Embed Size (px)
Citation preview

© 2003 Nature Publishing Group
REVIEWS
Just over 200 million years ago, archosaurs — theancestors of birds — evolved tympanic ears (includinga tympanic membrane and a middle ear), allowingthem to hear and to localize airborne sounds. Somemillion years later, early mammals independentlyevolved tympanic ears1,2 (FIG. 1). The subsequent evolu-tion of sophisticated auditory systems to allow theelaborate analysis of airborne sound occurred com-pletely independently in birds and mammals, althoughit was driven by the same evolutionary constraints,including the limited number of physical cues that areavailable to compute the position of a sound source.One of these cues is the tiny differences in the arrivaltime of a sound at the two ears, known as INTERAURAL
TIME DIFFERENCES (ITDs; FIG. 2a). The neural circuits thatencode these ITDs in birds and mammals exemplifyhow the evolution of the vertebrate brain came up withdifferent solutions for a single problem.
For both birds and mammals, ITD processing represents the ultimate challenge in temporal process-ing. No other neural system in vertebrates comes closeto the temporal resolution required to encode ITDs,except for the electric system in some fish. Consider
that the duration of an action potential is almost twoorders of magnitude greater than the minimal ITDsthat barn owls or humans can resolve (<10 µs).Particular structural and functional adaptations werenecessary to allow the successful processing and encod-ing of these ITDs. This fact makes the ITD-processingstructures in birds and mammals attractive model systems for studying the rules that underlie precisetemporal processing in the vertebrate brain in general,and it has stimulated many experimental studies andcomputational models.
The current textbook view of ITD processing resem-bles closely what Jeffress proposed more than 50 yearsago in his elegant model3. This model was based onthree main assumptions (BOX 1): bilateral, TIME-LOCKED orPHASE-LOCKED inputs into the ITD-processing system;COINCIDENCE DETECTION by ITD detector neurons; and an arrangement of delay lines to adjust coincidencedetector neurons to different preferred ITDs, creating atopographic representation of AZIMUTHAL SPACE.
The bird ITD-encoding system seems to have evolvedin a way that closely matches Jeffress’s predictions (FIG. 3a,b). Neurons in the nucleus laminaris, the first
NEW ROLES FOR SYNAPTICINHIBITION IN SOUNDLOCALIZATIONBenedikt Grothe
The arrival times of a sound at the two ears are only microseconds apart, but both birds andmammals can use these interaural time differences to localize low-frequency sounds.Traditionally, it was thought that the underlying mechanism involved only coincidence detection ofexcitatory inputs from the two ears. However, recent findings have uncovered profound roles forsynaptic inhibition in the processing of interaural time differences. In mammals, exquisitely timedhyperpolarizing inhibition adjusts the temporal sensitivity of coincidence detector neurons to thephysiologically relevant range of interaural time differences. Inhibition onto bird coincidencedetectors, by contrast, is depolarizing and devoid of temporal information, providing amechanism for gain control.
INTERAURAL TIME DIFFERENCE
(ITD). The difference in thearrival time of a sound at the twoears. Depending on the positionof the sound source and theindividual inter-ear distance,ITDs can be up to about 120 µsin the Mongolian gerbil, and upto about 650 µs in humans.
NATURE REVIEWS | NEUROSCIENCE VOLUME 4 | JULY 2003 | 1
Max-Planck-Institute ofNeurobiology,Auditory Processing Group,Am Klopferspitz 18a,D-82152 Martinsried,Germany.e-mail:[email protected]:10.1038/nrn1136
S E N S O R Y S Y S T E M S
© 2003 Nature Publishing Group
TIME-LOCKED
Action potentials of manyauditory neurons are locked tospecific events of acousticstimuli, such as onsets, offsets,prominent fluctuations infrequency or amplitude, or evena specific phase-angle ofsinusoidal low-frequencysounds.
PHASE-LOCKED
The most extreme case of time-locking in auditory neurons.Many low-frequency neurons(in mammals up to a few kHz, inbarn owls up to 8 kHz)synchronize their discharge to aspecific phase-angle of tones.
COINCIDENCE DETECTION
The activation of neurons not bysingle inputs, but only by thesimultaneous activity of severalinputs. Coincidence detectorneurons can be foundthroughout the nervous system.The most extreme case ofcoincidence detection is foundin the binaural auditory systemwhere the time windows forcoincidence detection are in therange of microseconds.
AZIMUTHAL SPACE
The definition of auditory spaceindependent of the elevation of asound source. The task oflocalizing a sound in azimuthalspace is often referred to as'lateralization'.
2 | JULY 2003 | VOLUME 4 www.nature.com/reviews/neuro
R E V I E W S
covering roughly the entire physiologically relevantrange13 (FIG. 3a,b). Accordingly, there is evidence for a systematic representation of azimuthal space at higherstations in the owl auditory system, the external nucleusof the auditory midbrain and the optic tectum. There,the best ITD of the neurons systematically shifts with theposition of the neurons, from rostral to caudal14–16. How-ever, our understanding of the bird ITD-encoding systemhas to be extended by one important factor: as I will dis-cuss later, GABA (γ-aminobutyric acid)-mediated inhibi-tion operates as a differential gain control system to keepthe coincidence detector in an appropriate working rangeand seems to compensate for other binaural influences.
The mammalian auditory system has also long beenconsidered as a ‘Jeffress-type’ system17–19. The evidence tosupport this view included the connection patterns withexcitatory inputs from both sides (details provided later),the cyclic nature of the ITD sensitivity when tested withpure tones (BOX 1), and the fact that the best ITD could bepredicted on the basis of the time delay of the response tothe monaural inputs from both ears17,20–22. Last but notleast, results from many human psychophysical studiescan be explained by the concept of binaural cross-correlation, as indicated by the Jeffress model23,24.
However, increasingly conflicting evidence began toraise considerable concerns about the validity of theJeffress model for the mammalian ITD encoder.A carefulanatomical reconstruction of single neurons that projectto the mammalian ITD detector, the medial superiorolive (MSO), revealed that far more anatomical projec-tion patterns did not fit the concept of delay lines thandid (see figure 6 in REF. 25; only 3 out of 16 MSO inputsresemble Jeffress-type delay lines. Note that the authorsof REF. 25 did not interpret their data to cast doubt onthe Jeffress model). Reconstructions from extracellular
station of binaural processing in the bird auditory brain-stem, act as coincidence detectors4–8 and receive theirexcitatory inputs through axons of systematically varyinglengths and with systematically varying interauraldelays5,9–12. As a consequence, neurons that are tuned tothe same characteristic (best) frequency show differentbest ITDs (the ITD at which a neuron fires maximally),
Cen
ozoi
cM
esoz
oic
Pal
aeoz
oic
Quaternary
Tertiary
Cretaceous
Jurassic
Triassic
Permian
Carboniferous
Devonian
1.8
65
146
208
250
290
360
410Rhipidistia
Early amphibians
DinosauriaPterosauriaPlesiosauriaIchthyosauria
AvesCrocodilia MammaliaSphenodontidae
Testudines SquamataAnuraUrodelaGymnophiona
'Stem reptiles'
Thecodontia
Pelycosauria
Therapsida
Amphibians
Appearance oftympanic ear
Archosaurs
Reptilians
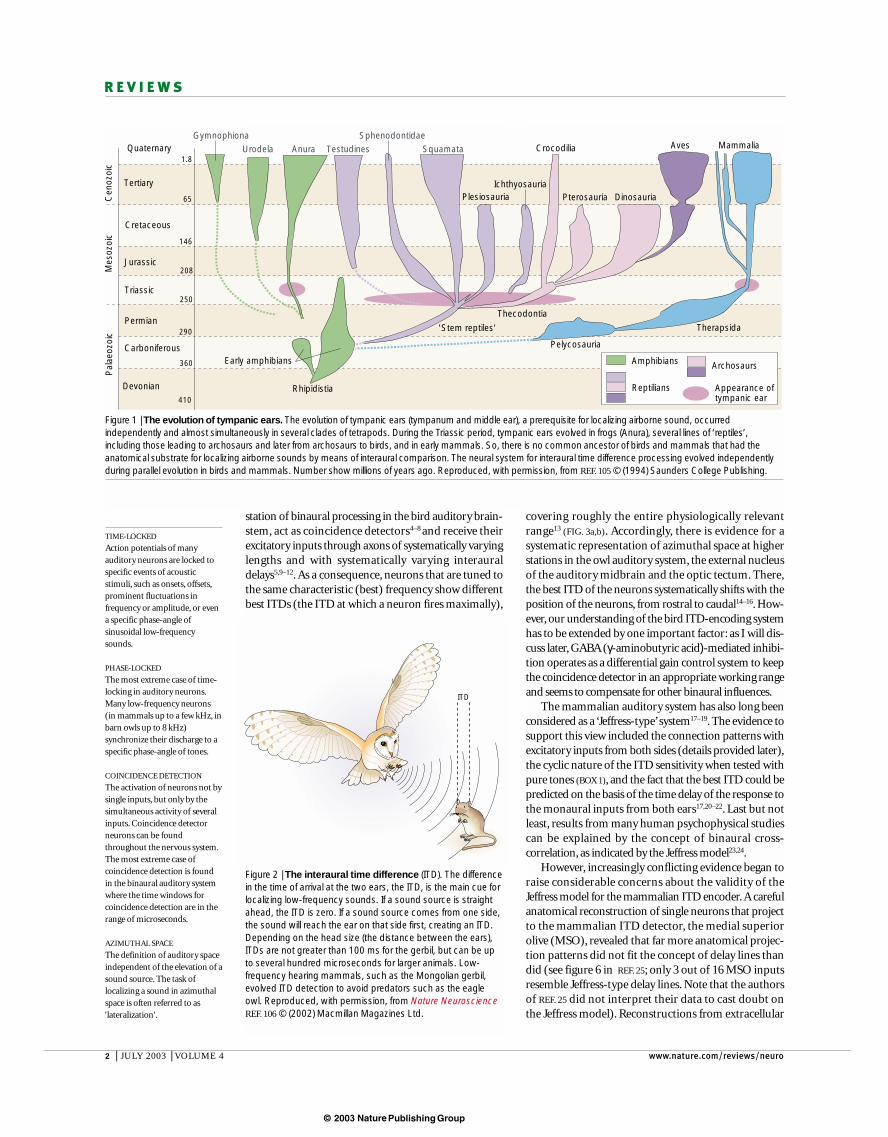
Figure 1 | The evolution of tympanic ears. The evolution of tympanic ears (tympanum and middle ear), a prerequisite for localizing airborne sound, occurredindependently and almost simultaneously in several clades of tetrapods. During the Triassic period, tympanic ears evolved in frogs (Anura), several lines of ‘reptiles’,including those leading to archosaurs and later from archosaurs to birds, and in early mammals. So, there is no common ancestor of birds and mammals that had theanatomical substrate for localizing airborne sounds by means of interaural comparison. The neural system for interaural time difference processing evolved independentlyduring parallel evolution in birds and mammals. Number show millions of years ago. Reproduced, with permission, from REF. 105 © (1994) Saunders College Publishing.
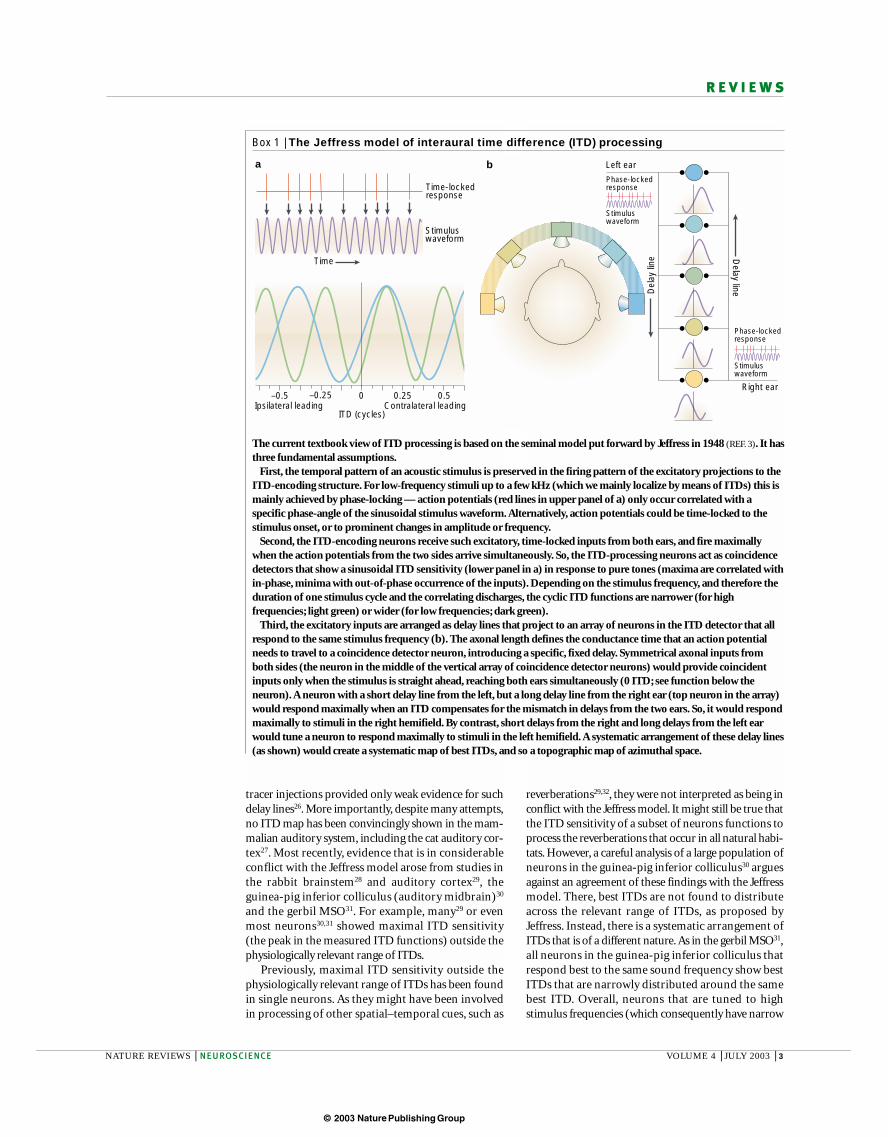
Figure 2 | The interaural time difference (ITD). The differencein the time of arrival at the two ears, the ITD, is the main cue forlocalizing low-frequency sounds. If a sound source is straightahead, the ITD is zero. If a sound source comes from one side,the sound will reach the ear on that side first, creating an ITD.Depending on the head size (the distance between the ears),ITDs are not greater than 100 ms for the gerbil, but can be upto several hundred microseconds for larger animals. Low-frequency hearing mammals, such as the Mongolian gerbil,evolved ITD detection to avoid predators such as the eagleowl. Reproduced, with permission, from Nature NeuroscienceREF. 106 © (2002) Macmillan Magazines Ltd.
ITD
© 2003 Nature Publishing Group
NATURE REVIEWS | NEUROSCIENCE VOLUME 4 | JULY 2003 | 3
R E V I E W S
reverberations29,32, they were not interpreted as being inconflict with the Jeffress model. It might still be true thatthe ITD sensitivity of a subset of neurons functions toprocess the reverberations that occur in all natural habi-tats. However, a careful analysis of a large population ofneurons in the guinea-pig inferior colliculus30 arguesagainst an agreement of these findings with the Jeffressmodel. There, best ITDs are not found to distributeacross the relevant range of ITDs, as proposed byJeffress. Instead, there is a systematic arrangement ofITDs that is of a different nature. As in the gerbil MSO31,all neurons in the guinea-pig inferior colliculus thatrespond best to the same sound frequency show bestITDs that are narrowly distributed around the samebest ITD. Overall, neurons that are tuned to high stimulus frequencies (which consequently have narrow
tracer injections provided only weak evidence for suchdelay lines26. More importantly, despite many attempts,no ITD map has been convincingly shown in the mam-malian auditory system, including the cat auditory cor-tex27. Most recently, evidence that is in considerableconflict with the Jeffress model arose from studies inthe rabbit brainstem28 and auditory cortex29, theguinea-pig inferior colliculus (auditory midbrain)30
and the gerbil MSO31. For example, many29 or evenmost neurons30,31 showed maximal ITD sensitivity (the peak in the measured ITD functions) outside thephysiologically relevant range of ITDs.
Previously, maximal ITD sensitivity outside thephysiologically relevant range of ITDs has been foundin single neurons. As they might have been involved in processing of other spatial–temporal cues, such as
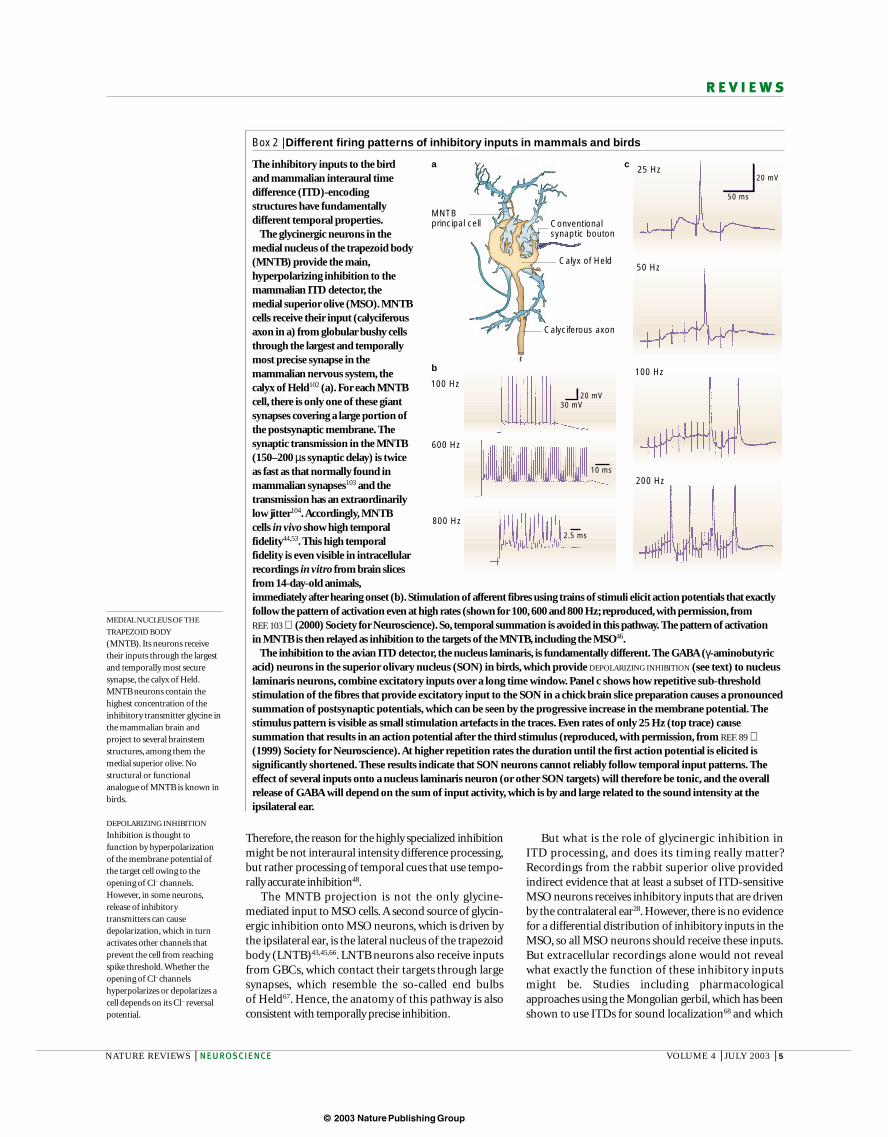
Box 1 | The Jeffress model of interaural time difference (ITD) processing
The current textbook view of ITD processing is based on the seminal model put forward by Jeffress in 1948 (REF. 3). It hasthree fundamental assumptions.
First, the temporal pattern of an acoustic stimulus is preserved in the firing pattern of the excitatory projections to theITD-encoding structure. For low-frequency stimuli up to a few kHz (which we mainly localize by means of ITDs) this ismainly achieved by phase-locking — action potentials (red lines in upper panel of a) only occur correlated with aspecific phase-angle of the sinusoidal stimulus waveform. Alternatively, action potentials could be time-locked to thestimulus onset, or to prominent changes in amplitude or frequency.
Second, the ITD-encoding neurons receive such excitatory, time-locked inputs from both ears, and fire maximallywhen the action potentials from the two sides arrive simultaneously. So, the ITD-processing neurons act as coincidencedetectors that show a sinusoidal ITD sensitivity (lower panel in a) in response to pure tones (maxima are correlated within-phase, minima with out-of-phase occurrence of the inputs). Depending on the stimulus frequency, and therefore theduration of one stimulus cycle and the correlating discharges, the cyclic ITD functions are narrower (for highfrequencies; light green) or wider (for low frequencies; dark green).
Third, the excitatory inputs are arranged as delay lines that project to an array of neurons in the ITD detector that allrespond to the same stimulus frequency (b). The axonal length defines the conductance time that an action potentialneeds to travel to a coincidence detector neuron, introducing a specific, fixed delay. Symmetrical axonal inputs fromboth sides (the neuron in the middle of the vertical array of coincidence detector neurons) would provide coincidentinputs only when the stimulus is straight ahead, reaching both ears simultaneously (0 ITD; see function below theneuron). A neuron with a short delay line from the left, but a long delay line from the right ear (top neuron in the array)would respond maximally when an ITD compensates for the mismatch in delays from the two ears. So, it would respondmaximally to stimuli in the right hemifield. By contrast, short delays from the right and long delays from the left earwould tune a neuron to respond maximally to stimuli in the left hemifield. A systematic arrangement of these delay lines(as shown) would create a systematic map of best ITDs, and so a topographic map of azimuthal space.
ITD (cycles)
0 0.25 0.5–0.25–0.5Ipsilateral leading Contralateral leading
Stimuluswaveform
Time-lockedresponse
Time
a b Left ear
Right ear
Phase-lockedresponse
Phase-lockedresponse
Stimuluswaveform
Stimuluswaveform
Del
ay li
ne
Delay line

© 2003 Nature Publishing Group
4 | JULY 2003 | VOLUME 4 www.nature.com/reviews/neuro
R E V I E W S
Glycine inhibition in the mammalian MSOThe major ITD-encoding structure in the mammalianauditory system, the MSO, receives direct excitatoryinputs from the ventral cochlear nuclei (VCN) on bothsides of the brainstem (FIG. 4a). The source of this bilateralexcitation is the spherical bushy cells (SBCs), which faithfully time-lock their discharges to the temporal pattern of sounds — for example, they provide phase-locked inputs in response to pure tones up to a fewkHz34,35. The SBCs project to the bipolar MSO cells, withipsilateral inputs making synapses on the lateral MSOdendrites and contralateral inputs making contact ontothe medial dendrites36–38. Such an arrangement isthought to improve binaural coincidence detection39.MSO cells show high-fidelity ITD sensitivity in all low-frequency hearing mammals tested so far17,20,22,28.
Although the first indirect evidence for inhibitionacting on low-frequency MSO cells goes back to the1960s20, undisputable anatomical evidence was not provided before the early 1990s40–44, and was shortly followed by direct physiological and pharmacologicalconfirmation in vitro45–47. The inhibition acting on MSOcells is predominantly, if not exclusively, mediated byglycine and is conveyed through two pathways. Thedominant pathway is from the MEDIAL NUCLEUS OF THE
TRAPEZOID BODY (MNTB; compare REF. 48; FIG. 4a). MNTBcells are driven by sound at the contralateral ear througha fast and effective pathway: they receive projectionsfrom globular bushy cells (GBCs) in the VCN, whichfollow the temporal structure of sounds with excep-tional precision (with high fidelity phase-locking to puretones49). The axon diameters of these projections are thelargest in the auditory brainstem50,51 and contact MNTBneurons through the largest, fastest and temporallymost secure synapses in the mammalian brain — thecalyces of Held52. As a result, MNTB cells also reliablyphase-lock to frequencies beyond 1 kHz44,53 (BOX 2).Therefore, they convert temporally precise excitationinto temporally precise inhibition.
Until recently, the MNTB represented an enigma,largely because the only target of MNTB cells that hasbeen intensively studied is the lateral superior olive(LSO), a nucleus juxtaposed to the MSO. The LSO is theinitial site of interaural intensity difference processing,which is used to localize high-frequency sounds.The underlying mechanism of this intensity differenceprocessing was considered to be pure subtraction ofcontralateral, glycine-mediated inhibition, provided bythe MNTB, from ipsilaterally driven excitation54,55. Withfew exceptions56,57, interaural intensity processing wasthought to require moderately fast but not outstandinglyprecise inhibition, and the existence of the calyx of Heldremained functionally unexplained. More recent studiesin the LSO show that timing does matter in adjustingthe binaural properties of these cells58,59 and there isindirect evidence that its kinetics are extraordinarilyfast28,60,61. But even more important is the finding thatMNTB neurons also project into two other areas, theventral nucleus of the lateral lemniscus62,63 and theMSO (see above), both of which are mainly, if notexclusively, involved in temporal processing53,64,65.
ITD functions) (BOX 1) prefer short ITDs. Neurons withlower best frequencies prefer longer ITDs. Across thepopulation of recorded cells, the mean best ITD corre-sponds to about 0.125 cycles of a sine wave at each neuron’s best frequency. This arrangement means thatthe steepest slope of ITD functions of most neuronsoccurs close to zero ITD and slightly shifted into thecontralateral hemifield. So, there are considerable problems with the idea of an azimuthal space map inmammals that is based on best ITDs as proposed byJeffress (FIG. 3c,d; compare REFS 27,33).
Not only is the representation of ITDs in mammalsinconsistent with the old textbook view, but there is alsoa conspicuous binaural inhibition innervating MSOneurons. Recent studies revealed a surprising role of thisinhibition in ITD processing that was not anticipated byany ITD model28,31.
–500 –200 0 200 500
–500 –200 0 200 500
ITD (µs)Left ear leadingRight ear leading
ITD (µs)Left ear leadingRight ear leading
Birds
Mammals
a b
c d
+ Excitation
Nature Reviews | Neuroscience
+
+++
+
+
+++
+
+
+––
– Inhibition
Figure 3 | Different strategies for encoding interaural time differences (ITDs). a | The birdITD-encoding structure, the nucleus laminaris (shown is the arrangement as found in the chick),operates in a way that is similar to the model suggested by Jeffress in 1948 (REF. 3). Arrays ofcoincidence detector neurons (coloured circles) on both sides of the brainstem receive excitatoryinputs (red lines) from the two ears (only inputs to the left nucleus laminaris are shown). Dependingon the axonal length (delay line) of the two inputs, each coincidence detector neuron respondsmaximally to a particular ITD, which compensates for differences in the internal neural delays. Asystematic arrangement of the delay lines from the contralateral ear creates a map of horizontalauditory space. b | The maxima of the ITD functions of different neurons in one nucleus laminarisdistribute across the physiologically relevant range (shaded area). The width of ITD functionsdepends on the stimulus frequency and, for lower frequencies, might be broader than shown. c | The mammalian ITD encoder is the medial superior olive (MSO; coloured circles), as described forguinea-pigs and gerbils. MSO neurons that are tuned to the same frequency preferentially respondto the same ITD. The ITD functions are adjusted to bring the maximal slope close to zero. Theopposite MSO is adjusted like a mirror image. Therefore, the relative activity of the entire populationof MSO cells, rather than the distribution within an MSO, represents the horizontal position of asound in space. Stimuli in the left hemifield mainly activate the right MSO, whereas stimuli in the righthemifield mainly activate the left MSO. Stimuli straight ahead activate both MSOs. The ratio of blueand yellow representing the auditory space in the figure indicates the relative activation of the left andright MSO. The ITD tuning is achieved by a complex interaction of binaural excitatory and inhibitoryinputs. d | ITD functions of MSO neurons are adjusted so that the maximal slope preferentiallyoccurs close to zero ITD. For a given frequency, the ITD tuning of the population of MSO neurons inthe left MSO mirrors that of the corresponding population of cells in the right MSO.
© 2003 Nature Publishing Group
NATURE REVIEWS | NEUROSCIENCE VOLUME 4 | JULY 2003 | 5
R E V I E W S
But what is the role of glycinergic inhibition in ITD processing, and does its timing really matter?Recordings from the rabbit superior olive providedindirect evidence that at least a subset of ITD-sensitiveMSO neurons receives inhibitory inputs that are drivenby the contralateral ear28. However, there is no evidencefor a differential distribution of inhibitory inputs in theMSO, so all MSO neurons should receive these inputs.But extracellular recordings alone would not revealwhat exactly the function of these inhibitory inputsmight be. Studies including pharmacologicalapproaches using the Mongolian gerbil, which has beenshown to use ITDs for sound localization68 and which
Therefore, the reason for the highly specialized inhibitionmight be not interaural intensity difference processing,but rather processing of temporal cues that use tempo-rally accurate inhibition48.
The MNTB projection is not the only glycine-mediated input to MSO cells. A second source of glycin-ergic inhibition onto MSO neurons, which is driven bythe ipsilateral ear, is the lateral nucleus of the trapezoidbody (LNTB)43,45,66. LNTB neurons also receive inputsfrom GBCs, which contact their targets through largesynapses, which resemble the so-called end bulbs of Held67. Hence, the anatomy of this pathway is alsoconsistent with temporally precise inhibition.
MEDIAL NUCLEUS OF THE
TRAPEZOID BODY
(MNTB). Its neurons receivetheir inputs through the largestand temporally most securesynapse, the calyx of Held.MNTB neurons contain thehighest concentration of theinhibitory transmitter glycine inthe mammalian brain andproject to several brainstemstructures, among them themedial superior olive. Nostructural or functionalanalogue of MNTB is known inbirds.
DEPOLARIZING INHIBITION
Inhibition is thought tofunction by hyperpolarizationof the membrane potential ofthe target cell owing to theopening of Cl– channels.However, in some neurons,release of inhibitorytransmitters can causedepolarization, which in turnactivates other channels thatprevent the cell from reachingspike threshold. Whether theopening of Cl– channelshyperpolarizes or depolarizes acell depends on its Cl– reversalpotential.
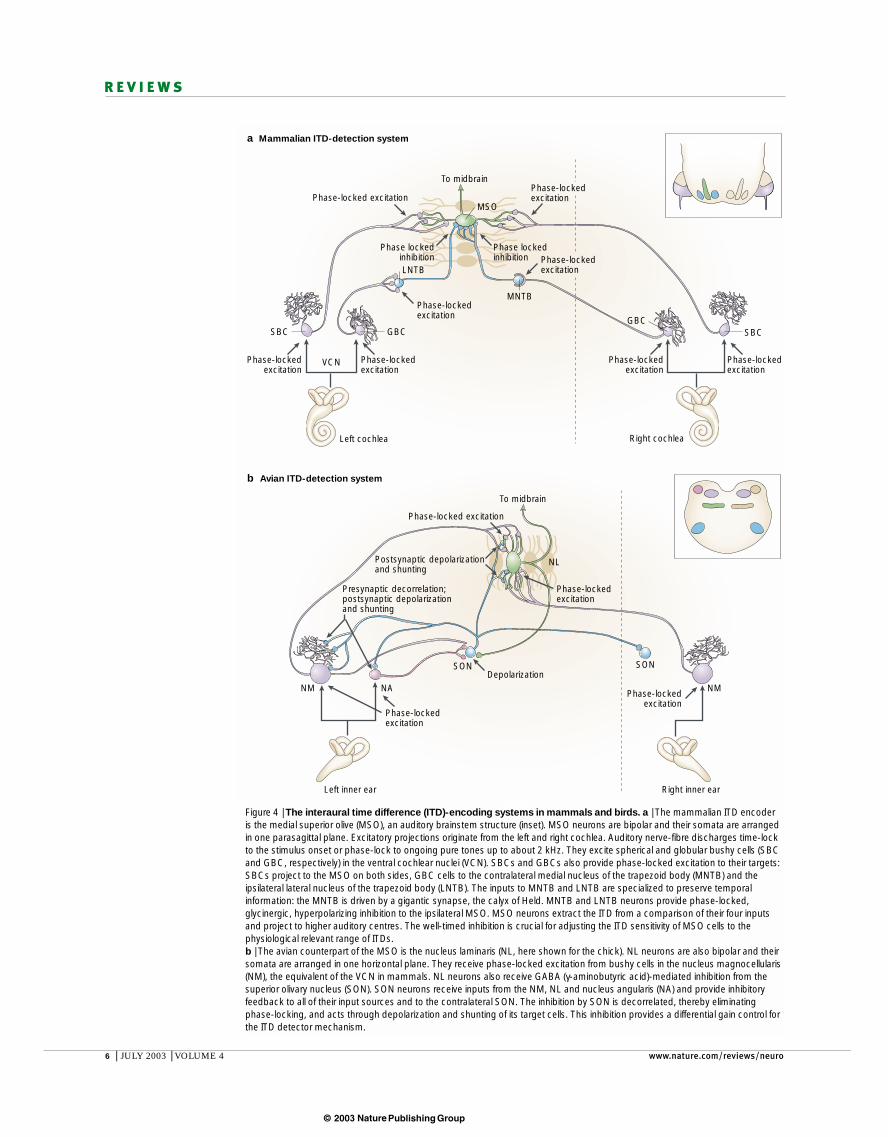
Box 2 | Different firing patterns of inhibitory inputs in mammals and birds
The inhibitory inputs to the birdand mammalian interaural timedifference (ITD)-encodingstructures have fundamentallydifferent temporal properties.
The glycinergic neurons in themedial nucleus of the trapezoid body(MNTB) provide the main,hyperpolarizing inhibition to themammalian ITD detector, themedial superior olive (MSO). MNTBcells receive their input (calyciferousaxon in a) from globular bushy cellsthrough the largest and temporallymost precise synapse in themammalian nervous system, thecalyx of Held102 (a). For each MNTBcell, there is only one of these giantsynapses covering a large portion ofthe postsynaptic membrane. Thesynaptic transmission in the MNTB(150–200 µs synaptic delay) is twiceas fast as that normally found inmammalian synapses103 and thetransmission has an extraordinarilylow jitter104.Accordingly, MNTBcells in vivo show high temporalfidelity44,53. This high temporalfidelity is even visible in intracellularrecordings in vitro from brain slicesfrom 14-day-old animals,immediately after hearing onset (b). Stimulation of afferent fibres using trains of stimuli elicit action potentials that exactlyfollow the pattern of activation even at high rates (shown for 100, 600 and 800 Hz; reproduced, with permission, from REF. 103 (2000) Society for Neuroscience). So, temporal summation is avoided in this pathway. The pattern of activation in MNTB is then relayed as inhibition to the targets of the MNTB, including the MSO46.
The inhibition to the avian ITD detector, the nucleus laminaris, is fundamentally different. The GABA (γ-aminobutyricacid) neurons in the superior olivary nucleus (SON) in birds, which provide DEPOLARIZING INHIBITION (see text) to nucleuslaminaris neurons, combine excitatory inputs over a long time window. Panel c shows how repetitive sub-thresholdstimulation of the fibres that provide excitatory input to the SON in a chick brain slice preparation causes a pronouncedsummation of postsynaptic potentials, which can be seen by the progressive increase in the membrane potential. Thestimulus pattern is visible as small stimulation artefacts in the traces. Even rates of only 25 Hz (top trace) causesummation that results in an action potential after the third stimulus (reproduced, with permission, from REF. 89 (1999) Society for Neuroscience). At higher repetition rates the duration until the first action potential is elicited issignificantly shortened. These results indicate that SON neurons cannot reliably follow temporal input patterns. Theeffect of several inputs onto a nucleus laminaris neuron (or other SON targets) will therefore be tonic, and the overallrelease of GABA will depend on the sum of input activity, which is by and large related to the sound intensity at theipsilateral ear.
a c
b
Calyx of Held
MNTB principal cell Conventional
synaptic bouton
Calyciferous axon
100 Hz
600 Hz
800 Hz
50 Hz
100 Hz
200 Hz
25 Hz 20 mV
50 ms
20 mV30 mV
10 ms
2.5 ms
© 2003 Nature Publishing Group
6 | JULY 2003 | VOLUME 4 www.nature.com/reviews/neuro
R E V I E W S
Phase lockedinhibition
Phase lockedinhibition
SBC SBCGBCGBC
VCN
To midbrain
To midbrain
MSO
Phase-lockedexcitation
Phase-lockedexcitation
Phase-lockedexcitation
Phase-lockedexcitation
Phase-lockedexcitation
Phase-lockedexcitation
Phase-lockedexcitation
Phase-lockedexcitation
Phase-lockedexcitation
Phase-lockedexcitation
Phase-locked excitation
Phase-locked excitation
LNTB
MNTB
Left cochlea
Left inner ear Right inner ear
Right cochlea
Depolarization
Presynaptic decorrelation;postsynaptic depolarizationand shunting
Postsynaptic depolarizationand shunting
NL
NM NMNA
SONSON
Mammalian ITD-detection systema
Avian ITD-detection systemb
Figure 4 | The interaural time difference (ITD)-encoding systems in mammals and birds. a | The mammalian ITD encoderis the medial superior olive (MSO), an auditory brainstem structure (inset). MSO neurons are bipolar and their somata are arrangedin one parasagittal plane. Excitatory projections originate from the left and right cochlea. Auditory nerve-fibre discharges time-lockto the stimulus onset or phase-lock to ongoing pure tones up to about 2 kHz. They excite spherical and globular bushy cells (SBCand GBC, respectively) in the ventral cochlear nuclei (VCN). SBCs and GBCs also provide phase-locked excitation to their targets:SBCs project to the MSO on both sides, GBC cells to the contralateral medial nucleus of the trapezoid body (MNTB) and theipsilateral lateral nucleus of the trapezoid body (LNTB). The inputs to MNTB and LNTB are specialized to preserve temporalinformation: the MNTB is driven by a gigantic synapse, the calyx of Held. MNTB and LNTB neurons provide phase-locked,glycinergic, hyperpolarizing inhibition to the ipsilateral MSO. MSO neurons extract the ITD from a comparison of their four inputsand project to higher auditory centres. The well-timed inhibition is crucial for adjusting the ITD sensitivity of MSO cells to thephysiological relevant range of ITDs. b | The avian counterpart of the MSO is the nucleus laminaris (NL, here shown for the chick). NL neurons are also bipolar and theirsomata are arranged in one horizontal plane. They receive phase-locked excitation from bushy cells in the nucleus magnocellularis(NM), the equivalent of the VCN in mammals. NL neurons also receive GABA (γ-aminobutyric acid)-mediated inhibition from thesuperior olivary nucleus (SON). SON neurons receive inputs from the NM, NL and nucleus angularis (NA) and provide inhibitoryfeedback to all of their input sources and to the contralateral SON. The inhibition by SON is decorrelated, thereby eliminatingphase-locking, and acts through depolarization and shunting of its target cells. This inhibition provides a differential gain control forthe ITD detector mechanism.

© 2003 Nature Publishing Group
NATURE REVIEWS | NEUROSCIENCE VOLUME 4 | JULY 2003 | 7
R E V I E W S
contralaterally driven excitation in a significant subset ofneurons64. In addition, ipsilaterally driven inhibition lagging the ipsilaterally driven excitation would shortenthe net excitatory potential that arises from ipsilateralstimulation, thereby adding to the effect of precedingcontralateral inhibition. Preliminary data confirm thatcontralateral inhibition precedes contralateral excita-tion and that ipsilateral inhibition is lagging ipsilateralexcitation71.
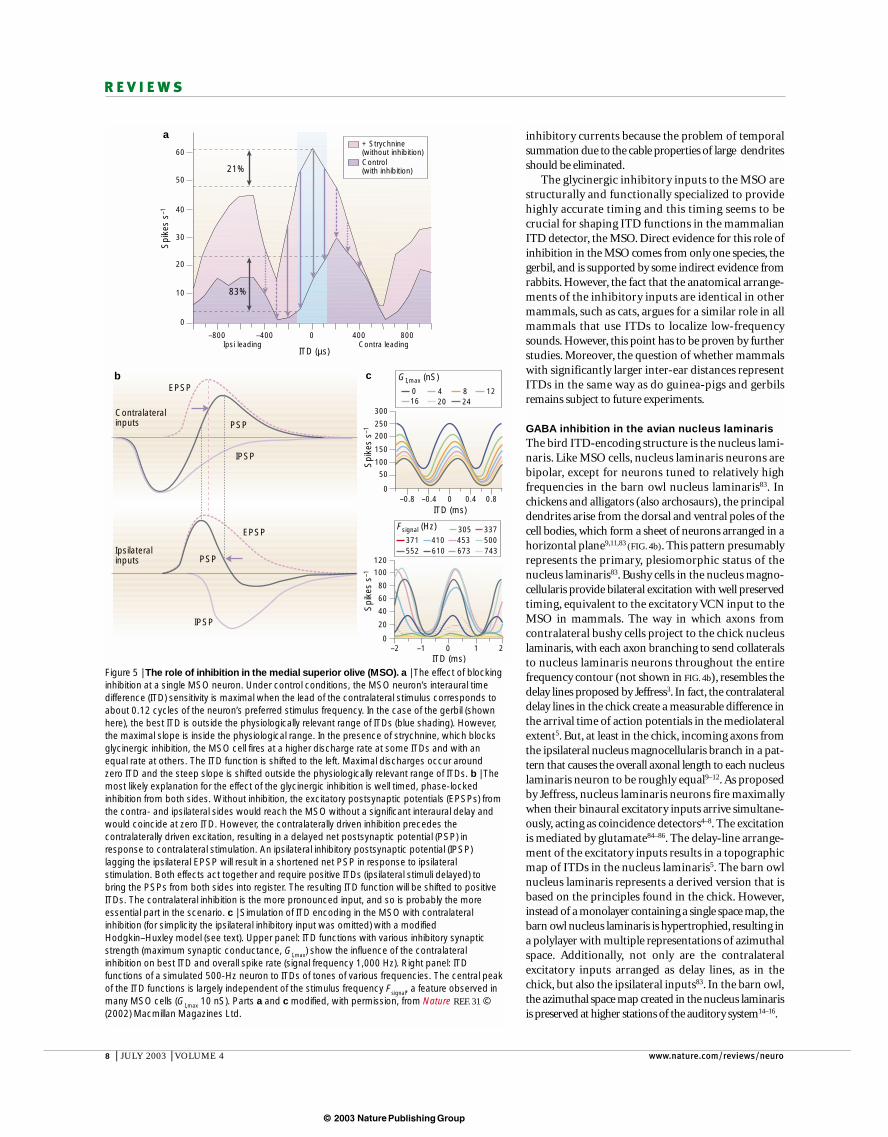
An earlier binaural model by Batra and colleagues28,and our own binaural model31, which was based on amodified Hodgkin–Huxley model72,73, incorporatedphase-locked inhibition from the MNTB. Such anarrangement could simulate the shift of the ITD functionas indicated earlier31. FIGURE 5c shows the results of one ofthese simulations. The crux of the models so far is thatone has to assume unusually fast conductance times forthe inhibitory input, with a time constant, τ
decay, in
the range of only a few hundred microseconds31.Estimations of in vitro recordings in rats indicate rela-tively fast conductance times, around 2 ms, for theglycinergic inhibition74. This is slower than required forthe proposed ITD-encoding mechanism, but there aretwo reasons to question whether this conductancereflects that in adult gerbils. First, the measurementswere performed in brain slices from juvenile animalsaround the time of hearing onset, several days before theinhibitory system in the superior olive matures69,75.Second, they were performed in rats, but it is unclearwhether rats use ITDs. The only published recordingsfrom rat MSO cells indicate that their temporal resolu-tion is in the range of a few milliseconds, by far inferior tothat in animals that are known to use ITDs76 and morelike that found in other non-ITD users, such as bats53.
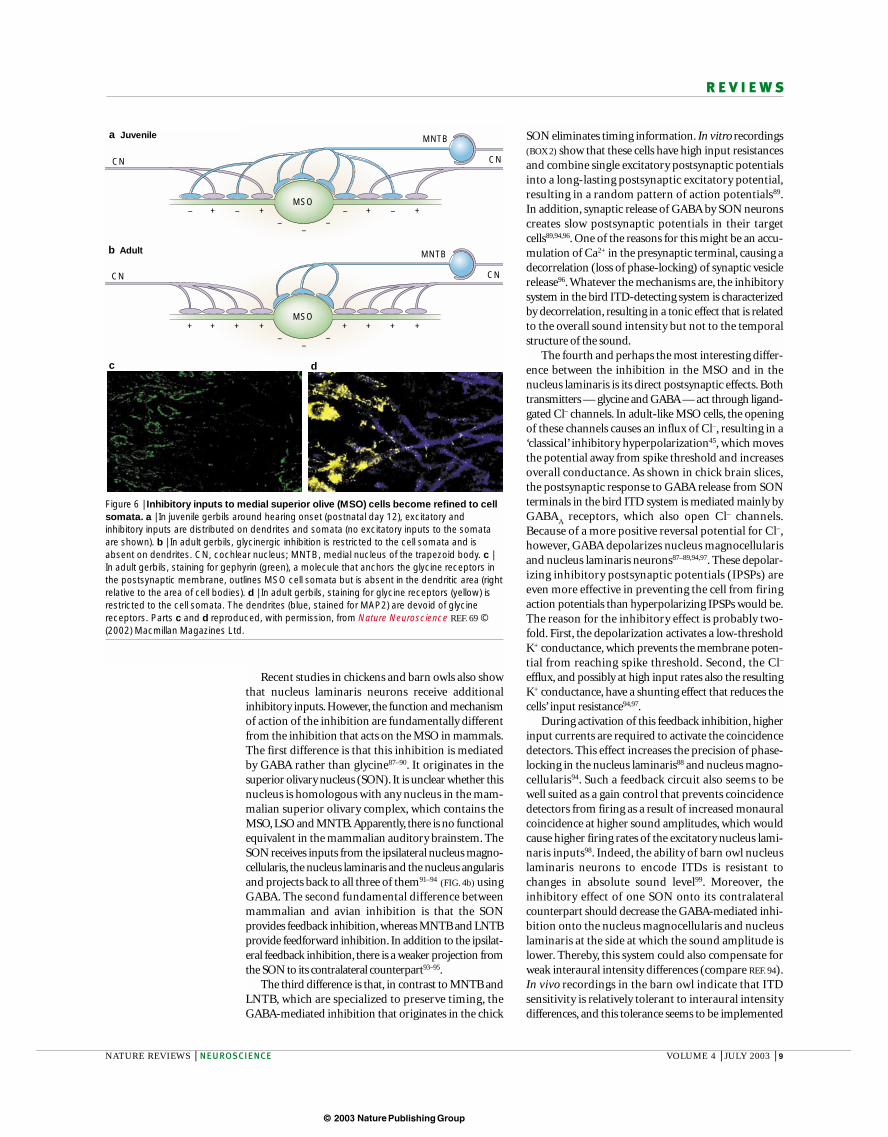
In accordance with this finding, the rat (and bat)MSO differs structurally from that in ITD-using mam-mals (such as gerbils or cats). The gerbil or cat MSOshows several anatomical specializations that probablyrepresent a structural correlate of, if not the basis for, thefunctional adaptation to process temporal informationin the microsecond range48,69. The most obvious struc-tural specialization is the strict alignment of the bipolarcell bodies in one sagittal plane, bringing the dendritesinto spatial register77. This might enable the axons togrow into the MSO in a uniform manner, to allow the‘right’ timing (which does not necessarily mean thatthey represent delay lines as proposed by Jeffress). Bycontrast, in non-ITD-using animals, MSO cells are notaligned in this way and the dendrites are therefore not in register48. The second structural specialization con-cerns the inhibitory inputs. In ITD-using mammals theglycine inputs are confined to the somata of MSOcells69,78–80, whereas they are evenly distributed acrosssomata and dendrites in non-ITD-using animals,including rats69. This confinement of inhibitory inputs isestablished only several days after hearing onset and is activity- and experience-dependent69 (FIG. 6). Thisdevelopment significantly departs from what is knownfrom the development of MNTB inputs to the LSO81,82.Functionally, the refinement of the inhibition on MSOneurons should increase the temporal acuity of the
has an MSO with all of the typical features of the mammalian ITD-processing system22,69, revealed anunexpected role of inhibition. In vitro recordings in a gerbil brainstem preparation had already shown that thetiming of the inhibition in the MSO is important for setting a time window for the coincidence detection ofthe excitatory inputs from both sides, and indicated a fastkinetic of the inhibitory postsynaptic potentials46. Theeffect of the inhibition, however, remained unclearbecause the in vitro approach did not allow the relativetiming of the four inputs to be assessed. Our recent in vivo recordings from single MSO neurons with andwithout pharmacological blockade of glycine-mediatedtransmission revealed a surprising function of the inhibition to the MSO31.
Under control conditions, the ITD sensitivity of mostneurons in the gerbil MSO was tuned as described earlier(FIG. 5a), with the maximal slope close to zero ITD and thebest ITDs corresponding to an interaural delay of 0.12cycles of the neurons’ best frequencies. The applicationof strychnine — an antagonist of glycinergic inhibition— caused an increase in MSO firing rates. Such anincrease in firing was expected, independent of any specific timing of the inhibition. However, this increasewas large at some ITDs but small or absent at otherITDs. As a consequence, the ITD functions shifted. Thenature of the shift was always the same: it moved themaximum of the ITD function close to zero ITD. As a consequence, the maximal slopes moved out of thephysiologically relevant range of ITDs (FIG. 5a).
From these results, two conclusions can be drawn.First, the excitatory inputs are generally adjusted so thatthe overall conductance delay from both sides is moreor less identical (how this is achieved during ontogenyis unknown). As a consequence, the MSO coincidencedetection based on excitation causes ITD functions topeak around zero ITD. Second, the glycinergic inhibitionadjusts the slope of the ITD function to the physiolo-gically relevant range, which gives a maximum amountof information because small changes in ITD causemaximal changes in discharge rates70. But how can thisbe explained in respect to the inhibitory inputs? A tonic, not phase-locked inhibition could account forsuch a shift only if the inhibition itself were already ITD sensitive. But the glycinergic inputs themselves aremonaural, so they are insensitive to ITDs. The onlyplausible explanation, therefore, is that the relative timing of one or both of the inhibitory inputs is thedefining factor.
FIGURE 5b shows how this timing could have the effectobserved in vivo. In this scenario, contralaterally driveninhibition precedes the excitation from the same side,thereby delaying the net excitatory postsynaptic potential.This would cause a lowering and a shift in the maximumof the ITD function to more positive ITDs (contralateralstimuli leading). Given the fast pathway from GBCsthrough the calyx synapse to the MNTB, which thencontacts only the MSO somata, such a temporal lead ofinhibition might be possible. In the bat MSO, the contralaterally driven glycinergic inhibition has beenshown to arrive simultaneously or even ahead of the
© 2003 Nature Publishing Group
8 | JULY 2003 | VOLUME 4 www.nature.com/reviews/neuro
R E V I E W S
inhibitory currents because the problem of temporalsummation due to the cable properties of large dendritesshould be eliminated.
The glycinergic inhibitory inputs to the MSO arestructurally and functionally specialized to providehighly accurate timing and this timing seems to be crucial for shaping ITD functions in the mammalianITD detector, the MSO. Direct evidence for this role ofinhibition in the MSO comes from only one species, thegerbil, and is supported by some indirect evidence fromrabbits. However, the fact that the anatomical arrange-ments of the inhibitory inputs are identical in othermammals, such as cats, argues for a similar role in allmammals that use ITDs to localize low-frequencysounds. However, this point has to be proven by furtherstudies. Moreover, the question of whether mammalswith significantly larger inter-ear distances representITDs in the same way as do guinea-pigs and gerbilsremains subject to future experiments.
GABA inhibition in the avian nucleus laminarisThe bird ITD-encoding structure is the nucleus lami-naris. Like MSO cells, nucleus laminaris neurons arebipolar, except for neurons tuned to relatively high frequencies in the barn owl nucleus laminaris83. Inchickens and alligators (also archosaurs), the principal dendrites arise from the dorsal and ventral poles of thecell bodies, which form a sheet of neurons arranged in ahorizontal plane9,11,83 (FIG. 4b). This pattern presumablyrepresents the primary, plesiomorphic status of thenucleus laminaris83. Bushy cells in the nucleus magno-cellularis provide bilateral excitation with well preservedtiming, equivalent to the excitatory VCN input to theMSO in mammals. The way in which axons from contralateral bushy cells project to the chick nucleuslaminaris, with each axon branching to send collaterals to nucleus laminaris neurons throughout the entire frequency contour (not shown in FIG. 4b), resembles thedelay lines proposed by Jeffress3. In fact, the contralateraldelay lines in the chick create a measurable difference inthe arrival time of action potentials in the mediolateralextent5. But, at least in the chick, incoming axons fromthe ipsilateral nucleus magnocellularis branch in a pat-tern that causes the overall axonal length to each nucleuslaminaris neuron to be roughly equal9–12. As proposedby Jeffress, nucleus laminaris neurons fire maximallywhen their binaural excitatory inputs arrive simultane-ously, acting as coincidence detectors4–8. The excitationis mediated by glutamate84–86. The delay-line arrange-ment of the excitatory inputs results in a topographicmap of ITDs in the nucleus laminaris5. The barn owlnucleus laminaris represents a derived version that isbased on the principles found in the chick. However,instead of a monolayer containing a single space map, thebarn owl nucleus laminaris is hypertrophied, resulting ina polylayer with multiple representations of azimuthalspace. Additionally, not only are the contralateral excitatory inputs arranged as delay lines, as in the chick, but also the ipsilateral inputs83. In the barn owl,the azimuthal space map created in the nucleus laminarisis preserved at higher stations of the auditory system14–16.
300
120
100
80
60
40
20
0
250
200
150
100
50
0
a
b c
60
50
40
30
20
10
0
–800 –400 0 400 800
Spi
kes
s–1
Spi
kes
s–1
Spi
kes
s–1
ITD (µs)
ITD (ms)
ITD (ms)
Ipsi leading Contra leading
21%
83%
+ Strychnine(without inhibition)Control(with inhibition)
EPSP
EPSP
IPSP
IPSP
PSP
PSP
Contralateralinputs
Ipsilateralinputs
016 20 24
4 8 12
305 337500453410371
552 610 673 743
GI,max (nS)
Fsignal (Hz)
–0.8 –0.4 0 0.4 0.8
–2 –1 0 1 2
Figure 5 | The role of inhibition in the medial superior olive (MSO). a | The effect of blockinginhibition at a single MSO neuron. Under control conditions, the MSO neuron’s interaural timedifference (ITD) sensitivity is maximal when the lead of the contralateral stimulus corresponds toabout 0.12 cycles of the neuron’s preferred stimulus frequency. In the case of the gerbil (shownhere), the best ITD is outside the physiologically relevant range of ITDs (blue shading). However,the maximal slope is inside the physiological range. In the presence of strychnine, which blocksglycinergic inhibition, the MSO cell fires at a higher discharge rate at some ITDs and with anequal rate at others. The ITD function is shifted to the left. Maximal discharges occur aroundzero ITD and the steep slope is shifted outside the physiologically relevant range of ITDs. b | Themost likely explanation for the effect of the glycinergic inhibition is well timed, phase-lockedinhibition from both sides. Without inhibition, the excitatory postsynaptic potentials (EPSPs) fromthe contra- and ipsilateral sides would reach the MSO without a significant interaural delay andwould coincide at zero ITD. However, the contralaterally driven inhibition precedes thecontralaterally driven excitation, resulting in a delayed net postsynaptic potential (PSP) inresponse to contralateral stimulation. An ipsilateral inhibitory postsynaptic potential (IPSP)lagging the ipsilateral EPSP will result in a shortened net PSP in response to ipsilateralstimulation. Both effects act together and require positive ITDs (ipsilateral stimuli delayed) tobring the PSPs from both sides into register. The resulting ITD function will be shifted to positiveITDs. The contralateral inhibition is the more pronounced input, and so is probably the moreessential part in the scenario. c | Simulation of ITD encoding in the MSO with contralateralinhibition (for simplicity the ipsilateral inhibitory input was omitted) with a modifiedHodgkin–Huxley model (see text). Upper panel: ITD functions with various inhibitory synapticstrength (maximum synaptic conductance, GI,max) show the influence of the contralateralinhibition on best ITD and overall spike rate (signal frequency 1,000 Hz). Right panel: ITDfunctions of a simulated 500-Hz neuron to ITDs of tones of various frequencies. The central peakof the ITD functions is largely independent of the stimulus frequency Fsignal, a feature observed inmany MSO cells (GI,max 10 nS). Parts a and c modified, with permission, from Nature REF. 31 ©(2002) Macmillan Magazines Ltd.
© 2003 Nature Publishing Group
NATURE REVIEWS | NEUROSCIENCE VOLUME 4 | JULY 2003 | 9
R E V I E W S
SON eliminates timing information. In vitro recordings(BOX 2) show that these cells have high input resistancesand combine single excitatory postsynaptic potentialsinto a long-lasting postsynaptic excitatory potential,resulting in a random pattern of action potentials89.In addition, synaptic release of GABA by SON neuronscreates slow postsynaptic potentials in their targetcells89,94,96. One of the reasons for this might be an accu-mulation of Ca2+ in the presynaptic terminal, causing adecorrelation (loss of phase-locking) of synaptic vesiclerelease96. Whatever the mechanisms are, the inhibitorysystem in the bird ITD-detecting system is characterizedby decorrelation, resulting in a tonic effect that is relatedto the overall sound intensity but not to the temporalstructure of the sound.
The fourth and perhaps the most interesting differ-ence between the inhibition in the MSO and in thenucleus laminaris is its direct postsynaptic effects. Bothtransmitters — glycine and GABA — act through ligand-gated Cl– channels. In adult-like MSO cells, the openingof these channels causes an influx of Cl–, resulting in a‘classical’ inhibitory hyperpolarization45, which movesthe potential away from spike threshold and increasesoverall conductance. As shown in chick brain slices,the postsynaptic response to GABA release from SONterminals in the bird ITD system is mediated mainly byGABA
Areceptors, which also open Cl– channels.
Because of a more positive reversal potential for Cl–,however, GABA depolarizes nucleus magnocellularisand nucleus laminaris neurons87–89,94,97. These depolar-izing inhibitory postsynaptic potentials (IPSPs) areeven more effective in preventing the cell from firingaction potentials than hyperpolarizing IPSPs would be.The reason for the inhibitory effect is probably two-fold. First, the depolarization activates a low-thresholdK+ conductance, which prevents the membrane poten-tial from reaching spike threshold. Second, the Cl–
efflux, and possibly at high input rates also the resultingK+ conductance, have a shunting effect that reduces thecells’ input resistance94,97.
During activation of this feedback inhibition, higherinput currents are required to activate the coincidencedetectors. This effect increases the precision of phase-locking in the nucleus laminaris88 and nucleus magno-cellularis94. Such a feedback circuit also seems to be well suited as a gain control that prevents coincidencedetectors from firing as a result of increased monauralcoincidence at higher sound amplitudes, which wouldcause higher firing rates of the excitatory nucleus lami-naris inputs98. Indeed, the ability of barn owl nucleuslaminaris neurons to encode ITDs is resistant tochanges in absolute sound level99. Moreover, theinhibitory effect of one SON onto its contralateralcounterpart should decrease the GABA-mediated inhi-bition onto the nucleus magnocellularis and nucleuslaminaris at the side at which the sound amplitude islower. Thereby, this system could also compensate forweak interaural intensity differences (compare REF. 94).In vivo recordings in the barn owl indicate that ITD sensitivity is relatively tolerant to interaural intensitydifferences, and this tolerance seems to be implemented
Recent studies in chickens and barn owls also showthat nucleus laminaris neurons receive additionalinhibitory inputs. However, the function and mechanismof action of the inhibition are fundamentally differentfrom the inhibition that acts on the MSO in mammals.The first difference is that this inhibition is mediated by GABA rather than glycine87–90. It originates in thesuperior olivary nucleus (SON). It is unclear whether thisnucleus is homologous with any nucleus in the mam-malian superior olivary complex, which contains theMSO, LSO and MNTB.Apparently, there is no functionalequivalent in the mammalian auditory brainstem. TheSON receives inputs from the ipsilateral nucleus magno-cellularis, the nucleus laminaris and the nucleus angularisand projects back to all three of them91–94 (FIG. 4b) usingGABA. The second fundamental difference betweenmammalian and avian inhibition is that the SON provides feedback inhibition, whereas MNTB and LNTBprovide feedforward inhibition. In addition to the ipsilat-eral feedback inhibition, there is a weaker projection fromthe SON to its contralateral counterpart93–95.
The third difference is that, in contrast to MNTB andLNTB, which are specialized to preserve timing, theGABA-mediated inhibition that originates in the chick
N t R i | N i
Juvenile MNTB
MNTB
a
Adultb
– + – + – + – +–
––
+ + + + + + + +–
––
CNCN
CN CN
MSO
MSO
dc
Figure 6 | Inhibitory inputs to medial superior olive (MSO) cells become refined to cellsomata. a | In juvenile gerbils around hearing onset (postnatal day 12), excitatory andinhibitory inputs are distributed on dendrites and somata (no excitatory inputs to the somataare shown). b | In adult gerbils, glycinergic inhibition is restricted to the cell somata and isabsent on dendrites. CN, cochlear nucleus; MNTB, medial nucleus of the trapezoid body. c |In adult gerbils, staining for gephyrin (green), a molecule that anchors the glycine receptors inthe postsynaptic membrane, outlines MSO cell somata but is absent in the dendritic area (rightrelative to the area of cell bodies). d | In adult gerbils, staining for glycine receptors (yellow) isrestricted to the cell somata. The dendrites (blue, stained for MAP2) are devoid of glycinereceptors. Parts c and d reproduced, with permission, from Nature Neuroscience REF. 69 ©(2002) Macmillan Magazines Ltd.

© 2003 Nature Publishing Group
10 | JULY 2003 | VOLUME 4 www.nature.com/reviews/neuro
R E V I E W S
1. Clack, J. A. The evolution of tetrapod ears and the fossilrecord. Brain Behav. Evol. 50, 198–212 (1997).An excellent review of our current knowledge of theindependent evolution of the tetrapod middle ears. It revises the old but still often stated view that themammalian three-ossicle middle ear derived from areptilian-like one-ossicle middle ear.
2. Clack, J. A. Patterns and processes in the early evolution ofthe tetrapod ear. J. Neurobiol. 53, 251–264 (2002).
3. Jeffress, L. A. A place theory of sound localization. J. Comp.Physiol. Psychol. 41, 35–39 (1948).One of the most elegant and influential models incomputational neuroscience. It provided theconceptual framework for several anatomical,physiological, psychophysical and theoreticalstudies related to sound localization in mammalsand birds.
4. Carr, C. E. & Konishi, M. A circuit for detection of interauraltime differences in the brain stem of the barn owl. J. Neurosci. 10, 3227–3246 (1990).
5. Overholt, E., Rubel, E. W. & Hyson, R. L. A circuit for codinginteraural time differences in the chick brainstem. J. Neurosci. 12, 1698–1708 (1992).
6. Joseph, A. W. & Hyson, R. L. Coincidence detection bybinaural neurons in the chick brain stem. J. Neurophysiol.69, 1197–1211 (1993).
7. Reyes, A. D., Rubel, E. W. & Spain, W. J. In vitro analysis ofoptimal stimuli for phase-locking and time-delayedmodulation of firing in avian nucleus laminaris neurons. J. Neurosci. 16, 993–1007 (1996).
8. Kuba, H., Koyano, K. & Ohmori, H. Development ofmembrane conductance improves coincidence detection inthe nucleus laminaris of the chicken. J. Physiol. (Lond.) 540,529–542 (2002).
9. Parks, T. N. & Rubel, E. W. Organization and development ofbrain stem auditory nuclei of the chicken: organization ofprojections from n. magnocellularis to n. laminaris. J. Comp.Neurol. 164, 435–448 (1975).
10. Rubel, E. W. & Parks, T. N. Organization and development ofbrain stem auditory nuclei of the chicken: tonotopicorganization of n. magnocellularis and n. laminaris. J. Comp.Neurol. 164, 411–433 (1975).
11. Young, E. D. & Rubel, E. W. Frequency-specific projectionsof individual neurons in chick brainstem auditory nuclei. J. Neurosci. 3, 1373–1378 (1983).
12. Young, E. D. & Rubel, E. W. Embryogenesis of arborizationpattern and topography of individual axons in N. laminaris of thechicken brain stem. J. Comp. Neurol. 245, 425–459 (1986).
13. Wagner, H., Mazer, J. A. & von Campenhausen, M.Response properties of neurons in the core of the centralnucleus of the inferior colliculus of the barn owl. Eur. J. Neurosci. 15, 1343–1352 (2002).
14. Knudsen, E. I. & Konishi, M. Center-surround organization ofauditory receptive fields in the owl. Science 202, 778–780(1978).
15. Brainard, M. S. & Knudsen, E. I. Experience-dependentplasticity in the inferior colliculus: a site for visual calibrationof the neural representation of auditory space in the barnowl. J. Neurosci. 13, 4589–4608 (1993).
16. Gold, J. I. & Knudsen, E. I. A site of auditory experience-dependent plasticity in the neural representation of auditoryspace in the barn owl’s inferior colliculus. J. Neurosci. 20,3469–3486 (2000).
17. Yin, T. C. & Chan, J. C. Interaural time sensitivity in medialsuperior olive of cat. J. Neurophysiol. 64, 465–488 (1990).A careful description of a significant number of MSOcells in vivo. It provided the strongest physiologicalevidence so far in favour of a Jeffress-type ITD-detection mechanism in the MSO.
18. Joris, P. X., Smith, P. H. & Yin, T. C. Coincidence detection inthe auditory system: 50 years after Jeffress. Neuron 21,1235–1238 (1998).
19. Moushegian, G., Rupert, A. L. & Gidda, J. S. Functionalcharacteristics of superior olivary neurons to binaural stimuli.J. Neurophysiol. 38, 1037–1048 (1975).
20. Goldberg, J. M. & Brown, P. B. Response of binauralneurons of dog superior olivary complex to dichotic tonalstimuli: some physiological mechanisms of soundlocalization. J. Neurophysiol. 32, 613–636 (1969).
21. Crow, G., Rupert, A. L. & Moushegian, G. Phase locking inmonaural and binaural medullary neurons: implications forbinaural phenomena. J. Acoust. Soc. Am. 64, 493–501(1978).
22. Spitzer, M. W. & Semple, M. N. Neurons sensitive tointeraural phase disparity in gerbil superior olive: diversemonaural and temporal response properties. J. Neurophysiol. 73, 1668–1690 (1995).
23. Stern, R. M. & Trahiotis, C. in Hearing (ed. Moore, B. C. J.)347–386 (Academic, New York, 1995).
24. Palmer, A. R. & Shackleton, T. M. The physiological basis forthe binaural masking level difference. Acta Acust. Unit.Acust. 88, 312–319 (2002).
25. Smith, P. H., Joris, P. X. & Yin, T. C. Projections ofphysiologically characterized spherical bushy cell axons fromthe cochlear nucleus of the cat: evidence for delay lines to themedial superior olive. J. Comp. Neurol. 331, 245–260 (1993).A thorough anatomical study addressing the questionof the existence of delay-lines in mammals (althoughB.G. does not agree with their conclusion).
26. Beckius, G. E., Batra, R. & Oliver, D. L. Axons fromanteroventral cochlear nucleus that terminate in medialsuperior olive of cat: observations related to delay lines. J. Neurosci. 19, 3146–3161 (1999).
27. Middlebrooks, J. C., Xu, L., Furukawa, S. & Macpherson, E. A.Cortical neurons that localize sounds. Neuroscientist 8,73–83 (2002).
28. Batra, R., Kuwada, S. & Fitzpatrick, D. C. Sensitivity tointeraural temporal disparities of low- and high-frequencyneurons in the superior olivary complex. I. Heterogeneity ofresponses. J. Neurophysiol. 78, 1222–1236 (1997).A detailed description of different types of ITDsensitivity in the superior olivary complex. This paperis particularly important because it comes from thefirst (and so far only) recordings from SONs in awakeanimals. Additionally, it provides the first indirect in vivo evidence for inhibitory effects on ITD functionsin a mammal that uses ITDs to localize low-frequencysounds.
29. Fitzpatrick, D. C., Kuwada, S. & Batra, R. Neural sensitivityto interaural time differences: beyond the Jeffress model. J. Neurosci. 20, 1605–1615 (2000).
30. McAlpine, D., Jiang, D. & Palmer, A. R. A neural code forlow-frequency sound localization in mammals. NatureNeurosci. 4, 396–401 (2001).A new way of analysing data from ITD-sensitiveneurons in the guinea-pig auditory midbrain provided
the basis for a new concept concerning therepresentation of ITDs in the mammalian auditorysystem.
31. Brand, A., Behrend, O., Marquardt, T., McAlpine, D. & Grothe,B. Precise inhibition is essential for microsecond interauraltime difference coding. Nature 417, 543–547 (2002).The first direct evidence for the involvement ofglycinergic inhibition in ITD processing in the MSO invivo. Pharmacological blockade of glycinergicinhibition caused a significant shift in the ITDsensitivity of single MSO cells.
32. Grothe, B. & Neuweiler, G. The function of the medialsuperior olive in small mammals: temporal receptive fields inauditory analysis. J. Comp. Physiol. A 186, 413–423 (2000).
33. McAlpine, D. & Grothe, B. Sound localisation and delay lines— do mammals fit the model? Trends Neurosci. (in the press).
34. Warr, W. B. Fiber degeneration following lesions in theanterior ventral cochlear nucleus of the cat. Exp. Neurol. 14,453–474 (1966).
35. Osen, K. K. Cytoarchitecture of the cochlear nuclei in thecat. J. Comp. Neurol. 136, 453–484 (1969).
36. Stotler, W. A. An experimental study of the cells andconnections of the superior olivary complex of the cat. J. Comp. Neurol. 98, 401–432 (1953).
37. Lindsey, B. G. Fine structure and distribution of axonterminals from the cochlear nucleus on neurons in themedial superior olivary nucleus of the cat. J. Comp. Neurol.160, 81–103 (1975).
38. Kitzes, L. M., Kageyama G. H., Semple, M. N. & Kil, J.Development of ectopic projections from the ventralcochlear nucleus to the superior olivary complex induced byneonatal ablation of the contralateral cochlea. J. Comp.Neurol. 353, 341–363 (1995).
39. Agmon-Snir, H., Carr, C. E. & Rinzel, J. The role of dendrites inauditory coincidence detection. Nature 393, 268–272 (1998).
40. Cant, N. B. in Neurobiology of Hearing: The Central AuditorySystem (eds Altschuler, R. A., Bobbin, R. P., Clopton, B. M.& Hoffman, D. W.) 99–119 (Raven, New York, 1991).
41. Covey, E., Vater, M. & Casseday, J. H. Binaural properties ofsingle units in the superior olivary complex of the mustachedbat. J. Neurophysiol. 66, 1080–1094 (1991).
42. Kuwabara, N. & Zook, J. M. Projections to the medialsuperior olive from the medial and lateral nuclei of thetrapezoid body in rodents and bats. J. Comp. Neurol. 324,522–538 (1992).
43. Cant, N. B. & Hyson, R. L. Projections from the lateralnucleus of the trapezoid body to the medial superior olivarynucleus in the gerbil. Hear. Res. 58, 26–34 (1992).
44. Smith, P. H., Joris, P. X. & Yin, T. C. Anatomy and physiologyof principal cells of the medial nucleus of the trapezoid body(MNTB) of the cat. J. Neurophysiol. 79, 3127–3142 (1998).
45. Grothe, B. & Sanes, D. H. Bilateral inhibition by glycinergicafferents in the medial superior olive. J. Neurophysiol. 69,1192–1196 (1993).
46. Grothe, B. & Sanes, D. H. Synaptic inhibition influences thetemporal coding properties of medial superior olivary neurons:an in vitro study. J. Neurosci. 14, 1701–1709 (1994).The first in vitro evidence for well-timed glycinergicinhibition that could influence the ITD tuning of MSOcells.
47. Smith, P. H. Structural and functional differences distinguishprincipal from nonprincipal cells in the guinea pig MSO slice.J. Neurophysiol. 73, 1653–1667 (1995).
at the level of the nucleus laminaris100.An open questionis whether GABA
B receptors might also contribute to
such gain controlin birds101.
ConclusionsIn contrast to the traditional textbook view, whichassumes that the ITD-processing system is alike in birdsand mammals, and functions according to the Jeffressmodel, recent evidence strongly indicates that birds andmammals have their own, profoundly different solutionsfor the same problem of how to localize sounds byusing ITDs. These differences concern the mode ofneural representation of ITDs as well as the role of inhi-bition in extracting ITDs. It will be important toexplain why the systems evolved in such different ways,and to what extent common precursor systems have
been crucial for this development. A system such as theITD-processing one — which performs such anextreme task and, therefore, has only a few degrees offreedom — might help us to understand why systemsevolve in one way and not in another, simply becausethe questions we can ask are unusually specific. Therewill also be the challenge to explain, at a cellular level,how the incredible specificity of the role of inhibition intemporal processing can be achieved and how it isadjusted during ontogeny. In particular, the role of audi-tory experience in the development of glycinergic inhi-bition in the MSO begs the question of how inhibitorysynapses can be selected on the basis of how they influ-ence temporal resolution in a sub-millisecond range. Tosolve these problems will be an important challenge inthe coming years of research in sensory systems.

© 2003 Nature Publishing Group
NATURE REVIEWS | NEUROSCIENCE VOLUME 4 | JULY 2003 | 11
R E V I E W S
48. Grothe, B. The evolution of temporal processing in themedial superior olive, an auditory brainstem structure. Prog.Neurobiol. 61, 581–610 (2000).This review addresses the question of the evolution ofITD processing in the mammalian auditory system. Itprovides an overview of the role of the MSO and itsinhibitory inputs in mammals that do not use ITDs forsound localization.
49. Smith, P. H., Joris, P. X., Carney, L. H. & Yin, T. C.Projections of physiologically characterized globular bushycell axons from the cochlear nucleus of the cat. J. Comp.Neurol. 304, 387–407 (1991).
50. Harrison, J. M. & Warr, W. B. A study of the cochlear nucleiand ascending auditory pathways of the medulla. J. Comp.Neurol. 119, 341–380 (1962).
51. Morest, D. K. The collateral system of the medial nucleus ofthe trapezoid body of the cat, its neuronal architecture andrelation to the olivo-cochlear bundle. Brain Res. 9, 288–311(1968).
52. von Gersdorff, H. & Borst, J. G. Short-term plasticity at thecalyx of Held. Nature Rev. Neurosci. 3, 53–64 (2002).
53. Grothe, B. & Park, T. J. Sensitivity to interaural time differencesin the medial superior olive of a small mammal, the Mexicanfree-tailed bat. J. Neurosci. 18, 6608–6622 (1998).
54. Boudreau, J. C. & Tsuchitani, C. Binaural interaction in the catsuperior olive S segment. J. Neurophysiol. 31, 442–454(1968).
55. Moore, M. J. & Caspary, D. M. Strychnine blocks binauralinhibition in lateral superior olivary neurons. J. Neurosci. 3,237–242 (1983).
56. Yin, T. C., Hirsch, J. A. & Chan, J. C. Responses of neuronsin the cat’s superior colliculus to acoustic stimuli. II. A modelof interaural intensity sensitivity. J. Neurophysiol. 53,746–758 (1985).
57. Pollak, G. D. Time is traded for intensity in the bat’s auditorysystem. Hear. Res. 36, 107–124 (1988).
58. Park, T. J., Grothe, B., Pollak, G. D., Schuller, G. & Koch, U.Neural delays shape selectivity to interaural intensitydifferences in the lateral superior olive. J. Neurosci. 16,6554–6566 (1996).
59. Irvine, D. R., Park, V. N. & McCormick, L. Mechanismsunderlying the sensitivity of neurons in the lateral superiorolive to interaural intensity differences. J. Neurophysiol. 86,2647–2666 (2001).
60. Joris, P. X. & Yin, T. C. Envelope coding in the lateral superiorolive. I. Sensitivity to interaural time differences. J. Neurophysiol. 73, 1043–1062 (1995).
61. Batra, R., Kuwada, S. & Fitzpatrick, D. C. Sensitivity tointeraural temporal disparities of low- and high-frequencyneurons in the superior olivary complex. II. Coincidencedetection. J. Neurophysiol. 78, 1237–1247 (1997).
62. Glendenning, K. K., Brunso-Bechtold, J. K., Thompson, G. C.& Masterton, R. B. Ascending auditory afferents to the nuclei ofthe lateral lemniscus. J. Comp. Neurol. 197, 673–703 (1981).
63. Covey, E. & Casseday, J. H. Connectional basis forfrequency representation in the nuclei of the laterallemniscus of the bat Eptesicus fuscus. J. Neurosci. 6,2926–2940 (1986).
64. Grothe, B. Interaction of excitation and inhibition inprocessing of pure tone and amplitude-modulated stimuli inthe medial superior olive of the mustached bat. J. Neurophysiol. 71, 706–721 (1994).
65. Covey, E. & Casseday, J. H. The monaural nuclei of thelateral lemniscus in an echolocating bat: parallel pathwaysfor analyzing temporal features of sound. J. Neurosci. 11,3456–3470 (1991).
66. Spirou, G. A. & Berrebi, A. S. Glycine immunoreactivity in thelateral nucleus of the trapezoid body of the cat. J. Comp.Neurol. 383, 473–488 (1997).
67. Spirou, G. A. & Berrebi, A. S. Organization of ventrolateralperiolivary cells of the cat superior olive as revealed by pep-19immunocytochemistry and nissl stain. J. Comp. Neurol.368, 100–120 (1996).
68. Heffner, R. S. & Heffner, H. E. Sound localization and use ofbinaural cues by the gerbil (Meriones unguiculatus). Behav.Neurosci. 102, 422–428 (1988).
69. Kapfer, C., Seidl, A. H., Schweizer, H. & Grothe, B.Experience-dependent refinement of inhibitory inputs toauditory coincidence-detector neurons. Nature Neurosci. 5,247–253 (2002).The study presents evidence for a structural changein the inhibitory MSO inputs after hearing onset. Itshows that this change depends on auditoryexperience during a critical period during ontogeny.
70. Skottun, B. C., Shackleton, T. M., Arnott, R. H. & Palmer, A. R.The ability of inferior colliculus neurons to signal differencesin interaural delay. Proc. Natl Acad. Sci. USA 98,14050–14054 (2001).
71. Brand, A. Precise Temporal Processing in the GerbilAuditory Brainstem. Thesis, Munich Univ. (2003).
72. Carney, L. H. A model for the responses of low-frequencyauditory-nerve fibers in cat. J. Acoust. Soc. Am. 93,401–417 (1993).
73. Brughera, A. R., Stutman, E. R., Carney, L. H. & Colburn, H. S.A model with exitation and inhibition for cells in the medialsuperior olive. Aud. Neurosci. 2, 219–233 (1996).
74. Smith, A. J., Owens, S. & Forsythe, I. D. Characterisation ofinhibitory and excitatory postsynaptic currents of the ratmedial superior olive. J. Physiol. (Lond.) 529, 681–698 (2000).
75. Sanes, D. H. The development of synaptic function andintegration in the central auditory system. J. Neurosci. 13,2627–2637 (1993).
76. Inbody, S. B. & Feng, A. S. Binaural response characteristicsof single neurons in the medial superior olivary nucleus of thealbino rat. Brain Res. 210, 361–366 (1981).
77. Ramón y Cajal, S. Histologie du Systeme Nerveux del’Homme et des Vertebrates (Malonie, Paris, 1907).
78. Clark, G. M. The ultrastructure of nerve endings in the medialsuperior olive of the cat. Brain Res. 14, 293–305 (1969).
79. Perkins, R. E. An electron microscopic study of synapticorganization in the medial superior olive of normal and experi-mental chinchillas. J. Comp. Neurol. 148, 387–415 (1973).
80. Russell, F. A. & Moore, D. R. Ultrastructural transynapticeffects of unilateral cochlear ablation in the gerbil medialsuperior olive. Hear. Res. 173, 43–61 (2002).
81. Sanes, D. H. & Friauf, E. Development and influence ofinhibition in the lateral superior olivary nucleus. Hear. Res.147, 46–58 (2000).
82. Kim, G. & Kandler, K. Elimination and strengthening ofglycinergic/GABAergic connections during tonotopic mapformation. Nature Neurosci. 6, 282–290 (2003).
83. Carr, C. E. & Soares, D. Evolutionary convergence andshared computational principles in the auditory system.Brain Behav. Evol. 59, 294–311 (2002).
84. Levin, M. D., Kubke, M. F., Schneider, M., Wenthold, R. &Carr, C. E. Localization of AMPA-selective glutamatereceptors in the auditory brainstem of the barn owl.J. Comp. Neurol. 378, 239–253 (1997).
85. Kubke, M. F. & Carr, C. E. Development of AMPA-selectiveglutamate receptors in the auditory brainstem of the barnowl. Microsc. Res. Tech. 41, 176–186 (1998).
86. Ravindranathan, A. et al. Contrasting molecular compositionand channel properties of AMPA receptors on chick auditoryand brainstem motor neurons. J. Physiol. (Lond.) 523,667–684 (2000).
87. Hyson, R. L., Reyes, A. D. & Rubel, E. W. A depolarizinginhibitory response to GABA in brainstem auditory neuronsof the chick. Brain Res. 677, 117–126 (1995).The first description of the depolarizing nature of theGABA-mediated inhibition in the bird nucleus laminaris.
88. Funabiki, K., Koyano, K. & Ohmori, H. The role ofGABAergic inputs for coincidence detection in the neuronesof nucleus laminaris of the chick. J. Physiol. (Lond.) 508,851–869 (1998).In vitro recordings indicate that the GABA-mediatedinhibition in the bird ITD detector increases themembrane conductance and thereby improves thecoincidence detection of the excitatory inputs.
89. Yang, L. C., Monsivais, P. & Rubel, E. W. The superior olivary nucleus and its influence on nucleus laminaris: a sourceof inhibitory feedback for coincidence detection in the avianauditory brainstem. J. Neurosci. 19, 2313–2325 (1999).
Anatomical and physiological characterization of theSON — the source of GABA-mediated inputs to the birdITD detector neurons in the nucleus laminaris. The dataindicate that the SON provides feedback inhibition thatdoes not follow the temporal structure of a sound, andis suited to function as a gain control and to improvethe resolution of coincidence detection.
90. Takahashi, Y. & Konishi, M. Manipulation of inhibition in theowl’s nucleus laminaris and its effects on optic tectumneurons. Neuroscience 111, 373–378 (2002).
91. Takahashi, T. T. & Konishi, M. Projections of nucleusangularis and nucleus laminaris to the lateral lemniscalnuclear complex of the barn owl. J. Comp. Neurol. 274,212–238 (1988).
92. Carr, C. E., Fujita, I. & Konishi, M. Distribution of GABAergicneurons and terminals in the auditory system of the barnowl. J. Comp. Neurol. 286, 190–207 (1989).
93. Lachica, E. A., Rubsamen, R. & Rubel, E. W. GABAergicterminals in nucleus magnocellularis and laminaris originatefrom the superior olivary nucleus. J. Comp. Neurol. 348,403–418 (1994).
94. Monsivais, P., Yang, L. & Rubel, E. W. GABAergic inhibition innucleus magnocellularis: implications for phase locking in theavian auditory brainstem. J. Neurosci. 20, 2954–2963 (2000).
95. Westerberg, B. D. & Schwarz, D. W. Connections of thesuperior olive in the chicken. J. Otolaryngol. 24, 20–30 (1995).
96. Lu, T. & Trussell, L. O. Inhibitory transmission mediated byasynchronous transmitter release. Neuron 26, 683–694(2000).
97. Monsivais, P. & Rubel, E. W. Accommodation enhancesdepolarizing inhibition in central neurons. J. Neurosci. 21,7823–7830 (2001).Functional analysis of the depolarizing GABA-mediated inhibition in the chick auditory brainstemindicates that this mechanism has an advantage overhyperpolarizing inhibition, at least in neurons withlarge excitatory conductances.
98. Reed, M. C. & Durbeck, L. Delay lines and auditoryprocessing. Comments Theor. Biol. 3, 441–461 (1995).
99. Pena, J. l., Viete, S., Albeck, Y. & Konishi, M. Tolerance tosound intensity of binaural coincidence detection in thenucleus laminaris of the owl. J. Neurosci. 16, 7046–7054(1996).
100. Viete, S., Pena, J. L. & Konishi, M. Effects of interauralintensity difference on the processing of interaural timedifference in the owl’s nucleus laminaris. J. Neurosci. 17,1815–1824 (1997).
101. Brenowitz, S., David, J. & Trussell, L. Enhancement ofsynaptic efficacy by presynaptic GABAB receptors. Neuron20, 135–141 (1998).
102. Morest, D. K. The growth of synaptic endings in themammalian brain: a study of the calyces of the trapezoidbody. Z. Anat. Entwicklungsgesch. 127, 201–220 (1968).
103. Taschenberger, H. & von Gersdorff, H. Fine-tuning anauditory synapse for speed and fidelity: developmentalchanges in presynaptic waveform, EPSC kinetics, andsynaptic plasticity. J. Neurosci. 20, 9162–9173 (2000).
104. Guinan, J. J. Jr & Li, R. Y. Signal processing in brainstemauditory neurons which receive giant endings (calyces ofHeld) in the medial nucleus of the trapezoid body of the cat.Hear. Res. 49, 321–334 (1990).
105. Walker, W. F. & Liem, K. F. Functional Anatomy ofVertebrates: An Evolutionary Perspective (Saunders CollegePublishing, Orlando, 1994).
106. Sanes, D. H. Right place at the right time. Nature Neurosci.5, 187–188 (2002).
Online links
FURTHER INFORMATIONEncyclopedia of Life Sciences: http://www.els.net/prey detection by bats and owlsBenedikt Grothe’s Lab:http://www.neuro.mpg.de/researchgr/audiproc/Access to this interactive links box is free online.













![Learning efficient task-dependent representations with synaptic … · 2020-06-25 · successfully explained features of early sensory representations [2, 3] in terms of the interaction](https://img.pdfslide.us/doc/110x75/5f3286a5106bbe4e15205811/learning-efficient-task-dependent-representations-with-synaptic-2020-06-25-successfully.jpg)















