Embed Size (px)
Citation preview

Contp. Biochem. Physiol., 1973, Vol. 46A, pp. 207 to 213. Pwgamon Press. Printed in Great Britain
SOME ALTERATIONS IN HEMOLYMPH PROTEINS OF HEMIGRAPSUS NUDUS ASSOCIATED WITH
TEMPERATURE ACCLIMATION*
RALPH I. PETERS, JR. and GARRELL E. LONG
Department of Zoology, Washington State University, Pullman, Washington 99163, U.S.A.
(Received 11 Se#tember 1972)
Abstract-l. The purple shore crab, Hemigrapsus nudus, was exposed to environmental temperatures 10 and 2O”C, and the time course of changes in hemolymph proteins and clotting time was recorded.
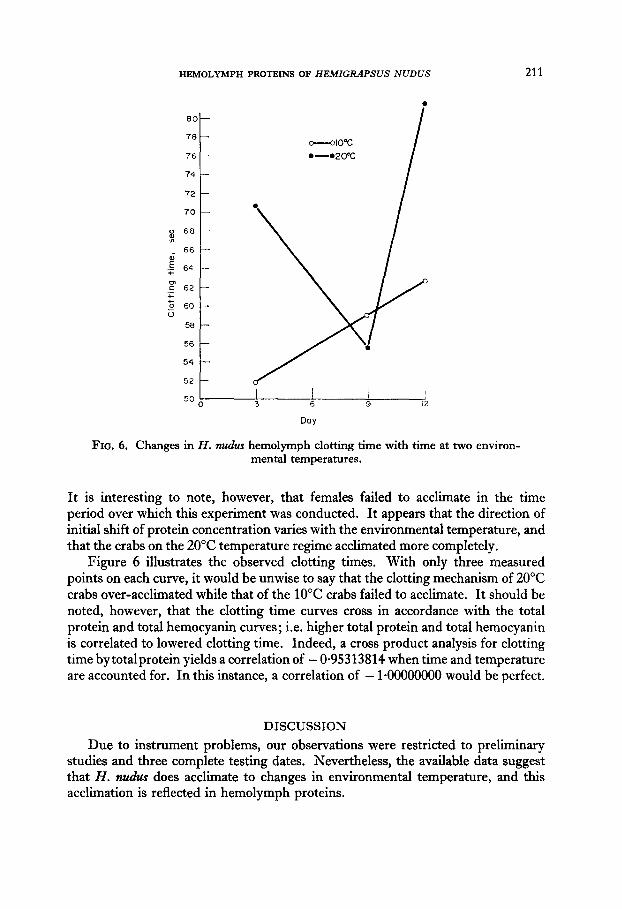
2. Results indicate that this animal exhibits responses to abrupt changes in environmental temperature characteristic of acclimation. Initial shifts of total hemocyanin concentrations of up to 54 per cent of initial value were followed by a return to within 2 per cent.
3. Total hemolymph protein and hemolymph clotting time exhibited similar responses, with a correlation between these two variables of - 0.95313814.
INTRODUCTION
SEPARATION and identification of blood proteins have been established as a useful tool in dealing with vertebrates in establishing or confirming phylogenetic relation- ships, and in providing an indication of the physiologic status of an animal, Attempts to confirm invertebrate phylogenetic relationships by comparison of hemolymph proteins have been largely unsuccessful (Manwell & Baker, 1963; Horn & Kerr, 1969). Recent work, however, indicates that the events associated with the female reproductive cycle (Kerr, 1969), and certain types of parasitization of crustaceans may be reflected in the hemolymph of an individual (Manwell & Baker, 1963). Differences in crab hemolymph with different environmental temperatures have been noted, as well as differences in hemolymph clotting time of crabs acclimated to different temperatures (Dean, 1966). To the best of our knowledge, there is no record of the time course of hemolymph protein alterations with acclimation to temperature, and no successful correlation of specific hemolymph protein altera- tions with clotting time changes. Such a correlation would greatly help to elucidate the mechanism of hemolymph coagulation.
Our intent is to illustrate: (1) whether or not the purple shore crab, Hemigrupsus nudus, exhibits differences in clotting time and hemolymph proteins with differences in environmental temperature, thus confirming previous observations and extending
* This work was partially supported by a National Institutes of Health Grant (No. GiV-01276).
207

208 RALPH I. PETERS, JR. AND GARRELL E. LONG
the phenomenon to another species; (2) the time course of these changes if they occur; (3) any variations in the direction or amount of change with the sex of the animal; (4) any correlation between the amount of a specific protein constituent and the clotting time of the hemolymph.
Previous investigators have noted an immense individual variation in crab hemolymph and for this reason it was decided to pool hemolymph samples from three individuals on one cellulose acetate strip. Individual *variation has been attributed to the sex of the individual (Horn & Kerr, 1963) ; if female, the stage of the reproductive cycle (Kerr, 1969); parasitization (Manwell & Baker, 1963); and/or the stage of the molt cycle of the individual (Passano, 1960; Laufer & McNamara, 1962).
In order to standardize sex-induced differences in hemolymph proteins, crabs from each environment were pooled according to sex. Random selection of individuals to be analyzed, coupled with the pooling procedure, was the most practicable means for standardizing variations due to parasitization and reproductive cycle differences. The crustacean molt cycle was a difficult variable to deal with, as many metabolic changes associated with pre- and post-molt are not displayed externally. It has been reported that a DD photoperiod arrests molt in some crustaceans (Passano, 1960). We utilized constant darkness environments in an attempt to arrest molting, and relied upon random sampling and sample pooling to standardize differences.
Most previous hemolymph protein separations have been accomplished by starch gel electrophoresis. However, we decided upon cellulose acetate electro- phoresis because of the relative speed, ease and accuracy it affords.
MATERIALS AND METHODS Approximately 500 specimens of Hemigrapsus nudus were collected from the Pacific
coast near Anacortes, Washington. Immediately after collection they were transported to our laboratory in Pullman and split into two populations, one being put into a 10°C controlled temperature chamber and the other into a 20°C chamber. Both populations were kept on a DD photoperiod. The animals were kept in sea water collected from their natural habitat and were not fed during the experiment to minimize any effects of temperature on appetite.
On days 3, 9 and 12, six males and six females were randomly selected from each temperature chamber and hemolymph was drawn by cardiac puncture.
The first drop of hemolymph was placed immediately on a clean glass slide and clotting time determined as follows: the slide was tipped once each 10 set, and clot formation was indicated when tipping of the slide resulted in no conformational change in the drop.
The remainder of the hemolymph was simultaneously mixed with 12.5 y0 sodium citrate as an anticoagulant (Stutman & Dolliver, 1968), and two drops from each of three crabs from the same temperature and of the same sex were pooled.
The pooled samples were centrifuged at high speed for 20 min and 4 ~1 of plasma from each pool was placed on a cellulose acetate strip (Sepraphore III, Gelman Instrument Co., Box 1449, Ann Arbor, Michigan 48106) after preliminary soaking of the strip in buffer. The strips were then electrophoresed at 220 V, 2 ma/strip for 30 min in 0.05 M Tris- barbital-sodium-barbital buffer (H. R. Buffer, Gelman Instrument Co., Box 1449, Ann Arbor, Michigan 48106) at pH 8.8. The strips were then stained for protein in a con- centrated solution of Ponceau S for 10 min. De-staining was accomplished by 10-r& baths

HRMOLYMPH PROTEINS OF HEMIGIUPSUS NUDUS 209
in three successive tanks of 10% acetic acid, and clearing was accomplished by a 2-min bath in fresh 10% acetic acid/ethanol followed by 10 min in an 80°C oven. The cleared strips were scanned with a densitometer and the curves integrated with a planimeter.
RESULTS
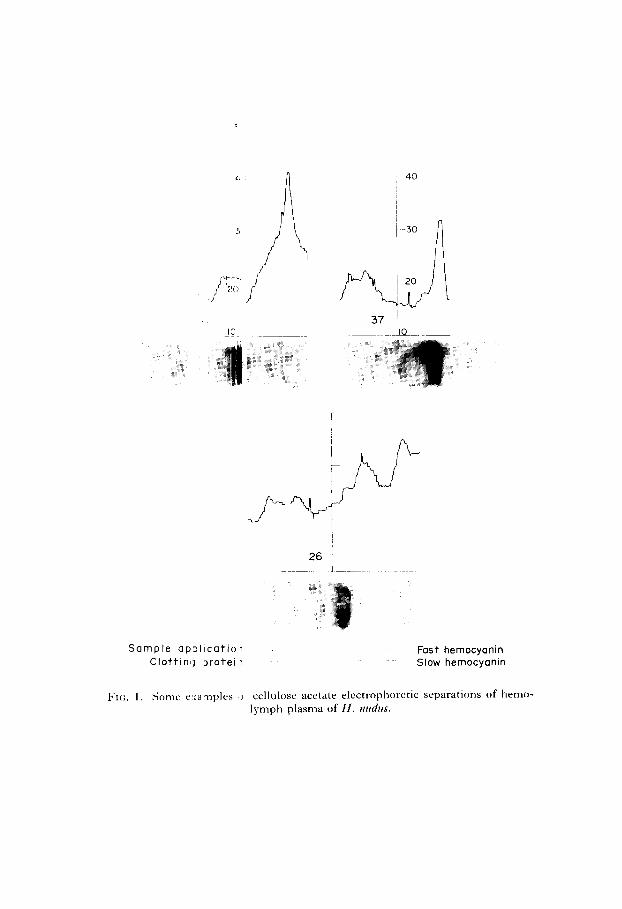
Several separations and their respective recordings are shown in Fig. 1. We found no published figures of cellulose acetate separations of hemolymph of H. 1zudus, and since time limitations prevented us from analyzing the observed bands, our designations of “clotting protein”, “slow hemocyanin” and “fast hemocyanin” may be questioned. It should be noted, however, that what we call “clotting protein” was not observed in sera in preliminary experiments and exhibits approximately the same mobility as the clotting protein observed by others (Horn & Kerr, 1963, 1969; Manwell & Baker, 1963). Likewise, our labeling of the two major bands as “slow hemocyanin” and “fast hemocyanin” is based on the resemblance between their mobility and the mobility of bands identified as hemocyanin by others (Horn & Kerr, 1963, 1969; Manwell SC Baker, 1963). As is evident in Fig. 1, fast and slow hemocyanin did not always separate completely enough for our densitometer to note a distinct line of demarcation. Thus, it is not possible to draw any accurate conclusions based on either of these fractions. Rather, total hemocyanin must be used in the analysis of our findings.
Indeed, total hemocyanin may be the most valuable variable in drawing conclusions from this experiment, as the occurrence of recognizable bands of clotting protein was so unpredictable. Only 62 per cent of the cold-acclimated crabs exhibited plasma containing detectable amounts of clotting protein, and warm- acclimated crabs were only slightly better, with 75 per cent of the individuals in this category. The concentration differences between the two temperature regimes were not appreciable. Detectable quantities of clotting protein appeared in 67 per cent of the males tested and 69 per cent of the females tested. The females showed slightly greater quantities of clotting protein, but the difference was not notable. An observation not completely explicable, but worth noting, is the increase in numbers of individuals exhibiting clotting protein throughout the experiment. On day 3, only 50 per cent of the individuals showed detectable amounts of clotting protein. This percentage rose to 78 per cent on day 9 and 83 per cent on day 12. Whether this increase was due to true metabolic adjustment, possibly a stress response to the sudden shift of environments, or merely to selection pressures eliminating individuals without this protein early in captivity, remains to be seen. The clotting protein concentrations varied so widely and apparently without pattern that no conclusions can safely be drawn.
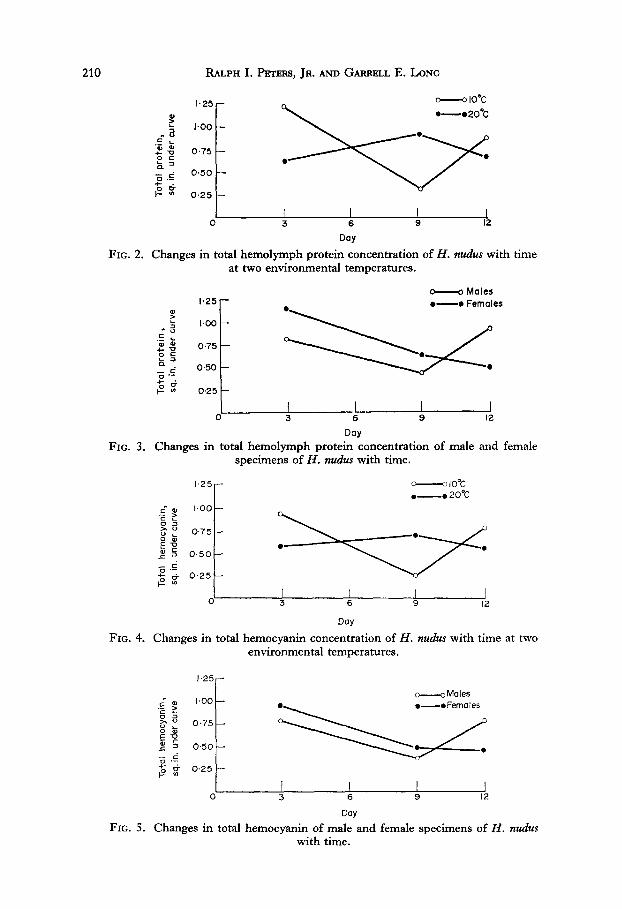
Figures 2-6 graphically represent our findings. Figures 2 and 4 indicate that cold-acclimated specimens of H. nudes appear to have more hemolymph protein than warm-acclimated individuals. This is in direct contradiction to Dean’s findings on Ucu pugihtor (Dean & Vernberg, 1966). Initially, females showed higher protein concentrations than males, in agreement with Horn & Kerr (1963).
210 RALPH I. F%TJW, JR. AND GARRELL E. LONG
$i! i
0.25 -
I I I 1 0 3 6 9 12 Day
FIG. 2. Changes in total hemolymph protein concentration of H. nudus with time at two environmental temperatures.
^ Males
I I I I I 0 3 6 9 12
Day FIG. 3. Changes in total hemolymph protein concentration of male and female
specimens of H. nudus with time.
I.25 r ---loot ,-_. 20%
FIG. 4. Changes in total hemocyanin concentration of H. nudus with time at two environmental temperatures.
-Males 2 m I.00 ‘E ? g: zs sz z .’
11’:’ 0-7
p:: 0.25 -
I I I 0 3 6 9 12
Day FIG. 5. Changes in total hemocyanin of male and female specimens of H. nudus
with time.
40
Sample ap:llicatioi Clottinlj 2rotel7
I
26 j
Fast hemocyanin Slow hemocyanin
cellulose acetate electrophoretic separations of hemo- lymph plasma of H. nztdzrs.


HEMOLYMPH PROTEINS OF HEMIGRAPSUS NUDUS 213
LAUFER H. & MCNAMARA T. (1962) Blood protein changes in crustacea. Biol. Bull., mar. biol. Lab., Woods Hole 123, 519.
MANWELL C. 8z BAKER C. M. A. (1963) Starch gel electrophoresis of sera from some marine arthropods: studies on the heterogeneity of hemocyanin and on a “ceruloplasmin-like protein”. Camp. Biochem. Physiol. 8, 193-208.
PASSANO L. M. (1960) Molting and its control. In The Physiology of Crustacea (Edited by WATERMAN T. H.), pp. 473-536. Academic Press, New York.
ROBERT M. & GRAY I. (1972) Enzymatic mechanisms during temperature acclimation of the blue crab, Callinectes sapidus-I. Oxygen consumption and activity of glucose-6- phosphodehydrogenase and 6-phosphogluconate dehydrogenase. Camp. Biochem. Physiol. 42B, 377-387.
STUTMAN L. J. & DOLLIVER M. (1968) Mechanism of coagulation in Gecarcinus lateralis. Am. 2001. 8, 481-489.
WOOD P. J. & KARPAWICH P. P. (1972) Studies on the coagulation process in the crayfish: attempts to identify a fibrinogen-like factor. Camp. Biochem. Physiol. 42B, 41-48.
Key Word Index-Hemigrapsus nudus; acclimation; hemolymph; blood proteins; blood clotting; coagulation; electrophoresis.





























