Embed Size (px)
Citation preview

doi:10.1006/jmbi.1999.3490 available online at http://www.idealibrary.com on J. Mol. Biol. (2000) 296, 851±861
Solution Structure of an 11-mer Duplex Containingthe 3,N4-ethenocytosine Adduct Opposite20-Deoxycytidine: Implications for the Recognitionof Exocyclic Lesions by DNA Glycosylases
David Cullinan, Francis Johnson and Carlos de los Santos*
Department of PharmacologicalSciences, State University ofNew York at Stony BrookStony Brook, NY 11794-8651, USA
E-mail address of the [email protected].
Abbreviations used: TSP, (2,2,3,3trimethylsilyl-propionate; NOESY,effect spectroscopy; COSY, correlatDQF-COSY, double quantum ®lterspectroscopy; HOHAHA, homonucspectroscopy; NOE, nuclear OverhHeteronuclear multiple quantum cspectroscopy; HPLC, high performchromatography; ppm, parts per metheno-20-deoxycytidine; dC, 20-deodeoxyadenosine; dG, 20-deoxyguanMD, molecular dynamics; RMSD, rdeviation; AAG, alkyladenine DNAmismatch-speci®c uracil glycosylasmismatch-speci®c thymine-DNA g
0022-2836/00/030851±11 $35.00/0
Lipid peroxidation products, as well as the metabolic products of vinylchloride, react with cellular DNA producing the mutagenic adduct 3,N4-etheno-20-deoxycytidine (edC), along with several other exocyclic deriva-tives. High-resolution NMR spectroscopy and restrained moleculardynamics simulations were used to establish the solution structure of an11-mer duplex containing an edC �dC base-pair at its center. The NMRdata suggested a regular right-handed helical structure having all resi-dues in the anti orientation around the glycosydic torsion angle and Wat-son-Crick alignments for all canonical base-pairs of the duplex.Restrained molecular dynamics generated a three-dimensional model inexcellent agreement with the spectroscopic data. The (edC �dC)-duplexstructure is a regular right-handed helix with a slight bend at the lesionsite and no severe distortions of the sugar-phosphate backbone. The edCadduct and its partner dC were displaced towards opposite grooves ofthe helix, resulting in a lesion-containing base-pair that was highlysheared but stabilized to some degree by the formation of a single hydro-gen bond. Such a sheared base-pair alignment at the lesion site was pre-viously observed for edC �dG and edC �T duplexes, and was also presentin the crystal structures of duplexes containing dG �T and dG �U mis-matches. These observations suggest the existence of a substrate struc-tural motif that may be recognized by speci®c DNA glycosylases duringthe process of base excision repair.
# 2000 Academic Press
Keywords: exocyclic DNA adducts; DNA structure; NMR spectroscopy;DNA recognition, MUG glycosylase
*Corresponding authoring author:
-d4) sodium 3-nuclear Overhauserion spectroscopy;ed correlationlear Hartmann-Hahn
auser effect; HMQC,orrelationance liquidillion; edC, 3,N4-xycytidine; dA, 20-osine; T, thymidine;oot mean square
glycosylase; MUG,e; hTDG, humanlycosylase.
Introduction
Since the mid-1970's, it has been known thatexposure to vinyl chloride, used in the manufac-ture of synthetic polymers, may cause human liversarcoma (Creech & Johnson 1974; Maltoni et al.,1974). It was later shown that vinyl chloride andrelated compounds are metabolized by the hepaticP450 system to produce 2-chlorooxirene and itsrearrangement product chloroacetaldehyde(Guengerich et al., 1979; Barbin et al., 1985; Schereret al., 1986). These bifunctional alkylating agentsreact with cellular DNA to produce several exocyc-lic lesions, including 3,N4-etheno-20-deoxycytidine(edC) (Barbin & Bartsch, 1986; Kusmierik & Singer,1992). Etheno, adducts also can be formed by thereaction of lipid peroxidation products withnucleotides (el Ghissassi et al., 1995; Cheng &
# 2000 Academic Press

Figure 1. Chemical structure of 3,N4-etheno-20-deoxy-cytidine; edC �dC duplex sequence and numberingscheme.
852 Structure and Recognition of Exocyclic DNA Lesions
Chung, 1996) and edC has been shown to be per-sistently present in human and rat liver DNA(Nair et al., 1995).
The mutagenic properties of edC have beeninvestigated in several laboratories. In vitro replica-tion of an edC-containing template using the Kle-now fragment of DNA polymerase I showedprimarily dA and T being incorporated oppositethe adduct (Singer & Spengler, 1986; Simha et al.,1991; Zhang et al., 1995a). Transformation ofEscherichia coli with vectors containing edC showedthat the lesion is weakly mutagenic in bacteria (2 %mutation frequency), with dA and T being themost commonly misincorporated bases (Palejwalaet al., 1991, 1993; Basu et al., 1993; Moriya et al.,1994). In vitro studies using mammalian poly-merases a and d showed that dA and T incorpor-ation predominates, while polymerase bincorporates primarily dC (Shibutani et al., 1996).Studies in mammalian cells revealed that edC ishighly mutagenic, with T and dA incorporationbeing most common (Moriya et al., 1994).
Enzymes that repair edC lesions in E. coli andhuman cells have been recently identi®ed(Saparbaev & Laval, 1998; Hang et al., 1998). Thesestudies found that the bacterial double-strandeduracil-DNA glycosylase (dsUDG), also known asmismatch speci®c uracil glycosylase (MUG), andthe human mismatch-speci®c thymine-DNA glyco-sylase (hTDG) remove edC from double-strandedDNA. In the case of the bacterial enzyme, removalof the edC DNA adducts is more ef®cient thanexcision of uracil from G �U mismatch-containingsubstrates. Furthermore, adduct excision rates aresimilar for DNA substrates differing only in theresidue positioned opposite the lesion. The crystalstructure of MUG complexed with DNA contain-ing a G �U mismatch has been reported (Barrettet al., 1998). The excised uracil residue was not Pre-sent in the crystal but the conformation of theDNA backbone suggested that MUG had extrudedit into its active-site pocket during the catalyticprocess. The depth of the pocket was almost twicethe size of a pyridine ring but speci®c contactsbetween the protein and the extruded uracil wereuncertain. The stability of the complex wasincreased by the insertion of two amino acid resi-dues in the cavity created by the everted residue.Both amino acids entered the duplex from theminor groove of the helix and formed hydrogenbonds with the unpaired guanine.
A thorough study of the thermodynamic proper-ties of edC has recently been performed (Gelfandet al., 1998). The presence of the lesion at the centerof a 13-mer duplex of DNA caused an average70 % loss in the Gibb's free energy, as compared tothe same duplex with an unmodi®ed base-pair.The signi®cant destabilization brought about bythe lesion was found to be mostly independent ofthe sequence context and of the base positionedopposite the lesion. However, the enthalpy wasshown to depend more on both of these factors.
For the sequence used in our study, the edC �dCduplex showed the smallest decrease in enthalpy.
We have reported previously the NMR struc-tures of DNA duplexes containing edC oppositedA, T, and dG (Korobka et al., 1996; Cullinan et al.,1996, 1997). A common characteristic of theseduplexes is the incorporation of edC inside quiteregular helical structures, which showed pertur-bation only at the lesion site. The edC �dA base-pair alignment is staggered, facilitating stackingbetween the edC and dA residues. By contrast, theedC �dG and edC �T alignments are co-planar andstabilized to some extent by hydrogen bonds.Using identical sequence context, we describe herethe determination of the edC �dC duplex structureby NMR spectroscopy and MD simulations. Inaddition, we report the results of a comparisonbetween the edC �dC duplex and the other mem-bers of the edC family as well as the dG �T mis-match duplex, supporting the idea of a substraterecognition motif for MUG and hTDG. The chemi-cal structure of edC and the sequence of the duplexused in our study are shown in Figure 1.
Results
NMR spectra of the eeedC �dC duplex
The one-dimensional proton spectra of theedC �dC duplex in 2H2O buffer (pH 6.8) 15 �C andin H2O buffer at 5 �C are characterized by the pre-sence of sharp signals and the absence of minor res-onances (Supplementary Material). Assignment ofthe base and sugar protons is based on the analysisof NOESY and COSY spectra following standardprocedures (Hare et al., 1983; WuÈ thrich, 1986). Asan example, Figure 2 shows contour plots of aregion of a NOESY spectrum (250 ms mixing time)recorded in 2H2O buffer (pH 6.8) 15 �C. NOE cross-peaks between the base protons and its own and
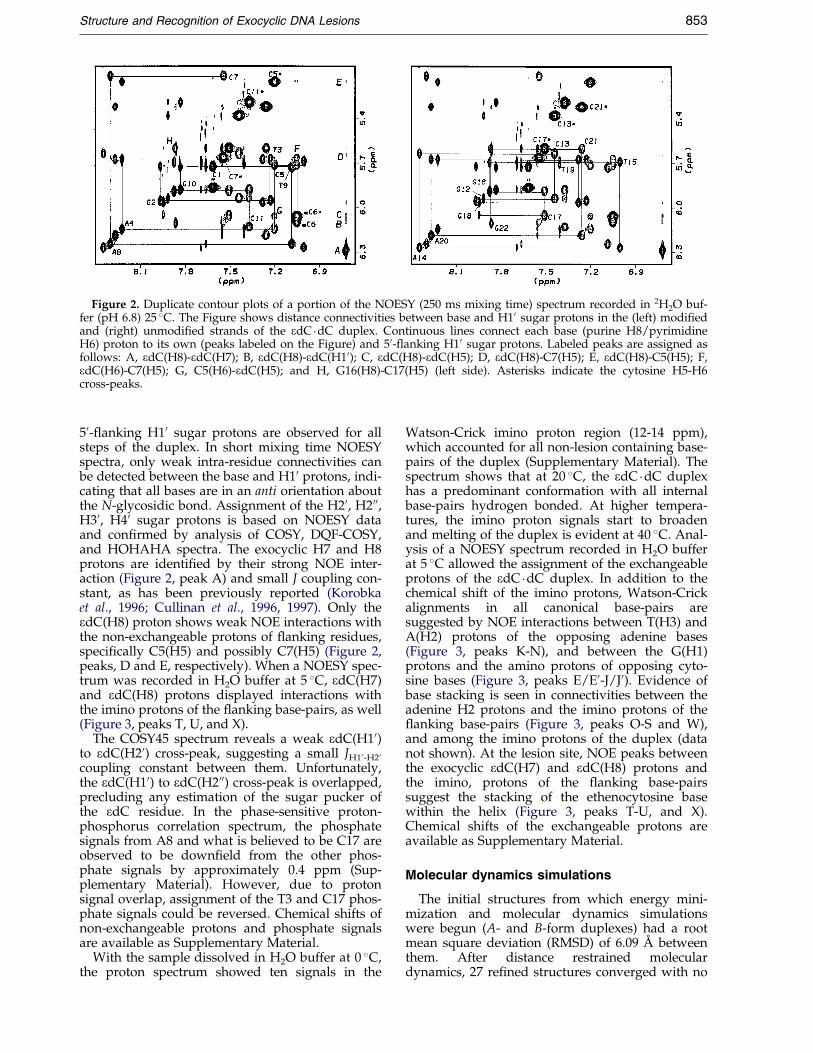
Figure 2. Duplicate contour plots of a portion of the NOESY (250 ms mixing time) spectrum recorded in 2H2O buf-fer (pH 6.8) 25 �C. The Figure shows distance connectivities between base and H10 sugar protons in the (left) modi®edand (right) unmodi®ed strands of the edC �dC duplex. Continuous lines connect each base (purine H8/pyrimidineH6) proton to its own (peaks labeled on the Figure) and 50-¯anking H10 sugar protons. Labeled peaks are assigned asfollows: A, edC(H8)-edC(H7); B, edC(H8)-edC(H10); C, edC(H8)-edC(H5); D, edC(H8)-C7(H5); E, edC(H8)-C5(H5); F,edC(H6)-C7(H5); G, C5(H6)-edC(H5); and H, G16(H8)-C17(H5) (left side). Asterisks indicate the cytosine H5-H6cross-peaks.
Structure and Recognition of Exocyclic DNA Lesions 853
50-¯anking H10 sugar protons are observed for allsteps of the duplex. In short mixing time NOESYspectra, only weak intra-residue connectivities canbe detected between the base and H10 protons, indi-cating that all bases are in an anti orientation aboutthe N-glycosidic bond. Assignment of the H20, H200,H30, H40 sugar protons is based on NOESY dataand con®rmed by analysis of COSY, DQF-COSY,and HOHAHA spectra. The exocyclic H7 and H8protons are identi®ed by their strong NOE inter-action (Figure 2, peak A) and small J coupling con-stant, as has been previously reported (Korobkaet al., 1996; Cullinan et al., 1996, 1997). Only theedC(H8) proton shows weak NOE interactions withthe non-exchangeable protons of ¯anking residues,speci®cally C5(H5) and possibly C7(H5) (Figure 2,peaks, D and E, respectively). When a NOESY spec-trum was recorded in H2O buffer at 5 �C, edC(H7)and edC(H8) protons displayed interactions withthe imino protons of the ¯anking base-pairs, as well(Figure 3, peaks T, U, and X).
The COSY45 spectrum reveals a weak edC(H10)to edC(H20) cross-peak, suggesting a small JH10-H20
coupling constant between them. Unfortunately,the edC(H10) to edC(H200) cross-peak is overlapped,precluding any estimation of the sugar pucker ofthe edC residue. In the phase-sensitive proton-phosphorus correlation spectrum, the phosphatesignals from A8 and what is believed to be C17 areobserved to be down®eld from the other phos-phate signals by approximately 0.4 ppm (Sup-plementary Material). However, due to protonsignal overlap, assignment of the T3 and C17 phos-phate signals could be reversed. Chemical shifts ofnon-exchangeable protons and phosphate signalsare available as Supplementary Material.
With the sample dissolved in H2O buffer at 0 �C,the proton spectrum showed ten signals in the
Watson-Crick imino proton region (12-14 ppm),which accounted for all non-lesion containing base-pairs of the duplex (Supplementary Material). Thespectrum shows that at 20 �C, the edC �dC duplexhas a predominant conformation with all internalbase-pairs hydrogen bonded. At higher tempera-tures, the imino proton signals start to broadenand melting of the duplex is evident at 40 �C. Anal-ysis of a NOESY spectrum recorded in H2O bufferat 5 �C allowed the assignment of the exchangeableprotons of the edC �dC duplex. In addition to thechemical shift of the imino protons, Watson-Crickalignments in all canonical base-pairs aresuggested by NOE interactions between T(H3) andA(H2) protons of the opposing adenine bases(Figure 3, peaks K-N), and between the G(H1)protons and the amino protons of opposing cyto-sine bases (Figure 3, peaks E/E0-J/J0). Evidence ofbase stacking is seen in connectivities between theadenine H2 protons and the imino protons of the¯anking base-pairs (Figure 3, peaks O-S and W),and among the imino protons of the duplex (datanot shown). At the lesion site, NOE peaks betweenthe exocyclic edC(H7) and edC(H8) protons andthe imino, protons of the ¯anking base-pairssuggest the stacking of the ethenocytosine basewithin the helix (Figure 3, peaks T-U, and X).Chemical shifts of the exchangeable protons areavailable as Supplementary Material.
Molecular dynamics simulations
The initial structures from which energy mini-mization and molecular dynamics simulationswere begun (A- and B-form duplexes) had a rootmean square deviation (RMSD) of 6.09 AÊ betweenthem. After distance restrained moleculardynamics, 27 re®ned structures converged with no
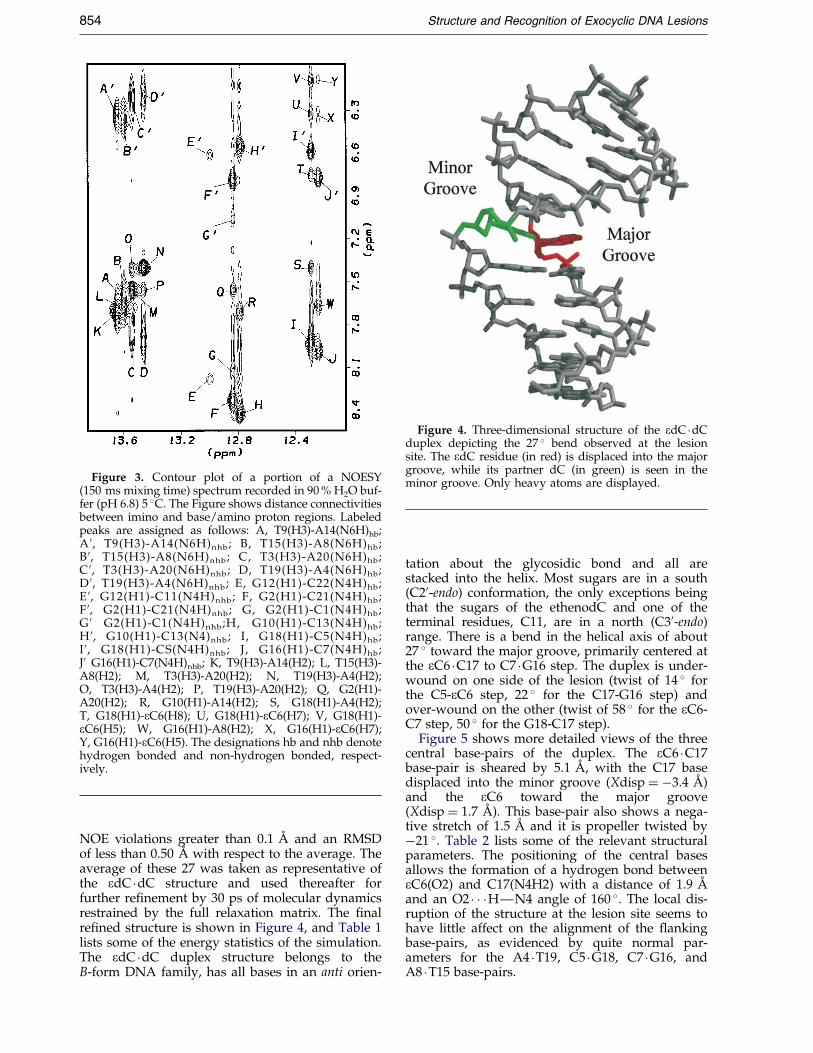
Figure 3. Contour plot of a portion of a NOESY(150 ms mixing time) spectrum recorded in 90 % H2O buf-fer (pH 6.8) 5 �C. The Figure shows distance connectivitiesbetween imino and base/amino proton regions. Labeledpeaks are assigned as follows: A, T9(H3)-A14(N6H)hb;A 0, T9(H3)-A14(N6H)nhb; B, T15(H3)-A8(N6H)hb;B 0, T15(H3)-A8(N6H)nhb; C, T3(H3)-A20(N6H)hb;C 0, T3(H3)-A20(N6H)nhb; D, T19(H3)-A4(N6H)hb;D 0, T19(H3)-A4(N6H)nhb; E, G12(H1)-C22(N4H)hb;E 0, G12(H1)-C11(N4H)nhb; F, G2(H1)-C21(N4H)hb;F 0, G2(H1)-C21(N4H)nhb; G, G2(H1)-C1(N4H)hb;G 0 G2(H1)-C1(N4H)nhb;H, G10(H1)-C13(N4H)hb;H 0, G10(H1)-C13(N4)nhb; I, G18(H1)-C5(N4H)hb;I 0, G18(H1)-CS(N4H)nhb; J, G16(H1)-C7(N4H)hb;J0 G16(H1)-C7(N4H)nhb; K, T9(H3)-A14(H2); L, T15(H3)-A8(H2); M, T3(H3)-A20(H2); N, T19(H3)-A4(H2);O, T3(H3)-A4(H2); P, T19(H3)-A20(H2); Q, G2(H1)-A20(H2); R, G10(H1)-A14(H2); S, G18(H1)-A4(H2);T, G18(H1)-eC6(H8); U, G18(H1)-eC6(H7); V, G18(H1)-eC6(H5); W, G16(H1)-A8(H2); X, G16(H1)-eC6(H7);Y, G16(H1)-eC6(H5). The designations hb and nhb denotehydrogen bonded and non-hydrogen bonded, respect-ively.
Figure 4. Three-dimensional structure of the edC �dCduplex depicting the 27 � bend observed at the lesionsite. The edC residue (in red) is displaced into the majorgroove, while its partner dC (in green) is seen in theminor groove. Only heavy atoms are displayed.
854 Structure and Recognition of Exocyclic DNA Lesions
NOE violations greater than 0.1 AÊ and an RMSDof less than 0.50 AÊ with respect to the average. Theaverage of these 27 was taken as representative ofthe edC �dC structure and used thereafter forfurther re®nement by 30 ps of molecular dynamicsrestrained by the full relaxation matrix. The ®nalre®ned structure is shown in Figure 4, and Table 1lists some of the energy statistics of the simulation.The edC �dC duplex structure belongs to theB-form DNA family, has all bases in an anti orien-
tation about the glycosidic bond and all arestacked into the helix. Most sugars are in a south(C20-endo) conformation, the only exceptions beingthat the sugars of the ethenodC and one of theterminal residues, C11, are in a north (C30-endo)range. There is a bend in the helical axis of about27 � toward the major groove, primarily centered atthe eC6 �C17 to C7 �G16 step. The duplex is under-wound on one side of the lesion (twist of 14 � forthe C5-eC6 step, 22 � for the C17-G16 step) andover-wound on the other (twist of 58 � for the eC6-C7 step, 50 � for the G18-C17 step).
Figure 5 shows more detailed views of the threecentral base-pairs of the duplex. The eC6 �C17base-pair is sheared by 5.1 AÊ , with the C17 basedisplaced into the minor groove (Xdisp � ÿ3.4 AÊ )and the eC6 toward the major groove(Xdisp � 1.7 AÊ ). This base-pair also shows a nega-tive stretch of 1.5 AÊ and it is propeller twisted byÿ21 �. Table 2 lists some of the relevant structuralparameters. The positioning of the central basesallows the formation of a hydrogen bond betweeneC6(O2) and C17(N4H2) with a distance of 1.9 AÊ
and an O2 � � �HÐN4 angle of 160 �. The local dis-ruption of the structure at the lesion site seems tohave little affect on the alignment of the ¯ankingbase-pairs, as evidenced by quite normal par-ameters for the A4 �T19, C5 �G18, C7 �G16, andA8 �T15 base-pairs.
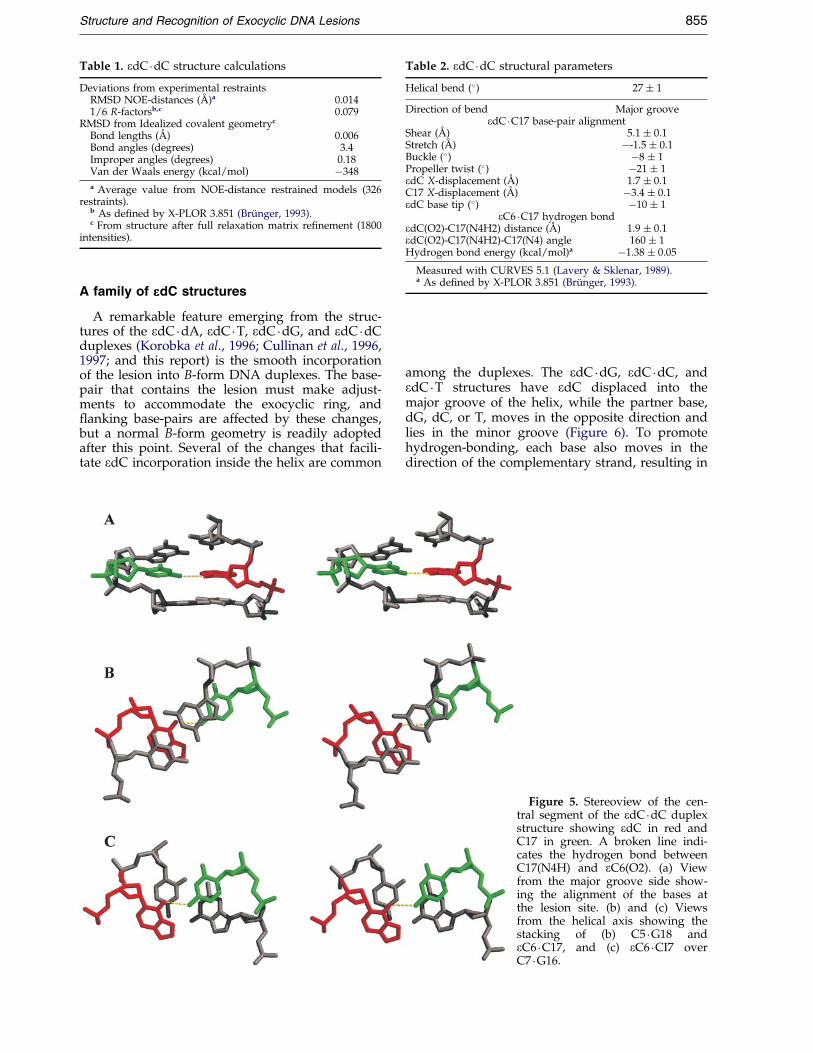
Table 1. edC �dC structure calculations
Deviations from experimental restraintsRMSD NOE-distances (AÊ )a 0.0141/6 R-factorsb,c 0.079
RMSD from Idealized covalent geometryc
Bond lengths (AÊ ) 0.006Bond angles (degrees) 3.4Improper angles (degrees) 0.18Van der Waals energy (kcal/mol) ÿ348
a Average value from NOE-distance restrained models (326restraints).
b As de®ned by X-PLOR 3.851 (BruÈ nger, 1993).c From structure after full relaxation matrix re®nement (1800
intensities).
Table 2. edC �dC structural parameters
Helical bend (�) 27 � 1
Direction of bend Major grooveedC �C17 base-pair alignment
Shear (AÊ ) 5.1 � 0.1Stretch (AÊ ) ÿ-1.5 � 0.1Buckle (�) ÿ8 � 1Propeller twist (�) ÿ21 � 1edC X-displacement (AÊ ) 1.7 � 0.1C17 X-displacement (AÊ ) ÿ3.4 � 0.1edC base tip (�) ÿ10 � 1
eC6 �C17 hydrogen bondedC(O2)-C17(N4H2) distance (AÊ ) 1.9 � 0.1edC(O2)-C17(N4H2)-C17(N4) angle 160 � 1Hydrogen bond energy (kcal/mol)a ÿ1.38 � 0.05
Measured with CURVES 5.1 (Lavery & Sklenar, 1989).a As de®ned by X-PLOR 3.851 (BruÈ nger, 1993).
Structure and Recognition of Exocyclic DNA Lesions 855
A family of eeedC structures
A remarkable feature emerging from the struc-tures of the edC �dA, edC �T, edC �dG, and edC �dCduplexes (Korobka et al., 1996; Cullinan et al., 1996,1997; and this report) is the smooth incorporationof the lesion into B-form DNA duplexes. The base-pair that contains the lesion must make adjust-ments to accommodate the exocyclic ring, and¯anking base-pairs are affected by these changes,but a normal B-form geometry is readily adoptedafter this point. Several of the changes that facili-tate edC incorporation inside the helix are common
among the duplexes. The edC �dG, edC �dC, andedC �T structures have edC displaced into themajor groove of the helix, while the partner base,dG, dC, or T, moves in the opposite direction andlies in the minor groove (Figure 6). To promotehydrogen-bonding, each base also moves in thedirection of the complementary strand, resulting in
Figure 5. Stereoview of the cen-tral segment of the edC �dC duplexstructure showing edC in red andC17 in green. A broken line indi-cates the hydrogen bond betweenC17(N4H) and eC6(O2). (a) Viewfrom the major groove side show-ing the alignment of the bases atthe lesion site. (b) and (c) Viewsfrom the helical axis showing thestacking of (b) C5 �G18 andeC6 �C17, and (c) eC6 �CI7 overC7 �G16.

Figure 6. Structure of the centralsegment of edC duplexes and aG �T mismatch (Hunter et al., 1987)shown with the major groove pro-minent. The structural family, com-posed by edC �T, edC �dC, edC �dG,and G �T duplexes, has the edC orT residues (in yellow) displacedinto the major groove and thepartner residues (in red) into theminor groove. All four duplexeshave hydrogen bonds at the lesionsite. These are: G(H1)-edC(O2),G17(N2H1)-C7(O40), G17(N2H2)-G18(O40) in the edC �dG duplex,T(H3)-edC(N4) in the edC �Tduplex, C17(N4H)-edC(O2) in theedC �dC duplex, and G(O6)-T(H3),G(N1H)-T(O2) in the G �T mis-match.
856 Structure and Recognition of Exocyclic DNA Lesions
a lesion-containing base-pair which is highlysheared, relatively coplanar, and has a negativestretch value. The helix is slightly bent (9-27 �) inthe direction of the major groove, and the minorgroove becomes wider than in canonical B-formDNA. The structural parameters that de®ne theedC family are listed in Table 3. Only the edC �dAduplex adopts a different conformation, character-ized by a highly staggered alignment at the lesionsite, such that the edC base stacks to the 30 side ofits partner dA, and the dA base to the 30 side ofthe adduct (Korobka et al., 1996). Each residuemoves in the direction of the complementarystrand, facilitating hydrophobic interaction withinthe base-pair but, in this case, there is no hydrogenbond present. This staggered conformation dra-
Table 3. Structural parameters edC-containing base-pairs
X-Displa. (AÊ ) Y-Displb. (AÊ )
edC X edC X
edC �dA ÿ1.1 0.4 ÿ1.4 ÿ1.2edC �dC 1.7 ÿ3.4 0.7 ÿ0.6edC �dG 1.1 ÿ1.7 0.0 ÿ1.4edC �T 4.5 ÿ1.0 0.0 ÿ0.1dG �Tf 0.7 ÿ1.7 ÿ0.2 ÿ0.1
a X-displacement describes base movement in the plane of the basment in the direction of the major groove.
b Y-displacement describes base movement in the plane of the bmovement in the direction of the opposing base.
c Shear is the X-displacements difference within the base-pair.d Stretch is the sum of Y-displacements within a base-pair.e Stagger is the relative movement in the direction of the helical a
X-displacement � ÿ0.71; Y-displacement, Shear, Stretch, and Staggerf Parameters for the T residue are given in the edC column.
matically increases rise values at the lesion site,resulting in a duplex that has an end to end dis-tance 3-4.5 AÊ greater than that of the other threeduplexes (Table 3).
The structures of G �T mismatch-containingduplexes have characteristics of the edC family.Independent of the DNA conformation (A or B-form duplexes), the changes induced by the pre-sence of the mismatch are always localized at thelesion site. In the X-ray structures, the mismatchedT is invariably displaced into the major groove ofthe helix while its partner G moves toward theminor groove (Kennard, 1985; Hunter et al., 1987).This sheared alignment, called wobble in the orig-inal report, is stabilized by the formation of twohydrogen bonds across the mismatched base-pair,
Base-base parameters (AÊ ) Axis parameters
Shearc Stretchd Staggere Bend (�)
ÿ1.5 ÿ2.7 ÿ3.4 16 (Minor groove)5.1 ÿ1.3 ÿ0.4 27 (Major groove)2.8 ÿ1.4 2.0 14 (Major groove)5.5 ÿ0.1 1.0 9 (Major groove)2.4 ÿ0.3 ÿ0.1 3 (Minor groove)
e-pair toward the DNA grooves. Positive values indicate move-
ase-pair toward the DNA backbone. Negative values indicate
xis of bases in a base-pair. Canonical values in B-form DNA are� 0.0.

Figure 7. Side by side compari-son of the edC �dG adduct-contain-ing (top structures) and dG �Tmismatch-containing (bottom struc-tures) duplexes. (a) Lateral view,depicting the edC and T residues(in yellow) in the major groove(right side of the picture); (b) lateralview from the minor groove side;and (c) view from the top (minorgroove towards the bottom of thebase-pair) showing the similitudebetween the wobble dG �T and theedC �dG base-pair alignments.
Structure and Recognition of Exocyclic DNA Lesions 857
and allows favorable stacking interactions with¯anking bases. Figure 7 compares the crystal struc-ture of a dG �T mismatch duplex at 2.5 AÊ resol-ution (Hunter et al., 1987) and the solutionstructure of a edC �dG duplex (Cullinan et al.,1997). The overall conformations of the sugar-phos-phate backbone in these duplexes are similar, asevidenced by pair-wise RMS deviation of 1.9 AÊ .Values of 1.7 AÊ and 2.0 AÊ are obtained when thedG �T mismatch is compared to the edC �T andedC �dC duplexes, respectively. The edC lesion andthe mismatched T are structurally equivalent in thesense that both residues are displaced towards themajor groove of the helix. At the same time, thepartner dG bases (T or dC residues in the edC �Tand edC �dC duplex, respectively), are displacedtowards the minor groove. No other structural par-ameter is accordingly changed in the edC and G �Tmismatched duplexes. The structural parameters ofthe dG �T alignment are also listed in Table 3.
Discussion
eeedC �dC duplex structure
The complete assignment of the NMR signals toa single set of protons, together with the obser-vation of sharp and well-resolved lines in the spec-tra, indicates that the duplex adopts a singleconformation in solution. This structure is that of aright-handed helix belonging to the B-form DNAfamily with a bend in the helical axis centeredbetween the edC �C17 and C7 �G16 base-pairs. TheedC sugar has a north conformation and points itsedC(H20) proton toward the C7 base, resulting in atilt of 13 � at the edC-C7 step. Watson-Crick hydro-gen bonding is preserved in all non-lesion-contain-ing base-pairs and base stacking is quite regularthroughout the duplex.
It is important to emphasize the experimentalevidence de®ning the alignment of the lesion-con-taining base-pair and the stacking of edC and C17
inside the helix. The NOE interactions of edC(H5)with C5(H6), edC(H6) with C7(H5), edC(H8) withthe H5 protons of C5 and C7 (Figure 2, peaks G, F,E, and D, respectively), and edC(H5, H7, and H8)with the imino protons of the ¯anking base-pairs(Figure 3, peaks T-V, X, and Y, respectively) leaveno doubt about the intrahelical location of edC. Bycontrast, the position of C17 opposite the lesion isless well established by the data. However, thequite normal NOE intensities through C17 and theproximity of G16(H8) and C17(H5) (Figure 2, leftside of peak H) argue for a stacked conformationof C17 in a standard anti conformation around theglycosylic torsion angle. The three-dimensionalstructure shows that edC and C17 are stacked with¯anking bases and displaced to opposite groovesof the helix. This creates a sheared base-pair thatallows the formation of a single hydrogen bond,which contributes stability to the duplex at thelesion site.
DNA recognition by MUG and hTDG
It is remarkable that such different DNA aberra-tions as G �T mismatches and edC lesions arerecognized and repaired by the same two DNAglycosylases, MUG and hTDG. In the case of themismatch, the enzymes can recognize and cleave athymine or uracil only when the base is pairedwith dG (Wiebauer & Jiricny, 1989) whereas, in thelatter case, they remove a highly modi®ed cytosinemoiety, edC, independent of the opposing base(Saparbaev & Laval, 1998). A clue to understand-ing how these glycosylases may accomplish suchspeci®city is the identi®cation of common structur-al features on the DNA substrates. The residuesthat will be removed by the enzymes, edC or T,are always displaced towards the major grooveand the partner base is displaced to the minorgroove. Other structural parameters, such as a heli-cal bend, twist, or groove dimensions, which arethought to facilitate DNA recognition, are notchanged systematically in the edC and G �T

858 Structure and Recognition of Exocyclic DNA Lesions
duplexes (Table 3). Therefore, we propose that thehighly sheared base-pair alignment, present in theG �T, edC �dG, edC �dC, and edC �T duplexes, is thesubstrate structural motif that facilitates their rec-ognition by MUG and hTDG.
The question as to how these proteins are able torecognize a single G �T mismatch or edC lesioncontained within a background of normal base-pairs still remains unanswered. The possibility ofdamage recognition via protein interactions withcharged groups of edC and G �T duplexes canimmediately be ruled out, because the distributionof polar groups at the lesion site is different for allknown substrates. It has been postulated thathuman AP endonuclease binds directly to a duplexsubstrate containing an everted abasic site(Gorman et al., 1997). Although our data do notrule out the Possibility that MUG and hTDGdirectly bind to a pre-existing ¯ipped-out residue,they suggest this is unlikely. While the thermalstability of edC-containing duplexes is diminished(Gelfand et al., 1998), all the structures showed apredominant conformation at 25 �C with the eCmoiety stacked inside the helix (Korobka et al.,1996; Cullinan et al., 1996, 1997, and Figure 2). Fur-thermore, the ef®ciency of MUG (kcat/Km � 0.38 minÿ1 nMÿ1 for a edC �dG duplex sub-strate) suggests an active participation of theenzyme during the base eversion process(Saparbaev & Laval, 1998).
In the case of edC and G �T duplexes, we postu-late that damage recognition and base extrusionare combined in a single process driven by thermo-dynamic properties of the duplex substrates andstructures of the kinetic intermediates. After initiallow af®nity binding of the protein to the DNA, theenzyme moves along the helical axis probing thestability of each base-pair with its recognitionwedge. Two concurrent conditions lead to the for-mation of a competent protein/DNA complex onlywhen MUG (hTDG) encounters a highly shearedbase-pair alignment. First, DNA duplexes contain-ing such an alignment have a reduced thermodyn-amic stability, diminishing the energetic costinvolved in the extrusion process. A recent studyestablished that the presence of a single edC-con-taining base-pair diminished the thermal stabilityof a 13-mer duplex by 10-18 �C and induced a ther-modynamic destabilization (��G) of ÿ13.8 kcal/mol (Gelfand et al., 1998). Similarly, replacement ofa G �C pair by a G �T mismatch reduced the stab-ility of the duplex DNA by 3.5 kcal/mol (Allawi &SantaLucia, 1997). Second, a sheared base-pairalignment with the scissile base displaced towardsthe major groove of the helix does not interferewith, but rather facilitates, the process of baseextrusion and the formation of a productive DNA/protein complex. MUG enzyme is less active inremoving edC lesions when paired with dAsuggesting that, while still possible, a staggerededC �dA alignment does not facilitate the extrusionprocess.
eeedC and alkyladenine DNA glycosylase (AAG)
AAG is another example of a nucleotide evertingenzyme that interacts with DNA substrates on theminor groove side of the helix. It removes 3-methy-ladenine, 7-methylguanine, 8-oxoguanine, 1,N6-ethenoadenine, N2,3-ethenoguanine, and 1,N2-ethe-noguanine adducts but, quite remarkably, is inac-tive against 3,N4-ethenocytosine-containingduplexes. It has been postulated that AAG recog-nizes DNA duplex substrates by sequentiallyextruding each residue from the helix and remov-ing any base that ®ts its catalytic pocket (Lau et al.,1998). In this model, DNA substrates have a pas-sive role and duplex structure is not relevant forthe recognition process. However, docking of edCinto the active site of AAG shows that the adduct®ts quite nicely into the catalytic pocket of theenzyme, suggesting that formation of a competentcomplex is sterically possible (data not shown). Webelieve that the lack of AAG activity on edClesions can be explained by considering that sub-strate structures play a role in the recognition pro-cess. The crystal structure of an AAG/enzymeinhibitor complex showed the widowed residuepositioned into the major groove of the helix (Lauet al., 1998), as opposed to the minor groove posi-tioning observed for MUG substrates. It is possiblethat AAG exerts a different mechanism of baseextrusion, which is blocked by the sheared base-pair alignment seen in edC duplexes. Other DNAglycosylases are likely to act on everted residuesand have a preferential set of substrates. Theknowledge of substrate structures, their thermo-dynamic properties, and the conformation of pro-tein/DNA complexes are necessary to identify allthe elements involved in substrate recognition andenzyme speci®city.
Materials and Methods
Sample preparation
The 3,N4-etheno-20-deoxycytidine adduct was syn-thesized as a 50-dimethoxytrityl-30-b-cyanoethylpho-sphoroamidite derivative (Zhang et al., 1995b) andincorporated into the oligodeoxynucleotide sequence bystandard phosphoramidite chemistry procedures. Thecrude synthesis product was treated with concentratedammonia for 46 hours at room temperature, to isolatesequences containing an O-50-dimethoxytrityl group, andpuri®ed by reverse phase HPLC on a preparative Dyna-max (300 mm � 25 mm) C4 column. The mobile phaseconsisted of solvent A (0.1 M triethylamine acetic acidbuffer, pH 6.8) and solvent B (acetonitrile). Using a lineargradient of 0 to 50 % of B over 50 minutes, the desiredsequence was eluted as a main fraction at 33 minutes.The O-50-dimethoxytrityl group was cleaved by treat-ment with 80 % acetic acid for 30 minutes, followed bytwo rounds of ether extraction before puri®cation withHPLC. Oligodeoxynucleotide sequences were desaltedby passing them through a Sephadex G-25 column andconverted to the sodium salt using a Dowex 50W cationexchange column. A 1:1 stoichiometric ratio of theduplex was obtained by monitoring the intensity of

Structure and Recognition of Exocyclic DNA Lesions 859
NMR proton signals during gradual addition of theunmodi®ed strand to the eC-containing strand at 55 �C.NMR samples consisted of 550 A260 of the duplex dis-solved in 0.8 ml of 50 mM phosphate buffer (pH 6.8)containing 50 mM NaCl and 0.5 mM EDTA in either99.96 % 2H2O or 90 % H2O-10 % 2H2O (v/v), correspond-ing to a concentration of approximately 6 mM. Sampleswere degassed under nitrogen before collection of theNMR data.
NMR experiments
One- and two-dimensional NMR experiments wererecorded on Varian Inova spectrometers operating at11.75 T and 14.1 T. Proton chemical shifts were refer-enced relative to TSP at 0.0 ppm. Phase-sensitive NOESY(50, 100, 150, 200, and 250 ms mixing times), COSY,DQF-COSY, and TOCSY spectra in 2H2O buffer were col-lected with a repetition delay of 1.5 seconds, duringwhich the residual water signal was suppressed by sat-uration. A NOESY (150 ms mixing time) spectrum in90 % H2O buffer was recorded using a jump-return read-ing pulse. Time domain data sets consisted of 2048 � 300complex data points in the t2 and t1 dimensions, respect-ively. A COSY45 spectrum was recorded using6144 � 1000 complex points. An inverse-detected proton-phosphorous correlation (HETCOR) spectrum wasrecorded using 4096 complex points � 150 increments.All 2H2O experiments were carried out at 15 �C, exceptthe proton phosphorus correlation, which was recordedat 25 �C. The NOESY experiment in 90 % H2O was per-formed at 5 �C. NMR data were processed using FELIX(Biosym Technologies, Inc.). Time domain data sets weremultiplied by 90 � shifted sinebell window function priorto Fourier transformation. No base line correction wasapplied to the transformed spectra.
Molecular dynamics simulations
Distance restraints were determined from NOE cross-peak volumes using MARDIGRAS (UCSF). Based on theconsistency of the determined distance through thedifferent mixing times, the average distances weredivided into three groups with distance bounds of�0.5 AÊ , �0.6 AÊ , or �0.8 AÊ . A total of 300 restraints wereused. Initial structures were created using INSIGHTII(Biosym Technologies, Inc.) from canonical A and B-form11-mer duplexes, adding an etheno bridge to the N4- andN3-nitrogen atoms of a deoxycytidine residue in the sixthposition of one strand, and replacing the guanine oppo-site it with a cytosine. The initial models were energyminimized without NOE-derived restraints, to relievesteric clashes, then minimized again with the distancerestraints present, prior to molecular dynamics simu-lations. Energy minimization and restrained moleculardynamics simulations were carried out in vacuo on Sili-con Graphics and Sun computers using X-PLOR 3.851(BruÈ nger, 1993). Partial atomic charges on phosphategroups were not reduced, resulting in residues with anet charge of ÿ1. The value of the dielectric constantwas set to 4 (Friedman & Honig, 1992). Experimentaldistance bounds were enforced using a square-wellpotential energy function with force constants of100 kcal/(mol AÊ 2) (heating stage) and 50 kcal/(mol AÊ 2)(cooling/dynamic stage). Watson-Crick hydrogen bondalignments were enforced by distance restraints on allcanonical base-pairs of the duplex. Backbone dihedralangles were restrained by a square-well potential energy
function with a width encompassing A and B-form DNAvalues. Distance restrained molecular dynamics simu-lations consisted of a heating stage, an equilibrationstage, a cooling stage, and a dynamics stage. At the heat-ing stage, the temperature of the system was broughtfrom 50, 100 or 150 K to 500 K over a period of 5 ps. Inthe equilibration stage, the temperature stayed at 500 Kfor 30, 31, 32, 33, or 34 ps, during which the scale of thedistance restraints was increased to its maximum valuein the ®rst 8 ps. At the end of the equilibration stage thesystem was cooled to 300 K over 5 ps and kept at thistemperature for 200 ps. Coordinates of the last 10 ps ofthe dynamics stage were averaged, and subjected to a2500 step energy minimization. A total of 30 distancerestrained structures were generated by using three start-ing temperatures, ®ve different lengths of the equili-bration stage, and two initial models. A new averagestructure, computed from the converging distance-restrained models, was then subjected to a further 30 psof molecular dynamics at 300 K restrained by the fullrelaxation matrix method (Nilges et al., 1991). A total of1800 experimental volumes measured from NOESY spec-tra were input as restraints with a 30 % error range. Agrid search determined that an isotropic correlation timeof 2.25 ns best ®t the experimental data, so this valuewas used throughout the re®nement. Coordinates duringthe last 2 ps of the re®nement were averaged and energyminimized, yielding the ®nal structure. The structureswere visualized using MIDAS PLUS (UCSF, ComputerGraphics Laboratory) and structural parameters wereobtained using CURVES (Lavery & Sklenar, 1989).
Note
After submission of this manuscript, the crystal struc-ture of MUG enzyme in a complex with an oligonucleo-tide containing a non-hydrolysable deoxyuridine analogmismatched with guanine was reported (Barrett et al.,1999). In the structure, the scissile uridine residue iseverted from the helix into a non-speci®c binding pocketof the enzyme. Protein residues occupy the cavity left inthe DNA duplex in a form similar to what was pre-viously observed in the MUG/abasic site complex(Barrett et al., 1998).
PDB Accession codes
Atomic coordinates have been deposited in the ProteinData Bank. ID codes are: 1B6Y for edC �dA, 1B6O foredC �dC, 1B6X for edC �dG, and 1B5 K for edC �T.
Acknowledgements
This research was supported by Grant CA47995 fromthe NIH. The NMR facility at SUNY Stony Brook is sup-ported by grants CHE8911350 and CHE9413510 from theNSF and 1S10RR554701 from the NIH.
We thank Dr Charles R. Iden, Mr Robert Rieger andMs Cecilia, Torres for the synthesis and puri®cation ofmodi®ed oligodeoxynucleotides, and Dr Arthur P. Groll-man for many discussions during the preparation of themanuscript.

860 Structure and Recognition of Exocyclic DNA Lesions
References
Allawi, H. T. & SantaLucia, J. Jr (1997). Thermodynamicsand NMR of internal G.T mismatches in DNA.Biochemistry, 36, 10581-10594.
Barbin, A. & Bartsch, H. (1986). The Role of Cyclic NucleicAcid Adducts in Carcinogenesis and Mutagenesis, No.70, pp. 345-358, International Agency for Researchon Cancer, Lyon, France.
Barbin, A., Besson, F. & Perrard, M. (1985). Induction ofspeci®c base-pair substitutions in E. coli trpAmutants by chloroethylene oxide, a carcinogenicvinyl chloride metabolite. Mutat. Res. 152, 147-156.
Barrett, T. E., Savva, R., Panayotou, G., Barlow, T.,Brown, T., Jiricny, J. & Pearl, L. H. (1998). Crystalstructure of a G:T/U mismatch speci®c DNA glyco-sylase: mismatch recognition by complementary-strand interactions. Cell, 92, 117-129.
Barrett, T. E., SchaÈrer, O. D., Savva, R., Brown, T.,Jiricny, J., Verdine, G. L. & Pearl, L. H. (1999). Crys-tal structure of a thwarted mismatch glycosylaseDNA repair complex. EMBO J. 18, 6599-6609.
Basu, A. K., Wood, M. L., Niedernhofer, L. J., Ramos,L. A. & Essigmann, J. M. (1993). Mutagenic andgenotoxic effects of three vinyl chloride-inducedDNA lesions: 1,N6-ethernoadenine, 3,N4-ethenocyto-sine, and 4-amino-5-(imidazol-2-yl)imidazole.Biochemistry, 32, 12793-12801.
BruÈ nger, A. (1993). X-PLOR, Version 3.1: A System for X-Ray Crystallography and NMR, Yale University Press,New Haven, CT.
Cheng, H.-J. C. & Chung, F.-L. (1996). Epoxidation oftrans-4-hydroxy-2-nonenal by fatty acid hydroperox-ides and hydrogen peroxide. Chem. Res. Toxicol. 9,306-312.
Creech, J. L. Jr & Johnson, M. N. (1974). Angiosarcomaof liver in the manufacture of polyvinyl chloride.J. Occup. Med. 16, 150-151.
Cullinan, D., Korobka, A., Grollman, A. P., Patel, D. J.,Eisenberg, M. & de los Santos, C. (1996). NMR sol-ution structure of an oligodeoxynucleotide duplexcontaining the exocyclic lesion 3,N4-etheno-20-deoxy-cytidine opposite thymidine: comparison with theduplex containing deoxyadenosine opposite theadduct. Biochemistry, 35, 13319-13327.
Cullinan, D., Johnson, F., Grollman, A. P., Eisenberg, M.& de los Santos, C. (1997). Solution structure of aDNA duplex containing the exocyclic lesion 3,N4-etheno-20-deoxycytidine opposite 20deoxyguanosine.Biochemistry, 36, 11933-11943.
el Ghissassi, F., Barbin, A., Nair, J. & Bartsch, H. (1995).Formation of 1,N6-ethenoadenine and 3,N4-etheno-cytosine by lipid peroxidation products and nucleicacid bases. Chem. Res. Toxicol. 8, 278-283.
Friedman, R. A. & Honig, B. (1992). The electrostaticcontribution to DNA base-stacking interactions.Biopolymers, 32, 145-159.
Gelfand, C. A., Plum, E., Grollman, A. P., Johnson, F. &Beslauer, K. J. (1998). The impact of an exocycliccytosine adduct on DNA duplex properties: signi®-cant thermodynamic consequences despite modestlesion-induced structural alterations. Biochemistry,37, 12507-12512.
Gorman, M. A., Morera, S., Rothwell, D. G., de LaFortelle, E., Mol, C. D., Tainer, J. A., Hickson, I. D.& Freemont, P. S. (1997). The crystal structure ofthe human DNA repair endonuclease HAP1suggests the recognition of extra-helical deoxyriboseat DNA abasic sites. EMBO J. 16, 6548-6558.
Guengerich, F. P., Crawford, W. M. & Hathaway, D. E.(1979). Activation of vinyl chloride to covalentlybound metabolites: roles of 2-chloroethylene oxideand 2-chloroacetaldehyde. Biochemistry, 18, 5177-5182.
Hang, B., Medina, M., Fraenkel-Conrat, H. & Singer, B.(1998). A 55-kDa protein isolated from human cellsshows DNA glycosylase activity toward 3,N4-ethe-nocytosine and the G/T mismatch. Proc. Natl Acad.Sci. USA, 95, 13561-13566.
Hare, D. R., Wemmer, D. E., Chou, S. H., Drobny, G. &Reid, B. (1983). Assignment of the non-exchangeableproton resonances of d(C-G-C-G-A-A-T-T-C-G-C-G)using two-dimensional nuclear magnetic resonancemethods. J. Mol. Biol. 171, 319-336.
Hunter, W. N., Brown, T., Kneale, G., Anand, N. N.,Rabinovich, D. & Kennard, O. (1987). The structureof guanosine-thymidine mismatches in B-DNA at2.5-AÊ resolution. J. Biol. Chem. 262, 9962-9970.
Kennard, O. J. (1985). Structural studies of DNA frag-ments: the G.T wobble base-pair in A, B and ZDNA; the G.A base-pair in B-DNA. Biomol. Struct.Dynam. 3, 205-226.
Korobka, A., Cullinan, D., Cosman, M., Grollman, A. P.,Patel, D. J., Eisenberg, M. & de los Santos, C.(1996). Solution structure of an oligodeoxynucleo-tide duplex containing the exocyclic lesions 3,N4-etheno-20-deoxycytidine opposite 20-deoxyadeno-sine, determined by NMR spectroscopy andrestrained molecular dynamics. Biochemistry, 35,13310-13318.
Kusmierik, J. T. & Singer, B. (1992). 1,N2-Ethenodeoxy-guanosine: properties and formation in chloroacetal-dehyde-treated polynucleotides and DNA. Chem.Res. Toxicol. 5, 634-638.
Lau, A. Y., ScaÈrer, O. D., Samson, L., Verdine, G. L. &Ellenberger, T. (1998). Crystal structure of a humanalkylbase-DNA repair enzyme complexed to DNA:mechanisms for nucleotide ¯ipping and base exci-sion. Cell, 95, 249-258.
Lavery, R. & Sklenar, H. (1989). De®ning the structureof irregular nucleic acids: conventions and prin-ciples. J. Biomol. Struct. Dynam. 6, 655-667.
Maltoni, C., Lefemine, G., Chieco, P. & Carretti, D.(1974). Vinyl chloride carcinogenesis: current resultsand perspectives. Med. Lavoro. 65, 421-444.
Moriya, M., Zhang, W., Johnson, F. & Grollman, A. P.(1994). Mutagenic potency of exocyclic DNAadducts: marked differences between Escherichia coliand simian kidney cells. Proc. Natl Acad. Sci. USA,91, 11899-11903.
Nair, J., Barbin, A., Guichard, Y. & Bartsch, H. (1995).1,N6-ethenodeoxyadenosine and 3,N4-ethenodeoxy-cytine in liver DNA from humans and untreatedrodents detected by immunoaf®nity/32P-postlabel-ing. Carcinogenesis, 16, 613-617.
Nilges, M., Habazettl, J., Brunger, A. T. & Holak, T. A.(1991). Relaxation matrix re®nement of the solutionstructure of squash trypsin inhibitor. J. Mol. Biol.219, 499-510.
Palejwala, V. A., Simha, D. & Humayun, M. Z. (1991).Mechanisms of mutagenesis by exocyclic DNAadducts. Transfection of M13 viral DNA bearing asite-speci®c adduct shows that ethenocytosine is ahighly ef®cient RecA-independent mutagenic nonin-structional lesion. Biochemistry, 30, 8736-8743.
Palejwala, V. A., Rzepka, R. W. & Humayun, M. Z.(1993). UV irradiation of Escherichia coli modulatesmutagenesis at a site-speci®c ethenocytosine residue

Structure and Recognition of Exocyclic DNA Lesions 861
on M13 DNA. Evidence for an inducible recA-inde-pendent effect. Biochemistry, 32, 4112-4120.
Saparbaev, M. & Laval, J. (1998). 3,N4-ethenocytosine, ahighly mutagenic adduct, is a primary substrate forEscherichia coli double-stranded uracil-DNA glycosy-lase and human mismatch-speci®c thymine-DNAglycosylase. Proc. Natl Acad. Sci. USA, 95, 8508-8513.
Scherer, E., Winterwerp, H. & Emmelot, P. (1986). TheRole of Cyclic Nucleic Acid Adducts in Carcinogenesisand Mutagenesis, No. 70, pp. 109-125, InternationalAgency for Research on Cancer, Lyon, France.
Shibutani, S., Suzuki, N., Matsumoto, Y. & Grollman,A. P. (1996). Miscoding properties of 3,N4-etheno-20-deoxycytidine in reactions catalyzed by mammalianDNA polymerases. Biochemistry, 35, 14992-14998.
Simha, D., Palejwala, V. A. & Humayun, M. Z. (1991).Mechanisms of mutagenesis by exocyclic DNAadducts, construction and in vitro template charac-teristics of an oligonucleotide bearing a single site-speci®c ethenocytosine. Biochemistry, 30, 8727-8735.
Singer, B. & Spengler, S. J. (1986). The Role of CyclicNucleic Acid Adducts in Carcinogenesis and Mutagen-esis, No. 70, pp. 345-358, International Agency forResearch on Cancer, Lyon, France.
Wiebauer, K. & Jiricny, J. (1989). In vitro correction ofG �T mispairs to G �C pairs in nuclear extracts fromhuman cells. Nature, 339, 234-236.
WuÈ thrich, K. (1986). NMR of Proteins and Nucleic Acids,John Wiley, New York.
Zhang, W., Johnson, F., Grollman, A. P. & Shibutani, S.(1995a). Miscoding by the exocyclic and relatedDNA adducts 3,N4-etheno-20-deoxycytidine, 3,N4-ethano-20-deoxycytidine, and 3-(2-hydroxyethyl)-20-deoxyuridine. Chem. Res. Toxicol. 8, 157-163.
Zhang, W., Rieger, R., Iden, C. & Johnson, F. (1995b).Synthesis of 3,N4-etheno, 3,N4-ethano, and 3-(2-hydroxyethyl) derivatives of 20-deoxycytidine andtheir incorporation into oligomeric DNA. Chem. Res.Toxicol. 8, 148-156.
Edited by I. Tinoco
(Received 29 October 1999; received in revised form 23December 1999; accepted 23 December 1999)
http://www.academicpress.com/jmb
Supplementary material comprising threeFigures and two Tables is available from JMBOnline






















![A role for the base excision repair enzyme NEIL3 in replication ...faculty.missouri.edu/~gatesk/gatespapers/2017_DNA_repair_review.pdf · bose backbone [82]. Various BER glycosylases](https://img.pdfslide.us/doc/110x75/5e7b7c07f1250763aa59d00b/a-role-for-the-base-excision-repair-enzyme-neil3-in-replication-gateskgatespapers2017dnarepairreviewpdf.jpg)






