Embed Size (px)
Citation preview

Spinal Projections of the Cat Parvicellular Red Nucleus
MILTON PONG, KRIS M. HORN, AND ALAN R. GIBSONDivision of Neurobiology, Barrow Neurological Institute, St. Joseph’s Hospital and Medical Center, Phoenix, Arizona 85013
Received 29 December 2000; accepted in final form 14 August 2001
Pong, Milton, Kris M. Horn, and Alan R. Gibson. Spinal projec-tions of the cat parvicellular red nucleus. J Neurophysiol 87: 453–468,2002; 10.1152/jn.00950.2000. Traditionally, the red nucleus of the catis divided into two parts: a large-celled, magnocellular, division(RNm) and a small-celled, parvicellular, division (RNp). The RNmprojects to the spinal cord and receives input from the cerebellarinterpositus nucleus. The RNp projects to the inferior olive andreceives input from the cerebellar dentate nucleus. In this report, wereexamine the connections of the red nucleus using the bidirectionaltracer wheat germ agglutinin-horseradish peroxidase (WGA-HRP).Our findings demonstrate that the cat RNp has a large caudal andlateral region that projects to contralateral spinal cord and not to theinferior olive. The spinally projecting region of RNp receives inputfrom the dentate nucleus and a lateral segment of anterior interpositus.Cervical projections from the red nucleus show a topography with therostral portion of RNp favoring upper segments and the caudal portionof RNm favoring lower segments. The results show that dentateoutput can influence spinal activity without passing through the ce-rebral cortex. For the control of movements such as reaching andgrasping, we suggest that RNp and dentate focus on the control ofproximal limb musculature, whereas RNm and the anterior interposi-tus focus on the control of distal limb musculature. We also suggestthat other species are likely to have a small-celled area of red nucleusprojecting to the spinal cord.
I N T R O D U C T I O N
Lesion or inactivation of the lateral cerebellum or its outputnucleus, dentate, impairs the ability to make accurate well-coordinated limb movements (Bastian and Thach 1995; Thachet al. 1992). It is generally assumed that the output of lateralcerebellum exerts its influence on limb movements by modi-fying activity in motor cortex via thalamic connections. How-ever, lesion of the cerebellar receiving area of the thalamus(Bastian and Thach 1995; Fabre-Thorpe and Levesque 1991;Ranish and Soechting 1976) has a relatively minor effect onlimb movements in comparison with cerebellar lesion. There-fore it is likely that lateral cerebellum can influence spinalactivity via brain stem connections, and physiological studiessuggest that this is the case (Hames et al. 1981). One of themajor brain stem targets of the dentate nucleus (DN) of the catis the parvicellular red nucleus (RNp). In this study, we reex-amine the connections of RNp to determine whether these cellscould provide an anatomical basis for the influence of lateralcerebellum on limb movements.
The mammalian red nucleus (RN) is typically divided into acaudal magnocellular region (RNm) and a rostral parvicellular
region (RNp). It is generally assumed that these cytoarchitec-tonic divisions differ also in connectivity. RNm receives inputfrom the cerebellar interpositus nucleus and projects to thecontralateral spinal cord, whereas RNp receives input from thecerebellar dentate nucleus and projects to the ipsilateral inferiorolive (Massion 1967). However, in the cat there is no clearseparation between large and small cells in the RN, and at leastsome small cells project to the contralateral spinal cord (Hol-stege and Tan 1988; Mussen 1927; Pompeiano and Brodal1957). Could the spinally projecting cells in RNp connect DNoutput to the spinal cord rather than to the inferior olive? In thisstudy, we address this question by placing injections of wheatgerm agglutinin-horseradish peroxidase (WGA-HRP), a bidi-rectional tracer, into physiologically identified regions of thecerebello-rubro-spinal pathway.
Our results demonstrate that most of the cat RNp projects tothe contralateral spinal cord. The spinally projecting cells arelargely, if not entirely, separate from cells projecting to theipsilateral inferior olive. Regions of RNp that project to thecontralateral spinal cord receive input from the contralateralcerebellar dentate and a lateral segment of anterior interpositus.For projections to cervical cord, RNp favors upper segments,whereas RNm favors lower segments.
The data do not support the prevalent view of RN connec-tivity and indicate that the lateral cerebellum can influencelimb movements via spinal connections through RNp. Theconnections of RNp and its physiological actions (see Horn etal. 2002) suggest that the pathway influences upper limb mus-culature more strongly than distal limb musculature. Dentatemay play a role in directing and stabilizing the limb duringreaching and grasping.
M E T H O D S
Fourteen male cats (3.5–4.5 kg) were used to trace connections ofthe red nucleus. Anesthesia consisted of an initial intramuscularinjection of ketamine hydrochloride (8 mg/kg) followed by intrave-nous doses (10 mg) of pentobarbital sodium. Cats were fastened intoa stereotaxic frame, and a craniotomy and/or laminectomy was per-formed to provide access to brain stem and spinal injection sites. Afterthe tracer injections, wounds were sutured, and the cats were placedon a heating pad. Anesthesia was maintained with iv infusion ofpentobarbital sodium (2 mg � kg�1 � h�1) for the duration of thetransport period (approximately 48 h). During the transport period,breathing and temperature were monitored, and the airway wascleared frequently with suction. Prior to perfusion, cats were admin-istered lethal doses of pentobarbital sodium (�250 mg iv as a rapid
Address for reprint requests: M. Pong, BNI-Neurobiology, St. Joseph’sHospital and Medical Center, 350 W. Thomas Rd., Phoenix, AZ 85013(E-mail: [email protected]).
The costs of publication of this article were defrayed in part by the paymentof page charges. The article must therefore be hereby marked ‘‘advertisement’’in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
J Neurophysiol87: 453–468, 2002; 10.1152/jn.00950.2000.
4530022-3077/02 $5.00 Copyright © 2002 The American Physiological Societywww.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

bolus). All procedures were approved by the St. Joseph’s HospitalInstitutional Animal Care and Use Committee and were in accordancewith National Institutes of Health guidelines.
Small pressure injections (0.008–0.032 �l) of WGA-HRP (Sigma)were made at physiologically identified sites. Injection sites wereinitially identified using tungsten microelectrodes. The injection pi-pettes and the microelectrodes were cross-referenced by using thesame optical zero point. Accuracy of final placement was confirmedby recording with the injection pipettes. Table 1 summarizes theinjection site or sites for each case.
Perfusion consisted of a saline rinse followed by two liters offreshly made paraformaldehyde (3–4%, depending on the case) and aseries of 10, 20, and 30% sucrose/phosphate buffer solutions (0.1 M,pH 7.4). Brains were sectioned at 50 �m in either the parasagittal orfrontal planes. Sections were processed with a modified tetramethylbenzidine (TMB) reaction (Gibson et al. 1984; Mesulam 1982) andthen stained with thionin.
The locations of anterogradely labeled terminals and fibers andretrogradely labeled cell bodies were plotted onto high-resolutiondigital images using a computerized plotting system (Image Tracer,Translational Technology). The plotting system registered microscopestage position (�1-�m resolution) to the digital image for accuratemarking of label location. All plotting was performed under high-powered (�100) observation using polarized light illumination.
Overlap between the label from two different cases was assessed byfirst creating computer images of the sections with the plotted label.Corresponding sections from the two cases were scaled to the samesize and manually overplotted with the aid of a computer drawingprogram (Canvas, Deneba). When the location of label from one casefell within 100 �m of label from the other case, the area was markedas a site of overlap. For the red nucleus, the location of the caudal andventral large cells provided the most reliable alignment between cases.Stereotaxic coordinates were assigned based on comparison of thesections with the cat atlas of Berman (1968).
The paraformaldehyde fixation produced severe and variable tissueshrinkage, so our estimates of cell sizes within RN are not comparableto those made by previous investigators. However, comparisons be-tween RN areas within a subject are still meaningful. To measure thesize of spinally projecting cells, high-power brightfield microscopicimages were made of selected regions within the nucleus. UsingImage Tracer, we then marked the retrogradely labeled cells in theimage under darkfield polarized light illumination. The width andlength of marked cell somas were then measured from the brightfieldimage. Measurements were orthogonal to each other and made at thepoints of maximal cell dimensions. When the section appeared toinclude the apical dendrite, an estimate of soma size was extrapolatedfrom cell-wall curvature. Attempts were made to measure every
labeled cell, but in some instances, cell membranes could not bevisualized. The product of each pair of measurements was used as anestimate of soma area.
R E S U L T S
Projections to the cervical cord
RED NUCLEUS LABELING FROM SPINAL INJECTIONS. Our firstobjective was to identify regions of the red nucleus that projectto the cervical spinal cord. A single rubrospinal fiber canterminate along several segments of the cord (Shinoda et al.1977), so we maximized the number of labeled rubrospinalcells by making closely spaced injections across several seg-ments. To simplify the study, we examined projections only tocervical cord. A series of WGA-HRP injections were madeinto the intermediate spinal gray every 2–3 mm from C3 to C6on the right side of the cord (case CN10, Table 1). Recordingsthrough the injection pipette helped distinguish cells in theintermediate laminae from the large motoneurons in the ventralhorn.
The frontal sections through the left RN in Fig. 1 werephotographed with a combination of semi-polarized darkfieldand transmitted light to visualize both labeled and unlabeledcells. At the caudal RN pole (Fig. 1, A3.8), the labeled cells areconfined to approximately the dorsal half of the nucleus. Spinalprojections of the RN have a well-recognized topography (Hol-stege and Tan 1988; Pompeiano and Brodal 1957; Robinson etal. 1987), and the unlabeled cells in the ventral half of thenucleus project to lower levels of the cord. Although thelabeled cells vary in size, the high percentage of large cellsidentifies this level as the RNm.
Rostrally (Fig. 1, A4.6), two groups of labeled cells areapparent. Lying medially and consisting mainly of large cells,one group appears to be a rostral continuation of RNm. Adifferent group of labeled cells can be seen lateral to RNm.Although this group contains cells of various sizes, the cellsappear to be smaller than those of the medial group.
Further rostrally (Fig. 1, A5.2) there is no clear RNm, but thesmaller celled lateral group is prominent. In this report, weconsider the rostral lateral group of cells as part of RNp, whichis consistent with their designation in the Berman (1968) catatlas.
Measurements of cell areas within the groups (see METHODS)supported our visual impressions. We compared the cells inFig. 1. Cells in RNp (lateral group) at A5.2 and A4.6 hadmedian soma areas of 275 and 273 �m2, respectively. Cells inRNm at A4.6 (medial group) and A3.8 (all cells) had mediansoma areas of 420 and 418 �m2, respectively. A one-wayANOVA between groups (Kruskal-Wallis ANOVA on ranks)indicated significant differences (P � 0.05), so we ran pairwisecomparisons (Dunn’s method) between the groups. Compari-sons between measurements of RNm and RNp cells weresignificantly different (P � 0.05), whereas comparisons withineither RNm or RNp were not significantly different (P � 0.05).
A complete representation of the retrograde labeling fromA3.1 to A6.9 for case CN10 is shown in Fig. 2, left. Thesmaller-celled lateral group becomes prominent at about A4.5and continues to A5.9. The medial RNm group extends fromthe caudal end of the nucleus (A3.1) to approximately A5.4.The spinal projection from both groups of cells is almostentirely contralateral. An outline of the labeled RN cells on the
TABLE 1. Injection sites and volumes
Cat Injection Site(s)WGA-HRP (1%) Injected,
�l
BRN1 RNp (left) and RNm (right) 0.008 (RNp) 0.008 (RNm)BRN3 RNp (left) 0.004 (2%)C4-1 C4 spinal segment (right) 0.016 � 4C4-2 C4 spinal segment (right) 0.008 � 2C8-1 C8 spinal segment (right) 0.008CN9 Dentate (right) 0.012CN10 C3–C6 spinal segments (right) 0.016 � 10, 0.020, 0.032CN11 Dentate (left) and IO (left) 0.014 (DN) 0.008/0.012 (IO)CN12 Dentate (bilateral) 0.008/0.008PS1 C1 spinal segment (right) 0.016 � 2, 0.008 � 3, 0.012SC9 C6 spinal segment (left) 0.012SC12 C1–C5 spinal segments (right) 0.016 � 20ZI1 Zona incerta (left) 0.012
RNm and RNp, magno- and parvicellular divisions, respectively, of the rednucleus; IO, inferior olive.
454 M. PONG, K. M. HORN, AND A. R. GIBSON
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

left (contralateral to the injection) has been transposed to theright to compare locations of cells projecting to the ipsilateraland contralateral cord. Only a few ipsilateral cells overlap withthe area of contralateral projection (Fig. 2, column 1, right sideof section). Most cells projecting to the ipsilateral cord are locatedin the interstitial nucleus of Cajal (ICA), the area between RNand ICA, and, more rostrally, the Fields of Forel (FF).
TOPOGRAPHY OF RN PROJECTIONS TO THE SPINAL CORD. Theresults from the spinal injection case (CN10) indicated thatdorsolateral RN contains smaller cells that project to the con-tralateral cord. Do the cells in dorsolateral RN terminate at thesame spinal levels as cells in medial RN? We compared thelocations of retrogradely labeled neurons in the RN between
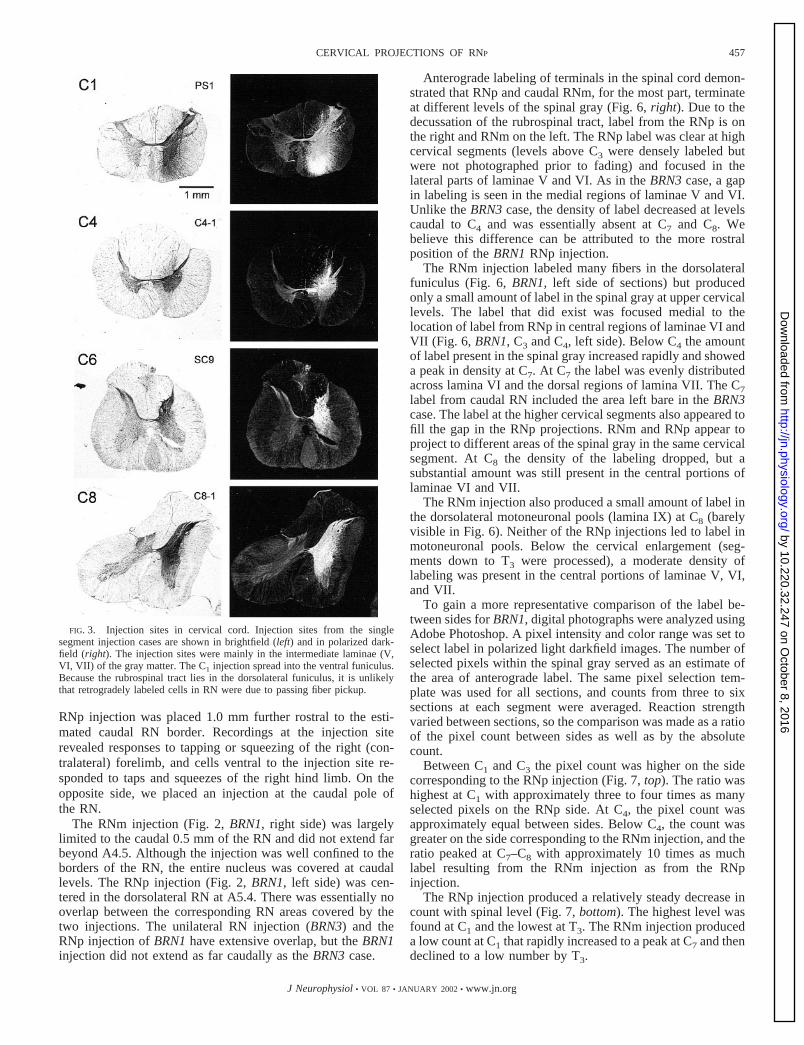
cases with injections restricted to one segment of the cervicalcord (Fig. 3). Although the injections sometimes includedfibers in the ventral funiculus (i.e., Fig. 3, C1), the dorsolateralfuniculus, where fibers from RN travel (Fig. 6), was not in-cluded by the injection sites. Therefore retrogradely labeledcells within the RN were due to pickup by terminals in thespinal gray and not by fibers of passage.
The C1 injection (Fig. 4, left) labeled cells across the medialto lateral extent of RN, although the largest number of labeledcells are in the most lateral section (L3.2). The C4 injectionproduced a relatively even distribution of labeled cells through-out the nucleus. (A replication of the C4 injection, case C4-2,resulted in the same distribution.) The C6 injection labeled alarge number of cells in the most medial section (L1.4), but fewcells were labeled in the lateral sections. The C8 injectionproduced a pattern similar to the C6 injection.
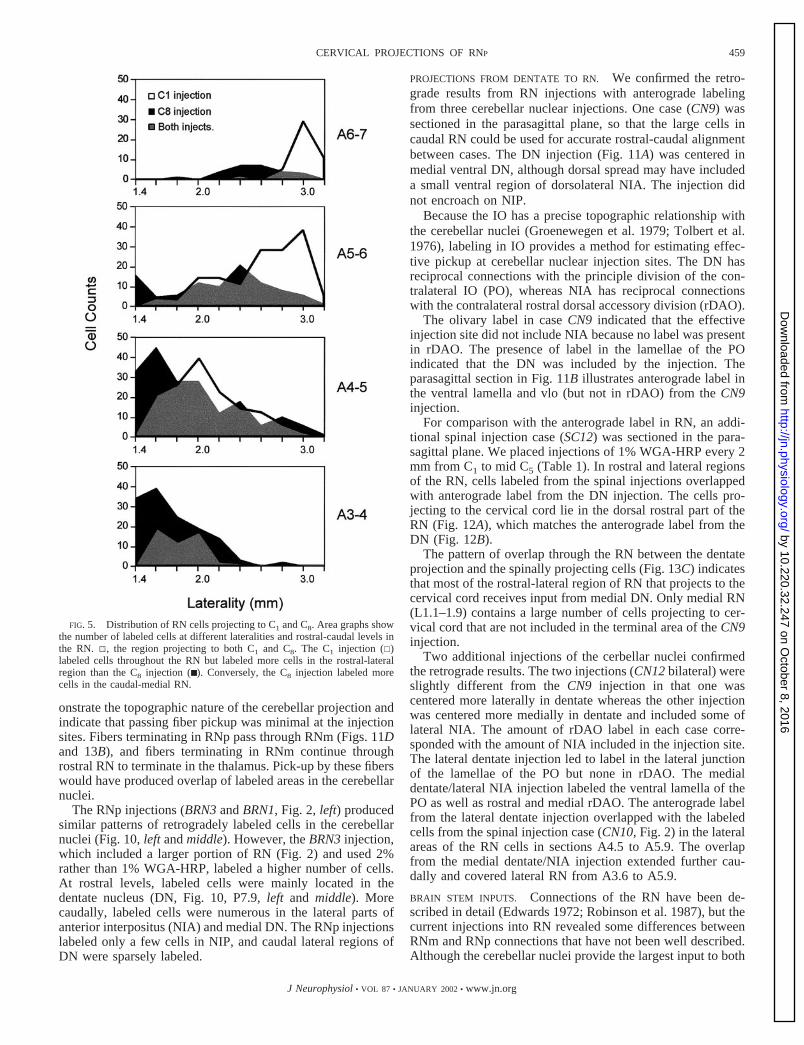
We compared the location of labeled RN cells from the C1and C8 injections in greater detail (Fig. 5). At caudal levels(A3-4), there is total overlap between C1 and C8 projectingcells with more cells labeled by the C8 injection. At morerostral levels, the distribution of labeled cells shifts laterallywith proportionately more resulting from the C1 injection. Thelargest amount of overlap occurs at mid-levels of the nucleus.The data indicate a modest topography in the cervical spinalprojection of RN. Although all segments seem to receive someinput from the entire RN, more cells in caudal and medial RN(RNm) project to C6 and C8, whereas more cells in rostral andlateral RN (RNp) project to C1. The intermediate segmentsshow more even distributions.
SPINAL LABELING FROM RED NUCLEUS INJECTIONS. To confirmthe pattern of RN projections to cervical cord, we made smallinjections of WGA-HRP into selected regions of the RN andtraced anterograde labeling in the cord. With anterograde trac-ing, both the laminar and segmental distribution of termina-tions can be determined.
To accurately place the injection sites, we first located thecaudal pole of RN by recording with a tungsten microelectrodeand used those coordinates to calculate the location of thedesired injection sites. In one case (BRN3, Table 1), we placeda single injection in the rostral RN on the left side 2 mm rostraland 0.5 mm lateral to the caudal pole. Cells at the injection sitefired during forelimb withdrawal in response to hard pinches ofthe paw.
The center of the BRN3 injection (Fig. 2, middle) waslocated in the lateral RN at A5.0 (reaction product above theinjection resulted from bleeding along the track). The injectionsite covered slightly more than the rostral half of the RN. Therewas little encroachment into the region dorsal and medial toRN that contains cells with ipsilateral spinal projections.
Contralateral to the injection, labeled fibers can be seen inthe dorsolateral funiculus (Fig. 6, left). Only a few labeledfibers could be found in the dorsolateral and ventral funiculi onthe ipsilateral (left) side, and only a small amount of terminallabel was present on this side. Most of the ipsilateral label waslocated in Rexed’s lamina VII (Rexed 1954) near the centralcanal and appeared to result from fibers crossing at segmentallevels.
On the contralateral side (right), many labeled fibers couldbe seen streaming from the dorsolateral funiculus into thespinal gray, and terminal label was present in laminae V, VI,
FIG. 1. Red nucleus (RN) neurons projecting to cervical spinal cord. Fron-tal sections illustrating wheat germ agglutinin-horseradish peroxidase (WGA-HRP)-labeled neurons with polarized darkfield illumination. At caudal levels(A3.8), predominately large cells in the dorsal half of the nucleus are labeled.More rostrally (A4.6), a second group of labeled cells appears dorsal andlateral in the nucleus. Although mixed in size, the cells are considerablysmaller than the caudal-medial group. Further rostrally (A5.2), the dorsolateralgroup is prominent, but the medial group has largely disappeared. Case CN10(Table 1; Fig. 2). Digitized 35-mm photographs, contrast adjusted digitally.
455CERVICAL PROJECTIONS OF RNP
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

and VII. At upper cervical levels (C1–C4), the labeled terminalsformed a focus in the lateral regions of lamina V and VI (nolabel was present in the lateral cervical nucleus). At C4, thelabeled terminals extended deeper into the dorsal part of laminaVII, and the projection to lamina VII was pronounced at C7and C8.
A gap in labeling can be seen in medial regions of laminaeVI and VII; this gap was most prominent at C7. The gap wasnot apparent at C8 (Fig. 6, BRN3). Below the cervical enlarge-ment (T2 was the lowest segment examined), the pattern ofterminal labeling resembled that seen at high cervical segmentswith most of the labeling confined to a lateral focus in laminaeV and VI. While the pattern of labeling was similar, the densityof labeling in the lower cervical and upper thoracic was sig-nificantly lower than at the higher cervical levels. No terminallabel was present in lamina IX (motoneuronal cell groups) atany level.COMPARISON OF SPINAL TERMINATIONS OF RNP TO RNM. TheRNp case (BRN3) confirmed a projection to the contralateralspinal cord. To compare projections of RNm and RNp, we
placed injections in different RN regions on opposite sides incase BRN1. The bilateral injection allowed us to compareacross sides at spinal levels so the relative density of labelingcould be judged in tissue processed under identical conditions.
Ipsilateral spinal projections from RN could confound bilat-eral injections. However, the results from CN10 and BRN3indicated that ipsilateral projections from rostral RN are insig-nificant. Furthermore, CN10 (Fig. 2) as well as the other spinalinjection cases (6 cases, Table 1) indicated that caudal RN andimmediately surrounding regions do not have a significantprojection to the ipsilateral cervical cord.
The lack of ipsilateral spinal projections from RNm agreeswell with previous studies (McCurdy et al. 1987; Pompeianoand Brodal 1957; Robinson et al. 1987). One case (1383 inHolstege 1987) has suggested ipsilateral spinal projectionsfrom RN; however, the injection site for this case included theregion dorsal and medial to rostral RN. Our data (i.e., CN10)also indicate an ipsilateral projection from this region, but itwas not included by our RN injections (Fig. 2).
The injection sites were identified as in BRN3 except that the
FIG. 2. RN cervical projection and RN injectionsites. Left: series of frontal sections illustrating labeledcells (black circles) resulting from injection into rightside of spinal segments C3–C6. A lateral and rostralextension of the nucleus extends from A4.5–A5.9. Anoutline (gray) of the labeled cells contralateral to theinjection has been added to the ipsilateral side (right)to show that very few cells within the RN project tothe ipsilateral cervical cord. A border indicating theventral edge of the RN has been added from A3.6 to5.4. Middle and right: injection sites of the unilateralcase BRN3 (0.004 �l, 2% WGA-HRP) and the bilat-eral case BRN1 (each injection, 0.008 �l, 1% WGA-HRP). Both injections on the left side covered therostral-lateral group of spinally projecting cells. Theinjection on the right of BRN1 included only thecaudal pole of the nucleus (RNm), which was notincluded by either of the left injections. None of thesites included areas with cells projecting to the ipsi-lateral cervical cord. Digital brightfield photographs.Injection sites reacted at the same strength as othersections.
456 M. PONG, K. M. HORN, AND A. R. GIBSON
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from
RNp injection was placed 1.0 mm further rostral to the esti-mated caudal RN border. Recordings at the injection siterevealed responses to tapping or squeezing of the right (con-tralateral) forelimb, and cells ventral to the injection site re-sponded to taps and squeezes of the right hind limb. On theopposite side, we placed an injection at the caudal pole ofthe RN.
The RNm injection (Fig. 2, BRN1, right side) was largelylimited to the caudal 0.5 mm of the RN and did not extend farbeyond A4.5. Although the injection was well confined to theborders of the RN, the entire nucleus was covered at caudallevels. The RNp injection (Fig. 2, BRN1, left side) was cen-tered in the dorsolateral RN at A5.4. There was essentially nooverlap between the corresponding RN areas covered by thetwo injections. The unilateral RN injection (BRN3) and theRNp injection of BRN1 have extensive overlap, but the BRN1injection did not extend as far caudally as the BRN3 case.
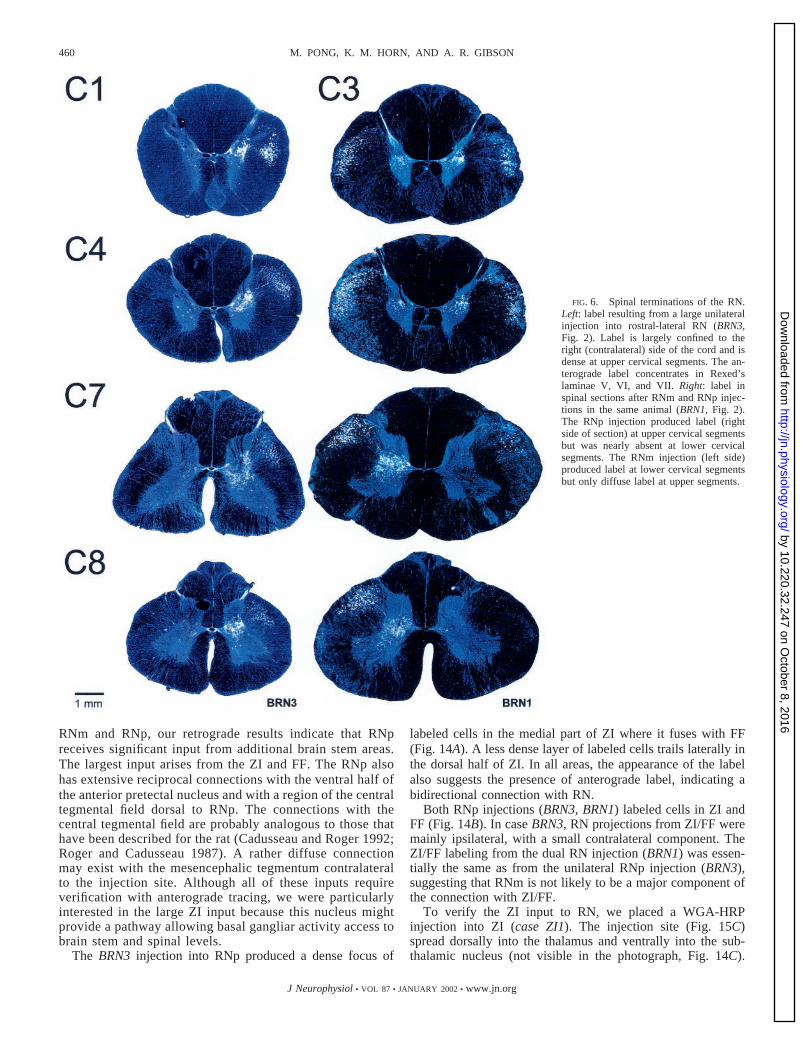
Anterograde labeling of terminals in the spinal cord demon-strated that RNp and caudal RNm, for the most part, terminateat different levels of the spinal gray (Fig. 6, right). Due to thedecussation of the rubrospinal tract, label from the RNp is onthe right and RNm on the left. The RNp label was clear at highcervical segments (levels above C3 were densely labeled butwere not photographed prior to fading) and focused in thelateral parts of laminae V and VI. As in the BRN3 case, a gapin labeling is seen in the medial regions of laminae V and VI.Unlike the BRN3 case, the density of label decreased at levelscaudal to C4 and was essentially absent at C7 and C8. Webelieve this difference can be attributed to the more rostralposition of the BRN1 RNp injection.
The RNm injection labeled many fibers in the dorsolateralfuniculus (Fig. 6, BRN1, left side of sections) but producedonly a small amount of label in the spinal gray at upper cervicallevels. The label that did exist was focused medial to thelocation of label from RNp in central regions of laminae VI andVII (Fig. 6, BRN1, C3 and C4, left side). Below C4 the amountof label present in the spinal gray increased rapidly and showeda peak in density at C7. At C7 the label was evenly distributedacross lamina VI and the dorsal regions of lamina VII. The C7label from caudal RN included the area left bare in the BRN3case. The label at the higher cervical segments also appeared tofill the gap in the RNp projections. RNm and RNp appear toproject to different areas of the spinal gray in the same cervicalsegment. At C8 the density of the labeling dropped, but asubstantial amount was still present in the central portions oflaminae VI and VII.
The RNm injection also produced a small amount of label inthe dorsolateral motoneuronal pools (lamina IX) at C8 (barelyvisible in Fig. 6). Neither of the RNp injections led to label inmotoneuronal pools. Below the cervical enlargement (seg-ments down to T3 were processed), a moderate density oflabeling was present in the central portions of laminae V, VI,and VII.
To gain a more representative comparison of the label be-tween sides for BRN1, digital photographs were analyzed usingAdobe Photoshop. A pixel intensity and color range was set toselect label in polarized light darkfield images. The number ofselected pixels within the spinal gray served as an estimate ofthe area of anterograde label. The same pixel selection tem-plate was used for all sections, and counts from three to sixsections at each segment were averaged. Reaction strengthvaried between sections, so the comparison was made as a ratioof the pixel count between sides as well as by the absolutecount.
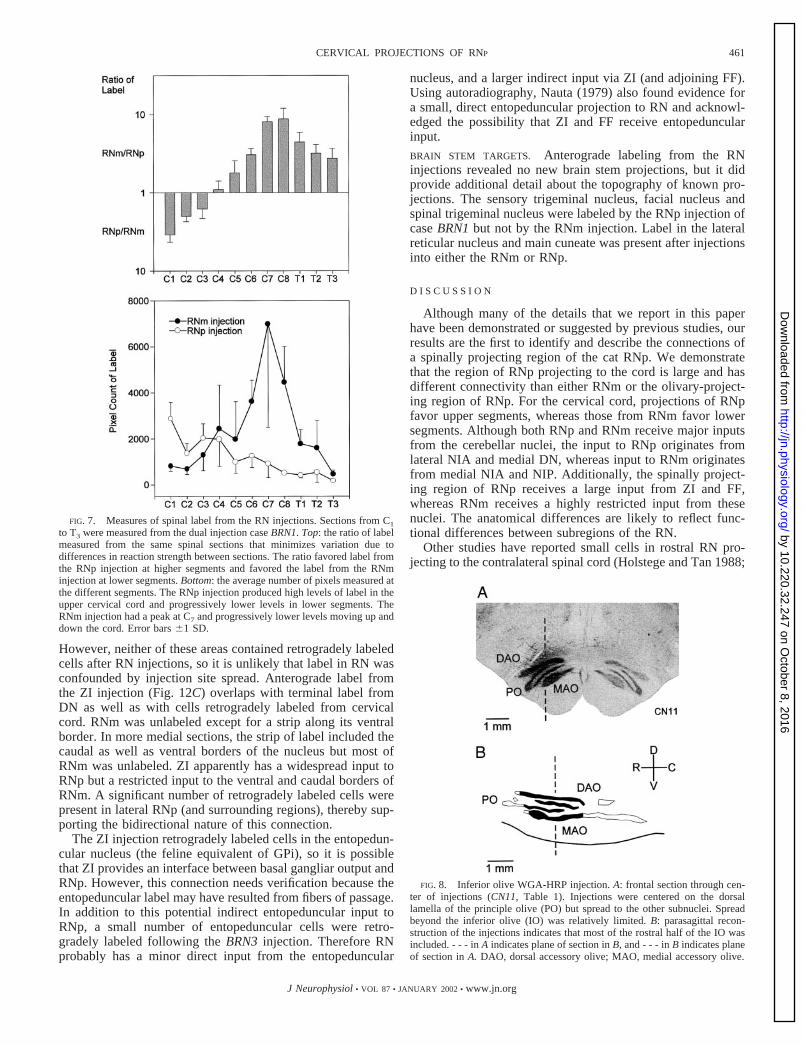
Between C1 and C3 the pixel count was higher on the sidecorresponding to the RNp injection (Fig. 7, top). The ratio washighest at C1 with approximately three to four times as manyselected pixels on the RNp side. At C4, the pixel count wasapproximately equal between sides. Below C4, the count wasgreater on the side corresponding to the RNm injection, and theratio peaked at C7–C8 with approximately 10 times as muchlabel resulting from the RNm injection as from the RNpinjection.
The RNp injection produced a relatively steady decrease incount with spinal level (Fig. 7, bottom). The highest level wasfound at C1 and the lowest at T3. The RNm injection produceda low count at C1 that rapidly increased to a peak at C7 and thendeclined to a low number by T3.
FIG. 3. Injection sites in cervical cord. Injection sites from the singlesegment injection cases are shown in brightfield (left) and in polarized dark-field (right). The injection sites were mainly in the intermediate laminae (V,VI, VII) of the gray matter. The C1 injection spread into the ventral funiculus.Because the rubrospinal tract lies in the dorsolateral funiculus, it is unlikelythat retrogradely labeled cells in RN were due to passing fiber pickup.
457CERVICAL PROJECTIONS OF RNP
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

The anterograde results are in good agreement with theretrograde results. Large cells at the caudal pole of the nucleushave a projection to laminae VI and VII at all cervical levels,but the projection is denser at lower segments. Smaller cells inthe rostral pole of RN project mainly to laminae V, VI, and VIIat upper cervical segments.
SPINAL VERSUS OLIVARY PROJECTIONS. Traditionally, RNp hasbeen thought to transmit information from the dentate nucleusof the cerebellum to the principle subnucleus of the ipsilateralinferior olive. Because both our retrograde and anterogradestudies indicated that a substantial portion of the cat RNpprojects to the contralateral spinal cord, could the same cellsprovide input to the inferior olive (IO)? The BRN3 injectionproduced some anterograde label in the ipsilateral IO (mostlyin the dorsal lamella of the principal olive), but the BRN1injections produced no visible label in the IO. Therefore it islikely that spinally projecting and olivary projecting cells inRNp are largely from different populations of neurons.
To determine the location of RN cells projecting to the IO,we made injections centered on the dorsal lamella of theprinciple subnucleus of the inferior olive (PO). Several studieshave indicated that the dorsal lamella receives input from theRN and surrounding regions (Edwards 1972; Holstege and Tan1988; Walberg 1956). Two injections separated by 1 mm in theA-P plane were placed into the dorsal lamella. At both injec-tion sites, hard squeezes to the limbs elicited discharge of theolivary neurons. Cells in the layer dorsal to the injection sitesresponded to light cutaneous stimulation of the forelimb, whichis characteristic of the rostral dorsal accessory olive (Gellmanet al. 1983). The spread of the injections included all divisionsof the rostral half of the IO but there was relatively little spreadinto the overlying reticular formation (Fig. 8A). Except for the
rostral tip, the injection site covered the rostral half of the IO(Fig. 8B).
Frontal sections from the olivary injection case (Fig. 9,CN11) were matched to those of the C3–C6 injection case (Fig.9, CN10) to compare the distributions of retrogradely labeledcells. The olivary injection resulted in no labeled cells in thecaudal half of the RN. In the rostral half, a few olivaryprojecting cells were located in medial portions of the nucleus.However, most cells projecting to the olive are found in cellgroups surrounding and including parts of the central gray (i.e.,nucleus Darkschewitsch, ND) and ventral regions of the pre-tectal nuclei.
The largest area of overlap between spinally and olivaryprojecting cells occurs at A5.4, where some cells in the mid-region of RN were labeled by the IO injection (Fig. 9, right).These cells probably account for the label seen in the dorsallamella for case BRN3 because that injection site includedmid-RN at this level. For most levels, however, there is nooverlap between olivary and spinally projecting cells. Thespinally projecting area of RNp appears to have little or noprojection to rostral IO.
Sources of input to the red nucleusLABELED CELLS IN THE CEREBELLAR NUCLEI. The retrogradelabeling from the RN injections allowed us to compare inputsto RNp and RNm. The heaviest source of input to the RN arisesfrom the deep cerebellar nuclei. Both dentate and interpositusprovide input to RN. The injection into the caudal pole of RNmlabeled a large number of cells in posterior interpositus (NIP)and the medial two-thirds of anterior interpositus (NIA) butfew cells in DN or lateral NIA (Fig. 10, right). In contrast, theRNp injections labeled cells in lateral NIA and DN (Fig. 10,left and middle). The mutually exclusive areas of label dem-
FIG. 4. Topography of RN projection tocervical cord. Retrogradely labeled RN cells(●) illustrated by a series of parasagittal sec-tions with injections at C1, C4, C6, and C8
(Fig. 3). The outline in each section indicatesthe borders of the RN for that particular sec-tion. The nucleus shifts to a rostral locationas it extends laterally. Although the casesindicate a great deal of overlap, the projec-tion of cells in lateral RN favors upper cer-vical levels, whereas the projection fromcells in medial RN favors lower cervical seg-ments. Ventral cells project to lower spinalsegments. The vertical line for each case rep-resents the caudal extent of the RN at approx-imately A3.0.
458 M. PONG, K. M. HORN, AND A. R. GIBSON
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from
onstrate the topographic nature of the cerebellar projection andindicate that passing fiber pickup was minimal at the injectionsites. Fibers terminating in RNp pass through RNm (Figs. 11Dand 13B), and fibers terminating in RNm continue throughrostral RN to terminate in the thalamus. Pick-up by these fiberswould have produced overlap of labeled areas in the cerebellarnuclei.
The RNp injections (BRN3 and BRN1, Fig. 2, left) producedsimilar patterns of retrogradely labeled cells in the cerebellarnuclei (Fig. 10, left and middle). However, the BRN3 injection,which included a larger portion of RN (Fig. 2) and used 2%rather than 1% WGA-HRP, labeled a higher number of cells.At rostral levels, labeled cells were mainly located in thedentate nucleus (DN, Fig. 10, P7.9, left and middle). Morecaudally, labeled cells were numerous in the lateral parts ofanterior interpositus (NIA) and medial DN. The RNp injectionslabeled only a few cells in NIP, and caudal lateral regions ofDN were sparsely labeled.
PROJECTIONS FROM DENTATE TO RN. We confirmed the retro-grade results from RN injections with anterograde labelingfrom three cerebellar nuclear injections. One case (CN9) wassectioned in the parasagittal plane, so that the large cells incaudal RN could be used for accurate rostral-caudal alignmentbetween cases. The DN injection (Fig. 11A) was centered inmedial ventral DN, although dorsal spread may have includeda small ventral region of dorsolateral NIA. The injection didnot encroach on NIP.
Because the IO has a precise topographic relationship withthe cerebellar nuclei (Groenewegen et al. 1979; Tolbert et al.1976), labeling in IO provides a method for estimating effec-tive pickup at cerebellar nuclear injection sites. The DN hasreciprocal connections with the principle division of the con-tralateral IO (PO), whereas NIA has reciprocal connectionswith the contralateral rostral dorsal accessory division (rDAO).
The olivary label in case CN9 indicated that the effectiveinjection site did not include NIA because no label was presentin rDAO. The presence of label in the lamellae of the POindicated that the DN was included by the injection. Theparasagittal section in Fig. 11B illustrates anterograde label inthe ventral lamella and vlo (but not in rDAO) from the CN9injection.
For comparison with the anterograde label in RN, an addi-tional spinal injection case (SC12) was sectioned in the para-sagittal plane. We placed injections of 1% WGA-HRP every 2mm from C1 to mid C5 (Table 1). In rostral and lateral regionsof the RN, cells labeled from the spinal injections overlappedwith anterograde label from the DN injection. The cells pro-jecting to the cervical cord lie in the dorsal rostral part of theRN (Fig. 12A), which matches the anterograde label from theDN (Fig. 12B).
The pattern of overlap through the RN between the dentateprojection and the spinally projecting cells (Fig. 13C) indicatesthat most of the rostral-lateral region of RN that projects to thecervical cord receives input from medial DN. Only medial RN(L1.1–1.9) contains a large number of cells projecting to cer-vical cord that are not included in the terminal area of the CN9injection.
Two additional injections of the cerbellar nuclei confirmedthe retrograde results. The two injections (CN12 bilateral) wereslightly different from the CN9 injection in that one wascentered more laterally in dentate whereas the other injectionwas centered more medially in dentate and included some oflateral NIA. The amount of rDAO label in each case corre-sponded with the amount of NIA included in the injection site.The lateral dentate injection led to label in the lateral junctionof the lamellae of the PO but none in rDAO. The medialdentate/lateral NIA injection labeled the ventral lamella of thePO as well as rostral and medial rDAO. The anterograde labelfrom the lateral dentate injection overlapped with the labeledcells from the spinal injection case (CN10, Fig. 2) in the lateralareas of the RN cells in sections A4.5 to A5.9. The overlapfrom the medial dentate/NIA injection extended further cau-dally and covered lateral RN from A3.6 to A5.9.
BRAIN STEM INPUTS. Connections of the RN have been de-scribed in detail (Edwards 1972; Robinson et al. 1987), but thecurrent injections into RN revealed some differences betweenRNm and RNp connections that have not been well described.Although the cerebellar nuclei provide the largest input to both
FIG. 5. Distribution of RN cells projecting to C1 and C8. Area graphs showthe number of labeled cells at different lateralities and rostral-caudal levels inthe RN. 1, the region projecting to both C1 and C8. The C1 injection (�)labeled cells throughout the RN but labeled more cells in the rostral-lateralregion than the C8 injection (■ ). Conversely, the C8 injection labeled morecells in the caudal-medial RN.
459CERVICAL PROJECTIONS OF RNP
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from
RNm and RNp, our retrograde results indicate that RNpreceives significant input from additional brain stem areas.The largest input arises from the ZI and FF. The RNp alsohas extensive reciprocal connections with the ventral half ofthe anterior pretectal nucleus and with a region of the centraltegmental field dorsal to RNp. The connections with thecentral tegmental field are probably analogous to those thathave been described for the rat (Cadusseau and Roger 1992;Roger and Cadusseau 1987). A rather diffuse connectionmay exist with the mesencephalic tegmentum contralateralto the injection site. Although all of these inputs requireverification with anterograde tracing, we were particularlyinterested in the large ZI input because this nucleus mightprovide a pathway allowing basal gangliar activity access tobrain stem and spinal levels.
The BRN3 injection into RNp produced a dense focus of
labeled cells in the medial part of ZI where it fuses with FF(Fig. 14A). A less dense layer of labeled cells trails laterally inthe dorsal half of ZI. In all areas, the appearance of the labelalso suggests the presence of anterograde label, indicating abidirectional connection with RN.
Both RNp injections (BRN3, BRN1) labeled cells in ZI andFF (Fig. 14B). In case BRN3, RN projections from ZI/FF weremainly ipsilateral, with a small contralateral component. TheZI/FF labeling from the dual RN injection (BRN1) was essen-tially the same as from the unilateral RNp injection (BRN3),suggesting that RNm is not likely to be a major component ofthe connection with ZI/FF.
To verify the ZI input to RN, we placed a WGA-HRPinjection into ZI (case ZI1). The injection site (Fig. 15C)spread dorsally into the thalamus and ventrally into the sub-thalamic nucleus (not visible in the photograph, Fig. 14C).
FIG. 6. Spinal terminations of the RN.Left: label resulting from a large unilateralinjection into rostral-lateral RN (BRN3,Fig. 2). Label is largely confined to theright (contralateral) side of the cord and isdense at upper cervical segments. The an-terograde label concentrates in Rexed’slaminae V, VI, and VII. Right: label inspinal sections after RNm and RNp injec-tions in the same animal (BRN1, Fig. 2).The RNp injection produced label (rightside of section) at upper cervical segmentsbut was nearly absent at lower cervicalsegments. The RNm injection (left side)produced label at lower cervical segmentsbut only diffuse label at upper segments.
460 M. PONG, K. M. HORN, AND A. R. GIBSON
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from
However, neither of these areas contained retrogradely labeledcells after RN injections, so it is unlikely that label in RN wasconfounded by injection site spread. Anterograde label fromthe ZI injection (Fig. 12C) overlaps with terminal label fromDN as well as with cells retrogradely labeled from cervicalcord. RNm was unlabeled except for a strip along its ventralborder. In more medial sections, the strip of label included thecaudal as well as ventral borders of the nucleus but most ofRNm was unlabeled. ZI apparently has a widespread input toRNp but a restricted input to the ventral and caudal borders ofRNm. A significant number of retrogradely labeled cells werepresent in lateral RNp (and surrounding regions), thereby sup-porting the bidirectional nature of this connection.
The ZI injection retrogradely labeled cells in the entopedun-cular nucleus (the feline equivalent of GPi), so it is possiblethat ZI provides an interface between basal gangliar output andRNp. However, this connection needs verification because theentopeduncular label may have resulted from fibers of passage.In addition to this potential indirect entopeduncular input toRNp, a small number of entopeduncular cells were retro-gradely labeled following the BRN3 injection. Therefore RNprobably has a minor direct input from the entopeduncular
nucleus, and a larger indirect input via ZI (and adjoining FF).Using autoradiography, Nauta (1979) also found evidence fora small, direct entopeduncular projection to RN and acknowl-edged the possibility that ZI and FF receive entopeduncularinput.
BRAIN STEM TARGETS. Anterograde labeling from the RNinjections revealed no new brain stem projections, but it didprovide additional detail about the topography of known pro-jections. The sensory trigeminal nucleus, facial nucleus andspinal trigeminal nucleus were labeled by the RNp injection ofcase BRN1 but not by the RNm injection. Label in the lateralreticular nucleus and main cuneate was present after injectionsinto either the RNm or RNp.
D I S C U S S I O N
Although many of the details that we report in this paperhave been demonstrated or suggested by previous studies, ourresults are the first to identify and describe the connections ofa spinally projecting region of the cat RNp. We demonstratethat the region of RNp projecting to the cord is large and hasdifferent connectivity than either RNm or the olivary-project-ing region of RNp. For the cervical cord, projections of RNpfavor upper segments, whereas those from RNm favor lowersegments. Although both RNp and RNm receive major inputsfrom the cerebellar nuclei, the input to RNp originates fromlateral NIA and medial DN, whereas input to RNm originatesfrom medial NIA and NIP. Additionally, the spinally project-ing region of RNp receives a large input from ZI and FF,whereas RNm receives a highly restricted input from thesenuclei. The anatomical differences are likely to reflect func-tional differences between subregions of the RN.
Other studies have reported small cells in rostral RN pro-jecting to the contralateral spinal cord (Holstege and Tan 1988;
FIG. 8. Inferior olive WGA-HRP injection. A: frontal section through cen-ter of injections (CN11, Table 1). Injections were centered on the dorsallamella of the principle olive (PO) but spread to the other subnuclei. Spreadbeyond the inferior olive (IO) was relatively limited. B: parasagittal recon-struction of the injections indicates that most of the rostral half of the IO wasincluded. - - - in A indicates plane of section in B, and - - - in B indicates planeof section in A. DAO, dorsal accessory olive; MAO, medial accessory olive.
FIG. 7. Measures of spinal label from the RN injections. Sections from C1
to T3 were measured from the dual injection case BRN1. Top: the ratio of labelmeasured from the same spinal sections that minimizes variation due todifferences in reaction strength between sections. The ratio favored label fromthe RNp injection at higher segments and favored the label from the RNminjection at lower segments. Bottom: the average number of pixels measured atthe different segments. The RNp injection produced high levels of label in theupper cervical cord and progressively lower levels in lower segments. TheRNm injection had a peak at C7 and progressively lower levels moving up anddown the cord. Error bars �1 SD.
461CERVICAL PROJECTIONS OF RNP
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

Mussen 1927; Pompeiano and Brodal 1957). However, ourdata indicate that the cells do not mix with cells projecting toIO but rather form a large homogeneous region. The spinallyprojecting region of RNp appears to be more extensive in ourstudy, but this is probably because we made a number ofclosely spaced WGA-HRP injections in upper cervical seg-ments. The closely spaced injections would result in moretracer pickup by the extensively branched RN axons (Shinodaet al. 1977, 1988), and RNp terminates most heavily at uppersegments, especially C1. Previous studies of RN spinal projec-tions would not have identified cells projecting selectively toupper cervical segments because they either sectioned the tractor injected tracers at spinal levels caudal to C1.
Our results indicate that the RNp populations projectingto the spinal cord and the IO are almost entirely separate,which is in good agreement with previous studies (Huismanet al. 1982; Spence and Saint-Cyr 1988). The mesencephalicinput to the IO in the cat arises from a complex of nuclei that
includes only a restricted rostral and medial portion of RNp(Onodera 1984; Saint-Cyr and Courville 1981; Walberg andNordby 1981). The separation of olivary and spinally pro-jecting neurons supports the suggestion by Holstege and Tan(1988) that RNp be divided into subregions. However, pro-jection targets are just one possible basis for subdividingRN. Equally important considerations are the source ofinputs to the subregions.
Using degeneration techniques, Voogd (1964) identified adorsolateral region about midway along the rostral caudalextent of the RN that receives input from the cerebellar dentatenucleus. Our data support his observation and also identify thisas a spinally projecting region of RN. These neurons maymediate the short latency activation of spinal neurons follow-ing stimulation of the dentate nucleus in the decerebrate cat(Hames et al. 1981).
Based on spinal connections, the smaller cells in lateral RNcould be grouped with RNm. However, based on cerebellar
FIG. 9. Limited overlap of olivary and spinallyprojecting cells. Left: cells (●) labeled after aC3–C6 spinal injection (case CN10, Fig. 2). Mid-dle: cells labeled after the IO injection (Fig. 5).Right: areas of potential overlap of olivary andspinally projecting cells. An outline of the RNprojection to the contralateral cord has been in-cluded in the enlarged series. The largest potentialfor overlap occurs at A5.4 but is modest even atthis level. FF, fields of Forel; FR, fasciculus ret-roflexus; ICA, interstitial nucleus of Cajal; ND,nucleus Darkshewitsch; PTA, anterior pretectalnucleus; PTP, posterior pretectal nucleus.
462 M. PONG, K. M. HORN, AND A. R. GIBSON
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

input, they would be grouped with RNp because RNp hastraditionally been considered a recipient of dentate input. Wechose to group the lateral spinally projecting cells with RNp,which is consistent with the Berman (1968) cat atlas (seehorizontal plates).
Figure 15 summarizes the major input-output relations ofspinally projecting regions of the RN. The figure shows twodifferent pathways from the cerebellum. The first pathway(light gray) starts from a strip that includes lateral NIA andmedial DN. The rostral-lateral RN receives this projection aswell as one from ZI and FF and then projects to the lateral areasof the upper cervical cord. The second pathway (black) origi-nates in medial NIA and then projects to the caudal-medial RN.Cells in this area of RN project to the lower cervical segmentsincluding motoneurons at C8. The topography consists of twopathways with an intermediate overlapping region (dark gray).Alternatively, the cerebellar nuclei from lateral to medial couldmap to the cervical cord from rostral to caudal in a continuum.
COMPARISONS BETWEEN SPECIES. Parvicellular RN projects tothe spinal cord in species other than cat. The most intensivelystudied species has been the rat (see Ruigrok and Cella 1995for review). As in the cat, the rat RN is not sharply divided intomagnocellular and parvicellular regions (Reid et al. 1975a,b),and at least some of the parvicellular neurons project to thespinal cord (Huisman et al. 1981, 1983; Shieh et al. 1983).Additionally, the rat RNp does not supply input to the inferiorolive (Ruigrok et al. 1993; Rutherford et al. 1984). Dorsolateralregions of the rat RNp receive input from caudal regions of thecerebellar lateral (dentate) nucleus (Angaut and Cicirata 1988;Ruigrok et al. 1993; Teune et al. 2000). Therefore a largeportion of the rat RNp receives input from the cerebellar lateralnucleus, has little or no projection to the IO, and has, at least,some projection to the spinal cord. It appears that the connec-tions are similar, perhaps identical, to the connections of thespinally projecting region of the cat RNp.
The RN of the North American opossum is organized muchlike the cat and rat. There is no distinct parvicellular division ofthe opossum red nucleus (King et al. 1971; Martin et al. 1974),and the cells projecting to the IO lie rostral and medial in theRN (Martin et al. 1983). As in the cat, there is only a smallregion of spatial overlap between the olivary and spinallyprojecting cells. Even in this overlap region, the populationsare distinct because very few cells project to both the spinalcord and IO (Martin et al. 1983).
Studies in monkey indicate spinal terminations similar to thecat, including direct connections to motorneuronal pools thatinnervate muscles that move the digits (Holstege et al. 1988;Ralston et al. 1988). Early studies using degeneration tracingtechniques identified small- and medium-sized cells in or nearlateral RN projecting to the contralateral spinal cord (Kuypersand Lawrence 1967; Poirier and Bouvier 1966); however, HRPtracing studies failed to demonstrate this population (Castigli-oni et al. 1978; Kneisley et al. 1978). More recently, Burmanet al. (2000) used WGA-HRP to confirm the existence of
FIG. 10. Cerebellar input to RNm and RNp. Series of frontal sectionsthrough interpositus and dentate indicating the locations of retrogradely la-beled cells resulting from the RN injections (BRN3 and BRN1). For compar-ison, the plots from the RNm injection (case BRN1, right) have been flipped sothat all nuclear outlines are in the same orientation, and the medial nucleus isnot represented because it contained no labeled cells. Additionally, the arealabeled by the RNm injection has been shaded on the plots from the RNpinjection cases. The RNp injections labeled cells in rostral and medial dentate(DN) and lateral anterior interpositus (NIA). The RNm injection labeled cellsthroughout the medial regions of NIA and posterior interpositus (NIP). Thereis little overlap between nuclear regions projecting to RNm and RNp.
FIG. 11. Dentate injection site and inferior oliveconnections. A: parasagittal section illustrating aninjection into medial DN (case CN9). - - -, area ten-tatively identified as lateral NIA; the dorsal edge ofthe injection spread slightly into this area. B: para-sagittal section illustrating label in IO following in-jection shown in A. Label is confined to the ventrallamella of the PO and the vlo, no label was apparentin rDAO. Panels are digitized 35-mm photographs.B was photographed using polarized darkfield illu-mination and has been digitally enhanced to visual-ize cellular grouping and label. Enhancements wereapplied to the entire tissue section.
463CERVICAL PROJECTIONS OF RNP
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

smaller cells dorsolateral to the RN that project to the con-tralateral cord. It seems likely that these smaller cells areanalogous to spinally projecting RNp cells of the cat.
In the human RN, there are few large cells, so most of thenucleus is presumed to be parvicellular and therefore olivaryprojecting (Massion 1967). However, as Pompeiano andBrodal (1957) pointed out, if many spinally projecting cells inthe human are small, the spinal projection may be greatlyunderestimated. Degeneration studies following brain stem le-sions in human infants support this suggestion (Papez andStottler 1940). The authors conclude that most of the rubro-spinal fibers in the human arise from small cells caudal andlateral in the nucleus. Such a projection fits well with the datafrom the degeneration studies in the monkey as well as with thedata from our study.
It is likely that species differences in RN connections are notas large as previously supposed. All of the aforementionedspecies have large cells projecting to the contralateral spinalcord that receive input from interpositus. All have small cellsin the rostral medial part of the nucleus that project to theipsilateral IO. Finally, we would like to suggest that all have alateral area consisting of smaller cells that project to the con-tralateral cord and receive input from DN.
The greatest confusion about RN connections may be theresult of a lack of correspondence between similarly namedregions. In the monkey, the area identified as RNp is probablyconfined to the olivary projecting region of RNp. In the rat andcat, the area identified as RNp includes both olivary andspinally projecting regions.
FUNCTIONAL IMPLICATIONS. Most RN output projects to inter-neurons, but, in some instances, the RST terminates amongmotoneurons. Probably the strongest motoneuronal projectionis to the facial nucleus (Courville 1966; Holstege et al. 1984;Robinson et al. 1987). Both of our RNp injections but not theRNm injection produced anterograde label in the facial nu-cleus. Inactivation of RN blocks the expression of a classicallyconditioned eye-blink response (Chapman et al. 1990) as doesinactivation of interpositus (Yeo et al. 1985), so it appears thatRNp provides lateral interpositus with a pathway to the facialnucleus.
The involvement of lateral NIA with facial movementsagrees well with its olivary connections. Medial regions of therDAO project to lateral NIA, which, in turn, projects to dor-solateral RN (Daniel et al. 1987; Gibson et al. 1987; Robinsonet al. 1987). Cells in medial rDAO (and the adjoining medialportion of the ventral lamella of the PO) respond to stimulationof the face and head (Gellman et al. 1983; Weiss et al. 1993)and receive input from the trigeminal nucleus (Berkley andHand 1978). The trigeminal nucleus also connects to a caudalregion of dorsolateral RN (Robinson et al. 1987). ThereforeRNp is likely to be involved in facial movements.
The only clear RN projection to spinal motor pools is tolateral pools at C8 (Fujito et al. 1991; Holstege 1987; McCurdyet al. 1987; Robinson et al. 1987). Our data indicate that RNmis the source of C8 motoneuronal terminations. Although theprojection to motor pools is small in comparison to projectionsto spinal interneurons, many studies have indicated that theRNm and interpositus are especially important for the control
FIG. 12. Inputs to RNp. Parasagittal sections (L2.6) compar-ing spinally projecting cells in RNp (A) with anterograde labelfrom dentate (B) and zona incerta (C). Each section is illustratedwith polarized darkfield illumination (left) to illustrate label andbrightfield illumination (right) to illustrate cellular grouping. A:the cervical injections labeled small- and medium-sized cells inrostral and dorsal RN. B: terminal label from medial dentate isrestricted to rostral and dorsal RN. Labeled fibers course throughcaudal regions of RN. C: terminal label from zona incerta injec-tion (Fig. 12B) is heaviest in the rostral half of RN, although astrip of labeled terminals extends caudally along the ventralborder of the nucleus.
464 M. PONG, K. M. HORN, AND A. R. GIBSON
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

of digit movements (Gibson et al. 1985, 1994; Jarratt andHyland 1999; Martin et al. 1993; Mason et al. 1998; Mewesand Cheney 1991; Sybirska and Gorska 1980; van Kan andMcCurdy 2001; van Kan et al. 1994; Whishaw and Gorny1996). Positioning of the digits appears to be an importantfunction of RNm and interpositus.
Despite the focus of RNm on digit control, inactivation andlesion studies (Bracha et al. 1999; Gibson et al. 1994; Law-rence and Kuypers 1968; Martin et al. 1993; Schrimsher andReier 1993; Sybirska and Gorska 1980) indicate that the RNinfluences proximal as well as distal limb musculature [al-though Levesque and Fabre-Thorpe (1990) report minimaldeficits in reaching with chemical lesions of the RN]. The RNpprojection to upper cervical levels suggests that this region maybe important for proximal limb control. Recordings at the RNpinjection sites indicated that the cells discharge during with-drawal of the forelimb, and the retrograde labeling indicatedthat they receive input from the cerebellar nuclei which inter-connect with the lamellae of the PO. Cells in the PO respondto stimulation of the limbs (Gellman et al. 1983), and it is likelythat RNp and DN form a second cerebello-spinal circuit in-volved in limb movement control. The findings reported in theaccompanying paper (Horn et al. 2002) indicate that the outputfrom RNp does have a strong influence on musculature of theshoulder.
RNp terminations at C1–C2 could be involved in the controlof head movements because motor neurons at these levelsinnervate neck muscles (Gordon and Richmond 1991; Kita-mura and Richmond 1994). However, RNp terminations favorlateral interneuronal regions, which project to motoneurons atother spinal levels. Interneuronal regions of C1–C2 receiveinput from many descending motor systems (Gibson et al.1998; Holstege and Kuypers 1982), including the tecto-spinalpathway (Cowie and Holstege 1992; Huerta and Harting 1982),which has been implicated in limb movement control (Werneret al. 1997). This region of interneurons may integrate a num-ber of inputs important for orienting a variety of body parts
FIG. 13. Dentate input to RN. Matched parasagittal sections illustrating over-lap of dentate terminals (A) with cells projecting to cervical cord (B). A: labeledterminals (small crosses) from a medial dentate injection (Fig. 11A, case CN9). Inall plots, the outlines indicate the borders of the RN for the individual sections. Thevertical line for each case represents the caudal extent of the RN at approximatelyA3.0. B: labeled cells (black circles) following a series of spinal injections from C1
to C5 (case SC12). C: overlap areas (gray) between A and B. The overlap is locatedin the rostral-lateral RN. Only medial sections (L1.0–1.5) contain a large numberof spinally projecting cells that do not overlap with dentate terminals.
FIG. 14. RN input from zona incerta (ZI)and fields of Forel (FF). A: polarized darkfieldphotograph of ZI shows labeled cells through-out the medial-lateral extent of the nucleusafter RNp injection. B: frontal sections throughZI and FF show locations of labeled cells (●)after RNp injection cases (BRN3 and BRN1).- - -, the laterality of the section shown in C(L5.2). C: parasagittal section shows the ZIinjection site that produced heavy terminal la-beling in RNp (Fig. 10C).
465CERVICAL PROJECTIONS OF RNP
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

such as the head, limbs, and trunk. Propriospinal interneuronsat C3–C4 are likely to be involved in limb control (Illert et al.1977) and interrupting propriospinal connections from thesesegments does produce deficits in reaching (Alstermark et al.1981).
Inactivation of the monkey cerebellar nuclei can also affectlimb guidance, although there is some debate about whichregions of the nuclei are critical. Both dentate (Thach et al.1992) and NIP inactivation (Mason et al. 1998) have beenreported to disrupt reach accuracy. If the topography of cere-bellar projections to RN is similar between cat and monkey,injections into NIP might also affect DN cells projecting toRNp. Alternatively, both NIP and DN might be important forreach accuracy. Posterior interpositus projects to the medialborder of the cat RN (Robinson et al. 1987), and our datademonstrate that cells in medial regions of RN project to allcervical levels. Therefore NIP may play an integrative roleinfluencing activity in both proximal and distal musculatureduring limb movements. Dentate may focus more on proximalmusculature, but deficits in either system could produce inac-curacy during reaching.
Humans with lesions of the lateral cerebellum reach inaccu-rately. In fact, lesions of the lateral cerebellum disrupt reachaccuracy more than lesions of the thalamus, which suggeststhat dentate output projects to areas important for reachingwithout passing through thalamus (Bastian and Thach 1995). Itis tempting to speculate that the small-celled lateral region of
human RN (Papez and Stottler 1940) allows dentate output toinfluence activity of the upper cervical cord.
Testing of the previous speculation will require a great dealof additional research. At least for the cat, it is likely that thespinal projections of RNp and RNm serve different roles in thecontrol of limb movements—it would be surprising if this werenot also the case for other mammalian species.
This work was supported by National Institute of Neurological Disordersand Stroke Grants NS-36820 to A. R. Gibson and NS-10726 to M. Pong.
REFERENCES
ALSTERMARK B, LUNDBERG A, NORRSELL U, AND SYBIRSKA E. Integration indescending motor pathways controlling the forelimb in the cat. XI. Differ-ential behavioural defects after spinal cord lesions interrupting definedpathways from higher centres to motoneurones. Exp Brain Res 42: 299–318,1981.
ANGAUT P AND CICIRATA F. The dentatorubral projection in the rat: an auto-radiographic study. Behav Brain Res 28: 71–73, 1988.
BASTIAN AJ AND THACH WT. Cerebellar outflow lesions: a comparison ofmovement deficits resulting from lesions at the levels of the cerebellum andthalamus. Ann Neurol 38: 881–892, 1995.
BERKLEY KJ AND HAND PJ. Projections to the inferior olive of the cat. II.Comparisons of input from the gracile, cuneate and the spinal trigeminalnuclei. J Comp Neurol 180: 253–264, 1978.
BERMAN AL. The Brain Stem of the Cat. Madison, WI: Univ. Wisconsin Press,1968.
BRACHA V, KOLB FP, IRWIN KB, AND BLOEDEL JR. Inactivation of interposednuclei in the cat: classically conditioned withdrawal reflexes, voluntary limbmovements and the action primitive hypothesis. Exp Brain Res 126: 77–92,1999.
BURMAN K, DARIAN-SMITH C, AND DARIAN-SMITH I. Macaque red nucleus:origins of spinal and olivary projections and terminations of cortical inputs.J Comp Neurol 423: 179–196, 2000.
CADUSSEAU J AND ROGER M. Connections of the posterior region of thethalamus in rats. J Hirnforsch 33: 19–35, 1992.
CASTIGLIONI AJ, GALLAWAY MC, AND COULTER JD. Spinal projections fromthe midbrain in monkey. J Comp Neurol 178: 329–346, 1978.
CHAPMAN PF, STEINMETZ JE, SEARS LL, AND THOMPSON RF. Effects of lido-caine injection in the interpositus nucleus and red nucleus on conditionedbehavioral and neuronal responses. Brain Res 537: 149–156, 1990.
COURVILLE J. Rubrobulbar fibres to the facial nucleus and the lateral reticularnucleus (nucleus of the lateral funiculus). An experimental study in the catwith silver impregnation methods. Brain Res 1: 317–337, 1966.
COWIE RJ AND HOLSTEGE G. Dorsal mesencephalic projections to pons, me-dulla, and spinal cord in the cat: limbic and non-limbic components. J CompNeurol 319: 536–559, 1992.
DANIEL H, BILLARD JM, ANGAUT P, AND BATINI C. The interposito-rubrospinalsystem. Anatomical tracing of a motor control pathway in the rat. NeurosciRes 5: 87–112, 1987.
EDWARDS SB. The ascending and descending projections of the red nucleus inthe cat: an experimental study using an autoradiographic tracing method.Brain Res 48: 45–63, 1972.
FABRE-THORPE M AND LEVESQUE F. Visuomotor relearning after brain damagecrucially depends on the integrity of the ventrolateral thalamic nucleus.Behav Neurosci 105: 176–192, 1991.
FUJITO Y, IMAI T, AND AOKI M. Monosynaptic excitation of motoneuronsinnervating forelimb muscles following stimulation of the red nucleus incats. Neurosci Lett 127: 137–140, 1991.
GELLMAN R, HOUK JC, AND GIBSON AR. Somatosensory properties of theinferior olive of the cat. J Comp Neurol 215: 228–243, 1983.
GIBSON AR, HANSMA DI, HOUK JC, AND ROBINSON FR. A sensitive low artifactTMB procedure for the demonstration of WGA-HRP in the CNS. Brain Res298: 235–241, 1984.
GIBSON AR, HORN KM, PONG M, AND VAN KAN PL. Construction of areach-to-grasp. Novartis Found Symp 218: 233–245, 1998.
GIBSON AR, HORN KM, AND VAN KAN PLE. Grasping cerebellar function. In:Insights into the Reach to Grasp Movement, edited by Bennet KMB andCastiello U. Amsterdam: Elsevier, 1994, p. 85–108.
GIBSON AR, HOUK JC, AND KOHLERMAN NJ. Magnocellular red nucleus activ-ity during different types of limb movement in the macaque monkey.J Physiol (Lond) 358: 527–549, 1985.
FIG. 15. Major elements of the cerebello-rubro-spinal pathways to cervicalcord. Two pathways from the cerebellar nuclei are shown. In one pathway,lateral cerebellum (dentate/lateral NIA, light gray) projects to rostral-lateralRNp, which then projects to the upper cervical cord. This pathway convergeswith input from ZI and FF. In the other pathway, output from medial NIA andNIP (black) projects to the medial-caudal RNm, which then projects to inter-mediate spinal regions and motoneurons at C8. The spinal projection is heavi-est at lower cervical. The darker gray indicates areas of potential overlap in RNand spinal cord.
466 M. PONG, K. M. HORN, AND A. R. GIBSON
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

GIBSON AR, ROBINSON FR, ALAM J, AND HOUK JC. Somatotopic alignmentbetween climbing fiber input and nuclear output of the cat intermediatecerebellum. J Comp Neurol 260: 362–377, 1987.
GORDON DC AND RICHMOND FJ. Distribution of motoneurons supplying dorsalsuboccipital and intervertebral muscles in the cat neck. J Comp Neurol 304:343–356, 1991.
GROENEWEGEN HJ, VOOGD J, AND FREEDMAN SL. The parasagittal zonationwithin the olivocerebellar projection. II. Climbing fiber distribution in theintermediate and hemispheric parts of cat cerebellum. J Comp Neurol 183:551–601, 1979.
HAMES EG, EBNER TJ, AND BLOEDEL JR. Electrophysiological demonstrationof a dentato-rubrospinal projection in cats. Neuroscience 6: 2603–2612,1981.
HOLSTEGE G. Anatomical evidence for an ipsilateral rubrospinal pathway andfor direct rubrospinal projections to motoneurons in the cat. Neurosci Lett74: 269–274, 1987.
HOLSTEGE G, BLOK BF, AND RALSTON DD. Anatomical evidence for rednucleus projections to motoneuronal cell groups in the spinal cord of themonkey. Neurosci Lett 95: 97–101, 1988.
HOLSTEGE G AND KUYPERS HG. The anatomy of brain stem pathways to thespinal cord in cat. A labeled amino acid tracing study. Prog Brain Res 57:145–175, 1982.
HOLSTEGE G AND TAN J. Projections from the red nucleus and surroundingareas to the brainstem and spinal cord in the cat. An HRP and autoradio-graphical tracing study. Behav Brain Res 28: 33–57, 1988.
HOLSTEGE G, TAN J, VAN HAM J, AND BOS A. Mesencephalic projections to thefacial nucleus in the cat. An autoradiographical tracing study. Brain Res311: 7–22, 1984.
HORN KM, PONG M, BATNI SR, LEVY SM, AND GIBSON AR. Functionalspecialization within the cat red nucleus. J Neurophysiol 87: 469–477,2002.
HUERTA MF AND HARTING JK. Tectal control of spinal cord activity: neuro-anatomical demonstration of pathways connecting the superior colliculuswith the cervical spinal cord grey. Prog Brain Res 57: 293–328, 1982.
HUISMAN AM, KUYPERS HG, CONDE F, AND KEIZER K. Collaterals of rubro-spinal neurons to the cerebellum in rat. A retrograde fluorescent doublelabeling study. Brain Res 264: 181–196, 1983.
HUISMAN AM, KUYPERS HG, AND VERBURGH CA. Quantitative differences incollateralization of the descending spinal pathways from red nucleus andother brain stem cell groups in rat as demonstrated with the multiplefluorescent retrograde tracer technique. Brain Res 209: 271–286, 1981.
HUISMAN AM, KUYPERS HG, AND VERBURGH CA. Differences in collateraliza-tion of the descending spinal pathways from red nucleus and other brainstem cell groups in cat and monkey. Prog Brain Res 57: 185–217, 1982.
ILLERT M, LUNDBERG A, AND TANAKA R. Integration in descending motorpathways controlling the forelimb in the cat. III. Convergence on proprio-spinal neurones transmitting disynaptic excitation from the corticospinaltract and other descending tracts. Exp Brain Res 29: 323–346, 1977.
JARRATT H AND HYLAND B. Neuronal activity in rat red nucleus duringforelimb reach-to-grasp movements. Neuroscience 88: 629–642, 1999.
KING JS, BOWMAN MH, AND MARTIN GF. The red nucleus of the opossum(Didelphis marsupialis virginiana): a light and electron microscopic study.J Comp Neurol 143: 157–184, 1971.
KITAMURA S AND RICHMOND FJ. Distribution of motoneurons supplying dorsaland ventral suboccipital muscles in the feline neck [published erratumappears in J Comp Neurol 351: 328, 1994]. J Comp Neurol 347: 25–35,1994.
KNEISLEY LW, BIBER MP, AND LAVAIL JH. A study of the origin of brain stemprojections to monkey spinal cord using the retrograde transport method.Exp Neurol 60: 116–139, 1978.
KUYPERS HG AND LAWRENCE DG. Cortical projections to the red nucleus andthe brain stem in the Rhesus monkey. Brain Res 4: 151–188, 1967.
LAWRENCE DG AND KUYPERS HG. The functional organization of the motorsystem in the monkey. II. The effects of lesions of the descending brain-stempathways. Brain 91: 15–36, 1968.
LEVESQUE F AND FABRE-THORPE M. Motor deficit induced by red nucleuslesion: re-appraisal using kainic acid destructions. Exp Brain Res 81: 191–198, 1990.
MARTIN GF, CABANA T, AND WALTZER R. Anatomical demonstration of thelocation and collateralization of rubral neurons which project to the spinalcord, lateral brainstem and inferior olive in the North American opossum.Brain Behav Evol 23: 93–109, 1983.
MARTIN GF, DOM R, KATZ S, AND KING JS. The organization of projectionneurons in the opossum red nucleus. Brain Res 78: 17–34, 1974.
MARTIN JH, COOPER SE, AND GHEZ C. Differential effects of local inactivationwithin motor cortex and red nucleus on performance of an elbow task in thecat. Exp Brain Res 94: 418–428, 1993.
MASON CR, MILLER LE, BAKER JF, AND HOUK JC. Organization of reachingand grasping movements in the primate cerebellar nuclei as revealed byfocal muscimol inactivations. J Neurophysiol 79: 537–554, 1998.
MASSION J. The mammalian red nucleus. Physiol Rev 47: 383–436, 1967.MCCURDY ML, HANSMA DI, HOUK JC, AND GIBSON AR. Selective projections
from the cat red nucleus to digit motor neurons [published erratum appearsin J Comp Neurol 273: 445, 1988]. J Comp Neurol 265: 367–379, 1987.
MESULAM MM. Principles of horseradish peroxidase neurochemistry and theirapplications for tracing neural pathways-axonal transport, enzyme histo-chemistry and light microscopic analysis. In: Tracing Neural Connectionswith Horseradish Peroxidase, edited by Mesulam MM. New York: Wiley,1982, p. 1–152.
MEWES K AND CHENEY PD. Facilitation and suppression of wrist and digitmuscles from single rubromotoneuronal cells in the awake monkey. J Neu-rophysiol 66: 1965–1977, 1991.
MUSSEN A. Experimental investigations on the cerbellum. Brain 50: 313–348,1927.
NAUTA HJ. Projections of the pallidal complex: an autoradiographic study inthe cat. Neuroscience 4: 1853–1873, 1979.
ONODERA S. Olivary projections from the mesodiencephalic structures in thecat studied by means of axonal transport of horseradish peroxidase andtritiated amino acids. J Comp Neurol 227: 37–49, 1984.
PAPEZ JW AND STOTTLER WA. Connections of the red nucleus. Arch NeurolPsychiatry 44: 776–791, 1940.
POIRIER LJ AND BOUVIER G. The red nucleus and its efferent nervous pathwaysin the monkey. J Comp Neurol 128: 223–244, 1966.
POMPEIANO O AND BRODAL A. Experimental demonstration of a somatotopicalorigin of rubrospinal fibers in the cat. J Comp Neurol 108: 225–251, 1957.
RALSTON DD, MILROY AM, AND HOLSTEGE G. Ultrastructural evidence fordirect monosynaptic rubrospinal connections to motoneurons in Macacamulatta. Neurosci Lett 95: 102–106, 1988.
RANISH NA AND SOECHTING JF. Studies on the control of some simple motortasks. Effects of thalamic and red nuclei lesions. Brain Res 102: 339–345,1976.
REID JM, FLUMERFELT BA, AND GWYN DG. An ultrastructural study of the rednucleus in the rat. J Comp Neurol 162: 363–385, 1975a.
REID JM, GWYN DG, AND FLUMERFELT BA. A cytoarchitectonic and Golgistudy of the red nucleus in the rat. J Comp Neurol 162: 337–361, 1975b.
REXED B. A cytoarchitectonic atlas of the spinal cord in the cat. J Comp Neurol100: 297–351, 1954.
ROBINSON FR, HOUK JC, AND GIBSON AR. Limb specific connections of the catmagnocellular red nucleus [published erratum appears in J Comp Neurol259: 622, 1987]. J Comp Neurol 257: 553–577, 1987.
ROGER M AND CADUSSEAU J. Anatomical evidence of a reciprocal connectionbetween the posterior thalamic nucleus and the parvocellular division of thered nucleus in the rat. A combined retrograde and anterograde study.Neuroscience 21: 573–583, 1987.
RUIGROK TJH AND CELLA F. Precerebellar nuclei and red nucleus. In: The RatNervous System, edtied by Paxinos G. San Diego: Academic, 1995, p.277–308.
RUIGROK TJH, KOEKKOEK B, AND KRUMM P. On the existence of cerebellarnucleo-olivary circuits—an anterograde and retrograde tracer study in therat. Eur J Neurosci Suppl 6: 125, 1993.
RUTHERFORD JG, ANDERSON WA, AND GWYN DG. A reevaluation of midbrainand diencephalic projections to the inferior olive in rat with particularreference to the rubro-olivary pathway. J Comp Neurol 229: 285–300, 1984.
SAINT-CYR JA AND COURVILLE J. Sources of descending afferents to theinferior olive from the upper brain stem in the cat as revealed by theretrograde transport of horseradish peroxidase. J Comp Neurol 198: 567–581, 1981.
SCHRIMSHER GW AND REIER PJ. Forelimb motor performance following dorsalcolumn, dorsolateral funiculi, or ventrolateral funiculi lesions of the cervicalspinal cord in the rat. Exp Neurol 120: 264–276, 1993.
SHIEH JY, LEONG SK, AND WONG WC. Origin of the rubrospinal tract inneonatal, developing, and mature rats. J Comp Neurol 214: 79–86, 1983.
SHINODA Y, FUTAMI T, MITOMA H, AND YOKOTA J. Morphology of singleneurones in the cerebello-rubrospinal system. Behav Brain Res 28: 59–64,1988.
SHINODA Y, GHEZ C, AND ARNOLD A. Spinal branching of rubrospinal axons inthe cat. Exp Brain Res 30: 203–218, 1977.
467CERVICAL PROJECTIONS OF RNP
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from

SPENCE SJ AND SAINT-CYR JA. Comparative topography of projections fromthe mesodiencephalic junction to the inferior olive, vestibular nuclei, andupper cervical cord in the cat. J Comp Neurol 268: 357–374, 1988.
SYBIRSKA E AND GORSKA T. Effects of red nucleus lesions on forelimbmovements in the cat. Acta Neurobiol Exp 40: 821–841, 1980.
TEUNE TM, VAN DER BURG J, VAN DER MOER J, VOOGD J, AND RUIGROK TJ.Topography of cerebellar nuclear projections to the brain stem in the rat.Prog Brain Res 124: 141–172, 2000.
THACH WT, GOODKIN HP, AND KEATING JG. The cerebellum and the adaptivecoordination of movement. Annu Rev Neurosci 15: 403–442, 1992.
TOLBERT DL, MASSOPUST LC, MURPHY MG, AND YOUNG PA. The anatomicalorganization of the cerebello-olivary projection in the cat. J Comp Neurol170: 525–544, 1976.
VAN KAN PL, HORN KM, AND GIBSON AR. The importance of hand use todischarge of interpositus neurones of the monkey [published erratum ap-pears in J Physiol (Lond) 481: 811, 1994]. J Physiol (Lond) 480: 171–190,1994.
VAN KAN PL AND MCCURDY ML. Role of primate magnocellular red nucleusneurons in controlling hand preshaping during reaching to grasp. J Neuro-physiol 85: 1461–1478, 2001.
VOOGD J. The Cerebellum of the Cat: Structure and Fibre Connexions.Philadelphia, PA: Davis, 1964.
WALBERG F. Descending connections to the inferior olive: an experimentalstudy in the cat. J Comp Neurol 104: 77–173, 1956.
WALBERG F AND NORDBY T. A re-examination of the rubro-olivary tract in thecat, using horseradish peroxidase as a retrograde and an anterograde neu-ronal tracer. Neuroscience 6: 2379–2391, 1981.
WEISS C, DISTERHOFT JF, GIBSON AR, AND HOUK JC. Receptive fields of singlecells from the face zone of the cat rostral dorsal accessory olive. Brain Res605: 207–213, 1993.
WERNER W, DANNENBERG S, AND HOFFMANN KP. Arm-movement-relatedneurons in the primate superior colliculus and underlying reticular forma-tion: comparison of neuronal activity with EMGs of muscles of the shoulder,arm and trunk during reaching. Exp Brain Res 115: 191–205, 1997.
WHISHAW IQ AND GORNY B. Does the red nucleus provide the tonic supportagainst which fractionated movements occur? A study on forepaw move-ments used in skilled reaching by the rat. Behav Brain Res 74: 79–90, 1996.
YEO CH, HARDIMAN MJ, AND GLICKSTEIN M. Classical conditioning of thenictitating membrane response of the rabbit. I. Lesions of the cerebellarnuclei. Exp Brain Res 60: 87–98, 1985.
468 M. PONG, K. M. HORN, AND A. R. GIBSON
J Neurophysiol • VOL 87 • JANUARY 2002 • www.jn.org
by 10.220.32.247 on October 8, 2016
http://jn.physiology.org/D
ownloaded from





























