Embed Size (px)
Citation preview

Spatial variation in landscape-level CO2 and CH4 fluxesfrom arctic coastal tundra: influence from vegetation,wetness, and the thaw lake cycleCOVE S . S TURTEVANT and WALTER C. OECHEL
Global Change Research Group, Department of Biology, San Diego State University, San Diego, CA, USA
Abstract
Regional quantification of arctic CO2 and CH4 fluxes remains difficult due to high landscape heterogeneity
coupled with a sparse measurement network. Most of the arctic coastal tundra near Barrow, Alaska is part of the
thaw lake cycle, which includes current thaw lakes and a 5500-year chronosequence of vegetated thaw lake basins.
However, spatial variability in carbon fluxes from these features remains grossly understudied. Here, we present
an analysis of whole-ecosystem CO2 and CH4 fluxes from 20 thaw lake cycle features during the 2011 growing
season. We found that the thaw lake cycle was largely responsible for spatial variation in CO2 flux, mostly due to
its control on gross primary productivity (GPP). Current lakes were significant CO2 sources that varied little. Veg-
etated basins showed declining GPP and CO2 sink with age (R2 = 67% and 57%, respectively). CH4 fluxes mea-
sured from a subset of 12 vegetated basins showed no relationship with age or CO2 flux components. Instead,
higher CH4 fluxes were related to greater landscape wetness (R2 = 57%) and thaw depth (additional R2 = 28%).
Spatial variation in CO2 and CH4 fluxes had good satellite remote sensing indicators, and we estimated the region
to be a small CO2 sink of �4.9 � 2.4 (SE) g C m�2 between 11 June and 25 August, which was countered by a
CH4 source of 2.1 � 0.2 (SE) g C m�2. Results from our scaling exercise showed that developing or validating
regional estimates based on single tower sites can result in significant bias, on average by a factor 4 for CO2 flux
and 30% for CH4 flux. Although our results are specific to the Arctic Coastal Plain of Alaska, the degree of land-
scape-scale variability, large-scale controls on carbon exchange, and implications for regional estimation seen here
likely have wide relevance to other arctic landscapes.
Keywords: Alaska, carbon dioxide, eddy covariance, methane, normalized difference red edge index, normalized difference
vegetation index, Tasseled Cap Transformation, upscaling
Received 24 January 2013; revised version received 27 April 2013 and accepted 29 April 2013
Introduction
The Arctic is rapidly changing. Atmospheric warming
in arctic regions has occurred at nearly double the glo-
bal rate (IPCC, 2007), thereby warming permafrost tem-
peratures (Shiklomanov et al., 2010), altering vegetation
communities (Sturm et al., 2001), and changing the
hydrological processes of many ecosystems (Hinzman
et al., 2005). Northern permafrost soils are estimated to
contain 1672 Pg of carbon, enough to more than double
the amount currently in the atmosphere (Zimov et al.,
2006; Tarnocai et al., 2009). The uncertain fate of this
large soil carbon pool necessitates accurate regional car-
bon source/sink estimation to track changes in the arc-
tic carbon balance and forecast future emissions.
However, developing regional estimates for the Arctic
are hampered by highly complex and heterogeneous
landscapes (McGuire et al., 2012).
High spatial variability in arctic land-atmosphere car-
bon exchange is due to the presence of impermeable
permafrost below a shallow summer thaw layer. This
results in water tables very close to the surface, and
therefore small variations in surface elevation (microto-
pography) lead to large differences in soil characteris-
tics, moisture, and vegetation over short spatial scales
(Brown et al., 1980). As these are some of the major fac-
tors controlling productivity and decomposition, the
variability in CO2 and CH4 fluxes at the ecosystem scale
or smaller can be similar to that over large latitudinal
and longitudinal transects (e.g., Morrissey & Living-
ston, 1992; Soegaard et al., 2000; McFadden et al., 2003).
Although the high spatial variability in arctic CO2
and CH4 flux rates is well known, few studies (espe-
cially for CH4) have measured and attributed this vari-
ability at the whole-ecosystem (eddy covariance tower)
level needed for improving regional estimates (but see
McFadden et al., 2003; Parmentier et al., 2011; Vourlitis
et al., 2003; Williams et al., 2000; Zona et al., 2010; Zulu-
eta et al., 2011). Therefore, validation of arctic regionalCorrespondence: C. S. Sturtevant, tel. + 303 489-8763,
fax + 619 594-7831, e-mail: [email protected]
© 2013 John Wiley & Sons Ltd 2853
Global Change Biology (2013) 19, 2853–2866, doi: 10.1111/gcb.12247

flux models often relies on a very sparse tower network
(e.g., Kimball et al., 2009) containing almost no informa-
tion about within-region variability.
On the Arctic Coastal Plain of Alaska near Barrow,
where this study is focused, the predominant landscape
variability consists of current and former thaw lakes.
Current lakes cover about 22% of the surface (Hinkel
et al., 2003). Another 50% is attributed to vegetated
thaw lake basins (VTLBs), which are previous thaw
lakes that drained at some point over that last ~5500years (Hinkel et al., 2003). The remaining 28% is
‘matrix’ tundra between the lakes and VTLBs. Thaw
lakes form via thermokarst processes (Cabot, 1947; Car-
son, 1968) and are shallow; 77% have depths <2.2 m
(Jeffries et al., 1996). Partial or total lake drainage can
occur at any time (Cabot, 1947; Carson, 1968). After
drainage, a slow process of ecosystem development
occurs over thousands of years until another lake
forms, often overlapping other VTLBs and restarting
the cycle (Billings & Peterson, 1980; Hinkel et al., 2003).
Although lakes and VTLBs cover 72% of the surface
and the ecological characteristics of VTLBs have been
well documented, landscape variability in CO2 and
CH4 fluxes arising from these features are not well
characterized. Kling et al.’s (1992) study of CO2 and
CH4 fluxes from lakes and rivers in Arctic Alaska sug-
gested that aquatic sources may reduce the regional
carbon sink by 20–50%. Despite this potentially large
importance, carbon flux estimates from thaw lakes on
the Arctic Coastal Plain of Alaska are few in number
and remain severely underrepresented in regional esti-
mation (for example, in McGuire et al., 2012). On the
terrestrial side, most CO2 flux studies in this region
have either focused on matrix tundra (e.g., Vourlitis &
Oechel, 1997; Kwon et al., 2006) or microform-level var-
iation (e.g., Oechel et al., 1995; Olivas et al., 2011). Only
two previous studies have addressed variability in CO2
flux along parts of the VTLB chronosequence (Zona
et al., 2010; Zulueta et al., 2011), but neither have
included variability from lakes or CH4 flux measure-
ments. Both these studies indicated that productivity
generally declines with age in the VTLBs, but much
uncertainty remains as to the influence of these features
on the net regional growing season CO2 balance. A few
VTLBs have been explicitly included in CH4 flux stud-
ies from this area (Zona et al., 2009; von Fischer et al.,
2010; Sturtevant et al., 2012), showing agreement with
the ample literature indicating soil moisture as a strong
driver of spatial variability (Morrissey & Livingston,
1992; Vourlitis et al., 1993; Christensen et al., 1995).
However, no studies have addressed the contribution
from the ecosystem development along the VTLB
chronosequence. Therefore, our objectives were to (i)
quantitatively determine how thaw lake cycle features
contribute to landscape-scale variability in net growing
season CO2 and CH4 fluxes; and (ii) determine whether
this knowledge can improve regional estimates. This
study represents one of very few arctic studies examin-
ing region-scale variability in whole-ecosystem CO2
flux and to our knowledge is the first of this breadth for
whole-ecosystem CH4 flux. Although the results pre-
sented here are specific to the Arctic Coastal Plain of
Alaska, the variability in carbon fluxes, large-scale con-
trols on carbon exchange, and implications for regional
scaling likely have wide relevance to other arctic land-
scapes.
Materials and methods
Site description
The wet meadow tundra near Barrow, Alaska lies within the
continuous permafrost zone, with maximum soil thaw reach-
ing 30–90 cm (Bockheim et al., 1999). Vegetation is dominated
by graminoids (incl. Arctophila fulva, Dupontia fisheri, Eriopho-
rum spp., Carex aquatilis), mosses (incl. Sphagnum, Dicranum
elongatum), lichens, and a few prostrate dwarf shrubs (Salix
spp., Cassiope tetragona) (Billings & Peterson, 1980; Hinkel et al.,
2003). The growing season extends from early June to late
August, with long-term (1971–2000) mean air temperature and
precipitation of 3.3 °C (�1.0 SD) and 56.5 mm (�26.8 SD),
respectively. The growing season of 2011 (the year of this
study) was warmer and slightly wetter than the long term
average (4.3 °C and 64.6 mm, respectively).
Previous work by Hinkel et al. (2003) classified the Barrow
area into five classes of the thaw lake cycle plus the remainder
of matrix tundra. We used this classification along with acces-
sibility by road or trail to identify sites for this study (Fig. 1;
Table S1). Thaw lake cycle classes include current lakes (Lake)
and four age classes of the ca. 5500 year VTLB chronose-
quence of ecosystem development after drainage (Young:
0–50 years, Medium: 50–300 years, Old: 300–2000 years, and
Ancient: 2000–5500 years.). Young, Medium, and Ancient
VTLBs are fewer in number, representing 4%, 9.2%, and 6.6%
of the land surface, respectively, while Old basins cover 30.4%
of the surface (Hinkel et al., 2003). Briefly, major VTLB
changes through time include: (i) general surface heave from
accumulation of ice and organic matter; (ii) increasing micro-
topography, polygon patterned ground and ponding (Fig. 1
inset) due to the expansion of ice wedges (which eventually
leads to the formation of a new lake); and (iii) a succession of
vegetation from fast growing pioneer species to longer lived,
low-vigor assemblages (Billings & Peterson, 1980; Hinkel et al.,
2003).
Study design
We used three portable eddy covariance towers deployed on
the tundra between 11 June and 27 August of 2011, all
equipped to measure CO2 flux and two of them equipped
to measure CH4 flux. As a result of the CH4 instrument
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
2854 C. S . STURTEVANT & W. C. OECHEL

limitation, we measured CO2 flux for all five classes of the
thaw lake cycle (Lake, Young, Medium, Old, Ancient), and
CH4 flux for a subset of VTLBs (Young, Medium, Old). To
measure spatial variation among the surface classes as well
as capture the seasonal pattern, we revisited a set of ‘Core’
thaw lake cycle sites (one of each class) during four periods
throughout the season while moving the towers to three
different ‘Spatial’ sets of thaw lake cycle sites during the three
intervening periods (see Fig. 1 timeline and full schedule in
Table S2). Thus, the Core set was always the same and the
three Spatial replicate sets were different each time, resulting
in 20 total sites measured for CO2 flux, and 12 of these for
CH4 flux. Within each period, the three towers were moved
among the five classes to obtain overlapping data for all clas-
ses (Table S2), ca. 6 days of data obtained for each site. The
exception to this was Lake sites, which were measured for ca.
3 days and typically overlapped only with Young and Med-
ium VTLBs. Actual period lengths were adjusted in the field to
achieve at least 4 days of high quality data for each VLTB site.
At Lake sites, the tower was placed at the edge of the water, as
most lakes of this region are <2.2 m deep and the flux foot-
print extended well into the interior of the lake. For VTLB
sites, the tower was placed as close to the center of the VTLB
as possible. Sites were named using the class of the thaw lake
cycle and whether the site was part of the Core set or one of
the Spatial replicates. For example, the Young VLTB part of
the Core set was named ‘Young Core’, while the three Spatial
replicates were named ‘Young 1’, ‘Young 2’, and ‘Young 3’.
Instrumentation
Net ecosystem exchange of CO2 (NEE), CH4 flux, and energy
fluxes were measured at a height of 1.9–2.4 m above the moss
layer or lake surface. Flux data were recorded at 10 Hz with a
datalogger (CR3000; Campbell Scientific, Logan, Utah, USA).
A sonic anemometer (CSAT3; Campbell Scientific, Logan,
Utah, USA) measured three-dimensional wind speed and vir-
tual sonic temperature. Molar densities of CO2 and H2O were
measured with an open-path infrared gas analyzer (LI-7500;
LI-COR Biosciences, Lincoln, NE, USA) inclined 15° from ver-
tical. Methane molar density was measured with a precom-
mercial version of the open-path LI-7700 methane analyzer
(LI-COR Biosciences, Lincoln, NE, USA). The gas analyzers
were calibrated at the beginning, middle, and end of the study
period using a 2-point linear equation and gases with certifica-
tion accuracy <1% as well as a dew point generator (LI-610;
LI-COR Biosciences, Lincoln, NE, USA).
Supporting meteorological and ground measurements were
sampled concurrently with flux measurements. Air tempera-
ture (Ta) and relative humidity were measured at ca. 1.7 m
height on the flux tower (HMP45C; Vaisala, Helsinki, Finland).
Atmospheric pressure was measured with the pressure trans-
ducer provided in the LI-7500 control box (absolute accu-
racy = 1.5%). Soil temperatures at 0 cm (within the moss
layer), 5 cm, and 10 cm depths were measured with type T
thermocouples attached to a wooden stick inserted into
the soil within the VTLB footprint. Net radiation (Rn) and
Fig. 1 Landsat TM 5 color infrared image of the study area. Measured sites are denoted with the colored symbols corresponding with
the thaw lake cycle class (see legend). The text inside each marker designates whether it was a Core site (‘C’) measured multiple times
over the season or whether it was a Spatial replicate (‘1’, ‘2’, or ‘3’) measured only once. The period(s) of measurement for each site are
indicated in the timeline below the image (underlined character in the timeline corresponds with the character in the marker). Dates
are in (mm/dd) format. The top left inset shows zoomed-in nadir views of an example surface within each thaw lake cycle class. The
white box within the main image indicates the region of extrapolated net growing season CO2 and CH4 fluxes presented in the results.
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
SPATIAL VARIATION IN ARCTIC CO2 AND CH4 FLUX 2855

photosynthetically active radiation (PAR) were measured on
a ca. 1 m high tripod adjacent to the tower with a net radiom-
eter (NR-Lite, Kipp, & Zonen, Delft, The Netherlands) and
quantum sensor (LI-190; LI-COR Biosciences, Lincoln, NE,
USA), respectively. Ground heat flux was averaged over 4–5
ground heat flux plates (HFT3; Campbell Scientific, Logan,
Utah, USA) buried at 2 cm depth within the tower footprint.
We also took measurements of soil temperature, net radiation,
and ground heat flux at Lake sites, placing the sensors just
inside the lake edge. However, we found that meteorological
measurements on the tower were more useful in explaining
the data, likely because the lake edge was unrepresentative.
Therefore, these measurements are not presented.
Volumetric soil moisture in the top 7 cm of soil (FieldScout
TDR 300, Spectrum, Inc., Plainfield, IL, USA) and thaw depth
(graduated rod) were sampled by hand every 10 m along two
to three 100 m transects within the footprint of the VTLBs each
time a tower was brought to or removed from a site. Volumet-
ric soil moisture measurements were converted to ‘landscape
wetness’ by calculating the percent of the field measurements
at or above saturation. This soil moisture metric is presented
because it was the most informative in analyses, providing a
more standardized metric of aerobic vs. anaerobic soil status
of the site. Daily thaw depth and landscape wetness were
obtained by linear interpolation between sample dates.
Flux processing
All computational procedures were coded in MATLAB
v. 7.12.0 (Mathworks, Natick, Massachusetts, USA). Half-
hourly flux calculations were made using the eddy covariance
method (Baldocchi et al., 1988). Raw 10 Hz data were despi-
ked (>6 standard deviations from the 1-min running mean)
and filtered for periods clearly demonstrating error. The sonic
anemometer coordinate frame was double rotated to align
with the mean wind streamline and signals from separate sen-
sors were lag-corrected. Corrections were applied for simulta-
neous sensible and latent heat fluxes (Webb et al., 1980;
McDermitt et al., 2010), high frequency spectral loss (Moore,
1986), and the LI-7500 self-heating correction (Burba et al.,
2008) (Method 4 and linear regression therein). Computed
fluxes indicating nonstationarity (Foken et al., 2004), low tur-
bulence conditions (friction velocity <0.1 m s�1), and footprint
outside the feature of interest were rejected. Quality-
controlled fluxes showed cospectra similar to Kaimal et al.
(1972). Energy budget closure averaged 81% for VTLB classes,
which is just above the FLUXNET average (79%) (Wilson et al.,
2002), and there were no substantial differences among
classes. Energy closure for Lake sites was not evaluated due to
the large unknown storage term. A footprint model (Hsieh
et al., 2000) indicated that under most observed conditions
80% of measured fluxes were attributed to the first 150 m and
300 m upwind from the tower for VTLBs and Lake sites,
respectively.
During each period, daily averages of NEE for VTLB sites
were obtained by gap-filling missing half-hourly measure-
ments using a semi-empirical light and temperature response
model (Falge et al., 2001). Days were gap-filled only if the
quality-controlled data for that day ensured a good fit for the
model. No gap-filling was performed for NEE at Lake sites or
CH4 flux at any site due to small observed diurnal patterns. In
these cases, daily averages were calculated with available
measurements as long as at least 4 h of quality-controlled
NEE or CH4 flux was present for the day. The number of daily
averages used for final analysis by site and period, as well as
the percentage of quality-controlled (nongap-filled) data cov-
erage for those days are given in Table S3.
The measured daily average NEE for VTLB sites was then
partitioned into gross primary productivity (GPP) and ecosys-
tem respiration (ER) via the equation:
NEE ¼ GPPþ ER
Positive values indicate sources to the atmosphere. ER for
each day was measured as the NEE at zero light, either
directly observed (at night) or estimated from the y-intercept
of each day’s PAR-NEE relationship with a linear equation for
PAR <300 lmol quanta m�2 s�1 as long as at least 6 points
were available and P < 0.05.
Modeling and analysis methods
Seasonal modeling. To compute cumulative seasonal fluxes
and present baselines from which to evaluate spatial variation,
we modeled the seasonal trends of daily CO2 and CH4 fluxes
from the Core sites. The purpose of this modeling was not to
strictly test physiological models published for other arctic
studies, but to provide the best possible extrapolation of Core
site fluxes during Spatial periods. Seasonal models for Lake
Core NEE as well as CH4 flux from all Core sites were mod-
eled solely according to correlations in the data and are pre-
sented in the Results. Seasonal NEE models for the Core
VTLBs were modified versions of those in Loranty et al.
(2011). Descriptions of the NEE models as well as coefficients
and performance plots are presented in Section 3 of the Sup-
porting Information. Monte Carlo simulation was performed
to determine statistical uncertainty on cumulative fluxes com-
puted from the seasonal models. Each modeled daily flux was
assigned a normally distributed random error with mean of 0
and standard deviation equal to the root mean square error
(RMSE) of the model. This procure was performed 1000 times
and the standard deviation of the resultant cumulative fluxes
computed.
Normalization. Since only three sites could be measured at
any one time, comparing fluxes and environmental measure-
ments among sites was sometimes confounded by daily varia-
tion and seasonal trends. Therefore, for measurements of thaw
depth, GPP, ER, and NEE we normalized Spatial site measure-
ments to baselines derived from interpolated or modeled
values from the Core sites to remove temporal variation and
isolate spatial differences. The mean thaw depth for each site
and period was converted to a percent difference from the
average Core VTLB thaw depth (interpolated) for the corre-
sponding days. Normalization of CO2 flux components for the
VTLB sites was done by calculating the relative magnitude of
the average daily flux for each site in relation to the minimum
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
2856 C. S . STURTEVANT & W. C. OECHEL

and maximum modeled Core VTLB fluxes for the correspond-
ing days via the equation:
Fnorm ¼ Fave � FCmin
FCmax � FCmin
F indicates the flux (NEE, GPP, or ER). Fnorm and Fave are the
normalized and nonnormalized average daily flux, respec-
tively, for a given site and period. FCmin and FCmax are the
minimum and maximum average modeled Core VTLB fluxes,
respectively, for the same days included in Fave. To retain gen-
eral sign conventions, GPPnorm and NEEnorm were subtracted
from 1 and the result multiplied by �1. A normalization
example using the data is given in Section 4 of the Supporting
Information.
Spatial analysis. After any normalization procedures, daily
measurements were averaged for each site and period (‘per-
iod-average’) for final analysis of spatial variation. The middle
two Core periods were averaged to provide one period-aver-
age value for each Core site since these two periods were clos-
est to the time frame of the Spatial measurements and showed
the best separability among sites. To analyze differences in
fluxes and environmental variables among sites, we used ANO-
VA and multiple regression. Post hoc pairwise comparisons for
ANOVA models were performed using Tukey’s Honestly Signifi-
cant Difference Criterion to control the Type I error rate (0.05).
To estimate the range of cumulative fluxes from all VTLB
sites, we substituted the normalized NEE values or period-
averaged CH4 values with cumulative season totals using the
relationships observed for the Core sites (shown in Section 5
of the Supporting Information). Cumulative season totals for
each of the Lake sites was estimated using the percent differ-
ence of each site from the Lake Core seasonal model. We used
the Monte Carlo procedure outlined above to determine the
statistical uncertainty on cumulative fluxes of the Spatial repli-
cate sites estimated via the relationship between normalized
or mid-season averages and cumulative fluxes for the Core
sites. In this case, the values in the regression were varied
within their individual uncertainties.
Regional scaling
We related our observations to established remote sensing
metrics acquired from satellite imagery and scaled our results
to a 170 km2 region surrounding our sites (white box in
Fig. 1). We used a high resolution (2 m) Worldview 2 image
(Basic LV1B product; Digital Globe Inc., Longmont, CO, USA)
from 21 July 2010 and a moderate resolution (30 m) Landsat
TM 5 image (Level 1T product) from 6 August 2011. Although
the Worldview 2 image was from 2010, we reasoned that spa-
tial differences among sites would remain similar between the
2010 and 2011 growing seasons, as the climatic conditions in
2010 and 2011 were not drastically different and ecosystem
change in this landscape occurs over much longer time scales.
We computed three physically based remote sensing met-
rics, including the Normalized Difference Vegetation Index
(‘NDVI’; Rouse et al., 1973), the Normalized Difference Red
Edge index (‘NDRE’; Barnes et al., 2000), and the Landsat TM
5 Tasseled Cap Transformation Wetness index (‘TCTw’; Crist
& Cicone, 1984). Higher NDVI values indicate more green,
healthy vegetation, and NVDI has shown good correlation
with arctic GPP and NEE (McMichael et al., 1999; Vourlitis
et al., 2000; Zulueta et al., 2011). NDRE is a close relative of
NDVI demonstrated to show enhanced sensitivity to plant
health (Tilling et al., 2007; Eitel et al., 2011). TCTw provides a
metric for surface water status, higher values indicating
greater surface moisture (Adams & Gillespie, 2006). Due to
sensor-specific bands, NDVI was calculated from both images,
NDRE from the WoldView 2 image only, and TCTw from the
Landsat TM 5 image only.
All image processing was performed in ENVI 5.0 (Exelis,
Inc., McLean, Virginia, USA). The Beaufort Sea, clouds, and
urban areas were masked out (3% of the WorldView 2 image,
7.8% of the Landsat TM 5 image). Remote sensing metrics
were computed from the digital numbers of the imagery with
no further radiometric or geometric corrections. Mean values
for each VTLB site were computed for the pixels within the
80% cumulative flux footprint. The borders of lakes and
VTLBs were digitized by hand. Lakes, VTLBs, and matrix tun-
dra accounted for 16%, 51%, and 34% of the 170 km2 region,
respectively.
We again used the Monte Carlo procedure outlined above
to determine the statistical uncertainty on regional fluxes com-
puted using regressions between remotely sensed indices and
observed fluxes.
Results
Environmental and site conditions
During the study period, air temperature showed a
broad peak in late July and early August (Fig. 2a), PAR
generally declined (Fig. 2b), and liquid precipitation
was intermittent (Fig. 2c). Ice-free dates of the Lake
measurement sites were estimated from Landsat imag-
ery: Lake Core on 21 June, Lake 3 on 27 June, Lake 1 on
5 July, and Lake 2 between 6 and 12 July.
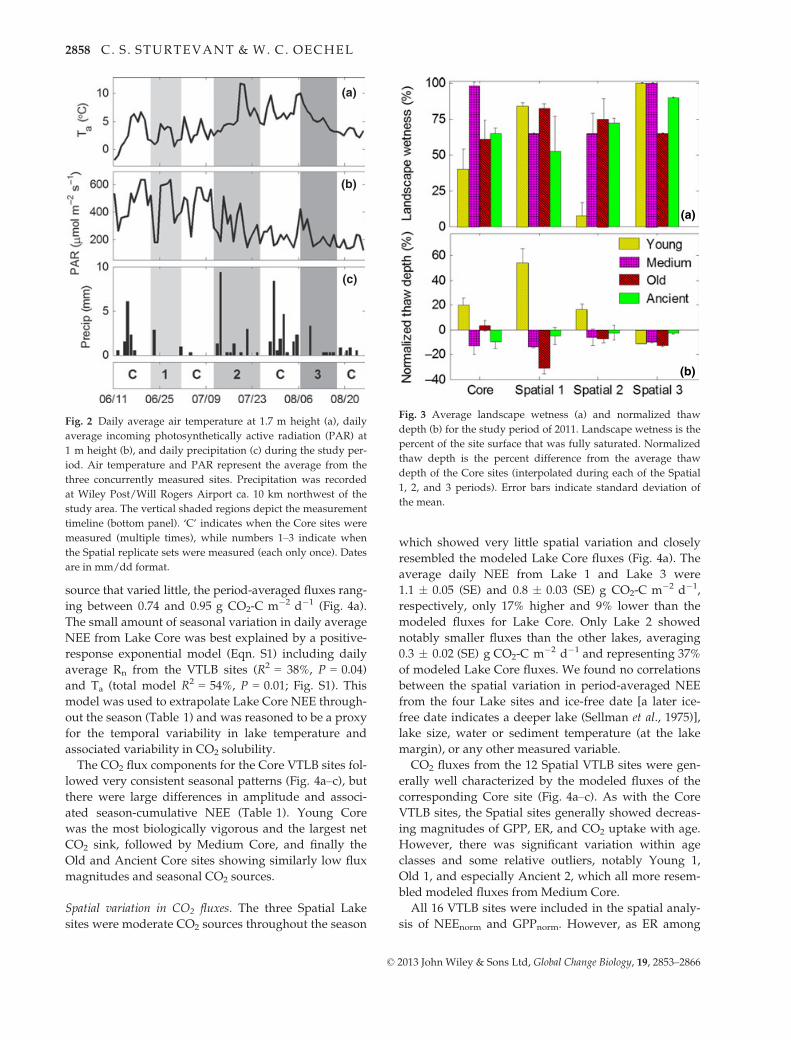
There was no consistent seasonal pattern in land-
scape wetness observed from the Core VTLBs. Includ-
ing the Spatial sites, period-average landscape wetness
ranged from near dry to completely flooded (Fig. 3a),
but landscape wetness was not associated with VTLB
age class (P = 0.54). In general, Young VTLBs exhibited
relatively higher thaw depth than the other age classes,
which showed no consistent differences (Fig. 3b;
P = 0.04). Other than thaw depth, no measured envi-
ronmental variables showed relative differences associ-
ated with VTLB age class (including soil temperature,
ground heat flux, and net radiation).
CO2 fluxes
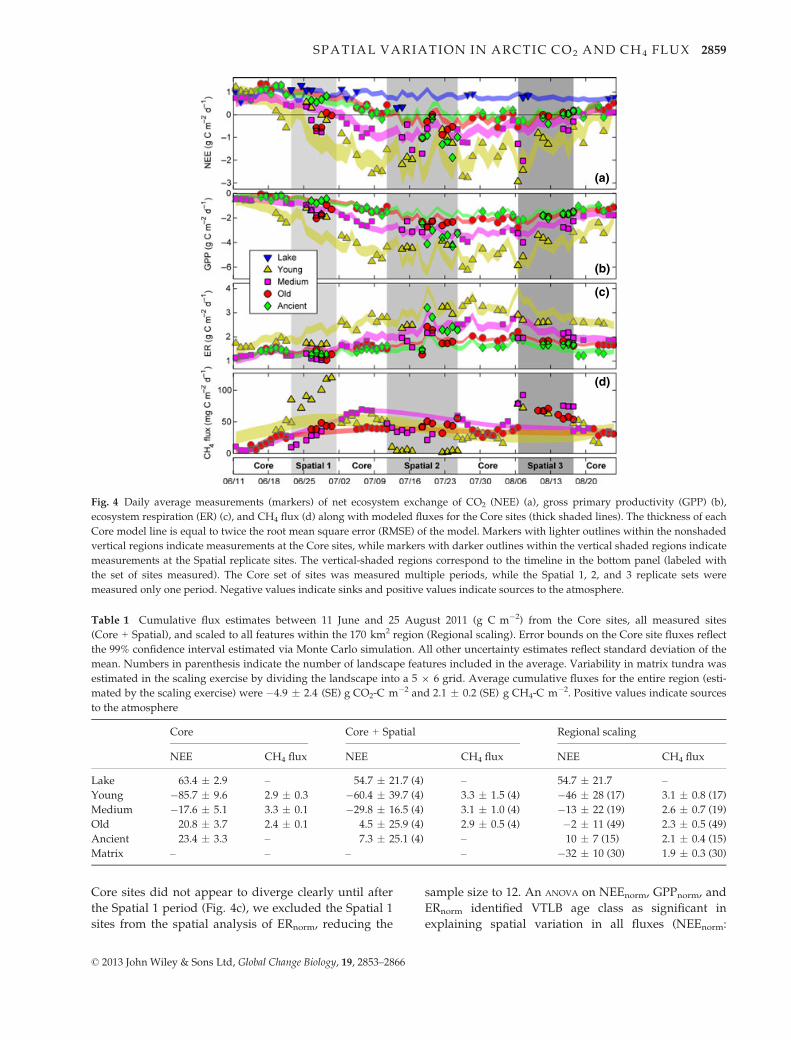
Seasonal variation in CO2 fluxes (Core sites). During the
growing season, Lake Core was a substantial CO2
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
SPATIAL VARIATION IN ARCTIC CO2 AND CH4 FLUX 2857
source that varied little, the period-averaged fluxes rang-
ing between 0.74 and 0.95 g CO2-C m�2 d�1 (Fig. 4a).
The small amount of seasonal variation in daily average
NEE from Lake Core was best explained by a positive-
response exponential model (Eqn. S1) including daily
average Rn from the VTLB sites (R2 = 38%, P = 0.04)
and Ta (total model R2 = 54%, P = 0.01; Fig. S1). This
model was used to extrapolate Lake Core NEE through-
out the season (Table 1) and was reasoned to be a proxy
for the temporal variability in lake temperature and
associated variability in CO2 solubility.
The CO2 flux components for the Core VTLB sites fol-
lowed very consistent seasonal patterns (Fig. 4a–c), butthere were large differences in amplitude and associ-
ated season-cumulative NEE (Table 1). Young Core
was the most biologically vigorous and the largest net
CO2 sink, followed by Medium Core, and finally the
Old and Ancient Core sites showing similarly low flux
magnitudes and seasonal CO2 sources.
Spatial variation in CO2 fluxes. The three Spatial Lake
sites were moderate CO2 sources throughout the season
which showed very little spatial variation and closely
resembled the modeled Lake Core fluxes (Fig. 4a). The
average daily NEE from Lake 1 and Lake 3 were
1.1 � 0.05 (SE) and 0.8 � 0.03 (SE) g CO2-C m�2 d�1,
respectively, only 17% higher and 9% lower than the
modeled fluxes for Lake Core. Only Lake 2 showed
notably smaller fluxes than the other lakes, averaging
0.3 � 0.02 (SE) g CO2-C m�2 d�1 and representing 37%
of modeled Lake Core fluxes. We found no correlations
between the spatial variation in period-averaged NEE
from the four Lake sites and ice-free date [a later ice-
free date indicates a deeper lake (Sellman et al., 1975)],
lake size, water or sediment temperature (at the lake
margin), or any other measured variable.
CO2 fluxes from the 12 Spatial VTLB sites were gen-
erally well characterized by the modeled fluxes of the
corresponding Core site (Fig. 4a–c). As with the Core
VTLB sites, the Spatial sites generally showed decreas-
ing magnitudes of GPP, ER, and CO2 uptake with age.
However, there was significant variation within age
classes and some relative outliers, notably Young 1,
Old 1, and especially Ancient 2, which all more resem-
bled modeled fluxes from Medium Core.
All 16 VTLB sites were included in the spatial analy-
sis of NEEnorm and GPPnorm. However, as ER among
(a)
(b)
(c)
Fig. 2 Daily average air temperature at 1.7 m height (a), daily
average incoming photosynthetically active radiation (PAR) at
1 m height (b), and daily precipitation (c) during the study per-
iod. Air temperature and PAR represent the average from the
three concurrently measured sites. Precipitation was recorded
at Wiley Post/Will Rogers Airport ca. 10 km northwest of the
study area. The vertical shaded regions depict the measurement
timeline (bottom panel). ‘C’ indicates when the Core sites were
measured (multiple times), while numbers 1–3 indicate when
the Spatial replicate sets were measured (each only once). Dates
are in mm/dd format.
(a)
(b)
Fig. 3 Average landscape wetness (a) and normalized thaw
depth (b) for the study period of 2011. Landscape wetness is the
percent of the site surface that was fully saturated. Normalized
thaw depth is the percent difference from the average thaw
depth of the Core sites (interpolated during each of the Spatial
1, 2, and 3 periods). Error bars indicate standard deviation of
the mean.
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
2858 C. S . STURTEVANT & W. C. OECHEL
Core sites did not appear to diverge clearly until after
the Spatial 1 period (Fig. 4c), we excluded the Spatial 1
sites from the spatial analysis of ERnorm, reducing the
sample size to 12. An ANOVA on NEEnorm, GPPnorm, and
ERnorm identified VTLB age class as significant in
explaining spatial variation in all fluxes (NEEnorm:
(a)
(b)
(c)
(d)
Fig. 4 Daily average measurements (markers) of net ecosystem exchange of CO2 (NEE) (a), gross primary productivity (GPP) (b),
ecosystem respiration (ER) (c), and CH4 flux (d) along with modeled fluxes for the Core sites (thick shaded lines). The thickness of each
Core model line is equal to twice the root mean square error (RMSE) of the model. Markers with lighter outlines within the nonshaded
vertical regions indicate measurements at the Core sites, while markers with darker outlines within the vertical shaded regions indicate
measurements at the Spatial replicate sites. The vertical-shaded regions correspond to the timeline in the bottom panel (labeled with
the set of sites measured). The Core set of sites was measured multiple periods, while the Spatial 1, 2, and 3 replicate sets were
measured only one period. Negative values indicate sinks and positive values indicate sources to the atmosphere.
Table 1 Cumulative flux estimates between 11 June and 25 August 2011 (g C m�2) from the Core sites, all measured sites
(Core + Spatial), and scaled to all features within the 170 km2 region (Regional scaling). Error bounds on the Core site fluxes reflect
the 99% confidence interval estimated via Monte Carlo simulation. All other uncertainty estimates reflect standard deviation of the
mean. Numbers in parenthesis indicate the number of landscape features included in the average. Variability in matrix tundra was
estimated in the scaling exercise by dividing the landscape into a 5 9 6 grid. Average cumulative fluxes for the entire region (esti-
mated by the scaling exercise) were �4.9 � 2.4 (SE) g CO2-C m�2 and 2.1 � 0.2 (SE) g CH4-C m�2. Positive values indicate sources
to the atmosphere
Core Core + Spatial Regional scaling
NEE CH4 flux NEE CH4 flux NEE CH4 flux
Lake 63.4 � 2.9 – 54.7 � 21.7 (4) – 54.7 � 21.7 –
Young �85.7 � 9.6 2.9 � 0.3 �60.4 � 39.7 (4) 3.3 � 1.5 (4) �46 � 28 (17) 3.1 � 0.8 (17)
Medium �17.6 � 5.1 3.3 � 0.1 �29.8 � 16.5 (4) 3.1 � 1.0 (4) �13 � 22 (19) 2.6 � 0.7 (19)
Old 20.8 � 3.7 2.4 � 0.1 4.5 � 25.9 (4) 2.9 � 0.5 (4) �2 � 11 (49) 2.3 � 0.5 (49)
Ancient 23.4 � 3.3 – 7.3 � 25.1 (4) – 10 � 7 (15) 2.1 � 0.4 (15)
Matrix – – – – �32 � 10 (30) 1.9 � 0.3 (30)
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
SPATIAL VARIATION IN ARCTIC CO2 AND CH4 FLUX 2859
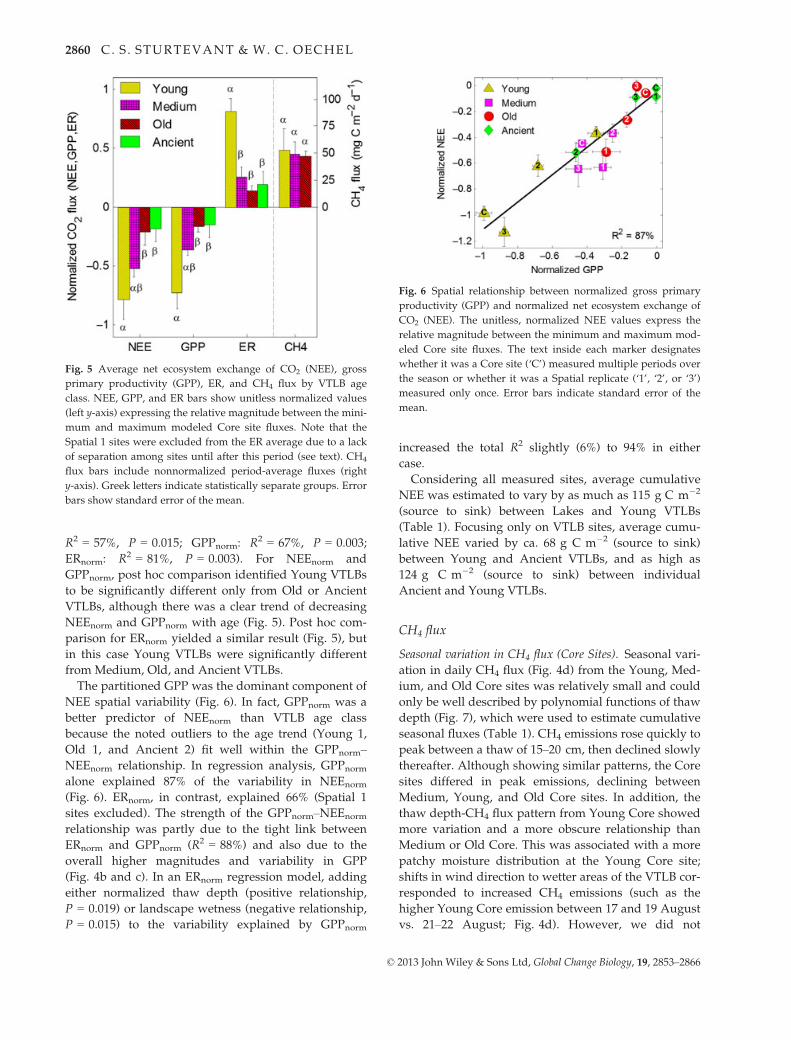
R2 = 57%, P = 0.015; GPPnorm: R2 = 67%, P = 0.003;
ERnorm: R2 = 81%, P = 0.003). For NEEnorm and
GPPnorm, post hoc comparison identified Young VTLBs
to be significantly different only from Old or Ancient
VTLBs, although there was a clear trend of decreasing
NEEnorm and GPPnorm with age (Fig. 5). Post hoc com-
parison for ERnorm yielded a similar result (Fig. 5), but
in this case Young VTLBs were significantly different
from Medium, Old, and Ancient VTLBs.
The partitioned GPP was the dominant component of
NEE spatial variability (Fig. 6). In fact, GPPnorm was a
better predictor of NEEnorm than VTLB age class
because the noted outliers to the age trend (Young 1,
Old 1, and Ancient 2) fit well within the GPPnorm–NEEnorm relationship. In regression analysis, GPPnorm
alone explained 87% of the variability in NEEnorm
(Fig. 6). ERnorm, in contrast, explained 66% (Spatial 1
sites excluded). The strength of the GPPnorm–NEEnorm
relationship was partly due to the tight link between
ERnorm and GPPnorm (R2 = 88%) and also due to the
overall higher magnitudes and variability in GPP
(Fig. 4b and c). In an ERnorm regression model, adding
either normalized thaw depth (positive relationship,
P = 0.019) or landscape wetness (negative relationship,
P = 0.015) to the variability explained by GPPnorm
increased the total R2 slightly (6%) to 94% in either
case.
Considering all measured sites, average cumulative
NEE was estimated to vary by as much as 115 g C m�2
(source to sink) between Lakes and Young VTLBs
(Table 1). Focusing only on VTLB sites, average cumu-
lative NEE varied by ca. 68 g C m�2 (source to sink)
between Young and Ancient VTLBs, and as high as
124 g C m�2 (source to sink) between individual
Ancient and Young VTLBs.
CH4 flux
Seasonal variation in CH4 flux (Core Sites). Seasonal vari-
ation in daily CH4 flux (Fig. 4d) from the Young, Med-
ium, and Old Core sites was relatively small and could
only be well described by polynomial functions of thaw
depth (Fig. 7), which were used to estimate cumulative
seasonal fluxes (Table 1). CH4 emissions rose quickly to
peak between a thaw of 15–20 cm, then declined slowly
thereafter. Although showing similar patterns, the Core
sites differed in peak emissions, declining between
Medium, Young, and Old Core sites. In addition, the
thaw depth-CH4 flux pattern from Young Core showed
more variation and a more obscure relationship than
Medium or Old Core. This was associated with a more
patchy moisture distribution at the Young Core site;
shifts in wind direction to wetter areas of the VTLB cor-
responded to increased CH4 emissions (such as the
higher Young Core emission between 17 and 19 August
vs. 21–22 August; Fig. 4d). However, we did not
Fig. 5 Average net ecosystem exchange of CO2 (NEE), gross
primary productivity (GPP), ER, and CH4 flux by VTLB age
class. NEE, GPP, and ER bars show unitless normalized values
(left y-axis) expressing the relative magnitude between the mini-
mum and maximum modeled Core site fluxes. Note that the
Spatial 1 sites were excluded from the ER average due to a lack
of separation among sites until after this period (see text). CH4
flux bars include nonnormalized period-average fluxes (right
y-axis). Greek letters indicate statistically separate groups. Error
bars show standard error of the mean.
Fig. 6 Spatial relationship between normalized gross primary
productivity (GPP) and normalized net ecosystem exchange of
CO2 (NEE). The unitless, normalized NEE values express the
relative magnitude between the minimum and maximum mod-
eled Core site fluxes. The text inside each marker designates
whether it was a Core site (‘C’) measured multiple periods over
the season or whether it was a Spatial replicate (‘1’, ‘2’, or ‘3’)
measured only once. Error bars indicate standard error of the
mean.
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
2860 C. S . STURTEVANT & W. C. OECHEL
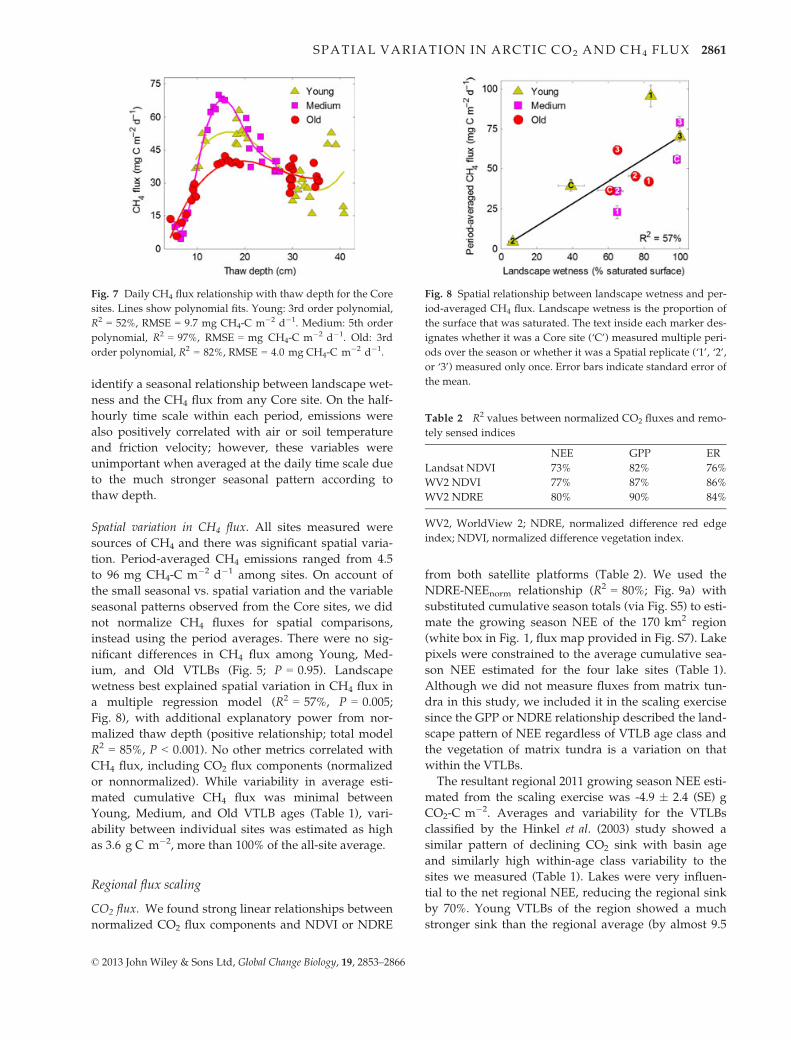
identify a seasonal relationship between landscape wet-
ness and the CH4 flux from any Core site. On the half-
hourly time scale within each period, emissions were
also positively correlated with air or soil temperature
and friction velocity; however, these variables were
unimportant when averaged at the daily time scale due
to the much stronger seasonal pattern according to
thaw depth.
Spatial variation in CH4 flux. All sites measured were
sources of CH4 and there was significant spatial varia-
tion. Period-averaged CH4 emissions ranged from 4.5
to 96 mg CH4-C m�2 d�1 among sites. On account of
the small seasonal vs. spatial variation and the variable
seasonal patterns observed from the Core sites, we did
not normalize CH4 fluxes for spatial comparisons,
instead using the period averages. There were no sig-
nificant differences in CH4 flux among Young, Med-
ium, and Old VTLBs (Fig. 5; P = 0.95). Landscape
wetness best explained spatial variation in CH4 flux in
a multiple regression model (R2 = 57%, P = 0.005;
Fig. 8), with additional explanatory power from nor-
malized thaw depth (positive relationship; total model
R2 = 85%, P < 0.001). No other metrics correlated with
CH4 flux, including CO2 flux components (normalized
or nonnormalized). While variability in average esti-
mated cumulative CH4 flux was minimal between
Young, Medium, and Old VTLB ages (Table 1), vari-
ability between individual sites was estimated as high
as 3.6 g C m�2, more than 100% of the all-site average.
Regional flux scaling
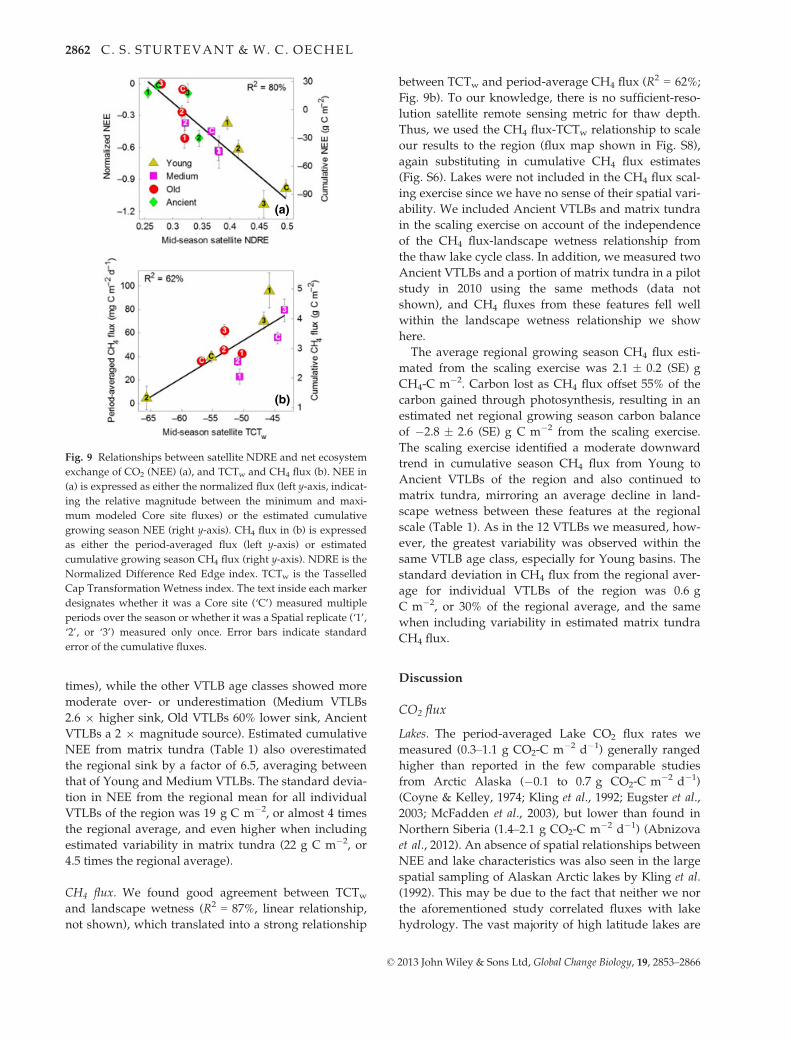
CO2 flux. We found strong linear relationships between
normalized CO2 flux components and NDVI or NDRE
from both satellite platforms (Table 2). We used the
NDRE-NEEnorm relationship (R2 = 80%; Fig. 9a) with
substituted cumulative season totals (via Fig. S5) to esti-
mate the growing season NEE of the 170 km2 region
(white box in Fig. 1, flux map provided in Fig. S7). Lake
pixels were constrained to the average cumulative sea-
son NEE estimated for the four lake sites (Table 1).
Although we did not measure fluxes from matrix tun-
dra in this study, we included it in the scaling exercise
since the GPP or NDRE relationship described the land-
scape pattern of NEE regardless of VTLB age class and
the vegetation of matrix tundra is a variation on that
within the VTLBs.
The resultant regional 2011 growing season NEE esti-
mated from the scaling exercise was -4.9 � 2.4 (SE) g
CO2-C m�2. Averages and variability for the VTLBs
classified by the Hinkel et al. (2003) study showed a
similar pattern of declining CO2 sink with basin age
and similarly high within-age class variability to the
sites we measured (Table 1). Lakes were very influen-
tial to the net regional NEE, reducing the regional sink
by 70%. Young VTLBs of the region showed a much
stronger sink than the regional average (by almost 9.5
Fig. 7 Daily CH4 flux relationship with thaw depth for the Core
sites. Lines show polynomial fits. Young: 3rd order polynomial,
R2 = 52%, RMSE = 9.7 mg CH4-C m�2 d�1. Medium: 5th order
polynomial, R2 = 97%, RMSE = mg CH4-C m�2 d�1. Old: 3rd
order polynomial, R2 = 82%, RMSE = 4.0 mg CH4-C m�2 d�1.
Fig. 8 Spatial relationship between landscape wetness and per-
iod-averaged CH4 flux. Landscape wetness is the proportion of
the surface that was saturated. The text inside each marker des-
ignates whether it was a Core site (‘C’) measured multiple peri-
ods over the season or whether it was a Spatial replicate (‘1’, ‘2’,
or ‘3’) measured only once. Error bars indicate standard error of
the mean.
Table 2 R2 values between normalized CO2 fluxes and remo-
tely sensed indices
NEE GPP ER
Landsat NDVI 73% 82% 76%
WV2 NDVI 77% 87% 86%
WV2 NDRE 80% 90% 84%
WV2, WorldView 2; NDRE, normalized difference red edge
index; NDVI, normalized difference vegetation index.
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
SPATIAL VARIATION IN ARCTIC CO2 AND CH4 FLUX 2861
times), while the other VTLB age classes showed more
moderate over- or underestimation (Medium VTLBs
2.6 9 higher sink, Old VTLBs 60% lower sink, Ancient
VTLBs a 2 9 magnitude source). Estimated cumulative
NEE from matrix tundra (Table 1) also overestimated
the regional sink by a factor of 6.5, averaging between
that of Young and Medium VTLBs. The standard devia-
tion in NEE from the regional mean for all individual
VTLBs of the region was 19 g C m�2, or almost 4 times
the regional average, and even higher when including
estimated variability in matrix tundra (22 g C m�2, or
4.5 times the regional average).
CH4 flux. We found good agreement between TCTw
and landscape wetness (R2 = 87%, linear relationship,
not shown), which translated into a strong relationship
between TCTw and period-average CH4 flux (R2 = 62%;
Fig. 9b). To our knowledge, there is no sufficient-reso-
lution satellite remote sensing metric for thaw depth.
Thus, we used the CH4 flux-TCTw relationship to scale
our results to the region (flux map shown in Fig. S8),
again substituting in cumulative CH4 flux estimates
(Fig. S6). Lakes were not included in the CH4 flux scal-
ing exercise since we have no sense of their spatial vari-
ability. We included Ancient VTLBs and matrix tundra
in the scaling exercise on account of the independence
of the CH4 flux-landscape wetness relationship from
the thaw lake cycle class. In addition, we measured two
Ancient VTLBs and a portion of matrix tundra in a pilot
study in 2010 using the same methods (data not
shown), and CH4 fluxes from these features fell well
within the landscape wetness relationship we show
here.
The average regional growing season CH4 flux esti-
mated from the scaling exercise was 2.1 � 0.2 (SE) g
CH4-C m�2. Carbon lost as CH4 flux offset 55% of the
carbon gained through photosynthesis, resulting in an
estimated net regional growing season carbon balance
of �2.8 � 2.6 (SE) g C m�2 from the scaling exercise.
The scaling exercise identified a moderate downward
trend in cumulative season CH4 flux from Young to
Ancient VTLBs of the region and also continued to
matrix tundra, mirroring an average decline in land-
scape wetness between these features at the regional
scale (Table 1). As in the 12 VTLBs we measured, how-
ever, the greatest variability was observed within the
same VTLB age class, especially for Young basins. The
standard deviation in CH4 flux from the regional aver-
age for individual VTLBs of the region was 0.6 g
C m�2, or 30% of the regional average, and the same
when including variability in estimated matrix tundra
CH4 flux.
Discussion
CO2 flux
Lakes. The period-averaged Lake CO2 flux rates we
measured (0.3–1.1 g CO2-C m�2 d�1) generally ranged
higher than reported in the few comparable studies
from Arctic Alaska (�0.1 to 0.7 g CO2-C m�2 d�1)
(Coyne & Kelley, 1974; Kling et al., 1992; Eugster et al.,
2003; McFadden et al., 2003), but lower than found in
Northern Siberia (1.4–2.1 g CO2-C m�2 d�1) (Abnizova
et al., 2012). An absence of spatial relationships between
NEE and lake characteristics was also seen in the large
spatial sampling of Alaskan Arctic lakes by Kling et al.
(1992). This may be due to the fact that neither we nor
the aforementioned study correlated fluxes with lake
hydrology. The vast majority of high latitude lakes are
(a)
(b)
Fig. 9 Relationships between satellite NDRE and net ecosystem
exchange of CO2 (NEE) (a), and TCTw and CH4 flux (b). NEE in
(a) is expressed as either the normalized flux (left y-axis, indicat-
ing the relative magnitude between the minimum and maxi-
mum modeled Core site fluxes) or the estimated cumulative
growing season NEE (right y-axis). CH4 flux in (b) is expressed
as either the period-averaged flux (left y-axis) or estimated
cumulative growing season CH4 flux (right y-axis). NDRE is the
Normalized Difference Red Edge index. TCTw is the Tasselled
Cap Transformation Wetness index. The text inside each marker
designates whether it was a Core site (‘C’) measured multiple
periods over the season or whether it was a Spatial replicate (‘1’,
‘2’, or ‘3’) measured only once. Error bars indicate standard
error of the cumulative fluxes.
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
2862 C. S . STURTEVANT & W. C. OECHEL

CO2 sources due to terrestrial carbon input from sur-
face runoff (Kling et al., 1992; Heikkinen et al., 2004;
Repo et al., 2007; Abnizova et al., 2012). Indeed, greater
lake flushing correlates with higher pCO2 (Sobek et al.,
2003; Tank et al., 2009). The lower emission from Lake 2
may have resulted from poor hydrological connectivity
with the landscape, as Lake 2 abuts the coast and does
not appear to be fed by any major surface flows.
VTLBs. The seasonal patterns and general magnitudes
of CO2 flux components we observed for the VTLB sites
were comparable to previous ecosystem-level studies in
arctic peatlands (e.g., Oechel et al., 1998a; Soegaard
et al., 2000; Vourlitis et al., 2000; Corradi et al., 2005;
Kwon et al., 2006; Loranty et al., 2011; Tagesson et al.,
2012). Our observations of declining productivity and
CO2 sink strength with VTLB age were also generally
consistent with the two previous regional flux studies
from these features (Zona et al., 2010; Zulueta et al.,
2011), although through the normalization and
increased sample size we show the trend to be much
clearer. Declining productivity with VTLB age is likely
a result of nutrients being locked up in slow decompos-
ing organic matter (Zona et al., 2010), as well as
increased ponding and associated strong CO2 source
(Coyne & Kelley, 1974; Abnizova et al., 2012). However,
there was also substantial variation within classes
observed in this and the two previous studies, exempli-
fied by the outliers we noted. However, aside from
Ancient 2 the CO2 fluxes from the outliers deviated by
only one age class, suggesting that other (unmeasured)
factors modify but generally do not replace the age
trend. As an example, the surprising productivity of
Ancient 2 might be caused by the drainage for Young 1
running directly through Ancient 2, possibly importing
nutrients and enhancing diffusion to roots via increased
soil water flow (Chapin et al., 1988; Oberbauer et al.,
1989; Rastetter et al., 2004).
It was surprising that large spatial variation in mois-
ture and thaw explained little spatial variation in CO2
flux components. Both these variables have a known
significant effect on decomposition rate in arctic soils
(e.g., Peterson et al., 1984; Oechel et al., 1998b; Olivas
et al., 2010) and greater wetness has been linked to
greater productivity in some studies (e.g., Oberbauer
et al., 1989; Sjogersten et al., 2006). We see two major
reasons for a lack of strong effect from either soil mois-
ture or thaw on spatial variation in NEE. First, the fac-
tors which influence ER (such as moisture and thaw)
can similarly affect GPP (Oberbauer et al., 2007; Olivas
et al., 2010). Second, the spatial variability in GPP was
very large, and since ER includes autotrophic respira-
tion to sustain plant biomass, variability in the auto-
trophic component of respiration (linked to GPP) likely
dominated the variability in ER. The strong relationship
we found between GPP and ER supports these link-
ages. Several other arctic studies have found that GPP
(or LAI as a proxy) controlled differences in NEE across
plant communities (Soegaard et al., 2000; McFadden
et al., 2003; Shaver et al., 2007; Lafleur et al., 2012). Thus,
the strong control of NEE variability by GPP is not
new, but we demonstrate that the ecosystem develop-
ment along the VTLB chronosequence largely drives
the spatial variability in GPP.
CH4 flux
The general flux rates, magnitude of seasonal variation,
and high spatial variability in CH4 emission we
observed from Young, Medium, and Old VTLBs were
consistent with previous arctic studies (e.g., Whalen &
Reeburgh, 1988; Vourlitis et al., 1993; Christensen et al.,
1995; Friborg et al., 2000; Sachs et al., 2008; Merbold
et al., 2009; von Fischer et al., 2010; Tagesson et al.,
2012). Surprising, however, was characterization of the
seasonal CH4 flux pattern only by thaw, which has been
found to limit emissions during the early season (e.g.,
Friborg et al., 2000; Tagesson et al., 2012) but to our
knowledge has not been explicitly linked to an early
peak and long decline. We suggest that the broad peak
in seasonal CH4 emission around 15–20 cm thaw could
reflect a lag in methanotrophy until higher soil methane
concentrations (Dedysh & Panikov, 1997; Segers, 1998;
Ambus & Petersen, 2005) or deeper thaw layers (Pani-
kov & Dedysh, 2000) were reached.
The strong control of spatial variability in CH4 flux
by landscape wetness and secondarily by relative thaw
is consistent with many smaller scale studies (e.g.,
Sebacher et al., 1986; Morrissey & Livingston, 1992;
Vourlitis et al., 1993; Christensen et al., 1995; Parmentier
et al., 2011). Water table position controls the volumes
of anaerobic and aerobic soil favorable for methane pro-
duction and oxidation, respectively (Bubier et al., 1993)
and thaw depth controls the total soil volume available
for decomposition. The most interesting aspect of this
result was the lack of relationship between CO2 and
CH4 fluxes as seen elsewhere (e.g., Whiting & Chanton,
1993; Tagesson et al., 2012), suggesting that even the
early stages of ecosystem development in this region
are not carbon limited.
Unfortunately, our limited resources prevented mea-
surement of spatial variation in Lake CH4 flux.
Although a few previous studies have found moder-
ately low lake CH4 emissions on the North Slope of
Alaska (Kling et al., 1992; Morrissey & Livingston,
1992), this is an important topic of future study, espe-
cially regarding the potential for large ebullitive CH4
emission (Walter et al., 2007).
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
SPATIAL VARIATION IN ARCTIC CO2 AND CH4 FLUX 2863

Regional estimates
The individual sites we measured along with our
scaling exercise suggest that single long term tower
measurements typically used to validate regional
models (e.g., Kimball et al., 2009) can be substantially
biased. Not only did we observe strong variability in
NEE among the classes of the thaw lake cycle (from
source to sink and an order of magnitude) but also
substantial variability within these classes (about
equal to the magnitude of the average flux). In addi-
tion, the high CO2 source of lakes resulted in a
strong (70%) reduction in the regional average CO2
sink, and should certainly be accounted for in regio-
nal estimates. Although bias in regional CH4 flux
estimates from single tower sites would not generally
be tied to the age sequence of VTLBs, variability in
wetness and associated CH4 flux across the landscape
due to the presence of any VTLB was quite strong.
Thus, any one tower site, although integrating the
variability within that specific ecosystem, will likely
not be representative of the regional average flux,
easily off by a factor 4 for NEE and 30% for CH4 flux
as indicated by our scaling exercise.
We think the information on spatial variability we
present here can improve regional growing carbon
exchange estimation for Alaskan Arctic coastal tundra
and possible many other arctic regions. We suggest that
accurate CO2 and CH4 flux scaling can be accomplished
with a few well-placed tower sites and satellite remote
sensing proxies for productivity and landscape wet-
ness, respectively, to calibrate the type of scaling rela-
tionships observed here for each growing season (and
region). Although the ecosystem succession along the
VTLB chronosequence was the mechanism largely
behind spatial variation in GPP (and hence NEE), we
did not need it to scale to the region because GPP (and
its proxies of NDRE or NDVI) reflected not only the age
trend but also deviations to that trend. Thus, although
the results presented here are specific to the Arctic
Coastal Plain of Alaska, the landscape-scale control of
productivity on NEE and wetness on CH4 flux likely
have wide applicability in regional estimation and scal-
ing across other arctic landscapes.
Finally, we recognize the limitations of this study.
Our measurements were confined to the growing sea-
son of 1 year. Therefore, more research on nonsummer
fluxes and interannual variability are needed. Future
study should also include variability within matrix tun-
dra as well as other landscape elements such as stream
systems (Kling et al., 1991) and areas of degrading per-
mafrost (Belshe et al., 2012) to test our assumptions that
the CO2 and CH4 flux scaling relationships extend
across these other landscape features.
Acknowledgements
We are grateful to our hard-working field assistants, Ryan Sher-man, and Timothy Hubbard. We thank LI-COR Biosciences forthe opportunity to use the newly developed LI-7700. We alsothank the three anonymous reviewers for their helpful sugges-tions to improve this manuscript. CH2M Hill Polar Services pro-vided logistic support. The WorldView 2 image was obtainedfrom the NSF-funded Polar Geospatial Data Center at the Uni-versity of Minnesota. Financial support was provided by the USDepartment of Energy (DOE) National Institute for ClimateChange Research (Grant No. DE-FC02-06ER64159), the NationalScience Foundation Graduate Research Fellowship (Grant No.DGE-0738622), the DOE Terrestrial Ecosystem Science program(Grant No. DE-SC005160), and NASA as part of the CARVEproject (JPL NASA Prime Contract Task Order 711140).
References
Abnizova A, Siemens J, Langer M, Boike J (2012) Small ponds with major impact: the
relevance of ponds and lakes in permafrost landscapes to carbon dioxide emis-
sions. Global Biogeochemical Cycles, 26, GB2041. doi: 10.1029/2011gb004237.
Adams JB, Gillespie AR (2006) Remote Sensing of Landscapes with Spectral Images: A
Physical Modeling Approach. Cambridge University Press, New York.
Ambus P, Petersen SO (2005) Oxidation of C-13-labeled methane in surface crusts of
pig-and cattle slurry. Isotopes in Environmental and Health Studies, 41, 125–133.
doi:10.1080/10256010500131783.
Baldocchi DD, Hicks BB, Meyers TP (1988) Measuring biosphere-atmosphere
exchanges of biologically related gases with micrometeorological methods. Ecol-
ogy, 69, 1331–1340.
Barnes EM, Clarke TR, Richards SE et al. (2000) Coincident detection of crop water
stress, nitrogen status and canopy density using ground-based multispectral data.
In: Proceedings of the 5th International Conference on Precision Agriculture (eds Robert
PC, Rust RH, Larson WE), pp. 1–15. ASA/CSSA/SSSA, Madison, WI, USA.
Belshe EF, Schuur EG, Bolker BM, Bracho R (2012) Incorporating spatial heterogene-
ity created by permafrost thaw into a landscape carbon estimate. Journal of
Geophysical Research-Biogeosciences, 117, G01026. doi: 10.1029/2011jg001836.
Billings WD, Peterson KM (1980) Vegetational change and ice-wedge polygons
through the thaw-lake cycle in Arctic Alaska. Arctic and Alpine Research, 12,
413–432.
Bockheim JG, Everett LR, Hinkel KM, Nelson FE, Brown J (1999) Soil organic carbon
storage and distribution in arctic tundra, Barrow, Alaska. Soil Science Society of
America Journal, 63, 934–940.
Brown J, Everett K, Webber P, MacLean S Jr, Murray D (1980) The coastal tundra at
Barrow. In: An Arctic Ecosystem: The Coastal Tundra at Barrow, Alaska (eds Brown J,
Miller PC, Tieszen LL, Bunnell FL), pp. 1–29. Hutchinson and Ross, Inc., Strouds-
burg, Pennsylvania, USA.
Bubier J, Costello A, Moore TR, Roulet NT, Savage K (1993) Microtopography and
methane flux in boreal peatlands, northern Ontario, Canada. Canadian Journal of
Botany/Revue Canadienne de Botanique, 71, 1056–1063.
Burba GG, McDermitt DK, Grelle A, Anderson DJ, Xu LK (2008) Addressing the
influence of instrument surface heat exchange on the measurements of CO2 flux
from open-path gas analyzers. Global Change Biology, 14, 1854–1876. doi:10.1111/j.
1365-2486.2008.01606.x.
Cabot EC (1947) The northern Alaskan coastal plain interpreted from aerial photo-
graphs. Geographical Review, 37, 639–648.
Carson CE (1968) Radiocarbon dating of lacustrine strands in Arctic Alaska. Arctic,
21, 12–26.
Chapin FS, Fetcher N, Kielland K, Everett KR, Linkins AE (1988) Productivity and
nutrient cycling of Alaskan tundra - Enhancement by flowing soil-water. Ecology,
69, 693–702.
Christensen TR, Jonasson S, Callaghan TV, Havstrom M (1995) Spatial variation in
high-latitude methane flux along a transect across Siberian and European tundra
environments. Journal of Geophysical Research-Atmospheres, 100, 21035–21045.
Corradi C, Kolle O, Walter K, Zimov SA, Schulze ED (2005) Carbon dioxide and
methane exchange of a north-east Siberian tussock tundra. Global Change Biology,
11, 1910–1925. doi:10.1111/j.1365-2486.2005.01023.x.
Coyne PI, Kelley JJ (1974) Carbon-dioxide partial pressures in arctic surface waters.
Limnology and Oceanography, 19, 928–938.
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
2864 C. S . STURTEVANT & W. C. OECHEL

Crist EP, Cicone RC (1984) A physically-based transformation of thematic mapper
data - the TM tasseled cap. Ieee Transactions on Geoscience and Remote Sensing, 22,
256–263. doi:10.1109/tgrs.1984.350619.
Dedysh SN, Panikov NS (1997) Effect of methane concentration on the rate of its oxi-
dation by bacteria in Sphagnum peat. Microbiology, 66, 470–475.
Eitel JUH, Vierling LA, Litvak ME et al. (2011) Broadband, red-edge information from
satellites improves early stress detection in a New Mexico conifer woodland.
Remote Sensing of Environment, 115, 3640–3646. doi:10.1016/j.rse.2011.09.002.
Eugster W, Kling G, Jonas T, McFadden JP, Wuest A, MacIntyre S, Chapin FS (2003)
CO2 exchange between air and water in an Arctic Alaskan and midlatitude Swiss
lake: importance of convective mixing. Journal of Geophysical Research-Atmospheres,
108, 4362. doi: 10.1029/2002jd002653.
Falge E, Baldocchi D, Olson R et al. (2001) Gap filling strategies for defensible annual
sums of net ecosystem exchange. Agricultural and Forest Meteorology, 107, 43–69.
von Fischer JC, Rhew RC, Ames GM, Fosdick BK, von Fischer PE(2010) Vegetation
height and other controls of spatial variability in methane emissions from the Arc-
tic coastal tundra at Barrow, Alaska. Journal of Geophysical Research, 115, G00I03,
doi: 10.1029/2009jg001283.
Foken T, Gockede M, Mauder M, Mahrt L, Amiro B, Munger W (2004) Post-field data
quality control. In: Handbook of Micrometeorology: A Guide For Surface Flux Measure-
ment And Analysis (eds Lee X, Massman WJ, Law BE), pp. 181–208. Kluwer
Academic Publishers, Dordrecht.
Friborg T, Christensen TR, Hansen BU, Nordstroem C, Soegaard H (2000) Trace gas
exchange in a high-arctic valley 2. Landscape CH4 fluxes measured and modeled
using eddy correlation data. Global Biogeochemical Cycles, 14, 715–723.
Heikkinen JEP, Virtanen T, Huttunen JT, Elsakov V, Martikainen PJ (2004) Carbon
balance in East European tundra. Global Biogeochemical Cycles, 18, GB1023. doi: 10.
1029/2003gb002054.
Hinkel KM, Eisner WR, Bockheim JG, Nelson FE, Peterson KM, Dai XY (2003) Spatial
extent, age, and carbon stocks in drained thaw lake basins on the Barrow Penin-
sula, Alaska. Arctic, Antarctic, and Alpine Research, 35, 291–300.
Hinzman LD, Bettex ND, Bolton WR et al. (2005) Evidence and implications of recent cli-
mate change innorthernAlaska and other arctic regions.Climatic Change, 72, 251–298.
Hsieh CI, Katul G, Chi T (2000) An approximate analytical model for footprint esti-
mation of scaler fluxes in thermally stratified atmospheric flows. Advances in Water
Resources, 23, 765–772.
IPCC (2007) Climate Change 2007: The Physical Science Basis. Cambridge University
Press, Cambridge, UK.
Jeffries MO, Morris K, Liston GE (1996) A method to determine lake depth and water
availability on the north slope of Alaska with spaceborne imaging radar and
numerical ice growth modelling. Arctic, 49, 367–374.
Kaimal JC, Wyngaard JC, Izumi Y, Cote R (1972) Spectral characteristics of surface-layer
turbulence.Quarterly Journal of the RoyalMeteorological Society, 98, 563–589.
Kimball JS, Jones LA, Zhang K, Heinsch FA, McDonald KC, Oechel WC (2009) A
satellite approach to estimate land-atmosphere CO2 exchange for boreal and arctic
biomes using MODIS and AMSR-E. Ieee Transactions on Geoscience and Remote Sens-
ing, 47, 569–587. doi: 10.1109/tgrs.2008.2003248.
Kling GW, Kipphut GW, Miller MC (1991) Arctic lakes and streams as gas conduits
to the atmosphere - implications for tundra carbon budgets. Science, 251, 298–301.
Kling GW, Kipphut GW, Miller MC (1992) The flux of CO2 and CH4 from lakes and
rivers in Arctic Alaska. Hydrobiologia, 240, 23–36.
Kwon HJ, Oechel WC, Zulueta RC, Hastings SJ (2006) Effects of climate variability on
carbon sequestration among adjacent wet sedge tundra and moist tussock tundra
ecosystems. Journal of Geophysical Research-Biogeosciences, 111, G03014. doi: 10.
1029/2005jg000036.
Lafleur PM, Humphreys ER, St Louis VL et al. (2012) Variation in peak growing sea-
son net ecosystem production across the Canadian Arctic. Environmental Science &
Technology, 46, 7971–7977. doi: 10.1021/es300500m.
Loranty MM, Goetz SJ, Rastetter EB, Rocha AV, Shaver GR, Humphreys ER, Lafleur
PM (2011) Scaling an instantaneous model of tundra NEE to the arctic landscape.
Ecosystems, 14, 76–93. doi:10.1007/s10021-010-9396-4.
McDermitt D, Burba G, Xu L et al. (2010) A new low-power, open-path instrument
for measuring methane flux by eddy covariance. Applied Physics B: Lasers and
Optics, 102, 391–405. doi: 10.1007/s00340-010-4307-0.
McFadden JP, Eugster W, Chapin FS (2003) A regional study of the controls on water
vapor and CO2 exchange in arctic tundra. Ecology, 84, 2762–2776.
McGuire AD, Christensen TR, Hayes D et al. (2012) An assessment of the carbon
balance of arctic tundra: comparisons among observations, process models, and
atmospheric inversions. Biogeosciences, 9, 3185–3204. doi:10.5194/bg-9-3185-2012.
McMichael CE, Hope AS, Stow DA, Fleming JB, Vourlitis G, Oechel W (1999)
Estimating CO2 exchange at two sites in arctic tundra ecosystems during the
growing season using a spectral vegetation index. International Journal of Remote
Sensing, 20, 683–698.
Merbold L, Kutsch WL, Corradi C et al. (2009) Artificial drainage and associated car-
bon fluxes (CO2/CH4) in a tundra ecosystem. Global Change Biology, 15, 2599–2614.
doi:10.1111/j.1365-2486.2009.01962.x.
Moore CJ (1986) Frequency-response corrections for eddy-correlation systems.
Boundary-Layer Meteorology, 37, 17–35.
Morrissey LA, Livingston GP (1992) Methane emissions from Alaska Arctic
tundra - an assessment of local spatial variability. Journal of Geophysical Research-
Atmospheres, 97, 16661–16670.
Oberbauer SF, Hastings SJ, Beyers JL, Oechel WC (1989) Comparative effects of
downslope water and nutrient movement on plant nutrition, photosynthesis, and
growth in Alaskan tundra. Holarctic Ecology, 12, 324–334.
Oberbauer SF, Tweedie CE, Welker J et al. (2007) Tundra CO2 fluxes in response to
experimental warming across latitudinal and moisture gradients. Ecological Mono-
graphs, 77, 221–238.
Oechel WC, Vourlitis GL, Hastings SJ, Bochkarev SA (1995) Change in arctic CO2 flux
over 2 decades - effects of climate-change at Barrow, Alaska. Ecological Applica-
tions, 5, 846–855.
Oechel WC, Vourlitis GL, Brooks S, Crawford TL, Dumas E (1998a) Intercomparison
among chamber, tower, and aircraft net CO2 and energy fluxes measured during
the Arctic System Science Land-Atmosphere-Ice Interactions (ARCSS-LAII) Flux
Study. Journal of Geophysical Research-Atmospheres, 103, 28993–29003.
Oechel WC, Vourlitis GL, Hastings SJ, Ault RP, Bryant P (1998b) The effects of water
table manipulation and elevated temperature on the net CO2 flux of wet sedge
tundra ecosystems. Global Change Biology, 4, 77–90.
Olivas PC, Oberbauer SF, Tweedie CE, Oechel WC, Kuchy A (2010) Responses of CO2
flux components of Alaskan Coastal Plain tundra to shifts in water table. Journal of
Geophysical Research-Biogeosciences, 115, G00I05, doi: 10.1029/2009jg001254.
Olivas PC, Oberbauer SF, Tweedie C, Oechel WC, Lin D, Kuchy A (2011) Effects of
fine-scale topography on CO2 flux components of Alaskan Coastal Plain tundra:
response to contrasting growing seasons. Arctic Antarctic and Alpine Research, 43,
256–266. doi: 10.1657/1938-4246-43.2.256.
Panikov NS, Dedysh SN (2000) Cold season CH4 and CO2 emission from boreal peat
bogs (West Siberia): winter fluxes and thaw activation dynamics. Global Biogeo-
chemical Cycles, 14, 1071–1080. doi:10.1029/1999gb900097.
Parmentier FJW, van Huissteden J, van der Molen MK, Schaepman-Strub G,
Karsanaev SA, Maximov TC, Dolman AJ (2011) Spatial and temporal dynamics in
eddy covariance observations of methane fluxes at a tundra site in northeastern
Siberia. Journal of Geophysical Research-Biogeosciences, 116, G03016. doi: 10.1029/
2010jg001637.
Peterson KM, Billings WD, Reynolds DN (1984) Influence of water-table and atmo-
spheric CO2 concentration on the carbon balance of arctic tundra. Arctic and Alpine
Research, 16, 331–335.
Rastetter EB, Kwiatkowski BL, Le Dizes S, Hobbie JE (2004) The role of down-slope
water and nutrient fluxes in the response of arctic hill slopes to climate change.
Biogeochemistry, 69, 37–62. doi: 10.1023/b:biog.0000031035.52498.21.
Repo ME, Huttunen JT, Naumov AV, Chichulin AV, Lapshina ED, Bleuten W, Marti-
kainen PJ (2007) Release of CO2 and CH4 from small wetland lakes in western
Siberia. Tellus Series B-Chemical and Physical Meteorology, 59, 788–796. doi: 10.1111/
j.1600-0889.2007.00301.x.
Rouse JW, Hass RH, Schell JA, Deering DW (1973) Monitoring vegetation
systems in the Great Plains with ERTS. 3rd ERTS Symposium, NASA SP-351 I, pp.
309–317.
Sachs T, Wille C, Boike J, Kutzbach L (2008) Environmental controls on ecosystem-scale
CH4 emission from polygonal tundra in the Lena River Delta, Siberia. Journal of
Geophysical Research-Biogeosciences, 113, G00a03. doi: 10.1029/2007jg000505.
Sebacher DI, Harriss RC, Bartlett KB, Sebacher SM, Grice SS (1986) Atmospheric
methane sources: Alaskan tundra bogs, an alpine fen, and a subarctic boreal
marsh. Tellus B, 38B, 1–10. doi: 10.1111/j.1600-0889.1986.tb00083.x.
Segers R (1998) Methane production and methane consumption: a review of pro-
cesses underlying wetland methane fluxes. Biogeochemistry, 41, 23–51. doi:10.1023/
a:1005929032764.
Sellman PV, Brown J, Lewellen RI, McKim H, Merry C (1975) The Classification and
Geomorhpic Implications of Thaw Lakes on the Arctic Coastal Plain, Alaska, Research
Report 344. U.S. Army Cold Regions Research and Engineering Laboratory, Hano-
ver, New Hampshire.
Shaver GR, Street LE, Rastetter EB, Van Wijk MT, Williams M (2007) Functional con-
vergence in regulation of net CO2 flux in heterogeneous tundra landscapes in
Alaska and Sweden. Journal of Ecology, 95, 802–817. doi:10.1111/j.1365-2745.2007.
01259.x.
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
SPATIAL VARIATION IN ARCTIC CO2 AND CH4 FLUX 2865

Shiklomanov NI, Streletskiy DA, Nelson FE et al. (2010) Decadal variations of active-
layer thickness in moisture-controlled landscapes, Barrow, Alaska. Journal of Geo-
physical Research-Biogeosciences, 115, G00i04. doi: 10.1029/2009jg001248.
Sjogersten S, van der Wal R, Woodin SJ (2006) Small-scale hydrological variation
determines landscape CO2 fluxes in the high Arctic. Biogeochemistry, 80, 205–216.
doi:10.1007/s10533-006-9018-6.
Sobek S, Algesten G, Bergstrom AK, Jansson M, Tranvik LJ (2003) The catchment and
climate regulation of pCO2 in boreal lakes. Global Change Biology, 9, 630–641.
doi:10.1046/j.1365-2486.2003.00619.x.
Soegaard H, Nordstroem C, Friborg T, Hansen BU, Christensen TR, Bay C (2000)
Trace gas exchange in a high-arctic valley 3. Integrating and scaling CO2 fluxes
from canopy to landscape using flux data, footprint modeling, and remote sensing.
Global Biogeochemical Cycles, 14, 725–744.
Sturm M, Racine C, Tape K (2001) Climate change - Increasing shrub abundance in
the Arctic. Nature, 411, 546–547.
Sturtevant CS, Oechel WC, Zona D, Kim Y, Emerson CE (2012) Soil moisture control
over autumn season methane flux, Arctic Coastal Plain of Alaska. Biogeosciences, 9,
1423–1440. doi:10.5194/bg-9-1423-2012.
Tagesson T, Molder M, Mastepanov M et al. (2012) Land-atmosphere exchange of
methane from soil thawing to soil freezing in a high-Arctic wet tundra ecosystem.
Global Change Biology, 18, 1928–1940. doi:10.1111/j.1365-2486.2012.02647.x.
Tank SE, Lesack LFW, Hesslein RH (2009) Northern delta lakes as summertime CO2
absorbers within the arctic landscape. Ecosystems, 12, 144–157. doi:10.1007/s10021-
008-9213-5.
Tarnocai C, Canadell JG, Schuur EAG, Kuhry P, Mazhitova G, Zimov S (2009) Soil
organic carbon pools in the northern circumpolar permafrost region. Global Biogeo-
chemical Cycles, 23, Gb2023. doi: 10.1029/2008gb003327.
Tilling AK, O’Leary GJ, Ferwerda JG, Jones SD, Fitzgerald GJ, Rodriguez D, Belford R
(2007) Remote sensing of nitrogen and water stress in wheat. Field Crops Research,
104, 77–85. doi:10.1016/j.fcr.2007.03.023.
Vourlitis GL, Oechel WC (1997) Landscape-scale CO2, H2O vapour and energy flux of
moist-wet coastal tundra ecosystems over two growing seasons. Journal of Ecology,
85, 575–590.
Vourlitis G, Oechel W, Hastings SJ, Jenkins M (1993) The effect of soil moisture and
thaw depth on CH4 flux from wet sedge tundra ecosystems on the North Slope of
Alaska. Chemosphere, 26, 329–337.
Vourlitis GL, Oechel W, Hope A et al. (2000) Physiological Models for Scaling Plot
Measurements of CO2 Flux Across an Arctic Tundra Landscape. Ecological Applica-
tions, 10, 60–72.
Vourlitis GL, Verfaillie J, Oechel W, Hope A, Stow D, Engstrom R (2003) Spatial
variation in regional CO2 exchange for the Kuparuk River Basin, Alaska over the
summer growing season. Global Change Biology, 9, 930–941.
Walter KM, Smith LC, Chapin FS (2007) Methane bubbling from northern lakes: pres-
ent and future contributions to the global methane budget. Philosphical Transactions
of the Royal Society A, 365, 1657–1676. doi: 10.1098/rsta.2007.2036.
Webb EK, Pearman GI, Leuning R (1980) Correction of flux measurements for density
effects due to heat and water-vapor transfer. Quarterly Journal of the Royal Meteoro-
logical Society, 106, 85–100.
Whalen SC, Reeburgh WS (1988) A methane flux time series for tundra environments.
Global Biogeochemical Cycles, 2, 399–409. doi:10.1029/GB002i004p00399.
Whiting GJ, Chanton JP (1993) Primary production control of methane emission from
wetlands. Nature, 364, 794–795.
Williams M, Eugster W, Rastetter EB, McFadden JP, Chapin FS III (2000) The controls
on net ecosystem productivity along an Arctic transect: a model comparison with
flux measurements. Global Change Biology, 6, 116–126.
Wilson K, Goldstein A, Falge E et al. (2002) Energy balance closure at FLUXNET sites.
Agricultural and Forest Meteorology, 113, 223–243.
Zimov SA, Schuur EAG, Chapin FS (2006) Permafrost and the global carbon budget.
Science, 312, 1612–1613. doi:10.1126/science.1128908.
Zona D, Oechel WC, Kochendorfer J et al. (2009) Methane fluxes during the initiation
of a large-scale water table manipulation experiment in the Alaskan Arctic tundra.
Global Biogeochemical Cycles, 23, GB2013, doi: 10.1029/2009GB003487.
Zona D, Oechel WC, Peterson KM, Clements RJ, Paw UKT, Ustin SL (2010) Character-
ization of the carbon fluxes of a vegetated drained lake basin chronosequence on
the Alaskan Arctic Coastal Plain. Global Change Biology, 16, 1870–1882. doi: 10.
1111/j.1365-2486.2009.02107.x.
Zulueta RC, Oechel WC, Loescher HW, Lawrence WT, Paw UKT (2011) Aircraft-
derived regional scale CO2 fluxes from vegetated drained thaw-lake basins and
interstitial tundra on the Arctic Coastal Plain of Alaska. Global Change Biology, 17,
2781–2802. doi: 10.1111/j.1365-2486.2011.02433.x.
Supporting Information
Additional Supporting Information may be found in theonline version of this article:
Appendix S1. Site coordinates and full measurement sche-dule.Appendix S2. Quality-controlled data coverage.Appendix S3. Seasonal NEE models for the Core sites.Appendix S4. A NEE normalization example.Appendix S5. Relationships used to estimate cumulativeseason fluxes for the Spatial sites.Appendix S6. 2011 regional flux maps derived from thescaling exercise.
© 2013 John Wiley & Sons Ltd, Global Change Biology, 19, 2853–2866
2866 C. S . STURTEVANT & W. C. OECHEL





























