Embed Size (px)
Citation preview

4.1
Enhanced environmental conditions cause changes in qualityand quantity of extruded compounds on the cell surface
Alicja M. Zobel1, Gosia Zobel2, Janina Turło3
1Spectroscopy Institute, Trent University, Peterborough, ON, Canada K9J 6Y1; 2 Department of Animal Welfare, University of British Columbia, Vancouver, BC, Canada V6T 1Z4; 3Medical Academy, Banacha 1, Warsaw, Polande-mail: Alicja Zobel <[email protected]>
Abstract: Ultraviolet radiation and environmental pollu-tion by metal elements caused an increase in concentra-tion of natural products extruded onto the exterior epi-cuticular waxes on the plant surface. Eighty-six per cent of the total furanocoumarins found were on these waxes, acting as a shield against ultraviolet radiation, while the remaining 14% were on the surface of cells adjacent to the intercellular spaces. Callus cells exposed to extreme environmental conditions extruded coumarins to the surface. Mineral elements caused extrusion of natural products onto the surface of tissue cultures of both plant and fungal cells. A variety of these natural products are incorporated in varying amounts into both human food and animal feed. Introduction: Since 1988 Zobel and Brown have investi-gated the surface compounds on shoots, seeds, fruit and roots. R. graveolens was the first model system, and even on its callus culture there were coumarins up to 86% of
the total. In the true plant the remaining 14% of the cou-marins were in the interior on the cell walls of the inter-cellular spaces, which need to be protected by antimitot-ics against bacteria and fungal hyphae migrating inside the plant through the stomata, and penetrating the root interior with migrating soil water. Thus it was no surprise that single callus cells had extruded coumarins on the surface, or that substantial deposits of a variety of natural products are on the root surface. Glycosides of secondary metabolites are located in the vacuoles and play impor-tant roles as antioxidants and even hormones.Materials and Methods: Surface compounds were re-moved simply by melting the surface wax by dipping the tissue for 1–2 seconds in almost-boiling water, thus extracting both water- and fat-soluble compounds. After the dipping, remaining compounds were removed by a modified Harborne’s method [2] with acidified ethanol, in which the concentration was lowered to 25%. After ten minutes’ boiling to kill the tissue the mixture was allowed to stand overnight. Compounds were thus removed from the cell walls and eluted from the vacuoles after plasma-lemma and tonoplasts had been destroyed. Localization of coumarins. Concentrations of psoralen, bergapten and xanthotoxin were found on and within fruits and seeds (Table 1). Xanthotoxin has been found as crystals on stored roots of parsnips [1]. We found that roots of three-month-old seedlings of Ruta graveolens, Pastinaca sativa and Angelica archangelica had xanthotoxin and bergapten on their surface. When seedlings of R. gra-veolens were irradiated by 366 nm UV, the roots reacted by lowering the total concentration of bergapten, xantho-toxin and imperatorin by over 50%. Interior concentra-
Table 1. Concentrations (μg/g fresh weight )of psoralen, bergapten, and xanthotoxin on and inside fruits or seeds.Species On surface Inside + surface % extruded on the surfaceUmbelliferae Angelica archangelica 14±0.7 2564±150 0.54Heracleum lanatum 20±1.2 1270±60 1.6Pastinaca sativa 40±2.0 3240±180 1.2Apium graveolens 15±0.7 74±3.1 20Pimpinella anisum 1.3±0.06 5.4±0.2 25Foeniculum vulgare 1.25±0.06 5.75±0.24 22Daucus carota 0.8±0.04 2.05±0.07 43Leguminosae Psoralea botuminosa 40±1.9 1390±70 3Rutaceae Evodia danielli 6.4±0.4 236.4±2.8 2.8Phellodendron chinensis 0.70±0.45 -- not avaibleRuta graveolens 0.039±0.002 10.0±0.7 0.38
Table 2. Concentration of furanocoumarins on and inside the roots of Pastinaca sativaSurface Interior % on surfacenot treated keptin
Cukeptin H2O
not treated
keptin Cu
keptin H2O
not treated keptin Cu
keptin H2O
ImperatorinBergaptenXanthotoxin
0.301.409.50
0.100.684.70
0.201.406.3
6.7016.088.0
21.519.0324
5570380
4.38.09.8
0.463.41.4
0.362.01.6
Session 4. Biotechnology and phytochemistry, environmental interactions

Vol. 56 63Bioactive Plant Compounds — Structural and Applicative Aspects
tions did not change, but surface concentrations of xan-thotoxin and bergapten declined several fold. When A. angelica seedlings (unpublished data) were irra-diated by 366 nm UV the opposite reaction was observed: total concentration doubled, with drastically increased extrusion and a parallel decline of interior concentra-tions. These two species reacted differently: thus, Ruta de-creased extrusion and maintained interior concentration, but Angelica increased extrusion and decreased interior concentration. In Ruta something as yet unexplained hap-pened to these compounds in the soil, where they were released immediately from the root surface. In Angelica higher deposits observed at first on the root surface re-mained there, but ultimately, of course, they would have been released into the soil. Roots of other plants were found to extrude scopoletin under stress conditions (un-published data). When such compounds are extruded to the surface of roots they have the potential to become in-volved in communication between plants, and in the rela-tionships among plants, soil, insects and microbes. Con-centration of furanocoumarins on and within the roots of Pastinaca sativa is shown in Table 2. References: Ceska O et al (1986) 42: 1302-11304. Harborne JB (1985). Ann Proc Phytochem Soc Europe 25: 393-408.
4.2
Composition of eggplant (Solanum melongena L.) leaf surface waxes as a factor affecting aphid feeding behaviour
Łukasz Haliński, Janusz Szafranek, Piotr Stepnowski
Department of Environmental Analysis, Faculty of Chemistry, University of Gdańsk, Sobieskiego 18, 80-952 Gdańsk, Polande-mail: Łukasz Haliński <[email protected]>
Eggplant (Solanum melongena L.) is an important vegeta-ble crop grown in tropics and subtropics. Due to its sus-ceptibility to a number of herbivores, feeding of a broad range of insect species inflicts heavy damage on the crops. As an alternative to the pesticide-based methods of eggplant protection, breeding of new lines and cultivars in respect to the natural plant resistance is of the highest importance. The profile of plant’s secondary metabolism is a feature that should be taken into account while exam-ining sources of plant resistance. As metabolites present on the surface of the plant (so-called cuticular or surface waxes) are the very first barrier, protecting plant from en-vironment, wax composition is often one of the factors, that influence insect settling and feeding behaviour. Plant colonisation and feeding behaviour of aphids (Myzus per-sicae and Macrosiphum euphorbiae) in relation to the total amount and chemical composition of eggplant leaf sur-face waxes were studied. Some morphological character-istics of plants were also taken into account. Additionally, the importance of the same factors in plant resistance to aphid feeding was analysed in case of the wild eggplant ancestor (Solanum incanum L.) in order to estimate the im-pact of domestication on the efficiency of plant resistance mechanisms. The objective of this study was, therefore, to evaluate the importance of eggplant’s leaf surface chem-istry in plant’s resistance to aphid feeding. Preliminary results presented here include biological observations and chemical analyses of waxes from nine eggplant culti-vars and one accession of the wild S. incanum. Plants were grown from seeds in greenhouse. Waxes from 15-week-old plants were extracted using organic solvent, fraction-ated by flash chromatography on silica gel and analysed by means of gas chromatography (GC) and GC coupled to mass spectrometer (GC-MS). Degree of aphid infesta-tion was estimated on the basis of number of insects on plants during the vegetation, changes in leaf colour and the presence of aphid honey-dew on the surface of the leaves. Results of quantitative analyses were processed using cluster analysis and principal component analysis. The most abundant wax fraction was composed of long-chain hydrocarbons, which accounted for approximately 80–90% of all compounds. Minor quantities of fatty ac-ids, aliphatic and triterpene alcohols, sterols and also trit-erpenyl, steryl and wax esters were detected. The most aphid-resistant plants (S. incanum and S. melongena cv. Black Beauty) were characterised by the highest total wax load (up to 16 μg cm–2), while the susceptible egg-plant cultivars had the lowest leaf wax content (8–11 μg cm–2). Resistant plants had also the highest amounts of cuticular triterpenes (430–710 ng cm–2), while the suscep-

64 2009Abstracts
tible cultivars were characterised by the lowest content of those compounds (10–110 ng cm–2). Plants that were moderately susceptible had an intermediate content of both triterpenes and total waxes. The high alkane and triterpene contents were negatively correlated with the average leaf area (ALA). The most resistant plants had the lowest ALA values and the most susceptible — the highest ones. The leaf area was the most important factor affecting triterpene content and quite important in case of hydrocarbons. The amounts of other wax components were not correlated with the leaf area. Relative composition of cuticular sterol fraction was found to be useful as a biochemical marker of insect-related plant damage. Degree of aphid infestation was positively correlated with the contribution of cholesterol to the total amount of cuticular sterols. Stigmasterol and β-sitosterol were main cucticular sterols in resistant and moderately susceptible cultivars, while cholesterol prevailed in frac-tion in case of plants strongly affected by aphids. How-ever, cholesterol was a minor sterol even in susceptible plants if they were grown in absence of insects.The similarity of S. incanum and the most resistant culti-vars of eggplant suggest the importance of the primitive traits in insect resistance. Low value of average leaf area and high content of triterpenes and hydrocarbons are possibly such primitive characteristics of species studied, which were lost in much extent during eggplant’s domes-tication. The presence of such traits in wild species and some eggplant cultivars may be useful in obtaining more resistant lines of eggplant and other Solanum crop spe-cies.
4.3
Phenolic plant compounds as the growth substrates in cometabolic degradation of xenobiotics by microorganisms
Izabela Greń, Danuta Wojcieszyńska, Urszula Guzik, Sylwia Łabużek
Department of Biochemistry, Faculty of Biology and Environmental Protection, Silesian University in Katowice, Jagiellońska 28, 40-032 Katowice, Polande-mail: Izabela Greń <[email protected]>
Xenobiotics of different chemical structure enter the en-vironment mainly as a result of human activity. Large group of xenobiotics constitutes aromatic compounds with hydroxyl group in the aromatic ring. Presence of hy-droxyl constituent increases solubility of aromatic hydro-carbons and makes their degradation by microorganisms easier. In the presence of hydroxyl group the electrophilic substitution of the sequential constituent(s) is facilitated, especially in the ortho and para positions of the aromatic ring. In order to carry out the hydroxylation of aromatic ring microorganisms exploit monooxygenases [EC 1.14.X.X], which catalyse this reaction in the presence of oxy-gen and NAD(P)H (Ulrich & Hofrichter, 2007). Structure of catechol or hydroquinone is indispensable for activ-ity of dioxygenases. This group of enzymes cleaves the ring destroying aromatic structure. Apart from catechol and hydroquinone dioxygenases [EC 1.13.11.X], in the cells of microorganisms protocatechuate and/or gentisate dioxygenase can be induced. Synthesis of the dioxygen-ases usually takes place in the presence of inducer (Vail-lancourt et al., 2006; Khomenkov et al. 2008). Toxicity of many xenobiotics of phenolic structure is increased by the presence of additional constituents such as e.g. chlo-rine or nitro- groups. Their number and position in the aromatic ring have large influence on xenobiotics’ biode-gradability as well as vitality of microorganisms in their surroundings. Lack of xenobiotic biodegradation can be due to absence of the appropriate enzymes induction or deficiency in reduced coenzymes. Weakened prolifera-tion of microorganisms in the presence of toxic xenobiotic can also influence on degree of its disappearance. Reso-lution of this problem in the environment can be appli-cation of cometabolic conditions for xenobiotic removal. Introduction of additional source of carbon and energy can facilitate the proliferation of microorganisms and the synthesis of essential coenzymes. If the chemical struc-ture of growth substrate is similar to xenobiotic one, it can also determine the induction of useful enzymes. So far the most often applied growth substrate in cometabolic deg-radation of xenobiotic was phenol (Hill et al., 1996; Hao et al., 2002; Leahy et al., 2003). Unfortunately introduction of additional amount of phenol determines escalation of the environment pollution. Application of phenolic com-pounds of natural origin will not intensify the problem of environment pollution and could be solution to increased contamination of the environment by aromatic hydrocar-bons. The aim of this research was to verify which sec-ondary aromatic plant metabolites can serve as the only source of carbon and energy for environmental strain

Vol. 56 65Bioactive Plant Compounds — Structural and Applicative Aspects
Stenotrophomonas maltophilia KB2, and how the presence of 4-hydroxybenzoic acid influences on removal of chlo-rophenols and maintenance of cells growth.Material and methods: Strain Stenotrophomonas mal-tophilia KB2 (NCBI DQ230920) belongs to the Collection of Microorganisms, Department of Biochemistry, Faculty of Biology and Environment Protection, Silesian Univer-sity in Katowice (Guzik et al., 2009). Strain was kept on nutrient agar slopes at 4°C and systematically transferred to the new ones every month. Adaptations of strain KB2 to degradation of different aromatic plant compounds were carried out according to the procedures described previously (Guzik et al., 2009). Cometabolic cultures were prepared using cell suspension prepared as follows: colo-nies were picked from nutrient agar and transferred to a 50-ml conical flask containing 20 ml of mineral salt me-dium with 3 mM of phenol. After 24 hours of incubation the suspension of the cells was transferred to a 250-ml flask containing 80 ml of mineral medium with 3 mM of phenol. The cells were allowed to grow the next 24 hours, and after that cells were harvested by centrifugation (5000 × g at 4°C for 15 min) and washed with fresh sterile medi-um. Such prepared cells were used as an inoculum in the amount to give the final optical density about 0.2 or 0.4 (depending on the non-growth substrate) in absorbance scale at λ=600 nm. Solutions of chlorophenols or phenolic plant compounds were prepared in Millipore water or NaOH solution, depending on their solubility. Growth of the cultures was determined by the measure of absorb-ance at 600 nm. The aromatic substrates were determined by a Merck HITACHI HPLC with a LiChromospher® RP-18 column (4 × 250 mm) and a DAD detector (Merck HITACHI). The wavelength of detection, composition of elution solvent and the flow rate were developed sepa-rately for each aromatic compound.Results and discussion: Identification of natural sub-stances that support growth of Stenotrophomonas mal-tophilia KB2 strain is the first step in devising a practical bioremediation strategy for in situ treatment of chlo-rophenols-contaminated environments. In this research a screening assay was carried out to identify plant com-pounds that might support growth of KB2 strain cells.
4-Hydroxybenzoic acid, protocatechuic acid, benzoic acid, vanillic acid and catechol could serve as the source of carbon and energy for Stenotrophomonas maltophilia KB2 strain preincubated with phenol. Time of each aromatic compound degradation depended on the chemical struc-ture and type of degradative pathways involved in its degradation. The longest time of degradation was deter-mined for vanillic acid (20 hours), which biodegradation required activity of demethylase (Morawski et al., 2000). Phenol and catechol in the concentration of 3 mM were degraded about 5 hours. Degradation time of benzoic acid and its hydroxyl derivatives was slightly elongated to 6 hours. It was correlated with induction of another cleaving enzymes, 1,2-catechol and 3,4-protocatechuate dioxygenase, respectively, in comparison to the induction of 2,3-catechol dioxygenases in the presence of phenol and catechol (Guzik et al., 2009). Salicylic acid, syringic acid, gallic acid, gentisic acid, cinnamic acid, p-coumaric acid, ferulic acid, sinapic acid, caffeic acid, resorcinol, hy-droquinone, thymol, p-cymene, carvacrol, and eugenol were not transformed by strain KB2. Lack of the ability to degrade these aromatic compounds might be due to their chemical structure and/or shortage of appropriate enzymes engaged in degradation of phenylopropanoid compounds or chemicals devoid of hydroxyl constitu-ent in the aromatic ring. Although the highest doses of aromatic hydrocarbons degraded by Stenotrophomonas maltophilia KB2 strain exceeded 3 mM (Guzik et al., 2009), in the cometabolic cultures that concentration of growth substrates were verified. When non-growth substrate (cometabolite) and growth substrate have the similar chemical structure, the competition inhibition could take place, and transformation of cometabolite can be slower and less effective (Hill et al., 1996; Hollender et al., 2000; Hao et al., 2002; Leahy et al., 2003).Biotransformation of 2-chlorophenol (2-CP), 3-chloroph-enol (3-CP), 4-chlorophenol (4-CP), 2,4-dichlorophenol (2,4-DCP), 2,4,5-trichlorophenol (2,4,5-TCP), and pen-tachlorophenol (PCP) in the concentration of 1 mM in the presence of 4-hydroxybenzoic acid (4-HB) was deter-mined. In addition phenol (Ph) as a control was also used as the growth substrate. These isomers of chlorophenols
Table 1. Percentage of removal of non-growth and growth substrates during the cometabolic cultures of Stenotrophomonas maltophilia KB2 strain, preincubated with phenol after 6 hours, 24 hours, and 2 days (for monochlorophenols and growth sub-strates)/14 days (for DCP, TCP and PCP); the results are an average of three independent cultures; the results from the abiotic cultures were taken under consideration during recalculations.
Cometabolic cultures
% of substrate removalnon-growth substrate growth substrate6 hours 24 hours 2/14 days 6 hours 24 hours 2/14 days
2-CP + 4-HB 71 ± 12 74 ± 9 82 ± 12 77 ± 10 100 ± 0 -3-CP + 4-HB 61 ± 16 64 ± 2 65 ± 2 75 ± 17 100 ± 0 -4-CP + 4-HB 21 ± 3 88 ± 3 100 ± 0 0 ± 0 100 ± 0 -2,4-DCP+ 4-HB 0 ± 0 0 ± 0 13 ± 3 0 ± 0 0 ± 0 0 ± 02,4,5-TCP + 4-HB 0 ± 0 0 ± 0 0 ± 0 0 ± 0 0 ± 0 0 ± 0PCP + 4-HB 0 ± 0 0 ± 0 0 ± 0 3.7 ± 0.5 100 ± 0 -2-CP + Ph 71 ± 17 75 ± 9 83 ± 8 44 ± 13 89 ± 0 100 ± 03-CP + Ph 22 ± 4 23 ± 4 23 ± 5 2,5 ± 0,5 7 ± 1 7 ± 14-CP + Ph 40 ± 3 100 ± 0 - 44 100 -2,4-DCP+ Ph 0 ± 0 0 ± 0 9 ± 4 0 ± 0 0 ± 0 0 ± 02,4,5-TCP + Ph 0 ± 0 0 ± 0 0 ± 0 0 ± 0 0 ± 0 0 ± 0PCP + Ph 0 ± 0 0 ± 0 0 ± 0 10,7 ± 3 99 ± 1,7 100

66 2009Abstracts
didn’t serve as a source of carbon and energy for KB2 strain (data not shown).Complete degradation of non-growth substrate was ob-served only for 4-chlorophenol, both, in the presence of 4-HB and phenol. Remaining isomers of monochloroph-enols were only partially transformed, and position ortho of the aromatic ring with chlorine substituent seemed to be less toxic than position meta (Table 1) (Zaitsev et al., 1995). 4-Hydroxybenzoic acid supported the growth of Stenotrophomonas maltophilia KB2 cells and was completely metabolized within 24 hours. Presence of monochloroph-enols in the culture only elongated time course of its deg-radation. It was probably due to the induction of other set of enzymes, connected with protocatechuic acid degrada-tion (data not shown). Decomposition of phenol needed the enzymes previously induced, and their activities were considerable inhibited in the presence of 3-chlorophenol. Chlorine substituent in the meta position according to the hydroxyl group of the aromatic ring is thought to limit the space for monooxygenase activity (Fava et al., 1995; Farrell & Quilty, 2002). Presence of two or more chlorines in the aromatic ring almost complete inhibited degrada-tion of non-growth and growth substrates (Table 1). The exception of this principle is pentachlorophenol in the presence of that 4-hydroxybenzoic acid and phenol were degraded. It can be assumed that molecule of pentachlo-rophenol is much larger than di- or trichlorophenol (due to the number of substituent) and doesn’t account for the spherical obstacle to activity of enzymes involved in 4-HB and/or phenol biodegradation. 2,4-DCP and 2,4,5-TCP completely inhibited the degradation of growth sub-strates as well as the vitality of cells (data not shown). One reason of this toxic influence could be the applied dose of these chlorophenols’ isomers. Marsolek et al. (2007) showed that 2,4,5-trichlorophenol is strongly toxic in the concentration of 40 μM, and Smith & Beadle (2007) determined the toxic dose of 2,4-dichlorophenol for Pseu-domonas cepacia 2a to be 0.35 mM. 4-Hydroxybenzoic acid as the hypothetical phenolic plant compound could sup-port the growth of Stenotrophomonas maltophilia KB2 cells and facilitate transformation of chlorophenols but its ac-tivity was dependent on the structure of xenobiotic and its toxicity. References: Farrell A, Quilty B (2002) J Ind Microbiol Biotechnol 28: 316-324.Fava F et al (1995) Lett Appl Microbiol 21: 307-312.Guzik U et al (2009) Braz J Microbiol 40: 285-291. Hao et al (2002) Chemosphere 46: 797-807. Hill GA et al (1996) Appl Microbiol Biotechnol 46: 163-168.Hollender J et al (1997) World J Microb Biotech 16: 445-450.Khomenkov VG et al (2008) Appl Biochem Microbiol 44: 117-135.Leahy JG et al (2003) FEMS Microbiol Ecol 43: 271-276.Marsolek DM et al (2007) Biodegradation 18: 351-358.Morawski B et al (2000) J Bacteriol 182: 1383-1389.Smith ARW, Beadle CA (2007) Biodegradation 19: 669-681.Ulrich R, Hofrichter M (2007) Cell Mol Life Sci 64: 271-293.Vaillancourt FH et al (2006) Crit Rev Biochem Mol Biol 41: 241-267.Zaitsev GM et al (1995) Appl Environ Microbiol 61: 4191-4201.
Acknowledgments:This work was supported by the Ministry of Science and Higher Education, project No 3626/B/P01/2007/33. Authors would like to thank Agnieszka Materna MSc. for the excellent technical as-sistance.
4.4
Evaluation of antifungal activity of pentacyclic triterpenoids isolated from Calendula officinalis L.
Ewa Wiktorowska, Marek Długosz, Wirginia Janiszowska
Department of Plant Biochemistry, Institute of Biochemistry, University of Warsaw, Miecznikowa 1, 02-096 Warsaw, Poland e-mail: Ewa Wiktorowska <[email protected]>
Calendula officinalis L., marigold (family Asteraceae), an an-nual plant has been used in traditional medicine for the treatment of various diseases and ailments since ancient times. Marigold contains two series of oleanolic acid (3β-hydroxy-olea-12-en-28-oic acid, OA) (Fig. 1) glycosides, namely „glucuronides” (derivatives of 3-O-monoglucuro-nide F) [1] and „glucosides” (derivatives of 3-O-monoglu-coside I) [2]. Pentacyclic triterpenoids are common phyto-chemicals, which are naturally occur in many medicinal plants and represent one of the most active class of com-pounds posses a wide spectrum of biological activities. Oleanolic acid glycosides isolated from marigold exhibit allelopathic, hemolytic, antibacterial and antiparasitic ac-tivities [3, 4]. However, no information is available about the antifungal activity of these compounds. Therefore, the aim of the present study was to evaluate the anti-fungal activity of pentacyclic triterpenoids isolated from C. officinalis against some plant pathogenic fungi. Materials and methods: Five fungi isolates (Alternaria brassicicola, Trichoderma viride, Fusarium oxysporum, Mu-cor hiemalis and Aspergillus niger) were obtained from the Institute of Fermentation Technology and Microbiology, Technical University of Lodz. For the antifungal bioassay, the required amount of triterpenoids (oleanolic acid, 3-O-
Figure 1. Structure of oleanolic acid and its glycosides from C. officinalis (Glur — glucuronic acid, Gal — galactose, Glu — glucose).
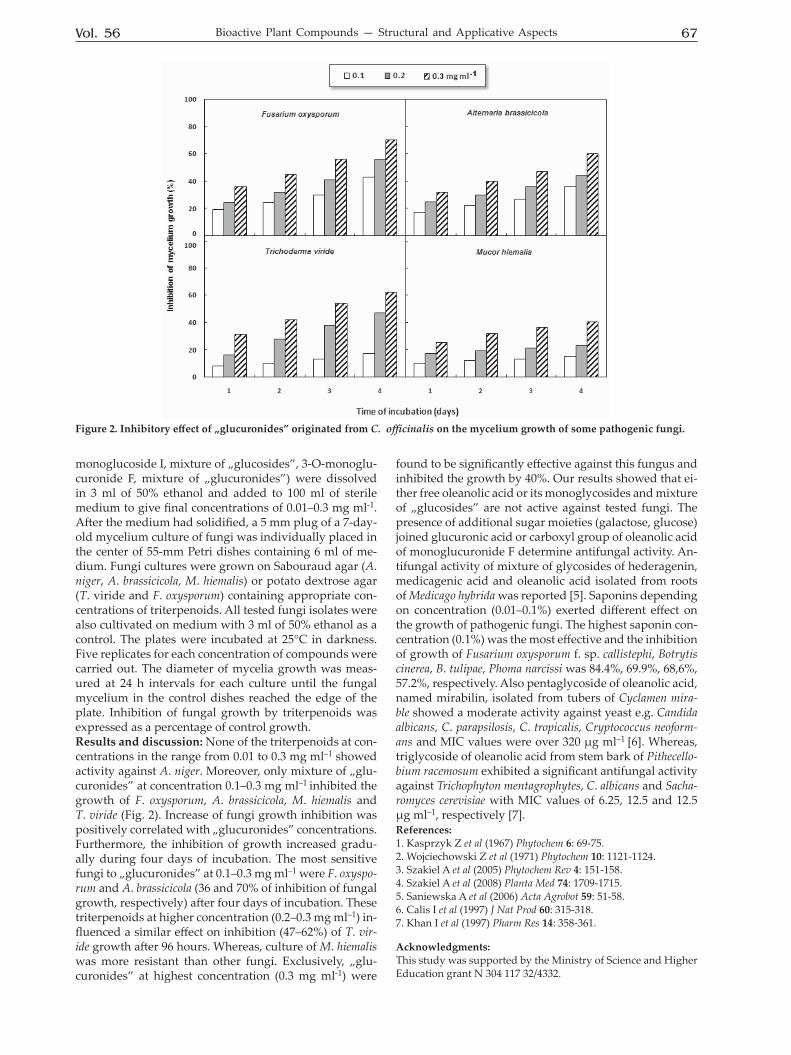
Vol. 56 67Bioactive Plant Compounds — Structural and Applicative Aspects
monoglucoside I, mixture of „glucosides”, 3-O-monoglu-curonide F, mixture of „glucuronides”) were dissolved in 3 ml of 50% ethanol and added to 100 ml of sterile medium to give final concentrations of 0.01–0.3 mg ml-1. After the medium had solidified, a 5 mm plug of a 7-day-old mycelium culture of fungi was individually placed in the center of 55-mm Petri dishes containing 6 ml of me-dium. Fungi cultures were grown on Sabouraud agar (A. niger, A. brassicicola, M. hiemalis) or potato dextrose agar (T. viride and F. oxysporum) containing appropriate con-centrations of triterpenoids. All tested fungi isolates were also cultivated on medium with 3 ml of 50% ethanol as a control. The plates were incubated at 25°C in darkness. Five replicates for each concentration of compounds were carried out. The diameter of mycelia growth was meas-ured at 24 h intervals for each culture until the fungal mycelium in the control dishes reached the edge of the plate. Inhibition of fungal growth by triterpenoids was expressed as a percentage of control growth. Results and discussion: None of the triterpenoids at con-centrations in the range from 0.01 to 0.3 mg ml–1 showed activity against A. niger. Moreover, only mixture of „glu-curonides” at concentration 0.1–0.3 mg ml–1 inhibited the growth of F. oxysporum, A. brassicicola, M. hiemalis and T. viride (Fig. 2). Increase of fungi growth inhibition was positively correlated with „glucuronides” concentrations. Furthermore, the inhibition of growth increased gradu-ally during four days of incubation. The most sensitive fungi to „glucuronides” at 0.1–0.3 mg ml–1 were F. oxyspo-rum and A. brassicicola (36 and 70% of inhibition of fungal growth, respectively) after four days of incubation. These triterpenoids at higher concentration (0.2–0.3 mg ml–1) in-fluenced a similar effect on inhibition (47–62%) of T. vir-ide growth after 96 hours. Whereas, culture of M. hiemalis was more resistant than other fungi. Exclusively, „glu-curonides” at highest concentration (0.3 mg ml-1) were
found to be significantly effective against this fungus and inhibited the growth by 40%. Our results showed that ei-ther free oleanolic acid or its monoglycosides and mixture of „glucosides” are not active against tested fungi. The presence of additional sugar moieties (galactose, glucose) joined glucuronic acid or carboxyl group of oleanolic acid of monoglucuronide F determine antifungal activity. An-tifungal activity of mixture of glycosides of hederagenin, medicagenic acid and oleanolic acid isolated from roots of Medicago hybrida was reported [5]. Saponins depending on concentration (0.01–0.1%) exerted different effect on the growth of pathogenic fungi. The highest saponin con-centration (0.1%) was the most effective and the inhibition of growth of Fusarium oxysporum f. sp. callistephi, Botrytis cinerea, B. tulipae, Phoma narcissi was 84.4%, 69.9%, 68,6%, 57.2%, respectively. Also pentaglycoside of oleanolic acid, named mirabilin, isolated from tubers of Cyclamen mira-ble showed a moderate activity against yeast e.g. Candida albicans, C. parapsilosis, C. tropicalis, Cryptococcus neoform-ans and MIC values were over 320 μg ml–1 [6]. Whereas, triglycoside of oleanolic acid from stem bark of Pithecello-bium racemosum exhibited a significant antifungal activity against Trichophyton mentagrophytes, C. albicans and Sacha-romyces cerevisiae with MIC values of 6.25, 12.5 and 12.5 μg ml–1, respectively [7]. References: 1. Kasprzyk Z et al (1967) Phytochem 6: 69-75.2. Wojciechowski Z et al (1971) Phytochem 10: 1121-1124.3. Szakiel A et al (2005) Phytochem Rev 4: 151-158.4. Szakiel A et al (2008) Planta Med 74: 1709-1715.5. Saniewska A et al (2006) Acta Agrobot 59: 51-58.6. Calis I et al (1997) J Nat Prod 60: 315-318.7. Khan I et al (1997) Pharm Res 14: 358-361.
Acknowledgments:This study was supported by the Ministry of Science and Higher Education grant N 304 117 32/4332.
Figure 2. Inhibitory effect of „glucuronides” originated from C. officinalis on the mycelium growth of some pathogenic fungi.

68 2009Abstracts
4.5
Molecular markers for powdery mildew (Podosphaera macularis) resistance in hop
Marcin Przybyś, Urszula Skomra
Department of Plant Breeding and Biotechnology, Institute of Soil Science and Plant Cultivation – State Research Institute, Puławy, Polande-mail: Marcin Przybyś <[email protected]>
Hop powdery mildew caused by Podosphaera macularis is one of the most important fungal disease causing drastic losses of yield and quality in hops worldwide. Growing resistant cultivars is the most economical and environ-mentally safe approach to eliminate the use of fungicides and to reduce production losses due to this disease. So far, eight different resistance genes in hops have been described. The most stable in the field conditions is the resistance based on the R2 gene, derived from the English cultivar Wye Target. The object of the work were two hop progenies from the crosses between hop cv. Wye Target, used as a source of powdery mildew resistance and two susceptible male plants. In each progeny all individuals were tested for their powdery mildew resistance using artificial inoculation of leaves in Petri dishes in control-led conditions. Random amplified polymorphic DNA (RAPD) markers were used to determine of presence re-sistance gene. Until now 40 primers were tested and two RAPD markers closely linked to a gene for resistance to Podosphera macularis were identified. Work to identify other molecular markers to powdery mildew resistance genes is still in progress.
4.6
Insect feeding deterrent activity of salicyl alcohol derivatives from Picris strigosa
Wanda Kisiel1, Klaudia Michalska1, Gerard Nowak2 1Institute of Pharmacology Polish Academy of Sciences, Department of Phytochemistry, 31-343 Kraków, Poland; 2Poznań University of Medical Sciences, Department of Medicinal and Cosmetic Natural Products, 60-623 Poznań, Polande-mail: Klaudia Michalska <[email protected]>
From roots of hitherto not studied Picris strigosa M. Bieb. (Asteraceae) three known guaiane-type sesquiterpene lactones, derivatives of zaluzanin C, were isolated, to-gether with seven known low molecular weight pheno-lics methyl p-hydroxyphenylacetate, syringaldehyde, di-hydrosyringin, benzyl-O-β-glucopyranoside, helicin (1), salicin (2) and isosalicin (3), the latter being the major sec-ondary metabolite of the plant material. The compounds were characterized by direct comparison of their spectra and HPLC retention time values with those of reference compounds from our collection.This is the first time sal-icin has been found in plants of the family Asteraceae. Preparations containing salicin from willow bark are popular herbal remedies for the treatment of rheumatism and pain. The influence of the salicyl alcohol derivatives helicin (1), salicin (2) and isosalicin (3) on insect feeding was tested using the three selected stored product insect pests Sitophilus granarius, Tribolium confusum and Trogo-derma granarium. The compounds showed different ef-fects on the insects varying from good feeding deterrent activities (i.e. helicin and isosalicin for T. confusum larvae) to feeding stimulant activities, especially for T. granarium larvae. Aside from the taxonomic benefits, the results of our feeding studies also contribute to an understanding of the function of low molecular phenolic glycosides.

Vol. 56 69Bioactive Plant Compounds — Structural and Applicative Aspects
4.7
Biological activities of lyoniside isolated from rhizomes of Vaccinium myrtillus L.
Anna Szakiel1, Max Henry2 1Department of Plant Biochemistry, Faculty of Biology, University of Warsaw, Miecznikowa 1, 02-096 Warszawa, Poland; 2Structure et Réactivité des Systèmes Moléculaires Complexes (SRSMC), Nancy-Université, UMR7565 CNRS-UHP, 5 rue Albert Lebrun, BP: 80403, 54001, Nancy cedex, Francee-mail: Anna Szakiel <[email protected]>
Lignans are a class of secondary plant metabolites pro-duced by oxidative dimerization of two phenylpropanoid units. They constitute a group of important plant phenols possessing a diverse spectrum of biological activities, in-cluding potent antimicrobial, antifungal, antiviral, anti-oxidant, insecticidal and antifeeding properties. They may also participate in the regulation of plant growth and development. Furthermore, the great significance of lignans is due to their possible application in the fields of pharmacy and nutrition utilizing their antitumor, antiin-flammatory and immunosuppressive effects. Lignans are widely distributed in the plant kingdom and have been found in species belonging to more than seventy families of pteridophytes, gymnosperms and angiosperms. So far several hundreds of lignans, occurring in wooden parts, roots, leaves, flowers, fruits and seeds, have been identi-fied (Slanina & Glantz, 2004; Willför et al., 2006). Lyoniside (Fig. 1) is a phenyltetralin lignan glycoside which was first isolated and identified as a major com-ponent of the wood of Lyonia ovalifolia (Ericaceae) (Yasue 1960) and afterwards its occurrence was also presented in other plants of various families. It was isolated mainly from woody parts (inner and stem barks, roots) but occa-sionally also from other organs, for example semperviv-ent leaves of Eriobotrya japonica (Jiang & Xuan, 2006). Re-cently for the first time we demonstrated the occurrence of this compound in bilberry, Vaccinium myrtillus (Szakiel et al., 2008). Bilberry is a plant abounded in phenolics found in all organs including leaves and fruits. How-ever, the occurrence of lyoniside is restricted only to the winter-persistent part of the plant, namely aboveground stems and underground rhizomes. Exclusive occurrence in winter-persistent organs suggests the role in chemical defence against pathogens and herbivores, protection against UV-radiation and allelopathic activity if leakage or exudation to the soil is possible. The aim of the present work was to investigate the antifungal and allelopathic properties of lyoniside. Methods: Isolation of lyoniside. Air-dried and crushed V. myrtillus rhizomes were extracted preliminarily with diethyl ether and afterwards with 70% ethanol in Soxh-let apparatus during 6 h. Obtained ethanolic extract was fractionated by droplet countercurrent chromatogra-phy. DCCC chromatograph was equipped with 95 tubes (400 × 2 mm) and operating with the solvent system CHCl3/MeOH/H2O (43:37:20, v/v) in the descending mode. 370 fractions of 7 ml each were collected during 40 h. The col-lected fractions were analysed by TLC (silica gel, CHCl3:
MeOH:H2O 61:32:7). Fractions containing compounds with similar Rf were combined. Fractions 50–71 contained pure lyoniside precipitating after concentration. Antifungal bioassays. Fungi strains, obtained from the Institute of Fermentation Technology and Microbiology, Technical University of Łódź, were grown on Sabouraud agar (Aspergillus niger, Alternaria brassicicola and Mucor hiemalis) or potato dextrose agar (Fusarium oxysporum and Trichoderma viridae). 55 mm Petri dishes filled with 6 ml of sterile medium containing lyoniside at concentrations of 20 and 50·μg·ml–1 were inoculated with 5 mm mycellium plug and incubated at 25°C in darkness for 5 days. Three replicates for each concentration were carried out. The ra-dial growth of mycelium (diameter of colony) was meas-ured after 1.5, 3 and 5 days for each culture and compared with control essays without lyoniside. Allelopathic bioassays. Sheets of Whatman No.1 filter paper were placed in 100 mm diameter Petri dishes and impregnated with 10 ml of ethanol containing lyoniside at concentration of 10 μl·ml–1. Solvent was evaporated and 30 seeds of tested plants: lettuce Lactuca sativa, cress Lepidium sativum, pine Pinus sylvestris, spruce Picea abies or larch Larix decidua were distributed evenly on the pre-pared sheets moisturized with 10 ml of pure water. For each plant control dishes were prepared without lyoni-side. Afterwards, all dishes were closed and placed in the dark in a thermostat (22oC). Germinating seeds of lettuce and cress were counted after 3 days and those of pine, spruce and larix after 7 days; the length of roots and hy-pocotyls was measured after 7 or 15 days (respectively) of experiment. Results and discussion: Fungicidal activity of lyoniside against 5 strains of Ascomycota icluding moulds Aspergil-lus niger and Trichoderma viride as well as phytopathogens Alternaria brassicicola, Fusarium oxysporum and Mucor hie-malis was investigated (Table 1). Lignans are potent an-timycotics influencing fungal cell synthesis (Hwang et al., 2007), nevertheless, various strains differ appreciably in their sensitivity to particular compounds. Mycelial growth of all tested fungi was markedly influenced by lyoniside, however, with different dynamics of inhibition. The growth of two moulds, A. niger and T.viride was al-most completely inhibited at the beginning of incubation (A. niger by 100%, T. viride by 96% at lyoniside concen-tration of 50 μl·ml–1 during first 3 days). Afterwards, the mycelium spread rapidly reaching almost half of diam-eter of dish during subsequent 2 days. Moreover, when the experiment was continued for longer time (data not shown), finally both moulds developed full colonies on the whole dish surface. In turn, the inhibition of growth of M. hiemalis was strong during first 3 days (92%), whereas it was weaker in the case of two other phytopathogens
H
OH
O O
OH
OCH3
H3CO
HO
H3CO
OCH3
HOOH
OH
Figure 1. The structure of lyoniside (9-O-β-D-xylopyranosyl(+)lyoniresinol).

70 2009Abstracts
A. brassicicola and F. oxysporum (76 and 78%, respective-ly). However, after 5 days of experiment the inhibitory ef-fect was still significant, equalling 64% for A. brassicicola, 80% for M. hiemalis and 78% for F. oxysporum. Prolonga-tion of experiment did not change these results, therefore lyoniside can be regarded as an antifungal agent active against tested phytopathogens. Allelopathic activity of lyoniside was estimated as the influence on seed germination and subsequent seedling growth of commonly used dicotyledon model plants (let-tuce and cress) but also on conifer trees (pine, spruce) which occur in forests with bilberry as dominant un-derstorey, and larch which is not the natural neighbour of bilberry in its environment. Applied concentration of lyoniside was comparable to that detected in the soil obtained from bilberry habitat. The results are shown in Table 2. Lyoniside exerted moderatory inhibiting effect on the germination of lettuce (34%) and even weaker on cress (18%), however, it strongly suppressed the seedling
root growth of both plants by 75%. With other phenolics, lignans are often regarded as effective allelochemicals ac-cording to their capacity of interference with all sorts of proteins: enzymes, transporters, ion-channels, receptors, thus being able to disturb the transport of compounds and biosynthetic pathways in target plants (Wink 2003). Among tested conifer trees, the strongest inhibitory effect was exerted on larch, both on germination (55%) and root and hypocotyl growth (50 and 46%, respectively). The in-fluence on pine and spruce was not so well expressed, with the weakest inhibition shown against pine (germi-nation by only 15%, growth of root by 22%, hypocotyl by 25%). The effects on spruce were stronger (germina-tion by 39%, growth by 32 and 35%) but less than that on larch. These results show that lyoniside alone cannot be responsible for known allelopathic potential of bilberry exerted against pine and spruce. In natural habitats re-lationships between plants are very complex due to the long-term coevolution and cannot rely exclusively on the
Table 1. The influence of lyoniside on mycelial growth of tested fungi. The growth of mycelium is measured as diameter of colony without inoculation plug. Values are means of 3 replicates ±S.D.
Fungi Lyoniside concentra-tion [μg·ml–1]
Diameter of colony [mm] Inhibition [%]Days of incubation
1.5 3 5 1.5 3 5Aspergillus niger control 15 ± 1.6 40 ± 1.10 50
20 4 ± 0.52 10 ± 0.95 33 ± 1.68 73 75 4450 0 0 19 ± 1.23 100 100 62
Trichoderma viride control 21 ± 2.05 50 5020 4 ± 0.90 6 ± 1.08 29 ± 3.05 81 88 4250 1 ± 0.2 2 ± 0.75 20 ± 1.86 95 96 60
Alternaria brassicicola control 35 ± 1.98 50 5020 2 ± 0.82 16 ± 1.28 21 ± 1.45 94 68 5850 0 12 ± 1.42 18 ± 1.98 100 76 64
Fusarium oxysporum control 14 ± 1.86 32 ± 3.78 5020 10 ± 1.04 18 ± 1.75 28 ± 2.06 29 44 4450 6 ± 1.25 7 ± 1.03 11 ± 1.63 57 78 78
Mucor hiemalis control 40 ± 2.36 50 5020 12 ± 1.46 20 ± 2.02 25 ± 3.78 70 60 5050 2 ± 0.10 4 ± 0.68 10 ± 2.10 95 92 80
Table 2. The influence of lyoniside on germination and growth of tested plants. Values are means of 3 replicates ±S.D.
Plant/compoundGermination Seedling growth
Number of germinating seeds
Inhibition (%)
Root length [mm]
Inhibition (%)
Hypocotyl length [mm]
Inhibition (%)
Lettucecontrol 29 ± 1 22.83 ± 2.01 10.67 ± 2.15lyoniside 19 ± 2 34 5.71 ± 2.24 75 8.14 ± 2.87 24Cresscontrol 28 ± 2 38.13 ± 2.05 24.5 ± 1.55lyoniside 23 ± 1 18 9.6 ± 0.74 75 10.4 ± 3.70 58Pinecontrol 26 ± 3 5 ± 2.8 2 ± 0.36lyoniside 23 ± 2 15 3.9 ± 0.63 22 1.5 ± 0.1 25Sprucecontrol 23 ± 2 6.28 ± 1.6 2.84 ± 0.62lyoniside 14 ± 1 39 4.24 ± 2.35 32 1.85 ± 0.2 35Larixcontrol 22 ± 2 7.12 ± 1.26 3.08 ± 0.89lyoniside 10 ± 1 55 3.54 ± 1.47 50 1.65 ± 0.32 46

Vol. 56 71Bioactive Plant Compounds — Structural and Applicative Aspects
action of one compound or even one type of compounds (Jha et al., 2006; Malik & Pellisier, 2000). Indeed, recently we demonstrated the synergism of lyoniside with trit-erpene acids leading to inhibition of gemination of pine seed by 80% and suppression of seedling growth by 70% (Szakiel & Henry, 2008). On the other hand, the strong ef-fect exerted on larch, not being the natural environmental neighbour of bilberry, shows that lyoniside is an effective allelochemical when it influences the plant without evo-luted mechanisms of tolerance or detoxification. The presented results show that lyoniside, possessing an-tifungal and allelopathic activities, plays significant role in chemical protection against phytopathogens and envi-ronmental interactions of bilberry. References: Hwang EI et al (2007) Planta Med 73: 679-682. Jha S, Jha PK, Gewali MB (2006) Pak J Pl Sci 12: 103-113. Jiang LL, Xuan LJ (2006) Chinese Chem Lett 17: 35-37. Mallik AU, Pellisier F (2000) J Chem Ecol 26: 2197-2209. Slanina J, Glatz Z (2004) J Chromatography B 812: 215-229. Szakiel A, Henry M (2008) Planta Med 74: 980. Szakiel A, Voutquenne-Nazabadioko L, Henry M (2008) Planta Med 74: 1065. Willför SM, Smeds AI, Holmbom BR (2006) J Chromatography A 1112: 64-77. Wink M (2003) Phytochemistry 64: 3-19. Yasue M (1960) Yakugaku Zasshi 80: 1013.
4.8
Rosmarinic acid accumulation in the cell suspension culture of Eryngium planum L.
Małgorzata Kikowska1, Barbara Thiem1, Aldona Krawczyk2
1Department of Pharmaceutical Botany and Plant Biotechnology, Poznań University of Medical Sciences, Św. Marii Magdaleny 14, 61-861 Poznań, Poland; 2Quality Control Laboratory of Phytopharm Klęka S.A., Klęka 1, 63-040 Nowe Miasto nad Wartą, Polande-mail: Małgorzata Kikowska <[email protected]>
Abstract: The objective of this report was to establish the cell suspension culture and demonstrate the presence and varied concentration of rosmarinic acid in the suspension culture growth phases. This paper is also focus on the ef-fect of methyl jasmonate on RA accumulation. The phe-nolic acids presence was established by RP HPLC. Introduction: Eryngium planum L., a species that belongs to the Apiaceae family and to the Saniculoideae subfamily, is used in folk medicine in Europe. The presence of the active constituents found in most of Sea Holly species in-cluding phenolic acids (rosmarinic acid – RA, chlorogenic acid – ChA) (Fig. 1), triterpene saponins, flavonoids, es-sential oils and coumarins determines their multidirec-tional pharmacological activity: diuretic, expectorant and antimycotic (Duke et al., 2002). Rosmarinic acid, repre-senting one of the most common caffeic esters in plants, is supposed to act an important role in defense mecha-nism against pathogens. RA is found in many species of the Lamiaceae, Boraginaceae and other families (including Apiaceae). RA has many biological activities: antioxida-tive, distinct antiseptic, antiviral, antibacterial, antiphlo-gistic, anti-inflamatory, which make it an important factor for the health promoting effects and a valuable product for cosmetic and food industries (Petersen et al. 1994; Le Claire et al., 2005; Park et al., 2008). Due to the presence of a high number of hydroxyl groups and a carboxyl moi-ety, RA and ChA have antioxidant activity (Matkowski, 2008). Elicitation with methyl jasmonate (MeJA) has been used to increase the accumulation of RA in plant tissue and cell cultures (Szabo et al., 1999). The jasmonates are possibly signal compounds in the elicitation process in-ducing transcriptional activation of genes involved in de novo formation of secondary metabolites (Yukimune et al., 1996). In our previous studies, it was shown, that the phenolic acids were accumulated in in vitro cultures of E. planum (Thiem et al., 2009). Numerous studies have been carried out using plant cell suspension cultures as a po-tential source of valuable constituents. Some difficulties correlated with low or varied concentration in the intact
Figure 1. Chemical structures of RA and ChA.
OH
OH
O
OH
OHO H
O
OH
OOH
OH
O
HOOC
OH OH
OH1
23
45
6
rosmarinic acid chlorogenic acid (5-O-caffeoylquinic acid)

72 2009Abstracts
plants can be overcome by using plant cell biotechnology. The use of cell suspension cultures gives advantages such as shorter cycles of growth, the possibility of optimising the processes of secondary metabolites accumulation and the maintenance under controlled conditions. The main problem is that the desired compound usually occur in a higher concentration in a short period of the culture growth. For that reason the shake-flask suspensions are analyzed in terms of assigning the proper phase over the course of typical growth cycle suitable for biotechnologi-cal methods applying. Material and Methods: Plant cell suspension culture. Callus culture of E. planum was initiated from seedling’s root explants by culturing it on MS basal medium (Mu-rashige & Skoog, 1962) supplemented with 1.0 mgl–1 2,4-D and 1.0 mgl–1 BAP. The 2/4/6 g of stabilized, friable callus was sieved through stainlesssteel sieve into 50 ml of fresh medium (MS) containing 0.5 mgl–1 2,4-D. Liquid suspension culture was established by continuous shak-ing at 110 rpm in the darkness, and maintained by sub-culturing at regular intervals of 8-10 days. The 5 ml inocu-lation of old culture was added to 50 ml of new medium to maintain cell vitality. The experiment was set up in a completely randomized design. Addition of MeJA. The filter-sterilised solution of MeJA (Sigma-Aldrich) in 96% ethanol was added to the culture medium resulting in a final concentration of 100μM on the day of linear phase of the cultivated period. Control experiment was run with only ethanol. Dry biomass The growth of cell suspen-sion was monitored by measurement of the fresh and dry biomass. For this aim the exact amounts of fresh biomass were dried in a dryer (60oC for 1h, followed by 120oC for 24h) to constant weight. Sample preparation procedure / RA determination. The 0.2 g of dried and powdered sus-pension biomass were extracted three times with 50% (v/v) methanol for 30 min at the boiling point temperature of the extractive mixture under a reflux. Cooled and filtered extract was next diluted with the methanol 50%(v/v) up to the volume of 50 or 100 ml. The solution was filtred through 0.2 μm filter (Schleicher & Schuell) and 10μl was analyzed by HPLC. The RA content in the metanolic extracts was determined by RP HPLC analysis on a Li-Chrospher 100 250×4 mm reversed phase column (RP 18e, 5 μm, Merck). The solvent system was a linear gradient of acetonitrile/H2O and phosphoric acid pH=2.2; ACN from 15% to 60% for 40 min, 60% for 15 min, from 60% to 15% for 1 min and 15% for 9 min. The flow rate was 1 ml/min and the effluent was monitored by UV detection at 320 nm. The identity of RA and ChA in the chromatograms were verified by comparison with pure standards. Results: Suspension culture presented a typical growth curve divided into four phases: lag, exponential, linear and stationary. A growth of cell population inoculated into a fresh medium did not ensue immediately. The cells were taken from the exponential phase so it is seem that the lag phase was determined to be brief. The cells reached the linear phase, performing the multiplication in geometric progression. At the and of the culture, cell suspension reached the stationary phase. Preliminary RP HPLC investigations have shown that extracts from cell suspension culture contain phenolic acids, mainly ros-marinic acid. The concentration of RA varied depending on the phases of the cycle. The highest concentration of
RA was found in 3-9 days of linear phase of suspension culture and decreased when its reached the stationary phase. In the most favourable phase the productivity of RA attained 0.093 g/100g of DW (Fig. 2). The amounts of RA increased significantly after elicitation with 100μM MeJA (Fig. 3). Content increased 1.69 and 2.21 — fold af-ter elicitation for 24 and 48h respectively, compared with the control. In contrast, the amount of ChA decreased with the addition of MeJA.Disscusion: Numerous studies have been carried out on RA accumulation in cell suspension culture in higher con-tent than in intact plants e.g.: Coleus blumei, Anchusa of-ficinalis, Eritrichum sericeum, Lithospermum erythrorrhizon, Lavandula vera, Ocimum basilicum, Salvia officinalis, Zataria multiflora (Matkowski, 2008). The first results showing the important quantitative and qualitative changes in RA accumulation were obtained from cell suspension cul-tures of Salvia officinalis. The larges RA production was reported for Coleus blumei cell cultures (Hippolyte et al., 1992). Rosmarinic acid production can be correlated with growth or begins only when growth has stopped. In our studies RA accumulation was enhanced during linear phase of the cell growth cycle. The first days are required to recover from stress of being transferred to fresh me-dium. The addition of MeJA (12th day) affected the RA increase in cell suspension culture compared with the control cell culture. Similarly, Ketchum (1999) suggested that the greater effectiveness of the elicitor on Taxus cell culture was achieved when the MeJA was added 7, 8 or 14 days of inoculation, rather than at time of inoculation. In conclusion, RA biosynthesis was stimulated in E.planum cell suspension culture by the presence of MeJA in the liquid medium. Our results show that the specific me-tabolites in plants can be modulated by an elicitor addi-tion to the stabilized and monitored line of cultured cells. Besides of the presence of rosmarinic acid in low content in cell biomass of E. planum, its activity can be correlated
Figure 2. HPLC chromatogram of RA and ChA in linear phase of non-elicited cell suspension culture.
Figure 3. Content of RA in elicited and non-elicited cell sus-pension cultures.
Minutes0 5 10 15 20 25 30 35 40
mAU
0
10
20
30
mAU
0
10
20
30
chlo
roge
nic
acid
rosm
arin
ic a
cid
Multi-Chrom 1 (1: 320 nm, 4 nm)Eryngium planum_5-Rep1
Name
%(m/m)0,09320342413,420RA
%(m/m)0,029775404,493ChA
UnitsAmountAreaTimeName
%(m/m)0,09320342413,420RA
%(m/m)0,029775404,493ChA
UnitsAmountAreaTimeName
00,05
0,10,15
0,20,25
0,30,35
0,4
without elicitation 24h of elicitation by (100µM) MeJA
48h of elicitation by (100µM) MeJA
cont
ent o
f RA
(g/1
00g)
Content of RA in cell suspension culture

Vol. 56 73Bioactive Plant Compounds — Structural and Applicative Aspects
with other bioactive compounds and make beneficial and health promoting effects. Acknowledgements:This work was supported by the Ministry of Science and Higher Education, Warsaw, Poland (grant no NN 405 065334) References:Duke JA et al (2002) Handbook of medicinal herbs. CRC Press 277-278.Hippolyte J et al (1992) Plant Cell Rep 11: 109-112.Ketchun REB et al (1999) Biotechnol Bioeng 62: 97-105.Le Claire E et al (2005) J Agric Food Chem 53:4367-4372.Matkowski A (2008) Biotechnol Adv 26: 548-560.Murashige T, Skoog F (1962) Physiol Plant 15: 473-97.Park SU et al (2008) African J Biotechnol 7: 4959-4965.Petersen M (1994) Plant Cell Tissue Organ Culture 38: 171-179.Park SU et al (2008) African J Biotechnol 7: 4959-4965.Szabo E et al (1999) Plant Cell Rep 18: 485-489.Thiem B et al (2009) PSE CSIC — Young Scientists Meeting, Los Narejos, Spain May 12th-14th
, 2009 Abstracts p.108.Yukimune Y et al (1996) Nat Biotechnol 14: 1129-1132.
4.9
Induction of hairy roots in Calendula officinalis capable of biosynthesising of oleanolic acid glycosides
Marek Długosz, Ewa Wiktorowska, Wirginia Janiszowska
Department of Plant Biochemistry, Institute of Biochemistry, Faculty of Biology, University of Warsaw, Miecznikowa 1, 02-096 Warsaw, Polande-mail: Marek Długosz <[email protected]>
Marigold (Calendula officinalis L.) seems to be one of the plants most exploited by pharmaceutical, cosmetic and food industries. The richness and diversity of biological active compounds that have been produced by marigold make this plant very attractive object for biotechnologi-cal application. Among these compounds pentacyclic triterpenes, i.e. oleanolic acid and his glycosides are fun-damental for numerous pharmacological properties: anti-inflammatory, anti-HIV, anti-tumor, cytotoxic, anti-bacte-rial and hepatoprotective (Liu et al. 2005). Plant in vitro cultures, although many imperfections (e.g. somaclonal variation, low efficiency metabolite synthesis), provide great potential as a production system of these valuable plant metabolites. Especially hairy root culture thanks to genetic/biochemical stability and higher productivity offer substantial advantages over any undifferentiated cultures. Growth of hairy roots can be scaled up using bioreactors and hence they can be exploited for commer-cial production of valuable plant metabolites (Guillon et al. 2006). Material and methods: Plant material. Calendula offici-nalis mature embryos were excised from seeds and steri-lized with a mixture “Domestos”: sterile water 1:3, v/v for 20 min. After three washes with sterile water embryos were placed on MS medium (Murashige & Skoog 1962) with half-reduced concentration of macroelements solid with 0.7% agar for germination. Seedlings with a fully de-veloped cotyledons and hypocotyl were used for trans-formation.Transformation and establishment of hairy roots cul-ture. Wild type Agrobacterium rhizogenes strain ATCC 15834 was used for hairy roots induction. The bacterial cultures were grown in liquid YEB medium until OD = 1.5 was achived. Then cultures were centrifugated at 5000 rpm for 20 min and resuspended with ½ MS liq-uid medium to OD = 0.3. The cotyledons or hypocotyls dissected from the seedlings were placed in Petri dish containing 30 ml medium with bacteria for 30 min. After inoculation all explants were removed to solid ½ MS me-dium and co-cultivated for 3 days in the dark. Next the bacteria were washed away from the explants with ½ MS medium supplemented with 300 mg dm-3 Timentin. The explants were placed on solid ½ MS supplemented with Timentin (300 mg dm–3). Single hairy roots growing at the cut surfaces were separated from explants and placed at the fresh solid medium supplemented with antibiotic. Es-tablished lines were maintained on ½ MS solid medium and subcultured every 4–8 weeks in complete darkness at

74 2009Abstracts
24oC. The hairy root cultures were also maintained on ½ MS liquid medium on a shaker at 120 rpm.Oleanolic acid analysis. Homogenized roots were ex-tracted two times with boiling methanol, while the me-dium was extracted four times with n-butanol. Extracts were evaporated to dryness and hydrolyzed in the Kiliani mixture. OA was extracted four times with ethyl ether, purified by TLC and methylated. The amount of oleanolic acid methyl ester was measured by GLC chromatography (Janiszowska & Kasprzyk 1977).Results and discussion: First hairy roots were observed after two weeks from placing the explants on the medium supplemented with antibiotics. Reactions were identical for the cotyledons and hypocotyls (Fig. 1) of seedlings: at the place of cutting the single root was sprouted. In some cases of cotyledons, two roots growing from lump of callus were observed. Identification of such roots as a single or two independent transgenic event was difficult. Roots 3 to 4 cm in length were cut off and removed on new plates separately and were marked as a line derived from single independent transgenic event (Fig. 2). So eve-ry explants gave at maximum two independent cultures of hairy roots.
In two independent experiments higher transformation efficiency of hypocotyls (66.6%), than cotyledons (33.8%), were initially observed. After separation roots from pri-mary explants and replacement to the fresh media 28 lines from cotyledons and only 13 lines from hypocotyls were obtained. Such situation could result from an aux-ins efflux after tissue cutting from the hypocotyls roots incorrectly classified as hairy roots. The dominance of cotyledons over hypocotyls for transformation efficiency was reported by Triplett et al. (2008) in cotton hairy roots producing gossypol. Authors used the same strain of A. rhizogenes. Strain 15834 was also exploited for transforma-tion of four Asteraceae species, but unsuccessful for C. of-ficinalis (Pellegrino et al. 1999). Among 41 lines of hairy root cultures 48.8% expressed intense growth: 13 lines originated from cotyledon and 7 from hypocotyl. For ob-taining better growth which enables biochemical analysis part of cultures were introduced into liquid medium and maintained in rotary shaker (Fig. 3). Recently two lines with the highest growth, one from each sort of explants, were selected for determination of oleanolic acid amount in both tissue and in the medium (Fig. 4). Line from coty-ledon (line C) contained comparable amounts of OA as a line from hypocotyls (lineH). Hairy roots contained 64 times more OA than embryogenic culture and 25 times more than root suspension culture (unpublished data). Interesting results were noticed in liquid media: line H re-leased 20-times more OA than line C. This feature opens excellent possibilities for biotechnological acquisition of such compounds. References: Guillon S et al. (2006) Trends Biotechnol 24: 403–409.Janiszowska W, Kasprzyk Z (1977) Phytochemistry 16: 1919–1923.Liu J (2005) J Ethnopharmacol 100: 92-94.Murashige, Skoog (1962) Physiol Plant 15: 473–497.Pellegrino AP (1999) Acta Hort 502: 299-302.Tripplet BA et al (2008) In Vitro Cell Dev Biol-Plant 44: 508-517.
Acknowledgements:Authors thank Dr Anita Wiśniewska from Department of Plant Physiology, Faculty of Agriculture and Biology, Warsaw Univer-sity of Life Science, for providing A. rhizogenes strain.
Figure 3. Hairy roots culture of C. officinalis after subculture in liquid medium at shaken flasks.
Figure 1. Hairy roots initiation in hypocotyls of C. officinalis after transformation of A. rhizogenes strain ATCC 1583.
Figure 2. Single line of hairy roots culture derived from inde-pendent transgenic events of hypocotyls of C. officinalis.
Figure 4. Oleanolic acid concentration in hairy root cultures orginating from hypocotyl and cotyledon explants.Data is presented as means of three (hypocotyl) and two (coty-ledons) replicates from single line. Bars represent standard error of the means.

Vol. 56 75Bioactive Plant Compounds — Structural and Applicative Aspects
4.10
Microbial transformation of lipophilic flavones as a way to obtain derivatives that are useful in pharmaceutical and cosmetic industry
Edyta Kostrzewa-Susłow, Jadwiga Dmochowska-Gładysz, Tomasz Janeczko
Department of Chemistry, Wrocław University of Environmental and Life Sciences, Wrocław, Polande-mail: Edyta Kostrzewa-Susłow <[email protected]>
Flavonoids are one of the largest groups of secondary metabolites which are found in a wide variety of plants. They are distributed in different parts of plants, includ-ing leaves, flowers, roots, fruits and seeds. The colour of flowers and leaves and its intensity is associated with the presence of these compounds. Flavonoids are involved in many aspects of plant growth and development, such as pathogen defense and UV-light protection, they act as natural inhibitors of auxin transport, activate formation of root nodules in papilionaceous plants, some of them also act as preinfectious compounds, attractants or feeding deterrents (Harborne & Williams, 2000; Winkel-Shirley, 2001). Flavonoids have not been found in animal and human or-ganisms, so far. They are defined as natural non-nutrient chemicals (NNC), the compounds that are a part of human diet and that may directly or non-directly affect homeos-tasis of the body. Due to their large content in plants and plant products, flavonoids are inherent ingredients of hu-man diet. The main sources of these compounds in food are vegetables, fruits, chocolate (cocoa), tea, red wine and beer (Troszyńska et al., 2000). Worldwide research revealed a wide range of valuable biological activities of these compounds, such as antiarte-riosclerotic, anticancerogenic, antidiabetic, antiallergic, antiinflammatory and antihepatotoxic properties (Di Car-lo et al., 1999; Hodek et al., 2002). The wide spectrum of pharmacological activities of flavonoids depends on their chemical structures. Especially important is the presence of a carbonyl group and a C2-C3 double bond, as well as the presence, number and location of hydroxyl and methoxyl groups (Rice-Evans et al., 1996). Flavonoids, due to their health-related activity of different kinds are important ingredients of many nutraceuticals or dietary supplements. Microbial transformations using natural enzymatic sys-tems of microorganisms have become an important tool for obtaining of new derivatives of antimutagenic or anticancerogenic activity. Microorganisms can perform different structural modifications of flavonoids, like in-troduction of new functional groups, which lead to new compounds characterized with increased antioxidant and lipophilic properties, possessing ability to chelate metal ions or characterized with sweet taste (Kostrzewa-Susłow et al., 2007). As a result, such compounds may find poten-tial application as food supplements, cosmetics or phar-maceuticals. As substrates for our research we used non-substituted flavone and its hydroxy and methoxy derivatives. Trans-formations were performed by several filamentous fungi
of genera: Aspergillus, Penicillium, Absidia, Coryneum, Nec-tria, Cryptosporiopsis, Spicaria, Piptoporus, and Chaetomium. In the first stage screening tests were carried out in order to select the most reactive bioreagents for the flavonoid substrates. Then biotransformations on a preparative scale were performed using the selected microorganisms. For the reactions we used whole cells of fungi and the cultures were shaken. Biotransformation products were isolated from the reaction mixtures, purified using chro-matographic methods under optimized conditions and identified by means of spectroscopic methods. In four cases crystallographic analysis was carried out so as to fully establish product structure. In the next stage of the study UV spectra of the substrates and the products were recorded. Effect of new functional groups (auxochromes) introduced in the biotransforma-tions on the shift of UV absorption maxima was analyzed and the changes of molar absorption coefficients were traced. Changes in antioxidant activity of the products compared to the substrates were measured by means of spectropho-tometric method using DPPH radical in order to find cor-relation between the presence and location of functional groups and antioxidant properties.As a result of our study we obtained in microbial trans-formations several products in very high yields (above 90%). Their synthesis either in microbial or in chemical way have not been known, so far. The main type of trans-formation of the tested flavones was hydroxylation at C-3, C-5, C-6, C-7, C-3’, C-4’, O-methylation at C-4’, and demethylation. Analysis of antioxidant properties of the substrates and the products indicate that such an activity depends on the structure and location of substituents in a molecule. The products obtained may help an immune system to provide protection against free radicals. Addi-tionally, some of them are able to chelate metal ions. Mi-crobial transformation of flavonoid compounds may be a new, natural and economic way to obtain new derivatives of valuable biological properties, antioxidant activity or of an interesting taste.References:Di Carlo G et al (1999) Life Sci 65: 337-353.Harborne JB, Williams ChA (2000) Phytochemistry 55: 481-504.Hodek P et al (2002) Chem Biol Interact 139: 1-21.Kostrzewa-Susłow E et al (2007) J Mol Catalysis B: Enzymatic 49: 13-17. Rice-Evans CA et al (1996) Free Radic Biol Med 20: 933-956.Troszyńska A et al (2000) Postępy Fitoterapii 2/2000 (in Polish).Winkel-Shirley B (2001) Plant Physiol 127: 1399-1404.

76 2009Abstracts
4.11
Triterpenoids in allelopathic potential of plants of Vaccinium genus
Anna Szakiel, Barbara Kabacińska
Department of Plant Biochemistry, Faculty of Biology, University of Warsaw, Miecznikowa 1, 02-096 Warszawa, Poland e-mail: Anna Szakiel <[email protected]>
Several plants of Vaccinium genus, including wild species as bilberry Vaccinium myrtillus and cowberry V. vitis-idaea as well as agriculturally important highbush blueberry V. corymbosum, exert significant allelopathic activity in sup-pression of germination and establishment of other plants in immediate environment. This activity seems to be a result of combined action of different classes of second-ary metabolites, mainly phenolics but also triterpenoids, namely oleanolic and ursolic acids and their derivatives (Szakiel & Mroczek, 2007; Szakiel & Henry, 2008). Allelopathy is often submitted for controversial discus-sions because it is difficult to demonstrate the process of release of potentially active chemicals from donor plant, their accumulation in the environment, uptake by target objects, direct harmful effects exerted on afflicted plant and molecular mechanisms of this action. Additionally, allelopathic influence might not involve exclusively di-rect plant-plant interference but also indirect interactions with abiotic and biotic enviromental factors (Inderjit and Weiner, 2001; Weir et al., 2004). The presence of investi-gated triterpenoids in the soil obtained from wild or cul-tivated habitats of Vaccinium plants and observed inhibi-tion of germination and growth of neighbouring plants in nature as well as tested plants in laboratory bioassays, suggest the direct allelopathic action of these compounds, although not excluding the possibility of other indirect effects.The aim of this work was to investigate the con-secutive phase of allelopathic influence exerted by triter-penoids, that is the uptake by target plants and the site of accumulation in their organs and cells. Materials and methods: Preparation of radioactive pre-cursor. [3-3H]Oleanolic acid was obtained by reduction of 3-keto-oleanolic acid with NaB3H4 (Janiszowska et al., 1988) and had a specific activity of 0.3 GBq·mmol-1. Allelopathic bioassays. Sheets of Whatman No.1 filter paper were placed in 100 mm diameter Petri dishes and impregnated with 10 ml of butanol extracts of V. myrtillus soil samples at concentration of 10 μl·ml-1, prepared as previously for V. vitis-idaea (Szakiel and Mroczek, 2007); and/or 10 ml of diethyl ether containing unlabelled or radioactive oleanolic acid at concentration of 10 μl·ml–1
(respectively 65 kBq per dish). Solvents were evaporated and 30 seeds of tested plants: cress Lepidium sativum or lettuce Lactuca sativa were distributed evenly on the pre-pared sheets moisturized with 10 ml of pure water. For each plant control dishes were prepared with pure water only. All dishes were closed and placed in the dark in a thermostat (22oC). Germinating seeds were counted after 3 days, the length of roots and hypocotyls was measured after 7 days and the radioactivity of cellular fractions were established after 15 days of experiment.
Preparation of the cellular subfractions. 15-days old seedlings were separated into seed coats and root and hypocotyl parts. Roots and hypocotyls were ground in a mortar and afterwards in a Potter homogenizer with 0.1 M Tris/HCl, pH 7.3 (3 ml·g–1 fr.w.). The homogenates were centrifuged at 2000 g to remove cell debris. Cytosolic and membranous fractions were obtained by centrifuga-tion at 105 000 × g during 3 h. Determination of radioactivity. Obtained root and hy-pocotyl cytosolic and membranous fractions as well as dried seed coats were extracted 3 times with diethyl ether. After evaporation of solvent, radioactivity of each fraction and seed coat extracts was measured in toluene-based scintillator in the LS 6000 IC Beckman counter. Results and discussion: Oleanolic acid (Fig. 1) was cho-sen as a tested compound in our experiments due to rela-tively simple methods of its obtaining in both unlabelled (Szakiel et al., 2008) and radioactive form. Allelopathic activity of oleanolic acid supplied individually or mixed with soil extract containing bilberry saponins was inves-tigated in biotests with the use of two model plants, cress Lepidium sativum and lettuce Lactuca sativa. Applied con-centration of oleanolic acid was comparable to the con-centration of the mixture of free triterpene acids, oleanol-ic and ursolic, detected in the soil obtained from natural habitat of bilberry. Bilberry is the only plant of Vaccinium genus in which oleanolic acid is the predominant triter-pene isomer (Szakiel et al., 2007). The results of bioassays, presented as compared to respective controls, are shown in Table 1. Oleanolic acid inhibited the germination of cress and lettuce seeds by 7 and 13%, respectively. This effect was more pronounced (14 and 24%) when the mixture of oleanolic acid with the fraction of bilberry saponins was applied. Generally, free triterpene acids are not consid-ered as strong agents suppressing germination (Ohara and Ohira, 2003). The influence of oleanolic acid on the seedling growth of the two tested plants exhibited slight-ly distinct pattern. The growth of cress and lettuce roots was decreased for both plants by 25% by oleanolic acid
Table 1. The influence of oleanolic acid on germination and growth of tested plants.Values are means ±S.D. of 3 replicates. OL – oleanolic acid, Sap – soil extract containing bilberry sa-ponins.
Plant/CompoundGermination Hypocotyl
lengthRoot length
% of controlCressOL 93 ± 1.6 87 ± 3.07 75 ± 4.98OL+Sap 86 ± 2.8 79 ± 3.96 64 ± 5.23LettuceOL 87 ± 2.4 96 ± 1.16 75 ± 2.98OL+Sap 76 ± 3.2 102 ± 1.14 68 ± 4.06
COOH
OHFigure 1. The structure of oleanolic acid.

Vol. 56 77Bioactive Plant Compounds — Structural and Applicative Aspects
and more, by 36% and 32%, respectively, by its mixture with saponins. Similarly, the hypocotyl growth of cress was suppressed in both tested conditions, respectively by 13 and 21%. However, the hypocotyl growth of lettuce was reduced scarsely, by 4% by oleanolic acid and even stimulated by 2% by mixed fractions, which is a typical phenomenon for many allelochemicals applied in low concentrations. Obtained results point to the moderatory allelopathic ac-tivity of oleanolic acid exerted against model dicotyledon plants. They are consistent with previous observations that diethyl ether extracts of soil obtained from natural habitat of cowberry V. vitis-idaea containing the mixture of ursolic and oleanolic acids slightly inhibited the germina-tion of lettuce and cress (by 6–10%), the root growth by 25–34%, shoot by 10–22%, and these effects were stronger in the presence of butanol extracts (Szakiel & Mroczek, 2007). The stronger inhibitory effect on roots compared with shoots, often observed in allelopathic bioassays on Petri dishes, might reflect the mechanism of allelopathic influence more significant in this organ but it can also be caused by the fact that roots had been in direct contact with supplied chemicals. After the administration of radioactive oleanolic acid to allelopathic bioassays, its incorporation into cytosolic and membranous fractions obtained from seedlings of tested target plants, divided into root and hypocotyl parts and separated from their seed coats, was investigated. The re-sults are presented in Table 2. Despite lipophilic properties of oleanolic acid and its con-sequent unsolubility in water, considerable amount of administrated radioactivity was found in plant material (15% in cress, 11% in lettuce). Some amount of supplied radioactive oleanolic acid was retained in the seed coats (26% in cress, 31% in lettuce). The majority of labelling incorporated into developed organs of seedlings of both plants was found in roots (86% of radioactivity detected in cress seedlings separated from seed coats, 80% — in lettuce seedlings), more precisely: in root membranous fraction (84% and 77% in cress and lettuce, respectively). These results point to the root membranes as the main site of action of oleanolic acid in target plants. Among trit-erpenoids, the compounds that are regarded as strongly
interacting with biomembranes are saponins with their amphiphilic structures and ability of pore-forming. How-ever, also lipophilic triterpene acids can cause distur-bance of membrane permeability leading to uncontrolled cell leakage (Wink 2003). Another mechanism of action assumed to triterpenoids is antiauxin and antigibberelin activity (Jha et al, 2006). Further studies are required to describe more precisely the function of triterpenoids in allelopathic interactions. References: Inderjit, Weiner J (2001) Perspectives in Plant Ecology, Evolution and Systematics 4: 3-12. Janiszowska W et al (1988) Pol J Chem 62: 403-407. Jha S et al (2006) Pak J Pl Sci 12: 103-113. Ohara S, Ohira T (2003) J Wood Sci 49: 59-64. Szakiel A et al (2007) Acta Biochim Polon 54: Suppl 4: 77. Szakiel A, Henry M (2008) Planta Med 74: 980. Szakiel A, Mroczek A (2007) Acta Biochim Polon 54: 733-740. Szakiel A et al (2008) Planta Med. 74: 1667-1768. Weir TL et al (2004) Curr Opin Plant Biol 7: 472-479.Wink M (2003) Phytochemistry 64: 3-19.
Table 2. The incorporation of [3-3H]oleanolic acid into seed-lings of cress and lettuce. Values are means ±S.D. of 3 inde-pendent experiments. Plant/Fraction Radioactivity [Bq] %CressSeed coat 2535 ± 223 26.0Hypocotyl membranes 670 ± 54 3.8Hypocotyl cytosol 368 ± 34 6.9Root membranes 6054 ± 745 62.0Root cytosol 123 ± 31 1.3Total 9750 100LettuceSeed coat 2216 ± 210 31.0Hypocotyl membranes 618 ± 59 4.8Hypocotyl cytosol 344 ± 25 8.6Root membranes 3789 ± 426 53.0Root cytosol 183 ± 27 2.6Total 7150 100

78 2009Abstracts
4.12
Hairy roots and untransformed root culture of Eryngium L. species
Barbara Thiem, Małgorzata Kikowska
Department of Pharmaceutical Botany and Plant Biotechnology, Poznań University of Medical Sciences, Św. Marii Magdaleny 14, 61-861 Poznań, Polande-mail: Barbara Thiem <[email protected]>
Introduction: Eryngium planum L., E. campestre L. and E. alpinum L. belong to the Apiaceae family and Sanicu-loideae subfamily. These species have been used in folk medicine as diuretic, expectorant and antimycotic rem-edies. The major active secondary metabolites are phe-nolic acids (rosmarinic acid), triterpene saponins and flavonoids (Duke et al., 2002; Matkowski, 2008). In our previous studies it was reported that the callus and cell suspension cultures of E. planum had demonstrated the ability to synthesize both — phenolic compounds and triterpene saponins (Thiem et al., 2009). The production of wide range of secondary metabolites by organ cultures — untransformed root cultures and hairy root cultures were attempted (Guillon et al., 2006). Hairy root cultures have been established for a variety of species and they offer many advantages over cell culture systems includ-ing fast auxin-independent growth, genetic stability and stable high-level production of secondary metabolites. Unlike plant cell cultures which often produce very small amounts of compounds, hairy root cultures can display high biosynthetic capability that is often comparable to those of normal roots from intact plants (Guillon et al., 2006). Accumulation of secondary metabolites by organ cultures is usually more efficient. Moreover, culture con-ditions or elicitors were able to enhance the content of desired compounds produced by root cultures without transformation (Kevers et al., 1999; Kim et al., 2003) and hairy root cultures (Satdive et al., 2007). These two type of root cultures can be useful for the production of biomass and for testing the production of secondary metabolites.Aim: The aim of present study was to establish the in vit-ro root cultures of three Eryngium species – E. planum L., E. campestre L. and E. alpinum L., without transformation and using strain of Agrobacterium rhizogenes.Materials and methods: Sterile seedlings and plant-lets, previously obtained (Thiem et al., 2006), were used for induction of root cultures. Agrobacterium rhizogenes (agropine strain A4), kept on agar YMB medium at 26oC for 48 h was used for transformation. Hairy roots of E. planum were induced by infection of leaves with peti-oles (Fig. 1). Wounded explants were cultured individual-ly on solid, phytohormone free medium MS (Murashige & Skoog, 1962) or MS supplemented with acetosyringone. The hairy roots derived after the infection with Ri T-DNA were transferred to 250 ml shake flasks with 100 ml of liquid MS or WP medium supplemented with ampicillin to eliminate bacteria. Results of transformation were de-tected by PCR. For comparison, also untransformed root cultures of E. planum were established.Untransformed root cultures of E. alpinum and E. planum were obtained from root tips of plantlets. Root segments (0.5–5 cm) were
inoculated into 300 ml flasks, containing 100 ml of liq-uid MS medium supplemented with 0.1–0.5 mgl–1 IAA, IBA or NAA, where their elongation and branching was observed (Figs. 2, 3).The root cultures were subcultured every 6 weeks and maintained in a growth chamber in the dark, at 24oC and kept on rotary shaker at 110 rpm. Growth activity of the root cultures were estimated by the growth index and moisture content in the cultured roots was determined.Results: Our preliminary phytochemical studies (TLC, HPLC) were showed that roots of E. planum and E. alpinum in in vitro cultures are able to produce the phenolic acids (chlorogenic and rosmarinic acids), flavonoids and trit-erpene saponins (Thiem et al., 2009a; Thiem et al., 2009b).Hairy root cultures were established for E. planum. Fre-quency of transformation from infected petioles was 40%. E. campestre and E. alpinum gave negative response to pro-duce hairy roots within 45 days after infection. Therefore, adventitious root cultures, without transformation with A. rhizogenes were established for E. alpinum. All media with tested auxins were suitable for root elongation and branching. The best growth of root culture was observed on MS supplemented with NAA 0.4 mgl–1 (growth index = 10.2 g/6 weeks). Conclusion: To summarize, the obtained root cultures were established for future elicitation with methyl jas-monate and sucrose, for enhancing the accumulation of triterpene saponins, rosmarinic acid and total phenolics.
Figure 1. Hairy roots induced from petioles of E. planum
Figure 2. Untransformed root culture of E. alpinum
Figure 3. Biomass of untransformed roots of E. alpinum

Vol. 56 79Bioactive Plant Compounds — Structural and Applicative Aspects
References:Duke JA et al (2002) Handbook of medicinal herbs CRC Press 277-278.Guillon S et al (2006) Curr Opin Plant Biol 9: 341-346.Kevers C et al (1999) Plant Growth Regul 27:173-178.Kim Y-S et al (2003) In Vitro Cell Dev Biol-Plant 39: 245-249.Matkowski A (2008) Biotechnol Adv 26: 548-560.Murashige T, Skoog F (1962) Physiol Plant 15: 473-497.Satdive RK et al (2007) JnBiotechnol 128: 281-289.Thiem B et al (2009a) PSE CSIC — Young Scientists Meeting Los Narejos Spain 12th–14th May 2009, Abstracts p 108.Thiem B et al (2009b) 5thGerman-Polish Symposium Poznan, 15–16 May 2009, Abstracts p 127.Thiem B et al (2006) XI Ogólnopolska Konferencja Kultur In Vitro i Biotechnologii Roślin, Międzyzdroje, 6–9 września 2006 p 89 (in Polish).
Acknowledgements:This work was supported by the Ministry of Science and Higher Education, Warsaw, Poland (grant NN 405 065334).
4.13
Rosmarinic acid production in basil under in vitro condition
Anna Czubacka, Justyna Krzyżanowska, Ireneusz Kapusta, Marcin Przybyś, Teresa Doroszewska, Wiesław Oleszek
Institute of Soil Science and Plant Cultivation, State ResearchInstitute, Czartoryskich 8, 24-100 Puławy, Polande-mail: Anna Czubacka <[email protected]>
Basil (Ocimum basilicum L.) is a species belonging to plant family Lamiaceae, subfamily Nepetoideae. Within the ge-nus Ocimum, the species Ocimum basilicum is the major essential crop cultured in many countries of the world. Basil is an annual, 20–60 cm long plant, with white-pur-ple flowers. It originally comes from India and other re-gions of Asia. Basil is commonly used as a culinary herb because of distinctive aroma of the leaves containing es-sential oils [1–3]. Fresh basil is widely used in the kitchen, especially in Mediterranean and Southeast Asian food [4]. Furthermore, it is in use for many years in the traditional medicine in the treatment of headaches and pains, kidney malfunction, coughs. Moreover, the high economic value of basil is due to the presence of rosmarinic acid (RA), which is an ester of caffeic acid and 3,4-dihydroxyphenyl-lactic acid [5]. Rosmarinic acid has a number of interesting biological activities, e.g. antiviral, antibacterial, anti-in-flammatory and antimutagen. Thanks to its antioxidative properties, it has application in cosmetic industry [2, 3, 6]. Rosmarinic acid is readily accumulated in undifferenti-ated plant cell cultures [5]. Therefore, RA production in basil may be increased in suspension culture. However, at first it is necessary to choose the cultivar which is the richest in this secondary metabolite. Material and methods: Plant material. The materials of the study were plants belonging to three cultivars of basil (Ocimum basilicum): sweet basil, lemon basil and red basil. Seeds were sterilized in 10% perhydrol, rinsed in sterile water and sown on LS medium [7]. After a few days sprouting plants were transferred on LS medium enriched with 0.2 mg/l NAA and 0.2 mg/l IAA. After a month aerial parts of plants were collected.Preparation of samples and analysis. Crude extracts of basil samples were obtained through the extraction of lyophilized plant material using an automated extraction device ASE 200. The ASE conditions were following: 70% ethanol as the extraction solvent, 1500-psi solvent pressure, 100°C cell temperature, flush 150% and 2 static cycles for 2 minutes each. Extracts-25 ml were collected in vials. In the next step the extracts were dried (ethanol was evaporated) at 40°C under reduced pressure in a rotary evaporator
Table 1. Concentration of rosmarinic acid in aerial parts of in vitro basil plants belonging to different cultivars Basil varieties Rosmarinic acid concentration (mg/g d. m.)red basil 11.749 lemon basil 26.836 sweet basil 29.856

80 2009Abstracts
(Laborota 4000, Heidolph). The dried extracts were dis-solved in water, then flavonoid fractions were purified by Solid Phase Extraction (SPE). These extracts were loaded on C18 cartridges (Sep-Pak, Waters) and washed with water to remove carbohydrates, to elute flavonoids the columns were washed with 40% ethanol. The phy-tochemical analysis was carried out using an ultra per-formance liquid chromatography technique. The analy-sis was performed on UPLC BEH C18 column (1.7 mm, 50 mm × 2.1 mm) utilizing a gradient elution profile of a mobile phase consisting of 0.1% CH3COOH in water and 40% AcN. The column was maintained at 50°C. Injection volume was 5 μl. Time of analysis was 6.5 min., at flow rate 0.350 ml/min. The UPLC analyses were monitored at 254 nm. Quantitative determination of rosmarinic acid was carried out using the external standard method and calibration curve. Results and discussion: The dominant compound ob-tained as a result of UPLC analysis was rosmarinic acid. Another phenolic compounds were present in tested samples in very little concentrations. Rosmarinic acid concentrations were different depending on basil culti-var. The highest level of that secondary metabolite was noticed in case of sweet basil — nearly 30 mg/g dry mat-ter (Table 1). Lemon basil showed to be a little poorer in rosmarinic acid — above 26 mg/ g d. m. The lowest pro-duction of that compound was observed in case of plants of red basil. Differences in production of essential oils in five cultivars of basil were showed by Czech researchers [2]. They ob-served that the pattern of producing analyzed compounds remained similar, while the concentrations of individual metabolites varied depending a cultivar. Rady and Nazif reported that level of rosmarinic acid production in Oci-mum americanum plants coming from in vitro culture was conditioned by a kind of phytohormones contained in a medium. The highest production of rosmarinic acid they observed in plants growing on MS medium with addi-tion of 1 mg/l BA and 0.25 mg/l IAA [8]. Our experiment showed that among three tested cultivars, sweet basil is the most effective as regards RA production under in vit-ro condition. Therefore this cultivar could be material for further investigations. References:1. Özcan M et al (2005) J Food Engin 69: 375-379.2. Klimankova E et al (2008) Food Chem 107: 464-472.3. Opalchenova G Obreshkova D (2003) J Microbiol Methods 54: 105-110.4. Juntachote T, Berghofer E (2005) Food Chem 92: 193-202.5. Petersen M, Simmonds MSJ (2003) Phytochemistry 62: 121-125.6. Pereira P et al (2005) Pharmacol Res 52: 199-203.7. Linsmayer EM, Skoog F (1965) Physiol Plantarum 18: 100-127. 8. Rady MR, Nazif NM (2005) Fitoterapia 76: 525-533 (in Polish).





























