Embed Size (px)
Citation preview

This article was downloaded by: [Dr Favio Vossler]On: 25 March 2015, At: 13:05Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Click for updates
GranaPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/sgra20
Small pollen grain volumes and sizes dominatethe diet composition of three South Americansubtropical stingless beesFavio Gerardo Vosslera
a Laboratorio de Actuopalinología, CICyTTP-CONICET/FCyT-UADER, Diamante, ArgentinaPublished online: 23 Jul 2014.
To cite this article: Favio Gerardo Vossler (2015) Small pollen grain volumes and sizes dominate the diet composition ofthree South American subtropical stingless bees, Grana, 54:1, 68-81, DOI: 10.1080/00173134.2014.932838
To link to this article: http://dx.doi.org/10.1080/00173134.2014.932838
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose ofthe Content. Any opinions and views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be reliedupon and should be independently verified with primary sources of information. Taylor and Francis shallnot be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and otherliabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

Small pollen grain volumes and sizes dominate the diet composition ofthree South American subtropical stingless bees
FAVIO GERARDO VOSSLER
Laboratorio de Actuopalinología, CICyTTP-CONICET/FCyT-UADER, Diamante, Argentina
AbstractThe pollen preferences of three Meliponini species (Geotrigona argentina, Scaptotrigona jujuyensis and Tetragonisca fiebrigi)were studied in the Chaco forest of South America. Surprisingly, a large number of pollen types with small volumes and sizewere found in pollen masses stored in the 18 nests studied. Small pollen grains are commonly found in small flowers withshort pistils, a flower morphology adapted to the pollination by these small- to medium-sized bees. The prevalence of grainswith small volumes may be due to the preference of Meliponini bees for small flowers with short pistils, supporting apreviously suggested hypothesis of diffuse co-evolution between mass-flowering trees and these tiny bees in Neotropicalhabitats. However, the opportunistic foraging habit of these generalist bees would suggest that the large number of grainswith small size is due to their greater availability in the Chaco forest nearby the nests. This study also provides measurementsand descriptions of pollen cytoplasmic shapes as well as a new classification of pollen volumes based from measurements ofthe five pollen size categories using non-acetolyzed grains.
Keywords: corbiculate bee, cytoplasmic volume, Geotrigona argentina, opportunistic bee, pollen size category,Scaptotrigona jujuyensis, Tetragonisca fiebrigi
The Meliponini bee fauna of the subtropical DryChaco forest of Argentina is composed of at leastseven species (Arenas 2003; Vossler 2012; Roig-Alsina et al. 2013). To maintain their perennial colo-nies with hundreds to thousands of individuals andhigh rates of offspring, these bees have to forage largeamounts of food from a great number of plants fromdiverse families (Roubik 1989; Ramalho et al. 2007).
It is well known that other corbiculate bees such asthe widely studied Apis mellifera L. forage on a broadspectrum of plants. For instance, O’Rourke andBuchmann (1991) found that A. mellifera gatheredsimilar amounts of large, medium and small sizes ofpollen grains in a study carried out in an arid region ofthe Sonoran Desert in Arizona, but Biesmeijer et al.(1992) found that it foraged mainly on small pollengrains in Suriname. Unfortunately, few studies haverecorded the volume and size of the pollen grainsforaged by Meliponini bees to make comparisons.
For instance, it has been reported that the BrazilianTrigona spinipes (Fabricius) forage on a broad spec-trum of pollen sizes from 10 to > 100 µm, whileA. mellifera forage on a smaller range (10 to ≤ 71 µm)in the same area (Cortopassi-Laurino & Ramalho1988). Marques-Souza et al. (2002) reported that pol-len grains of diverse sizes are equally present amongthe pollen gathered by the Brazilian AmazonianFrieseomelitta, Melipona and Scaptotrigona. In the DryChaco forest, Vossler et al. (2010) reported thatGeotrigona argentina Camargo et Moure intensivelyforaged (≥ 5%) on pollen types of very small to med-ium volumes (from 65 to 34 782 µm3).In view of the polylectic specialization category of all
members of this bee tribe, it is expected that the threesubtropical Meliponini species studied forage on awide spectrum of flower sizes having pollen grainsfrom very small (< 10 µm and < 524 µm3) to verylarge (> 100 µm and > 523 583 µm3) both in size and
Correspondence: Favio Gerardo Vossler, Laboratorio de Actuopalinología, CICyTTP-CONICET/FCyT-UADER, Dr Materi y España, E3105BWA,Diamante, Entre Ríos, Argentina. E-mail: [email protected]
(Received 16 June 2013; accepted 1 June 2014)
Grana, 2015Vol. 54, No. 1, 68–81, http://dx.doi.org/10.1080/00173134.2014.932838
© 2014 Collegium Palynologicum Scandinavicum
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015

volume. For this reason, the goals of this study wereto establish the cytoplasmic volume and size categoriesof the pollen grains foraged by three species ofMeliponini bees in the South American Chaco forest,particularly those intensively foraged, to detect a pos-sible relationship between pollen size/volume andflower size, and to give possible explanations aboutwhy certain pollen grains are highly abundant in thesecolonies.
Material and methods
Dry Chaco forest vegetation
The vegetation units of the study area are ‘Quebrachal’of three ‘quebrachos’ belonging to the central ChacoForests, and ‘Quebrachal’ of two ‘quebrachos’ and‘Palosantales’ of Bulnesia sarmientoi Lorentz ex Griseb.both belonging to the western Chaco Forests, sensuPrado (1993). The two former units are characterisedby the dominance of Schinopsis lorentzii Engl. (Anacar-diaceae), Schinopsis balansae Engl. and Aspidospermaquebracho-blanco Schldtl. (Apocynaceae), and thelatter by Bulnesia sarmientoi Lorentz ex Griseb.(Zygophyllaceae). Other floristic elements are Ziziphusmistol Griseb. (Rhamnaceae), Sideroxylon obtusifolium(Roem. et Schult.) T.D. Penn. (Sapotaceae), severalspecies of Prosopis (Fabaceae, Mimosoideae), Caesal-
pinia paraguariensis (D. Parodi) Burkart (Fabaceae,Caesalpinioideae), Pterogyne nitens Tul. (Fabaceae,Caesalpinioideae), Tabebuia nodosa (Griseb.) Griseb.(Bignoniaceae), Aspidosperma triternatum Rojas Acosta(Apocynaceae), Trithrinax schizophylla Drude (Are-caceae), Castela coccinea Griseb. (Simaroubaceae),Achatocarpus praecox Griseb. (Achatocarpaceae), Celtisspp. (Celtidaceae), Schinus fasciculatus (Griseb.) I.M.Johnst. var. arenicola (Hauman) F.A. Barkley (Anacar-diaceae), Capparis atamisquea Kuntze (Capparidaceae),Maytenus vitis-idaea Griseb. (Celastraceae), Moya spi-nosa Griseb. (Celastraceae), Mimosa detinens Benth.(Fabaceae, Mimosoideae), Stetsonia coryne (Salm-Dyck) Britton et Rose (Cactaceae) and several speciesof Capparis (Capparidaceae) and Acacia (Fabaceae,Mimosoideae) (Prado 1993; Vossler 2013).
Pollen sampling and processing
Pollen masses (the pollen content of each cerumenpot) were extracted from one pot if it was completeor closed or a few if they were broken and/or opened.As each nest is an independent sample, pollenmasses taken from the same nest were consideredsubsamples.A total of 86 subsamples of pollen masses from
ten nests were analysed for Tetragonisca fiebrigi
Figure 1. Map depicting the distribution of the subtropical Chaco forest of South America indicating the sites where nest pollen sampleswere obtained. 1, Juan José Castelli (25° 56′ S; 60° 37′ W); 2, Villa Río Bermejito (25° 37′ S; 60° 15′ W); 3, Miraflores (25° 29′ S; 61° 01′W); 4, El Espinillo (25° 24′ S; 60° 27′ W); 5, El Sauzalito (24° 24′ S; 61° 40′ W).
Pollen grains in Chaquenian Meliponini diet 69
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015
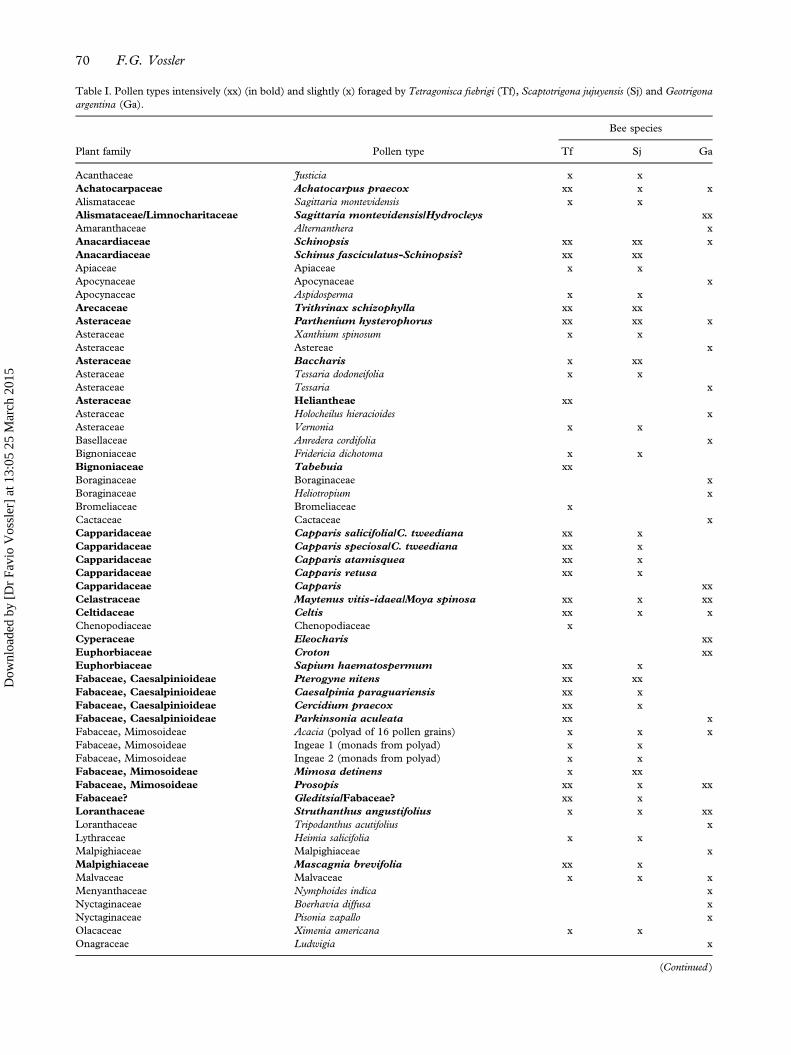
Table I. Pollen types intensively (xx) (in bold) and slightly (x) foraged by Tetragonisca fiebrigi (Tf), Scaptotrigona jujuyensis (Sj) and Geotrigonaargentina (Ga).
Bee species
Plant family Pollen type Tf Sj Ga
Acanthaceae Justicia x xAchatocarpaceae Achatocarpus praecox xx x xAlismataceae Sagittaria montevidensis x xAlismataceae/Limnocharitaceae Sagittaria montevidensis/Hydrocleys xxAmaranthaceae Alternanthera xAnacardiaceae Schinopsis xx xx xAnacardiaceae Schinus fasciculatus-Schinopsis? xx xxApiaceae Apiaceae x xApocynaceae Apocynaceae xApocynaceae Aspidosperma x xArecaceae Trithrinax schizophylla xx xxAsteraceae Parthenium hysterophorus xx xx xAsteraceae Xanthium spinosum x xAsteraceae Astereae xAsteraceae Baccharis x xxAsteraceae Tessaria dodoneifolia x xAsteraceae Tessaria xAsteraceae Heliantheae xxAsteraceae Holocheilus hieracioides xAsteraceae Vernonia x xBasellaceae Anredera cordifolia xBignoniaceae Fridericia dichotoma x xBignoniaceae Tabebuia xxBoraginaceae Boraginaceae xBoraginaceae Heliotropium xBromeliaceae Bromeliaceae xCactaceae Cactaceae xCapparidaceae Capparis salicifolia/C. tweediana xx xCapparidaceae Capparis speciosa/C. tweediana xx xCapparidaceae Capparis atamisquea xx xCapparidaceae Capparis retusa xx xCapparidaceae Capparis xxCelastraceae Maytenus vitis-idaea/Moya spinosa xx x xxCeltidaceae Celtis xx x xChenopodiaceae Chenopodiaceae xCyperaceae Eleocharis xxEuphorbiaceae Croton xxEuphorbiaceae Sapium haematospermum xx xFabaceae, Caesalpinioideae Pterogyne nitens xx xxFabaceae, Caesalpinioideae Caesalpinia paraguariensis xx xFabaceae, Caesalpinioideae Cercidium praecox xx xFabaceae, Caesalpinioideae Parkinsonia aculeata xx xFabaceae, Mimosoideae Acacia (polyad of 16 pollen grains) x x xFabaceae, Mimosoideae Ingeae 1 (monads from polyad) x xFabaceae, Mimosoideae Ingeae 2 (monads from polyad) x xFabaceae, Mimosoideae Mimosa detinens x xxFabaceae, Mimosoideae Prosopis xx x xxFabaceae? Gleditsia/Fabaceae? xx xLoranthaceae Struthanthus angustifolius x x xxLoranthaceae Tripodanthus acutifolius xLythraceae Heimia salicifolia x xMalpighiaceae Malpighiaceae xMalpighiaceae Mascagnia brevifolia xx xMalvaceae Malvaceae x x xMenyanthaceae Nymphoides indica xNyctaginaceae Boerhavia diffusa xNyctaginaceae Pisonia zapallo xOlacaceae Ximenia americana x xOnagraceae Ludwigia x
(Continued )
70 F.G. Vossler
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015
(Schwarz), 18 subsamples from four nests forScaptotrigona jujuyensis (Schrottky), and 63 fromfour nests for Geotrigona argentina (Vossler et al.2010; Vossler 2013). Pollen types were classified intwo categories according to their abundance: ≤ 10%and > 10% (Table I) following Ramalho et al.(1985), Cortopassi-Laurino and Ramalho (1988),and Ramalho (1990). A total of 32 pollen typeswere intensively foraged (> 10%) by the threeMeliponini species studied. Tetragonisca fiebrigiintensively foraged on 24 out of the 52 foraged pol-len types, S. jujuyensis on seven out of 45 and G.argentina on nine out of 41 (Table I). From the 78pollen types identified in all subsamples, 37 of themwere taken at random for both diameter and volumemeasurement.
Pollen type nomenclature follows the recommen-dations of Birks (1973), Birks and Birks (1980),Joosten and De Klerk (2002), and De Klerk andJoosten (2007). In most cases, pollen volumes andsize were measured from reference pollen and there-fore authority was not deleted.
The reference pollen collection was made fromflower buds of plant species collected in variouslocalities from the Chaco province of Argentina(Juan José Castelli [25° 56′ S; 60° 37′ W], Villa RíoBermejito [25° 37′ S; 60° 15′W], El Espinillo [25°24′ S; 60° 27′ W], Miraflores [25° 29′ S; 61° 01′ W]and El Sauzalito [24° 24′ S; 61° 40′W]; Figure 1).
The plant specimens were pressed, dried, identifiedby the author and deposited in the herbaria of theMuseo de La Plata (LP), La Plata, and the MuseoArgentino de Ciencias Naturales ‘BernardinoRivadavia’ (BA), Buenos Aires, Argentina. Plantnomenclature follows Zuloaga et al. (2008). Freshpollen grains from flowers were dyed using fuchsine(but not acetolysed), mounted on slides using a gly-cerine-gelatine mixture and measured under a NikonEclipse E200 light microscope at 1000× magnifica-tion. When pollen from flowers was not available,pollen grains from the colonies were measured. Beeswere identified by Arturo Roig-Alsina and depositedin the Entomology Collection of the MuseoArgentino de Ciencias Naturales ‘BernardinoRivadavia’.
Categories of volume and size of pollen grains
Ten to 15 measurements of both polar and equator-ial axes of pollen grain cytoplasms per each referenceplant were taken. The measurements were fromequatorial view for spherical and ellipsoidal shapes,and from two different views for prism shapes.The pollen size categories were taken from
Wodehouse (1935) and Hesse et al. (2009), beingvery small (< 10 µm), small (10–25 µm), medium(26–50 µm), large (51–100 µm) and very large(> 100 µm). To allocate a particular pollen type to
Table I. (Continued ).
Bee species
Plant family Pollen type Tf Sj Ga
Polygonaceae Polygonum x xPolygonaceae Ruprechtia triflora x xPortulacaceae Portulaca xRhamnaceae Ziziphus mistol xx x xxSantalaceae Acanthosyris falcata x xSapindaceae Urvillaea chacöensis x xSapotaceae Sideroxylon obtusifolium xx x xScrophulariaceae Scoparia xSimaroubaceae Castela coccinea x x xxTyphaceae Typha xUlmaceae Phyllostylon rhamnoides xxVerbenaceae Phyla reptans x x xVerbenaceae Verbena xZygophyllaceae Bulnesia sarmientoi x? Tetrad x x? Unidentified 1 x x? Unidentified 2 x x? Unidentified 3 x? Unidentified 4 x
Number of pollen types ≤ 10% (x) 28 38 32Number of pollen types > 10% (xx) 24 7 9Total number of pollen types per bee species 52 45 41
Note: The number of foraged pollen types varied among these species. These three bee species foraged intensively on a total of 32 pollen types.
Pollen grains in Chaquenian Meliponini diet 71
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015

each size category, the longest value of any of the twocytoplasmic diameters was used to establish the cate-gories of whole pollen grains (cytoplasm plus walls),as suggested by Hesse et al. (2009).
To estimate the cytoplasmic volume, pollen grainswere compared to similar geometric shapes: sphere,ellipsoid and triangular base prism. Based on the sizeclasses, five pollen volume categories are used in thepresent study: very small volume (< 524 µm3), smallvolume (524–8181 µm3), medium volume (8182–65 448 µm3), large volume (65 449–523 583 µm3) andvery large volume (> 523 583 µm3). The calculationsof volume categories are based on spherical pollengrains.
Pollen shapes
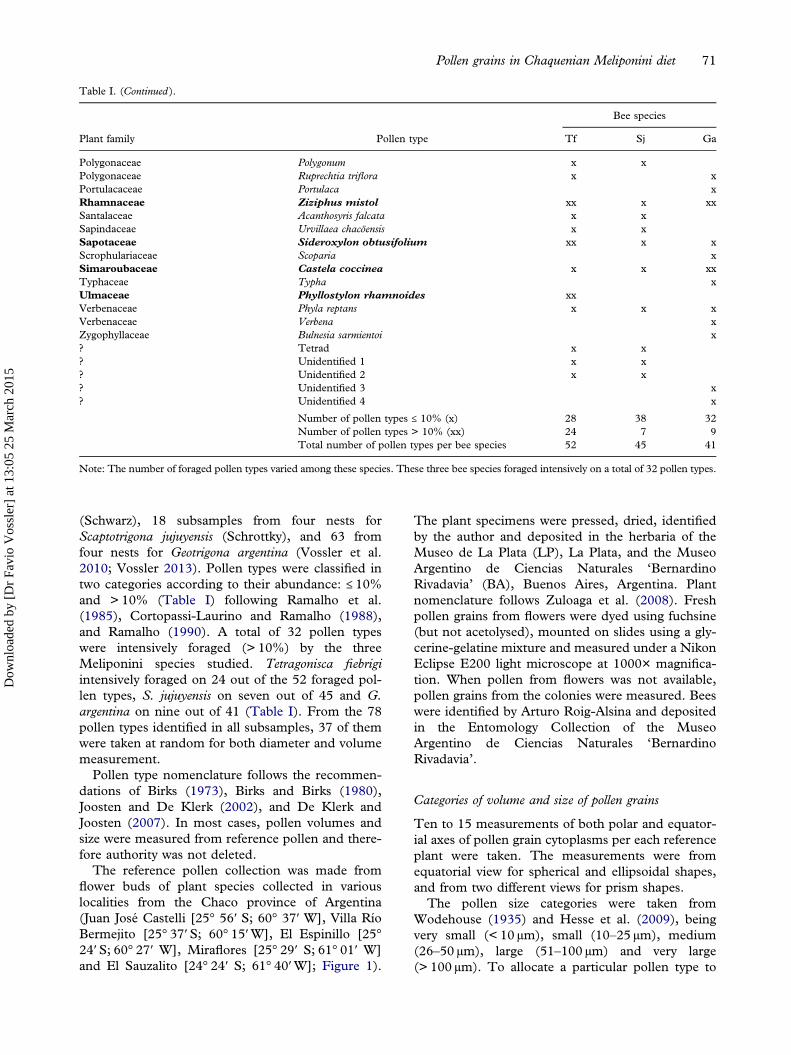
All shapes found in the samples were identified andmeasured in equatorial view as follows (seeFigure 2A).
Prolate ellipsoid (and prolate ellipsoid polyads) (Figure2A, B). — Main axis p (polar axis) and two minoraxis e of the same length (equatorial axes; e.g.Pterogyne nitens and Mimosa detinens polyad). Theformula is 1/6 π p e2. In the same way, the axis p ofthe prolate ellipsoid polyad was considered the axisof greater length, while e the minor axes.
Flat oblate ellipsoid (and flat polyads) (Figure2A, B). — Minor axis p (polar axis) and two axes e(equatorial axes), they can be of the same length (e2;e.g. Acacia praecox Griseb. and Acacia sp. polyads), orof different lengths (e1 and e2; e.g. polyad ofEnterolobium contortisiliquum (Vell.) Morong). The for-mulae are 1/6 π p e2 or 1/6 π p e1 e2, respectively. In flat
polyads, their thickness (corresponding to the minoraxis) was considered as p axis, while both other majoraxis (height and width of the polyad) were consideredas e axes (or e1 and e2 if they are different in length).
Oblate ellipsoid (Figure 2A). — Similar to prolateellipsoid shape with a rotation of 180°. The mainaxis e (long equatorial axis) and two axes p of thesame length (polar axis and short equatorial axis; e.g.Salvia). The formula is 1/6 π e p2.
Sphere (Figure 2A). — A polar axis p and an equa-torial axis e of approximately the same length (e.g.Polygonum). The ratio between the mean values ofthese two axes must be within the 0.99 to 1.01 range.The formula is 1/6 π p3. In cases where spherical andprolate or flat oblate ellipsoidal pollen grains werefound in the same pollen type, the prolate ellipsoidformula was applied (Figure 2A).
Triangular base prism (Figure 2A). — Being s any ofthe base sides and h the height of the prism(O’Rourke & Buchmann 1991; e.g. Acanthosyris fal-cata Griseb). As both values could not be measuredin the same grain, s and h were measured in differentgrains. The formula is ¼ s2 h √3.
Other shapes such as cubic shapes or those foundin heteropolar pollen grains were not present amongthe pollen grains foraged by Meliponini bees fromthe Dry Chaco forest. Although the oblate ellipsoidshape was not found in the samples, it was describedto avoid mistakes in volume calculations as it is verysimilar to the prolate ellipsoid shape. However, incases where the oblate ellipsoid shape is present, itsvolume should be calculated according to Figure 2A
Figure 2. A. Pollen shapes (a–e) found in the pollen samples studied. Location of axis e and p (of ellipsoidal and spherical shapes) and h ands (of prismatic shape) is provided. B. Scanning photo-micrograph of grains found in pollen masses of Meliponini bees, showing differentshapes and sizes. Scale bar – 50 µm.
72 F.G. Vossler
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015
(c). In the same way, Enterolobium contortisiliquum(Vell.) Morong was not found among pollen samplesas a whole polyad, but only as isolated monads.However, due to the fact that this acalymmate polyadcould be broken during foraging or palynologicalprocessing, the formula to describe flat polyads isgiven.
Flower size
Flower measurements were made on those plantspecies whose pollen grains were measured for sizeand volume. Maximum size of flowers was estab-lished mainly from fresh flowers and photographstaken during fieldwork as well as from literature onthe local flora. According to their maximum size,flowers were classified as either small (≤ 1 cm), med-ium (1–2 cm) or large (≥ 2 cm).
Statistical analysis
Correlation (Pearson coefficient) and linear regres-sion analyses were used to examine the dataset.Shapiro–Wilk and Kolmogorov–Smirnov tests fornormality were applied to the data set. Mean pollenvolume, maximum pollen size and maximum flowersize were log-transformed prior to analysis to meetthe assumptions of parametric tests. To investigatewhether the distribution of the pollen types byvolume/size category differed in nest stores and for-est, the null hypothesis was that there is no differencebetween the two distributions. This was tested using
a χ2 goodness-of-fit test, at 5% significance level.The categories very small and very large had anexpected frequency count of less than five. For thisreason, they had to be added to small and large,respectively, as required by this test. For both statis-tical analyses, the program package Statgraphics®Centurion XVI (StatPoint Technologies 2010) wasused.
Results
Volume range of the pollen foraged by Meliponini bees inthe Dry Chaco forest
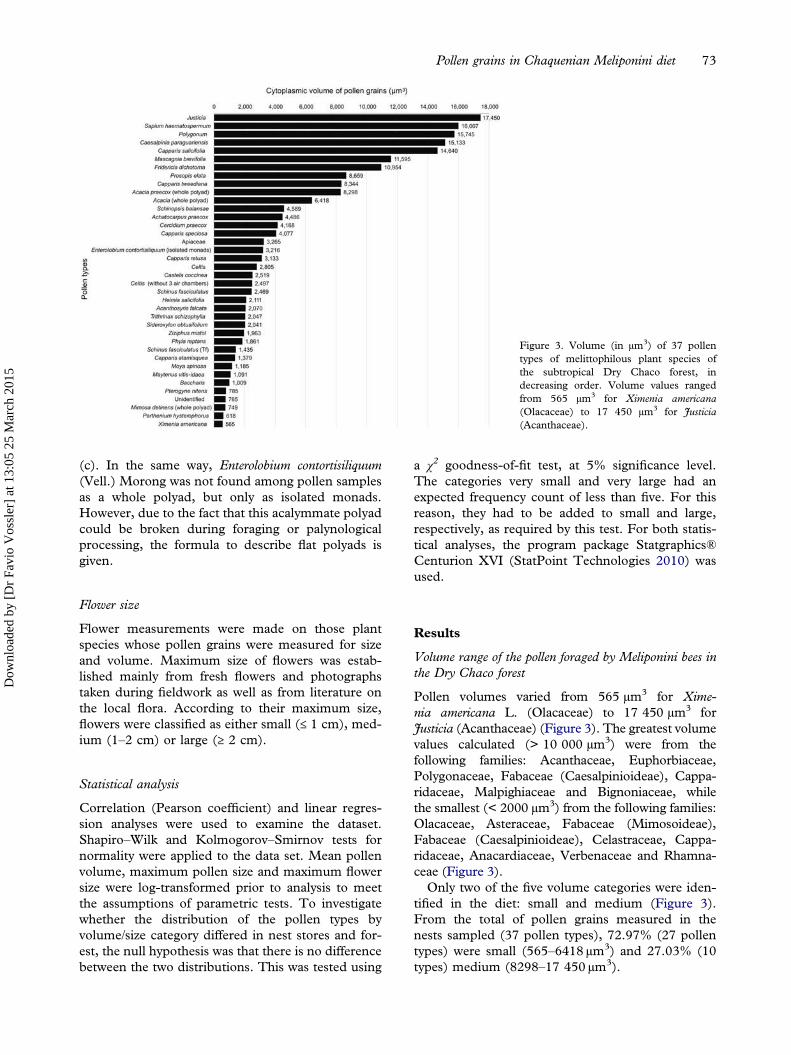
Pollen volumes varied from 565 µm3 for Xime-nia americana L. (Olacaceae) to 17 450 µm3 forJusticia (Acanthaceae) (Figure 3). The greatest volumevalues calculated (> 10 000 µm3) were from thefollowing families: Acanthaceae, Euphorbiaceae,Polygonaceae, Fabaceae (Caesalpinioideae), Cappa-ridaceae, Malpighiaceae and Bignoniaceae, whilethe smallest (< 2000 µm3) from the following families:Olacaceae, Asteraceae, Fabaceae (Mimosoideae),Fabaceae (Caesalpinioideae), Celastraceae, Cappa-ridaceae, Anacardiaceae, Verbenaceae and Rhamna-ceae (Figure 3).Only two of the five volume categories were iden-
tified in the diet: small and medium (Figure 3).From the total of pollen grains measured in thenests sampled (37 pollen types), 72.97% (27 pollentypes) were small (565–6418 µm3) and 27.03% (10types) medium (8298–17 450 µm3).
Figure 3. Volume (in µm3) of 37 pollentypes of melittophilous plant species ofthe subtropical Dry Chaco forest, indecreasing order. Volume values rangedfrom 565 µm3 for Ximenia americana(Olacaceae) to 17 450 µm3 for Justicia(Acanthaceae).
Pollen grains in Chaquenian Meliponini diet 73
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015
Pollen volume of the intensively foraged pollen types
The volume values of 25 out of the 32 intensivelyforaged pollen types ranged from 618 µm3 forParthenium hysterophorus L. (Asteraceae) (smallvolume) to 16 007 µm3 for Sapium haematospermumMüll. Arg. (Euphorbiaceae) (medium volume).When only those pollen types that were intensivelyforaged (> 10% frequency) are considered, smallpollen volumes are also highly represented (76%)(19 pollen types).
Pollen size of the foraged pollen grains
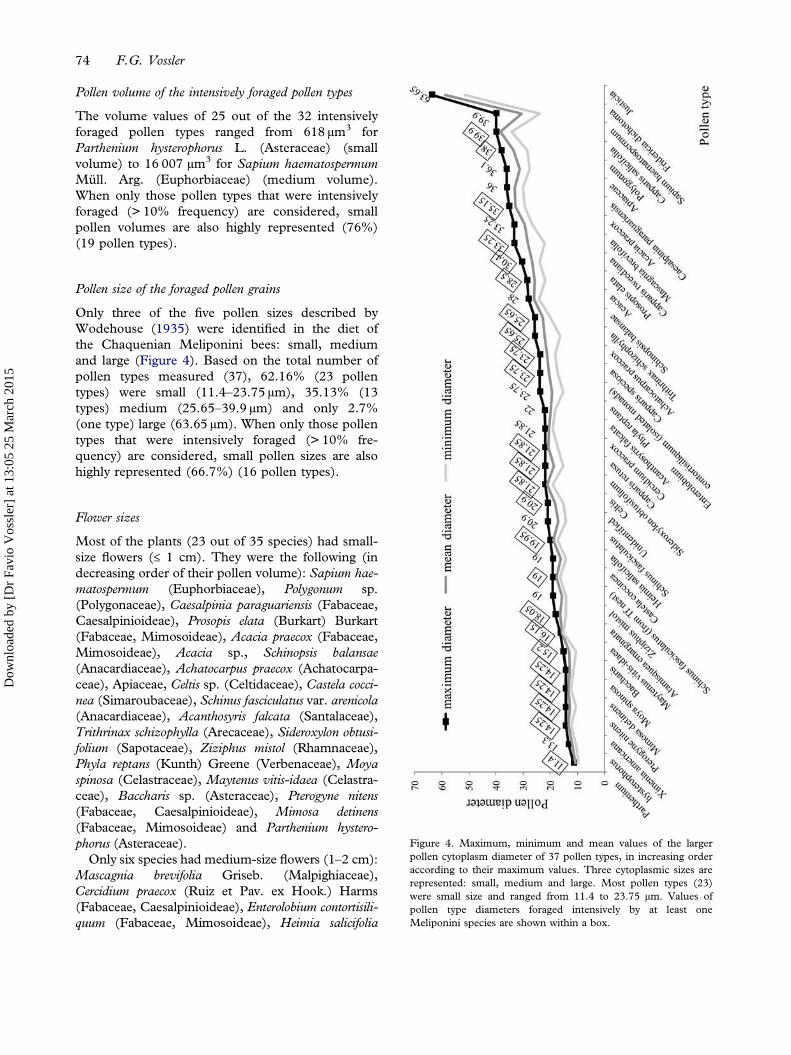
Only three of the five pollen sizes described byWodehouse (1935) were identified in the diet ofthe Chaquenian Meliponini bees: small, mediumand large (Figure 4). Based on the total number ofpollen types measured (37), 62.16% (23 pollentypes) were small (11.4–23.75 µm), 35.13% (13types) medium (25.65–39.9 µm) and only 2.7%(one type) large (63.65 µm). When only those pollentypes that were intensively foraged (> 10% fre-quency) are considered, small pollen sizes are alsohighly represented (66.7%) (16 pollen types).
Flower sizes
Most of the plants (23 out of 35 species) had small-size flowers (≤ 1 cm). They were the following (indecreasing order of their pollen volume): Sapium hae-matospermum (Euphorbiaceae), Polygonum sp.(Polygonaceae), Caesalpinia paraguariensis (Fabaceae,Caesalpinioideae), Prosopis elata (Burkart) Burkart(Fabaceae, Mimosoideae), Acacia praecox (Fabaceae,Mimosoideae), Acacia sp., Schinopsis balansae(Anacardiaceae), Achatocarpus praecox (Achatocarpa-ceae), Apiaceae, Celtis sp. (Celtidaceae), Castela cocci-nea (Simaroubaceae), Schinus fasciculatus var. arenicola(Anacardiaceae), Acanthosyris falcata (Santalaceae),Trithrinax schizophylla (Arecaceae), Sideroxylon obtusi-folium (Sapotaceae), Ziziphus mistol (Rhamnaceae),Phyla reptans (Kunth) Greene (Verbenaceae), Moyaspinosa (Celastraceae), Maytenus vitis-idaea (Celastra-ceae), Baccharis sp. (Asteraceae), Pterogyne nitens(Fabaceae, Caesalpinioideae), Mimosa detinens(Fabaceae, Mimosoideae) and Parthenium hystero-phorus (Asteraceae).
Only six species had medium-size flowers (1–2 cm):Mascagnia brevifolia Griseb. (Malpighiaceae),Cercidium praecox (Ruiz et Pav. ex Hook.) Harms(Fabaceae, Caesalpinioideae), Enterolobium contortisili-quum (Fabaceae, Mimosoideae), Heimia salicifolia
Figure 4. Maximum, minimum and mean values of the largerpollen cytoplasm diameter of 37 pollen types, in increasing orderaccording to their maximum values. Three cytoplasmic sizes arerepresented: small, medium and large. Most pollen types (23)were small size and ranged from 11.4 to 23.75 µm. Values ofpollen type diameters foraged intensively by at least oneMeliponini species are shown within a box.
74 F.G. Vossler
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015
(Kunth) Link (Lythraceae), Capparis atamisquea(Capparidaceae) and Ximenia americana (Olacaceae).
Six had large-size flowers (≥ 2 cm): Justicia sp.(Acanthaceae), Capparis salicifolia Griseb. (Cappari-daceae), Fridericia dichotoma (Jacq.) L.G. Lohmann(Bignoniaceae), Capparis tweediana Eichler, Capparisspeciosa Griseb. and Capparis retusa Griseb.
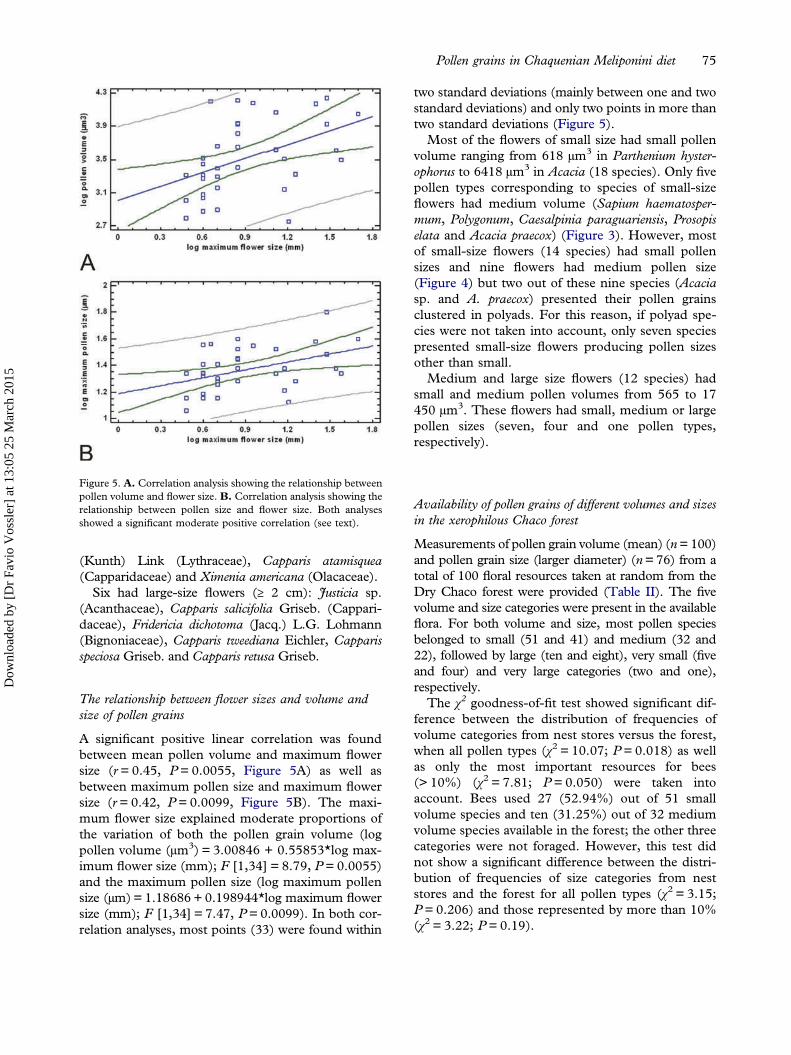
The relationship between flower sizes and volume andsize of pollen grains
A significant positive linear correlation was foundbetween mean pollen volume and maximum flowersize (r = 0.45, P = 0.0055, Figure 5A) as well asbetween maximum pollen size and maximum flowersize (r= 0.42, P = 0.0099, Figure 5B). The maxi-mum flower size explained moderate proportions ofthe variation of both the pollen grain volume (logpollen volume (µm3) = 3.00846 + 0.55853*log max-imum flower size (mm); F [1,34] = 8.79, P = 0.0055)and the maximum pollen size (log maximum pollensize (µm) = 1.18686 + 0.198944*log maximum flowersize (mm); F [1,34] = 7.47, P= 0.0099). In both cor-relation analyses, most points (33) were found within
two standard deviations (mainly between one and twostandard deviations) and only two points in more thantwo standard deviations (Figure 5).Most of the flowers of small size had small pollen
volume ranging from 618 µm3 in Parthenium hyster-ophorus to 6418 µm3 in Acacia (18 species). Only fivepollen types corresponding to species of small-sizeflowers had medium volume (Sapium haematosper-mum, Polygonum, Caesalpinia paraguariensis, Prosopiselata and Acacia praecox) (Figure 3). However, mostof small-size flowers (14 species) had small pollensizes and nine flowers had medium pollen size(Figure 4) but two out of these nine species (Acaciasp. and A. praecox) presented their pollen grainsclustered in polyads. For this reason, if polyad spe-cies were not taken into account, only seven speciespresented small-size flowers producing pollen sizesother than small.Medium and large size flowers (12 species) had
small and medium pollen volumes from 565 to 17450 µm3. These flowers had small, medium or largepollen sizes (seven, four and one pollen types,respectively).
Availability of pollen grains of different volumes and sizesin the xerophilous Chaco forest
Measurements of pollen grain volume (mean) (n=100)and pollen grain size (larger diameter) (n=76) from atotal of 100 floral resources taken at random from theDry Chaco forest were provided (Table II). The fivevolume and size categories were present in the availableflora. For both volume and size, most pollen speciesbelonged to small (51 and 41) and medium (32 and22), followed by large (ten and eight), very small (fiveand four) and very large categories (two and one),respectively.The χ2 goodness-of-fit test showed significant dif-
ference between the distribution of frequencies ofvolume categories from nest stores versus the forest,when all pollen types (χ2 = 10.07; P = 0.018) as wellas only the most important resources for bees(> 10%) (χ2 = 7.81; P = 0.050) were taken intoaccount. Bees used 27 (52.94%) out of 51 smallvolume species and ten (31.25%) out of 32 mediumvolume species available in the forest; the other threecategories were not foraged. However, this test didnot show a significant difference between the distri-bution of frequencies of size categories from neststores and the forest for all pollen types (χ2 = 3.15;P = 0.206) and those represented by more than 10%(χ2 = 3.22; P = 0.19).
Figure 5. A. Correlation analysis showing the relationship betweenpollen volume and flower size. B. Correlation analysis showing therelationship between pollen size and flower size. Both analysesshowed a significant moderate positive correlation (see text).
Pollen grains in Chaquenian Meliponini diet 75
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015
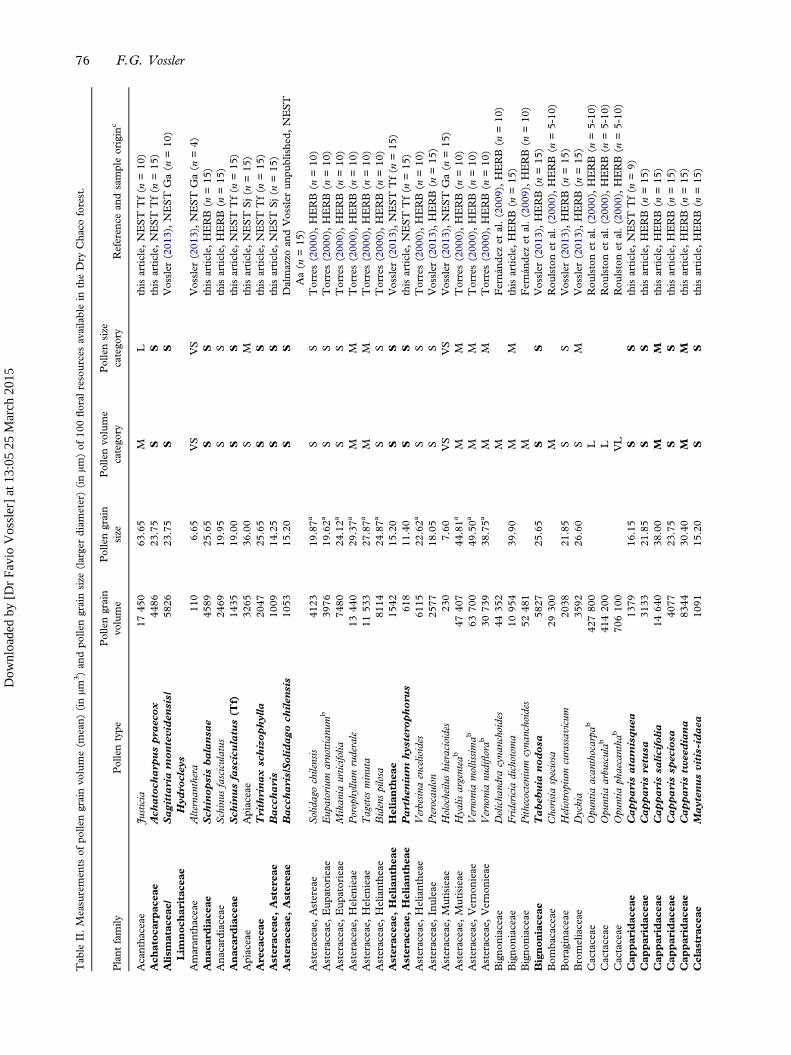
Tab
leII.Measuremen
tsof
polle
ngrainvo
lume(m
ean)
(inµm
3)an
dpo
llengrainsize
(largerdiam
eter)(inµm
)of
100floral
resourcesavailablein
theDry
Cha
coforest.
Plant
family
Pollentype
Pollengrain
volume
Pollengrain
size
Pollenvo
lume
catego
ryPollensize
catego
ryReferen
cean
dsampleorigin
c
Acantha
ceae
Justicia
1745
063
.65
ML
thisarticle,
NEST
Tf(n
=10
)Ach
atoc
arpac
eae
Ach
atoc
har
puspra
ecox
4486
23.75
SS
thisarticle,
NEST
Tf(n
=15
)Alism
atac
eae/
Lim
noc
har
itac
eae
Sag
ittariam
onteviden
sis/
Hyd
rocley
s58
2623
.75
SS
Vossler
(201
3),NEST
Ga(n
=10
)
Amaran
thaceae
Alternan
thera
110
6.65
VS
VS
Vossler
(201
3),NEST
Ga(n
=4)
Anac
ardiace
aeSch
inop
sisbalan
sae
4589
25.65
SS
thisarticle,
HERB
(n=15
)Ana
cardiaceae
Schinus
fasciculatus
2469
19.95
SS
thisarticle,
HERB
(n=15
)Anac
ardiace
aeSch
inusfasciculatus(T
f)14
3519
.00
SS
thisarticle,
NEST
Tf(n
=15
)Apiaceae
Apiaceae
3265
36.00
SM
thisarticle,
NEST
Sj(n
=15
)Areca
ceae
Trithrinax
schizop
hylla
2047
25.65
SS
thisarticle,
NEST
Tf(n
=15
)Asterac
eae,
Asterea
eBac
char
is10
0914
.25
SS
thisarticle,
NEST
Sj(n
=15
)Asterac
eae,
Asterea
eBac
char
is/Solidag
och
ilen
sis
1053
15.20
SS
Dalmazzo
andVossler
unpu
blishe
d,NEST
Aa(n
=15
)Asteraceae,
Astereae
Solidagochilensis
4123
19.87a
SS
Torres(200
0),HERB
(n=10
)Asteraceae,
Eup
atorieae
Eup
atorium
arnottian
umb
3976
19.62a
SS
Torres(200
0),HERB
(n=10
)Asteraceae,
Eup
atorieae
Mikan
iaurticifolia
7480
24.12a
SS
Torres(200
0),HERB
(n=10
)Asteraceae,
Helen
ieae
Porophyllum
ruderale
1344
029
.37a
MM
Torres(200
0),HERB
(n=10
)Asteraceae,
Helen
ieae
Tagetes
minuta
1153
327
.87a
MM
Torres(200
0),HERB
(n=10
)Asteraceae,
Helianthe
aeBidenspilosa
8114
24.87a
SS
Torres(200
0),HERB
(n=10
)Asterac
eae,
Helianthea
eHelianthea
e15
4215
.20
SS
Vossler
(201
3),NEST
Tf(n
=15
)Asterac
eae,
Helianthea
ePar
then
ium
hys
tero
phor
us
618
11.40
SS
thisarticle,
NEST
Tf(n
=15
)Asteraceae,
Helianthe
aeVerbesina
encelioides
6115
22.62a
SS
Torres(200
0),HERB
(n=10
)Asteraceae,
Inuleae
Pterocaulon
2577
18.05
SS
Vossler
(201
3),HERB
(n=15
)Asteraceae,
Mutisieae
Holocheilu
shieracioides
230
7.60
VS
VS
Vossler
(201
3),NEST
Ga(n
=15
)Asteraceae,
Mutisieae
Hyalis
argentea
b47
407
44.81a
MM
Torres(200
0),HERB
(n=10
)Asteraceae,
Verno
nieae
Vernoniamollissimab
6370
049
.50a
MM
Torres(200
0),HERB
(n=10
)Asteraceae,
Verno
nieae
Vernonianu
diflorab
3073
938
.75a
MM
Torres(200
0),HERB
(n=10
)Bigno
niaceae
Dolicha
ndra
cyna
nchoides
4435
2M
Ferná
ndez
etal.(200
9),HERB
(n=10
)Bigno
niaceae
Fridericiadichotom
a10
954
39.90
MM
thisarticle,
HERB
(n=15
)Bigno
niaceae
Pith
ecoctenium
cyna
nchoides
5248
1M
Ferná
ndez
etal.(200
9),HERB
(n=10
)Bignon
iace
aeTab
ebuia
nod
osa
5827
25.65
SS
Vossler
(201
3),HERB
(n=15
)Bom
bacaceae
Chorisiaspeciosa
2930
0M
Rou
lstonet
al.(200
0),HERB
(n=5-10
)Boraginaceae
Heliotropium
curassav
icum
2038
21.85
SS
Vossler
(201
3),HERB
(n=15
)Bromeliaceae
Dyckia
3592
26.60
SM
Vossler
(201
3),HERB
(n=15
)Cactaceae
Opu
ntia
acan
thocarpa
b42
780
0L
Rou
lstonet
al.(200
0),HERB
(n=5-10
)Cactaceae
Opu
ntia
arbusculab
41420
0L
Rou
lstonet
al.(200
0),HERB
(n=5-10
)Cactaceae
Opu
ntia
phaecantha
b70
610
0VL
Rou
lstonet
al.(200
0),HERB
(n=5-10
)Cap
par
idac
eae
Cap
par
isatam
isqu
ea13
7916
.15
SS
thisarticle,
NEST
Tf(n
=9)
Cap
par
idac
eae
Cap
par
isretusa
3133
21.85
SS
thisarticle,
HERB
(n=15
)Cap
par
idac
eae
Cap
par
issa
licifolia
1464
038
.00
MM
thisarticle,
HERB
(n=15
)Cap
par
idac
eae
Cap
par
issp
eciosa
4077
23.75
SS
thisarticle,
HERB
(n=15
)Cap
par
idac
eae
Cap
par
istw
eediana
8344
30.40
MM
thisarticle,
HERB
(n=15
)Celastrac
eae
May
tenusvitis-idae
a10
9115
.20
SS
thisarticle,
HERB
(n=15
)
76 F.G. Vossler
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015

Celastrac
eae
Moy
asp
inos
a11
8514
.25
SS
thisarticle,
HERB
(n=13
)Celtidac
eae
Celtis
2805
20.90
SS
thisarticle,
NEST
Tf(n
=15
)Con
volvulaceae
Evolvulus
sericeus
2967
9M
Ferná
ndez
etal.(200
9),HERB
(n=10
)Con
volvulaceae
Ipom
oeaalba
109
514
414
4.40
VL
VL
Dalmazzo
andVossler
unpu
blishe
d,NEST
Aa(n
=15
)Con
volvulaceae
Ipom
oeacairica
6247
651
.30
ML
Dalmazzo
andVossler
unpu
blishe
d,NEST
Aa(n
=15
)Con
volvulaceae
Ipom
oeahieronym
ib23
170
6L
Ferná
ndez
etal.(200
9),HERB
(n=10
)Cyp
erac
eae
Eleoc
har
is47
013
.30
VS
SVossler
(201
3),NEST
Ga(n
=10
)Euphor
biace
aeCro
ton
3231
845
.60
MM
Vossler
(201
3),NEST
Ga(n
=15
)Eup
horbiaceae
Croton
7015
351
.17a
LL
Silv
eira
1991
),NEST
Am
(n=25
)Eup
horbiaceae
Croton/Man
ihot
grah
amiib
23759
390
.25
LL
Dalmazzo
andVossler
unpu
blishe
d,NEST
Aa(n
=12
)Euphor
biace
aeSap
ium
hae
matos
permum
1600
739
.90
MM
thisarticle,
NEST
Tf(n
=15
)Fab
acea
e,Cae
salpinioidea
eCae
salpinia
par
aguar
iensis
1513
335
.15
MM
thisarticle,
HERB
(n=15
)
Fab
acea
e,Cae
salpinioidea
eCercidium
pra
ecox
4168
21.85
SS
thisarticle,
NEST
Tf(n
=9)
Fab
acea
e,Cae
salpinioidea
eGleditsiaam
orphoides
5213
26.60
SM
Vossler
(201
3),HERB
(n=15
)
Fab
acea
e,Cae
salpinioidea
ePtero
gyneniten
s78
514
.25
SS
thisarticle,
HERB
(n=15
)
Fab
aceae,
Caesalpinioideae
Senna
occidentalis
2190
0M
Rou
lstonet
al.(200
0),HERB
(n=5-10
)Fab
aceae,
Mim
osoide
aeAcaciapraecox
8298
33.25
MM
thisarticle,
HERB
(n=15
)Fab
aceae,
Mim
osoide
aeAcacia
6418
28.00
SM
thisarticle,
NEST
Sj(n
=15
)Fab
aceae,
Mim
osoide
aeAlbizia
inun
data
4625
766
.50
ML
Vossler
unpu
blishe
d,NEST
Mo(n
=7)
Fab
aceae,
Mim
osoide
aeEnterolobium
(isolatedmon
ad)
3216
23.75
SS
thisarticle,
HERB
(n=15
)Fab
aceae,
Mim
osoide
aeEnterolobium
(who
lepo
lyad
)11
143
286
.45
LL
thisarticle,
HERB
(n=8)
Fab
acea
e,Mim
osoidea
eMim
osadetinen
s74
914
.25
SS
thisarticle,
NEST
Sj(n
=15
)Fab
acea
e,Mim
osoidea
ePro
sopis
elata
8659
28.50
MM
thisarticle,
HERB
(n=15
)Fab
aceae,
Pap
ilion
oide
aeVicia
gran
diflorab
9100
MRou
lstonet
al.(200
0),HERB
(n=5-10
)Lam
iaceae
Salvia
6200
SRou
lstonet
al.(200
0),HERB
(n=5-10
)Lor
anth
acea
eStruthan
thusura
guen
sis
8266
34.20
MM
Vossler
(201
3),NEST
Ga(n
=10
)Lythraceae
Heimia
salicifolia
2111
19.00
SS
thisarticle,
HERB
(n=15
)M
alpighiace
aeMas
cagn
iabrevifolia
1159
533
.25
MM
thisarticle,
NEST
Tf(n
=15
)M
alvaceae
Gossypium
thurberib
19530
0L
Rou
lstonet
al.(200
0),HERB
(n=5-10
)M
alvaceae
Pav
onia
21315
180
.75
LL
Dalmazzo
andVossler
unpu
blishe
d,NEST
Aa(n
=8)
Malvaceae
Sph
aeralcea
ambiguab
4160
0M
Rou
lstonet
al.(200
0),HERB
(n=5-10
)M
alvaceae
Sph
aeralcea
cordobensisb
2380
7M
Ferná
ndez
etal.(200
9),HERB
(n=10
)Nyctagina
ceae
Pisonia
zapa
llo55
6628
.50
SM
Vossler
unpu
blishe
d,NEST
Mo(n
=15
)Olacaceae
Xim
enia
american
a56
513
.30
SS
thisarticle,
NEST
Tf(n
=7)
Passifloraceae
Passifloracaerulea
b70
997
LFerná
ndez
etal.(200
9),HERB
(n=10
)Plantaginaceae
Scopa
ria
465.70
VS
VS
Vossler
(201
3),NEST
Ga(n
=6)
Polygon
aceae
Polygonum
1574
536
.10
MM
thisarticle,
NEST
Sj(n
=15
)
(Contin
ued)
Pollen grains in Chaquenian Meliponini diet 77
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015
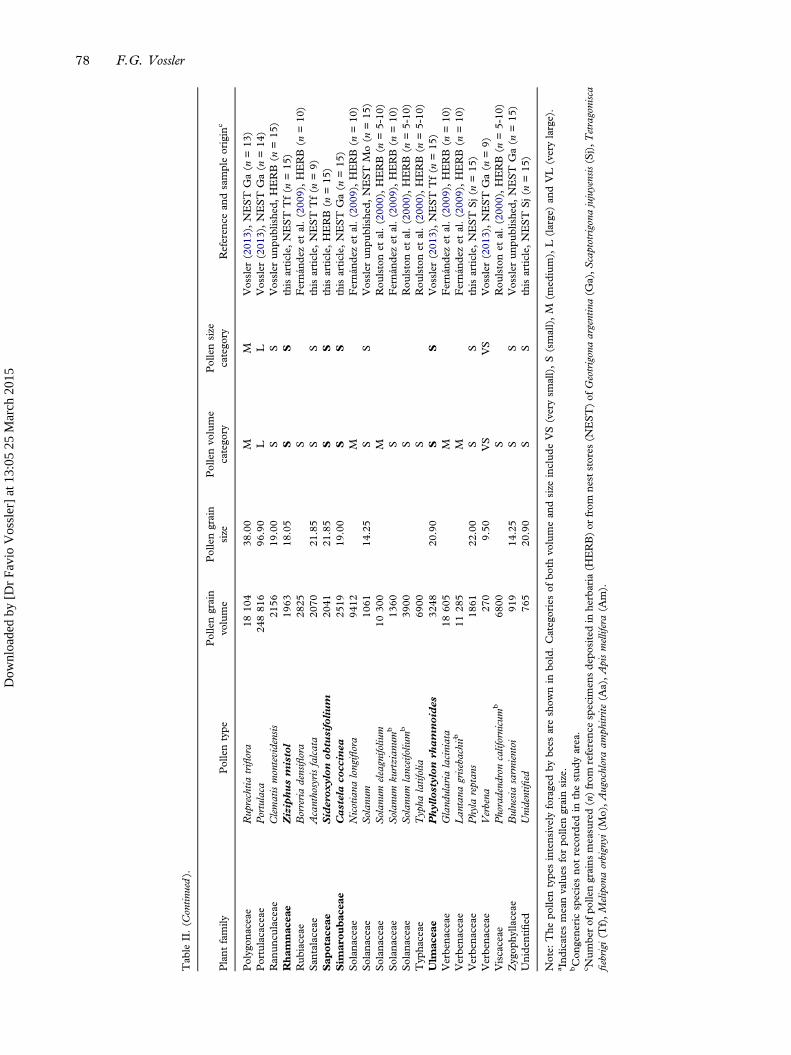
Tab
leII.(C
ontin
ued).
Plant
family
Pollentype
Pollengrain
volume
Pollengrain
size
Pollenvo
lume
catego
ryPollensize
catego
ryReferen
cean
dsampleorigin
c
Polygon
aceae
Rup
rechtia
triflora
1810
438
.00
MM
Vossler
(201
3),NEST
Ga(n
=13
)Portulacaceae
Portulaca
24881
696
.90
LL
Vossler
(201
3),NEST
Ga(n
=14
)Ran
uncu
laceae
Clematismontevidensis
2156
19.00
SS
Vossler
unpu
blishe
d,HERB
(n=15
)Rham
nac
eae
Ziziphusm
istol
1963
18.05
SS
thisarticle,
NEST
Tf(n
=15
)Rub
iaceae
Borreriadensiflora
2825
SFerná
ndez
etal.(200
9),HERB
(n=10
)San
talaceae
Acanthosyrisfalcata
2070
21.85
SS
thisarticle,
NEST
Tf(n
=9)
Sap
otac
eae
Siderox
ylon
obtusifolium
2041
21.85
SS
thisarticle,
HERB
(n=15
)Sim
arou
bac
eae
Cas
tela
coccinea
2519
19.00
SS
thisarticle,
NEST
Ga(n
=15
)Solan
aceae
Nicotiana
longiflora
9412
MFerná
ndez
etal.(200
9),HERB
(n=10
)Solan
aceae
Solan
um10
6114
.25
SS
Vossler
unpu
blishe
d,NEST
Mo(n
=15
)Solan
aceae
Solan
umeleagnifoliu
m10
300
MRou
lstonet
al.(200
0),HERB
(n=5-10
)Solan
aceae
Solan
umku
rtzian
umb
1360
SFerná
ndez
etal.(200
9),HERB
(n=10
)Solan
aceae
Solan
umlanceifoliu
mb
3900
SRou
lstonet
al.(200
0),HERB
(n=5-10
)Typ
haceae
Typha
latifolia
6900
SRou
lstonet
al.(200
0),HERB
(n=5-10
)Ulm
acea
ePhyllostylon
rham
noides
3248
20.90
SS
Vossler
(201
3),NEST
Tf(n
=15
)Verbe
naceae
Gland
ularia
laciniata
1860
5M
Ferná
ndez
etal.(200
9),HERB
(n=10
)Verbe
naceae
Lan
tana
grisebachiib
1128
5M
Ferná
ndez
etal.(200
9),HERB
(n=10
)Verbe
naceae
Phyla
reptan
s18
6122
.00
SS
thisarticle,
NEST
Sj(n
=15
)Verbe
naceae
Verbena
270
9.50
VS
VS
Vossler
(201
3),NEST
Ga(n
=9)
Viscaceae
Phoradend
roncalifornicum
b68
00S
Rou
lstonet
al.(200
0),HERB
(n=5-10
)Zygop
hylla
ceae
Bulnesiasarm
ientoi
919
14.25
SS
Vossler
unpu
blishe
d,NEST
Ga(n
=15
)Unide
ntified
Unidentified
765
20.90
SS
thisarticle,
NEST
Sj(n
=15
)
Note:
The
polle
ntype
sintensivelyforagedby
bees
areshow
nin
bold.Categoriesof
both
volumean
dsize
includ
eVS(verysm
all),S(small),M
(med
ium),L(large)an
dVL(verylarge).
a Ind
icates
meanvalues
forpo
llengrainsize.
bCon
gene
ricspeciesno
treco
rded
inthestud
yarea.
c Num
berof
polle
ngrains
measured(n)from
referenc
especim
ensde
positedin
herbaria
(HERB)or
from
neststores
(NEST)of
Geotrigonaargentina(G
a),S
captotrigona
jujuyensis(Sj),T
etragonisca
fiebrigi
(Tf),Meliponaorbignyi
(Mo),Augochloraam
phitrite
(Aa),Apismellifera(A
m).
78 F.G. Vossler
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015

Discussion
A wide spectrum of pollen types in colonies of threesubtropical Meliponini bees
Meliponini bees carry pollen grains in their corbi-cula, the most specialized structure for transportingpollen among bees (Michener et al. 1978).Corbiculate bees (tribes Apini, Meliponini,Bombini and Euglossini) as well as those with scopaewith sparse hairs moisten pollen grains with nectar.This enables them to increase the size of the pollenload beyond the scopal hair length and favours thecollection of a wide spectrum of pollen sizes (Thorp2000). Leioproctus (Colletidae) for example collectslarge pollen grains (80–90 µm) from Conospermum(Proteaceae) (Houston 1989). The fact thatMeliponini bees moisten their pollen loads with nec-tar could account for the great variety of pollen grainvolumes and sizes foraged by the three species stu-died in the Chaco forest. Thus, pollen from a broadspectrum of plant species of several families can besuccessfully foraged allowing these eusocial bees tosurvive in different seasons and environments.
Small bees, small flowers and small volume and size ofpollen grains
Although the three Meliponini species foraged on alarge number of plants, those of small volume andsize pollen grains were the most intensively foraged,followed by medium grains. Large pollen grains werescarcely gathered. Although large grains supplies lar-ger nutrient biomass (in volume) than small ones(O’Rourke & Buchmann 1991; Vossler et al. 2010),these bees concentrated their foraging on the latter.
It is known that pollen grain volume is positivelycorrelated to the pistil length of the flowers, due tothe important role of the cytoplasm nutrients in thegrowth of the pollen tube, essential to fertilise theovules (Schnack & Covas 1945; Torres 2000). Plantswith small pollen grains produce greater number ofgrains per anther than those with large grains (SubbaReddi & Reddi 1986; Tormo Molina et al. 1996),having a higher number of male gametes available tolocate a flower stigma. Meliponini bees have beenconsidered by some authors as poor pollinatorsbecause they moisten with nectar the transportedpollen load, reducing the chances of losing indivi-dual pollen grains during floral visitation and ofeventually making contact with a stigma (Thorp2000). However, the high number of small grainstransported per corbicula compared to large grainsizes should increase the chance of pollen beingtransferred to floral stigma surface during floral vis-its. Moreover, Meliponini bees appear to be ade-quate pollinators of small flowers clustered in dense
inflorescences, due to the fact that they forage slowlyon a high number of flowers in the same trip.The floral syndromes associated to the small- to
medium-sized Meliponini bees include those flowerssmall in size, with radial symmetry and with easyaccess to pollen and nectar, clustered in dense inflor-escences (Ramalho 1990, 2004). Although thesehighly generalist bees visit a broad spectrum offlower sizes (collecting a broad spectrum of pollenvolumes and sizes), they prefer small flower sizessimilar to their body size, being good candidates aspollinators for this kind of flowers (Ramalho 2004).From this scenario, it appears that the high abun-dance of small pollen grains stored in colonies of thethree studied Chaquenian species of Meliponini isdue to their preference for small flowers of shortpistils. Wilms et al. (1997) and Ramalho (2004)suggested that the tight relationship betweenMeliponini and small flowers clustered in rich inflor-escences is part of a diffuse co-evolutionary processand it seems to be the cause for the high abundanceof mass-flowering trees in Neotropical areas. In thepresent study, a significant positive linear correlationbetween maximum flower size and both mean pollenvolume and maximum pollen size was also found(Figure 5). This tight relationship betweenMeliponini bees and mass flowerings has beenobserved in tropical areas and now can also beextended to South American subtropical xerophilousDry Chaco forest.
Availability of pollen grains of different volumes and sizesin the xerophilous Chaco forest
As it has been observed for other Neotropical envir-onments, the eusocial Meliponini bees forage mainlyon the most locally abundant plants (Ramalho et al.1990) to supply their large colonies throughout theyear. This opportunistic foraging habit would suggestthat the large number of small pollen grains found inthe nest samples of the present study is due to theirhigh availability in this subtropical forest. Moreover, astudy on floral visitation of six Meliponini species inthis region showed that they mainly foraged on themost diverse plant families Fabaceae (subfamiliesMimosoideae and Caesalpinioideae) and Asteraceae(Vossler 2012).These Meliponini bees foraged mainly on the trees
and perennial woody shrubs from the forest, nearby totheir nests. Moreover, this woody vegetation provides ahigher number of flowers per individual compared toherbaceous plants from open areas and the lower stra-tumof the forest (grasslands, forest edges, swarms, etc.),where their preferred nesting substrates (large treetrunk) are scarce or absent. For instance, melitophilousmembers of Asteraceae, Malvaceae, Cactaceae,
Pollen grains in Chaquenian Meliponini diet 79
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015

Acanthaceae, Fabaceae (Papilionoideae), Bromeliaceaeand Convolvulaceae, among others, have large to verylarge pollen grains and were highly available in openareas and the lower stratum of the forest but poorlyrepresented or absent in nest pollen stores.
The comparison of volume categories between neststores and Chaco vegetation demonstrates that foragedpollen types do not follow the abundance patternfound in the available flora. This result indicates thatbees preferred certain volume categories (small andmedium volumes) that are present in small flowers ofshort pistils. This finding is in agreement with thehypothesis of Wilms et al. (1997) and Ramalho(2004) for Neotropical habitats. However, when thefive pollen size categories were compared between neststores and the Chaco flora, preference was not found.Therefore, the abundance of small pollen types in nestsamples could be interpreted as opportunism, i.e.Meliponini bees forage floral resources in proportionto their availability in the surrounding environment.This opportunism hypothesis would indicate that ahigh number of plant species having pollen grains ofsmall to medium volume and size is present in thevegetation of the subtropical Dry Chaco forest. Asthese two results are contradictory, both hypothesesremain open.
Conclusion
In the subtropical Dry Chaco forest, the stinglessbees Tetragonisca fiebrigi, Scaptotrigona jujuyensis andGeotrigona argentina foraged on a large number ofplants, mainly those having pollen grains of smallvolumes and sizes, followed by medium ones. Mostof the plant species (23 out of 35 species) had small-size flowers (≤ 1 cm). Most of the flowers of smallsize had both small pollen volume ranging from 618µm3 in Parthenium hysterophorus to 6418 µm3 inAcacia and small pollen sizes. A significant positivelinear correlation was found between mean pollenvolume and maximum flower size as well as betweenmaximum pollen size and maximum flower size.
Two hypotheses are suitable to explain why thesebees mainly foraged on small pollen grains. The firsthypothesis (preference) is in agreement with thefindings of Wilms et al. (1997) and Ramalho(2004) for Meliponini bees in Neotropical habitats.However, the second hypothesis (opportunisticbehaviour) seems to support the absence of prefer-ence for particular pollen size, indicating that thesebees forage on floral resources according to theiravailability in the forest. The opportunism hypoth-esis would indicate that a high number of plantspecies having pollen grains of small to mediumvolume and size is present in the vegetation of thesubtropical Dry Chaco forest.
Acknowledgements
The author thanks Nora Brea, María CelesteScattolini and Anabela Plos for providing sugges-tions and comments on the manuscript. This studywas supported by CONICET (Consejo Nacional deInvestigaciones Científicas y Técnicas).
References
Arenas P. 2003. Etnografía y alimentación entre los toba-ñachilamoleek y wichí lhuku’tas del Chaco Central (Argentina).Buenos Aires, Argentina.
Biesmeijer JC, Van Marwiik B, Deursen K, Punt W, SommeijerMJ. 1992. Pollen sources for Apis mellifera L (Hym, Apidae) inSurinam, based on pollen grain volume estimates. Apidologie23: 245–256. doi:10.1051/apido:19920308.
Birks HJB. 1973. Past and present vegetation of the Isle of Skye – Apalaeoecological study. London: Cambridge University Press.
Birks HJB, Birks HH. 1980. Quaternary palaeoecology. London:Edward Arnold.
Cortopassi-Laurino M, Ramalho M. 1988. Pollen harvest byAfricanized Apis mellifera and Trigona spinipes in São Paulo –
Botanical and ecological views. Apidologie 19: 1–24.doi:10.1051/apido:19880101.
De Klerk P, Joosten H. 2007. The difference between pollen typesand plant taxa: A plea for clarity and scientific freedom.Eiszeitalter und Gegenwart/Quaternary Science Journal 56:162–171.
Fernández VA, Galetto L, Astegiano J. 2009. Influence of flowerfunctionality and pollination system on the pollen size-pistillength relationship. Organisms, Diversity & Evolution 9: 75–82. doi:10.1016/j.ode.2009.02.001.
Hesse M, Zetter R, Halbritter H, Weber M, Buchner R, Frosch-Radivo A, Ulrich S. 2009. Pollen terminology. An illustratedhandbook. Vienna: Springer.
Houston TF. 1989. Leioproctus bees associated with WesternAustralian smoke bushes (Conospermum spp.) and their adapta-tions for foraging and concealment (Hymenoptera: Colletidae:Paracolletini). Records of the Western Australian Museum 14:275–292.
Joosten H, De Klerk P. 2002. What’s in a name? Some thoughtson pollen classification, identification, and nomenclature inQuaternary palynology. Review of Palaeobotany andPalynology 122: 29–45. doi:10.1016/S0034-6667(02)00090-8.
Marques-Souza AC, Miranda IPA, Moura CO, Rabelo A, BarbosaEM. 2002. Características morfológicas e bioquímicas do pólencoletado por cinco espécies de meliponíneos da AmazôniaCentral. Acta amazônica 32: 217–229.
Michener CD, Winston ML, Jander R. 1978. Pollen manipulationand related activities and structures in bees of the familyApidae. University of Kansas Science Bulletin 51: 575–601.
O’Rourke MK, Buchmann SL. 1991. Standardized analyticaltechniques for bee-collected pollen. EnvironmentalEntomology 20: 507–513.
Prado DE. 1993. What is the Gran Chaco vegetation in SouthAmerica? I. A review. Contribution to the study of flora andvegetation of the Chaco. V. Candollea 48: 145–172.
Ramalho M. 1990. Foraging by stingless bees of the genusScaptotrigona (Apidae, Meliponinae). Journal of ApiculturalResearch 29: 61–67.
Ramalho M. 2004. Stingless bees and mass flowering trees in thecanopy of Atlantic Forest: A tight relationship. Acta BotanicaBrasilica 18: 37–47. doi:10.1590/S0102-33062004000100005.
80 F.G. Vossler
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015

Ramalho M, Imperatriz-Fonseca VL, Kleinekt-Giovannini A,Cortopassl-Laurino M. 1985. Exploitation of floral resourcesby Plebeia remotaHolmberg (Apidae, Meliponinae). Apidologie16: 307–330. doi:10.1051/apido:19850306.
Ramalho M, Kleinert-Giovannini A, Imperatriz-Fonseca VL.1990. Important bee plants for stingless bees (Melipona andTrigonini) and Africanized honeybees (Apis mellifera) in neotro-pical habitats: A review. Apidologie 21: 469–488. doi:10.1051/apido:19900508.
Ramalho M, Silva MD, Carvalho CAL. 2007. Dinâmica de uso defontes de pólen por Melipona scutellaris Latreille (Hymenoptera:Apidae): Uma análise comparativa com Apis mellifera L.(Hymenoptera: Apidae), no Domínio Tropical Atlântico.Neotropical Entomology 36: 38–45. doi:10.1590/S1519-566X2007000100005.
Roig-Alsina A, Vossler FG, Gennari GP. 2013. Stingless bees inArgentina. In: Vit P, Pedro SRM, Roubik DW, eds. Pot honey:A legacy of stingless bees, 125–134. New York: Springer.
Roubik DW. 1989. Ecology and natural history of tropical bees.New York: Cambridge University Press.
Roulston TH, Cane JH, Buchmann SL. 2000. What governs pro-tein content of pollen: Pollinator preferences, pollen-pistil inter-actions, or phylogeny? Ecological Monographs 70: 617–643.
Schnack B, Covas G. 1945. Hibridación interespecífica enGlandularia (Verbenáceas). Darwiniana 7: 71–79.
Silveira da FA. 1991. Influence of pollen grain volume on theestimation of the relative importance of its source to bees.Apidologie 22: 495–502. doi:10.1051/apido:19910502.
StatPoint Technologies. 2010. Statgraphics® Centurion XVI.StatPoint Technologies, Inc., Warrenton, VA. http://www.stat-graphics.com.
Subba Reddi C, Reddi NS. 1986. Pollen production in someanemophilous angiosperms. Grana 25: 55–61. doi:10.1080/00173138609429933.
Thorp RW. 2000. The collection of pollen by bees. PlantSystematics and Evolution 222: 211–223. doi:10.1007/BF00984103.
Tormo Molina R, Muñoz Rodríguez A, Silva Palaciso I, GallardoLópez F. 1996. Pollen production in anemophilous trees.Grana 35: 38–46. doi:10.1080/00173139609430499.
Torres C. 2000. Pollen size evolution: Correlation between pollenvolume and pistil length in Asteraceae. Sexual PlantReproduction 12: 365–370. doi:10.1007/s004970000030.
Vossler FG. 2012. Flower visits, nesting and nest defence behaviourof stingless bees (Apidae: Meliponini): Suitability of the beespecies for meliponiculture in the Argentinean Chaco region.Apidologie 43: 139–161. doi:10.1007/s13592-011-0097-6.
Vossler FG. 2013. Estudio palinológico de las reservas alimentar-ias (miel y masas de polen) de “abejas nativas sin aguijón”(Hymenoptera, Apidae, Meliponini): Un aporte al conoci-miento de la interacción abeja-planta en el Chaco Seco deArgentina. La Plata: Universidad Nacional de La Plata,Argentina, PhD Diss.
Vossler FG, Tellería MC, Cunningham M. 2010. Floral resourcesforaged by Geotrigona argentina (Apidae, Meliponini) in theArgentine Dry Chaco forest. Grana 49: 142–153.doi:10.1080/00173131003694274.
Wilms W, Ramalho M, Wendel L. 1997. Stingless bees andAfricanized honey bees in the Mata Atlântica rainforest ofBrazil. 30th International Apicultural Congress of Apimondia,Antwerp, Belgium, 1–6 September 1997, 167–170.
Wodehouse RP. 1935. Pollen grains. New York and London:McGraw-Hill.
Zuloaga FO, Morrone O, Belgrano MJ. 2008. Catálogo de lasplantas vasculares del Cono Sur (Argentina, Sur de Brasil,Chile, Paraguay y Uruguay). Volumes 1–3. Monographs inSystematic Botany from the Missouri Botanical Garden 107:1–983, 985–2286, 2287–3348.
Pollen grains in Chaquenian Meliponini diet 81
Dow
nloa
ded
by [
Dr
Favi
o V
ossl
er]
at 1
3:05
25
Mar
ch 2
015















![[Chaco] Cloud Computing – Cecilia Conci](https://img.pdfslide.us/doc/110x75/546c2b3cb4af9f702c8b4f50/chaco-cloud-computing-cecilia-conci.jpg)












