Embed Size (px)
Citation preview

a PATTERNS & PHENOTYPES
Simultaneous Activation of SHR and ATHB8Expression Defines Switch to PreprocambialCell State in Arabidopsis Leaf DevelopmentJason Gardiner,† Tyler J. Donner,† and Enrico Scarpella*
The processes underlying the formation of leaf vascular networks have long captured the attention of de-velopmental biologists, especially because files of elongated vascular-precursor procambial cells seem todifferentiate from apparently equivalent, isodiametric ground cells. In Arabidopsis leaves, ground cellsthat have been specified to vascular fate engage expression of ARABIDOPSIS THALIANA HOMEOBOX8(ATHB8). While definition of the transcriptional state of ATHB8-expressing ground cells would be particu-larly informative, no other genes have been identified whose expression is initiated at this stage. Here weshow that expression of SHORT-ROOT (SHR) is activated simultaneously with that of ATHB8 in leaf devel-opment. Congruence between SHR and ATHB8 expression domains persists under conditions of manipu-lated vein patterning, suggesting that inception of expression of SHR and ATHB8 identifies transition toa preprocambial cell state that presages vein formation. Our observations further characterize the molec-ular identity of cells at anatomically inconspicuous stages of leaf vein development. DevelopmentalDynamics 240:261–270, 2011. VC 2010 Wiley-Liss, Inc.
Key words: Arabidopsis thaliana; leaf development; vascular patterning; vein formation; procambium; auxintransport; ATHB8; SHR; SCL29; SCL32
Accepted 1 November 2010
INTRODUCTION
The vascular system of plants is a net-work of veins that extends throughoutall organs (Esau, 1965). Veins trans-port water and nutrients, and aresource of signals that act locally, toassign identity to surrounding cells,and systemically, to coordinate initia-tion of new shoot organs with that ofnew roots (Berleth and Sachs, 2001).Sites of vein formation are foreshad-owed by appearance of files of elon-gated procambial cells, which in leafdevelopment seem to emerge denovo from within a homogeneous
population of isodiametric groundcells (Louis, 1935; Esau, 1943; Foster,1952).
The molecular events that lead to
acquisition of procambial cell identity
during leaf development are not
entirely clear, but available evidence
supports a decisive role for transport
and transduction of the plant signal-
ing molecule auxin in specifying paths
of leaf vein formation. Auxin applica-
tion to leaf primordia induces forma-
tion of new veins (Sachs, 1975, 1989;
Scarpella et al., 2006), and chemical
inhibition of auxin transport during
leaf development severely disturbs
vein patterning (Mattsson et al., 1999;
Sieburth, 1999). Consistent with these
observations, mutants impaired in
auxin biosynthesis, response, or trans-
port display diagnostic alterations in
leaf vein patterns (Przemeck et al.,
1996; Mattsson et al., 1999; Alonso-
Peral et al., 2006; Cheng et al., 2006).
During leaf development, ground cells
are directed toward procambial fate
through induction of wide domains
of expression of the PIN-FORMED1
(PIN1) auxin exporter and of the
auxin response transcription factor
Additional Supporting Information may be found in the online version of this article.
Department of Biological Sciences, University of Alberta, Edmonton AB, CanadaGrant sponsor: the Natural Sciences and Engineering Research Council of Canada (NSERC); Grant number: NSERC RGPIN 311819-05.†J. Gardiner and T. J. Donner contributed equally in this work.*Correspondence to: Enrico Scarpella, Department of Biological Sciences, University of Alberta, CW-405 BiologicalSciences Building, Edmonton AB, Canada T6G 2E9. E-mail: [email protected]
DOI 10.1002/dvdy.22516Published online 3 December 2010 in Wiley Online Library (wileyonlinelibrary.com).
DEVELOPMENTAL DYNAMICS 240:261–270, 2011
VC 2010 Wiley-Liss, Inc.
Dev
elop
men
tal D
ynam
ics

MONOPTEROS (MP; Hardtke and
Berleth, 1998; Scarpella et al., 2006;
Sawchuk et al., 2007; Wenzel et al.,
2007; Donner et al., 2009). Cessation
of PIN1 and MP expression occurs in
some of the cells, as fields of PIN1 and
MP expression become restricted to
individual lines of elongating procam-
bial cells (Scarpella et al., 2006; Wen-
zel et al., 2007; Donner et al., 2009).While initiation of PIN1 and MP
expression identifies a reversible statein leaf vein formation, files of PIN1-and MP-expressing ground cells thatare stabilized toward procambial fateactivate expression of the class IIIHOMEODOMAIN-LEUCINE ZIPPER(HD-ZIP III) gene ARABIDOPSISTHALIANA HOMEOBOX8 (ATHB8;Baima et al., 1995; Kang and Dengler,2004; Scarpella et al., 2004; Sawchuket al., 2007; Donner et al., 2009). Onsetof ATHB8 expression is directly con-trolled by MP (Donner et al., 2009),and identifies transition to a typicallyirreversible ‘‘preprocambial’’ cell statethat accurately predicts sites of leafvein formation (e.g., Koizumi et al.,2000; Carland and Nelson, 2004; Kangand Dengler, 2004; Scarpella et al.,2004, 2006; Pineau et al., 2005;Alonso-Peral et al., 2006; Cnops et al.,2006; Petricka and Nelson, 2007; Saw-chuk et al., 2007, 2008; Donner et al.,2009; Carland et al., 2010). Therefore,characterization of the transcriptionalprofile of ground cells that haveswitched to preprocambial state wouldbe particularly desirable as it may pro-vide insight into the molecular path-ways controlling vein formation. How-ever, as of yet, no genes have beenidentified whose expression in vein de-velopment is initiated simultaneouslywith that ofATHB8.
In this study, we searched for geneexpression patterns associated withearly stages of vein development inArabidopsis leaves. We found thatonset of expression of SHORT-ROOT(SHR), which encodes a transcriptionfactor of the GRAS family (afterGIBBERELLIC ACID INSENSITIVE,REPRESSOR OF gibberellic acid1–3and SCARECROW; Di Laurenzioet al., 1996; Peng et al., 1997; Silver-stone et al., 1998; Pysh et al., 1999;Helariutta et al., 2000), coincides withthat of ATHB8 during undisturbedleaf development. Parallel initiation of
expression of SHR and ATHB8 per-sisted under conditions of experimen-tally manipulated leaf vascular pat-terning, suggesting that synchronousactivation of expression of SHR andATHB8 operationally defines a repro-ducible cell state that presages veinappearance. While the ATHB8 proteinremained confined to leaf vascularcells, however, the SHR protein addi-tionally localized to adjacent, perivei-nal positions, suggesting functions ofpreprocambial cells beyond vein for-mation. Our observations assist inthe molecular characterization ofcell state at morphologically indis-tinguishable, preprocambial stages ofleaf vein formation.
RESULTS
In Arabidopsis leaves, veins arearranged in a ramified pattern thatlargely reflects the shape of the leaf(Nelson and Dengler, 1997; Candelaet al., 1999; Dengler and Kang, 2001;Fig. 1A). Lateral veins depart from ei-ther side of a conspicuous central vein(midvein), extend along the leaf mar-gin and connect to distal veins to formprominent closed loops. A series ofhigher-order veins branch from mid-vein and loops, and can either termi-nate in the lamina or join two veins.Veins of succeeding orders become rec-ognizable progressively later in thesame area of the developing leaf pri-mordium, and veins of the same orderappear in a tip-to-base sequence dur-ing leaf development (Telfer and Poet-hig, 1994; Kinsman and Pyke, 1998;Candela et al., 1999; Mattsson et al.,1999; Sieburth, 1999; Kang and Den-gler, 2002, 2004; Steynen and Schultz,2003; Scarpella et al., 2004; Fig. 1B–D). The illustrations in Figure 1 (Fig.1A–D) schematically depict the tempo-ral sequence of vein formation eventsin Arabidopsis leaf development, anddefine stages and terminology towhich we refer throughout this study(for additional details on nomencla-ture, see the Experimental Proceduressection).
Leaf Expression of Root
Vascular Markers
All the genes whose expression haspreviously been assigned to earlystages of leaf vein development have
also been reported to be expressed inthe root procambium (e.g., Baimaet al., 1995; Hardtke and Berleth,1998; Steinmann et al., 1999; Kangand Dengler, 2004; Scarpella et al.,2004, 2006; Alonso-Peral et al., 2006;Konishi and Yanagisawa, 2007; Wen-zel et al., 2007; Carland and Nelson,2009; Gardiner et al., 2010), and iden-tification of leaf vascular gene expres-sion profiles based on root procambialexpression has proved to be an effec-tive strategy (Gardiner et al., 2010).Reporter gene expression in the J2501and Q0990::mGFP5er enhancer-traplines and in transcriptional fusionsto SHR or to WOODENLEG/CYTO-KININ RESPONSE1/ARABIDOPSISHISTIDINE KINASE4 (Mahonenet al., 2000; Inoue et al., 2001; Suzukiet al., 2001; WOL hereafter) has con-sistently been used as reliable markerof root procambial cells (e.g., Benkovaet al., 2003; Birnbaum et al., 2003;Wang et al., 2005; Zhang et al., 2005;Dello Ioio et al., 2007; Hirota et al.,2007; Mustroph et al., 2009; Peterssonet al., 2009; Fig. 2A–D). Activation ofQ0990::mGFP5er expression in theleaf coincides with acquisition of pro-cambial cell identity (Sawchuk et al.,2007; Fig. 2E), further supporting thevalue of root procambial expression fil-tering for discovery of leaf vascularexpression patterns. Therefore, toidentify new preprocambial expres-sion profiles, we asked whether re-porter gene expression in J2501::mGFP5er and in transcriptionalfusions to SHR or WOL retained, likeQ0990::mGFP5er, vascular specificityin the leaf. To address this question,we visualized fluorescence protein ac-tivity in J2501::mGFP5er and in tran-scriptional fusions of SHR or WOL tonuclear localized YFP or GFP(HTA6:EYFP or HTA6:EGFP; Zhanget al., 2005), and compared it with thatof Q0990::mGFP5er, in first leaves ofseedlings 4 days after germination(DAG) as their venation is predomi-nantly preprocambial and procambial(Sawchuk et al., 2007; Donner et al.,2009; Fig. 1D).While, in agreement with previous
observations (Sawchuk et al., 2007;Donner et al., 2009), Q0990::mGFP5er signals in 4-DAG leaveswere restricted to procambial midveinand first loops (Fig. 2E), expression ofJ2501::mGFP5er was not detected
262 GARDINER ET AL.
Dev
elop
men
tal D
ynam
ics
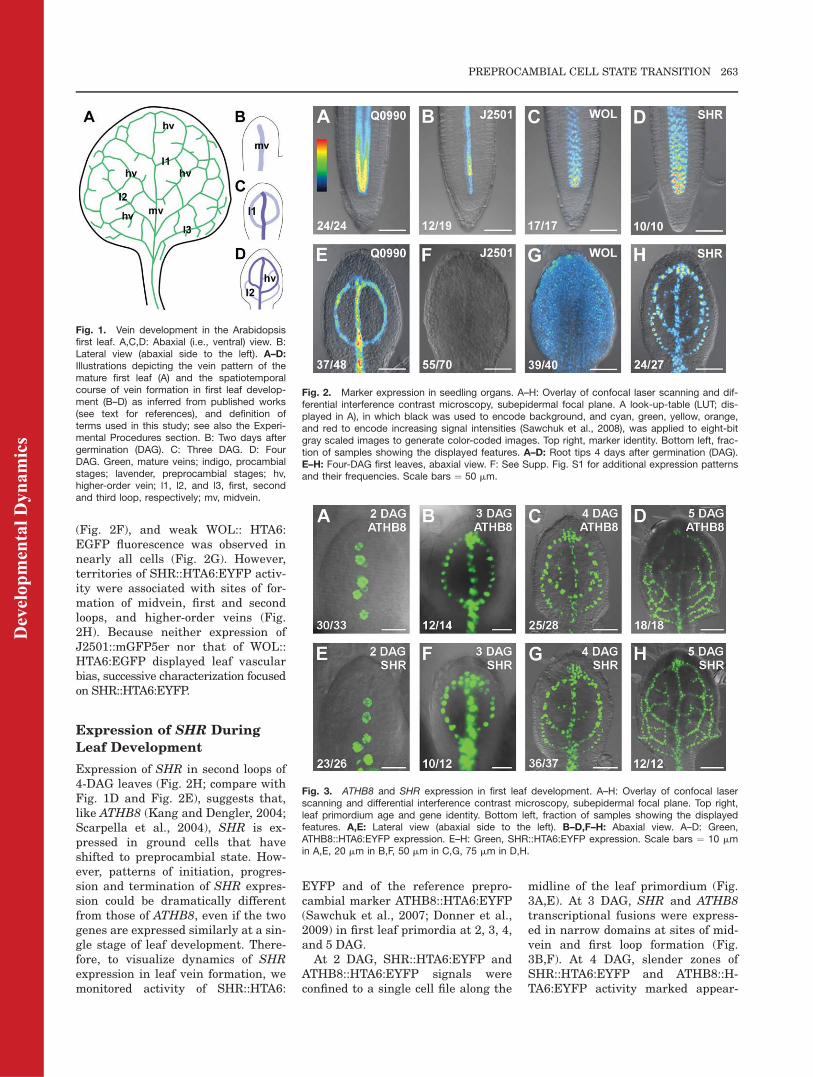
(Fig. 2F), and weak WOL:: HTA6:EGFP fluorescence was observed innearly all cells (Fig. 2G). However,territories of SHR::HTA6:EYFP activ-ity were associated with sites of for-mation of midvein, first and secondloops, and higher-order veins (Fig.2H). Because neither expression ofJ2501::mGFP5er nor that of WOL::HTA6:EGFP displayed leaf vascularbias, successive characterization focusedon SHR::HTA6:EYFP.
Expression of SHR During
Leaf Development
Expression of SHR in second loops of4-DAG leaves (Fig. 2H; compare withFig. 1D and Fig. 2E), suggests that,like ATHB8 (Kang and Dengler, 2004;Scarpella et al., 2004), SHR is ex-pressed in ground cells that haveshifted to preprocambial state. How-ever, patterns of initiation, progres-sion and termination of SHR expres-sion could be dramatically differentfrom those of ATHB8, even if the twogenes are expressed similarly at a sin-gle stage of leaf development. There-fore, to visualize dynamics of SHRexpression in leaf vein formation, wemonitored activity of SHR::HTA6:
EYFP and of the reference prepro-cambial marker ATHB8::HTA6:EYFP(Sawchuk et al., 2007; Donner et al.,2009) in first leaf primordia at 2, 3, 4,and 5 DAG.
At 2 DAG, SHR::HTA6:EYFP andATHB8::HTA6:EYFP signals wereconfined to a single cell file along the
midline of the leaf primordium (Fig.3A,E). At 3 DAG, SHR and ATHB8transcriptional fusions were express-ed in narrow domains at sites of mid-vein and first loop formation (Fig.3B,F). At 4 DAG, slender zones ofSHR::HTA6:EYFP and ATHB8::H-TA6:EYFP activity marked appear-
Fig. 1. Vein development in the Arabidopsisfirst leaf. A,C,D: Abaxial (i.e., ventral) view. B:Lateral view (abaxial side to the left). A–D:Illustrations depicting the vein pattern of themature first leaf (A) and the spatiotemporalcourse of vein formation in first leaf develop-ment (B–D) as inferred from published works(see text for references), and definition ofterms used in this study; see also the Experi-mental Procedures section. B: Two days aftergermination (DAG). C: Three DAG. D: FourDAG. Green, mature veins; indigo, procambialstages; lavender, preprocambial stages; hv,higher-order vein; l1, l2, and l3, first, secondand third loop, respectively; mv, midvein.
Fig. 2. Marker expression in seedling organs. A–H: Overlay of confocal laser scanning and dif-ferential interference contrast microscopy, subepidermal focal plane. A look-up-table (LUT; dis-played in A), in which black was used to encode background, and cyan, green, yellow, orange,and red to encode increasing signal intensities (Sawchuk et al., 2008), was applied to eight-bitgray scaled images to generate color-coded images. Top right, marker identity. Bottom left, frac-tion of samples showing the displayed features. A–D: Root tips 4 days after germination (DAG).E–H: Four-DAG first leaves, abaxial view. F: See Supp. Fig. S1 for additional expression patternsand their frequencies. Scale bars ¼ 50 mm.
Fig. 3. ATHB8 and SHR expression in first leaf development. A–H: Overlay of confocal laserscanning and differential interference contrast microscopy, subepidermal focal plane. Top right,leaf primordium age and gene identity. Bottom left, fraction of samples showing the displayedfeatures. A,E: Lateral view (abaxial side to the left). B–D,F–H: Abaxial view. A–D: Green,ATHB8::HTA6:EYFP expression. E–H: Green, SHR::HTA6:EYFP expression. Scale bars ¼ 10 mmin A,E, 20 mm in B,F, 50 mm in C,G, 75 mm in D,H.
PREPROCAMBIAL CELL STATE TRANSITION 263
Dev
elop
men
tal D
ynam
ics

ance of midvein, first and secondloops, and higher-order veins (Fig.3C,G). Finally, at 5 DAG, SHR andATHB8 promoters directed expressionin developing midvein, first, second,and third loops, and higher-orderveins (Fig. 3D,H). However, whileATHB8::HTA6:EYFP expression hadsubsided from the apical portion ofmidvein and first loops (Fig. 3D), theSHR transcriptional fusion wasevenly active throughout the leaf vas-culature (Fig. 3H).
In summary, expression of SHRseemed to be tightly associated withregions of ATHB8-labeled vein forma-tion throughout leaf development.
Stage-Specific SHR
Expression in Leaf Vein
Formation
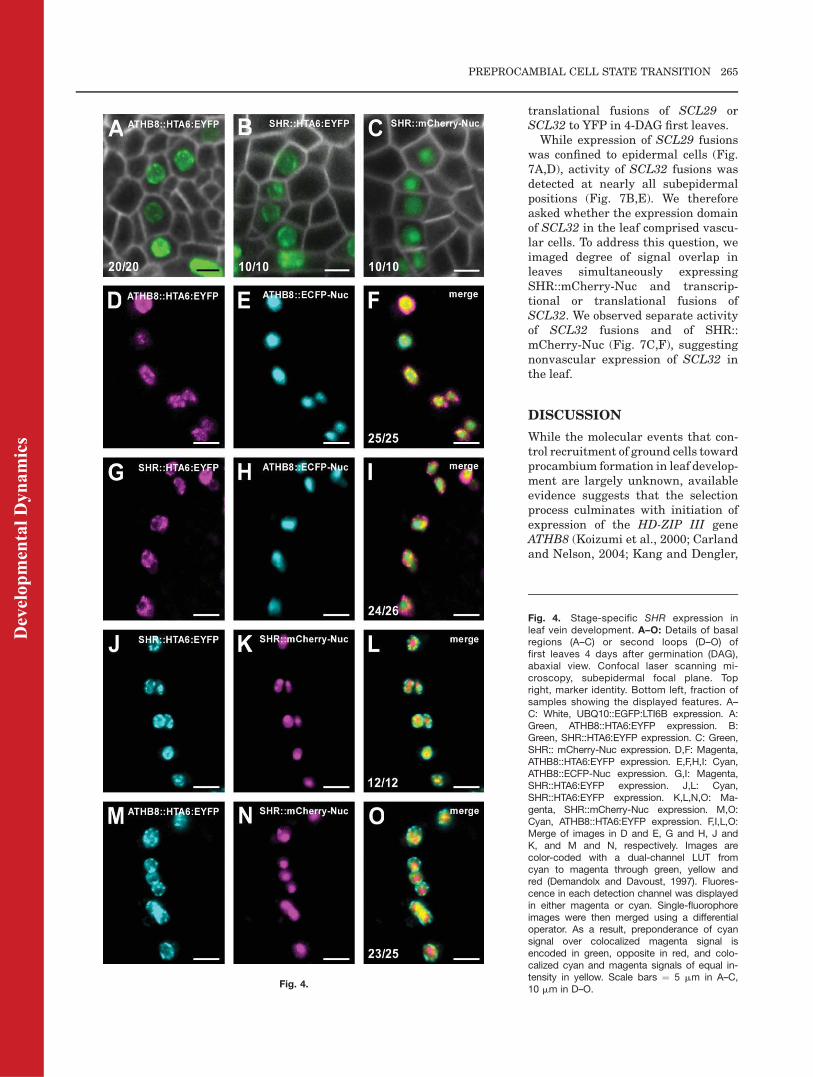
Comparison between SHR andATHB8 expression profiles during leafdevelopment (Fig. 3) suggests thatexpression of SHR is initiated as earlyas that of ATHB8, and that thereforeSHR expression could be assigned toground cells that have switched to pre-procambial state. We adopted two cri-teria to test such a hypothesis: (1) visu-alization of shape of cells expressingSHR; (2) detection of SHR and ATHB8expression within the same sample.
Simultaneous imaging of activity ofSHR transcriptional fusions andplasma-membrane-localized GFP(Sawchuk et al., 2008) in basalregions of 4-DAG first leaves showedthat, like ATHB8 (Kang and Dengler,2004; Scarpella et al., 2004; Fig. 4A),SHR is expressed in isodiametric cells(Fig. 4B,C), suggesting that SHRexpression is initiated in ground cells.
Covisualization of signals of SHR::HTA6:EYFP and ATHB::ECFP-Nuc(Sawchuk et al., 2007) in second loopsof 4-DAGfirst leaves showedmatchingexpression of fluorescent reporters(Fig. 4G–I), suggesting that expres-sion of SHR is initiated simultane-ously with thatATHB8. To test for pos-sible artifacts induced by fluorophoreintrinsic properties (e.g., differentmaturation times and stabilities ofHTA6:EYFP and ECFP-Nuc) or detec-tion parameters (e.g., suboptimal exci-tation wavelengths and emissionintervals), we visualized extent ofcoexpression between SHR::mCherry-
Nuc and ATHB8::HTA6:EYFP signals.The reproducible coincidence of fluo-rescence in reciprocal permutations ofSHR and ATHB8 regulatory regionswith YFP and CFP, or mCherry (com-pare Fig. 4M–O to Fig. 4G–I), suggeststhat our covisualization data are fluo-rophore independent, further support-ing that expression of SHR andATHB8 is simultaneously activated inground cells that have transitioned topreprocambial state.
SHR Expression in Auxin
Transport-Inhibited Leaves
Domains of SHR expression may berigidly specified in leaf developmentand only incidentally matching withzones of vein appearance. Therefore,we asked whether fields of SHRexpression remained associated withareas of leaf vein formation upon ex-perimental interference with vascularpatterning. Auxin transport has beenshown to define sites of vein appear-ance in developing leaf primordia(Mattsson et al., 1999; Sieburth,1999). Therefore, we grew seedlingsharboring the SHR and ATHB8 tran-scriptional fusions in the presence ofthe auxin transport inhibitor 1-N-naphthylphthalamic acid (NPA) andimaged fluorescent protein expressionin first leaves at 3, 4, and 5DAG.
Leaves of plants germinated andgrown in the presence of auxin trans-port inhibitors are characterized byseveral reproducible, distinct abnor-malities in vein network configura-tion; most conspicuously, great num-bers of broad vein loops that fuse alongthe entire edge of the leaf, to give riseto awidemarginal zone of vascular dif-ferentiation, and that extend parallelto one another at the centre of the leaf,to give rise to a laterally expandedmidvein (Mattsson et al., 1999; Sie-burth, 1999). As shown in Figure 5,domains of SHR::mCherry-Nuc andATHB8::HTA6:EYFP expression re-tained their tight relation to sites ofvein formation throughout develop-ment of auxin transport-inhibitedleaves (Fig. 5A–F). Furthermore,strict congruence between regions ofSHR and ATHB8 promoter activitywas preserved under conditions ofreduced auxin transport (Fig. 5G–I).However, as observed in undisturbedleaf development, SHR::mCherry-Nuc
signals persisted at later stages of veindifferentiation, while expression of theATHB8 transcriptional fusion haddeclined (Fig. 5H,I).In conclusion, association between
SHR expression domains with areasof ATHB8-marked vein formationobserved under undisturbed condi-tions persisted in auxin transport-inhibited leaves, suggesting noncir-cumstantial correlation between SHRexpression and leaf vein emergence.
SHR Expression in Leaf Vein
Development
In the root, SHR transcription is re-stricted to the procambium, but SHRprotein is additionally localized to thecell layer surrounding the root vascu-lature (Helariutta et al., 2000; Naka-jima et al., 2001). We therefore askedwhether SHR displayed similarbehavior in the leaf. To address thisquestion, we visualized expression ofa translational fusion of SHR to YFPin 4-DAG first leaves, and comparedit to expression of the nonmobileATHB8::ATHB8:mCherry transla-tional fusion (Donner et al., 2009).In agreement with previous obser-
vations (Donner et al., 2009), expres-sion of the fluorescently taggedATHB8 protein mimicked ATHB8 pro-moter activity in leaf vascular cells(Fig. 6A–C). In contrast, SHR::SHR:EYFP signals were further detected incells adjacent the preprocambial andprocambial domains of expression ofthe SHR transcriptional fusion (Fig.6D–I). However, while SHR::SHR:EYFP fluorescence was distributed inboth nucleus and cytoplasm of cellswithin the vascular expression terri-tory, fusion protein localization in theperiveinal cell layer was markedly nu-clear (Fig. 6D–I).
Leaf Expression of
SHR-Related Genes
SHR belongs to a small clade ofGRAS genes that includes SCARE-CROW-LIKE29 (SCL29) and SCL32(Bolle, 2004; Lee et al., 2008). There-fore, we asked whether SCL29 andSCL32 were expressed in the leaf in apattern similar to that of SHR. Toaddress this question, we visualizedexpression of transcriptional and
264 GARDINER ET AL.
Dev
elop
men
tal D
ynam
ics
translational fusions of SCL29 orSCL32 to YFP in 4-DAG first leaves.While expression of SCL29 fusions
was confined to epidermal cells (Fig.7A,D), activity of SCL32 fusions wasdetected at nearly all subepidermalpositions (Fig. 7B,E). We thereforeasked whether the expression domainof SCL32 in the leaf comprised vascu-lar cells. To address this question, weimaged degree of signal overlap inleaves simultaneously expressingSHR::mCherry-Nuc and transcrip-tional or translational fusions ofSCL32. We observed separate activityof SCL32 fusions and of SHR::mCherry-Nuc (Fig. 7C,F), suggestingnonvascular expression of SCL32 inthe leaf.
DISCUSSION
While the molecular events that con-trol recruitment of ground cells towardprocambium formation in leaf develop-ment are largely unknown, availableevidence suggests that the selectionprocess culminates with initiation ofexpression of the HD-ZIP III geneATHB8 (Koizumi et al., 2000; Carlandand Nelson, 2004; Kang and Dengler,
Fig. 4.
Fig. 4. Stage-specific SHR expression inleaf vein development. A–O: Details of basalregions (A–C) or second loops (D–O) offirst leaves 4 days after germination (DAG),abaxial view. Confocal laser scanning mi-croscopy, subepidermal focal plane. Topright, marker identity. Bottom left, fraction ofsamples showing the displayed features. A–C: White, UBQ10::EGFP:LTI6B expression. A:Green, ATHB8::HTA6:EYFP expression. B:Green, SHR::HTA6:EYFP expression. C: Green,SHR:: mCherry-Nuc expression. D,F: Magenta,ATHB8::HTA6:EYFP expression. E,F,H,I: Cyan,ATHB8::ECFP-Nuc expression. G,I: Magenta,SHR::HTA6:EYFP expression. J,L: Cyan,SHR::HTA6:EYFP expression. K,L,N,O: Ma-genta, SHR::mCherry-Nuc expression. M,O:Cyan, ATHB8::HTA6:EYFP expression. F,I,L,O:Merge of images in D and E, G and H, J andK, and M and N, respectively. Images arecolor-coded with a dual-channel LUT fromcyan to magenta through green, yellow andred (Demandolx and Davoust, 1997). Fluores-cence in each detection channel was displayedin either magenta or cyan. Single-fluorophoreimages were then merged using a differentialoperator. As a result, preponderance of cyansignal over colocalized magenta signal isencoded in green, opposite in red, and colo-calized cyan and magenta signals of equal in-tensity in yellow. Scale bars ¼ 5 mm in A–C,10 mm in D–O.
PREPROCAMBIAL CELL STATE TRANSITION 265
Dev
elop
men
tal D
ynam
ics
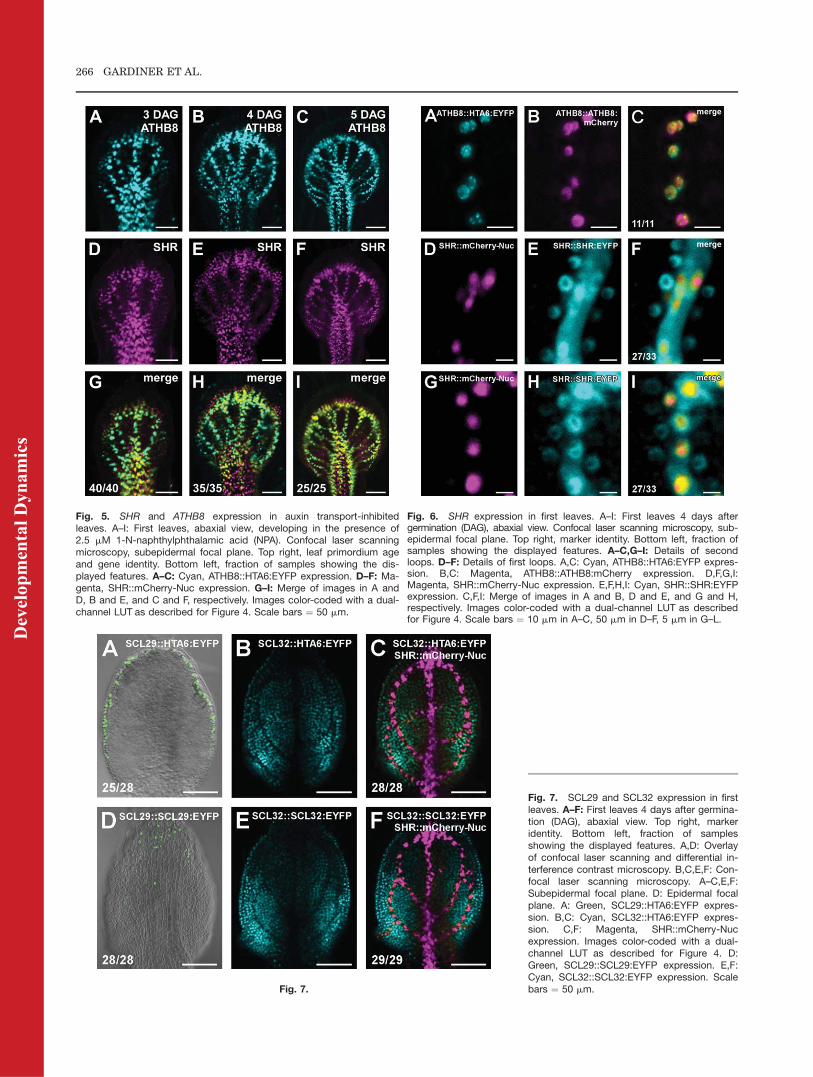
Fig. 5. SHR and ATHB8 expression in auxin transport-inhibitedleaves. A–I: First leaves, abaxial view, developing in the presence of2.5 mM 1-N-naphthylphthalamic acid (NPA). Confocal laser scanningmicroscopy, subepidermal focal plane. Top right, leaf primordium ageand gene identity. Bottom left, fraction of samples showing the dis-played features. A–C: Cyan, ATHB8::HTA6:EYFP expression. D–F: Ma-genta, SHR::mCherry-Nuc expression. G–I: Merge of images in A andD, B and E, and C and F, respectively. Images color-coded with a dual-channel LUT as described for Figure 4. Scale bars ¼ 50 mm.
Fig. 6. SHR expression in first leaves. A–I: First leaves 4 days aftergermination (DAG), abaxial view. Confocal laser scanning microscopy, sub-epidermal focal plane. Top right, marker identity. Bottom left, fraction ofsamples showing the displayed features. A–C,G–I: Details of secondloops. D–F: Details of first loops. A,C: Cyan, ATHB8::HTA6:EYFP expres-sion. B,C: Magenta, ATHB8::ATHB8:mCherry expression. D,F,G,I:Magenta, SHR::mCherry-Nuc expression. E,F,H,I: Cyan, SHR::SHR:EYFPexpression. C,F,I: Merge of images in A and B, D and E, and G and H,respectively. Images color-coded with a dual-channel LUT as describedfor Figure 4. Scale bars ¼ 10 mm in A–C, 50 mm in D–F, 5 mm in G–L.
Fig. 7.
Fig. 7. SCL29 and SCL32 expression in firstleaves. A–F: First leaves 4 days after germina-tion (DAG), abaxial view. Top right, markeridentity. Bottom left, fraction of samplesshowing the displayed features. A,D: Overlayof confocal laser scanning and differential in-terference contrast microscopy. B,C,E,F: Con-focal laser scanning microscopy. A–C,E,F:Subepidermal focal plane. D: Epidermal focalplane. A: Green, SCL29::HTA6:EYFP expres-sion. B,C: Cyan, SCL32::HTA6:EYFP expres-sion. C,F: Magenta, SHR::mCherry-Nucexpression. Images color-coded with a dual-channel LUT as described for Figure 4. D:Green, SCL29::SCL29:EYFP expression. E,F:Cyan, SCL32::SCL32:EYFP expression. Scalebars ¼ 50 mm.
266 GARDINER ET AL.
Dev
elop
men
tal D
ynam
ics

2004; Scarpella et al., 2004, 2006;Pineau et al., 2005; Alonso-Peral et al.,2006; Cnops et al., 2006; Petricka andNelson, 2007; Sawchuk et al., 2007,2008; Donner et al., 2009; Carlandet al., 2010). Activation of ATHB8expression defines transition to a mor-phologically inconspicuous prepro-cambial cell state that preludes to pro-cambium appearance. Therefore,characterization of themolecular iden-tity of ground cells that have switchedto preprocambial state would be par-ticularly informative as it may provideinsight into the molecular circuits con-trolling vein formation.
In this study, we searched for geneexpression profiles associated withpreprocambial stages of vein develop-ment in Arabidopsis leaves. We foundthat expression of SHR, which enco-des a member of the GRAS family ofplant-specific transcription factors (DiLaurenzio et al., 1996; Pysh et al.,1999; Helariutta et al., 2000),emerges in synchrony with that ofATHB8 in leaf development, suggest-ing that parallel activation of expres-sion of SHR and ATHB8 identifies apreprocambial cell state that announ-ces vein formation. However, whileATHB8 protein expression remainedconfined to developing veins, the SHRprotein expression domain furtherincluded a contiguous, perivascularcell layer, suggestive of activities ofprocambium-precursor cells beyondvein formation.
Transition to Preprocambial
Cell State
During leaf development, SHR andATHB8 were expressed in seeminglyoverlapping subepidermal domainsand with amazingly comparable dy-namics. Expression of both SHR andATHB8 was initiated in narrowdomains that became associated withsites of vein formation. Further, vein-associated expression fields of SHRand ATHB8 emerged in the same tem-poral sequence: midvein, first loops,second loops and higher-order veins,third loops. Finally, vein order-specificexpression domains of SHR andATHB8 became apparent at the samestage of leaf development. However,expression of SHRwas sustained at allstages of vein formation, while that ofATHB8 became dissipated at later
stages of vascular differentiation, inagreement with previous reports(Kang and Dengler, 2002, 2004; Scar-pella et al., 2004). Expression of bothSHR and ATHB8 was initiated in filesof polygonal, isodiametric groundcells, and positions of activation ofSHR expression overlapped with sitesof initiation of ATHB8 expression, sug-gesting that SHR is expressed at pre-procambial stages of vein develop-ment. Moreover, that SHR andATHB8 preprocambial expressiondomains reproducibly coincided withone another suggests that expressionof SHR is initiated concurrently withtransition to ATHB8 preprocambialcell state.
If coincidence between expression ofSHR and ATHB8 were merely circum-stantial, one would not expect suchassociation to endure under conditionsof manipulated ATHB8 expression.Behavior of SHR expression in leavesdeveloping under conditions of re-duced auxin transport, which dramati-cally changes the architecture ofATHB8 expression domains and ofvein networks (Mattsson et al., 1999;Sieburth, 1999; Gardiner et al., 2010),was comparable to that observedunder undisturbed vein patterning.All aspects of SHR expression, includ-ing relation to ATHB8 expression andassociation with positions of vein for-mation, proved to be highly reproduci-ble under all experimental conditions.We therefore suggest that, togetherwith ATHB8, activation of expressionof SHR defines switch to a morphologi-cally inconspicuous transcriptionalstate that foreshadows procambialdevelopment.
Unlike ATHB8, the SHR protein isadditionally localized to a layer of non-vascular cells that surrounds leafveins. This observation is consistentwith events occurring in root develop-ment, where SHRmovement from vas-cular to neighboring cells is requiredfor the formation of the cell sheaththat envelops the single vein (Helar-iutta et al., 2000; Nakajima et al.,2001; Gallagher et al., 2004). Leafveins have long been suspected to pro-vide positional cues that control differ-entiation of adjoining photosyntheticcell types (Langdale and Nelson,1991), and the pattern of SHR expres-sion in the leaf suggests that suchorganizing influence may arise simul-
taneously with transition to prepro-cambial cell state.Correct initiation of ATHB8 expres-
sion at preprocambial stages of leafvein development strictly depends onthe presence of a TGTCTG regulatoryelement in the ATHB8 promoter (Don-ner et al., 2009). The SHR promoterdoes not contain any TGTCTG ele-ment, suggesting an independentmechanism controlling onset of SHRexpression. It will be interesting to un-derstand the molecular basis of SHRpreprocambial expression; neverthe-less, our findings already contribute tomolecularly define cells at incipientstages of leaf vascular development.
Complementary Leaf
Expression Profiles of
SHR-Related Genes
Members of gene families frequentlydisplay overlapping expression pro-files (e.g., Mason et al., 2004; Tsuchi-saka and Theologis, 2004; Sawchuket al., 2008). In contrast, the expres-sion of the related SHR, SCL29, andSCL32 genes defines complementaryterritories of cells in the leaf.Epidermal domains of SCL29 pro-
moter activity become further com-partmentalized by presence of the in-tronless SCL29 coding sequence.Reports of regulatory elements withinthe coding region are not unprece-dented (e.g., Ito et al., 2003), and vari-ous post-transcriptional control mech-anisms have been described that couldaccount for the differential behavior ofSCL29 transcriptional and transla-tional fusions, including regulated nu-clear export (Bailey-Serres et al.,2009), mRNA decay (Belostotsky andSieburth, 2009), and intercellularmRNA trafficking (Ueki and Citovsky,2000).Subepidermal cells that express ei-
ther type of SCL32 fusion lack expres-sion of the preprocambial markergene SHR, and mutual exclusivity ofSCL32 and SHR expression domainsis consistent with the view that photo-synthetic and vascular cell identityacquisition represent antagonisticpathways in leaf subepidermal tissueontogeny (Scarpella et al., 2004; Kanget al., 2007; Sawchuk et al., 2008).Tissue-specific expression data are
available for 21 of the 32 GRAS genes
PREPROCAMBIAL CELL STATE TRANSITION 267
Dev
elop
men
tal D
ynam
ics

in Arabidopsis, but function is onlyknown for 10 of them (Di Laurenzioet al., 1996; Peng et al., 1997; Silver-stone et al., 1998; Pysh et al., 1999;Bolle et al., 2000; Dill et al., 2001; Dilland Sun, 2001; Lee et al., 2002; Wenand Chang, 2002; Greb et al., 2003;Fu et al., 2004; Tyler et al., 2004;Torres-Galea et al., 2006; Fode et al.,2008; Lee et al., 2008). While it will beinteresting to learn whether the nono-verlapping expression patterns ofSHR, SCL29, and SCL32 are associ-ated with equally distinct functions,our results already assist in the char-acterization of a family of plant-spe-cific transcription factors in leafdevelopment.
EXPERIMENTAL
PROCEDURES
Terminology and Notation
We apply the generic term ‘‘subepi-dermal’’ to all positions of the leafbeneath the epidermis. We refer to‘‘ground cells’’ as polygonal, isodia-metric, subepidermal cells of the leaf.We use the terms ‘‘procambial’’ and‘‘procambium’’ to indicate morphologi-cally identifiable vascular cell precur-sors. We designate as ‘‘preprocambial’’all stages of vein development beforeprocambium formation. We adopt the‘‘::’’ and ‘‘:’’ symbols to denote tran-scriptional and translational genefusions, respectively.
Vector Construction
All amplifications were performed onArabidopsis (Arabidopsis thaliana)ecotype Col-0 genomic DNA usingFinnzymes Phusion high-fidelity DNApolymerase (New England BiolabsInc., Ipswich, MA) and gene-specificprimers (Supp. Table S1, which isavailable online). To generate theSHR::HTA6:EYFP construct, the2487-bp region from �2503 to �16 ofthe SHR gene (AT4G37650) wasrecombined into the pFYTAG vector(Zhang et al., 2005). To generate theSHR::mCherry-Nuc construct, the2494-bp region of the SHR gene from�2504 to �10 was cloned upstream ofa translational fusion of the mCherrycoding sequence (Shaner et al., 2004)to the 3xSV40 nuclear localization sig-nal from pEYFP-Nuc (Clontech Labo-ratories, Mountain View, CA). To gen-
erate the SHR::SHR:EYFP construct,the 4107-bp region of the SHR genefrom �2514 to þ1593 was clonedupstream of the EYFP codingsequence (Clontech) using an Asp-Pro-Gly linker, as described in Gallagheret al., 2004. To generate theSCL29::HTA6:EYFP construct, the1679-bp region from �1686 to �7 ofthe SCL29 gene (AT3G13840) wasrecombined into the pFYTAG vector(Zhang et al., 2005). To generate theSCL29::SCL29:EYFP construct, the3227-bp region of the SCL29 genefrom �1697 to þ1530 was clonedupstream of the EYFP codingsequence (Clontech) using a Pro-Asp-Pro-Gly linker. To generate theSCL32::HTA6:EYFP construct, the2886-bp region from �2888 to �2 ofthe SCL32 gene (AT3G49950) wasrecombined into the pFYTAG vector(Zhang et al., 2005). To generate theSCL32::SCL32:EYFP construct, the4169-bp of the SCL32 gene from�2940 to þ1229 was cloned upstreamof the EYFP coding sequence (Clon-tech) using an Asp-Pro-Gly linker.
Plant Material and Growth
Conditions
The J2501 and Q0990::mGFP5erenhancer-trap lines of the Haseloff col-lection (Haseloff, 1999) were obtainedfrom the Arabidopsis Biological Re-source Center. TheWOL::HTA6:EGFPline was a generous gift of David Gal-braith’s. The origins of the ATHB8::HTA6:EYFP, UBQ10::EGFP:LTI6B,ATHB8::ECFP-Nuc and ATHB8::ATHB8:mCherry lines have been des-cribed (Sawchuk et al., 2007, 2008;Donner et al., 2009). Seeds were steri-lized and germinated, and seedlingsand plants were grown, transformed,and selected as described (Sawchuket al., 2007, 2008). For SHR::HTA6:EYFP, SHR::mCherry-Nuc, SCL29::HTA6:EYFP, SCL32::HTA6:EYFP,SHR::SHR: EYFP, SCL29::SCL29:EYFP and SCL32::SCL32:EYFP, theprogeny of 10 to 26 independenttransgenic lines were inspected toidentify the most representativeexpression pattern. Successive expres-sion analysis was performed on theprogeny of at least three lines per con-struct, which were selected because ofstrong fluorescent protein expressionthat was emblematic of the expression
profile observed across the entire se-ries of transgenic lines and thatresulted from single insertion of thetransgene. In genetic crosses, theprogeny of at least two independentlines per construct were examined.For auxin transport inhibition, seedswere germinated on growth mediumsupplemented with 2.5 mM NPA(Chem Service Inc., West Chester, PA).We define ‘‘days after germination’’(DAG) as days following exposure ofimbibed seeds to light.
Microscopy and Image
Processing
Dissected seedling organs weremounted and imaged as described(Sawchuk et al., 2007, 2008; Donneret al., 2009). Brightness and contrastwere adjusted through linear stretch-ing of the histogram in ImageJ(National Institutes of Health, http://rsb.info.nih.gov/ij). Signal levels andcolocalization were visualized asdescribed (Sawchuk et al., 2008).
ACKNOWLEDGMENTSWe thank the Arabidopsis BiologicalResource Center, Robert Campbell,David Galbraith, and Roger Tsien forkindly providing seeds and plasmids;and Megan Sawchuk and Osama Odatfor critically reading the manuscript.This work was supported by a Discov-ery Grant of the Natural Sciences andEngineering Research Council of Can-ada (NSERC). E.S. was supported bythe Canada Research Chairs Program.T.J.D. was supported by an NSERCCGS-M Scholarship, an NSERC CGS-D Scholarship and an Alberta Ingenu-ity Student Scholarship.
REFERENCES
Alonso-Peral MM, Candela H, del PozoJC, Martinez-Laborda A, Ponce MR,Micol JL. 2006. The HVE/CAND1 geneis required for the early patterning ofleaf venation in Arabidopsis. Develop-ment 133:3755–3766.
Bailey-Serres J, Sorenson R, JuntawongP. 2009. Getting the message across:cytoplasmic ribonucleoprotein com-plexes. Trends Plant Sci 14:443–453.
Baima S, Nobili F, Sessa G, Lucchetti S,Ruberti I, Morelli G. 1995. The expres-sion of the Athb-8 homeobox gene is re-stricted to provascular cells in Ara-bidopsis thaliana. Development 121:4171–4182.
268 GARDINER ET AL.
Dev
elop
men
tal D
ynam
ics

Belostotsky DA, Sieburth LE. 2009. Killthe messenger: mRNA decay and plantdevelopment. Curr Opin Plant Biol 12:96–102.
Benkova E, Michniewicz M, Sauer M,Teichmann T, Seifertova D, Jurgens G,Friml J. 2003. Local, efflux-dependentauxin gradients as a common modulefor plant organ formation. Cell 115:591–602.
Berleth T, Sachs T. 2001. Plant morpho-genesis: long-distance coordination andlocal patterning. Curr Opin Plant Biol4:57–62.
Birnbaum K, Shasha DE, Wang JY, JungJW, Lambert GM, Galbraith DW, BenfeyPN. 2003. A gene expression map of theArabidopsis root. Science 302:1956–1960.
Bolle C. 2004. The role of GRAS proteinsin plant signal transduction and devel-opment. Planta 218:683–692.
Bolle C, Koncz C, Chua NH. 2000. PAT1,a new member of the GRAS family, isinvolved in phytochrome A signal trans-duction. Genes Dev 14:1269–1278.
Candela H, Martinez-Laborda A, MicolJL. 1999. Venation pattern formation inArabidopsis thaliana vegetative leaves.Dev Biol 205:205–216.
Carland FM, Nelson T. 2004. Cotyledonvascular pattern2-mediated inositol(1,4,5) triphosphate signal transductionis essential for closed venation patternsof Arabidopsis foliar organs. Plant Cell16:1263–1275.
Carland F, Nelson T. 2009. CVP2- andCVL1-mediated phosphoinositide sig-naling as a regulator of the ARF GAPSFC/VAN3 in establishment of foliarvein patterns. Plant J 59:895–907.
Carland F, Fujioka S, Nelson T. 2010. Thesterol methyltransferases SMT1, SMT2,and SMT3 influence Arabidopsis devel-opment through nonbrassinosteroidproducts. Plant Physiol 153:741–756.
Cheng Y, Dai X, Zhao Y. 2006. Auxin bio-synthesis by the YUCCA flavin monoox-ygenases controls the formation of floralorgans and vascular tissues in Arabi-dopsis. Genes Dev 20:1790–1799.
Cnops G, Neyt P, Raes J, Petrarulo M,Nelissen H, Malenica N, Luschnig C,Tietz O, Ditengou F, Palme K, Azmi A,Prinsen E, Van Lijsebettens M. 2006.The TORNADO1 and TORNADO2genes function in several patterningprocesses during early leaf developmentin Arabidopsis thaliana. Plant Cell 18:852–866.
Dello Ioio R, Linhares FS, Scacchi E, Casa-mitjana-Martinez E, Heidstra R, Costan-tino P, Sabatini S. 2007. Cytokininsdetermine Arabidopsis root-meristem sizeby controlling cell differentiation. CurrBiol 17:678–682.
Demandolx D, Davoust J. 1997. Multicol-our analysis and local image correlationin confocal microscopy. J Microsc 185:21–36.
Dengler N, Kang J. 2001. Vascular pat-terning and leaf shape. Curr OpinPlant Biol 4:50–56.
Di Laurenzio L, Wysocka-Diller J, Mal-amy JE, Pysh L, Helariutta Y, Freshour
G, Hahn MG, Feldmann KA, BenfeyPN. 1996. The SCARECROW gene reg-ulates an asymmetric cell division thatis essential for generating the radial or-ganization of the Arabidopsis root. Cell86:423–433.
Dill A, Sun T. 2001. Synergistic derepres-sion of gibberellin signaling by remov-ing RGA and GAI function inArabidopsis thaliana. Genetics 159:777–785.
Dill A, Jung HS, Sun TP. 2001. TheDELLA motif is essential for gibberel-lin-induced degradation of RGA. ProcNatl Acad Sci U S A 98:14162–14167.
Donner TJ, Sherr I, Scarpella E. 2009. Reg-ulation of preprocambial cell state acqui-sition by auxin signaling in Arabidopsisleaves. Development 36:3235–3246.
Esau K. 1943. Origin and development ofprimary vascular tissues in plants. BotRev 9:125–206.
Esau K. 1965. Vascular differentiation inplants. New York: Holt, Rinehart andWinston. 170 p.
Fode B, Siemsen T, Thurow C, Weigel R,Gatz C. 2008. The Arabidopsis GRASprotein SCL14 interacts with class IITGA transcription factors and is essen-tial for the activation of stress-induciblepromoters. Plant Cell 20:3122–3135.
Foster AS. 1952. Foliar venation in angio-sperms from an ontogenetic standpoint.Am J Bot 39:752–766.
Fu X, Richards DE, Fleck B, Xie D, Bur-ton N, Harberd NP. 2004. The Arabi-dopsis mutant sleepy1gar2–1 proteinpromotes plant growth by increasingthe affinity of the SCFSLY1 E3 ubiqui-tin ligase for DELLA protein sub-strates. Plant Cell 16:1406–1418.
Gallagher KL, Paquette AJ, Nakajima K,Benfey PN. 2004. Mechanisms regulat-ing SHORT-ROOT intercellular move-ment. Curr Biol 14:1847–1851.
Gardiner J, Sherr I, Scarpella E. 2010.Expression of DOF genes identifies earlystages of vascular development in Arabi-dopsis leaves. Int JDevBiol 54:1389–1396.
Greb T, Clarenz O, Schafer E, Muller D,Herrero R, Schmitz G, Theres K. 2003.Molecular analysis of the LATERALSUPPRESSOR gene in Arabidopsisreveals a conserved control mechanismfor axillary meristem formation. GenesDev 17:1175–1187.
Hardtke CS, Berleth T. 1998. The Arabi-dopsis gene MONOPTEROS encodes atranscription factor mediating embryoaxis formation and vascular develop-ment. EMBO J 17:1405–1411.
Haseloff J. 1999. GFP variants for multi-spectral imaging of living cells. Meth-ods Cell Biol 58:139–151.
Helariutta Y, Fukaki H, Wysocka-Diller J,Nakajima K, Jung J, Sena G, HauserMT, Benfey PN. 2000. The SHORT-ROOT gene controls radial patterningof the Arabidopsis root through radialsignaling. Cell 101:555–567.
Hirota A, Kato T, Fukaki H, Aida M,Tasaka M. 2007. The auxin-regulatedAP2/EREBP gene PUCHI is requiredfor morphogenesis in the early lateral
root primordium of Arabidopsis. PlantCell 19:2156–2168.
Inoue T, Higuchi M, Hashimoto Y, Seki M,Kobayashi M, Kato T, Tabata S, Shino-zaki K, Kakimoto T. 2001. Identificationof CRE1 as a cytokinin receptor fromAra-bidopsis. Nature 409:1060–1063.
Ito T, Sakai H, Meyerowitz EM. 2003.Whorl-specific expression of the SUPER-MAN gene of Arabidopsis is mediated bycis elements in the transcribed region.Curr Biol 13:1524–1530.
Kang J, Dengler N. 2002. Cell cycling fre-quency and expression of the homeoboxgene ATHB-8 during leaf vein develop-ment in Arabidopsis. Planta 216:212–219.
Kang J, Dengler N. 2004. Vein pattern de-velopment in adult leaves of Arabidopsisthaliana. Int J Plant Sci 165:231–242.
Kang J, Mizukami Y, Wang H, Fowke L,Dengler NG. 2007. Modification of cellproliferation patterns alters leaf veinarchitecture in Arabidopsis thaliana.Planta 226:1207–1218.
Kinsman EA, Pyke KA. 1998. Bundlesheath cells and cell-specific plastid de-velopment in Arabidopsis leaves. Devel-opment 125:1815–1822.
Koizumi K, Sugiyama M, Fukuda H.2000. A series of novel mutants of Ara-bidopsis thaliana that are defective inthe formation of continuous vascularnetwork: calling the auxin signal flowcanalization hypothesis into question.Development 127:3197–3204.
Konishi M, Yanagisawa S. 2007. Sequen-tial activation of two Dof transcriptionfactor gene promoters during vasculardevelopment in Arabidopsis thaliana.Plant Physiol Biochem 45:623–629.
Langdale JA, Nelson T. 1991. Spatial reg-ulation of photosynthetic developmentin C4 plants. Trends Genet 7:191–196.
Lee MH, Kim B, Song SK, Heo JO, YuNI, Lee SA, Kim M, Kim DG, Sohn SO,Lim CE, Chang KS, Lee MM, Lim J.2008. Large-scale analysis of the GRASgene family in Arabidopsis thaliana.Plant Mol Biol 67:659–670.
Lee S, Cheng H, King KE, Wang W, He Y,Hussain A, Lo J, Harberd NP, Peng J.2002. Gibberellin regulates Arabidopsisseed germination via RGL2, a GAI/RGA-like gene whose expression is up-regulated following imhibition. GenesDev 16:646–658.
Louis J. 1935. L’ontogenese du systemeconducteur dans la pousse feuillee desDicotyles et des Gymnospermes. La Cel-lule 44:87–172.
Mahonen AP, Bonke M, Kauppinen L, Riiko-nen M, Benfey PN, Helariutta Y. 2000. Anovel two-component hybrid molecule reg-ulates vascular morphogenesis of the Ara-bidopsis root. Genes Dev 14:2938–2943.
Mason MG, Li J, Mathews DE, Kieber JJ,Schaller GE. 2004. Type-B responseregulators display overlapping expres-sion patterns in Arabidopsis. PlantPhysiol 135:927–937.
Mattsson J, Sung ZR, Berleth T. 1999.Responses of plant vascular systems toauxin transport inhibition. Develop-ment 126:2979–2991.
PREPROCAMBIAL CELL STATE TRANSITION 269
Dev
elop
men
tal D
ynam
ics

Mustroph A, Zanetti ME, Jang CJ, HoltanHE, Repetti PP, Galbraith DW, Girke T,Bailey-Serres J. 2009. Profiling transla-tomes of discrete cell populationsresolves altered cellular priorities dur-ing hypoxia in Arabidopsis. Proc NatlAcad Sci U S A 106:18843–18848.
Nakajima K, Sena G, Nawy T, BenfeyPN. 2001. Intercellular movement ofthe putative transcription factor SHRin root patterning. Nature 413:307–311.
Nelson T, Dengler N. 1997. Leaf vascularpattern formation. Plant Cell 9:1121–1135.
Peng J, Carol P, Richards DE, King KE,Cowling RJ, Murphy GP, Harberd NP.1997. The Arabidopsis GAI gene definesa signaling pathway that negativelyregulates gibberellin responses. GenesDev 11:3194–3205.
Petersson SV, Johansson AI, KowalczykM, Makoveychuk A, Wang JY, Moritz T,Grebe M, Benfey PN, Sandberg G,Ljung K. 2009. An auxin gradient andmaximum in the Arabidopsis root apexshown by high-resolution cell-specificanalysis of IAA distribution and synthe-sis. Plant Cell 21:1659–1668.
Petricka JJ, Nelson TM. 2007. Arabidop-sis nucleolin affects plant developmentand patterning. Plant Physiol 144:173–186.
Pineau C, Freydier A, Ranocha P, Jau-neau A, Turner S, Lemonnier G, RenouJP, Tarkowski P, Sandberg G, JouaninL, Sundberg B, Boudet AM, Goffner D,Pichon M. 2005. hca: an Arabidopsismutant exhibiting unusual cambial ac-tivity and altered vascular patterning.Plant J 44:271–289.
Przemeck GK, Mattsson J, Hardtke CS,Sung ZR, Berleth T. 1996. Studies onthe role of the Arabidopsis geneMONOPTEROS in vascular develop-ment and plant cell axialization. Planta200:229–237.
Pysh LD, Wysocka-Diller JW, Camilleri C,Bouchez D, Benfey PN. 1999. TheGRAS gene family in Arabidopsis:sequence characterization and basicexpression analysis of the SCARE-
CROW-LIKE genes. Plant J 18:111–119.Sachs T. 1975. Control of Differentiationof Vascular Networks. Ann Bot 39:197–204.
Sachs T. 1989. The development of vascu-lar networks during leaf development.Curr Top Plant Biochem Physiol 8:168–183.
Sawchuk MG, Head P, Donner TJ, Scar-pella E. 2007. Time-lapse imaging ofArabidopsis leaf development showsdynamic patterns of procambium forma-tion. New Phytol 176:560–571.
Sawchuk MG, Donner TJ, Head P, Scar-pella E. 2008. Unique and overlappingexpression patterns among members ofphotosynthesis-associated nuclear genefamilies in Arabidopsis. Plant Physiol148:1908–1924.
Scarpella E, Francis P, Berleth T. 2004.Stage-specific markers define early stepsof procambium development in Arabidop-sis leaves and correlate termination ofvein formation with mesophyll differen-tiation. Development 131:3445–3455.
Scarpella E, Marcos D, Friml J, BerlethT. 2006. Control of leaf vascular pat-terning by polar auxin transport. GenesDev 20:1015–1027.
Shaner NC, Campbell RE, Steinbach PA,Giepmans BN, Palmer AE, Tsien RY.2004. Improved monomeric red, orangeand yellow fluorescent proteins derivedfrom Discosoma sp. red fluorescent pro-tein. Nat Biotechnol 22:1567–1572.
Sieburth LE. 1999. Auxin is required forleaf vein pattern in Arabidopsis. PlantPhysiol 121:1179–1190.
Silverstone AL, Ciampaglio CN, Sun T.1998. The Arabidopsis RGA gene enco-des a transcriptional regulator repres-sing the gibberellin signal transductionpathway. Plant Cell 10:155–169.
Steinmann T, Geldner N, Grebe M, MangoldS, Jackson CL, Paris S, Galweiler L,Palme K, Jurgens G. 1999. Coordinatedpolar localization of auxin efflux carrierPIN1 by GNOM ARF GEF. Science 286:316–318.
Steynen QJ, Schultz EA. 2003. TheFORKED genes are essential for distal
vein meeting in Arabidopsis. Develop-ment 130:4695–4708.
Suzuki T, Miwa K, Ishikawa K, YamadaH, Aiba H, Mizuno T. 2001. The Arabi-dopsis sensor His-kinase, AHK4, canrespond to cytokinins. Plant Cell Phys-iol 42:107–113.
Telfer A, Poethig RS. 1994. Leaf develop-ment in Arabidopsis. In: MeyerowitzEM, Somerville CR, editors. Arabidop-sis. New York: Cold Spring HarborPress. p 379–401.
Torres-Galea P, Huang LF, Chua NH, BolleC. 2006. The GRAS protein SCL13 is apositive regulator of phytochrome-de-pendent red light signaling, but can alsomodulate phytochrome A responses. MolGenet Genomics 276:13–30.
Tsuchisaka A, Theologis A. 2004. Uniqueand overlapping expression patternsamong the Arabidopsis 1-amino-cyclo-propane-1-carboxylate synthase genefamily members. Plant Physiol 136:2982–3000.
Tyler L, Thomas SG, Hu J, Dill A, AlonsoJM, Ecker JR, Sun TP. 2004. Della pro-teins and gibberellin-regulated seedgermination and floral developmentin Arabidopsis. Plant Physiol 135:1008–1019.
Ueki S, Citovsky V. 2000. IntercellularRNA transport in plants. Curr TopPlant Biol 2:167–179.
Wang JW, Wang LJ, Mao YB, Cai WJ,Xue HW, Chen XY. 2005. Control ofroot cap formation by MicroRNA-tar-geted auxin response factors in Arabi-dopsis. Plant Cell 17:2204–2216.
Wen CK, Chang C. 2002. Arabidopsis RGL1encodes a negative regulator of gibberellinresponses. Plant Cell 14:87–100.
Wenzel CL, Schuetz M, Yu Q, Mattsson J.2007. Dynamics of MONOPTEROS andPIN-FORMED1 expression during leafvein pattern formation in Arabidopsisthaliana. Plant J 49:387–398.
Zhang C, Gong FC, Lambert GM, Gal-braith DW. 2005. Cell type-specificcharacterization of nuclear DNA con-tents within complex tissues andorgans. Plant Methods 1:7.
270 GARDINER ET AL.
Dev
elop
men
tal D
ynam
ics




















![[Manual] SHR 1041K](https://img.pdfslide.us/doc/110x75/547ca951b4af9f8a138b45c5/manual-shr-1041k.jpg)








