Embed Size (px)
Citation preview

www.elsevier.com/locate/brainres
Brain Research 1019 (2004) 64–67
Research report
Serotonergic collateralized projections from Barrington’s nucleus to the
medial preoptic area and lumbo-sacral spinal cord
Antonella Russoa,*, Sebastiana Monacoa, Rosa Romeob, Rosalia Pellitteric, Stefania Stanzania
aDepartment of Physiological Sciences, University of Catania, Viale A. Doria 6, 95125 Catania, ItalybDepartment of Anatomy, Diagnostic Pathology, Phorens Medicine, Hygiene and Publich Health, University of Catania, Via Santa Sofia, Catania, Italy
c Institute of Neurological Sciences, National Research Counsil, Viale R. Margherita 6, 95123, Catania, Italy
Accepted 10 March 2004
Available online 8 July 2004
Abstract
In this study, we employed triple fluorescent labelling to reveal the distribution of the direct serotonergic neurons within Barrington’s
nucleus (BN) that supply branching collateral input to the medial preoptic area (MPA) and to the lumbo-sacral spinal cord (LSC).
Immunocytochemical detection of the monoclonal antibody raised against serotonin was used for identification of the neurons. The
projections were defined by injections of two retrograde tracers: fluoro gold and rhodamine in the MPA and LSC, respectively. The aim of
this study is to identify the direct projections to BN and MPA and/or LSC. The present study confirms findings of others describing BN–LSC
projections and extends previous findings by demonstrating an single or collateralized fibers with MPA, and serotonergic immunoreactive
fibers.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Barrington’s nucleus; Medial preoptic area; Lumbo-sacral spinal cord; Serotonergic neuron; Fluoro gold; Rhodamine
1. Introduction strated that the neurons of Barrington’s nucleus (BN)
Micturition is a spino-bulbo-spinal reflex: a coordinated
action between the detrusor muscle of the bladder and the
external striated urethral sphincter. In adult mammals, the
area responsible for the synergistic action of both muscles
(detrusor–sphincter) is identifiable in the brainstem. The
region involved is located in the dorsolateral pons and is
known as Barrington’s area, M-region or pontine micturi-
tion center in different species [1–3,8,9]. This area is
indicated as a small group of neurons lying just ventrome-
dial to the rostral pole of the nucleus locus coeruleus (LC),
and receives afferent fibers from brainstem nuclei and
forebrain limbic structures. Electrical or chemical stimula-
tion of this region in rats elicits bladder contraction and
increases discharge of bladder postganglionic nerve
[11,15]. Moreover, lesions to this nucleus or applications
of opiates inhibit micturition [19]. Anatomical studies
utilizing anterograde and retrograde tracers have demon-
0006-8993/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.brainres.2004.03.080
* Corresponding author. Tel.: +39-95-7384041; fax: +39-95-330645.
E-mail addresses: [email protected] (A. Russo),
[email protected] (S. Stanzani).
project to the intermediolateral column of the sacral spinal
cord in the region of preganglionic neurons that innervate
the bladder [5,6,16]. It has been suggested that the spinal
neurons convey bladder-filling information directly to BN;
there have been some studies on this pathway with trans-
neuronal virus labelling [10], c-fos expression [4], and
physiological techniques [11]. In addition, BN receives
widespread afferents from the brain including hypothala-
mus [2] and, in particular, the medial preoptic area (MPA)
that can be involved in the modulation of the micturition
reflex, in the rat [7]; in addition, BN contains not only
projecting neurons to the spinal cord, but also some
projecting neurons to the paraventricular thalamic nucleus,
periacqueductal gray and other regions, through axon
collaterals [12,17]. In contrast to the numerous studies of
the BN and spinal cord, little information exists regarding
the connections of BN with supraspinal structures. Some
studies have involved the BN in the simultaneous trans-
mission of signals [17] with corticotropin-releasing hor-
mone (CRH) release. Nonetheless, there have been no
systematic investigations focusing on various neurotrans-
mitters in BN [14,18]. Here, we raise the question of
whether single neurons in BN sending projection fibers to

A. Russo et al. / Brain Research 1019 (2004) 64–67 65
supraspinal region also project directly to the spinal cord,
and what kind of neurotransmitter these fibers contain.
In this study, we used two retrograde tracers to eluci-
date the circuitry of BN regulation to MPA and lumbo-
sacral spinal cord (LSC). Furthermore, to determine which
neurotransmitter is present in BN neurons, we initiated a
systematic investigation using immunocytochemical 5-HT
(serotonin) detection, and later on, we could assay other
specific antibodies. The aim of this study was to demon-
strate the presence and distribution of BN neurons that
project via axon collaterals to the medial preoptic area
(MPA) and lumbo-sacral spinal cord (LSC), and the
participation of serotonin in these pathways, presumed to
controlling micturition.
2. Materials and methods
All animal experiments were carried out in accordance
with current institutional guidelines for the care and use of
experimental animals. Experiments were performed on 12
adult male Wistar rats weighing 250–300 g (Morini, Italy),
maintained under controlled conditions of room temperature
(23F 1 jC) and lighting (lights on 07:00–19:00 h); labo-
ratory chow diet and water were available ad libitum; the in
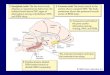
Fig. 1. Injection zones in the CNS. Microphotographs showing injection zones: (A
(B) FG injection site and drawing in LSC (black area), scale bar = 90 Am.
vivo experimental procedure was performed during the
daytime (10:00–13:00 h). Animals were anaesthetized with
chloral hydrate (400 mg/kg i.p.). Briefly, 0.08 Al of rhoda-mine-labelled bead (RLB) were injected into MPA (stereo-
taxic planes: AP=� 0.30; L= 0.5; V=� 8.5) [13], whereas
0.04 Al of FG solution (6%in saline) were injected into the
ventral portion of LSC monolaterally. Both tracers were
pressure-injected at a rate of 50 nl/min using 1 Al Hamilton
micro-syringe.
Seven days after injections, the animals were reanaes-
thetized and perfused through ascending aorta with 60 ml
saline solution, followed by 300 ml ice-cold 4% parafor-
maldeyde phosphate buffer (pH 7.4). The brains were
removed, immersed in the same fixative for 3–4 h, and
cryo-protected overnight in phosphate-buffered 20% su-
crose solution. Two series of coronal sections (40 Am) were
cut on a cryostat and collected in phosphate buffer (PBS, pH
7.4, 0.1 M). One series was serially mounted on microscope
slides and immediately observed with a Reichert fluores-
cence microscope. After the first observation of BN region,
we employed the second series for immunocytochemical
processing. The series designated for immunocytochemical
visualization of serotonin was incubated as free-floating
sections for 16–18 h with anti-mouse monoclonal anti-
serotonin antibody (Chemicon). The primary antibody was
) RLB injection site and drawing in MPA (black area), scale bar = 400 Am;

Fig. 2. Microphotographs of triple-labelled neurons FG-RLB-FITC in
Barrington’s nucleus (BN). (A) Cell stained positively for FG (excitation
wavelength 330 nm); (B) the same cell stained positively for RLB
(excitation wavelength 560 nm) indicating the existence of a collateral
axon; (C) the same cell is positive to the serotonin (FITC, excitation
wavelength 450 nm). Scale bar = 20 Am. (D) Schematic drawing of frontal
brainstem section including BN region: the black dots show the labelled
neurons within the BN.
Table 1
Ipsilateral and contralateral labelled cells in the whole Barrington’s nucleus
Ipsilateral Contralateral
FG 17.5F 0.1 12.8F 0.3
RLB 8.8F 1.7 6.0F 0.8
FG+RLB 5.4F 1.0 2.1F 0.3
FG+ 5-HT 10.3F 1.0 8.4F 0.4
RLB+ 5-HT 6.1F 0.2 5.3F 0.2
FG+RLB+5-HT 1.8F 0.2 No neurons
The results (cells number per section) are summarized in this table.
A. Russo et al. / Brain Research 1019 (2004) 64–6766
diluted (1:200) in a solution of 0.3% Triton X-100 in PBS.
After a 15-min rinse in PBS, the sections were incubated for
30 min with fluorescein isothiocyanate (FITC) conjugated to
sheep anti-mouse Ig G (1:100; Boehringer). The sections
were air-dried, mounted and observed with a Reichert
fluorescence microscope equipped with filter combinations
revealing red (RLB), yellow (FG) or green (FITC) fluores-
cence. To determine the number of neurons containing both
retrograde tracers, cell count was performed on the series
that was not stained immunocytochemically. The incidence
of triple-labelled cells was estimated directly from the series
processed immunocytochemically by sequentially viewing
tissue with the three different filters. For every animal, three
non-adjacent sections were evaluated and the labelled cells
were plotted onto schematic drawings of the BN region
level. Thus, cell numbers were expressed as the average
number/section calculated from these three sections.
3. Results
All injections of the retrograde tracers remained relatively
localized as has previously reported. Fig. 1 shows micro-
photographs and schematic drawings example of the injec-
tion site of the retrograde tracers in the MPA and LSC; only
those cases where microscopic analysis of the injection site
revealed that the tracer deposits were correctly positioned
were included in this study. Injection of RBL into MPA and
of FG into LSC resulted in a large number of retrogradely
labelled neurons in the whole ipsi- and contralateral BN
region (stereotaxic planes: � 9.16/� 9.80 [13]; Fig. 2D).
More BN neurons from the ipsilateral LSC and MPA
were labelled than contralateral (Table 1).
The fluorescence microscopy revealed a substantial
number of double-labelled neurons, thus providing evidence
of collateralization to the MPA and LSC. These neurons
were generally of small size (25–30 Am).
The multiple staining protocol followed in the present
study evidences patterns of neurons in the brainstem, dem-
onstrating serotonin-like immunoreactivity (5-HT) together
with retrograde labelling by each of the tracers employed.
Only a limited number of labelled cells (FG +RLB) were
found to bifurcate to the contralateral MPA and LSC (10%),
whereas a consistent population of cells were found to
branch to MPA and LSC ipsilaterally (20%).
A relatively low number of FG/RLB/FITC triple labelled
(Fig. 2A–C) were scattered mainly at the ipsilateral BN
level (33.33% of the total immunoreactive population).
The results (BN cells number per section) are summa-
rized in Table 1.
Our results show that: (1) BN single neurons directly
project to MPA, and confirm that BN single neurons directly
project to LSC [7]; (2) by separate counting, the RLB-5-HT-
positive neurons are about 88.33% ipsilateral, and 69.31%

A. Russo et al. / Brain Research 1019 (2004) 64–67 67
contralateral; whereas the FG-5-HT neurons are 65.62%
ipsilateral and 58.85% contralateral; (3) BN neurons supply,
via collaterals, branching inputs to the MPA and LSC; (4)
about one third of these double-labelled neurons in BN are
serotonergic.
4. Discussion
These results implicate the distribution of neuronal single
projection in the BN; in particular, they confirm other
findings [5–7,16] regarding the projections to LSC and
visualize new projections to MPA. This result is very
interesting because the demonstration of direct MPA pro-
jection exists [7]; in addition, the presence of collateralized
fibers of the BN neurons includes the possibility of the
ascending and descending simultaneous control. The dis-
covery that BN laterodorsal tegmental nucleus modulates
micturition has been defined [1–3,8,9]. BN receives blad-
der-filling information through ascending projection path-
ways, directly or indirectly via periacqueductal gray.
Besides, the direct projections from MPA to the BN neurons
directly projecting to the LSC [7] were found. This result
involves the MPA function in control of BN that regulates
the micturition.
5. Conclusion
In conclusion, this study reveals the presence of a
projection from the Barrington’s nucleus to MPA and this
ascending serotonergic pathway may close a control loop.
Furthermore, the BN neurons send projection fibers to both
the MPA and LSC by way of serotonergic and non-seroto-
nergic axon collaterals. Future researches will include in-
vestigating the other possible neurochemical nature of these
BN projections.
Acknowledgements
This study was supported by MIUR. We thank Mr. Silvio
Bentivegna for help in adjustment of figure.
References
[1] F.J.F. Barrington, The effect of lesions of the hind- and midbrain on
micturition in the cat, Q.J. Exp. Physiol. Cogn. Med. 15 (1925)
81–102.
[2] B.F. Blok, Central pathways controlling micturition and urinary con-
tinence, Urology 59 (2002) 13–17.
[3] B.F. Blok, G. Holstage, The pontine micturition center in rat receives
direct lumbosacral input. An ultrastructural study, Neurosci. Lett. 282
(2000) 29–32.
[4] K. Bon, M. Lanteri-Minet, J. De Pommery, J.F. Michiels, D.
Menetrey, Cyclophosphamide cystitis as a model of visceral pain in
rats. A survey of hindbrain structures involved in visceroception and
nociception using the expression of c-Fos and Krox-24 proteins,
Exp. Brain Res. 108 (1996) 404–416.
[5] G. Cano, J.P. Card, L. Rinaman, A.F. Sved, Connections of Barring-
ton’s nucleus to the sympathetic nervous system in rats, J. Auton.
Nerv. Syst. 79 (2000) 117–128.
[6] Y.Q. Ding, H.X. Zheng, L.W. Gong, Y. Lu, H. Zhao, B.Z. Qin, Direct
projections from the lumbosacral spinal cord to Barrington’s nucleus
in the rat: a special reference to micturition reflex, J. Comp. Neurol.
389 (1997) 149–160.
[7] Y.Q. Ding, D. Wang, X. Jun-Qing, J. Gong, Direct projections from
the medial preoptic area to spinally-projecting neurons in Barrington’s
nucleus: an electron microscope study in the rat, Neurosci. Lett. 271
(1999) 175–178.
[8] G. Holstage, D. Griffiths, H. DeWall, E. Dalm, Anatomical and phys-
iological observations on supraspinal control of bladder and urethral
sphincter muscles in the cat, J. Comp. Neurol. 250 (1986) 449–461.
[9] A.D. Loewy, C.B. Saper, R.P. Baker, Descending projections from the
pontine micturition center, Brain Res. 172 (1979) 533–539.
[10] I. Nadelhaft, P.L. Vera, Central nervous system infected by pseu-
dorabies virus injected into the rat urinary bladder following unilat-
eral transection of the pelvic nerve, J. Comp. Neurol. 359 (1995)
443–456.
[11] H. Noto, J.R. Roppolo, W.D. Steers, W.C. De Groat, Excitatory and
inhibitory influences on bladder activity elicited by electrical stimu-
lation in the pontine micturition center in the rat, Brain Res. 492
(1988) 99–115.
[12] K. Otake, Y. Nakamura, Single neurons in Barrington’s nucleus pro-
jecting to both the paraventricular thalamic nucleus and the spinal
cord by way of axonal collaterals: a double labeling study in the
rat, Neurosci. Lett. 209 (1996) 97–100.
[13] G. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates,
Academic Press, Sydney, 1986.
[14] D.A Ruggiero, N.D. Underwood, P.M. Rice, J.J. Mann, V. Arango,
Corticotropic-releasing hormone and serotonin interact in the human
brainstem: behavioral implications, Neuroscience 91 (4) (1999)
1343–1354.
[15] M. Sasaki, Bladder contractility-related neurons in Barrington’s nu-
cleus: axonal projections to the spinal cord in the cat, J. Comp. Neu-
rol. 449 (2002) 355–363.
[16] D.J. Tracey, Ascending and descending pathways in the spinal cord,
in: G. Paxinos (Ed.), The Rat Nervous System, 2nd ed., Academic
Press, San Diego, CA, 1995, pp. 67–75.
[17] R.J. Valentino, L.A. Pavcovich, H. Hirata, Evidence for corticotro-
pin-releasing hormone projections from Barrington’s nucleus to the
periaqueductal gray and dorsal motor nucleus of the vagus in the rat,
J. Comp. Neurol. 363 (1995) 402–422.
[18] R.J. Valentino, S. Chen, Y. Zhu, G. Aston-Jones, Evidence for diver-
gent projections to the brain noradrenergic system and the spinal
parasympathetic system from Barrington’s nucleus, Brain Res. 732
(1996) 1–15.
[19] R.N. Willette, S. Morrison, H.N. Sapru, D.J. Reis, Stimulation of
opiate receptors in the dorsal pontine tegmentum inhibits reflex con-
traction of the urinary bladder, Pharmacol. Exp. Ther. 244 (1988)
403–409.