Embed Size (px)
Citation preview

trees often overtop the adjacent canopy. The crowns of these trees are dominated by much larger limbs than are found in cylindrical crowns. This spherical form of crown seems more characteristic of old-growth stands in the Coast Ranges than of those on the west side of the Cas- cade Range and may reflect crowns developed in a denser stand (see footnote 7).
Few old-growth Douglas-firs have vertical trunks. The lower trunk leans away from the hillside but be- comes nearly vertical where it ex- tends above the surrounding canopy. Trunks on level sites appear to slope almost at random. Even a slight inclination of a trunk results in an important differentiation of habitat on its two sides. The upper
side gets almost all the moisture, both from direct precip.itation and from stem flow and throughfall. Con- sequently, it is colonized by epi- phytic plants (plants that grow on other plants) with relatively high moisture requirements, chiefly mosses. The lower side is a “desert” occupied by scattered col- onies of lichens (Pike et al. 1975) that form a crust over the bark sur- face. The bark on the wet upper side is soft and easily eroded, sometimes appearing to be held in place by its mantle of mosses and lichens, whereas the bark on the lower side is hard and deeply furrowed, indicat- ing that it remains in place for longer periods.
Old specimens of other tree species can play a role comparable to that of Douglas-fir to at least some degree, although none have been as thoroughly studied (fig. 13). Sitka spruce attains comparable sizes in coastal regions; irregular crown sys- tems and heavy, epiphyte-laden branch systems are characteristic of
older specimens. Noble fir and west- ern white pine are subalpine species with some, but not all, of the distinc- tive characteristics of Douglas-fir, as is sugar pine in southwestern Oregon. The so-called cedars-west- ern redcedar, Alaska-cedar, Port- Orford-cedar, and incense- cedar-are capable of attaining sizes and fulfilling roles comparable to Douglas-fir in their respective types. These species have the addl- tional advantage of fostering im- proved soil conditions through their base-rich litter. The major climax species-western hemlock, Pacific silver fir, and grand fir-appear, on the other hand, to lack the ability to completely fulfill the ecological roles of these long-lived pioneers.
23
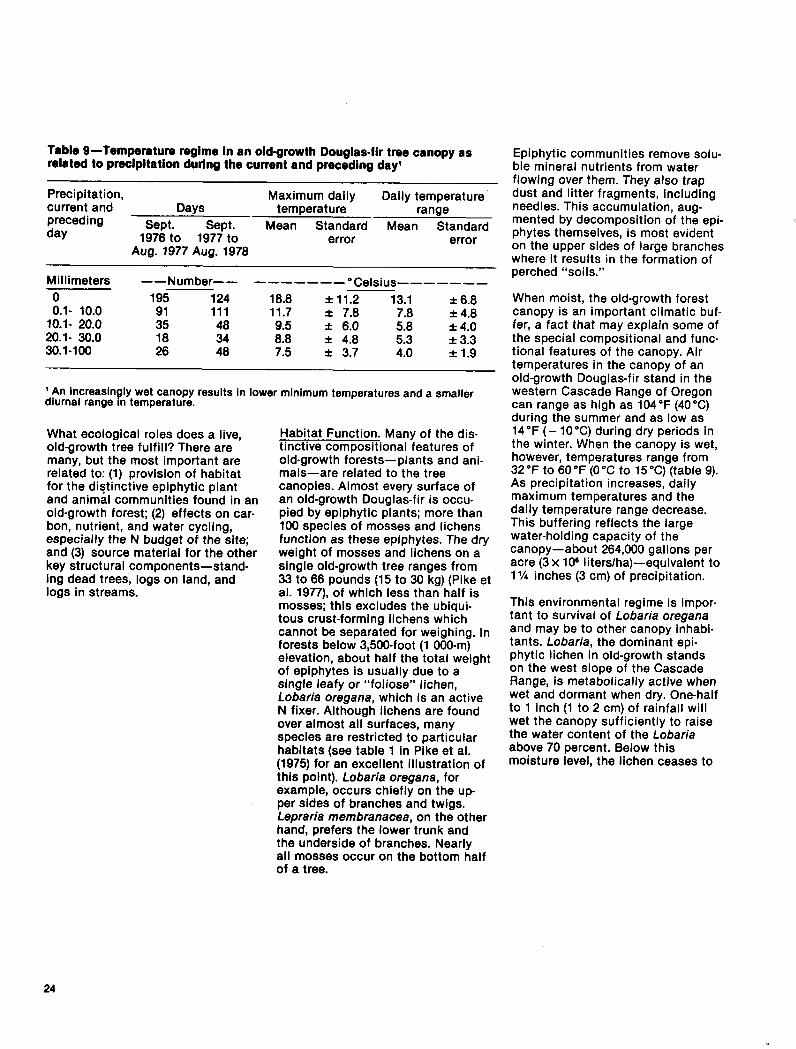
Table Q-Temperature regime in an old-growth Douglas-fir tree canopy as related to precipitation during the current and preceding day’
Precipitation, current and
Maximum daily Days
Daily temperature temperature
preceding range
day Sept. Sept. Mean Standard Mean Standard
1976 to 1977 to error error Aug. 1977 Aug. 197%
Millimeters
0 0.1. 10.0
10.1. 20.0 20.1. 30.0 30.1-100
--Number-- ------_ “Celsius-------
195 124 18.8 f 11.2 13.1 f 6.8 91 111 11.7 f 7.8 f 4.8 35 48 9.5 f 6.0 i-s8 18 34 8.8 f 4.8 5:3
f 4.0 f 3.3
26 48 7.5 f 3.7 4.0 f 1.9
’ An increasin ly wet canopy results in lower minimum temperatures and a smaller diurnal range n temperature. B
What ecological roles does a live, old-growth tree fulfill? There are many, but the most important are related to: (1) provision of habitat for the distinctive epiphytic plant and animal communities found in an old-growth forest; (2) effects on car- bon, nutrient, and water cycling, especially the N budget of the site; and (3) source material for the other key structural components-stand- ing dead trees, logs on land, and logs in streams.
Habitat Function. Many of the dis- tinctive compositional features of old-growth forests-plants and ani- mals-are related to the tree canopies. Almost every surface of an old-growth Douglas-fir is occu- pied by epiphytic plants; more than 100 species of mosses and lichens function as these epiphytes. The dry weight of mosses and lichens on a single old-growth tree ranges from 33 to 66 pounds (15 to 30 kg) (Pike et al. 1977), of which less than half is mosses; this excludes the ubiqui- tous crust-forming lichens which cannot be separated for weighing. In forests below 3,500-foot (1 000-m) elevation, about half the total weight of epiphytes is usually due to a single leafy or “foliose” lichen, Lobaria oregana, which is an active N fixer. Although lichens are found over almost all surfaces, many species are restricted to particular habitats (see table 1 in Pike et al. (1975) for an excellent illustration of this point). Lobaria oregana, for example, occurs chiefly on the up- per sides of branches and twigs. Lepraria membranacea, on the other hand, prefers the lower trunk and the underside of branches. Nearly all mosses occur on the bottom half of a tree.
Epiphytic communities remove solu- ble mineral nutrients from water flowing over them. They also trap dust and litter fragments, including needles. This accumulation, aug- mented by decomposition of the epi- phytes themselves, is most evident on the upper sides of large branches where it results in the formation of perched “soils.”
When moist, the old-growth forest canopy is an important climatic buf- fer, a fact that may explain some of the special compositional and func- tional features of the canopy. Air temperatures in the canopy of an old-growth Douglas-fir stand in the western Cascade Range of Oregon can range as high as 104°F (40°C) during the summer and as low as 14°F (- 10°C) during dry periods in the winter. When the canopy is wet, however, temperatures range from 32 “F to 60 “F (0 “C to 15 “C) (table 9). As precipitation increases, daily maximum temperatures and the daily temperature range decrease. This buffering reflects the large water-holding capacity of the canopy-about 264,000 gallons per acre (3 x lOa liters/ha)-equivalent to 1% inches (3 cm) of precipitation.
This environmental regime is impor- tant to survival of Lobaria oregana and may be to other canopy inhabi- tants. Lobaria, the dominant epi- phytic lichen in old-growth stands on the west slope of the Cascade Range, is metabolically active when wet and dormant when dry. One-half to 1 inch (1 to 2 cm) of rainfall will wet the canopy sufficiently to raise the water content of the Lobaria above 70 percent. Below this moisture level, the lichen ceases to
24
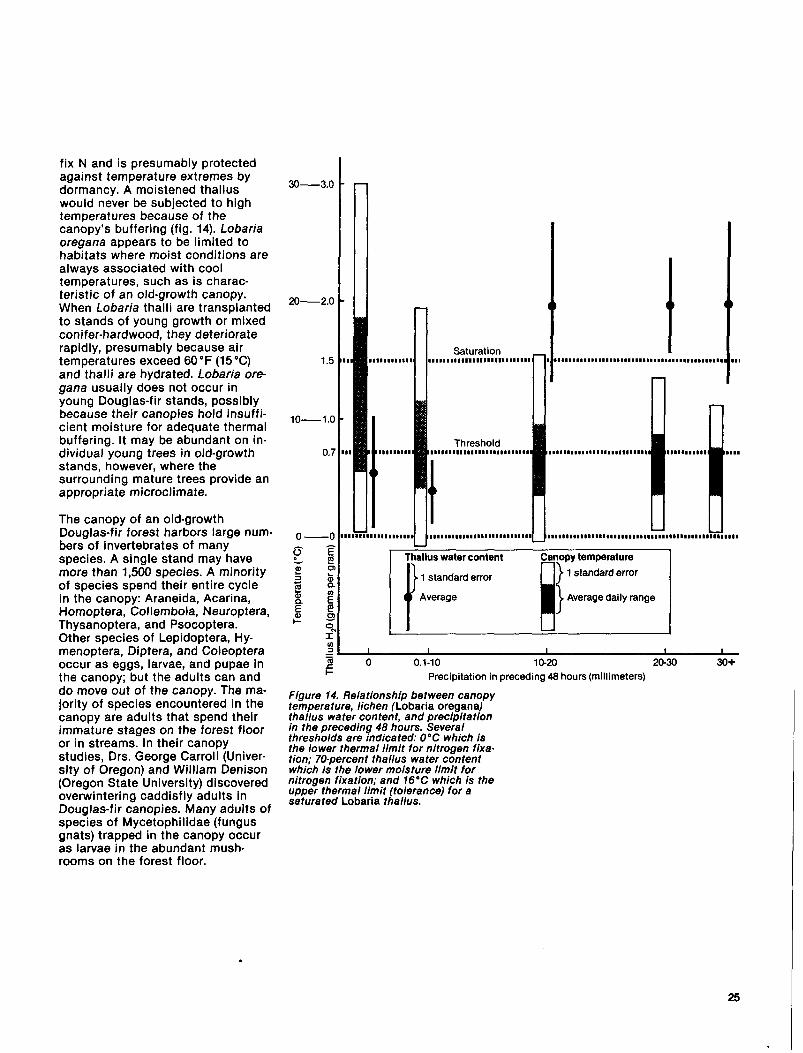
fix N and is presumably protected against temperature extremes by dormancy. A moistened thallus would never be subjected to high temperatures because of the canopy’s buffering (fig. 14). Lobaria oregana appears to be limited to habitats where moist conditions are always associated with cool temperatures, such as is charac- teristic of an old-growth canopy. When Lobaria thalli are transplanted to stands of young growth or mixed conifer-hardwood, they deteriorate rapidly, presumably because air temperatures exceed 60 ‘F (15 “C) and thalli are hydrated. Lobaria ore- gana usually does not occur in young Douglas-fir stands, possibly because their canopies hold insuffi- cient moisture for adequate thermal buffering. It may be abundant on in- dividual young trees in old-growth stands, however, where the surrounding mature trees provide an appropriate microclimate.
The canopy of an old-growth Douglas-fir forest harbors large num- bers of invertebrates of many species. A single stand may have more than 1,500 species. A minority of species spend their entire cycle in the canopy: Araneida, Acarina, Homoptera, Collembola, Neuroptera, Thysanoptera, and Psocoptera. Other species of Lepidoptera, Hy- menoptera, Diptera, and Coleoptera occur as eggs, larvae, and pupae in the canopy; but the adults can and do move out of the canopy. The ma- jority of species encountered in the canopy are adults that spend their immature stages on the forest floor or in streams. In their canopy studies, Drs. George Carroll (Univer- sity of Oregon) and William Denison (Oregon State University) discovered overwintering caddisfly adults in Douglas-fir canopies. Many adults of species of Mycetophilidae (fungus gnats) trapped in the canopy occur as larvae in the abundant mush- rooms on the forest floor.
30-3.0
i
Thallus water content
/;;;;;;;d error [/EInge
I I I I I
0 0.1-10 1 O-20 20-30 30+ Precipitation in preceding 48 hours (millimeters)
Figure 14. Relationship between canopy temperature, lichen (Lobaria oregana) thallus water content, and precipitation in the preceding 48 hours. Several thresholds are indicated: 0°C which is the lower thermal limit for nitrogen fixa- tion; 70-percent thallus water content which is the lower moisture limit for nitrogen fixation; and 76’C which is the upper thermal limit (tolerance) for a saturated Lobaria thaflus.
.

Although primary consumers (in-sects-such as sawflies, scales, oraphids-which feed on foliage orbeetles which feed on wood) do oc-cur in the canopy, they are not abun-dant. The most abundant arthropodsare predaceous spiders, whichbelong to families such as Salti-cidae and Thomisidae. The largenumbers of flies found in thecanopy probably provide food for thespiders. Other arthropods feed ondebris or on bacteria and fungi onsurfaces of the canopy or are preda-tors of other invertebrates. Duringone sampling period, invertebrateswashed from foliage samplesincluded:
Food sourceNeedles
Invertebrates1 species of scale1 species of mealybug1 species ofLepidoptera
Bacteria and 6 species of mitesfungi living 4 species of flieson needles (as larvae)
5 species ofCollembola
Other 2 species of mitesinvertebrates 2 species of spiders
The canopy of an old-growth forestprovides several insect habitats,both vertically and horizontally.Some species are found in the uppercanopy, others in the lower; somespecies occur on major limbs andothers among twigs and foliage.
Several vertebrates depend heavilyon the old-growth canopy as sitesfor nesting, feeding, and protection.Well-known examples are the north-ern spotted owl, northern flyingsquirrel, and red tree vole. The volemay live for many generations in thesame tree. The role that the largebranch systems and organic accum-ulations play in providing suitablehabitat should not be overlooked.
Cycling Function. Old-growth treesare one of the primary sites forphotosynthesis, or production of thefood base, on which the rest of thesystem depends. In this sense, theyare the same as younger trees, ex-cept that each tree represents alarge accumulation of organic mate-rial and nutrients (a “sink” in theshort run and a “storehouse” in thelong run) as well as a large photo-synthetic factory. A single old-growth tree can have over 60 millionindividual needles with a cumulativeweight of 440 pounds (200 kg) and asurface area of 30,000 square feet(2 800 m2) (Pike et al. 1977). Totalleaf areas in old-growth stands areprobably not much different fromthose in younger stands, but theleaves are concentrated on fewer in-dividuals. Fluctuations in productionand live biomass strongly reflectmortality of these large dominanttrees, which are both factory andstorehouse, and the rate at whichother trees occupy the vacatedspace.
A distinctive and unusual functionalrole of an old-growth tree is its con-tribution to the nitrogen economy oflow-elevation to midelevation sites.Lichens that inhabit the canopy fixsignificant amounts of N which ulti-mately become available to thewhole forest through leaching, litterfall, and decomposition. Estimatesof fixed N range from 2.5 to 4.5pounds per acre (3 to 5 kg/ha) peryear. Most of the fixation is accom-plished by Lobaria oregana,although several other large folioselichens, such as L. pulmonaria,Pseudocyphellaria rainierensis, andPeltigera aphthosa, are also azoto-desmic8 and, therefore, capable offixing atmospheric N. Lobaria ore-gana accounts for half the total epi-phytic biomass in the western Ore-gon Douglas-fir stands that havebeen studied. Lobaria and mostN-fixing epiphytes are not commonin young-growth stands, and thismay be related to the microclimateof the old-growth forest canopy. Sig-nificant epiphytic inputs of N are,therefore, largely confined to oldgrowth. Nitrogen-fixing bacteria onDouglas-fir foliage have not beenfound in the Pacific Northwest, eventhough they have been reported inEurope.
8 Azotodesmic lichens contain a blue-green alga, either as a primary plantsymbiont or a secondary one, andtherefore are capable of fixing N.Nonazotodesmic lichens contain a greenalga as the sole algal symbiont and arenot capable of fixing N.

Standing Dead Trees or Snags. In any old-growth stand there are sub- stantial numbers of standing dead trees or snags (fig. 15). Indeed, snags were the first dead compo- nent of natural forests of which foresters were made aware-initially because of the fire and safety hazard they represent and, more recently, because of their value to wildlife (Bull and Meslow 1977, Bull 1978, Thomas et al. 1979a, Mannan et al. 1980). Some representative data for old-growth stands are pro- vided in table 10. The only compre- hensive study on dynamics of snags is by Cline et al. (1980), who studied 30 stands from 5 to 440 Years old in the Coas Densitie! stand ag increase 72 cm) bl 200; largl Cline et ( ties of SI d.b.h. at 7.3 per a and 200 ] These va (model) f acre (2311 are substantially below the densities in table 10; all six of the old-growth stands of Cline et al. (1980) are located in the Coast Ranges.
Figure 15. Large numbers of standing dead trees or snags are characteristic of old-growth forests. A. The volume and numbers of standing dead trees may not be apparent to the casual observer in this 250-year-old Douglas-fir stand in the Bagby Research Natural Area, Mount Hood National Forest; dead stems are marked with an X. ~3. Heavily decom- posed snags in old-growth Douglas- fir-western hemlock stand.
27
c
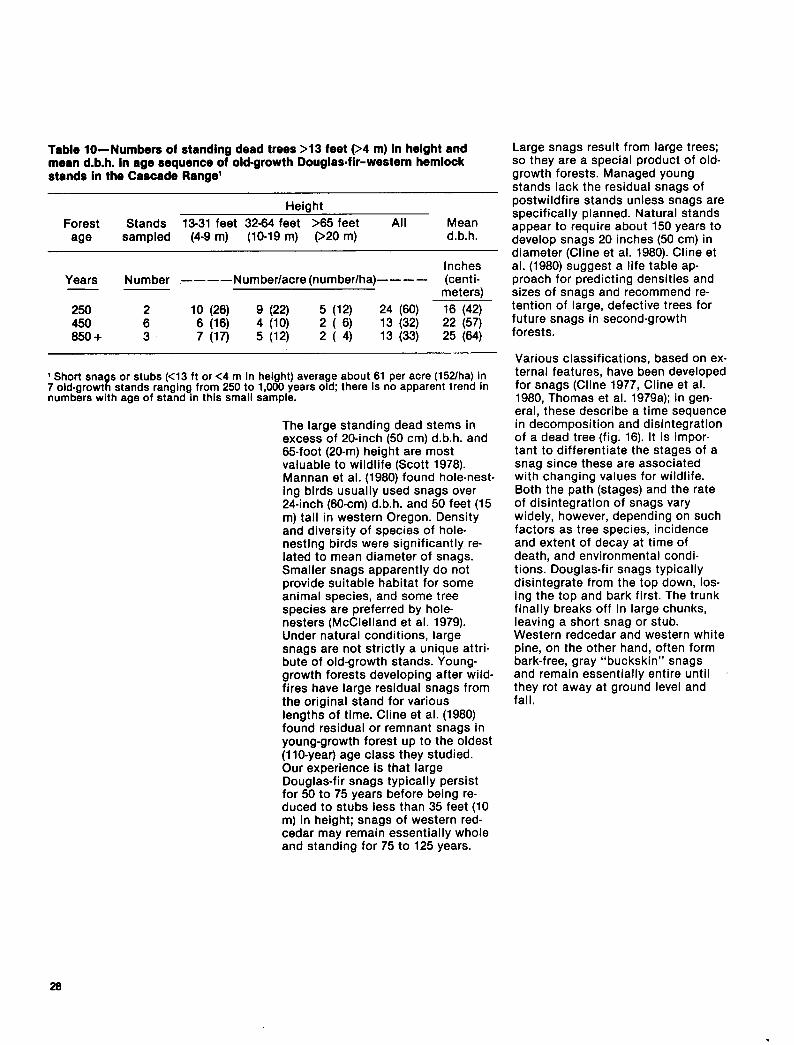
Table lo-Numbers of standing dead trees >13 feet 94 m) in height and mean d.b.h. in age sequence of old-growth Douglasfir-western hemlock stands in the Cascade Range’
Height
Forest Stands 13-31 feet 32-64 feet >65 feet All Mean age sampled (4-9 m) (lo-19 m) 920 m) d.b.h.
Inches Years Number ----Number/acre(number/ha)---- (centi- -- meters) 250 2 10 (26) 9 (22) 5 (12) 24 (60) 16 (42) 450 i 6 (16) 4 (10) 2 ( ‘3) 13 (32) 22 (57) 650 + 7 (17) 5 (12) 2 ( 4) 13 (33) 25 (64)
( Short snags or stubs (<13 ft or <4 m in height) average about 61 per acre (1Wha) in 7 old-growth stands ranging from 250 to 1,000 years old; there is no apparent trend in numbers with age of stand in this small sample.
The large standing dead stems in excess of 20-inch (50 cm) d.b.h. and 65-foot (20-m) height are most valuable to wildlife (Scott 1976). Mannan et al. (1980) found hole-nest- ing birds usually used snags over 24-inch (so-cm) d.b.h. and 50 feet (15 m) tall in western Oregon. Density and diversity of species of hole- nesting birds were significantly re- lated to mean diameter of snags. Smaller snags apparently do not provide suitable habitat for some animal species, and some tree species are preferred by hole- nesters (McClelland et al. 1979). Under natural conditions, large snags are not strictly a unique attri- bute of old-growth stands. Young- growth forests developing after wild- fires have large residual snags from the original stand for various lengths of time. Cline et al. (1980) found residual or remnant snags in young-growth forest up to the oldest (IlO-year) age class they studied. Our experience is that large Douglas-fir snags typically persist for 50 to 75 years before being re- duced to stubs less than 35 feet (10 m) in height; snags of western red- cedar may remain essentially whole and standing for 75 to 125 years.
Large snags result from large trees; so they are a special product of old- growth forests. Managed young stands lack the residual snags of postwildfire stands unless snags are specifically planned. Natural stands appear to require about 150 years to develop snags 20 inches (50 cm) in diameter (Cline et al. 1980). Cline et al. (1980) suggest a life table ap- proach for predicting densities and sizes of snags and recommend re- tention of large, defective trees for future snags in second-growth forests.
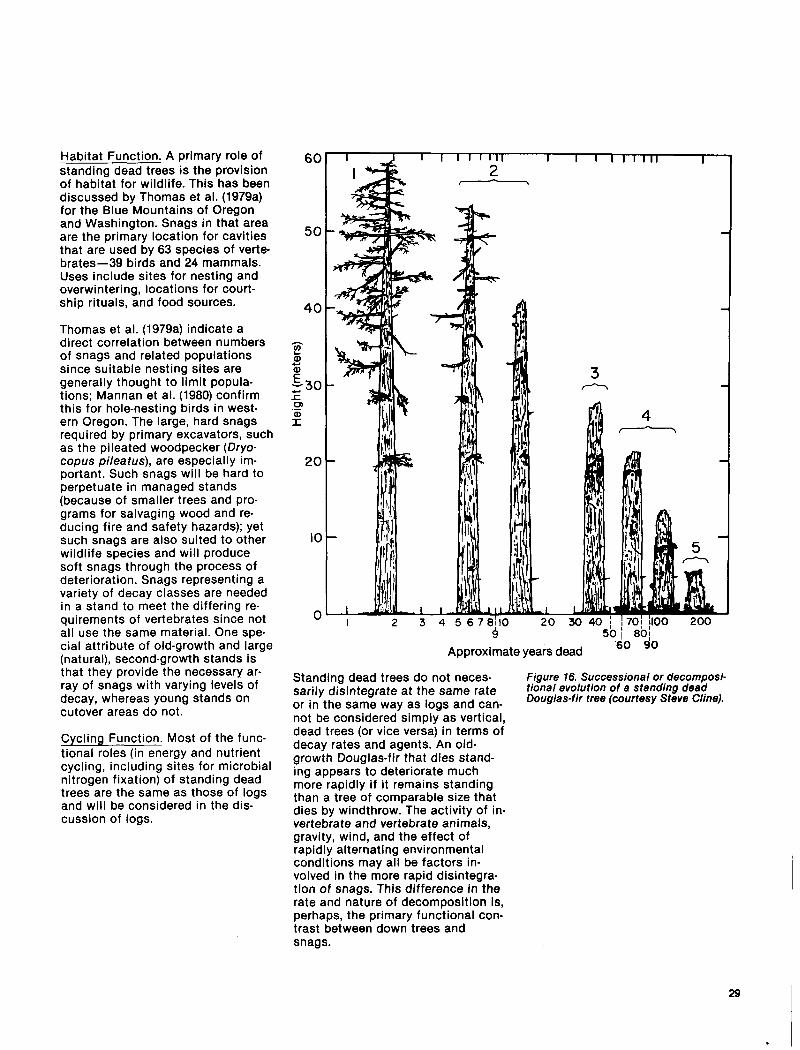
Various classifications, based on ex- ternal features, have been developed for snags (Cline 1977, Cline et al. 1980, Thomas et al. 1979a); in gen- eral, these describe a time sequence in decomposition and disintegration of a dead tree (fig. 16). It is impor- tant to differentiate the stages of a snag since these are associated with changing values for wildlife. Both the path (stages) and the rate of disintegration of snags vary widely, however, depending on such factors as tree species, incidence and extent of decay at time of death, and environmental condi- tions. Douglas-fir snags typically disintegrate from the top down, los- ing the top and bark first. The trunk finally breaks off in large chunks, leaving a short snag or stub. Western redcedar and western white pine, on the other hand, often form bark-free, gray “buckskin” snags and remain essentially entire until they rot away at ground level and fall.
28
Habitat Function. A primary role of standing dead trees is the provision of habitat for wildlife. This has been discussed by Thomas et al. (1979a) for the Blue Mountains of Oregon and Washington. Snags in that area are the primary location for cavities that are used by 63 species of verte- brates-39 birds and 24 mammals. Uses include sites for nesting and overwintering, locations for court- ship rituals, and food sources.
Thomas et al. (1979a) indicate a direct correlation between numbers of snags and related populations since suitable nesting sites are generally thought to limit popula- tions; Mannan et al. (1980) confirm this for hole-nesting birds in west- ern Oregon. The large, hard snags required by primary excavators, such as the pileated woodpecker (Dryo- copus pileatus), are especially im- portant. Such snags will be hard to perpetuate in managed stands (because of smaller trees and pro- grams for salvaging wood and re- ducing fire and safety hazards); yet such snags are also suited to other wildlife species and will produce soft snags through the process of deterioration. Snags representing a variety of decay classes are needed in a stand to meet the differing re- quirements of vertebrates since not all use the same material. One spe- cial attribute of old-growth and large (natural), second-growth stands is that they provide the necessary ar- ray of snags with varying levels of decay, whereas young stands on cutover areas do not.
Cycling Function. Most of the func- tional roles (in energy and nutrient cycling, including sites for microbial nitrogen fixation) of standing dead trees are the same as those of logs and will be considered in the dis- cussion of logs.
60
50
40
6 $ ‘;
g30 E 0 .- I”
20
IO
0
I I I 111111~ I I 1 IIIIII I
5’0 / 86:
Approximate years dead ‘60 d0
Standing dead trees do not neces- Figure 16. Successional or decomposi- sarily disintegrate at the same rate tional evolution of a stending deed
or in the same way as logs and can- Douglas-fir tree (courtesy Steve Cline).
not be considered simply as vertical, dead trees (or vice versa) in terms of decay rates and agents. An old- growth Douglas-fir that dies stand- ing appears to deteriorate much more rapidly if it remains standing than a tree of comparable size that dies by windthrow. The activity of in- vertebrate and vertebrate animals, gravity, wind, and the effect of rapidly alternating environmental conditions may all be factors in- volved in the more rapid disintegra- tion of snags. This difference in the rate and nature of decomposition is, perhaps, the primary functional con- trast between down trees and snags.

Figure 17. Large masses of logs can be a dominant feature of old-growth forests, as illustrated in these stands with near- maximal accumulations: A. Midelevation stand of old-growth Douglas-fir in the western Cascade Range of Oregon. 8. Old-growth stand of noble fir near Mount St. Helens, Washington.
.
30
i

Logs on Land. Logs, also describa- ble as down dead trees or coarse woody debris, are nearly as conspic- uous as the large, live trees. Large masses of logs can be the dominant feature of old-growth forests (fig. 17), and, in numbers, volume, and weight of organic matter, they con- stitute an important component. From 38 to 85 tons per acre (85 to 190 tonnes/ha) are typical values that have been reported. Down logs averaged 85 tons per acre over a 25acre (lo-ha) watershed covered with old-growth Douglas-fir-western hemlock forest (Grier and Logan 1977). Amounts within the watershed ranged widely -the lightest weights (24 tons/acre or 55 tonnes/ha) on a dry ridgetop and the heaviest (259 tons/acre or 581 tonnes/ha) on a lower slope, streamside area. Losses by downslope transfer had occurred on the ridgetop, and substantial amounts of debris had accumulated on the lower slope. In a lo-acre (4-ha) midelevation stand of Douglas-fir, western hemlock, and true firs, there were 82 tons of debris per acre (182 tonnes/ha),* 55 percent as recently fallen trees. Logs occupied 29 percent of the forest floor in this stand (fig. 18).
B MacMillan, Paul C., Joseph E. Means, Kermit Cromack, Jr., and Glenn M. Hawk. Douglas-fir decomposition, biomass, and nutrient capital in the western Cascades, Oregon. 65 p. Unpublished manuscript on file at Forestry Sciences Laboratory, Cor- vallis, Oreg.
The average weight of down logs in seven old-growth stands, from 250 to over 900 years old, was 53 tons per acre (118 tonnes/ha);1° the range was 38 to 70 tons per acre (85 to 156 tonnes/ha).” The largest accumula- tion of down wood recorded for a stand thus far is in the Carbon River Valley at Mouni Rainier National Park; a hectare plot contains 188 tons per acre (418 tonnes/ha) of logs that covered 23 percent of the plot.
Logs are also major pools of impor- tant nutrients, such as N and phos- phorus (P). In the old-growth water- shed, the log component contained 192 pounds per acre (215 kg/ha) of N and 6.0 pounds per acre (6.7 kg/ha) of P (Grier and Logan 1977). In the
Decay class: ET12 04 l Stump or snag 03 q Is 0 Live tree
midelevation stand, coarse woody debris contained 485 pounds per
Figure 18. Down logs in midelevation stand of old growth in the H. J. Andrews
acre (544 kg/ha) of N (see footnote 9). Experimental Forest. Logs occupied 29 percent of the forest floor in this stand.
lo Unpublished data on file at Forestry Sciences Laboratory, Corvallis, Oreg. Stands are at low to middle elevations in the northern Oregon and southern Wash- ington Cascade Range (from H. J. An- drews Experimental Forest, Oregon, to Mount Rainier Natioral Park,
,Washington).
It Weights of down logs and stand age are only loosely correlated. Natural young Douglas-fir stands (about 100 yearsold), surveyed at the same time as the old growth, had masses of logs as larae as some found in old-arowth stands-primarily material Earried over from previous stands as snags and logs. Large volumes of coarse woody debris are apparently characteristic of our natural forest ecosystems, adding credence to the concept of coarse woody debris as a mechanism to provide continuity of habitat from one forest generation to another (Maser et al. 1979) and for conserving large masses of organic matter and nutrients in major disturbances. It also suggests that the long-term ecological effects of nearly complete removal of woody debris in cutover stands and the prevention of new accumulations in intensively man- aged stands should be carefully examined.
31
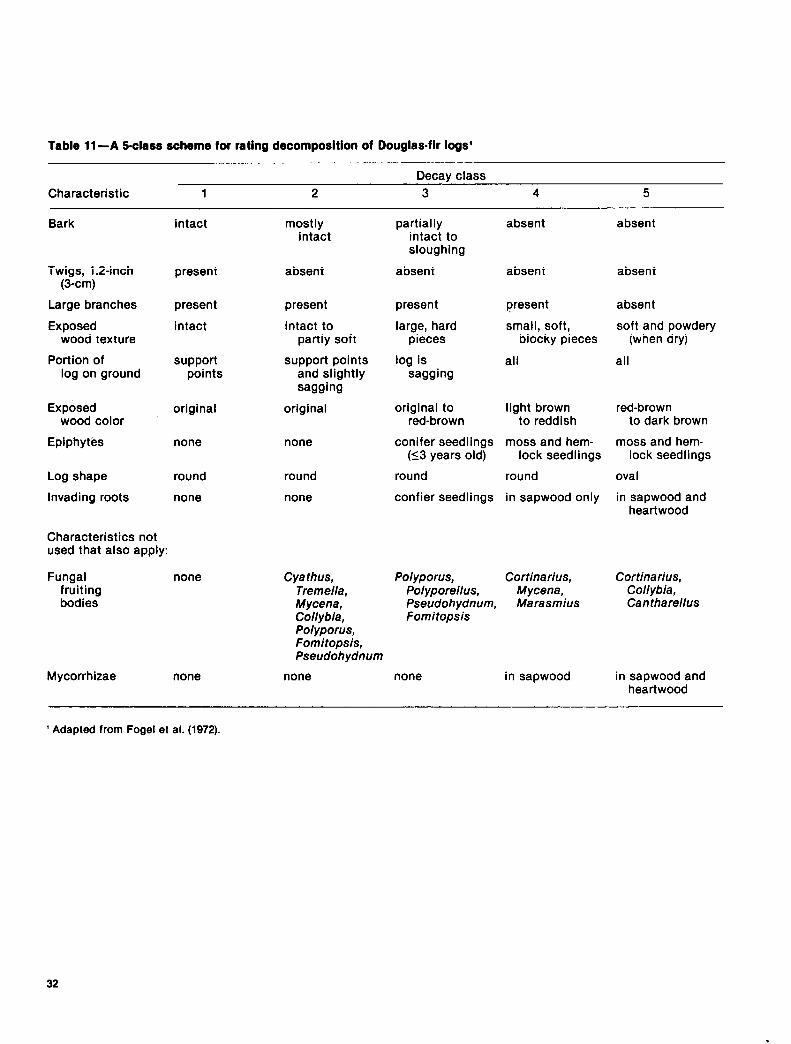
Table 11-A S-class scheme for rating decomposition of Douglasfir logs’
Characteristic 1 2 Decay class
3 4 5
Bark
Twigs, 1.2-inch (3-cm)
Large branches
Exposed wood texture
present
present
intact
Portion of support log on ground points
Exposed wood color
Epiphytes
Log shape
Invading roots
Characteristics not used that also apply:
Fungal fruiting bodies
Mycorrhizae
intact
original
none
round
none
none
none
mostly intact
absent
present
intact to partly soft
support points and slightly sagging
original
round
none
cya thus, Tremella, Mycena, Collybia, Polyporus, Fomitopsis, Pseudohydnum
none
partially intact to sloughing
absent
absent
absent
absent
absent
present
large, hard pieces
log is sagging
original to light brown red-brown to reddish
conifer seedlings moss and hem- (53 years old) lock seedlings
round round
confier seedlings in sapwood only
present
small, soft, blocky pieces
all
absent
soft and powdery (when dry)
all
red-brown to dark brown
moss and hem- lock seedlings
oval
in sapwood and heartwood
Polyporus, Cortinarius, Cortinarius, Polyporellus, Mycena, Collybia, Pseudohydnum, Marasmius Cantharellus Fomitopsis
none in sapwood in sapwood and heartwood
’ Adapted from Fogel et al. (1972).
32
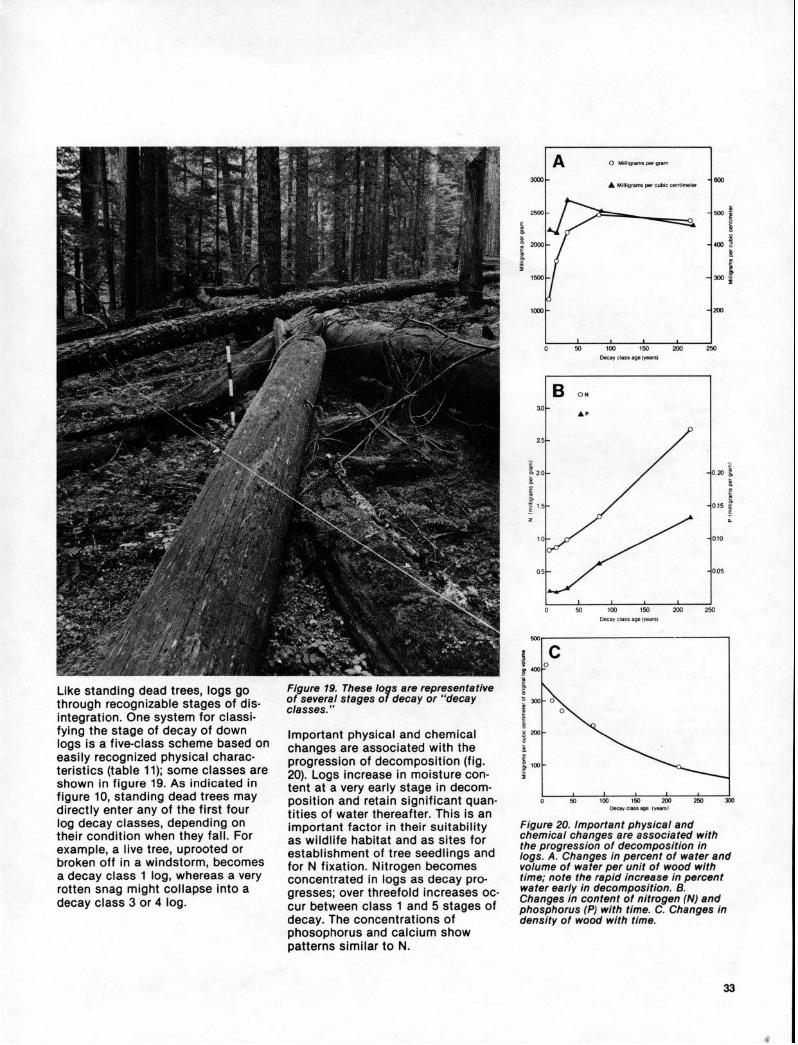
Like standing dead trees, logs go through recognizable stages of dis- integration. One system for classi- fying the stage of decay of down logs is a five-class scheme based on easily recognized physical charac- teristics (table 11); some classes are shown in figure 19. As indicated in figure 10, standing dead trees may directly enter any of the first four log decay classes, depending on their condition when they fall. For example, a live tree, uprooted or broken off in a windstorm, becomes a decay class 1 log, whereas a very rotten snag might collapse into a decay class 3 or 4 log.
Figure 79. These lo s are representative of several stages o 9 decay or “decay classes.”
Important physical and chemical changes are associated with the progression of decomposition (fig. 20). Logs increase in moisture con- tent at a very early stage in decom- position and retain significant quan- tities of water thereafter. This is an important factor in their suitability as wildlife habitat and as sites for establishment of tree seedlings and for N fixation. Nitrogen becomes concentrated in logs as decay pro- gresses; over threefold increases oc- cur between class 1 and 5 stages of decay. The concentrations of phosophorus and calcium show patterns similar to N.
Figure 20. Important physical and chemical changes are associated with the progression of decomposition in logs. A. Changes in percent of water and volume of water per unit of wood with time; note the rapid increase in percent water early in decomposition. B. Changes in content of nitrogen (N) and phosphorus (P) with time. C. Changes in density of wood with time.
33

Large logs disappear slowly. In oneold-growth Douglas-fir stand at mid-elevation, for example, a class 5 loghad charcoal on surfaces in contactwith mineral soil, suggesting that ithad fallen at the time of the wildfirethat initiated the present stand-470years ago! Decomposition rates canbe expressed as decreases in logdensity (fig. 20c). Logs lose onlyabout 40 percent of their originaldensity after 150 to 200 years.Based on decomposition models,480 to 580 years are estimated for a30-inch (80-cm) diameter Douglas-firlog to become 90 percent decayed.‘?
Habitat Function. Logs provideessential habitat for a variety of in-vertebrates (Deyrup 1975) and verte-brates (Maser et al. 1979). They areused as sites for lookouts, feedingand reproduction, protection andcover, sources and storage of food,and bedding. The high moisture con-tent of logs makes them particularlyimportant as habitat for amphibians.
Maser et al. (1979) reported that 178vertebrates use logs in the BlueMountains-14 amphibians and rep-tiles, 115 birds, and 49 mammals;they tabulated use by log decayclasses for each species. Logs areconsidered important in early suc-cessional stages as well as in old-growth forests. The persistence oflarge logs has special importance inproviding wildlife with habitat con-tinuity over long periods andthrough major disturbances.
12 Means, Joseph E., Kermit Cromack, Jr.,Paul E. MacMillan, and Alfred T. Brown.Models of Douglas-fir loin a 450-year-old forest. 25 p. Unpubl-lished manuscript on file at ForestrySciences Laboratory, Corvallis, Oreg.
Logs may contribute significantly toreestablishment of animal popula-tions by providing pathways alongwhich small mammals can ventureinto clearcuts and other bare areas.This has relevance to the rees-tablishment of tree seedlings onbared areas since survival andgrowth of new trees depend on de-velopment of appropriate mycor-rhizal associations. Surprisingly,fungal symbionts apparently disappear from cutover areas shortly aftertheir host trees are removed (Harveyet al. 1978a), and the sites must bereinoculated with their spores. Manymycosymbionts have undergroundfruiting bodies and completely de-pend on animals for disseminationof spores. Small mammals are thevectors. They consume the fungusand carry spores to new areas,thereby inoculating tree seedlings(Maser et al. 1978a, 1978b; Trappeand Maser 1978).
Logs also serve as sites for repro-duction of tree species, especiallywestern hemlock (fig. 21). This isclearly an important function innatural stands since these seedlingsand saplings supply replacementsas openings appear in the overstorycanopy. In one old-growth stand atmidelevation in the Cascade Range,over 64 percent of the western hem-lock and 4 percent of the Pacificsilver fir reproduction were rooted inrotten wood.13 The phenomenon ofnurse logs is widespread in theforest types of the Pacific North-west. Minore (1972) found that seed-lings of both Sitka spruce andwestern hemlock were morenumerous and taller on rotten logsthan on the adjacent forest floor atCascade Head Experimental Forest
in the Sitka Spruce Zone (Franklinand Dyrness 1973) on the Oregoncoast. In the South Fork Hoh RiverValley of Olympic National Park,also in the Sitka Spruce Zone, repro-duction of spruce and hemlock isessentially confined to rotten woodsubstrates;14 different species oflogs also vary in their suitability asnurse logs as evidenced by differ-ences in densities of seedlings. Insubalpine environments, such as thePacific Silver Fir Zone, successfulreproduction of western hemlock isinevitably associated with rottenlogs which are also heavily colon-ized by Pacific silver fir (Thornburgh1989, Franklin 1988). Rotten logs asseedbeds or “nurseries” may havepractical significance in a variety ofsituations; for example, as sites fornatural reproduction after shelter-wood or selective cuttings or forplanting in cutover areas. Wood sub-strates appear to have particularsilvicultural importance in coastalenvironments and where reproduc-tion of western hemlock is desired.Rotten logs can also be of key sig-nificance in perpetuating camp-grounds in old-growth forests in theCascade and the Coast Ranges byproviding seedbeds for treereproduction.15
14 LaRoi, George H., and Jerry F. Frank-lin. 1979. Picea sitchensis and Tsugaheterophylla recruitment and survivor-ship in alluvial rain forests of the HohRiver Valley, Olympic National Park. Un-published manuscript on file at ForestrySciences Laboratory, Corvallis, Oreg.15 Moehring, Janis, and Jerry F. Franklin.1977. Ohanapecosh camp roundrehabilitation study. 51 p. Unpublishedreport for the National Park Service,Mount Rainier National Park.
13 Means, Joseph E. Personal com-munication. USDA Forest Service,Forestry Sciences Laboratory, Corvallis,Oreg.
34

Rotten wood is also critical as sub- strate for ectomycorrhizal formation. In one coniferous forest stand, over 95 percent of all active mycorrhizae were in organic matter-of which 21 percent were in decayed wood (Harvey et al. 1976b). In another study in the northern Rocky Moun- tains, decayed wood in soil was im- portant in moist, mesic, and arid habitat types (Harvey et al. 1979); it was the most frequent substrate for active ectomycorrhizae on the dry site, probably because of high mois- ture levels in the wood. Mycorrhizal fungi can colonize logs, presumably using them as sources of water and
nutrients (Harvey et al. 1978). The Figure 21. Decayfng logs are important mycorrhizal relationships may be im- habitat for the reproduction of tree
portant factors in establishment of species; this “nurse log” has seedlings
seedlings on nurse logs; they are of western hemlock, western redcedar,
also important to mature trees. and Sitka spruce.
Just as quality and special proper- ties of wood products vary by tree species, the natural ecological characteristics of logs also vary by species. Average size of logs and slow rates of decay make some species, such as Douglas-fir and redcedar, more persistent. Differ- ences in value of species as nurse logs may relate to physical and chemical properties of the log or the type of wood rot infesting it; the in- ferior performance of western hemlock in the role of nurse log is one example.
35

Cycling Function. Based on currentknowledge, the most important cy-cling functions of logs are as sinksor storage compartments for energyand nutrients and as sites for N fixa-tion. In addition, logs may providephysical stability, protecting the sitefrom surface erosion.
The accumulations of carbon andnutrients represented by logs can bevery large. In the short-term view,this material is a sink since it ismade available so slowly. On theother hand, it is a significant sourceof stored energy and nutrients andone that can “bridge” major distur-bances (can be continuous from anold-growth forest through a wildfireto a young-growth stand). Trunks oflive trees, snags, and logs are struc-tures in which N is retained througha wildfire; whereas in more easilyburned organic components (leaves,forest floor) substantial N is vola-tilized and lost from the system.
The discovery of significant bac-terial N fixation in coarse woodydebris is recent; it occurred almostsimultaneously in the forests of theNortheastern, Southeastern, andNorthwestern United States. Themost thorough study reported todate is Roskoski’s (1977) in north-eastern hardwood forests. Greaterdecay and higher moisture contentswere associated with a higher inci-dence of N fixation in woody debris.Fixation occurred in an average of25 percent of the wood samples>0.4 inch (1 cm) in diameter. Largerwoody debris was probably morefavorable for N fixation because ofbetter moisture conditions (and con-sequent low oxygen levels) and be-cause larger pieces last longer,disintegrate more slowly, and, there-fore, provide greater opportunity forinoculation by the appropriatebacteria. Larsen et al. (1978) andRoskoski (1977) found higher ratesof N fixation in logs at advancedstages of decay and higher moisturecontent.
Roskoski (1977) estimated total Nfixation in hardwood stands of dif-ferent ages. The largest amountswere in the youngest (14 yearsold-l.25 + 1.80 kg/ha per year) andoldest (200 years old-0.96 + 0.77kg/ha per year) stands. The amountsof N fixed were directly related tothe weight of coarse woody debriswhich was 17 tons per acre (38tonnes/ha)-all from the previousold-growth forest-and 15 tons peracre (34 tonnes/ha) in the 4- and200-year-old stands, respectively.
Amounts of N fixation have not yetbeen estimated for old-growth con-iferous forests in the Pacific North-west, although N fixation has beendetected. Substantially greateramounts are expected than thoseestimated for eastern hardwoodforests, based on the large tonnageof woody debris present (5 to 15times that reported by Roskoski1977), and the much larger averagesize of material in the Douglas-firregion. We estimate approximately 4pounds per acre (4.4 kg/ha) per yearof N fixed in an average old-growthstand;16 a daily rate from Larsen etal. (1978) of 2.0 x 10-9 moles pergram of logs is assumed in alldecay classes throughout the yearin the more moderate year-round cli-mate on the west coast.
16 Our typical old-growth stand has 65tons per acre (145 tonnes/ha) of logs and20 tons per acre (45 tonnes/ha) of snags.
Decaying logs and decayed wood insoil are also of overwhelming impor-tance as sites for nonsymbiotic Nfixation in forests in the northernRocky Mountains (Harvey et al.1978); this is especially true of drysites and during dry periods. Nitro-gen fixation may occur in standingdead trees, even with their widerfluctuations in moisture content andaeration, but the amounts areunknown. Fixation is also asso-ciated with development of heartrots in live trees (Aho et al. 1974).Harvey et al. (1978) provide ex-amples of both situations-N fixa-tion associated with Fomes pinicola(a saprophytic fungus) in deadDouglas-fir and with Echinodontiumtinctorium (a heart rot) in livewestern hemlock.
Logs have numerous important rolesin cycling nutrients and maintainingsite productivity. Complete removalof wood residues by harvesting andbroadcast burning eliminates logsas reservoirs of nutrients and waterand as entities whose physical ef-fects may have a positive influenceon the quality of a site (Jurgensonet al. 1977). Increased commercialuse of wood residues or completeyarding of unmerchantable residuesreduces the amount of sound or par-tially decayed logs. Since the prod-ucts of decay and fire (humus,decayed wood, and charcoal) are im-portant to quality and function offorest soils (Harvey et al. 1978,Jenny 1980), future forest manage-ment must consider logs as “parentmaterial” for soil organic matter.

Logs in Streams. Large, woody debris in streams is a dominant element in aquatic ecosystems of old-growth forests. Debris largely controls the distribution of aquatic habitats, stability of streambeds and streambanks, and routing of sedi- ments and water through the stream system.
Large, organic debris enters streams by a variety of mechanisms, some of which are interrelated and occur as a chain reaction. Principal mecha- nisms of input are blowdown, slides, avalanches, deep-seated mass movements from adjacent slopes, and undercutting of streambanks. Large debris is exported from sec- tions of streams by flotation at high streamflow; in massive debris tor- rents involving the rapid down- channel movement of a slurry of soil, alluvium, and large organic debris; and by movement of fine par- ticulates and dissolved material pro duced by biological and physical breakdown of logs.
The general character of large debris in small (first to second order) and intermediate-size (third to fourth order) streams in old-growth Douglas-fir is shown in figures 22 and 23. The first-order stream (fig. 22) is choked with debris, located essentially where it fell. The third- order stream (fig. 23) is large enough to float and redistribute some of the debris, forming accumulations along the channel. The third-order stream is still narrow relative to the length of individual pieces of debris, so ac- cumulations of debris affect the width of the entire channel. In fifth- order and larger rivers, debris from old-growth forests accumulates high on streambanks at high flow, and, therefore, plays only a minor direct role in the physical and biological character of the river (Keller and Swanson 1979).
- \I y Waterflow ‘* Mlnlmum time at site (years)
0 4 Log, height above @ Largerock
METERS low water (meters) ----- Mldsummer channel
-$= /Root wad Floated organic debris
cyi, Potential stream debris, height above channel (meters)
13 Trapped sediment - Channel boundary
Figure 22. Small, first-order streams in old-growth forests are often choked with coarse woody debris as in these mapped sections of a stream in the H. J. Andrews Experimental Forest, western Oregon Cascade Range.
Figure 23. Third-order streams are large enough to float and redistribute much of the woody debris, forming distinct ac- cumulations as in this section of Mack Creek in the H. J. Andrews Experimental Forest, western Oregon Cascade Range.
37
+.
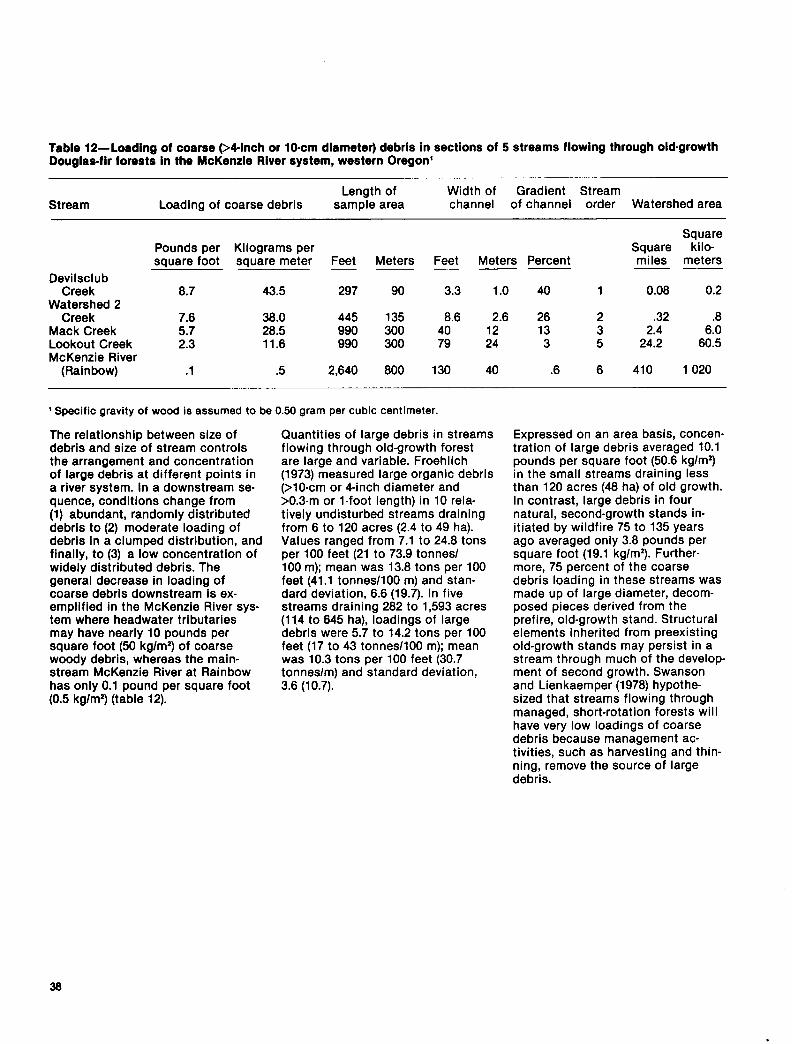
Table 12-Loadlng of coarse 94.inch or lo-cm diameter) debris in sections of 5 streams flowing through old.growth Douglas-fir forests in the McKenzie River system, western Oregon’
Stream Loading of coarse debris Length of
sample area Width of Gradient Stream channel of channel order Watershed area
Square Pounds per Kilograms per Square kilo- square foot square meter Feet Meters Feet Meters Percent miles meters - - - -~
Devilsclub Creek 0.7 43.5 297 90 3.3 1.0 40 1 0.08 0.2
Watershed 2 Creek 7.8 38.0 445 135 8.6 2.6 26 2 .32 .8
Mack Creek Z:i 28.5 990 300 40 12 13 3 2.4 6.0 Lookout Creek 11.6 990 300 79 24 3 5 24.2 60.5 McKenzie River
(Rainbow) .l .5 2,640 800 130 40 .6 6 410 1 020
( Specific gravity of wood is assumed to be 0.50 gram per cubic centimeter.
The relationship between size of debris and size of stream controls the arrangement and concentration of large debris at different points in a river system. In a downstream se- quence, conditions change from (1) abundant, randomly distributed debris to (2) moderate loading of debris in a clumped distribution, and finally, to (3) a low concentration of widely distributed debris. The general decrease in loading of coarse debris downstream is ex- emplified in the McKenzie River sys- tem where headwater tributaries may have nearly 10 pounds per square foot (50 kg/m2) of coarse woody debris, whereas the main- stream McKenzie River at Rainbow has only 0.1 pound per square foot (0.5 kg/m*) (table 12).
Quantities of large debris in streams flowing through old-growth forest are large and variable. Froehlich (1973) measured large organic debris (>lO-cm or 4-inch diameter and >0.3-m or l-foot length) in 10 rela- tively undisturbed streams draining from 6 to 120 acres (2.4 to 49 ha). Values ranged from 7.1 to 24.8 tons per 100 feet (21 to 73.9 tonnes/ 100 m); mean was 13.8 tons per 100 feet (41.1 tonnes/l00 m) and stan- dard deviation, 6.6 (19.7). In five streams draining 282 to 1,593 acres (114 to 645 ha), loadings of large debris were 5.7 to 14.2 tons per 100 feet (17 to 43 tonnes/l00 m); mean was 10.3 tons per 100 feet (30.7 tonnes/m) and standard deviation, 3.6 (10.7).
Expressed on an area basis, concen- tration of large debris averaged 10.1 pounds per square foot (50.6 kg/m’) in the small streams draining less than 120 acres (48 ha) of old growth. In contrast, large debris in four natural, second-growth stands in- itiated by wildfire 75 to 135 years ago averaged only 3.8 pounds per square foot (19.1 kg/m2). Further- more, 75 percent of the coarse debris loading in these streams was made up of large diameter, decom- posed pieces derived from the prefire, old-growth stand. Structural elements inherited from preexisting old-growth stands may persist in a stream through much of the develop ment of second growth. Swanson and Lienkaemper (1978) hypothe- sized that streams flowing through managed, short-rotation forests will have very low loadings of coarse debris because management ac- tivities, such as harvesting and thin- ning, remove the source of large debris.

Loading of debris may be abruptlyreduced by debris torrents, or“sluice-outs.” Debris torrents arerare occurrences in old-growth eco-systems; the return period of tor-rents in a 25-acre (10-ha), old-growthwatershed in the H. J. Andrews Ex-perimental Forest has been esti-mated to be from 500 to 600 years.Torrents may reduce loading ofdebris to less than 1.0 ton per 100feet (3.0 tonnes/100 m) of stream. Astorrents move past forested stream-side areas, however, banks areundercut, destabilizing trees andleading to a several-decade post-torrent period of relatively highinputs of coarse debris and reestab-lishment of stable debris dams andassociated stream habitat. Wherestreamside stands are very young(less than 30 or 40 years), pieces ofdebris after a torrent may not belarge enough to form stable accu-mulations and associated habitat.
The history of large debris instreams has been examined bydendrochronological methods(Swanson et al. 1976, Swanson andLienkaemper 1976). Large pieces ofdebris commonly have been instreams for 25 to more than 100years and have, therefore, weatheredextreme events, such as the Decem-ber 1964 flood in the Pacific North-west. Western redcedar is particu-larly long lasting, followed, in orderof increasing rate of breakdown, byDouglas-fir, western hemlock, andriparian hardwoods. This long resi-dence time results from both thecharacteristic predominance ofslowly decomposing Douglas-fir andwestern redcedar and the largedebris provided by old-growthstands.
Large debris in streams strongly in-fluences morphology of the channeland routing of sediment and water.In first-order through about third-order streams, debris helps to forma stepped longitudinal stream pro-file. Much of the energy of a streamis dissipated at falls or cascadescreated by debris. This pattern ofdissipation in a small proportion oftotal stream length results in lessenergy available for erosion of bedand banks, more storage of sedi-ment in the channel, slower routingof organic detritus, and greater di-versity of habitat than in straight,even-gradient channels.
In small and intermediate streams inthe Pacific Northwest, large debrismay be the principal factor deter-mining the characteristics ofaquatic habitats. The important roleof debris in creating habitat for fishhas been reviewed by Narver (1971),Hall and Baker (1975), and others.The wood itself is a habitat or sub-strate for much biological activity bymicrobial, invertebrate, and verte-brate organisms.
The general influence of woodydebris on aquatic habitat has beenmeasured in several streams in theH. J. Andrews Experimental Forest(Swanson and Lienkaemper 1976). Inan 800-foot (240-m) section of MackCreek, which flows through oldgrowth, 11 percent of the streamarea is woody debris, 16 percent iswood-created habitat (primarilydepositions of sediments behindwoody debris), and 73 percent isnonwoody habitat-mainly cascadesdominated by boulders. This sectionoccurs in a third-order stream chan-nel draining about 1,500 acres (600ha). Woody debris occupies 25 per-cent and wood-created habitatanother 21 percent of the streamarea of Devilsclub Creek, a first-order tributary of Mack Creek drain-ing 25 acres (10 ha). Much of thebiological activity in the processingof detritus and other consumerorganisms is concentrated in thewood and wood-created habitats.
Over half the inputs of N are asso-ciated with woody debris, includingN fixation on wood. Quantitatively,woody debris is also the majorenergy input.
Distinctions between streamecosystems in old-growth andsecond-growth stands are less clearthan those between stream orders.Large organic debris plays the domi-nant role in streams in old-growthforests and clearly does not inmanaged second-growth forests.Natural, second-growth stands haveresidual debris dams as well asresidual large stems for new damsthat help span the period until the,young stand begins producing largerdiameter woody debris. Analysis ofthe overall response of aquatic com-munities to long-term changes instructure and nutrient sources, suchas those resulting from reduced con-centrations of large, organic debris,is an important research problem.
In summary, coarse woody debris Isextremely important to streams Inold-growth coniferous forests.Debris dams and associated plungepools and trapped sediments, suchas gravel bars, provide a great diver-sity of habitats for organisms. Theresulting stepped stream profilesprovide for greater physical and bio-logical stability by dissipatingenergy otherwise used in cuttingchannels and moving sediment.Debris dams also slow the routingof other organic inputs, allowingorganisms time to more fully proc-ess these materials before they areexported downstream. Finally, thewoody debris is itself a majorsource of energy and nutrients forthe stream ecosystem.
39

Managing for Old-GrowthForests and Attributes
Perpetuating or Re-creatingOld-Growth Forests
Old-growth forests provide highlyspecialized habitats and are neitherdecadent, unproductive ecosystemsnor biological deserts. In this paper,we have contrasted the composi-tional, functional, and structuralfeatures of old growth with those ofyoung-growth forests wherever datapermit. It is clear that there are ma-jor contrasts between old-growthand managed, young-growth stands;structures, species, and patternsand rates of material (carbon andnutrient) flows and cyclescharacteristic of old growth will beabsent from managed lands unlessspecifically provided for insilvicultural prescriptions. Dif-ferences between old-growth standsand natural, second-growth standsoften appear to be one of degree;however, this is often because ofmaterial carried over from the old tothe new stand, such as large snagsand logs, which provide woodydebris and animal habitat and serveother functions.
Alternatives for managing old-growth forest ecosystems or at-tributes include management to: (1)perpetuate existing old-growthstands, (2) recreate ecosystemswith old-growth characteristics bylong rotations, and (3) provide for in-dividual features of old-growthforests. The first two alternatives in-volve a relatively small percentageof the commercial forest land in thePacific Northwest; various acreageshave been proposed in Federal land--use planning exercises, includingspotted owl management plans,generally totaling 5 percent or lessof the land base. Perpetuation of ex-isting stands is the surest coursesince the old-growth conditions ex-ist and can be expected to persist
for several additional centuries. Thisassumes viable sites (appropriatesize and shape with boundaries thatcan be protected) are selected andcatastrophic fire and windthrow areavoided. Recreation of old-growthconditions through long rotations istheoretically possible but clearly un-proved at this time. Site selection isimportant, though perhaps not ascritical as it is where perpetuationof an existing old-growth forest isthe objective.
The third alternative-providing forindividual features-involves apply-ing the information from this reportto the main body of commercialforest land in the Pacific Northwest,not simply to the small enclavesmanaged for old growth. Ourknowledge of old-growth forests andhow they work can be used to ad-vantage in managing millions ofacres of forest for timber productionby improving their ability to provideadditional ecological benefits. Someof the most important ecologicalfeatures of old-growth forests, and,indeed, the entire natural forestsere, can be duplicated on theselands with relatively small sacrificeof production of timber. Knowledgeof old growth, such-as that aboutsnags and coarse woody debris, iswidely applicable to forestmanagement.
If the forest manager decides to re-tain, maintain, or recreate someforest ecosystems with old-growthcharacteristics several questions re-quire attention. How should the old-growth allocation be distributed overthe landscape? Which managementpractices should be followed inareas selected for retention of oldgrowth and on areas to be managedon long rotations? Size and shape ofold-growth enclaves are, distribu-tional concerns. Structural featuresare the management key in eitherperpetuating or recreating old-growth stands.
Distribution of Old-Growth Manage-ment Areas. Having decided tomaintain a certain percentage of amanagement unit as old-growthforest, a land manager must decidehow it is most effectivelydistributed. Should a small stand bemaintained in each drainage or com-partment or fewer large tracts re-served? Where are boundaries bestlocated? The history of naturalwildfires is somewhat instructive, atleast in indicating the landscapepatterns in which existing old-growth stands originated. Standswere typically established in blocksof hundreds of acres. Boundariesmost often were along topographicfeatures, such as ridges or streams.“Feathering” of boundaries, result-ing in large areas of mixed stands(residual old growth scatteredthrough a young stand), was com-mon. Fires often skipped largepatches of trees, particularly onlower slopes and stream bottoms, orstands protected by natural barriers.
How large a drainage unit or standis essential for a viable old-growthecosystem is a difficult question.Generally, an area of 300 to 500acres (120 to 200 ha) is sufficient formost plants and animals. This sizehas been suggested as essential toa pair of breeding northern spottedowls, for example, one of the mostwide-ranging species dependent onold growth. McClelland et al. (1979)

suggest 50 to 100 acres (20 to 40 ha)for the nesting and feeding areas ofcavity-dwelling birds. Generally, 500to 1,000 acres (200 to 400 ha) areneeded for a third-order streamdrainage. Areas much smaller than500 acres (200 ha) can, on the otherhand, preserve many attributes ofold-growth forests, particularly if theboundaries chosen for the area pre-vent rapid deterioration of the stand.
Entire drainage basins, even forsmall first- or second-order streams,are ecologically the most desirableunits for old-growth forest manage-ment. Such drainages have naturaltopographic boundaries that oftenprovide superior protection fromwindthrow and other external influ-ences for the reserved stand. A vari-ety of ecological conditions arepresent, along with a stream systemin which natural land-water inter-actions (for example, woody debrisinputs to the stream) can continue.Plant and animal diversity will alsobe higher in a drainage basin thanin an isolated upland forest stand ofthe same size.
Another useful location for old-growth management areas is alongstreams. Streamside strips (bufferzones) were originally conceived forshading streams and minimizing in-creases in water temperatures; amore valuable function in streamsup through at least third order is inproviding essential energy, struc-tural inputs (for debris dams), andstability of banks. Debris dams mustbe continually created to replacebroken dams; this is particularly im-portant after infrequent debris tor-rents that remove all, or most,pieces of large debris. Streamsidestrips of old-growth forest will pro-vide for continued physical and bio-logical stability of the aquaticecosystem.
Streamside and roadside strips ofold growth have the additionaladvantage of providing migrationroutes for organisms dependent onmature forests. The strips skirt man-aged stands and provide continuitybetween otherwise isolated pocketsof natural and old-growth forest.Such migration routes may beimportant for avoiding loss ofspecies as “islands” of habitatsuited to a species become morelimited and isolated MacClintock etal. 1977). The protected travel routesprovided by these strips allow or-ganisms to migrate in response toshifts in location of suitable habitat.
Reserve strips must be in appropri-ate locations and of sufficient sizeto survive normal windstorms.Streamside strips have often beennarrow (since shading the streamwas the primary objective) and sharpedged; consequently, they areextremely susceptible to blowdown.The wetter soils on lower slopes andstreamsides do not make reten-tion of strips any easier, and upslope harvesting often intensifiesthis problem. Regular programs tosalvage logs on roadside or stream-side strips are inappropriate be-cause they accelerate deteriorationof the stand and remove the essen-tial structural components. Reten-tion of dead wood is especiallyimportant for streams where it is thesource of stabilizing debris dams.Salvage logging may be appropriateif losses from catastrophic wind-throw or other causes occur. Eventhen, the salvage should not becomplete; some down materialshould be selected and left on theland and in the stream. A 200-foot-wide (60-m) streamside or roadsidestrip is not a viable unit in mostcases; considerable ingenuity andeffort will be necessary to identifyand lay out viable reserve strips.
Areas with problems that limitpotential for management may alsobe appropriate sites for perpetuatingold-growth ecosystems and orga-nisms. An example could be steeplandslide-prone headwall areas thatdepend on a strong and continuingroot mantle for stability.
Structural Attrlbutes in Perpetuatingor Re-creating Old Growth. The dis-tinctive structural features-thelarge, old-growth trees, snags, andlogs on land and in streams-pro-vide the major key to managementstrategies. The unique, importantcompositional and functional fea-tures of an old-growth forest usuallyaccompany these structuralelements.
An old-growth forest is much morethan simply a collection of largetrees. The dead, organic componentis as important as the highly indi-vidualistic, large trees. Decayingsnags and logs, particularly instreams, are beneficial and must beprovided for in managementschemes; they should not be viewedsolely as waste, fire hazards, or im-pediments to management. Snagsand logs play important roles ashabitat for various organisms and inconserving and cycling nutrientsand energy. To a large degree, suc-cess in managing forests for old-growth attributes will depend onlearning to manage the dead,organic material as cleverly as thelive trees.
There are implications here for man-agement of old-growth stands se-lected for perpetuation. Salvage log-ging is inappropriate since itremoves at least two of the majorstructural components-dead anddown-that are key elements of thesystem. In all likelihood, some of themore decadent, live trees would alsobe removed. Salvage logging is alsoinappropriate because of the dam-age inevitably done to root systemsand trunks of the residual standwhich results in accelerated mortal-ity of trees and overall deteriorationof the stand.17
17 When stands are selected for preserva-tion (for example, as roadside or stream-side strips), the first (and frequently re-peated) management activity is often asalvage program. If a manager wants toretain an old-growth ecosystem or amature forest stand, entries should beavoided or at least minimized. Treesviewed as safety or fire hazards may bet-ter be felled and left in place than
41

There are also implications if themanager wishes to create an old-growth environment (after a stand iscut) by using a long rotation. Ini-tially, foresters may retain largeramounts of woody residues, espe-cially down logs, from the previousstand. Retention of scattered indi-vidual, old-growth trees may be use-ful as sources of epiphytic flora andeventually of large, dead standingtrees and down logs.
Rapid development of large, long-crowned trees as early as possibleis a key objective of managementthat can be aided several ways.Selection of understocked stands ofreproduction as sites for creatingold-growth stands is one approachsince individuals will grow fasterand lose lower branches moreslowly under open-grown conditions.Many existing old-growth standsmay have regenerated slowly (Frank-lin and Waring 1980); growth pat-terns of individual trees suggestgrowing conditions essentially freefrom competition for a century ormore. If initial densities of standsare moderate-at current recom-mended levels for managed stands-precommercial and commercialthinnings will be necessary duringthe first 100 years of a long-rotationforest management cycle. Growthrates of individual trees will be toolow at high densities, or at moder-ate densities on less productivesites, to produce desired sizes ofstems even after 200 years; thin-nings and partial cuttings are essen-tial under those conditions. Greatcare must be taken, however, tominimize damage to residual trees.
Creation of appropriate types andamounts of standing dead and downtrees is a specific managementobjective. Snags and logs from theoriginal stand should be avoidedduring intermediate cuttings. Up toabout 100 years, the size of snagsand logs produced by the youngstand is probably not of particularecological importance. Some of thismaterial could be removed alongwith excess live trees-those thatwill die before reaching significantdiameter (50 cm or 20 inches). Open-ings for development of shade-toler-ant species can also be created thisway; if these species do not come innaturally, they could be artificiallyintroduced, possibly by underplant-ing. The large snags, logs, and anylive old growth left from the originalstand should not be removed duringsalvage operations.
After about 100 years, partial cuttingof any type becomes increasinglyinappropriate. There are fewer livedominants, and their loss, eitherdirectly by cutting or graduallythrough damage to roots and trunks,is undesirable. Standing dead treesand logs now being recruited fromthe live stands are of sufficient sizeto fully perform desired habitat andcycling functions.
To summarize, if the objective isperpetuation of an old-growth forestecosystem, a minimum amount ofdisturbance should be allowed.Snags and logs perform importantfunctions and are essential struc-tures. When the objective is tocreate an old-growth forest fromscratch, large individual trees withlarge crowns should be grown asquickly as possible. Scattered old-growth trees and rotten logs fromthe original stand should be re-tained and reproduction of westernhemlock, western redcedar, andother shade-tolerant associatesunder the Douglas-fir canopiesencouraged. Partial cuttings may beuseful and will be necessary inmoderate to heavily stocked standsof reproduction if large trees are tobe attained as quickly as possible.After trees are about 100 years old,such cuttings are increasinglyinappropriate, however.
For multiple-use objectives, anincreased awareness of the natureand nontimber value of individualtrees is important; for example,potential or current value as habitatfor epiphytic communities and wild-life. Knowledge of the ecologicalroles of standing, dead trees andlogs beyond their value as wildlifehabitat is also desirable.
42

3Managing for individual Old-GrowthAttributes
There is considerable logic in man-aging entire stands or small drain-ages for old-growth attributes. Theold-growth ecosystem is a system ofmany interlinked components, in-cluding organisms. The serial rela-tionship of the key structural com-ponents has been discussed-froma large, old-growth tree to a nearlydecomposed, rotten log (fig. 10). Fur-ther, some organisms or functionsmay depend on an intact old-growthforest for their perpetuation.
Nevertheless, a forester may wish tomanage for some individual old-growth attributes. This is, in fact,how the forester can put some ofthe information on old growth towork to increase ecological benefitsfrom intensively managed timber-lands. The structural componentsagain provide the key. Perhaps mostobvious is providing for large snagsand logs. This can be done in thefirst-generation managed forest byretaining some of this material fromthe virgin stand. The tendency hasbeen to remove all such materialsas a safety measure and to reducelogging residues, which are viewedas fire hazards and impediments inregeneration and other siivicuituraiactivities. in second- and third-generation stands, a forester willhave to create appropriate materialssince neither large snags nor logswill usually be present.
The need for snags was recognizedfirst by wildlife managers, and theyhave more recently recognized thevalue of logs (Maser et al. 1979,Thomas et al. 1979a). Thomas et al.(1979a) led in developing guidelineson sizes and numbers of such mate-rial needed to provide for verte-brates; although their research wasconducted in the Blue Mountains ofeastern Oregon and Washington, thesame principles apply on the westside of the Cascade Range, asshown by Mannan et al. (1980) andCline et al. (1980). These authorssuggest that snags be created fromdefective, living trees and urge main-tenance of large snags covering thespectrum of decomposition. Densi-ties of snags in natural, old-growthstands are proposed as an interimmanagement guide until more dataare developed. Cline et al. (1980)also suggest leaving snags ingroups to reduce problems of safetyand fire control. It is important toremember that much more thanhabitat for vertebrate animals isinvolved in preserving snags; stand-ing dead trees and logs serve otherfunctions as well.18
18 The role of dead wood in cycling andconserving nutrients, especially N, is anoutstanding example. Ten years agonothing was known about sources of Nin old-growth stands, other than the at-mospheric input. In the interim, epiphyticlichens and wood-dwelling bacteria havebeen identified as significant sites forfixation. There are several importantsources for additions of N in both theearly stages of forest succession-nonleguminous N fixers, such as alder(Alnus spp.) and ceanothus (Ceanothusspp.)-and in old-growth forests. Ex-isting management strategies call forquick establishment of conifer canopiesand short rotations-which effectivelyeliminate these additions of N to inten-sively managed sites.
There are currently no good guidesto the number and sizes of logs thatshould be left on cutover areas.Removal of ail coarse woody debrisis not the best ecological practice.Costs and benefits of some prac-tices (such as yarding unmerchant-able material) are not known; nega-tive impacts on long-term site pro-ductivity, wildlife, and erosion mayoffset the benefits to fire protectionand ground accessibility. it doesappear that at least several largerlogs per acre are needed for wildlife,especially small mammals. Definingthe types and sizes of logs andother woody debris desired in man-aged stands is a major problem forresearch.
Retention of small groups of old-growth trees, or scattered individualtrees, may be a useful practice. Thiswas conceived as a technique forproviding a source of epiphytic"inoculum” for adjacent youngtrees. Lack of such a source may bea factor in the absence of the N-fix-ing epiphytes on trees less than 150years old. Leaving occasional old-growth trees has another advantage-it will, in the long run, provide asource of large snags and logs. Thismay be the easiest strategy for per-petuating these structural compo-nents into second- and even third-generation managed stands.
4 3

Trees Mentioned in This Report Acknowledgments
Common nameAlaska-cedarCoast redwoodDouglas-firGrand firIncense-cedarNoble firPacific silver firPort-Orford-cedarSitka spruceSugar pineWestern hemlockWestern redcedarWestern white pine
Scientific nameChamaecyparis nootkatensis (D. Don) SpachSequoia sempervirens (D. Don) Endl.Pseudotsuga menziesii (Mirb.) FrancoAbies grandis (Dougl. ex D. Don) Lindl.Libocedrus decurrens Torr.Abies procera Rehd.Abies amabilis (Dougl.) ForbesChamaecyparis lawsoniana (A. Murr.) Parl.Picea sitchensis (Bong.) CarrPinus lambertiana Dougl.Tsuga heterophylla (Raf.) Sarg.Thuja plicata DonnPinus monticola Dougl. ex D. Don
William E. Emmingham and Lewis F.Roth provided data and ideas at awork session on old-growth forest,sponsored by the USDA Forest Serv-ice. Gerald N. Ptachen AssistantSupervisor of the Willamette Na-tional Forest, provided encourage-ment and many useful ideas fromthe perspective of a resourcemanager. William H. Moir, Glenn M.Hawk, T. B. Thomas, and Joseph E.Means provided useful ideas, data,and assistance in completion of thisreport. Much of the information isfrom the Coniferous Forest Biome(U.S. International Biological Pro-gram) research group at the H. J.Andrews Experimental Forest. Thisreport is Contribution No. 356 fromthe Coniferous Forest Biome.

Literature Cited
Aho, P. E., R. J. Seidler, H. J. Evans,and P. N. Raju.
1974. Distribution, enumeration,and identification of nitrogen-fixing bacteria associated withdecay in living white fir trees.Phytopathology 64(11):1413-1420.
Aho, R. S.1976. A population study of thecutthroat trout in an unshadedand shaded section of stream.MS. thesis. Oreg. State Univ.,Corvallis. 87 p.
Anderson, H. W., M. D. Hoover, andK. G. Reinhart.
1976. Forests and water: Effectsof forest management on floods,sedimentation, and water supply.USDA For. Serv. Gen. Tech. Rep.PSW-18, 115 p. Pac. SouthwestFor. and Range Exp. Stn., Berke-ley, Calif.
Anderson, N. H., and J. R. Sedell.1979. Detritus processing bymacroinvertebrates in stream eco-systems. Annu. Rev. Entomol.24:351-377.
Berntsen, C. M.1960. Productivity of a matureDouglas-fir stand. USDA For. Serv.Pac. Northwest For. and RangeExp. Stn. Res. Note 188, 4 p. Port-land, Oreg.
Boyce, J. S., and J. W. Bruce Wagg.1953. Conk rot of old-growthDouglas-fir in western Oregon.Oreg. For. Prod. Lab. Bull. 4, 96 p.
Bull, E. L.1978. Specialized habitat require-ments of birds: Snag manage-ment, old-growth, and riparianhabitat. Proceedings of the work-shop on nongame bird habitatmanagement in the coniferous for-ests of the Western United States.USDA For. Serv. Gen. Tech. Rep.PNW-64, p. 74-82. Pac. NorthwestFor. and Range Exp. Stn., Port-land, Oreg.
Bull, E. L., and E. C. Meslow1977. Habitat requirements of thepileated woodpecker in northeast-ern Oregon. J. For. 75(6):335-337.
Cline, S. P.1977. The characteristics anddynamics of snags in Douglas-firforests of the Oregon CoastRange. M.S. thesis. Oreg. StateUniv., Corvallis. 106 p.
Cline, S. P., A. B. Berg, and H. M.Wight.
1980. Snag characteristics anddynamics in Douglas-fir forests,western Oregon. J. Wildl. Manage.44(4):773-786.
DeGraaf R. M., Tech. Coord.1978. Proceedings of the work-shop on nongame bird habitatmanagement in the coniferousforests of the Western UnitedStates. USDA For. Serv. Gen.Tech. Rep. PNW-64, 100 p. Pac.Northwest For. and Range Exp.Stn., Portland, Oreg.
Deyrup, M. A.1975. The insect community ofdead and dying Douglas-fir. I. TheTymenoptera. Coniferous For.Biome Bull. 6, 104 p. Univ. Wash.,Seattle.
Dyrness, C. T., J. F. Franklin, W. H.Moir.
1974. A preliminary classificationof forest communities in the cen-tral portion of the western Cas-cades in Oregon. Coniferous For.Biome Bull. 4, 123 p. Univ. Wash.Coll. For. Resour., Seattle.
Edgerton, P. J., and J. W. Thomas.1978. Silvicultural options andhabitat values in coniferous for-ests. In Proceedings of the work-shop on nongame bird habitatmanagement in the coniferous for-ests of the Western United States.USDA For. Serv. Gen. Tech. Rep.PNW-64, p. 56-65. Pac. NorthwestFor. and Range Exp. Stn., Port-land, Oreg.
Fogel, R., M. Ogawa, and J. M.Trappe.
1972. Terrestrial decomposition: Asynopsis. Coniferous For. BiomeIntern. Rep. 135, 12 p. Univ. Wash.Coll. For. Resour., Seattle.
Franklin, J. F.1966. Vegetation and soils in thesubalpine forests of the southernWashington Cascade Range.Ph. D. thesis. Wash. State Univ.,Pullman. 132 p.
Franklin, J. F., and C. T. Dyrness.1973. Natural vegetation of Ore-gon and Washington. USDA For.Serv. Gen. Tech. Rep. PNW-80, 417p. Pac. Northwest For. and RangeExp. Stn., Portland, Oreg.
Franklin, J. F., and R. H. Waring.1980. Distinctive features of thenorthwestern coniferous forest:Development, structure, and func-tion. In Forests: Fresh perspec-tives from ecosystem analysis.R. H. Waring, ed. Proc. 40th Annu.Biol. Colloq. 1979:59-86. Oreg.State Univ. Press, Corvallis.
Fredriksen, R. L.1970. Erosion and sedimentationfollowing road construction andtimber harvest on unstable soilsin three small western Oregonwatersheds. USDA For. Serv. Res.Pap. PNW-104, 15 p. Pac. North-west For. and Range Exp. Stn.,Portland, Oreg.
45

Fredriksen, R. L.1972. Nutrient budget of a Doug-las-fir forest on an experimentalwatershed in western Oregon. InProceedings: Research on conifer-ous forest ecosystems-a sympo-sium, p. 115-131. USDA For. Serv.Pac. Northwest For. and RangeExp. Stn., Portland, Oreg.
Fredriksen, R. L., D. G. Moore, andL. A. Norris.
1975. The impact of timber har-vest, fertilization and herbicidetreatment on streamwater qualityin western Oregon and Washing-ton. In Forest soils and forestland management, p. 283-313. LesPresses de I’Universite Laval,Quebec.
Froehlich, H. A.1973. Natural and man-causedslash in headwater streams. Log-gers Handb. Vol. 33, 8 p. Pac.Logging Conf., Portland, Oreg.
Furman, T. E., and J. M. Trappe.1971. Phylogeny and ecology ofmycotrophic achlorophyllousangiosperms. Q. Rev. Biol.46(3):219-225.
Gashwiler, J. S.1970. Plant and mammal changeson a clearcut in west-central Ore-gon. Ecology 51(6):1018-1026.
Gholz, H. L., F. K. Fitz, and R. H.Waring.
1976. Leaf area differences asso-ciated with old-growth forest com-munities in the western OregonCascades. Can. J. For. Res.6(1):49-57.
Grier, C. C., and R. S. Logan.1977. Old-growth Pseudotsugamenriesii communities of a west-ern Oregon watershed: Biomassdistribution and production bud-gets. Ecol. Monogr. 47(4):373-400.
Hall, J. D., and C. O. Baker.1975. Biological impacts oforganic debris in Pacific North-west streams. Debris in streams,workshop 1, Sept. 1975, 13 p.Oreg. State Univ., Corvallis.
Harvey, A. E., M. F. Jurgensen, andM. H. Larsen.
1976a. Intensive fiber utilizationand prescribed fire: Effects on themicrobial ecology of forests.USDA For. Serv. Gen. Tech. Rep.INT-28, 46 p. Intermt. For. andRange Exp. Stn., Ogden, Utah.
Harvey, A. E., M. F. Jurgensen, andN. J. Larsen.
1978. Role of residue in andimpacts of its management of for-est soil biology. 8th World For.Congr. FAO Spec. Pap., 11 p.
Harvey, A. E., M. J. Larsen, and M. F.Jurgensen.
1976b. Distribution of ectomycor-rhizae in a mature Douglas-fir/larch forest soil in western Mon-tana. For. Sci. 22(4):393-398.
Harvey, A. E., M. J. Larsen, and M. F.Jurgensen.
1979. Comparative distribution ofectomycorrhizae in soils of threewestern Montana forest habitattypes. For. Sci. 25(1):350-358.
Jenny, H.1980. Alcohol or humus? Science2909(4455):444.
Jurgenson, M. F., M. J. Larson, andA. E. Harvey.
1977. Effects of timber harvestingon soil biology. Proc. Soc. Am.For. Annu. Conv., p. 244-250.
Keller, E. A., and F. J. Swanson.1979. Effects of large organicmaterial on channel form and flu-vial processes. Earth Surf. Pro-cesses 4:361-380.
Larsen, M. J., M. F. Jurgensen, andA. E. Harvey.
1978. N2 fixation associated withwood decayed by some commonfungi in western Montana. Can. J.For. Res. 8(3):341-345.
Long, J. M., and J. Turner.1975. Above ground biomass ofunderstory and overstory in anage sequence of four Douglas-firstands. J. Appl. Ecol. 12:179-188.
Long, J. N.1977. Trends in plant species di-versity associated with develop-ment in a series of Pseudotsugamenziesii/Gaultheria shallonstands. Northwest Sci. 51(2):119-130.
Lyford, J. H., and S. V. Gregory.1975. The dynamics and structureof periphyton communities inthree Cascade Mountain streams.Verh. Int. Ver. Limnol. 19:1810-1818.
McClelland, B. R., Frissell, S. S.,Fischer, W. C., and C. H. Halvorson.
1979. Habitat management forhole-nesting birds in forests ofwestern larch and Douglas-fir.J. For. 77(8):480-483.
MacClintock L., R. F. Whitcomb,and B. L. Whitcomb.
1977. Island biogeography and“habitat islands” of eastern for-ests. II. Evidence for the value ofcorridors and minimization of iso-lation in preservation of bioticdiversity. Am. Birds 31(1):6-12.
Mannan R. W., E. C. Meslow, andH. M. Wight.
1980. Use of snags by birds, Doug-las-fir forests, western Oregon.J. Wildl. Manage. 44(4):787-797.
King, J. P.1981. Growth and mortality in theWind River Natural Area. J. For.59(10):768-769.
46

Maser, C., R. Anderson, K. Cromack,Jr., J. T. Williams, and R. E. Martin.
1979. Dead and down woody ma-terial. Chapter 6. In Wildlife habi-tats in managed forests: The BlueMountains of Oregon and Wash-ington. J. W. Thomas, tech. ed.U.S. Dep. Agric. Agric. Handb. 553,p. 78-95.
Maser, C., J. M. Trappe, and R. A.Nussbaum.
1978a. Fungal-small mammalinterrelationships with emphasison Oregon coniferous forests.Ecology 59(4):799-809.
Maser, C., J. M. Trappe, and D. C.Ure.
1978b. Implications of small mam-mal mycophagy to the manage-ment of western coniferous for-ests. 43d North Am. Wildl. andNat. Resour. Conf. Trans., p. 78-88.
Meslow E. C.1978. The relationship of birds tohabitat structure-plant communi-ties and successional stages. InProceedings of the workshop onnongame bird habitat manage-ment in the coniferous forests ofthe Western United States. USDAFor. Serv. Gen. Tech. Rep.PNW-64, p. 12-18. Pac. NorthwestFor. and Range Exp. Stn., Port-land, Oreg.
Minore, D.1972. Germination and earlygrowth of coastal tree species onorganic seed beds. USDA For.Serv. Res. Pap. PNW-135, 18 p.Pac. Northwest For. and RangeExp. Stn., Portland, Oreg.
Murphy, M. L.1979. Predator assemblages inold-growth and logged sections ofsmall Cascade streams. M.S. the-sis. Oreg. State Univ., Corvallis.72 p.
Narver, D. W.1971. Effects of logging debris onfish production. In Forest landuses and stream environment,p.100-111. J. T. Krygier and J. D.Hall, eds. Oreg. State Univ., Cor-vallis.
Odum, E. P.1971. Fundamentals of ecology.3d ed. 574 p. W. B. Saunders Co.,Philadelphia.
Pike, L. H., W. C. Denison, D. M.Tracy, M. A. Sherwood, and F. M.Rhoades.
1975. Floristic survey of epiphyticlichens and bryophytes growingon old-growth conifers in westernOregon. Bryologist 78(4):389-402.
Pike, L. H., R. A. Rydell, and W. C.Denison.
1977. A 400-year-old Douglas-firtree and its epiphytes: Biomass,surface area, and their distribu-tions. Can. J. For. Res. 7(4):680-699.
Roskoski, J. 1977. Nitrogen fixation in northernhardwood forests. Ph. D. thesis.Yale Univ., New Haven, Conn.112 p.
Scott, V. E.1978. Characteristics of ponder-osa pine snags used by cavitynesting birds in Arizona. J. For.76(1):26-28.
Shidei, T., and T. Kira.1977. Primary productivity ofJapanese forests-productivity ofterrestrial communities. JIBPSynth.; Vol. 16, 289 p. Univ. TokyoPress.
Sollins, P., C. C. Grier, F. M.McCorison K. Cromack, Jr.,R. Fogel, and R. L. Fredriksen.
1980. The internal element cyclesof an old-growth Douglas-fir eco-system in western Oregon. Ecol.Monogr. 50(3):261-285.
Strahler, A. N.1957. Quantitative analysis ofwatershed geomorphology. Am.Geophys. Union Trans. (EOS)38:913-920.
Swanson, F. J., R. L. Fredriksen, andF. M. McCorison.
1981a. Material transfer in a west-ern Oregon forested watershed. InThe natural behavior and re-sponse to stress of western conif-erous forests. R. L. Edmunds, ed.Dowden Hutchinson & Ross, Inc.,Stroudsburg, Pa.
Swanson, F. J., S. V. Gregory, J. R.Sedell, and A. S. Campbell.
1981b. Land-water interaction: Theriparian zone. In The natural be-havior and response to stress ofwestern coniferous forests. R. L.Edmunds, ed. Dowden Hutchin-son & Ross, Inc., Stroudsburg, Pa.
Swanson, F. J., and G. W. Lien-kaemper.
1978. Physical consequences oflarge organic debris in PacificNorthwest streams. USDA For.Serv. Gen. Tech. Rep. PNW-69, 12 p.Pac. Northwest For. and RangeExp. Stn., Portland, Oreg.
Swanson, F. J., G. W. Lienkaemper,and J. Sedell.
1976. History, physical effects andmanagement implications of largeorganic debris in western Oregonstreams. USDA For. Serv. Gen.Tech. Rep. PNW-56, 15 p. Pac.Northwest For. and Range Exp.Stn., Portland, Oreg.
47

Thomas, J. W., Tech. Ed.1979. Wildlife habitats in managedforests: The Blue Mountains ofOregon and Washington. U.S. Dep.Agric. Agric. Handb. 553, 512 p.
Thomas, J. W., R. G. Anderson,C. Maser, and E. L. Bull.
1979a. Snags. Chapter 5. In Wild-life habitats in managed forests:The Blue Mountains of Oregonand Washington. J. W. Thomas,tech. ed. U.S. Dep. Agric. Agric.Handb. 553, p. 60-77.
Thomas, J. W., C. Maser, and J. E.Rodiek.
1979b. Riparian zones. Chapter 3.In Wildlife habitats in managedforests: The Blue Mountains ofOregon and Washington. J. W.Thomas, tech. ed. U.S. Dep. Agric.Agric. Handb. 553, p. 40-47.
Thomas, J. W., R. J. Miller, C. Maser,R. G. Anderson, and B. E. Carter.
1979c. Plant communities andsuccessional stages. Chapter 2.Wildlife habitats in managed for-ests: The Blue Mountains of Ore-gon and Washington. J. W.Thomas, tech. ed. U.S. Dep. Agric.Agric. Handb. 553, p. 22-39.
Thornburgh, D. A.1969. Dynamics of the true fir-hemlock forests of western Wash-ington. Ph. D. thesis. Univ. Wash.,Seattle. 192 p.
Trappe, J. M., and C. Maser.1978. Ectomycorrhizal fungi: Inter-actions of mushrooms and truf-fles with beasts and trees. InFungi and man, p. 163-169. A. B.Walters, ed. Linn-Benton Comm.Coll., Albany, Oreg.
U.S. Department of Agriculture, For-est Service Region 6.
1977. Draft environmental state-ment for Hebo planning unit ofthe Siuslaw National Forest,Oregon. USDA For. Serv. Reg. 6,176 p., illus. Pac. Northwest Reg.,Portland, Oreg.
Waring, R. H., and J. F. Franklin.1979. The evergreen coniferousforests of the Pacific Northwest.Science 204(4400):1380-1386.
Williamson, R. L., and F. E. Price.1971. Initial thinning effects in 70-to 150-year-old Douglas-fir-western Oregon and Washington.USDA For. Serv. Res. Pap.PNW-117, 15 p. Pac. NorthwestFor. and Range Exp. Stn., Port-land, Oreg.
*U.S . GOVERNMENT PRINTING OFFICE: 1990.0-791.171/20024

Franklin, Jerry F., Kermit Cromack, Jr., William Denison ArthurMcKee, Chris Maser, James Sedeii, Fred Swanson, and GlenJuday.
1981. Ecological characteristics of old-growth Douglas-firforests. USDA For. Serv. Gen.Tech. Rep. PNW-118, 48 p. Pac.Northwest For. and Range Exp. Stn., Portland, Oreg.
Old-growth coniferous forests differ significantly from younggrowth forests in species composition, function (rate and pathsof energy flow and nutrient and water cycling), and structure.Most differences can be related to four key structural com-ponents of old growth: large live trees, large snags, large logs onland, and large logs in streams. Foresters wishing to maintainold-growth forest ecosystems can key management schemes tothese structural components.
Keywords: Ecosystems, old-growth stands, stand composition,stand structure, Douglas-fir, Pseudotsuga menziesii, westernhemlock, Tsuga heterophylla.

The Forest Service of the U.S. Department ofAgriculture is dedicated to the principle of multipleuse management of the Nation’s forest resources.for sustained yields of wood, water, forage, wildlife,and recreation. Through forestry research,cooperation with the States and private forestowners, and management of the National Forestsand National Grasslands, it strives - as directed byCongress - to provide increasingly greater serviceto a growing Nation.
The U.S. Department of Agriculture is an EqualOpportunity Employer. Applicants for all Departmentprograms will be given equal consideration withoutregard to age, race, color, sex, religion, or nationalorigin.











![Conifer expansion [Chapter M.] - fs.fed.us](https://img.pdfslide.us/doc/110x75/61cace1c98ffe14ced450e01/conifer-expansion-chapter-m-fsfedus.jpg)