete’s buffering capacity when using the system for sport activity. Once more, the rest period
is of key importance; greater increases in glycolytic enzymes and buffering have been measured
when high-intensity intervals are separated by appropriate recovery. Rest intervals that are too
short are far less beneficial.
The actions that occur in the anaerobic systems are often the event-deciding plays for most
sports, but aerobic capacity remains vital to intermittent sports such as soccer, basketball,
hockey and rugby. Repeat sprint ability and high-velocity actions are sustained by oxygen-
supported recovery. Likewise, sports that may be performed over prolonged periods of time,
such as tennis, require adequate aerobic efficiency to spare energy. This being said, aerobic
benefits should be derived from anaerobic training because the improvements in sports come
from the interaction of the systems – not simply elevated oxygen consumption. An important
concept here is aerobic metabolism, while integral from a support perspective, can be detrimental
to force if trained as an energy system preference. Coaches must carefully manage conditioning
so aerobic metabolism functions as an interactive system and does not compromise anaerobic
functions. Over-emphasizing aerobic training will cause deleterious effects on force and power
output due to recruitment and morphological changes in the motor units. Therefore, team sport
athletes should not be conditioned using long-duration, steady-state training. By training with
anaerobic intervals (shorter work bouts with higher rest periods), problems that affect force
production and power including motor unit recruitment, firing asynchronicity and negative
enzyme changes can be properly accounted for in the program. Using the glycolytic system to
promote aerobic conditioning requires higher-intensity activity with work-to-rest ratios of 1:1
to 1:6. The intensities achievable through anaerobic energy systems promote improvements in
stroke volume and capillary density as well as enzymatic adjustments that more closely mimic
sport conditions. This allows the athlete to transport oxygen into muscle cells more efficiently;
improving sprint recovery and longer-duration play at higher intensities.
The Balance of Physiological Disruption and RecoveryThe concept of training quality over quantity extends to pre-exercise
nutrition and post-exercise recovery. It is important to recognize that the
quality of an exercise bout is dependent on relative fluid and energy stores
in the body at the initiation of training. Performing while in a fatigued state
can have injurious effects and result in muscle damage if the intensity and
duration is too aggressive or prolonged. Exaggerated levels of strain are
often the result of accumulative stress from improper doses of loading, poor
management in the quantity and type of tension (eccentric vs. concentric
volume) and inadequate recovery. Athletes are at an elevated risk for system
Event-deciding plays during competition areoften anaerobically-driven, but a high aerobiccapacity remains vital to intermittent sports suchas soccer, basketball, hockey and rugby.
The combination of significant contractile forces, inadequate recovery, andmetabolic disruption can create a deleterious environment.

202
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism
strain and injury during higher volumes of training, such as those seen in the hypertrophy and
strength phases of periodized programs. These higher-volume phases often cause strain-
associated muscle fatigue. As stated earlier, “more is better” is a flawed concept when applied to
loading conditions, particularly when neuromuscular stress is acutely or chronically too high.
Special consideration should be applied to programs simultaneously employing plyometrics,
ballistics, resisted sprints, and heavy compound exercises. Coaches should plan bouts based on
recovery, and during longer training cycles, plan for periods of unloading.
Training volume is not the only consideration when analyzing the risk for muscle damage.
The collective stress associated with muscle tension and inadequate recovery becomes more of
an issue when combined with glycogen depletion, high levels of reactive oxygen species (ROS)
associated with hypoxic states, and inadequate nutrition (including calcium, sodium and
potassium intake). It is commonly multi-factorial stress that promotes or exacerbates muscle
damage. Therefore, exercise selection and volumes should be considerately applied and balanced
with recovery measures; including sound nutritional strategies. To this point glycogen-loading
and adequate hydration are collectively associated with reductions in tissue damage when training
volumes are matched. A checks and balance approach should be used to evaluate and edit
programs that have become overzealous in any particular area. Stress is often accumulative, so
coaches should evaluate all associated risks for overtraining (e.g., sleep quality, glycogen stores,
hydration, heart rate variability), rather than just the few considered to be the most detrimental
(i.e., high volume, eccentric work).
Training Threshold & Recovery
The idea of using thresholds as a guide for training and recovery is by no means novel, but
in some cases the nomenclature can create confusion in application. For this reason, thresholds
should be regarded from two points of view; one is conceptual and the other is operational. From
a conceptual perspective, there are two ventilatory thresholds (VT1 and VT2) and two metabolic
thresholds (lactate threshold; LT1 and LT2) [21]. While clear physiological boundaries or
“thresholds” are nearly impossible to determine; these thresholds represent the beginning and
the end of the aerobic-anaerobic transition. Essentially, it should be considered a metabolic shift,
as no metabolic pathway is ever completely shut down. Similarly to a car, the body can downshift
to gain power, and upshift to gain efficiency. Lower gears of a car use the most fuel to produce
the most force, and consequently produce the most exhaust. Similarly, the higher the intensity
experienced by the body the more metabolic exhaust (lactate). The concentration of metabolic
“exhaust” triggers a corresponding response to maintain homeostasis. These metabolic changes
in the system reflect the lactate thresholds, whereas the ventilatory responses to those conditions
represent the ventilatory thresholds. VT1 is demonstrative of increased ventilation to mediate
lactate in a process of buffering. VT2 is characterized by a loss of lactate steady-state resulting in
lactate build-up, metabolic acidosis and a disproportional rise in ventilation.
Key elements of chronic muscle strain:
Excess loading without adequate recovery
Repeated isolated loading
High volume eccentric contractions
Excessive hip-trunk extension
Poor lifting technique – load over quality movement
Overall high volume of physical activity
Lack of proper nutrition and hydration
Inadequate sleep
DEFINITIONS
Reactive oxygen species –
Chemically-reactive moleculescontaining oxygen ions and peroxides;are a natural byproduct of oxygenmetabolism and serve important roles in cell signaling and homeostasis
DEFINITIONS
Lactate threshold –
Describes a work intensity where therate of lactate accumulation matcheslactate removal; any further increasewill cause excessive lactate accumu -lation and force the participant toreduce the workload or stop within agiven period of time due to acuteacidosis

203
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
The concept of anaerobic lactic capacity is more accurate than lactate threshold when used
for program design. When considering lactate metabolism, glycolysis is not very efficient and
results in greater H+ production than when sugar is used for aerobic metabolism. Glycolysis allows
for higher-intensity work, but at the same time creates increased quantities of byproduct. As a
result, the entire ATP production process slows down, acidity increases and muscles fatigue.
Previous text has identified the inability of enzymes to function in acidic environments. Therefore,
unmanaged byproducts like H+ limits enzyme activity to a point that promotes muscle fatigue, or
forced volitional failure. An individual in better metabolic shape, which combines all energy
systems, is able to produce more force for longer periods of time and repeat the actions with limited
rest. This represents the lay concept of being “in shape”. The unification of more efficient energy
systems reduces deleterious metabolic products in the environment, spares energy, and increases
repeated force capabilities. Therefore, repeated force is just as much metabolic as it is neuro -
muscular in nature; supporting a balance within strength and conditioning methodology.
Metabolic Conditioning
If an athlete needs to function at higher blood lactate levels for improved performance
metabolic conditioning must: 1) impart a stress that increases lactate production to higher levels
and 2) control rest intervals to force improvements in buffering capacity from appropriate levels
of exposure. In terms of training, this can be done in both the weight room and on the field. In
the weight room, anaerobic capacity is usually pushed at an intensity range of 50-70%1RM using
12-25 reps. The volume is high at 30 to 45 sets in a training session, with short rest intervals of
up to 30 seconds. Sprinting, rowing or cycling at maximum capacity for 30 seconds to 1 minute
has also been shown to increase anaerobic lactic capacity. Since Type IIA and IIX fibers produce
more lactate, higher activation of these fibers will result in enhanced adaptations. This is
accomplished, by maximizing movement velocity over specific distances. Select track distances,
weighted locomotion drills (including stair work), loaded/unloaded ballistic work, and novel
activities such as battle rope, kettlebell or sledgehammer drills can serve this purpose. The use
of supersets, tri-sets, metabolic circuits and on-field stations function well for improving
anaerobic capacity as long as the rest interval is appropriately short. Due to the high fatigue rate,
selected exercises should be uncomplicated to promote technique adherence. For obvious
reasons, Olympic lifts should not be used for these purposes. When speed, agility, and quickness
(SAQ) activities are used for conditioning, neural work involving finite change of direction (COD)
drills or agilities should precede gross COD/agilities to reflect fatigue-based technique adherence.
When employing on-field/court conditioning, multidirectional drills should be used over linear
drills as they enhance lactate production over a broader range of musculature; further promoting
fatigue consistent with sport applications. Changes in the center of mass (directionally and
vertically) cause dramatic increases in the activity of postural muscles and global stabilizers to
create a much better overall conditioning approach compared to linear running alone.
Types of FatigueFatigue is often defined as a general reduction in motor unit force output, but has impli -
cations in the psycho-emotional state of an athlete as well. This suggests that fatigue is not simply
metabolic in nature but also impacts the nervous system. Fatigue cannot be viewed as a general
concept in athletics as it is sport specific; factors related to successful competitive performance
are different for every sport and impart different issues during competition. For example, aFatigue is not simply metabolic in nature but alsoimpacts the nervous system.

204
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism
football coach may be concerned with decreased muscular force and power in the defensive line
during the fourth quarter of a game, whereas a golfer might be more negatively affected by a
decline in technique and finite skill performance at the closing round of a three-day tournament.
When addressing fatigue, it is important to understand the root cause of the problem, which
suggests an analysis of both metabolic and neural factors. The neuromuscular junction (NMJ)
is the physiological formation that allows the electrical signal (action potential) from the nerve
to innervate the muscle cell via a chemical conversion. Central (neurological) fatigue refers to
the reduction in all actions that occur on the neuron side of the NMJ, while peripheral fatigue
relates to those that impact the muscle contractile function.
Intense training and inadequate recovery can negatively impact the nervous side of the
contractile system. This combination can reduce brain motor signaling due to poor neuro trans -
mitter function and immune cell activity from high-stress exposure. Brain activity has also been
shown to be affected by ammonia accumulation from protein breakdown in the liver when sugar
supply is inadequate. These factors fatigue the CNS; affecting both the generation of action
potentials as well as mental sharpness during training/competition. At the muscle cell level the
NMJ also experiences fatigue. This can cause a slowdown in electrical-chemical transmission;
impeding force proficiency. Likewise, acute changes associated with high-tension training occur
at the NMJ; further slowing the contraction rate. From a practical sense however, it is important
to recognize central fatigue is most often associated with total-body depletion of carbohydrates
(glycogen) common of intense training without adequate nutritional support [22].
Peripheral fatigue, or muscle fatigue, refers to aspects that affect the metabolic properties
of tissue, and consequently the function of actin and myosin in the sarcomere. These include
reduced energy availability from immediate and intermediate sources, energy production
inhibition from byproduct accumulation, and muscle contraction limitations due to acidosis and
dehydration. When the cell experiences acute dysfunction due to pH changes that cause acidosis,
it is termed acute peripheral fatigue. The word acute is used because the environment can be
buffered to re-initiate work capacity during the training bout. When the fatigue occurs due to
lack of energy provisions in the cell, from either low pre-exercise stores or localized depletion of
glycogen in the muscle from ongoing work, it is termed general peripheral fatigue. General
peripheral fatigue does not improve with an acute rest interval because sugar is no longer available
to support the force demands. At this point, ongoing training could be viewed as detrimental
and therefore the exercise bout should be ended.
The key factors that need to be collectively controlled to prevent or delay fatigue are the
initial carbohydrate storage, the intensity and duration of activity, the recruitment patterns
involved in the movements and the work-to-rest ratio. Each of these factors is controllable and
therefore should be accounted for in the program. Coaches should ensure that athletes are 1)
adequately fueled and hydrated, 2) adequately exposed to metabolic-specific stress, and 3)
perform sport-specific movements at the intensity and durations associated with the sport.
These have collectively demonstrated athletic improvements in reaction time, agility and repeat
sprint endurance throughout the duration of competition. By suppressing peripheral fatigue,
athletes perform sharper movements and experience a reduced risk for injury in the final quarter
or period of a game. Additionally, athletes who are also exposed to concurrent heavy compound
lifts and ballistic training are better able to repeat high-level contractile force when compared
to athletes who have not experienced the same preparatory stress. Appropriate balance of sport-
specific power and conditioning work aimed at exploiting the PCr and glycolytic systems will
DEFINITIONS
Acute peripheral fatigue –
Occurs when cells experiencedysfunction due to a metabolicreduction in pH; acid limits enzymeactivity requiring buffering compoundsbefore work can be re-initiated
General peripheral fatigue –
Occurs with a lack of energy in the celldue to low pre-exercise stores orlocalized depletion of glycogen; acuterest intervals will not help as sugar is nolonger available

205
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
allow the body to endure higher tension for a longer period of time. When the environmental
stress is familiar, albeit internal or external, it is much easier for the body to deal with in a
successful manner.
Muscle DamageMuscle damage occurs as a result of excess physical stress placed on muscle fibers and
metabolic stress experienced during training and competition. As mentioned earlier, the
meta bolic contributions to muscle damage include insufficient energy storage, inefficient manage -
ment of byproducts due to inadequate recovery, depleted provisional states, and premature
exposure to progressive training volumes. The end result of each is exercise-induced muscle
weakness and damage. An added concern is insufficient fluid balance to maintain the metabolic
environment as cells cannot function without proper cellular fluid volume. The combination of
problems leading to muscle damage are most commonly associated with low oxygen levels, low
carbohydrate availability and insufficient metabolic water (electrolyte balance); all of which are
controllable factors.
Metabolic disruptions leading to muscle damage during exercise are associated with a
number of compounding factors. However, most problems seem to be tied to inadequate oxygen
availability from intense conditions. As mentioned previously, exercise creates hypoxic conditions
as a result of reduced oxygen supply to tissues. This leads to the proliferation of ROS, or free
radicals, as detailed earlier. These chemically-reactive molecules are a natural byproduct of
oxygen metabolism and serve important roles in cell signaling and homeostasis. ROS also play
an integral role in the inflammatory process leading to muscle adaptations, and their presence
promotes positive responses in skeletal muscle healing when training and recovery is properly
matched. However, during times of very high stress, such as heavy training in high heat, ROS
levels can increase dramatically; resulting in significant damage to cellular structures. Research
has shown that ROS production is associated with both intense aerobic and anaerobic work. In
fact, ROS have been directly associated with excessive muscle damage after resistance exercise [23].
Hypoxia and ROS have a direct effect on all metabolic systems; however, two mechanisms
seem to be most important. The first surrounds the fact that exercise during acute hypoxia
increases muscle and blood lactate levels more than during exercise with normal oxygen supply.
This means that hypoxia causes a greater than normal decrease in pH; thereby increasing potential
muscle damage. The second mechanism involves enzymes, as oxygen deprivation can cause
a number of enzymes to become less efficient. Glycogen phosphorylase, the enzyme which
promotes glycogenolysis in working muscle, has been shown to slow down in strained environ -
ments. Creatine kinase activity has also been found to be altered. In fact, rhabdomyolysis, or
the breakdown of injured muscle tissue and the release of myoglobin into circulation, is usually
diagnosed through creatine kinase serum levels.
It has been established that requisite levels of glycolytic conditioning is necessary to manage
hypoxic conditions and improve buffering during intense training. However, adaptations take
time and should be progressive. If athletes are exposed to excessive exercise durations or inten -
sities without adequate recovery in attempts to increase adaptation rates, they may be injured or
experience acute rhabdomyolysis. This is also true in trained athletes, even if temperature and
hydration are accounted for during the training. Although numerous documented cases indicate
hydration levels and temperature as key factors, this is not always the case with rhabdomyolysis.
Metabolic disruptions leading to muscle damageduring exercise are associated with a number ofcompounding factors. However, most problemsseem to be tied to inadequate oxygen availability.
DEFINITIONS
Rhabdomyolysis –
A potentially life-threatening issue;involves the breakdown of injuredmuscle and the release of myoglobin intocirculation following extreme metabolicand mechanical stress

206
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism
For instance, a healthy, well-trained 18-year-old NCAA Division 1-A football player was diagnosed
with rhabdomyolysis after executing a training program that included 10 sets of 30 squats and
Romanian deadlifts with one-minute rest periods, using two 40 lbs dumbbells [24]. This case was
important because it showed that rhabdomyolysis is not only a risk with excessive heat and
dehydration, but also aggressive training plans. Empirical evidence also implicates excessive
eccentric loading and high-volume training without physical readiness as risk factors for
rhabdomyolysis.
Role of Anabolic HormonesFundamentally, hormones are chemical communicators that either cause or inhibit cellular
actions. These biochemical regulators are released from specialized glands in order to change the
action of cells and tissues for growth, maintenance and repair, or function to maintain metabolic
homeostasis. When considered for training purposes, the anabolic hormones including growth
hormone (hGH), testosterone and insulin-like growth factor (IGF-1) as well as the adrenal
hormones epinephrine and cortisol warrant attention. The type of program employed deter -
mines the associated response, which is generally tied to the adaptation. This occurs because
each hormone has an affinity for a particular type of stress. For instance, when oxygen is needed
adrenal hormones trigger an increase in heart excitation to increase supply, whereas when sugar
regulation becomes relevant hGH plays a greater role.
During exercise, hormones function as messengers that provide a specific directive. Chem -
ical messages are sent to specific receptors in tissue to provide information for an action. These
messages can be clear and well-received, or incomplete; consequently impacting the effectiveness
of the response. The endocrine system has the ability to adapt to chronic stress; suggesting
training can cause both an improvement in the signaling and message, as well as the tissue’s
affinity for the message and response. This partially explains differences between novice and
experienced athletes performing the same program.
In the case of anabolic hormones, most studies show adaptations are greatest when training
requires the use of more muscle mass and a high number of motor units. The total mass recruited
for these types of exercises promotes increased concentration of circulating hormone, which has
an impact on the whole body. When experienced weightlifters performed unaccus tomed leg
training exercises in addition to their normal training in a given study, they experienced a 27%
greater increase in their biceps brachii strength compared to those who continued to only train
their upper body [25]. The increased activation of total mass and greater loading associated with
lower body, compound exercises improves the overall hormonal response and adaptations across
the body. This outcome is demonstrative of the relationship of mass and load on circulating
anabolic hormones as well as the adaptation effects of hormone on trained tissue.
Growth Hormone (hGH)
Growth hormone is highly regarded for sport metabolism and has been used as an ergogenic
aid by both athletes and non-athletes alike. Somatotropin, or hGH, is vital as it initiates growth
in body tissues, increases cell size and potentiates mitosis (cell division). The regenerative func-
tions of the hormone are also imperative to recovery from fatigue and injuries. Additionally,
growth hormone is involved in metabolic functions and energy provision; suggesting it is both
an anabolic and metabolic hormone.
DEFINITIONS
Insulin-like growth factor 1 (IGF-1) –
Currently considered to be a central“signaling” hormone that initiatesmuscle growth following resistanceexercise; it can also directly signal cellsto reproduce and grow

207
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
Growth hormone is involved to some degree during most exercise regimens, but is especially
partial to higher-stress environments. Maximal strength training programs (PCr/glycolytic) con-
ducted at 75-95% 1RM, hypertrophy training programs (glycolytic) at 70-85% 1RM, and power
training either at 30-50% 1RM (glycolytic) or 60-95% 1RM (PCr) have all showed positive effects
on hGH activation when rest intervals are appropriately applied. In addition, it appears that
muscular endurance at 50-70% 1RM has minimal but positive effects on the production of
growth hormone when short rest intervals are used. In sports that benefit from additional mus-
cle mass, hGH is relevant but its response is very specialized. Growth hormone serves as a trigger
for important liver hormones, particularly IGF-1, when rep-load-recovery time is properly bal-
anced. The stress causes a reduction in blood pH and high lactate levels to stimulate cortisol and
epinephrine release from the adrenal glands. A humoral (hormone to hormone) pathway with
hGH relays signaling to increase anabolic activity from the liver which causes the release of IGF-
1. The ideal recipe seems to be 8-12 repetitions, using 75-80% 1RM, with 30-60 second rest
intervals within a high-volume training program. Bodybuilders will also employ these tactics
with muscle isolation to further fuel localized muscle recruitment and inflammation for increases
in size.
Testosterone
Testosterone serves several functions at the genetic level among males and is produced in
lesser quantities in females. In addition to sexual functions and development, testosterone is
intimately tied to specific utilities in the tissue and overall health in males. In females, testosterone
serves as a homeostatic hormone in balance with estrogen. Testosterone is most cited for its
impressive affects on growth and metabolic enhancements; stimulating increased mass with
subsequent reductions in fat. Individuals that supplement testosterone experience muscle growth,
improvements in body composition, and enhanced recovery rates. Natural methods to increase
testosterone levels seem to occur in response to large muscle activation with high loads, as both
the back squat and deadlift yield desirable responses in blood concentration during bouts of heavy
strength training. Testosterone release is also consistent with the aforementioned training stress
that stimulates hGH. While women generally demonstrate about 1/10th the concentration of the
average male, positive testosterone responses following resistance exercise has been identified.
A study examining 47 women, with an average age of 22 years, showed that total testosterone
increased by 25% following 6 sets of 10 maximal effort squats separated by a 2-minute rest
period [26]. Testosterone levels have also been associated with performance variations in master’s
level athletes. Expectedly, athletes with higher testosterone levels outperformed their age-
equivalent counterparts. Due to reductions in the testosterone precursor DHEA during
andro pause, male athletes over 40 require modifications in training to promote higher circulating
concentrations.
Insulin-like Growth Factor (IGF-1)
IGF-1 is currently considered to be a central “signaling” hormone that initiates muscle
growth following resistance exercise. The term “signaling” refers to IGF’s capacity to generate
action from other molecules that ultimately result in the stimulation of muscle protein synthesis.
It can also directly signal cells to reproduce and grow. Due to the apparent role of IGF-I up-
regulation in muscle remodeling, IGF-1 mRNA (or mechano-growth factor (MGF)), is considered
a key element in mass gains. MGF is almost exclusively released from trained tissue which may
explain why mass gains in training-tenured athletes are greater than in novice athletes. Therefore,
Natural methods to increase testosterone levelsseem to occur in response to large muscleactivation with high loads as both the back squatand deadlift yield impressive responses in bloodconcentration associated with heavy strengthtraining.

208
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism
it appears that muscle growth depends on the presence of MGF. IGF-1 is clearly elicited by
hypertrophy-type training and is fairly exclusive to the glycolytic system. As mentioned pre -
viously, multiple-sets of moderate-high intensity with shortened rest intervals seems to be most
effective technique. The goal is to appropriately push the glycolytic pathway to its tension-time
limit. The rest period selected is very important for hormonal activation because it is the
metabolic byproducts that ultimately trigger the cascade of events leading to IGF-1 release. This
underscores the importance of tracking rest intervals with a stop watch. High-volume training
with rest periods of 30-60 seconds promotes the desired blood chemistry to trigger the signaling
response needed to stimulate anabolic hormones. Higher loads with longer rest intervals do not
create the same concentration of IGF-1 and consequently yield reduced signaling for mass gains.

209
NCSF Advanced Concepts of Strength & Conditioning Chapter 6
Sport Metabolism
REFERENCES:
1. Hirvonen, J., Rehunen, S., Rusko, H., & Härkönen, M. (1987). Breakdown of high-energy
phosphate compounds and lactate accumulation during short supramaximal exercise.
European Journal of Applied Physiology and Occupational Physiology, 56(3), 253-259.
2. Sahlin, K., Soderlund, K., Tonkonogi, M. I. K. A. E. L., & Hirakoba, K. (1997).
Phosphocreatine content in single fibers of human muscle after sustained submaximal
exercise. American Journal of Physiology-Cell Physiology, 273(1), C172-C178.
3. K. Van Someron, “The physiology of anaerobic training,” in The Physiology of Training,
G. Whyte, Ed., pp. 85–115, Elsevier, Oxford, UK, 2006.
4. Forbes, S. C., Slade, J. M., & Meyer, R. A. (2008). Short-term high-intensity interval
training improves phosphocreatine recovery kinetics following moderate-intensity
exercise in humans. Applied Physiology, Nutrition, and Metabolism, 33(6), 1124-1131.
5. Wong, P. L., Chaouachi, A., Chamari, K., Dellal, A., & Wisloff, U. (2010). Effect of pre-
season concurrent muscular strength and high-intensity interval training in professional
soccer players. The Journal of Strength & Conditioning Research, 24(3), 653-660.
6. Scates, A. E., Linn, M., & Kowalick, V. (2003). Complete Conditioning for Volleyball.
Human Kinetics.
7. Wells, G. D., Selvadurai, H., & Tein, I. (2009). Bioenergetic provision of energy for
muscular activity. Paediatric Respiratory Reviews, 10(3), 83-90
8. Norrbrand, L., Fluckey, J. D., Pozzo, M., & Tesch, P. A. (2008). Resistance training using
eccentric overload induces early adaptations in skeletal muscle size. European Journal of
Applied Physiology, 102(3), 271-281.
9. Harber, M. P., Gallagher, P. M., Trautmann, J., & Trappe, S. W. (2002). Myosin heavy chain
composition of single muscle fibers in male distance runners. International Journal of
Sports Medicine, 23(7), 484-488.
10. Behm, D. G. (1995). Neuromuscular implications and applications of resistance training.
The Journal of Strength & Conditioning Research, 9(4), 264-274.
11. Kamen, G., & Knight, C. A. (2004). Training-related adaptations in motor unit discharge
rate in young and older adults. The Journals of Gerontology Series A: Biological Sciences
and Medical Sciences, 59(12), 1334-1338.
12. Patten, C., Kamen, G., & Rowland, D. M. (2001). Adaptations in maximal motor unit
discharge rate to strength training in young and older adults. Muscle & Nerve, 24(4), 542-
550.
13. Van Cutsem, M., Duchateau, J., & Hainaut, K. (1998). Changes in single motor unit
behaviour contribute to the increase in contraction speed after dynamic training in
humans. The Journal of Physiology, 513(1), 295-305.
14. Saplinskas, J. S., Chobotas, M. A., & Yashchaninas, I. I. (1980). The time of completed
motor acts and impulse activity of single motor units according to the training level and
sport specialization of tested persons. Electromyography and Clinical Neurophysiology,
20(6), 529.

210
Chapter 6 NCSF Advanced Concepts of Strength & Conditioning
Sport Metabolism
15. Aagaard, P., Simonsen, E. B., Andersen, J. L., Magnusson, P., & Dyhre-Poulsen, P. (2002).
Increased rate of force development and neural drive of human skeletal muscle following
resistance training. Journal of Applied Physiology, 93(4), 1318-1326.
16. Newton, R. U., Kraemer, W. J., & Haekkinen, K. E. I. J. O. (1999). Effects of ballistic
training on preseason preparation of elite volleyball players. Medicine and Science in
Sports and Exercise, 31, 323-330.
17. Burd, N. A., Andrews, R. J., West, D. W., Little, J. P., Cochran, A. J., Hector, A. J., ... &
Phillips, S. M. (2012). Muscle time under tension during resistance exercise stimulates
differential muscle protein sub-fractional synthetic responses in men. The Journal of
Physiology, 590(2), 351-362.
18. Suzuki, S., Hayami, A., Suzuki, M., Watanabe, S., & Hutton, R. S. (1990). Reductions in
recruitment force thresholds in human single motor units by successive voluntary
contractions. Experimental Brain Research, 82(1), 227-230.
19. Haseler, L. J., Hogan, M. C., & Richardson, R. S. (1999). Skeletal muscle phosphocreatine
recovery in exercise-trained humans is dependent on O2availability. Journal of Applied
Physiology, 86(6), 2013-2018.
20. Bishop, D., Edge, J., Thomas, C., & Mercier, J. (2008). Effects of high-intensity training on
muscle lactate transporters and postexercise recovery of muscle lactate and hydrogen ions
in women. American Journal of Physiology-Regulatory, Integrative and Comparative
Physiology, 295(6), R1991-R1998.
21. Westhoff, M., Rühle, K. H., Greiwing, A., Schomaker, R., Eschenbacher, H., Siepmann, M.,
& Lehnigk, B. (2013). Ventilatorische und metabolische (Laktat-) Schwellen. DMW
Deutsche Medizinische Wochenschrift, 138(06), 275-280.
22. Davis, J. M., & Bailey, S. P. (1997). Possible mechanisms of central nervous system fatigue
during exercise. Medicine and Science in Sports and Exercise, 29(1), 45-57.
23. Uchiyama, S., Tsukamoto, H., Yoshimura, S., & Tamaki, T. (2006). Relationship between
oxidative stress in muscle tissue and weight-lifting-induced muscle damage. Pflügers
Archiv, 452(1), 109-116.
24. Moeckel-Cole, S. A., & Clarkson, P. M. (2009). Rhabdomyolysis in a collegiate football
player. The Journal of Strength & Conditioning Research, 23(4), 1055-1059.
25. Hansen, E. A., Andersen, J. L., Nielsen, J. S., & Sjøgaard, G. (2002). Muscle fibre type,
efficiency, and mechanical optima affect freely chosen pedal rate during cycling. Acta
Physiologica Scandinavica, 176(3), 185-194.
26. Nindl, B. C., Kraemer, W. J., Gotshalk, L. A., Marx, J. O., Volek, J. S., Bush, F. A., ... & Fleck,
S. J. (2001). Testosterone responses after resistance exercise in women: influence of
regional fat distribution. International Journal of Sport Nutrition and Exercise Metabolism,
11(4), 451.





























