Embed Size (px)
Citation preview

682
Environmental Toxicology and Chemistry, Vol. 23, No. 3, pp. 682–690, 2004q 2004 SETAC
Printed in the USA0730-7268/04 $12.00 1 .00
SILVER UPTAKE BY A MARINE DIATOM AND ITS TRANSFER TO THE COASTALCOPEPOD ACARTIA SPINICAUDA
YAN XU and WEN-XIONG WANG*Department of Biology, The Hong Kong University of Science and Technology, Clear Water Bay, Kowloon, Hong Kong, Republic of China
(Received 11 December 2002; Accepted 24 July 2003)
Abstract—Silver (Ag) is an important metal contaminant in many coastal waters and often is accompanied by high nutrientconcentrations in the effluent outfall. The biological uptake of Ag by the marine diatom Thalassiosira pseudonana at various levelsof nitrate, ammonium, and phosphate was examined under different growth conditions of the cells. The biological uptake of Agincreased significantly with increasing concentrations of nitrate, ammonium, and phosphate, presumably because of the increase inalgal growth rate or increasing sulfur-containing ligands available for Ag transport. The calculated Ag uptake rate constants increasedby 3 to 16 times with increasing nitrate and ammonium concentration from 5.88 mM to 176 mM. The assimilation efficiency (AE)of Ag by the coastal copepod Acartia spinicauda was quantified under different ecological and chemical conditions of the diatomprey. The Ag AEs were in the range of 3 to 23%, and increased with decreasing diatom food concentrations or when the diatomswere grown at a lower nutrient level. Significant correlations were demonstrated between the AE and the distribution of Ag in thediatom’s cytoplasm, the Ag concentration factor in the diatoms, Ag retention in the particles during the feeding period, and the Aggut passage time in the copepods. Desorption within the copepod’s gut appears to play a critical role in Ag assimilation and partiallyaccounts for the variability of Ag AEs under different food and geochemical conditions. Our study highlights that several geochemicaland physiological processes all significantly affect Ag trophic transfer in marine copepods. Given the dependence of Ag transferon its concentration in ingested particles and food concentration, dietary uptake of Ag is probably variable in natural environments.The influence of eutrophication on Ag trophic transfer is dependent on the degree to which each kinetic parameter is affected bynutrient enrichments.
Keywords—Silver Macronutrients Plankton Uptake Trophic transfer
INTRODUCTION
Extensive studies on metal accumulation and trophic trans-fer in marine plankton have been conducted over the past 10years [1–8]. Factors potentially influencing metal accumula-tion and trophic transfer in marine copepods have been ex-tensively considered. Among the many environmental factors,the influences of macronutrients on metal uptake in marinephytoplankton and subsequent transfer to marine copepodshave been examined only recently [9,10]. These earlier studiesfocused on a few oceanographically and environmentally im-portant metals such as Cd, Fe, Se, and Zn. Macronutrients,particularly nitrate, were found to substantially affect the up-take of Cd, Fe, and Zn by marine phytoplankton. Metals con-sidered in these previous studies (Fe, Se, and Zn) are knownto be essential to marine phytoplankton. Cadmium has alsobeen shown to be essential for the marine diatom Thalassiosiraweissflogii under Zn-limited conditions [11]. Whether the mac-ronutrients affect the uptake of nonessential metals such as Agis a matter of speculation.
Silver is highly toxic to marine phytoplankton and zoo-plankton [12–15]. The acute toxicity concentration of Ag tosensitive marine species such as bivalve larvae and copepodsis on the order of 100 nM [16,17]. Silver toxicity to marineinvertebrates may be manifested by reproductive failure[18,19]. Hook and Fisher [19] showed that dietary Ag reducedegg production of copepods at a much lower environmentalconcentration than that causing toxicity from the aqueousphase. Reproductive success of copepods decreased by .50%when the algal diets were exposed to only 1 nM Ag as com-
* To whom correspondence may be addressed ([email protected]).
pared to a 50% lethal concentration of 400 nM from the aque-ous phase. Clearly, Ag toxicology is influenced by the routesof Ag accumulation in the animals, and a considerable needexists to understand the exposure of zooplankton to Ag fromdifferent environmental phases [20,21]. In the marine copepodTemora longicornis, Wang and Fisher [4] modeled Ag ex-posure and indicated that about 20 to 50% of Ag was accu-mulated from dietary exposure.
Silver is an important pollutant in effluent discharge[22,23]. Because of its strong particle reactivity, Ag can beused as a tracer of particles in effluent discharge. Effluentdischarge often contains relatively high concentrations of mac-ronutrients such as N and P, but the interactions between thesemacronutrients and Ag in the marine environment are un-known. Furthermore, the bioaccumulation of Ag in the marinefood chain is not well understood despite its environmentalrelevance. Transfer of Ag from phytoplankton prey to zoo-plankton has only been sporadically examined [1,2,4]. Amongthe many processes possibly affecting Ag concentration inzooplankton, assimilation is an important process leading topotential Ag dietary accumulation. Factors affecting metal di-etary exposure in marine zooplankton include metal partition-ing in the algal cytoplasm [1,9], metal gut passage time, andfood passage time [6]. The relative importance of each processin the assimilation of a specific metal is unknown, but suchinformation is important for modeling metal transport in theenvironment. More recent studies suggest that metal assimi-lation in marine copepods is rather complicated and is gov-erned by a combination of different geochemical and physi-ological processes [6,7,9].
In this study, we quantified the biological uptake of Ag by

Silver uptake by marine plankton Environ. Toxicol. Chem. 23, 2004 683
the diatom Thalassiosira pseudonana at different nutrient con-ditions. The transfer of diatom-bound Ag to the copepod Acar-tia spinicauda was further quantified by measurements of Agassimilation efficiency (AE). The Ag AE was determined underdifferent diatom concentrations and nutritional conditions. Wefurther examined whether the particle reactivity of Ag in thediatoms affected its assimilation by the copepods, by usingdiatoms spiked with Ag for different durations. Several pro-cesses pertaining to Ag assimilation were specifically consid-ered in this study, including the Ag subcellular cytoplasmicdistributions, Ag gut passage time in the copepods, Ag con-centration factor in the food particles, as well as Ag retentionin food particles during radioactive feeding periods. Theseconcurrent measurements can provide insights into the mech-anisms of Ag assimilation in copepods and can help furtherunderstand the significance of Ag dietary uptake in copepods,which may have a contrasting toxicity when compared withaqueous Ag.
MATERIALS AND METHODS
Phytoplankton and copepods
The coastal diatom T. pseudonana (clone 3H) was obtainedfrom the Provasoli-Guillard Phytoplankton Collection Center(West Boothbay Harbor, ME, USA) and was maintained inaxenic conditions in an f/2 medium [24] at 188C with a lightillumination of 100 mmol photons/m2/s in a 14:10 h light:darkcycle. Seawater was collected 10 km east of Hong Kong, re-mote from any anthropogenic nutrient input. Adult copepods(A. spinicauda) were collected by plankton tow (250-mm meshsize) from Port Shelter (Clear Water Bay, Hong Kong) andwere maintained under laboratory conditions for 1 d and fedthe algal food T. pseudonana before the experiments.
Silver uptake by diatoms under different nutrientconditions
Radiotracer 110mAg (in 0.1 HNO3, with a specific activityof 106 mBq/mg Ag, Riso National Laboratory, Roskilde, Den-mark) was used to trace Ag uptake by the diatoms. We firstquantified the efficiency of the stable Ag rinse (100 nM)[25,26] in removing extracellular Ag from the diatoms to quan-tify the intracellular uptake. The diatom cells were exposed to110mAg for 1 min, and a 10-ml aliquot was filtered onto a 1-mm polycarbonate membrane, rinsed with filtered seawater,and then rinsed with stable Ag (100 nM) for 3 min. After therinse, the radioactivity of the cells on the membrane was gam-ma counted for 3 min. Results indicated that about .74% ofsurface-adsorbed 110mAg was removed by 100 nM stable Agrinse.
All uptake experiments were conducted in 150 ml of 0.2-mm-filtered seawater held in acid-cleaned polycarbonate flasksand maintained at 188C with a light illumination of 100 mmolphotons/m2/s. To measure the Ag uptake, the cells inoculatedunder different nutrient conditions (see below) were collectedonto 3-mm polycarbonate membranes, rinsed, and resuspendedin 0.22-mm-filtered seawater. The cells were then added to 150ml of 0.22-mm-filtered seawater containing 110mAg and differ-ent additions of nutrients. Any trace metals in the nutrientstock had been removed by passing it through a Chelex ionexchange resin (BioRad Laboratories, Hercules, CA, USA).Radioisotope addition corresponded to an Ag concentration of3.1 nM. Only the experimental macronutrient was added intothe medium for Ag uptake measurement. The cell density in
the medium was 5 3 105 cells/ml for all the uptake experi-ments. Each experimental treatment had two replicated bottles.At 0.5, 1, 1.5, and 2 h (during which time the nutrient con-centrations decreased by ,20%), a 10-ml aliquot was filteredonto a 1-mm polycarbonate membrane, rinsed with filteredseawater, and then soaked with 100 nM stable Ag for 3 min,after which the radioactivity of the cells was counted. At theend of the uptake experiment, a 10-ml aliquot was filtered ontoa 1 mm polycarbonate membrane, rinsed with filtered seawater,and its radioactivity was counted (representing the Ag accu-mulation in the whole cell). Another 10-ml aliquot was filtered,placed in freshwater, and then frozen. Changes in nutrientconcentrations during the 2-h exposure period were not quan-tified. The distribution of Ag in the diatom’s cytoplasm waslater quantified by differential centrifugation technique, as de-scribed by Reinfelder and Fisher [1] and Xu and Wang [6].Changes in cell density during the 2-h radioactive exposureperiod were determined by a Coulter counter (Fullerton, CA,USA). To measure the dry weight of the cells, the cells werefiltered onto a preweighed glass fiber filter, rinsed with 0.5 Mammonium formate, and dried at 808C for 1 d. The dry weightconcentration factor (DCF) was calculated as the ratio of Agintracellular accumulated concentrations (disintegrations perminute [dpm]/kg dry wt) divided by the Ag concentration inthe dissolved phase (dpm/L).
Three experiments were conducted to determine the influ-ences of macronutrients on the Ag uptake by the diatoms. Thediatom cells were grown at f/10 levels (i.e., five times lowerthan the f/2 levels) of the targeted nutrient (176 mM nitrate,7.24 mM phosphate, or 176 mM ammonium) and f/2 levels ofother nutrients [24]. For all experiments, cell growth was mon-itored on a daily basis. In the first experiment, we examinedthe uptake of Ag by diatoms grown under semicontinuousconditions (e.g., transferring the cultures at time intervals) atdifferent nitrate, phosphate, or ammonium concentrations.Cells in the exponential growth phase were filtered and trans-ferred to seawater containing different nitrate (5.88, 35.3, and176 mM), phosphate (0.24, 1.45, and 7.24 mM), or ammonium(5.88, 35.3, and 176 mM) concentrations in the semicontinuousculture. Other macronutrients were added at f/10 levels. Back-ground nutrient concentrations in the seawater were ,3.5 mMfor nitrate and ,0.2 mM for phosphate. The cells were trans-ferred to a new medium containing the same macronutrientconcentration when they reached the midexponential growthphase (every 1–1.5 d). After three transfers (ensuring that thecells were acclimated to the specific nutrient conditions), thecells were filtered again and resuspended in filtered seawater,and Ag uptake was determined at each macronutrient level asdescribed above. In the second experiment, we determined Aguptake by diatoms grown at different nitrate or ammoniumconcentrations for a relatively shorter period. The late expo-nentially growing diatom cells inoculated in f/10 nutrient me-dium were transferred to different nitrate (5.88, 35.3, and 176mM) or ammonium (5.88, 35.3, and 176 mM) concentrationsfor 1 d and Ag uptake was determined. In the third experiment,we determined the influence of nutrient (nitrate or phosphate)starvation on Ag uptake in diatom cells. The late exponentiallygrowing diatom cells inoculated in f/10 nutrient medium werefiltered and resuspended into either nutrient-depleted seawateror nutrient-repleted seawater (added at 176 mM nitrate or 7.24mM phosphate and other nutrients at the f/2 levels). After 2 dof starvation for nitrate or of 3 d starvation for phosphate (i.e.,no cell growth occurred afterwards), both the starved and en-

684 Environ. Toxicol. Chem. 23, 2004 Y. Xu and W.-X. Wang
Fig. 1. Intracellular accumulation (quantified as the dry wt concen-tration factor [DCF]) of Ag by the diatom Thalassiosira pseudonanamaintained at different nitrate concentrations under semicontinuous(A), short-term acclimation (B), and starvation (C) conditions. 2N/2N 5 N-starved cells in N-depleted medium; 2N/1N 5 N-starvedcells in N-repleted medium; 1N/2N 5 N-repleted cells in N-depletedmedium; 1N/1N 5 N-repleted cells in N-repleted medium. Valuesare means 6 semirange (n 5 2).
riched cells were collected and resuspended in both nutrient-depleted and nutrient-enriched filtered seawater and Ag uptakewas measured.
Trophic transfer of silver in marine copepods
Assimilation and elimination of Ag in the marine copepodsA. spinicauda feeding on diatom cells maintained under dif-ferent nutrient conditions or diatom concentrations were de-termined. The diatom cells were first maintained in semicon-tinuous culture (5.88, 35.3, and 176 mM for nitrate, and 0.24,1.45, and 7.24 mM for phosphate), or under starvation con-ditions, as described above. After three transfers, the cells werecollected and resuspended in 100 ml of filtered seawater con-taining the same nutrient level as that used during the accli-mation. The culture was then spiked with 110mAg and the cellswere radiolabeled for 2 h. Different amounts of radioactivitywere added to result in a comparable Ag cellular concentrationamong the different experimental treatments. The second ex-periment quantified Ag assimilation at different diatom con-centrations. Diatom cells in the late exponential growth phasewere filtered and resuspended into filtered seawater containingf/2 levels of N, P, Si, and vitamins and f/20 levels of Mn, Co,Fe, and Mo. The cells were radiolabeled with 110mAg for 1 d.The third experiment determined the influence of Ag contacttime with diatoms on Ag assimilation. Cells in the late ex-ponential growth phase were filtered and resuspended into 100ml of filtered seawater containing the nutrients as describedabove. The cells were then radiolabeled with radiotracer for1, 2, 8, and 24 h. The spikes were conducted at different timessuch that the assimilation experiment was carried out simul-taneously for different treatments. All the radiolabeled cellswere soaked with 100 nM stable Ag for 3 min and then rinsedwith filtered seawater to remove extracellular Ag before beingfed to copepods.
The radiolabeled diatoms were pulse-fed to the copepodsfor 30 min in the dark at a cell density of 200,000 cells/ml,except for the cell density experiment, in which the cell den-sities were 4,500, 20,000, 80,000, and 200,000, cells/ml (cor-responding to biomass of 0.10, 0.44, 1.76, and 4.40 mg/L,respectively). Copepod density was 0.7 individuals/ml. Afterthe pulse radioactive feeding, the copepods were collected ontoa 250-mm nylon mesh, rinsed with filtered seawater, and theirradioactivity was counted immediately. Any feces producedduring the radioactive feeding period were collected onto a20-mm nylon mesh, rinsed with seawater, and counted for ra-dioactivity. A 10-ml aliquot of the feeding seawater was fil-tered onto a 1-mm polycarbonate membrane, rinsed with fil-tered seawater, and counted for radioactivity to determine thepercentage of Ag retained on the cells after the 30-min radio-active feeding. The copepods were placed in filtered seawaterwith the presence of the nonradiolabeled diatom cells to dep-urate for 24 h. At time intervals (every 2–4 h), the radioactivityretained in the copepods and in the feces produced duringdepuration were determined. The seawater and food were re-newed each time when the radioactivity of the copepods wascounted.
The AE and the elimination rate were calculated as the y-intercept and the slope of the linear regression between thenatural log of the percentage of metals retained in the copepodsand the time of depuration (between 8 and 24 h), respectively.The gut passage time was calculated as the time at which 90%of unassimilated Ag was recovered in the feces, assuming acomplete digestion in 24 h. Radioactivity of 110mAg was de-
termined by a Wallac gamma detector (Turku, Finland) at 658keV. Counting times of the samples were 3 min, and the prop-agated errors were generally ,3%.
RESULTS
Silver uptake in diatoms under different macronutrientconditions
Intracellular uptake of 110mAg by the diatoms under differentnitrate, ammonium, and phosphate conditions is shown in Fig-ures 1 to 3. With a few exceptions, an approximate linearpattern of Ag uptake over 0.5 and 2 h was found during the2-h exposure period. Both semicontinuous and 1-d acclimationexperiments were consistent (Fig. 1). Diatom cells had a higheruptake with increasing nitrate level. For example, the calcu-lated concentration factor (DCF) after 2 h of exposure was 6.9times and 8.9 times higher at 176 mM than at 5.88 mM in thesemicontinuous experiment and 1-d acclimation experiment,respectively. A clear difference also was found between theN-starved and N-repleted cells (Fig. 1). By the end of the 2-h exposure, the DCFs were 8.5 to 11.8 times higher for N-repleted cells than for N-depleted cells. Difference in nitrateconcentrations in the exposed medium appeared to have littleeffect on subsequent Ag uptake, although the N-repleted cellshad a somewhat higher DCF without nitrate addition than withnitrate addition.
In the ammonium experiments (Fig. 2), diatom cells ex-posed to the same level of nitrate exhibited a somewhat higher
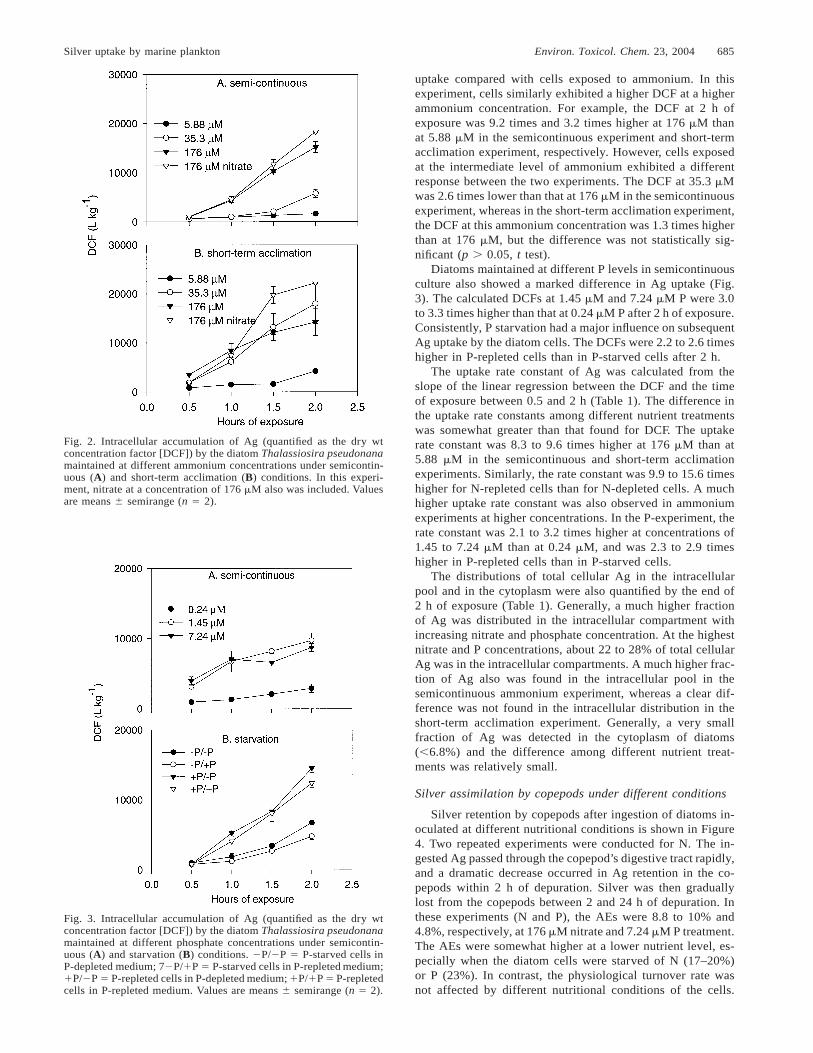
Silver uptake by marine plankton Environ. Toxicol. Chem. 23, 2004 685
Fig. 2. Intracellular accumulation of Ag (quantified as the dry wtconcentration factor [DCF]) by the diatom Thalassiosira pseudonanamaintained at different ammonium concentrations under semicontin-uous (A) and short-term acclimation (B) conditions. In this experi-ment, nitrate at a concentration of 176 mM also was included. Valuesare means 6 semirange (n 5 2).
Fig. 3. Intracellular accumulation of Ag (quantified as the dry wtconcentration factor [DCF]) by the diatom Thalassiosira pseudonanamaintained at different phosphate concentrations under semicontin-uous (A) and starvation (B) conditions. 2P/2P 5 P-starved cells inP-depleted medium; 72P/1P 5 P-starved cells in P-repleted medium;1P/2P 5 P-repleted cells in P-depleted medium; 1P/1P 5 P-repletedcells in P-repleted medium. Values are means 6 semirange (n 5 2).
uptake compared with cells exposed to ammonium. In thisexperiment, cells similarly exhibited a higher DCF at a higherammonium concentration. For example, the DCF at 2 h ofexposure was 9.2 times and 3.2 times higher at 176 mM thanat 5.88 mM in the semicontinuous experiment and short-termacclimation experiment, respectively. However, cells exposedat the intermediate level of ammonium exhibited a differentresponse between the two experiments. The DCF at 35.3 mMwas 2.6 times lower than that at 176 mM in the semicontinuousexperiment, whereas in the short-term acclimation experiment,the DCF at this ammonium concentration was 1.3 times higherthan at 176 mM, but the difference was not statistically sig-nificant (p . 0.05, t test).
Diatoms maintained at different P levels in semicontinuousculture also showed a marked difference in Ag uptake (Fig.3). The calculated DCFs at 1.45 mM and 7.24 mM P were 3.0to 3.3 times higher than that at 0.24 mM P after 2 h of exposure.Consistently, P starvation had a major influence on subsequentAg uptake by the diatom cells. The DCFs were 2.2 to 2.6 timeshigher in P-repleted cells than in P-starved cells after 2 h.
The uptake rate constant of Ag was calculated from theslope of the linear regression between the DCF and the timeof exposure between 0.5 and 2 h (Table 1). The difference inthe uptake rate constants among different nutrient treatmentswas somewhat greater than that found for DCF. The uptakerate constant was 8.3 to 9.6 times higher at 176 mM than at5.88 mM in the semicontinuous and short-term acclimationexperiments. Similarly, the rate constant was 9.9 to 15.6 timeshigher for N-repleted cells than for N-depleted cells. A muchhigher uptake rate constant was also observed in ammoniumexperiments at higher concentrations. In the P-experiment, therate constant was 2.1 to 3.2 times higher at concentrations of1.45 to 7.24 mM than at 0.24 mM, and was 2.3 to 2.9 timeshigher in P-repleted cells than in P-starved cells.
The distributions of total cellular Ag in the intracellularpool and in the cytoplasm were also quantified by the end of2 h of exposure (Table 1). Generally, a much higher fractionof Ag was distributed in the intracellular compartment withincreasing nitrate and phosphate concentration. At the highestnitrate and P concentrations, about 22 to 28% of total cellularAg was in the intracellular compartments. A much higher frac-tion of Ag also was found in the intracellular pool in thesemicontinuous ammonium experiment, whereas a clear dif-ference was not found in the intracellular distribution in theshort-term acclimation experiment. Generally, a very smallfraction of Ag was detected in the cytoplasm of diatoms(,6.8%) and the difference among different nutrient treat-ments was relatively small.
Silver assimilation by copepods under different conditions
Silver retention by copepods after ingestion of diatoms in-oculated at different nutritional conditions is shown in Figure4. Two repeated experiments were conducted for N. The in-gested Ag passed through the copepod’s digestive tract rapidly,and a dramatic decrease occurred in Ag retention in the co-pepods within 2 h of depuration. Silver was then graduallylost from the copepods between 2 and 24 h of depuration. Inthese experiments (N and P), the AEs were 8.8 to 10% and4.8%, respectively, at 176 mM nitrate and 7.24 mM P treatment.The AEs were somewhat higher at a lower nutrient level, es-pecially when the diatom cells were starved of N (17–20%)or P (23%). In contrast, the physiological turnover rate wasnot affected by different nutritional conditions of the cells.

686 Environ. Toxicol. Chem. 23, 2004 Y. Xu and W.-X. Wang
Table 1. Calculated Ag uptake rate constants and the subcellular distribution of Ag (intracellular and cytoplasmic) in the diatom Thalassiosirapseudonana after 2 h of exposure under different nutrient conditions. The growth rate constant of the diatoms during acclimation also is shown.
Values are mean 6 semirange (n 5 2)
Nutrient condition
Growth rateconstant during
acclimation (d21)
Upake rateconstant(L/kg/h)
% Ag in intracellularpool after 2 h
% Ag in cytoplasmafter 2 h
NitrateSemicontinuous
5.88 mM35.3 mM
176 mM
0.521.021.10
1,698 6 2805,609 6 171
14,044 6 361
23.5 6 2.530.5 6 0.035.1 6 1.1
1.7 6 0.32.4 6 0.31.0 6 0.1
Short-term acclimation5.88 mM
35.3 mM176 mM
0.981.251.16
1,594 6 655,179 6 21
15,388 6 376
12.1 6 0.414.3 6 0.528.0 6 0.5
4.4 6 1.42.0 6 0.22.1 6 0.2
Starvation1N/1N1N/2N2N/1N2N/2N
0.860.860.290.29
8,237 6 1,31311,688 6 907
526 6 1401,175 6 168
21.8 6 1.127.4 6 2.6
8.3 6 1.314.8 6 1.0
2.1 6 0.32.1 6 0.23.1 6 0.74.3 6 1.5
PhosphateSemicontinuous
0.24 mM1.45 mM7.24 mM
0.761.201.07
1,332 6 5314,247 6 692,763 6 695
9.7 6 1.318.8 6 1.222.9 6 1.0
1.9 6 0.01.7 6 0.54.7 6 0.3
Starvationb
1P/1P1P/2P2P/1P2P/2P
0.610.610.110.11
7,750 6 6358,831 6 2532,700 6 4243,771 6 0.22
26.4 6 0.925.6 6 2.116.1 6 1.018.1 6 0.3
2.6 6 0.02.2 6 0.16.8 6 1.35.0 6 0.1
AmmoniumSemicontinuous
5.88 mM35.3 mM
176 mM176 mM (NO3)
1.031.531.801.75
631 6 1003,336 6 4669,705 6 855
11,775 6 182
11.5 6 0.116.9 6 2.327.7 6 0.429.3 6 1.1
2.3 6 0.41.8 6 0.51.7 6 0.13.0 6 0.0
Short-term acclimation5.88 mM
35.3 mM176 mM176 mM (NO3)
1.211.481.401.25
2,071 6 9911,072 6 1952
7,155 6 1,43814,657 6 331
22.7 6 1.228.2 6 5.725.7 6 3.533.2 6 0.2
3.2 6 0.41.8 6 0.52.6 6 0.32.4 6 0.5
a 1N/1N 5 N-repleted cells in N-repleted medium; 1N/2N 5 N-repleted cells in N-depleted medium; 2N/1N 5 N-starved cells in N-repletedmedium; 2N/2N 5 N-starved cells in N-depleted medium; 1P/1P 5 P-repleted cell in P-repleted medium; 1P/2P 5 P-repleted cells in P-depleted medium; 2P/1P 5 P-starved cells in P-repleted medium; 2P/2P 5 P starved cells in P-deployed medium.
When both N and P experiments were combined together, asignificant correlation was found between the Ag AE and Agdistribution in the cytoplasm of the cells (Fig. 5). Furthermore,a significant negative correlation was found between the AgAEs and the Ag concentration factors in the diatom (measuredafter the radiolabeling) or the Ag partitioning in the diatomsduring the radioactive feeding period. A significant positivecorrelation between the Ag AE and its gut passage time in thecopepods also was observed.
A similar pattern of Ag retention was also found in thediatom concentration experiment (Fig. 6). A higher fractionof Ag was retained by the copepods at a lower cell concen-tration. The calculated Ag AEs decreased significantly withincreasing diatom concentration (p , 0.001, one-way analysisof variance). At the highest cell biomass examined (4.40 mg/L), the Ag AE was as low as 2.6%. The physiological turnoverrate was independent of the food concentration. A positiverelationship consistently was found between the Ag AE andthe Ag gut passage time in the copepods and a negative re-
lationship was found between the Ag AE and the Ag parti-tioning in the diatom cells during the radioactive feeding pe-riod (Fig. 6).
Retention of Ag in copepods after ingestion of diatomsradiolabeled with Ag for different durations is indicated inFigure 7. The calculated Ag AE decreased with increasingduration of Ag exposure to the diatoms. In this experiment,the dry weight concentration factor increased greatly with in-creasing Ag exposure time (Table 2). Similarly, the Ag AEincreased with increasing Ag distribution in the diatom’s cy-toplasm, and was negatively related to the Ag DCF in diatomsand the partitioning in diatoms during the radioactive feedingperiod (Fig. 7).
DISCUSSION
Macronutrient influences on silver uptake in diatoms
In this study, we used stable Ag (100 nM) to remove theextracellular-bound Ag and quantified the biological intracel-

Silver uptake by marine plankton Environ. Toxicol. Chem. 23, 2004 687
Fig. 4. Retention of Ag in the copepod Acartia spinicauda after theingestion of radiolabeled diatoms inoculated at different N (A and B)and P concentrations (C). Two replicated experiments were conductedfor N. 2N 5 N-starvation; 2P 5 P-starvation. Values are means 6standard deviation (n 5 3).
Fig. 6. (A) Retention of Ag in the copepod Acartia spinicauda atdifferent diatom concentrations. Relationships between the Ag assim-ilation efficiency (AE) in the copepods and (B) diatom biomass, (C)Ag partitioning in diatoms, and (D) Ag gut passage time (GPT) inthe copepods. Values are means 6 standard deviation (n 5 3).
Fig. 5. Relationships between Ag assimilation efficiency in the co-pepod Acartia spinicauda and (A) Ag subcellular distribution in di-atoms, (B) the Ag dry weight concentration factor (DCF), (C) par-titioning in diatoms, and (D) Ag gut passage time (GPT) in the co-pepods. Values are means 6 standard deviation (n 5 3).
Fig. 7. (A) Retention of Ag in the copepod Acartia spinicauda afteringestion of diatoms radiolabeled for different durations. Relation-ships between the Ag assimilation efficiency (AE) and (B) Ag sub-cellular distribution in diatoms, (C) Ag dry weight concentration fac-tor (DCF), and (D) partitioning in diatoms. Values are means 6 stan-dard deviation (n 5 3).
lular uptake by the diatom T. pseudonana. This technique hasbeen used to remove extracellular Ag in freshwater phyto-plankton [25,26]. Our study demonstrated a significant influ-ence of N and P on Ag uptake by the diatoms. Significantlyhigher uptake of Ag was documented with increasing nitrate,ammonium, or phosphate concentrations. The uptake rate in-creased by 1.4 to 16 times over the concentration ranges tested
in this study. These experimental results were consistent withour recent studies on other metals including Cd, Zn, and Fe[9,10]. For example, Wang et al. [9,10] found that the uptakeof Cd and Zn by the diatom T. pseudonana increased by 4.1times and 1.5 times, respectively, with increasing nitrate con-centration from 17.6 mM to 176 mM. Based on the uptake rateconstant, Ag apparently was the metal most affected by mac-ronutrients among the metals examined thus far (Cd, Fe, andZn). We also found a significant influence of P concentrationon Ag uptake by the diatoms, whereas Cd and Zn uptake wasnot related to the ambient P concentrations [9,10]. However,P starvation facilitated the biological uptake of Zn by thediatom, presumably because of the induction of surface stressproteins [10].
The mechanisms of macronutrient influence on Ag uptakeremain to be further explored. Silver has one of the highestbinding stability constants with sulfur-containing ligand [27].

688 Environ. Toxicol. Chem. 23, 2004 Y. Xu and W.-X. Wang
Table 2. Subcellular distribution (intracellular and cytoplasmic) and dry weight concentration factor(DCF) of Ag in the diatom Thalassiosira pseudonana inoculated under different conditions, and theAg assimilation efficiency (AE) and elimination rate constant (k) in the copepod Acartia spinicauda.
Data are mean 6 semirange or standard deviation (n 5 2–3)
Experiment DCF (L/kg)% Ag in
intracellular pool
% Ag in dia-tom
cytoplasm AE (%) k (d21)
N (Experiment 1) (mm)0 (starvation)5.88
35.3176
1,078 6 162,338 6 313,223 6 750
13,580 6 2,813
18.8 6 0.320.4 6 1.421.5 6 4.833.1 6 5.1
3.5 6 0.03.1 6 0.22.0 6 0.51.1 6 0.1
16.7 6 2.710.4 6 2.010.9 6 2.4
8.8 6 1.2
0.72 6 0.230.44 6 0.200.83 6 0.421.06 6 0.17
N (Experiment 2) (mm)0 (starvation)5.88
35.3176
1,985 6 321,691 6 933,619 6 172
13,837 6 1,794
31.5 6 1.121.4 6 1.823.1 6 1.928.0 6 2.6
2.1 6 0.32.9 6 0.72.2 6 0.21.5 6 0.1
20.4 6 5.715.6 6 3.412.0 6 2.310.0 6 1.1
0.68 6 0.060.70 6 0.190.59 6 0.120.66 6 0.06
P (mm)0 (starvation)0.241.457.24
3,821 6 33810,101 6 22721,273 6 19225,601 6 1,931
58.2 6 4.359.1 6 0.071.1 6 9.472.1 6 9.3
3.8 6 0.71.5 6 0.20.8 6 0.11.0 6 0.2
23.4 6 2.413.3 6 0.3
6.2 6 0.74.8 6 0.6
0.76 6 0.090.90 6 0.030.74 6 0.120.64 6 0.17
Cell density (mg/L)0.100.441.764.40
50,606 6 1,752 36.0 6 5.4 0.9 6 0.1 21.9 6 2.911.1 6 2.4
3.5 6 1.82.6 6 0.7
0.47 6 0.130.80 6 0.250.54 6 0.120.53 6 0.15
Labeling time (h)128
24
2,431 6 28311,504 6 42226,951 6 2,72459,656 6 3,300
29.8 6 2.142.9 6 1.155.4 6 4.370.5 6 0.8
6.6 6 0.24.3 6 0.15.2 6 0.82.7 6 0.4
11.4 6 0.45.9 6 0.14.7 6 0.22.8 6 0.4
0.67 6 0.100.75 6 0.030.84 6 0.050.65 6 0.23
After N starvation or N inoculation at a lower N level, theprotein concentrations were much lower than in those cellsinoculated at higher N levels, as shown by both protein contentmeasurements [9] and sodium dodecylsulfate–polyacrylamidegel electrophoresis (S.-W. Sam and W.-X. Wang, unpublisheddata). Because Ag is considered as a class B metal [28] andsulfur-containing ligand is important in Ag transport in thediatoms, the higher nitrate or ammonium concentration mayhave facilitated the synthesis of these sulfur-containing li-gands, leading to increasing Ag uptake with increasing nutrientconcentrations.
Another mechanism, which still remains to be tested [29],is the control of Ag uptake rate by the growth rate of the cellsas influenced by macronutrient additions. Because different Pconcentrations and P starvation presumably did not greatlyaffect the ligand available for Ag transport, the higher uptakewith increasing nutrient concentration may not necessarily besolely caused by a change in binding sites. A difference ingrowth rate also may be responsible for the increasing Aguptake. Although Ag has no biological function and is notbiologically essential, increasing growth rate may result inincreasing requirements for other essential metals or elements,and, under such circumstances, Ag may be accidentally trans-ported by the phytoplankton. Accidental uptake of Ag has beenshown to be the main transport mechanism in phytoplankton[30], but AgCl complex (because of the high Cl2 concentra-tion) was also found to be the principal species of inorganicAg bioavailable to marine phytoplankton [31,32]. Another pos-sibility for the higher uptake with increasing growth rate maybe due to the smaller sizes of the cells, leading to an increase
in the surface-area-to-volume ratio and thus a higher uptakepotential.
Whether or not Ag concentration in diatoms will increaseafter nutrient enrichments depends on the relative change ofgrowth rate and uptake rate. Silver concentration in diatomsunder steady-state conditions can be calculated as the uptakerate constant divided by the Ag efflux rate constant plus thegrowth rate constant. Assuming that the efflux rate constantis much smaller than the growth rate constant, the Ag con-centration can be simply calculated as the uptake rate constantdivided by the growth rate constant. Given the dramatic in-crease in the Ag uptake rate with increasing nitrate or am-monium concentration, it is likely that the Ag concentrationin the diatoms under steady-state conditions will increase withincreasing nutrient levels. Such changes in Ag concentrationsin phytoplankton will have important implications for Ag tro-phic transfer to the higher trophic levels (see below).
Silver transfer from diatoms to copepods
The Ag AEs in A. spinicauda measured in this study werein the range of 3 to 22%, consistent with several previousmeasurements in other species of copepods. Reinfelder andFisher [1] found an Ag AE of 15% in the copepod Acartiatonsa feeding on the diatom T. pseudonana. The AEs foranother copepod, Temora longicornis, feeding on two diatomdiets (T. pseudonana and Thalassiosira weissflogii) and nat-ural seston were 8 to 19% [4]. These AEs were generallyamong the lowest quantified for different metals [33], indi-cating a lower bioavailability of Ag from ingested food sourcesto marine herbivores. However, Wang and Fisher [4] showed

Silver uptake by marine plankton Environ. Toxicol. Chem. 23, 2004 689
that dietary uptake contributed appreciably to the overall Agaccumulation in marine copepods (largely because of the par-ticle reactivity in the food particles), although the relative sig-nificance of dietary uptake was generally smaller than theaqueous uptake. Our study demonstrated that the assimilationof Ag is a rather dynamic process and many factors can con-tribute to the variability of Ag AE in the copepods.
Our study shows that a few important processes are re-sponsible for Ag assimilation, including Ag distribution indifferent subcellular fractions (e.g., cytoplasmic), Ag concen-tration factor in the diatoms and its particle reactivity (e.g.,tendency to bind with particles), and its gut passage time. Allthese factors interact to result in a variable Ag assimilation inthe copepods as influenced by various environmental factorssuch as food concentration and nutritional stoichiometry ofthe food particles. All these factors are interrelated and thusit is difficult to conclude that a single process is wholly re-sponsible for Ag assimilation. Values of Ag AE as low as 3%were found in some treatments, suggesting that Ag can beessentially inert to the copepods under certain circumstances.Therefore, understanding the processes controlling Ag assim-ilation presents an important step in predicting Ag transferdynamics in marine ecosystems.
Reinfelder and Fisher [1] first found a linear 1:1 relationshipbetween metal AE in a marine copepod A. tonsa and the dis-tribution in the diatom’s cytoplasm when different metals wereconsidered together. Demonstration of a significant relation-ship has been shown for a specific metal, including Fe [34]and Cd [6,9]. Our study presents the experimental evidencethat the Ag AE also is dependent on its subcellular partitioningin the algal cytoplasm, but such a relationship was not 1:1. Inour study, we found that only a small fraction of Ag wasdistributed in the diatom’s cytoplasm (maximum of 7%, pre-sumably due to the short radiolabeling time, ,1 d), whereasthe AE could be higher than 30%. This experimental resultstrongly suggests that the noncytoplasmic component of thediatoms also can be assimilated by the copepods. This is con-sistent with our recent finding that metals (Cd, Se, and Zn)bound with the noncytoplasmic fraction can be available tomarine copepods [7]. In that study, the AEs of Cd and Znbound with the noncytoplasmic fraction (i.e., the cell wall)was in fact assimilated at a comparable efficiency as that fromthe living phytoplankton particles. In the estuarine amphipodLeptocheirus plumulosus, Ag and Cd assimilation efficienciesfrom phytoplankton were low and not related to the proportionof metal associated with cell cytosol or cytoplasm [35].
Given the significant relationships between the Ag AE andthe Ag DCF and Ag partitioning in the particles, our studystrongly implies that Ag assimilation in marine copepods islikely controlled by the particle reactivity of Ag in diatoms,in addition to Ag subcellular distribution. This is the first ex-perimental evidence suggesting a significant relationship be-tween metal AE and metal DCF or metal partitioning in thefood particles. A much higher particle reactivity results instronger binding with the diatoms, and less Ag will be ableto desorb from the particles, resulting in a lower AE. All theselines of evidence (DCF and partitioning in the particles) pointto the potential control of metal desorption within the cope-pod’s gut (or solubilization) in metal assimilation. Among themany metals that have been examined thus far, desorption hasbeen shown to account for the assimilation of Cd in severalinvertebrates such as mussels [36] and sipunculans [37]. Gag-non and Fisher [38] also found a significant correlation be-
tween Ag assimilation in mussels from ingested sediments andthe desorption of Ag from the sediment particles at pH 5, nearthe gut pH of the mussels. Particle reactivity and desorptionwere previously implied to play a critical role on the transferof Ag to marine herbivores in several studies [39,40]. Forexample, Connell et al. [39] demonstrated that Ag tightlybound to the diatoms and the feeding and digestion by inver-tebrates are not likely to dislodge Ag from biotic particles;thus, the food-borne transfer of Ag within the aquatic foodchains is unlikely to occur in estuarine invertebrates (e.g., thegrass shrimp Palaemonetes pugio).
The higher assimilation of Ag at the lower nutritional con-ditions of the cells was rather unexpected from our study. Ourprevious studies on Cd and Zn generally indicated that theirassimilation increased with increasing the N status of the di-atom cells, as a result of increased partitioning of these metalsin the diatom cytoplasm [9,10]. In this study, the cytoplasmicfraction of Ag was higher at lower nutrient concentrations.Furthermore, diatoms grown under nutrient-enriched condi-tions were able to bind more strongly with Ag than cells grownat lower nutrient concentrations and their Ag concentrationfactors were accordingly higher. This experimental evidenceindicated that the relatively higher Ag AE with decreasingnutrient concentrations was a result of the weaker binding ofAg with the diatoms, leading to an increasing potential of Agdesorption or solubilization, and the increased partitioning ofAg in the diatom’s cytoplasm.
The increasing Ag AE with decreasing diatom food con-centration is consistent with our recent study on another co-pepod, Calanus sinicus [6]. The Ag AE (21%) measured atthe lowest food concentration was indeed the highest foundamong the different experiments. Because the subcellular dis-tribution of Ag in the diatoms was comparable among thedifferent food concentrations, the physiological process waspresumably more important in contributing to the increasingAg AE with decreasing diatom cell concentration. Our studyindicated that such a high AE was related to the longer Aggut passage time and presumably also to the longer retentionof food particles in the gut. These experimental results at leastsuggest that digestion in copepod’s gut is flexible. However,given the significant difference in Ag partitioning in the foodparticles at different concentrations during the radioactivefeeding period, particle reactivity and thus desorption also maycontribute to the difference in Ag AE among the different foodconcentration treatments. Clearly, digestive physiology mayinteract closely with the geochemical processes in the foodparticles and contribute to the variability of Ag assimilationin marine copepods.
The Ag concentrations in copepods under steady-state con-ditions in response to nutrient enrichment likely will be de-pendent on the different kinetic parameters as influenced bynutrients. Metal concentration in copepods as a result of trophictransfer only can be calculated by the following equation:
(AE· IR·C )fC 5ke
where C is the metal concentration in the copepods understeady-state conditions (mg/g), AE is the metal assimilationefficiency from the ingested particles, IR is the ingestion rateof the copepods (% body wt/d), Cf is the metal concentrationin ingested particles (mg/g), and ke is the efflux rate constant(d21). Assuming that the efflux rate constant was independentof the nutritional conditions [9,10], any change in C will be

690 Environ. Toxicol. Chem. 23, 2004 Y. Xu and W.-X. Wang
determined by the relative changes of AE, IR, and Cf as in-fluenced by the macronutrients. Under nutrient-enrichmentconditions, Cf and IR likely will increase, but the change inAg AE is much more complicated because it can be influencedby both food concentrations and nutrient conditions. Clearly,prediction of Ag trophic transfer and its concentration in co-pepods under eutrophication conditions is not straightforward,and Ag AE is the key parameter affecting its trophic transferof Ag. Field experiments are required to test such influences,which can be very specific under different environmental con-ditions.
Acknowledgement—We thank the two anonymous reviewers for theircomments on this work. Support from the Hong Kong Research GrantCouncil (NpHKUST603/01M, HKUST6118/01M) to W.-X. Wang isgratefully acknowledged.
REFERENCES
1. Reinfelder JR, Fisher NS. 1991. The assimilation of elementsingested by marine copepods. Science 251:794–796.
2. Fisher NS, Breslin VT, Levandowsky M. 1995. Accumulation ofsilver and lead in estuarine microzooplankton. Mar Ecol ProgSer 116:207–215.
3. Wang WX, Reinfelder JR, Lee BG, Fisher NS. 1996. Assimilationand regeneration of trace elements by marine copepods. LimnolOceanogr 41:70–81.
4. Wang WX, Fisher NS. 1998. Accumulation of trace elements ina marine copepod. Limnol Oceanogr 43:273–283.
5. Connell DB, Sanders JG. 1999. Variation in cadmium uptake byestuarine phytoplankton and transfer to the copepod Eurytemoraaffinis. Mar Biol (Berl) 133:259–265.
6. Xu Y, Wang WX. 2001. Individual responses of metal assimilationand regeneration by copepod Calanus sinicus to changes in foodquantity. Mar Ecol Prog Ser 218:227–238.
7. Xu Y, Wang WX. 2002. The assimilation of detritus-bound metalsby marine copepod Acartia spinicauda. Limnol Oceanogr 47:604–610.
8. Wang WX. 2002. Interactions of trace metals and different marinefood chains. Mar Ecol Prog Ser 243:295–309.
9. Wang WX, Dei RCH, Xu Y. 2001. Cadmium uptake and trophictransfer in coastal plankton under contrasting nitrogen regimes.Mar Ecol Prog Ser 211:293–298.
10. Wang WX, Dei RCH, Xu Y. 2001. The responses of Zn assim-ilation by coastal plankton to macronutrients. Limnol Oceanogr46:1524–1534.
11. Cullen JT, Lane TW, Morel FMM, Sherrell RM. 1999. Modulationof cadmium uptake in phytoplankton by seawater CO2 concen-tration. Nature (Lond) 402:165–167.
12. Sanders JG, Cibik SJ. 1988. Response of Chesapeake Bay phy-toplankton communities to low levels of toxic substances. MarPollut Bull 19:439–444.
13. Sanders JG, Abbe GR, Riedel GF. 1990. Silver uptake and sub-sequent effects on growth and species composition in an estuarinecommunity. Sci Total Environ 97/98:761–769.
14. Eilser R. 1996. Silver hazards to fish, wildlife and invertebrates:A synoptic review. Contaminant Hazard Reviews Report 32. Na-tional Biological Survey, U.S. Department of the Interior, Wash-ington, DC.
15. Ratte HT. 1999. Bioaccumulation and toxicity of silver com-pounds: A review. Environ Toxicol Chem 18:89–108.
16. Luoma SN, Ho YB, Bryan GW. 1995. Fate, bioavailability andtoxicity of silver in estuarine environments. Mar Pollut Bull 31:44–54.
17. Wood CM, LaPoint TW, Armstrong DE, Birge WJ, Brauner CJ,Brix KV, Call DJ, Crecelius EA, Davies PH, Gorsuch JW, Hogs-trand C, Mahony JD, McGeer JC, O’Connor TP. 2002. Biologicaleffects of silver. In Andren AW, Bober TW, eds, Silver in theEnvironment: Transport, Fate, and Effects. SETAC, Pensacola,FL, USA, pp 27–63.
18. Hornberger MI, Luoma SN, Cain DJ, Parchaso F, Brown CL,
Bouse RM, Wellise C, Thompson JK. 2000. Linkage of bioac-cumulation and biological effects to changes in pollutant loadsin South San Francisco Bay. Environ Sci Technol 34:2401–2409.
19. Hook SE, Fisher NS. 2001. Sublethal effects of silver in zoo-plankton: Importance of exposure pathways and implications fortoxicity testing. Environ Toxicol Chem 20:568–574.
20. Fisher NS, Wang WX. 1998. Trophic transfer of silver to marineherbivores: A review of recent studies. Environ Toxicol Chem17:562–571.
21. Luoma SN, Hogstrand C, Bell RA, Bielmyer GK, Galvez F,LeBlanc GA, Lee BG, Purcell TW, Santore RC, Santshi PH, ShawJR. 2002. Biological processes. In Andren AW, Bober TW, eds,Silver in the Environment: Transport, Fate, and Effects. SETAC,Pensacola, FL, USA, pp 65–95.
22. Sanudo-Wilhelmy S, Flegal R. 1992. Anthropogenic silver in theSouthern California Bight: a new tracer of sewage in coastalwaters. Environ Sci Technol 26:2147–2151.
23. Smith GJ, Flegal AR. 1993. Silver in San Francisco Bay estuarinewaters. Estuaries 16:547–558.
24. Guillard RRL, Ryther JH. 1962. Studies on marine planktonicdiatoms. I. Cyclotella nana Hustedt and Detonula confervacea(Cleve) Gran. Can J Microbiol 8:229–239.
25. Fortin C, Campbell PGC. 2000. Silver uptake by the green alga,Chlamydomonas reinhardtii, in relation to chemical speciation:Influence of chloride. Environ Toxicol Chem 19:2769–2778.
26. Fortin C, Campbell PGC. 2001. Thiosulfate enhances silver up-take by a green alga: Role of anion transporters in metal uptake.Environ Sci Technol 35:2214–2218.
27. Hook SE, Fisher NS. 2002. Relating the reproductive toxicity offive ingested metals in calanoid copepods with sulfur affinity.Mar Environ Res 53:161–174.
28. Nieboer E, Richardson DH. 1980. The replacement of the non-descript term ‘heavy metals’ by a biological and chemically sig-nificant classification of metal ions. Environ Pollut Ser B ChemPhys 1:3–26.
29. Wang WX, Dei RCH. 2001. Effects of major nutrient additionson metal uptake in phytoplankton. Environ Pollut 111:233–240.
30. Campbell PGC, Errecalde O, Fortin C, Hiriart-Baer VP, VigneaultB. 2002. Metal bioavailability to phytoplankton—Applicabilityof the biotic ligand model. Comp Biochem Physiol C 133:189–206.
31. Engel DW, Sunda WG, Fowler BA. 1981. Factors affecting tracemetal uptake and toxicity to estuarine organisms. 1. Environ-mental parameters. In Vernberg JF, Calabrese A, Thurberg FP,Vernberg WB, eds, Biological Monitoring of Marine Pollutants.Academic, New York, NY, USA, pp 127–144.
32. Reinfelder JR, Chang SI. 1999. Speciation and microalgal bio-availability of inorganic silver. Environ Sci Technol 33:1860–1863.
33. Wang WX, Fisher NS. 1999. Assimilation efficiencies of chemicalcontaminants in aquatic invertebrates: A synthesis. Environ Tox-icol Chem 18:2034–2045.
34. Hutchins DA, Wang WX, Fisher NS. 1995. Copepods grazingand the biogeochemical fate of diatom iron. Limnol Oceanogr40:989–994.
35. Schlekat CE, Decho AW, Chandler GT. 2000. Bioavailability ofparticle-associated silver, cadmium, and zinc to the estuarine am-phipod Leptocheirus plumulosus through dietary ingestion. Lim-nol Oceanogr 45:11–21.
36. Wang WX, Fisher NS. 1996. Assimilation of trace elements bythe mussel, Mytilus edulis: Effects of diatom chemical compo-sition. Mar Biol (Berl) 125:715–724.
37. Yan QL, Wang WX. 2002. Metal exposure and bioavailability toa marine deposit-feeding sipuncula Sipunculus nudus. EnvironSci Technol 36:40–47.
38. Gagnon C, Fisher NS. 1997. The bioavailability of sediment-bound Cd, Co, and Ag to the mussel, Mytilus edulis. Can J FishAquat Sci 54:147–156.
39. Connell DB, Sanders JG, Riedel GF, Abbe GR. 1991. Pathwaysof silver uptake and trophic transfer in estuarine organisms. En-viron Sci Technol 25:921–924.
40. Abbe GR, Sanders JG. 1990. Pathways of silver uptake and ac-cumulation by the American oyster (Crassostrea virginica) inChesapeake Bay. Estuarine Coast Shelf Sci 31:113–123.





























