Embed Size (px)
Citation preview

Indirect interactions between alien and native
Senecio species as mediated by insects
Evelyn White (B. App. Sci. Hons)
PhD candidate
2008
School of Natural Resource Sciences
Queensland University of Technology


3
Statement of original authorship
To the best of my knowledge and belief, the work contained in this thesis has not
previously been submitted to meet requirements for an award at this or any other
higher education institution. The thesis contains no material previously published or
written by another person, except where due reference is made. All chapters
presented are published or submitted manuscripts, each of which has multiple
authors. The roles of the co-authors on the manuscripts, which vary from project
supervision through to active data collection, are acknowledged at the start of each
chapter.
Signature
Date


5
Abstract
The studies described in this thesis investigate the role of indirect effects in
invasion biology. The Introduction provides a brief overview of indirect effects and
an outline of the thesis structure. The role of indirect effects in the context of
invasion biology is addressed in an in-depth published literature review that
comprises the second chapter, providing a theoretical background for the
subsequent empirical studies. Chapters Three to Six are comprised of manuscripts
that have been published or are under review or in press, which describe studies
that investigate the importance of indirect effects in invasion biology using a model
system consisting of the alien Asteraceae Senecio madagascariensis, a closely-
related native, Senecio pinnatifolius, and the insect species with which they
interact. Senecio madagascariensis and S. pinnatifolius occur in a similar
geographic range in eastern Australia and these studies were conducted in mixed
and pure populations of the two species. The herbivore and floral visitor
assemblages of the two Senecio species at seven field sites in South-east
Queensland were compared using sweep-net sampling, manual searching and
floral visitor observation techniques. The floral visitor assemblages were similar
between the two species, comprised largely of species of Syrphidae and the
European honeybee, Apis mellifera. Herbivore assemblages, however, were highly
variable both between species and between sites, with greater herbivore
abundance and diversity recorded on the native S. pinnatifolius than its alien
congener. The most commonly recorded herbivores were sap-sucking species
such as Myridae. The magpie moth, Nyctemera amica was the most common
folivore on both Senecio species and laboratory studies demonstrated a clear
preference by ovipositing females and feeding larvae of this species for the native
Senecio species, over the alien. Field surveys supported these findings, recording

6
greater leaf damage on the native species than the invader. Herbivory levels were
lower, rather than higher, in mixed populations than in pure populations, thus there
was no evidence that the presence of one species enhanced herbivory in the other.
Field pollination trials were conducted to determine whether competition for
pollinators or facilitation of pollination occurred in mixed Senecio populations. The
presence of the native S. pinnatifolius affected pollinator visitation rates to the alien
Senecio; bee visits to S. madagascariensis were significantly reduced by the
presence of S. pinnatifolius, whilst syrphid visits increased. However, altered
visitation rates were not reflected in seed set. The presence of the alien species
had no impact on pollinator visits to the native. Surprisingly, S. pinnatifolius seed
set was higher in mixed populations than in pure populations. This might be due to
abiotic factors, lower rates of herbivory at these sites or transfer of pollen between
species resulting in the production of hybrid seed (if S. madagascariensis has
greater male fitness). Hybridisation in the field was investigated using AFLP
techniques. No mature hybrid plants were recorded in mixed populations, but
hybrid seeds were produced by both species. Senecio pinnatifolius maternal
parents produced higher numbers of hybrid seed than expected based on the
relative frequencies of the two species, whilst hybridisation in S. madagascariensis
was lower than expected. This may indicate greater male fitness of the invader.
A range of complex indirect interactions can occur between invasive and native
species, with these interactions having the potential to influence the success or
failure of the invader and its impacts on co-occurring natives. The Discussion
addresses the findings of the studies described here in the context of invasion
biology theory.

7
Keywords
Exotic; herbivory; higher order interactions; hybridisation; indirect effects; insect-
plant interactions; invasion biology; invasive species; pollination; weed.


9
Table of Contents
List of Tables and Figures.....................................................................................13
Acknowledgements...........................................................................................15
Chapter 1 - Introduction ........................................................................................17
1.1 Description of research problem investigated ........................................19
1.1.1 Background ...................................................................................19
1.1.2 The role of biotic indirect effects in invasion biology.......................20
1.2 Overall objectives of the study..............................................................22
1.2.1 A model system: alien and native Senecio .....................................22
1.2.2 Specific aims of the study ..............................................................24
1.3 Thesis outline and presentation.............................................................25
1.3.1 Thesis presentation........................................................................25
1.3.2 Account of research progress linking the research papers.............26
Chapter 2 - Biotic indirect effects: a neglected concept in invasion biology ....27
2.1 Abstract .................................................................................................29
2.2 Introduction............................................................................................29
2.3 Mechanisms for indirect effects in biological invasions ..........................33
2.3.1 Apparent competition .....................................................................33
2.3.2 Indirect mutualism / facilitation .......................................................44
2.3.3 Exploitative competition .................................................................45
2.3.4 Trophic cascades...........................................................................47
2.4 Discussion.............................................................................................48
Chapter 3 - Diversity and abundance of arthropod floral visitor and herbivore
assemblages on alien and native Senecio species .............................................55
3.1 Abstract .................................................................................................57
3.2 Introduction............................................................................................58
3.3 Materials and Methods ..........................................................................60

10
3.3.1 Study species................................................................................ 60
3.3.2 Study sites .................................................................................... 61
3.3.3 Survey Methodology...................................................................... 62
3.4 Results.................................................................................................. 65
3.4.1 Floral visitor assemblages and abundance.................................... 65
3.4.2 Herbivore assemblages and abundance ....................................... 69
3.5 Discussion ............................................................................................ 72
3.5.1 Conclusions................................................................................... 74
Chapter 4 - A test of the enemy release hypothesis: The native magpie moth
prefers a native fireweed (Senecio pinnatifolius) to its introduced congener
(S. madagascarensis)............................................................................................ 77
4.1 Abstract ................................................................................................ 79
4.2 Introduction........................................................................................... 80
4.3 Materials and Methods.......................................................................... 82
4.3.1 Larval Feeding and Survival .......................................................... 82
4.3.2 Larval Preference.......................................................................... 83
4.3.3 Oviposition Preference .................................................................. 84
4.3.4 Foliage damage ............................................................................ 84
4.3.5 Plant characteristics ...................................................................... 86
4.4 Results.................................................................................................. 86
4.4.1 Larval feeding and survival............................................................ 86
4.4.2 Larval preference .......................................................................... 87
4.4.3 Oviposition preference .................................................................. 87
4.4.4 Foliage damage ............................................................................ 88
4.4.5 Plant characteristics ...................................................................... 88
4.5 Discussion ............................................................................................ 89
4.5.1 Conclusions................................................................................... 92

11
Chapter 5 - Plant-pollinator interactions in sympatric exotic and native
Senecio species: Is facilitation or competition for pollinators occurring?........93
5.1 Abstract .................................................................................................95
5.2 Introduction............................................................................................95
5.3 Materials and Methods ..........................................................................98
5.3.1 Study species ................................................................................98
5.3.2 Study sites .....................................................................................99
5.3.3 Methods.......................................................................................100
5.4 Results ................................................................................................104
5.4.1 Quantity of pollen on insects ........................................................104
5.4.2 Vegetation structure.....................................................................104
5.4.3 Visitation rates and plant characteristics ......................................106
5.4.4 Pollinator visits.............................................................................106
5.4.5 Seed set ......................................................................................109
5.5 Discussion...........................................................................................110
5.5.1 Conclusions .................................................................................112
Chapter 6 - Can hybridisation cause local extinction: the case for
demographic swamping of the Australian native, Senecio pinnatifolius, by
the invasive S. madagascariensis? ....................................................................115
6.1 Abstract ...............................................................................................117
6.2 Introduction..........................................................................................118
6.3 Materials and Methods ........................................................................121
6.3.1 Study species ..............................................................................121
6.3.2 Study sites and sample collections ..............................................121
6.3.3 Reciprocal crossing experiments .................................................123
6.3.4 Relative densities of plants and flowers .......................................125
6.3.5 AFLP profiling ..............................................................................125
6.3.6 Data analysis – population level...................................................126

12
6.3.7 Data analysis – individual level.................................................... 126
6.3.8 Risk posed by hybridisation......................................................... 128
6.4 Results................................................................................................ 129
6.4.1 Reciprocal crossing experiments................................................. 129
6.4.2 Relative densities of plants and flowers....................................... 129
6.4.3 Genetic diversity and population differentiation ........................... 130
6.4.4 Risk posed by hybridisation......................................................... 134
6.5 Discussion .......................................................................................... 137
6.5.1 The incidence of hybridisation and fate of hybrids ....................... 137
6.5.2 Long-term population impacts of hybridisation – genetic diversity
and differentiation ....................................................................................... 138
6.5.3 What does the future hold for S. pinnatifolius? ............................ 139
Chapter 7 – General Discussion........................................................................ 143
7.1 Pollinator-mediated indirect interactions.............................................. 145
7.2 Herbivore-mediated indirect interactions ............................................. 149
7.3 Implications for management of invasive species................................ 151
7.4 Conclusions ........................................................................................ 154
Appendices.......................................................................................................... 157
Appendix A..................................................................................................... 157
Appendix B..................................................................................................... 161
Appendix C..................................................................................................... 165
Appendix D..................................................................................................... 167
References........................................................................................................... 169

13
List of Tables and Figures
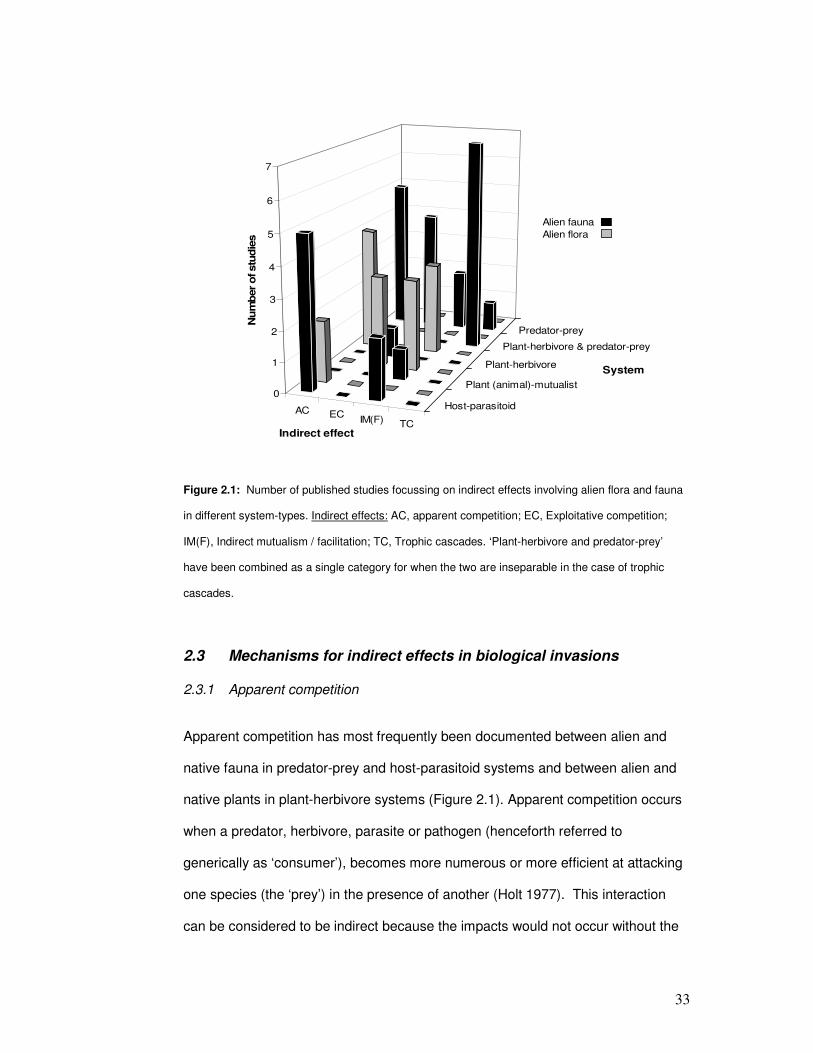
Figure 2.1: Number of published studies focussing on indirect effects involving alien flora &
fauna. ………………………………………………………………………………………………..
35
Table 2.1: Studies documenting indirect effects between invasive & native species.…………. 39
Figure 3.1: Mean number of floral visitors per plant per five min time period……………………. 67
Figure 3.2: Mean floral visitor species richness per plant ………………………………………….. 68
Figure 3.3: Dendrogram: Hierarchical, average linkage cluster analysis for floral visitor
assemblages………………………………………………………………………………………..
69
Table 3.1: Sørenson similarity indices for floral visitor & herbivore assemblages between
populations of Senecio pinnatifolius & S. madagascariensis………. …………………………
70
Figure 3.4: Mean number of herbivores per plant in four Senecio pinnatifolius populations …. 71
Figure 3.5: Mean herbivore species richness per plant……………………….…………………….. 72
Figure 3.6: Dendrogram: hierarchical, average linkage cluster analysis for arthropod herbivore
assemblages…………………………………...........................................................................
73
Figure 4.1: Mean proportion of feeding Nyctemera amica larvae on two Senecio species…….. 89
Figure 4.2: Mean proportion of damaged leaves per plant in each population type………… 90
Table 4.1: Vegetation structure in pure & mixed stands……………………………………………. 91
Table 5.1: Vegetation structure in pure & mixed stands……………..……………………………… 106
Table 5.2: Results of stepwise regression analyses for (i) amount of time spent per plant & (ii)
visitation rate, by bees & syrphids to Senecio plants in mixed & pure stands. ………...........
107
Figure 5.1: Bee visits per plant in pure & mixed stands……………………………………………. 109
Figure 5.2: Syrphid visits per plant in pure & mixed stands………………………………………… 110
Figure 5.3: Seeds set per capitulum in pure & mixed stands………………………………………. 111
Table 6.1: Population locations & relative densities of plants & flowers of Senecio pinnatifolius
& S. madagascariensis……………………………………………………………………………..
126
Table 6.2: Seed viability & amount of seed produced from reciprocal crosses between Senecio
pinnatifolius & S. madagascariensis. …………………………………………………………….
132
Figure 6.1: Unrooted neighbour-joining phenogram based on pairwise FST distances among
AFLP profiles for Senecio in sympatric & allopatric sites……………………………………….
133
Figure 6.2: Clustering of Senecio pinnatifolius & S. madagascariensis in sympatric & allopatric
sites…………………………………………………………………………………….………….
134

14
Figure 6.3: Percentage of plants of each species, capitula produced by each species & hybrid
& non-hybrid progeny produced by Senecio pinnatifolius & Senecio madagascariensis …
135
Figure 6.4: Annual viable seed production of Senecio pinnatifolius & S. madagascariensis in
sympatric sites……………………………………………………………………………………….
136
Table 6.3: Annual viable seed production by Senecio pinnatifolius & S. madagascariensis in
sympatric populations for a range of hybridisation scenarios…………………………………..
138

Acknowledgements
Thanks to postgraduate students and staff from the QUT School of Natural
Resource Sciences, particularly: Mike Duffy, Liz Dunlop and Alexis Wilson, for
assistance with fieldwork, as well as Nikki Sims, Amy Lawson, Helen Nahrung,
Mark Schutz and Peter Prentis for their great contribution to lab work and
assistance in the field. Thanks also to landholders and their families: the O’Reillys,
Helen Hall, Ernest Diamond, Sue Gordon, Ken Hack, Phil Curtis, Ray Cavanaugh,
Paul and Kylie Stumkat for granting me permission to work on their land and
assisting with locating plant populations. Ailsa Holland (Queensland Herbarium,
EPA), Ian Radford (Department of Environment and Conservation, WA) and
Rachel McFadyen (Weeds CRC) provided information and advice about Senecio
species and study sites. I gratefully acknowledge suggestions and comments made
on thesis drafts by Tanya Scharaschkin and Peter Mather (both of QUT) and on
submitted manuscripts by Rachel McFadyen (Weed CRC), Michael Bull (Flinders
University), Andy Shepherd (CSIRO) and several anonymous reviewers.
In particular I am very grateful to my supervisors, John Wilson and Tony Clarke for
their support, advice, and constructive input at every stage of the project.


Chapter 1 - Introduction


19
Chapter 1 – Introduction
1.1 Description of research problem investigated
1.1.1 Background
Increasing numbers of species are invading new environments worldwide. In
addition to having an economic impact on agriculture, invaders often have adverse
impacts on the biodiversity and functioning of ecological systems (Mooney and
Cleland 2001). In order to improve our understanding of invasions, thereby
allowing us to better predict, prevent and manage invasions, many workers have
addressed the questions: What makes a species invasive? What makes a system
invasible? And which species are likely to have the greatest impact on the native
biota (Crawley 1987; Mooney and Cleland 2001; Crooks 2002; Chornesky and
Randall 2003; Levine et al. 2003; Ricciardi 2003)?1
Various authors (starting with Darwin 1859) have argued that alien species more
distantly related to the native species in an area should be more successful as
invaders than aliens closely related to native species. This is in part because the
former are likely to be different in their resource utilisation, thereby avoiding
competition with natives (Simberloff 1986; Rejmánek 1998). In keeping with this
view, most empirical studies that have addressed biotic interactions associated
with invasions have concentrated on the more ‘obvious’ direct interactions between
alien and native species, such as competition (Fogarty and Facelli 1999; Jensen et
al. 2002; Cadi and Joly 2003; Kolb and Alpert 2003; Corbin and D’Antonio 2004;
Miller and Gorchov 2004) and predation (Savidge 1987; Dickman 1996; Wilson et
al. 1998; Kinnear et al. 2002; Kinzler and Maier 2003).
1 Terminology varies somewhat between the manuscripts comprising this thesis. Within these
manuscripts the terms ‘invasive species’,’introduced species’, ‘exotic’ and ‘alien’ all refer to any species that has established outside of its natural range and whose presence results in environmental or economic damage (Csurhes 1995; Davis and Thompson 2000, 2001).

20
However, Richardson et al. (2000) argue that mainstream ecology over
emphasises the role of negative interactions, particularly competition, in invasion
biology. They provide empirical examples in support of the view that positive biotic
interactions are of greater importance than competition, and that alien species
closely related to native species are more likely to be successful than
taxonomically isolated invaders, due to their ability to utilise local mutualists,
including pollinators, dispersers and mycorrhizal fungi.
There is, therefore, a lack of consensus on the factors that determine a species’
invasion potential and the impacts of an invader on native species. Clearly the
search for a single, simple explanation (for instance the competitive ability of a
potential invader in a certain situation; or a simple set of species traits) is unlikely to
provide a full elucidation of the invasion process or the subsequent impacts on
native biota (Mack 1996; Lavorel et al. 1999). Several workers have suggested that
the mechanisms of invasion are far more complex than has been acknowledged to
date, involving interactions between multiple species traits and multiple features of
the new system (Schierenbeck et al. 1994; Richardson et al. 2000; White et al.
2006).
1.1.2 The role of biotic indirect effects in invasion biology
There is increasing empirical evidence in support of the view that both the invasion
potential of a species, and its impacts on native biota, are influenced by a range of
complex biotic interactions, including those collectively termed ‘indirect effects’ (or
indirect interactions). Little is known about the role of indirect effects in structuring
communities, and even less is known about their role in invasions. Indirect effects
involve more than two species and are generally defined as ‘how one species
alters the effect that another species has on a third’ (Strauss 1991). Specifically, an
indirect effect is occurring when the presence of one species alters the abundance

21
or behaviour of a second intermediary species, which in turn has an impact on the
abundance, per capita growth rate, or genotype of a third species (Strauss 1991).
Such interactions include trophic cascades, apparent competition, indirect
mutualisms and some kinds of exploitative competition, all of which have been
documented in plant-herbivore and plant-pollinator systems.
Plant-herbivore interactions can affect both invasion success and the impacts of an
invader on a system. For instance, selective herbivory on one plant species can
give another plant species a competitive advantage (Brown 1994; Carson and Root
2000; Centre et al. 2005). When a native species is grazed in preference to a co-
occuring alien species, an invasion can be indirectly facilitated (Cross 1981;
Edwards et al. 2000). Conversely, native plant species may benefit through
preferential grazing of alien plants (Zancola et al. 2000). Furthermore, established
native plant-herbivore interactions in a system may be altered by the presence of a
new plant species, with potentially negative consequences for native plants. This
can occur via ‘apparent competition’, which occurs when a predator or herbivore
becomes more numerous or more effective at consuming one species in the
presence of another (Holt 1977). This topic is discussed in detail in Chapter Two.
Plant-pollinator interactions can also influence invasion success, and the impacts
of an alien plant species on a system. Richardson et al. (2000) argue that
invasions are rarely constrained by lack of required mutualisms (e.g. by pollinator
limitation), since most such relationships are generalised rather than being tightly
co-evolved. However, as yet, little research has been directed towards this area of
invasion biology. There is no doubt however, that once an invasive plant species
has established in an area, pollinators can mediate indirect interactions between
the alien and co-occurring natives. For instance, alien species can compete with
natives for the services of pollinators (Chittka and Schurkens 2001); interspecific

22
pollen transfer can result in reduced seed set for one or both plant species (Brown
and Mitchell 2001); and when pollen is transferred between closely related alien
and native species, hybridisation may occur (Vila et al. 2003), resulting in a range
of consequences for native biota.
Only in the last decade or so, have researchers begun to recognise the importance
of indirect effects in structuring ecological systems, and the potential role of these
complex interactions in invasion biology. Further work is required for us to gain a
more complete understanding of indirect interactions in invasion biology, and to
improve our ability to predict the full range of impacts of an alien species on natives
in its new range. The studies described in this thesis address indirect interactions
using a model system comprised of a native and an alien Senecio species, and the
insects with which they interact.
1.2 Overall objectives of the study
1.2.1 A model system: alien and native Senecio
Senecioneae is one of the largest tribes in the family Asteraceae, containing
around 150 genera, and over 3,000 species. The Senecioneae, described in detail
by Thomspon (2006), are herbs or shrubs with distinctive involucral bract
morphology. Around 50% of species in this tribe are currently assigned to the
genus Senecio (Pelser et al. 2006). Senecio species tend to be cosmopolitan,
inhabiting disturbed areas (Lawrence 1985), and are found as weeds in various
parts of the world (Fernandez and Verona 1984; Marohasy 1989; Garcia-Serrano
et al. 2004, 2005).
A number of studies have addressed interactions between invasive and native
Senecio species, investigating relative competitive abilities under different levels of
abiotic resources (Garcia-Serrano et al. 2007), comparative life-history traits

23
(Radford and Cousens 2000; Sans et al. 2004; Garcia-Serrano et al. 2005), and
habitat requirements (Garcia-Serrano et al. 2004). However, to my knowledge, no
published study has thus far explicitly addressed the occurrence of indirect effects
between an invasive and native Senecio species.
In Australia, the genus Senecio is represented by 87 native, and ten alien species
(Thompson 2006). Senecio madagascariensis Poiret (fireweed) is a widespread
weed in Australia, which invades arable land and grassland (Sindel et al. 1996). A
herbaceous annual or short-lived perennial, native to South Africa, S.
madagascariensis was first recorded in Australia in the Hunter Valley, New South
Wales (NSW) in 1918 (Radford et al. 1995a). In the last thirty years it has
increased its range to include all regions of coastal NSW, as well as southern
Queensland, and is still spreading (Radford and Cousens 2000). Senecio
madagascariensis also occurs as a weed in Hawaii (Le Roux et al. 2006) and parts
of South America (Fernández and Verona 1984). Research concerning S.
madagascariensis has focussed largely on basic biology and life history
characteristics (Fernández and Verona 1984; Sindel and Michael 1992, 1996;
Sindel et al. 1998), as well as control using herbicides (Anderson and Panetta
1995) and biological control agents (Marohasy 1989; Radford 1997).
Senecio madagascariensis is closely related to the native Senecio pinnatifolius A.
Rich, an herbaceous perennial (Ali 1966) which occurs in a similar range to S.
madagascariensis in south-eastern Australia. The native S. pinnatifolius tends to
form small scattered populations, in contrast with the large continuous populations
of alien S. madagascariensis (Radford 1997). Senecio pinnatifolius ssp.
lanceolatus, the focal subspecies in this thesis, inhabits a narrow border along the
edges of rainforest and wet sclerophyll forest in southeast Queensland. It is often
found within several metres of, and sometimes partially mixed with, S.

24
madagascariensis populations (Radford 1997). Senecio pinnatifolius spp.
lanceolatus (henceforth referred to simply as ‘S. pinnatifolius’) and S.
madagascariensis have coinciding flowering periods, S. madagascariensis
flowering from March to December in south eastern Australia and S. pinnatifolius
flowering between January and June (Radford 1997). The two species are
morphologically similar: both produce similar-sized yellow capitula which occur in
clusters on the plant, although mature S. pinnatifolius are often larger than S.
madagascariensis. Previous studies suggest that both species are self-
incompatible and are likely to rely on insects as pollinators (Ali 1966, Lawrence
1985). They are also known to share a number of insect herbivores (Holtkamp
and Hosking 1993), but neither plant-herbivore, nor plant-pollinator interactions in
these species have been studied in detail.
The two Senecio species and their associated invertebrate fauna provide an ideal
model system in which to investigate indirect interactions between an invasive and
a native species. Combined factors including the existence of sympatric
populations of the two species, their close relatedness, their overlapping flowering
periods, and the fact that they are likely to share insect pollinators and herbivores,
all create the potential for the occurrence of a range of indirect effects mediated by
their associated insect fauna. As illustrated by the examples provided in Chapter
Two, these interactions can be highly variable and difficult to predict, and as such
their outcomes might include positive, negative or neutral effects for either the
native or alien species. These indirect effects are the focus of the studies
contained in this thesis.
1.2.2 Specific aims of the study
The specific aims of this study are:

25
1. To synthesise the current published information on indirect interactions
between invasive and native species in a published literature review.
2. To determine the degree to which S. pinnatifolius and S. madagascariensis
share insect herbivores and pollinators and to identify the insect species
that are most likely to act as mediators of indirect interactions between the
two plant species.
3. To investigate herbivore preference and apparent competition between the
two Senecio species, focussing on one of the most important shared
folivores, Nyctemera amica (White) (Lepidoptera: Arctiidae).
4. To determine whether either plant species either facilitates or competes for
pollinator visits to the other in mixed populations, and if so, to determine
whether altered visitation rates have an impact on seed set in either
species.
5. To determine the degree of hybridisation occurring between the two species
in sympatric populations, as a result of transfer of pollen between species.
1.3 Thesis outline and presentation
1.3.1 Thesis presentation
The structure of this thesis follows QUT rules for a PhD by publication, which
allows thesis examination to be based on the presentation of a body of related
published or submitted works, linked together with abbreviated introduction and
discussion chapters. Rules can be found at www.rsc.qut.edu.au. Only minor
formatting changes have been made to the published or submitted works
comprising each chapter for the sake of consistency. These include:
standardisation of numbering of headings, tables and figures, standardisation of
citation style, incorporation of figures and tables into text, and compilation of all
cited works into a single reference list at the end of the thesis.

26
1.3.2 Account of research progress linking the research papers
The occurrence of indirect effects, mediated by insects, is, by definition, dependent
on the two Senecio species sharing insect herbivores and pollinators. The degree
of sharing of these faunal groups between the species has not been previously
quantified. Following the literature review (Chapter Two), the third chapter
addresses this issue and identifies the insect species that are most likely to act as
mediators of indirect interactions between S. pinnatifolius and S.
madagascariensis. Chapter Four focuses on N. amica, one of the more important
shared folivores identified in Chapter Three, and investigates herbivore preference
and apparent competition between the two Senecio species. In addition to
identifying shared herbivores, Chapter Three also identified a number of common,
shared floral visitors. Competition for pollinators between the alien and native
Senecio potentially could have a negative impact on seed set in either species;
alternatively the presence of one species might facilitate pollinator visits to, and
seed set in, the other. This subject is investigated in Chapter Five, whilst Chapter
Six examines the individual and population-level consequences of hybridisation
occurring between the two species in sympatric populations, as a result of transfer
of pollen between species. The concluding discussion (Chapter Seven) addresses
the outcomes of the current project in the context of invasion biology theory and
recommends directions for future research.

Chapter 2 - Biotic indirect effects: a neglected
concept in invasion biology
The following chapter was recently published as:
White, E., Wilson, J.C. and Clarke, A.R. (2006) Biotic indirect
effects: A neglected concept in invasion biology. Diversity and
Distributions 12: 443-455.
The roles of co-authors are as follows:
E.M. White: Responsible for conceptual basis of the chapter, conducted the
literature searches and wrote the paper.
J.C. Wilson: Project supervisor, made comments on drafts.
A.R. Clarke: Project supervisor, assisted with writing later drafts of the manuscript.


29
Chapter 2 – Biotic indirect effects: A neglected concept in invasion biology
2.1 Abstract
Indirect effects involve more than two species and are defined as how one species
alters the effect that another species has on a third. These complex interactions
are often overlooked in studies of interactions between exotic and native species
and their role in influencing biological invasions has been rarely considered. Based
on a comprehensive review of the invasion biology literature, we examine the
evidence for the occurrence of four of the most commonly documented indirect
effects (apparent competition, indirect mutualism/commensalism, exploitative
competition and trophic cascades) in the invasion process. Studies investigating
indirect effects in the context of invasion biology were found to be rare, but there
are sufficient examples to indicate that this kind of interaction is likely to be more
common than is currently recognised. Based on the known role of indirect effects
in structuring ecological communities, it is highly likely that indirect effects may
influence the course of a biological invasion. Whether indirect interactions are
mediated by an exotic or a native species, and whether they occur between
ecologically similar or dissimilar exotic and native species, depends in part on the
type of interaction considered and no predictable patterns were detected in the
literature. Further research is required in order to determine if any predictable
patterns do exist and whether knowledge of such may lead to a better
understanding of the potential impacts of an invasive species.
2.2 Introduction
Colonisation of new areas by plants and animals is a naturally occurring process.
However, this process has been accelerated by anthropogenic activities over the
last century with increasing rates of invasion of ecosystems by new species (i.e.
aliens, sensu Pysek et al. 2004). In many cases such invasions result in alterations

30
to the biodiversity and functioning of ecological systems (Mooney and Cleland
2001).
Many studies have investigated the impacts of alien species on native biota but the
mechanisms by which the impacts occur, although frequently speculated upon,
often remain unconfirmed by rigorous testing. Levine et al. (2003) reviewed 150
papers examining the impacts of alien plants. Approximately half of the studies
reviewed investigated effects on community structure (species diversity and
composition), while the remainder examined effects on ecosystem processes
(nutrient cycling, hydrology etc). Surprisingly, Levine et al. noted that fewer than
5% of studies confirmed the mechanism (e.g. competition, allelopathy) responsible
for the impacts, although competition was often hypothesized to be important.
Documented impacts of aliens include their direct negative effects on native
species via mechanisms such as predation (Savidge 1987; Dickman 1996; Wilson
et al. 1998; Kinnear et al. 2002; Kinzler and Maier 2003) or competition (Fogarty
and Facelli 1999; Jensen et al. 2002; Cadi and Joly 2003; Kolb and Alpert 2003;
Corbin and D’Antonio 2004; Miller and Gorchov 2004), as well as system-level
impacts, which alter abiotic processes (e.g. nutrient cycling, fire frequency,
hydrology) (Crooks 2002; Chornesky and Randall 2003). However, a native
species can also be affected if the presence of an alien species results in changes
to interactions between the native species and a third (either native or alien)
species within the invaded system. Such interactions are known as indirect effects.
In Simberloff and Von Holle’s (1999) review of 254 studies providing evidence of
facilitative interactions between alien species, only three papers discussed indirect
effects, the remainder focussed on direct interactions.
Indirect effects can be complex and difficult to predict, detect and quantify. These
interactions involve more than two species and are defined as ‘how one species

31
alters the effect that another species has on a third’ (Strauss 1991). The term
‘indirect effects’ has been used to refer to a variety of interactions described by a
range of names including apparent competition, trophic cascades, indirect (or
apparent) mutualism / facilitation, exploitative competition and interaction
modifications (Strauss 1991; Wootton 1994). Interactions arising through changes
in an abiotic resource (‘ecosystem engineering’) are also often described as
indirect effects (Strauss 1991; Lenz et al. 2003). There is no doubt that abiotic
resource modification can have major impacts on an invaded system, as
demonstrated by numerous recent studies (e.g. Fogarty and Facelli 1999; Lenz et
al. 2003; Standish et al. 2001; Wolf et al. 2004; Yelenik et al. 2004) (see Crooks
(2002) for a comprehensive review of this topic). The current review, however,
focuses on purely biotic indirect effects which, as defined above, require the
presence of a third species through which they are mediated. The third species,
which may be either native or alien, is henceforth referred to as the mediator or
mediating species.
Based on a comprehensive review of the invasion biology literature, this paper
examines the evidence for the occurrence of indirect effects between alien and
native species and their potential impacts on the invasion process. Literature
searches for publications released between the years 1994 and 2005 were
conducted using a wide range of search-terms including combinations of ‘indirect
effects’, ‘indirect interactions’, ‘higher-order interactions’, ‘multi-species
interactions’, tri-trophic interactions, ‘biotic interactions’ ‘exotic’, ‘alien’, ‘invasive
species’, ‘invader’, ‘trophic cascades’, ‘apparent competition’, ‘herbivory’,
‘competition’, ‘mutualism’, ‘indirect mutualism’, ‘indirect commensalism’,
‘facilitation’ and ‘pollination’. Searches were performed using databases ‘Biological
Abstracts’, ‘Ovid’, ‘Current Contents Connect’ and ‘Web of Science’, then cross-

32
searches were conducted to locate relevant papers published earlier than this
timeframe.
The core of the paper focuses on the four most commonly documented types of
indirect effect: apparent competition, indirect mutualism / facilitation, exploitative
competition (in situations in which the limiting resource (the mediator) is another
species, such as a prey species, rather than an abiotic resource) and trophic
cascades. Each of these interaction types is developed to present theoretical
examples of how such interactions might modify the invasion process and are
supported, where available, by one or two illustrative examples drawn from
empirical studies involving alien species. A comprehensive list (Table 2.1) of
documented biotic indirect effects from the invasion biology literature further
supports this section of the review. The discussion focuses on major patterns
concerning indirect effects and invasions which emerge from a synthesis of the
literature and stresses the importance of directly testing for indirect effects in future
invasion biology studies.
33
AC ECIM(F) TC
Host-parasitoid
Plant (animal)-mutualist
Plant-herbivore
Plant-herbivore & predator-prey
Predator-prey
0
1
2
3
4
5
6
7
Nu
mb
er
of stu
die
s
Indirect effect
System
Alien fauna
Alien flora
Figure 2.1: Number of published studies focussing on indirect effects involving alien flora and fauna
in different system-types. Indirect effects: AC, apparent competition; EC, Exploitative competition;
IM(F), Indirect mutualism / facilitation; TC, Trophic cascades. ‘Plant-herbivore and predator-prey’
have been combined as a single category for when the two are inseparable in the case of trophic
cascades.
2.3 Mechanisms for indirect effects in biological invasions
2.3.1 Apparent competition
Apparent competition has most frequently been documented between alien and
native fauna in predator-prey and host-parasitoid systems and between alien and
native plants in plant-herbivore systems (Figure 2.1). Apparent competition occurs
when a predator, herbivore, parasite or pathogen (henceforth referred to
generically as ‘consumer’), becomes more numerous or more efficient at attacking
one species (the ‘prey’) in the presence of another (Holt 1977). This interaction
can be considered to be indirect because the impacts would not occur without the

34
presence of the consumer mediating the interaction, which may itself be either
alien or native. Apparent competition could occur between an alien and native
species in a number of ways:
1) Co-occurring alien and native species provide different types of resource for the
consumer mediating the interaction.
a) An alien species provides a consumer with a non-food resource, such as
shelter, allowing the consumer population to increase or spread in
distribution subsequently having a negative impact on native prey species
(e.g. Sessions and Kelly 2002, Table 2.1).
b) The alien species provides a food-limited consumer with a food resource
which is required at a particular stage in the consumer’s lifecycle, allowing
the population to increase or spread in distribution and have a negative
impact on a native prey species which is utilised at a different stage in the
consumer’s lifecycle. For example, adult moths might be attracted to the
nectar of an alien plant species, but oviposit on a neighbouring native
species, which thus experiences increased larval herbivory in the presence
of the alien plant. This type of interaction has been demonstrated to occur
between native insect and plant species (Thomas 1986; Karban 1997). It
has not, to our knowledge, been recorded between an alien and native
plant species, but we see no reason why it could not occur, given the ability
of many invertebrates to use alien plant species as hosts (e.g. Louda et al.
2005; Russell and Louda 2005).
2) Co-occurring alien and native species both provide a food resource to a food-
limited consumer. The increased resource availability (provided by the alien),
allows the consumer mediating the interaction to increase in abundance and

35
consequently have a greater negative impact on sympatric native prey. This
could occur in the following situations:
a) Differential attack rates: The native species experiences a proportionally
greater attack rate than the alien (e.g. Settle and Wilson 1990; Benson et
al. 2003). This is likely when the mediating consumer species is native and
exhibits a preference for prey with which it evolved (Settle and Wilson 1990;
Meng and Orsi 1991; Schierenbeck et al. 1994; Brown et al. 1995; MacNeil
et al. 2003; Gamboa et al. 2004). Differential attack rates might also be
observed if one prey species is more easily captured than the other
(Courchamp et al. 2000) or via prey-switching. An example of the latter
may arise if the alien prey population (the presence of which has permitted
the consumer population to increase) experiences a dramatic decline in
abundance, resulting in the consumer switching to an alternate native prey
species (e.g. Norbury 2001, Table 2.1).
b) Different levels of tolerance: Attack rates are similar between the two prey
species, but the addition of a similar amount of new mortality has a greater
impact on one species than the other. This could occur if the native and
alien species differ in life history characteristics (Taylor 1979; Roane et al.
1986; van Riper 1986; Smith and Quin 1996; Courchamp et al. 2000;
Roemer et al. 2002; Holt and Barfield 2003; Grosholz 2005). In fact it is
common that native species have relatively low fecundity or growth rates, or
higher mortality rates compared to related or ecologically similar co-
occurring aliens (Noble 1989; Byers 2000a; Gamboa et al. 2004; Roemer et
al. 2002; Siemann and Rogers 2003) and thus may suffer differentially due
to increased predation. As an example, feral pigs in the California Channel
Islands have had an indirect negative impact on the native island fox by
acting as an abundant food resource for golden eagles, enabling them to

36
colonise the islands (Roemer et al. 2002). The eagles also preyed on the
native island fox, driving it towards extinction. The authors concluded that
the differential impact of predation between the pigs and the fox was due to
life history differences, the feral pig having higher fecundity and a larger
body size than the native fox.
There is sufficient evidence to indicate that apparent competition between alien
and native species has the potential to have significant negative consequences for
native species (see examples given above and Table 2.1). Because the outcome
of apparent competition can be the same as that of competition, it is possible that
the impact might be attributed to competition if alternative potential mechanisms,
including the involvement of a mediating species, behind the impact aren’t
investigated. It is vital to know whether apparent competition (as opposed to
resource competition) is occurring, as management strategies will have to take into
account the role of the consumer. Management is likely to vary depending on
characteristics of the consumer, for instance the issue may be complicated if the
consumer is a native species of conservation importance.
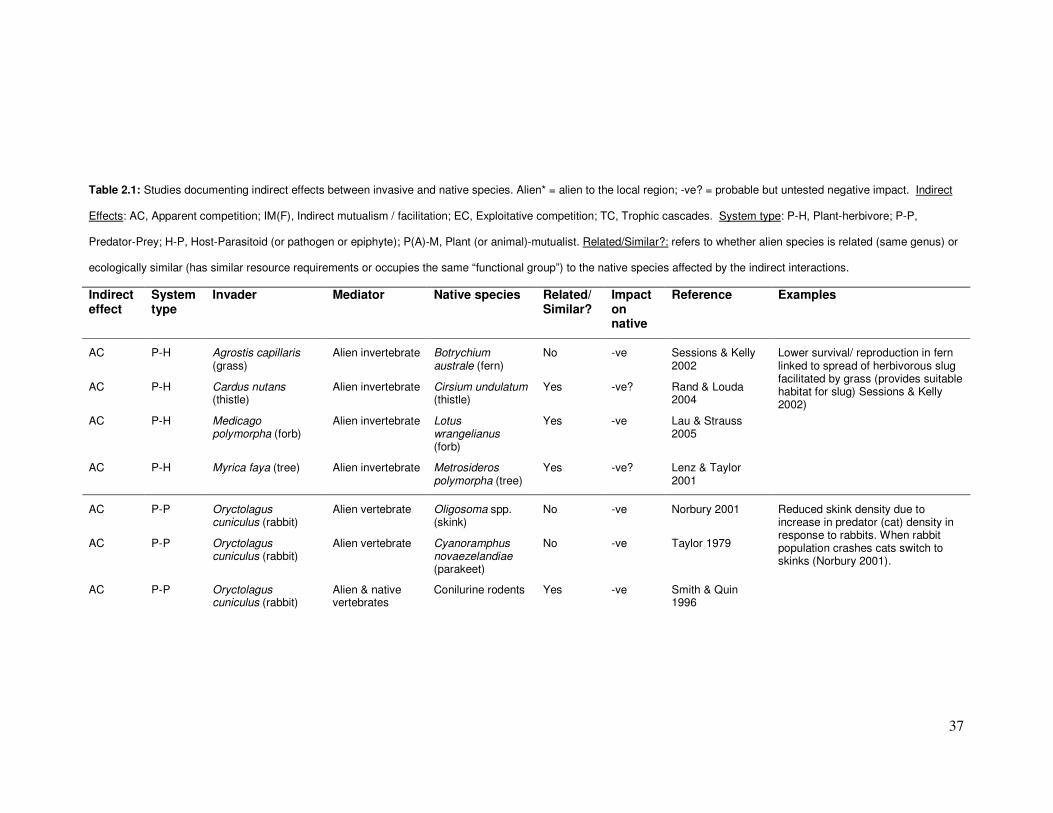
37
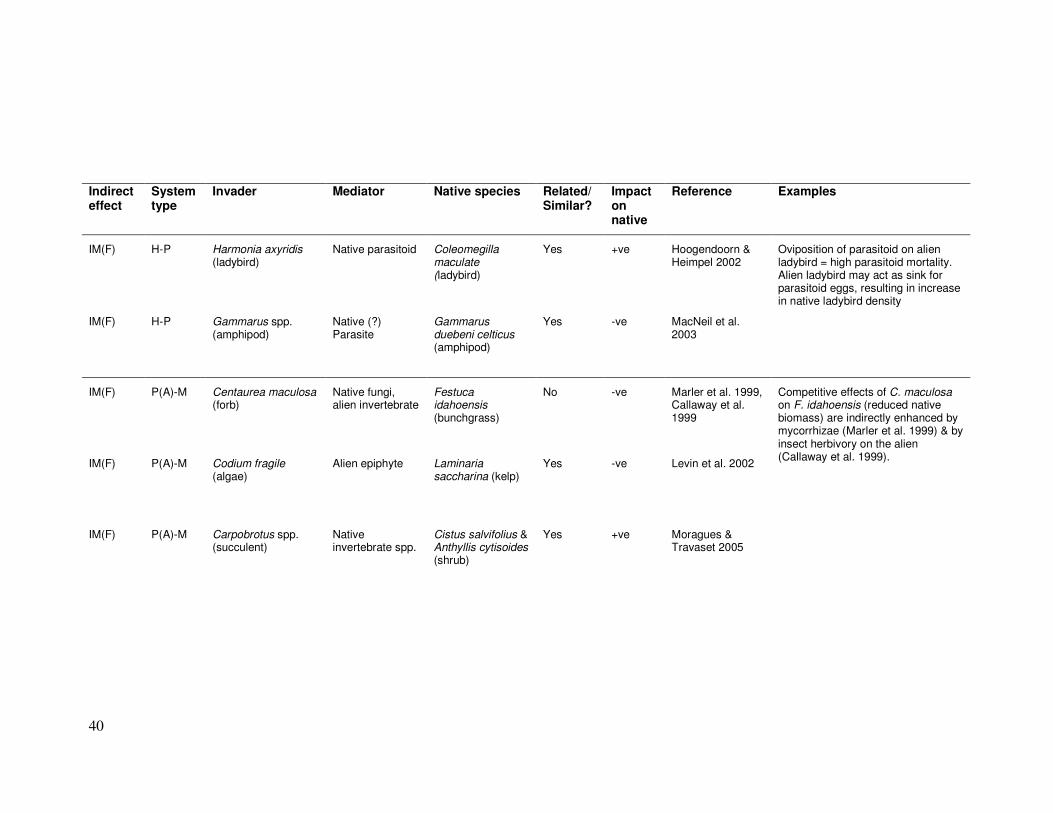
Table 2.1: Studies documenting indirect effects between invasive and native species. Alien* = alien to the local region; -ve? = probable but untested negative impact. Indirect
Effects: AC, Apparent competition; IM(F), Indirect mutualism / facilitation; EC, Exploitative competition; TC, Trophic cascades. System type: P-H, Plant-herbivore; P-P,
Predator-Prey; H-P, Host-Parasitoid (or pathogen or epiphyte); P(A)-M, Plant (or animal)-mutualist. Related/Similar?: refers to whether alien species is related (same genus) or
ecologically similar (has similar resource requirements or occupies the same “functional group”) to the native species affected by the indirect interactions.
Indirect effect
System type
Invader Mediator Native species Related/ Similar?
Impact on native
Reference Examples
AC P-H Agrostis capillaris (grass)
Alien invertebrate Botrychium australe (fern)
No -ve Sessions & Kelly 2002
AC P-H Cardus nutans (thistle)
Alien invertebrate Cirsium undulatum (thistle)
Yes -ve? Rand & Louda 2004
AC P-H Medicago polymorpha (forb)
Alien invertebrate Lotus wrangelianus (forb)
Yes -ve Lau & Strauss 2005
AC P-H Myrica faya (tree) Alien invertebrate Metrosideros polymorpha (tree)
Yes -ve? Lenz & Taylor 2001
Lower survival/ reproduction in fern linked to spread of herbivorous slug facilitated by grass (provides suitable habitat for slug) Sessions & Kelly 2002)
AC P-P Oryctolagus cuniculus (rabbit)
Alien vertebrate Oligosoma spp. (skink)
No -ve Norbury 2001
AC P-P Oryctolagus cuniculus (rabbit)
Alien vertebrate Cyanoramphus novaezelandiae (parakeet)
No -ve Taylor 1979
AC P-P Oryctolagus cuniculus (rabbit)
Alien & native vertebrates
Conilurine rodents Yes -ve Smith & Quin 1996
Reduced skink density due to increase in predator (cat) density in response to rabbits. When rabbit population crashes cats switch to skinks (Norbury 2001).
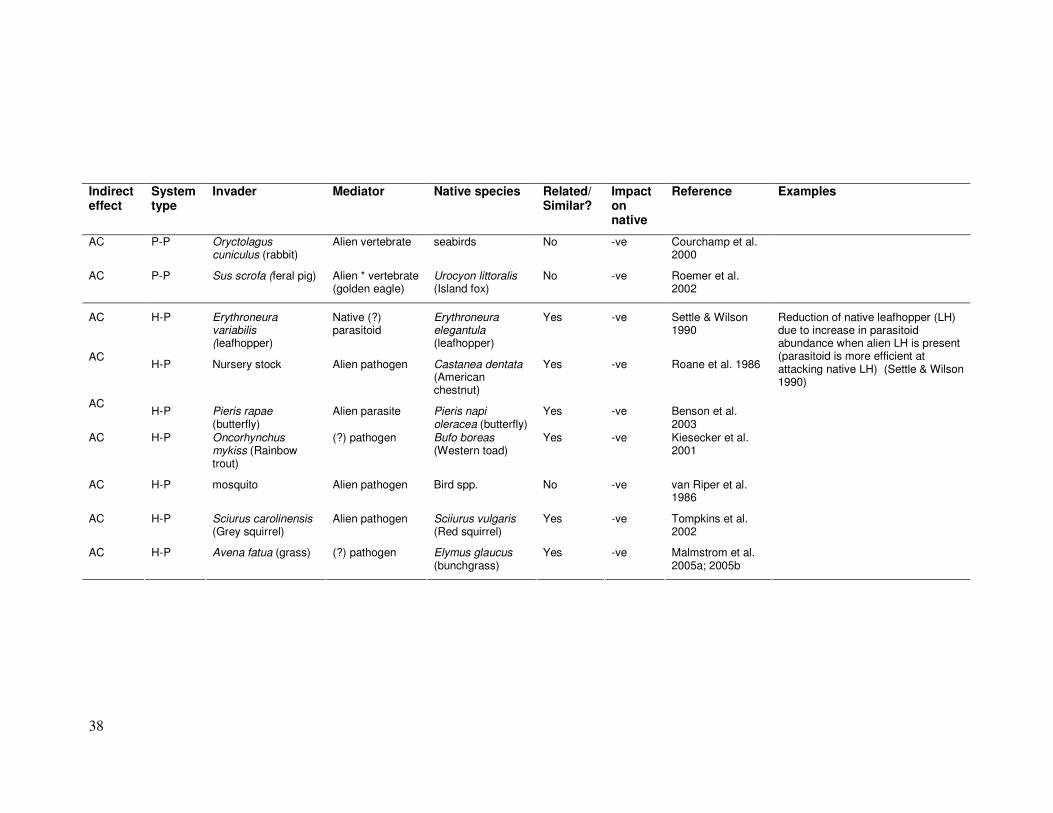
38
Indirect effect
System type
Invader Mediator Native species Related/ Similar?
Impact on native
Reference Examples
AC P-P Oryctolagus cuniculus (rabbit)
Alien vertebrate seabirds No -ve Courchamp et al. 2000
AC P-P Sus scrofa (feral pig) Alien * vertebrate (golden eagle)
Urocyon littoralis (Island fox)
No -ve Roemer et al. 2002
AC H-P Erythroneura variabilis (leafhopper)
Native (?) parasitoid
Erythroneura elegantula (leafhopper)
Yes -ve Settle & Wilson 1990
AC H-P Nursery stock Alien pathogen Castanea dentata
(American chestnut)
Yes -ve Roane et al. 1986
AC H-P Pieris rapae
(butterfly) Alien parasite Pieris napi
oleracea (butterfly) Yes -ve Benson et al.
2003 AC H-P Oncorhynchus
mykiss (Rainbow trout)
(?) pathogen Bufo boreas (Western toad)
Yes -ve Kiesecker et al. 2001
AC H-P mosquito Alien pathogen Bird spp. No -ve van Riper et al. 1986
AC H-P Sciurus carolinensis (Grey squirrel)
Alien pathogen Sciiurus vulgaris (Red squirrel)
Yes -ve Tompkins et al. 2002
AC H-P Avena fatua (grass) (?) pathogen Elymus glaucus (bunchgrass)
Yes -ve Malmstrom et al. 2005a; 2005b
Reduction of native leafhopper (LH) due to increase in parasitoid abundance when alien LH is present (parasitoid is more efficient at attacking native LH) (Settle & Wilson 1990)
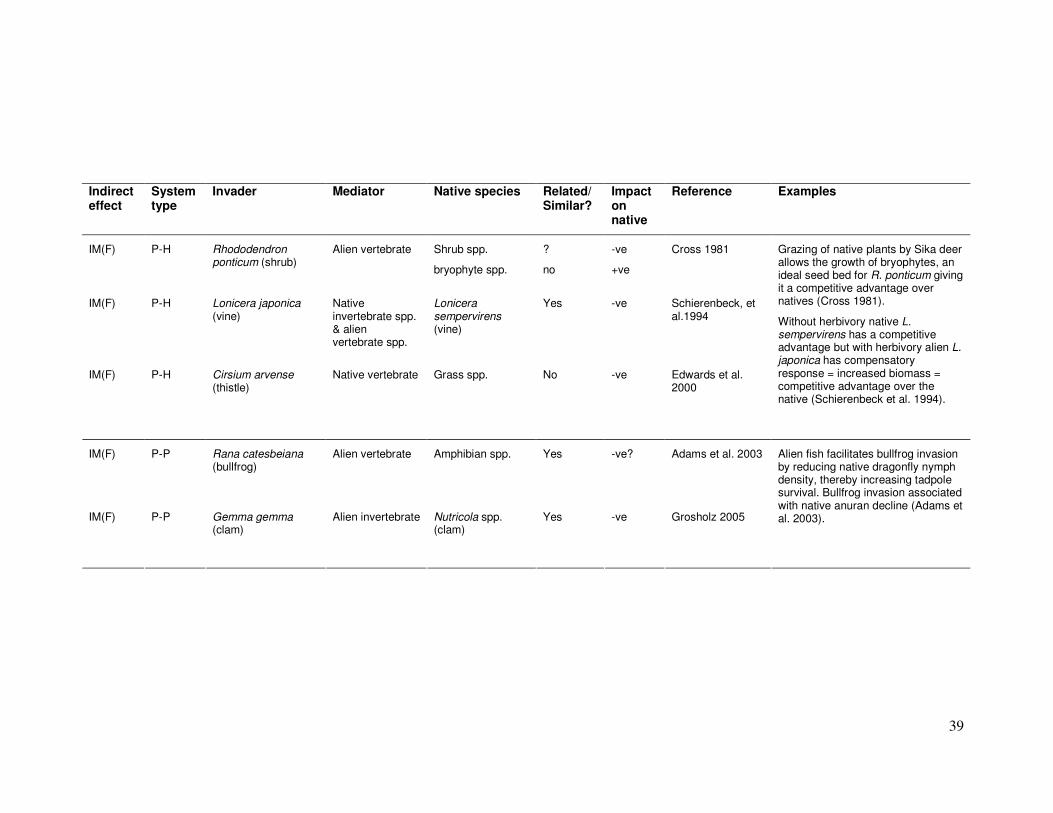
39
Indirect effect
System type
Invader Mediator Native species Related/ Similar?
Impact on native
Reference Examples
IM(F) P-H Rhododendron ponticum (shrub)
Alien vertebrate Shrub spp.
bryophyte spp.
?
no
-ve
+ve
Cross 1981
IM(F) P-H Lonicera japonica (vine)
Native invertebrate spp. & alien vertebrate spp.
Lonicera sempervirens (vine)
Yes -ve Schierenbeck, et al.1994
IM(F) P-H Cirsium arvense (thistle)
Native vertebrate Grass spp. No -ve Edwards et al. 2000
Grazing of native plants by Sika deer allows the growth of bryophytes, an ideal seed bed for R. ponticum giving it a competitive advantage over natives (Cross 1981).
Without herbivory native L. sempervirens has a competitive advantage but with herbivory alien L. japonica has compensatory response = increased biomass = competitive advantage over the native (Schierenbeck et al. 1994).
IM(F) P-P Rana catesbeiana (bullfrog)
Alien vertebrate Amphibian spp. Yes -ve? Adams et al. 2003
IM(F) P-P Gemma gemma (clam)
Alien invertebrate Nutricola spp. (clam)
Yes -ve Grosholz 2005
Alien fish facilitates bullfrog invasion by reducing native dragonfly nymph density, thereby increasing tadpole survival. Bullfrog invasion associated with native anuran decline (Adams et al. 2003).
40
Indirect effect
System type
Invader Mediator Native species Related/ Similar?
Impact on native
Reference Examples
IM(F) H-P Harmonia axyridis (ladybird)
Native parasitoid Coleomegilla maculate (ladybird)
Yes +ve Hoogendoorn & Heimpel 2002
Oviposition of parasitoid on alien ladybird = high parasitoid mortality. Alien ladybird may act as sink for parasitoid eggs, resulting in increase in native ladybird density
IM(F) H-P Gammarus spp. (amphipod)
Native (?) Parasite
Gammarus duebeni celticus (amphipod)
Yes -ve MacNeil et al. 2003
IM(F) P(A)-M Centaurea maculosa (forb)
Native fungi, alien invertebrate
Festuca idahoensis (bunchgrass)
No -ve Marler et al. 1999, Callaway et al. 1999
IM(F) P(A)-M Codium fragile (algae)
Alien epiphyte Laminaria saccharina (kelp)
Yes -ve Levin et al. 2002
IM(F) P(A)-M Carpobrotus spp. (succulent)
Native invertebrate spp.
Cistus salvifolius & Anthyllis cytisoides (shrub)
Yes +ve Moragues & Travaset 2005
Competitive effects of C. maculosa on F. idahoensis (reduced native biomass) are indirectly enhanced by mycorrhizae (Marler et al. 1999) & by insect herbivory on the alien (Callaway et al. 1999).
41
Indirect effect
System type
Invader Mediator Native species Related/ Similar?
Impact on native
Reference Examples
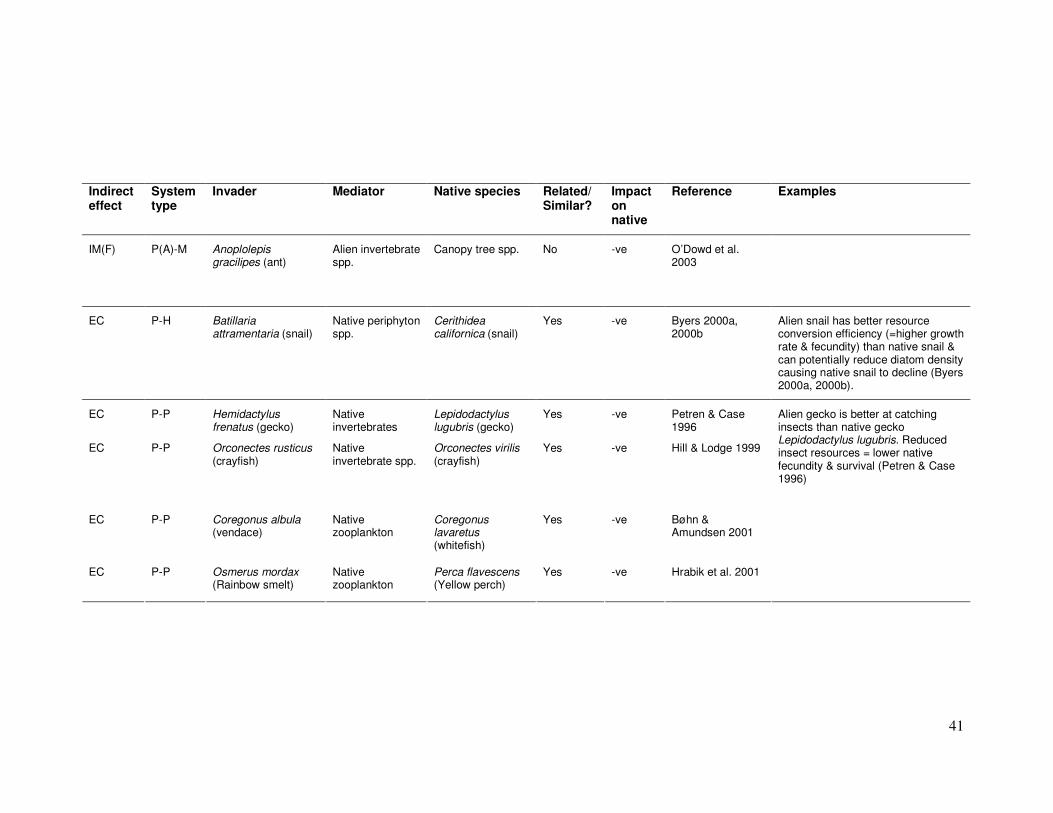
IM(F) P(A)-M Anoplolepis gracilipes (ant)
Alien invertebrate spp.
Canopy tree spp. No -ve O’Dowd et al. 2003
EC P-H Batillaria attramentaria (snail)
Native periphyton spp.
Cerithidea californica (snail)
Yes -ve Byers 2000a, 2000b
Alien snail has better resource conversion efficiency (=higher growth rate & fecundity) than native snail & can potentially reduce diatom density causing native snail to decline (Byers 2000a, 2000b).
EC P-P Hemidactylus frenatus (gecko)
Native invertebrates
Lepidodactylus lugubris (gecko)
Yes -ve Petren & Case 1996
EC P-P Orconectes rusticus (crayfish)
Native invertebrate spp.
Orconectes virilis (crayfish)
Yes -ve Hill & Lodge 1999
EC P-P Coregonus albula (vendace)
Native zooplankton
Coregonus lavaretus (whitefish)
Yes -ve Bøhn & Amundsen 2001
EC P-P Osmerus mordax (Rainbow smelt)
Native zooplankton
Perca flavescens (Yellow perch)
Yes -ve Hrabik et al. 2001
Alien gecko is better at catching insects than native gecko Lepidodactylus lugubris. Reduced insect resources = lower native fecundity & survival (Petren & Case 1996)
42
Indirect effect
System type
Invader Mediator Native species Related/ Similar?
Impact on native
Reference Examples
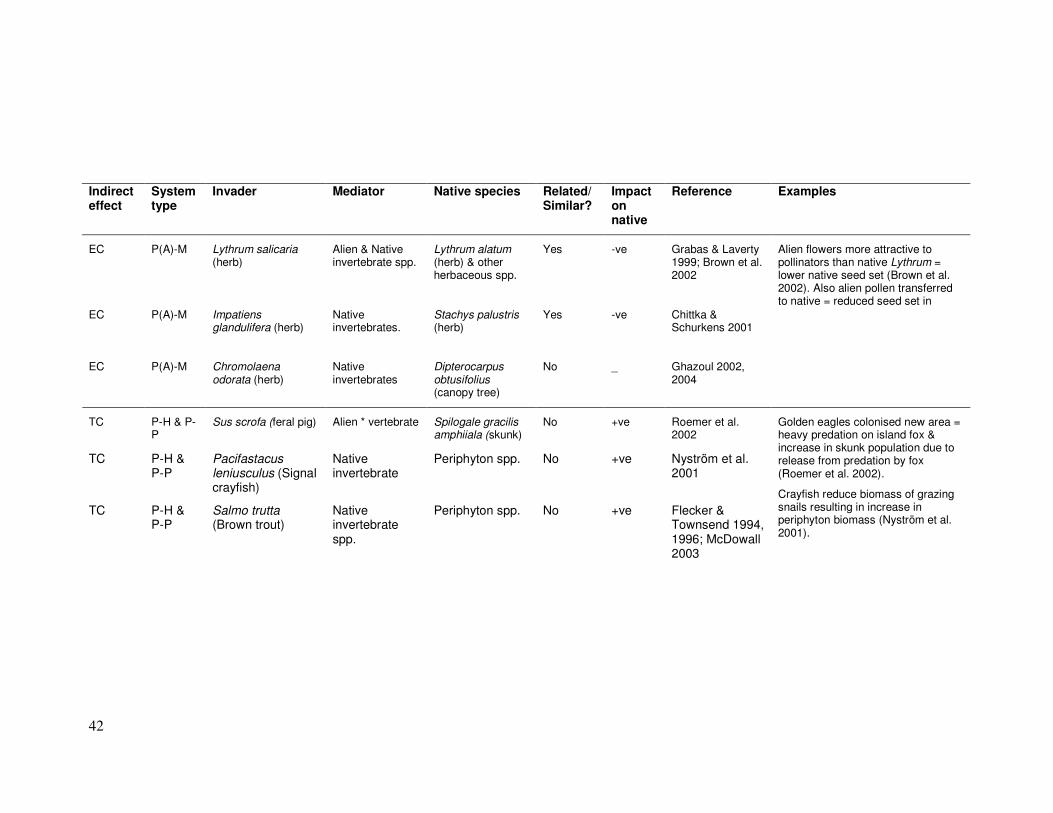
EC P(A)-M Lythrum salicaria (herb)
Alien & Native invertebrate spp.
Lythrum alatum (herb) & other herbaceous spp.
Yes -ve Grabas & Laverty 1999; Brown et al. 2002
EC P(A)-M Impatiens glandulifera (herb)
Native invertebrates.
Stachys palustris (herb)
Yes -ve Chittka & Schurkens 2001
EC P(A)-M Chromolaena odorata (herb)
Native invertebrates
Dipterocarpus obtusifolius (canopy tree)
No _ Ghazoul 2002, 2004
Alien flowers more attractive to pollinators than native Lythrum = lower native seed set (Brown et al. 2002). Also alien pollen transferred to native = reduced seed set in
TC P-H & P-P
Sus scrofa (feral pig) Alien * vertebrate Spilogale gracilis amphiiala (skunk)
No +ve Roemer et al. 2002
TC P-H & P-P
Pacifastacus leniusculus (Signal crayfish)
Native invertebrate
Periphyton spp. No +ve Nyström et al. 2001
TC P-H & P-P
Salmo trutta (Brown trout)
Native invertebrate spp.
Periphyton spp. No +ve Flecker & Townsend 1994, 1996; McDowall 2003
Golden eagles colonised new area = heavy predation on island fox & increase in skunk population due to release from predation by fox (Roemer et al. 2002).
Crayfish reduce biomass of grazing snails resulting in increase in periphyton biomass (Nyström et al. 2001).
43
Indirect effect
System type
Invader Mediator Native species Related/ Similar?
Impact on native
Reference Examples
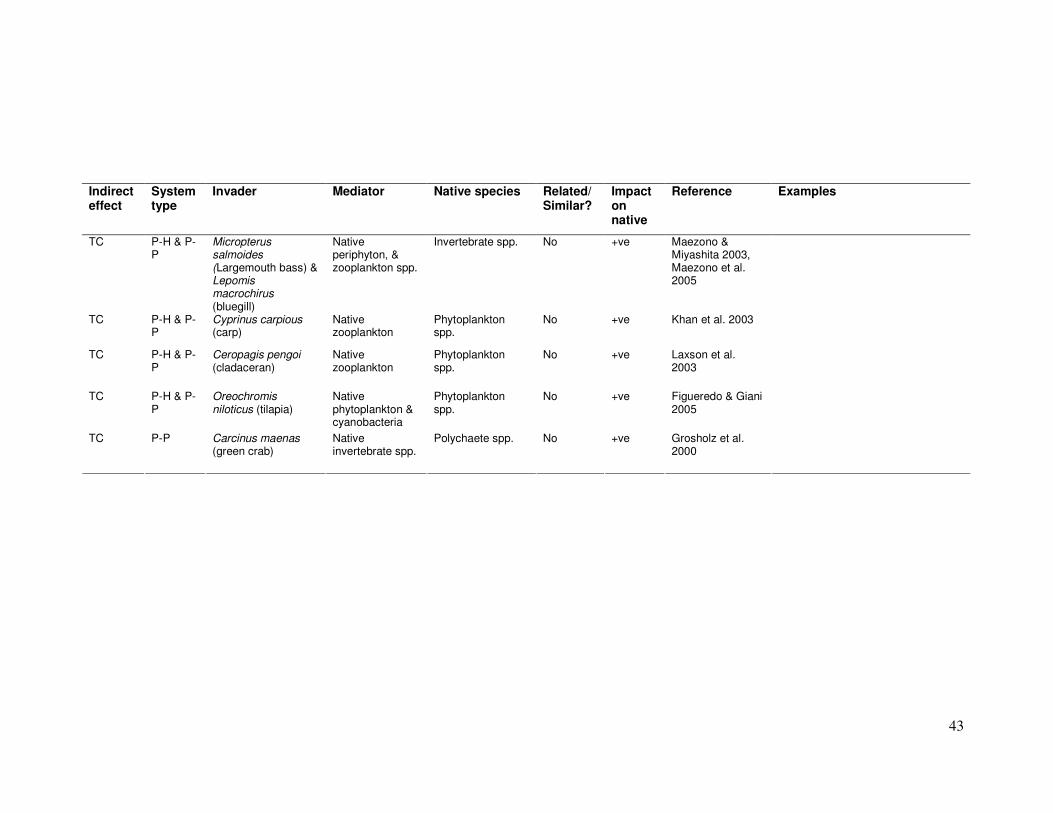
TC P-H & P-P
Micropterus salmoides (Largemouth bass) & Lepomis macrochirus (bluegill)
Native periphyton, & zooplankton spp.
Invertebrate spp. No +ve Maezono & Miyashita 2003, Maezono et al. 2005
TC P-H & P-P
Cyprinus carpious (carp)
Native zooplankton
Phytoplankton spp.
No +ve Khan et al. 2003
TC P-H & P-P
Ceropagis pengoi (cladaceran)
Native zooplankton
Phytoplankton spp.
No +ve Laxson et al. 2003
TC P-H & P-P
Oreochromis niloticus (tilapia)
Native phytoplankton & cyanobacteria
Phytoplankton spp.
No +ve Figueredo & Giani 2005
TC P-P Carcinus maenas (green crab)
Native invertebrate spp.
Polychaete spp. No +ve Grosholz et al. 2000

44
2.3.2 Indirect mutualism / facilitation
Indirect mutualism / facilitation has been recorded in a range of system types, and
can occur between either alien flora or fauna and other resident native or alien
species (Figure 2.1). Whilst apparent competition results in a negative impact on
the focal species, the contrary effect can also occur, with the presence of one
species having a positive indirect effect on another species, usually as a
consequence of direct negative impacts on a third mediator species. Indirect
mutualism (or facilitation) - the positive indirect effect of one species on another
(Schoener 1993) – has been hypothesised to be extremely important in structuring
some communities (Levine 1980; Bascompte et al. 2003). Two of the main ways in
which it can occur include:
1) In consumer-prey interactions via mechanisms such as prey switching,
consumer satiation (Abrams and Masuda 1996), or by a consumer targeting
a more abundant prey species, ignoring the less abundant prey (Abrams
1987). Indirect mutualism or facilitation could benefit either a native or alien
species involved in the interaction; for instance in Ireland invasion by the
alien sika deer indirectly benefits the alien shrub Rhododendron ponticum.
The mediating species in this interaction are native shrubs and bryophytes.
Disturbance to native vegetation caused by preferential grazing by deer
allows the establishment of a bryophyte carpet, an ideal seed bed for
Rhododendron (Cross 1981). Thus interactions between the deer and the
native vegetation indirectly benefit alien Rhododendron, allowing it to
outcompete native vegetation. This kind of indirect interaction is probably
quite common in invaded systems.
Although positive indirect effects of an alien on a native species are less
frequently reported, this does not mean they do not occur. Hoogendoorn

45
and Heimpel (2002) demonstrated that the presence of an alien ladybird
benefits a native ladybird in a host-parasitoid system, by acting as a sink for
parasitoid eggs, resulting in increase in native ladybird density.
2) Indirect mutualism / facilitation can also occur between competing species
when one species benefits another by suppressing a third shared
competitor (Miller 1994; Stone and Roberts 1991). This has been
demonstrated between co-occurring native species (Levine 1999; Callaway
and Pennings 2000), and there is the potential for such a scenario to
involve alien species though to our knowledge this not yet been
documented.
The most frequently documented scenario of indirect mutualism / facilitation
between alien and native species involves one alien species indirectly benefiting
another by interfering in some way with native species, altering the existing
competitive dominance hierarchies. In order to implement appropriate
management strategies it is important to understand the role of all species involved
in such interactions – for example in Ireland, attempts to control Rhododendron
would greatly benefit by simultaneous control of the deer population which
facilitates Rhododendron invasion.
2.3.3 Exploitative competition
Competitive interactions are often not acknowledged as indirect effects (Strauss
1991; Wootton 2002). However, because exploitative competition can involve the
effect of one species on another mediated through changes in abundance of a third
species (as in indirect mutualism, the mediator is usually a shared prey species) in
many cases it is, by definition, an indirect effect. Numerous studies have focussed
on competition for abiotic resources between alien and native plant species (e.g.

46
Fogarty and Facelli 1999; Blicker et al. 2003; Kolb and Alpert 2003; Vila et al.
2003; Corbin and D’Antonio 2004; Fehmi et al. 2004; Miller and Gorchov 2004) and
between alien and native fauna (e.g. Bryce et al. 2002; Jensen et al. 2002;
Landwer and Ferguson 2002; Wauters et al. 2002; Cadi and Joly 2003).
Surprisingly though, whilst exploitative competition is often hypothesised to be the
mechanism behind the impacts of alien species (Eguchi and Amano 1999; Kido et
al. 1999; Talman and Keough 2001; Lorenzoni et al. 2002; Kane et al. 2003; Mistri
et al. 2004; Raikow 2004), relatively few studies have quantified the impact of an
alien on a native species mediated by changes in abundance of a shared prey
species. Perhaps this is due to the difficulties associated with manipulation and
quantification of population dynamics of three interacting species, which are likely
to make this type of experiment more complex than an investigation of competition
for an abiotic resource.
If an alien is more efficient than a native competitor at finding or utilising a biotic
resource, the native may experience an indirect negative impact as a consequence
of the reduced density of the shared resource. The superior ability of an alien to
exploit a resource can be a result of:
a) Better harvesting ability: For example, the alien gecko Hemidactylus frenatus is
larger and faster and therefore better at catching insects (the mediating species)
than the sympatric native gecko, Lepidodactylus lugubris. This results in a
reduction in the prey resource, which has a negative impact on native gecko
survival and fecundity (Petren and Case 1996).
b) Superior resource conversion efficiency. The alien snail, Batillaria attramentaria,
can out-compete the native snail, Cerithidea californica, because of its superior
growth response at any given level of the mediating prey species (diatom species)
(Byers 2000a). The larger body size translates to higher fecundity, thus the alien

47
eventually reaches a higher density than the native snail. Byers’ model predicts
that this will result in the diatoms being driven below the equilibrium density set by
the native snail, causing the native to decline and be replaced by Batillaria.
For exploitative competition to be considered a true biotic indirect interaction, it
must be mediated by a third species, thus situations that involve competition for an
abiotic resource cannot be considered to be an indirect interaction. It might
therefore be expected that this type of interaction will be restricted to cases of alien
fauna (consumers). However competition among alien and native plants for
mutualists - namely pollinators - has also been documented (Rathke 1983) (see
Table 2.1 and Figure 2.1). It is this kind of scenario that may be particularly
challenging to identify since interactions between plants and their associated
mutualists may not be as immediately obvious as consumer-prey interactions.
2.3.4 Trophic cascades
Trophic cascades occur with the introduction of a new consumer into a food-web
system (Figure 2.1). The term ‘trophic cascades’ describes the way in which a
species at a lower trophic level (usually a plant) is indirectly affected by the
predation of its consumers (the mediators of this interaction) by species belonging
to higher trophic levels (Strong 1992). Most documented examples of trophic
cascades come from relatively low-complexity freshwater and marine systems
(Strong 1992; Shurin et al. 2002), for example in New Zealand the alien brown
trout Salmo trutta reduces the abundance of grazing invertebrates in rivers,
resulting in an increase in algal biomass (Townsend 1996; Simon and Townsend
2003). It has been argued that trophic cascades are more important in aquatic than
terrestrial systems because the link between consumers and primary producers is
stronger than in terrestrial systems (Strong 1992; Shurin et al. 2002). Wootton

48
(1994) however suggests that relatively few examples of trophic cascades come
from terrestrial environments simply because experimental manipulations of top
consumers are more difficult in such systems. A rare example of a trophic cascade
in an invaded terrestrial system comes from the California Channel Islands.
Roemer et al. (2002) recorded colonisation of the islands by golden eagles (a
species that was alien to the area), which preyed heavily on the island fox,
resulting in an increase in the native skunk population due to release from
predation by foxes.
Trophic cascades, initiated by the introduction of a new faunal species into aquatic
systems have been relatively well-documented, but virtually nothing is known about
the importance of trophic cascades when alien animals invade terrestrial system
invasions. Considering the potential for wide-ranging impacts on multiple trophic
levels, this is an area that warrants further investigation.
2.4 Discussion
As yet we don’t have a definitive set of generalisations to enable us to predict the
full range of impacts an alien may have on a new system. The difficulties
associated with predicting invasion success and impact may arise from the fact
that researchers have often sought a single explanation (e.g. a simple set of
species traits, or the outcome of pair-wise interactions between species) when
mechanisms of invasion and impact are likely to be far more subtle and complex
than we realise (Mack 1996; Lavorel et al. 1999), involving interactions between
multiple species traits and multiple features of the new system (Schierenbeck et al.
1994; Richardson et al. 2000). Many studies have successfully identified a range
of impacts of alien species, but far fewer have ascertained the major mechanism(s)
behind the impacts (Levine et al. 2003). A more thorough understanding of the

49
mechanisms will give us better predictive ability in regards to the potential impacts
of alien invaders.
There has been much discussion regarding the importance of indirect effects in
structuring ecological systems (Holt 1977; Holt 1984; Holt and Kotler 1987; Bonsall
and Hassell 1997; Strauss 1991; Wootton 1994; Cheng and Xu 2003), with some
authors arguing that indirect effects may in some cases be the most important
factor influencing the success of a species, allowing species ‘not only to feed
efficiently and protect themselves from predators, but also to modify their
community in ways that loop back beneficially to them’ (Wilson 1986). Many
empirical studies have demonstrated the occurrence of indirect effects between
sympatric native species (e.g. Levin and Anderson 1970; Campbell 1985; Thomas
1986; Mothershead and Marquis 2000; Veech 2000; Adler et al. 2001; Morris 2002;
Webster and Almany 2002; LoGuidice 2003; Lombadero et al. 2003; Rand 2003;
Rooney and Waller 2003; Morris et al. 2004), but until recently few empirical
studies had investigated such interactions in the context of invasion biology. If
these interactions are as important in structuring communities as is speculated by
some authors (Holt 1977; Wilson 1986; Holt and Kotler 1987; Miller 1994; Bonsall
and Hassell 1997; Lortie et al. 2004), they are likely to play a vital (though as yet
little known) role in influencing invasion success as well as determining the impact
of an alien invader on a system.
Many authors (starting with Darwin (1859)) have argued that potential invaders that
lack closely related (or ecologically similar) native species at a site should be more
successful than those which are similar to natives, partly because the former are
likely to be different in their resource utilisation and will therefore more easily
escape competition with natives. In contrast, Richardson et al. (2000) argue that
species that are similar to natives have a high chance of successful invasion

50
because they can easily utilise native mutualists. This raises the further question:
Once an invasion has occurred, how will indirect interactions and their impacts vary
depending on the degree of similarity (or relatedness) between the alien species
and natives with which it is interacting?
Aliens that are ecologically similar or related to native species should quickly
develop interactions with mutualists such as pollinators, dispersers and mycorrhizal
fungi (Richardson et al. 2000). The development of these direct interactions
provides the alien not only with the opportunity to establish successfully and
interact directly with native species, but also to interact with natives via indirect
effects. Certain types of indirect effect are more likely to occur between closely
related or ecologically similar species. In seven of the eight papers reviewed here
which demonstrated the occurrence of exploitative competition (Table 2.1), the
alien and native species between which the interaction occurred were either
related or could be considered to be ecological equivalents. It seems likely that the
greater the degree of similarity between two species, the more probable it is that
they will utilise the same resources, creating the potential for exploitative
competition to occur. Indirect effects are also more likely to be observed between
similar rather than dissimilar species when occurring between two hosts, mediated
by a parasite or pathogen, due to the relatively specialised nature of this type of
interaction. Apparent competition and indirect mutualism / facilitation have been
demonstrated to occur between alien and native species, mediated by parasites or
pathogens and the majority of cases (8/9 papers reviewed here, Table 2.1) are
reported between similar or related host species.
On the other hand, invasion by a species which lacks ecological equivalents
among the existing natives might also have a large impact on natives via other
indirect mechanisms. It has been suggested that the impact of some alien species

51
might be due to the novel mechanisms of interaction they bring to a system which
lacks indigenous related or ecologically similar species (Callaway and Aschehoug
2000; Ricciardi and Atkinson 2004). This is likely to be particularly pronounced for
alien fauna; for example the introduction of a top predator to a system that
previously lacked such species can have dramatic consequences for species at all
trophic levels via trophic cascades or apparent competition, in addition to the
obvious direct effects of predation (e.g. Roemer et al. 2002). By definition,
multitrophic interactions such as trophic cascades occur between ecologically
dissimilar species (because they occupy different trophic levels) and are restricted
to predator-prey and plant-herbivore systems (Figure 2.1). This type of interaction
has been relatively well documented in aquatic environments between alien and
native species and is usually reported to be mediated by a native, rather than alien
prey species.
Other types of interaction, namely apparent competition, mediated by a shared
herbivore or predator, seem equally likely to occur between similar or dissimilar
alien and native species.
Some types of indirect interaction are more frequently reported to be mediated by
an alien, rather than native species. In thirteen of the fifteen papers reviewed here
which investigated apparent competition (Table 2.1), the interaction was mediated
by an alien predator, herbivore or parasite species. This offers support to the
invasional meltdown hypothesis (Simberloff and Von Holle 1999; Ricciardi 2001),
which suggests that facilitative interactions between alien species are a common
occurrence (predicted to result in an accelerating accumulation of introduced
species and their impacts). In most, if not all, reported cases of apparent
competition between an alien and native species, the alien has particular life
history characteristics, such as greater fecundity or lower rates of mortality, than

52
the native species, allowing it to withstand higher rates of predation and support
higher densities of the predator or herbivore than would normally be supported by
the native prey. This may be a result of co-evolution between an introduced prey
species and its introduced predator (the mediator of this interaction).
In contrast to apparent competition, interactions in which the alien species
occupies a higher trophic level than the species mediating the interaction – i.e.
exploitative competition and trophic cascades – are more frequently reported to be
mediated by native species than aliens. The reason for this is unclear, but may
simply reflect the approach of the studies, which initially focused on direct
interactions between the alien and native species.
Invasion biology studies are commonly designed to determine the impact of one
species on another, but not to determine the mechanism responsible for the
impact. Thus indirect effects – if identified at all - may be discovered incidentally in
studies which weren’t originally designed to detect them (e.g. Sessions and Kelly
2002). As a result, indirect effects might be presumed or hypothesized to be
responsible for an observed effect (e.g. Sessions and Kelly 2002), but are rarely
directly tested for. Conversely, there are studies in which the potential mechanism
of an indirect effect is detected but the impact remains untested (Adams et al.
2003; Lenz and Taylor 2001). The abundance of leafhopper Sophonia rufofascia,
for example, is shown to be higher on the native Hawaiian tree Metrosideros
polymorpha where the exotic tree Myrica faya is present (Lenz and Taylor 2001)
but the resulting impact (if any) on the native tree species is unknown. Future
studies designed to determine both the impact and the mechanism behind the
impact of an invader on a native species will provide an insight into the relative
importance of direct and indirect effects in structuring ecological systems.

53
Empirical evidence is mounting to suggest that indirect effects may be important in
influencing the outcome of invasions and the impacts of an invader on native
species. It is likely that in some cases observed impacts that are attributed to more
obvious direct interactions (such as interspecific competition) might in fact be
caused by other indirect effects (e.g. apparent competition), and these more
complex interactions should be taken into consideration when conducting
experiments designed to investigate impacts of exotic on native species. Further
investigation of indirect effects will give us a better understanding and predictive
ability of the range of potential impacts of an invasive species and might assist us
in designing management strategies both for invasive and native species.
Furthermore, from a theoretical viewpoint, a system that has experienced a
species addition can provide the ideal opportunity to investigate the importance of
indirect effects in structuring ecological systems.


Chapter 3 - Diversity and abundance of
arthropod floral visitor and herbivore
assemblages on alien and native Senecio
species
The following chapter is currently in press as:
White, E.M. and Clarke, A.R. (in press) Diversity and abundance
of arthropod floral visitor and herbivore assemblages on alien
and native Senecio species. Plant Protection Quarterly
The roles of co-authors are as follows:
E.M. White: Designed the experiment, conducted all fieldwork, and data analysis
and wrote the manuscript.
A.R. Clarke: Project supervisor, provided advice on experimental design and data
analysis and assisted with writing drafts of the manuscript.


57
Chapter 3 – Diversity and abundance of arthropod floral visitor and herbivore
assemblages on alien and native Senecio species
3.1 Abstract
The enemy release hypothesis predicts that native herbivores prefer native, rather
than exotic plants, giving invaders a competitive advantage. In contrast, the biotic
resistance hypothesis states that many invaders are prevented from establishing
because of competitive interactions, including herbivory, with native fauna and
flora. Success or failure of spread and establishment might also be influenced by
the presence or absence of mutualists, such as pollinators. Senecio
madagascariensis (fireweed), an annual weed from South Africa, inhabits a similar
range in Australia to the related native S. pinnatifolius. The aim of this study was
to determine, within the context of invasion biology theory, whether the two
Senecio species share insect fauna, including floral visitors and herbivores.
Surveys were carried out in south-east Queensland on allopatric populations of the
two Senecio species, with collected insects identified to morphospecies. Floral
visitor assemblages were variable between populations. However, the two Senecio
species shared the two most abundant floral visitors, honeybees and hoverflies.
Herbivore assemblages, comprising mainly hemipterans of the families
Cicadellidae and Miridae, were variable between sites and no patterns could be
detected between Senecio species at the morphospecies level. However, when
insect assemblages were pooled (i.e. community level analysis), S. pinnatifolius
was shown to host a greater total abundance and richness of herbivores. Senecio
madagascariensis is unlikely to be constrained by lack of pollinators in its new
range and may benefit from lower levels of herbivory compared to its native
congener S. pinnatifolius.

58
3.2 Introduction
It is widely accepted that biotic interactions between alien and native species can
influence the success (or otherwise) of an invasive species (Colautti et al. 2004;
Richardson et al. 2000; Stastny et al. 2005). The enemy release hypothesis (ERH)
(Darwin 1859; Elton 1958; Keane and Crawley 2002) posits that alien plant species
benefit in their area of introduction because they have escaped their natural
enemies (including herbivores, parasites and pathogens). This hypothesis is based
on the assumptions that: (1) natural enemies regulate plant populations; (2)
enemies prefer native over alien species; and (3) plants introduced to a new area
benefit from reduced attack by enemies (Keane and Crawley 2002). In contrast,
the biotic resistance hypothesis (Elton 1958; Keane and Crawley 2002) states that
many potential invaders fail to establish because of strong interactions, such as
competition, parasitism and herbivory, with native species in the new area.
Evidence in support of the ERH has been provided by a number of studies that
have shown that: (i) alien plants host fewer species of insect herbivore than do
congeneric natives in their new range (Olckers and Hulley 1991; van der Putten et
al. 2005); (ii) alien species experience lower rates of herbivory than do congeneric
or ecologically similar natives (Dietz et al. 2004; Olckers and Hulley 1991;
Schierenbeck et al. 1994; Siemann and Rogers 2003; White et al. in press); (iii)
invaders are better able to compensate for the effects of herbivory than are
congeneric natives (Rogers and Siemann 2002); and (iv) invaders host fewer
species of phytophagous insects in a new range than in their native range (Wolfe
2002). Other studies, in contrast, have supported the biotic resistance hypothesis,
showing that: (i) the abundance of insect herbivores on alien plants to be equal to
or greater than that on congeneric natives (Burki and Nentwig 1997; Frenzel and
Brandl 2003; Novotny et al. 2003; Torrusio et al. 2002); (ii) some invaders
experience levels of herbivory equal to or greater than that of related native

59
species (Agrawal and Kotanen 2003); and (iii) herbivory can have a strong
negative effect on invader establishment (Levine et al. 2004).
Whilst the enemy release and biotic resistance hypotheses focus on negative
interactions, other authors have highlighted the importance of positive biotic
interactions in influencing invasion success or failure (Larson et al. 2002; Parker
1997; Parker and Haubensak 2002; Parker 2001; Richardson et al. 2000). The
establishment and spread of an invasive plant might depend in part on the
presence of mutualists, including seed dispersers (Constible et al. 2005;
Richardson et al. 2000), mycorrhiza (Parker 2001), or pollinators (Byers et al.
2002; Larson et al. 2002). The spread of an invader may be limited by the lack of
pollinators at the edge of its new range (Larson et al. 2002; Parker 1997; Parker
and Haubensak 2002), although given the generalist nature of many plant-
pollinator interactions (Herrera 2005; Vazquez and Aizen 2004), this is probably
the exception rather than the rule (Herrera 2005; Richardson et al. 2000; Vazquez
and Aizen 2004). In fact, several studies have implicated a relationship between
the presence of generalist pollinators (often alien honeybees, Apis mellifera, which
are widespread and abundant in many areas) and the spread of invasive plant
species (Goulson and Derwent 2004; Hanley and Goulson 2003; Stout et al. 2002;
Turner and Conran 2004).
Since generalist pollinators are common in natural systems (Richardson et al.
2000), it seems probable that an invasive species will be visited by a similar suite
of pollinators to that visiting the flowers of native species. As demonstrated by the
conflicting results of studies of herbivory on alien species however (e.g. Burki and
Nentwig 1997; Frenzel and Brandl 2003; Olckers and Hulley 1991; van der Putten
et al. 2005), it can be more difficult to predict the degree of utilisation of alien plants
by native herbivores. Owing to subtle physical and chemical differences in plant

60
tissue, or learned behaviour in insects, even two closely related, morphologically
similar plant species can vary in their relative attractiveness to arthropod
herbivores (Cunningham and West 2001; Foss and Rieske 2003; Ladner and
Altizer 2005; Olckers and Hulley 1991).
Several studies (as mentioned above) have compared herbivore assemblages and
impact of herbivory between invasive and native plant species, whilst others have
focussed on pollinators shared by invasive and native species. However, to our
knowledge, no single study has simultaneously compared these two faunal groups
between an alien and native species. The aim of this study is to determine the
degree of similarity in arthropod herbivores and floral visitors between native
Senecio. pinnatifolius A. Rich and the closely related and morphologically similar
alien S. madagascariensis Poiret, in eastern Australia. Sharing of arthropod
herbivores and pollinators may have implications for the spread of the invasive
species, as well creating the potential for the occurrence of indirect interactions,
such as competition for pollinators, hybridisation and apparent competition,
between the two species.
3.3 Materials and Methods
3.3.1 Study species
Senecio madagascariensis (fireweed), an annual weed from South Africa, was first
recorded in Australia in 1918 and has since invaded large areas of farmland and
grassland in south-eastern Australia (Radford 1997; Radford et al. 1995). In south-
eastern Australia, S. madagascariensis flowers between the months of March and
December (Radford 1997).
Closely related to S. madagascariensis is a group of sub-species belonging to the
Australian native S. pinnatifolius complex. Senecio pinnatifolius is a herbaceous

61
perennial (Ali 1966), which is found in a similar range to that of S.
madagascariensis in Australia but generally occurs in smaller, more scattered
populations than the exotic (Radford 1997). Senecio pinnatifolius ssp. lanceolatus,
the focus of this study, inhabits disturbed areas and pasture usually close to the
edge of rainforest or moist eucalypt forest and flowers between January and June
in south-eastern Australia (Radford 1997). There is a four month period of overlap
between the flowering periods of the exotic and native Senecio. Senecio
pinnatifolius and S. madagascariensis are both believed to be self-incompatible
and are likely to rely on insects as pollinators (Ali 1966; Radford 1997).
3.3.2 Study sites
Insect sampling was conducted at seven locations in south-east Queensland,
encompassing four populations of S. pinnatifolius and three populations of S.
madagascariensis. Populations of the former were Swanfels 1 (28o 07’S, 152o
23’E), Swanfels 2 (28o 08’S, 152o 23’E), Bunya Mountains (Bunya) (26o 53’S, 151o
35’E) and Hampton (27o 22’S, 152o 10’E), while populations of the latter were
Springbrook (28o 11’S, 153o 16’E), Tamborine (27o 58’S, 153o12’E) and Beechmont
(28o 07’S, 153o 10’E). All sites, with the exception of Bunya, occur within an
approximately 120 km length of the “Border Ranges”, a group of linked mountain
ranges running along the eastern portion of the Queensland/NSW state border.
The Bunya Mountains are approximately 200 km north-west of the nearest
neighbouring site (Hampton) and are separated from the Border ranges by farmed
plain-lands. All populations, regardless of site, occurred within a similar altitudinal
range (between 550 m and 700 m ASL), had similar types of neighbouring
vegetation (pasture and moist eucalypt forest or rainforest), and were surveyed
between March and May when both species were flowering.

62
3.3.3 Survey Methodology
In order to collect as wide a range of insect species as possible, three different
collection techniques, described below, were used on all plants.
Floral visitor observations
Thirty random plants per population were used for floral visitor observations. Two
observers each monitored one plant at a time, recording the species, where
possible, and number of insects visiting flowers on a plant during a five minute
observation period, before moving to another plant. Each observer conducted a
total of six, five-minute observations per hour, between the hours of 8 am and 4
pm. This process was repeated over two days at each of the seven sites on sunny,
windless days during which the temperature in the shade ranged between 17 and
23oC. The height of each plant was also recorded as an index of plant size.
Manual search
Thirty random plants per population were searched by hand. For each plant, stems
and upper and lower surfaces of the newest 20 leaves on 10 randomly selected
branches were examined and identity and abundance of all arthropod species
occurring on the plant was recorded. When identification was impossible in the field
insect specimens were collected for later identification. The height of each plant
was again recorded.
Sweep-net sampling
Using the same 30 plants per site as were surveyed during the manual search,
arthropods were collected with a sweepnet using three sweeps per plant. Insects
were held in specimen jars for later identification.

63
Specimen identification
All specimens collected were initially categorised to morphospecies level and then
identified to family using keys in Naumann et al. (1991). Except for very common
species, e.g. Apis mellifera, formal species identification was not undertaken. Since
this experiment was conducted as part of a broader study requiring continual
monitoring of floral visitors, which would have been disrupted by collecting every
individual that landed on a plant, specimens were not collected for some of the less
common floral visitors. Consequently, some morphospecies of uncommon
lepidopteran floral visitors were identified only to ordinal level. For similar reasons,
individual syrphid (Diptera: Syrphidae) species were identified only to family level
as accurate morphospecies identification for flies on the wing was found to be
unachievable. Despite our efforts to include as many arthropod groups as possible
by using a range of sampling procedures, some of the groups of smaller
arthropods (e.g. aphids) may have been under sampled.
Data analysis
Data analyses were performed with SPSS Vs 12.0.1. When data were not normally
distributed, variables were log (n+1) transformed. In order to test whether sites
varied in terms of herbivore abundance and species richness, one-way analyses of
variance (ANOVA) were performed using morphospecies abundance (or species
richness) per plant as the dependent variable and site as the independent variable
(Fowler and Cohen 1990).
Sørensen’s similarity indices, using presence/absence data were used to assess
the degree of similarity in arthropod assemblages between sites, and dendrograms
were created using hierarchical, average linkage cluster analyses based on
Sørensen’s similarity coefficients.

64
Senecio madagascariensis and S. pinnatifolius differ in size, with the latter species
generally being larger: within sites, there is also variation in individual plant size. In
order to determine if we needed to correct for size variation prior to analyse,
spearman rank correlation was performed on herbivore abundance and floral
visitor data for all sites to determine whether there existed a relationship between
plant size (using height as an index) and herbivore or floral visitor abundance. We
found no significant correlations between plant height and herbivore abundance at
any site, and a significant correlation between plant height and floral visitors for
only two sites, one of each Senecio species (authors’ unpublished data). We
therefore concluded that plant size was not a driver of invertebrate abundance in
this system and we did not correct for plant size in subsequent analyses.
a
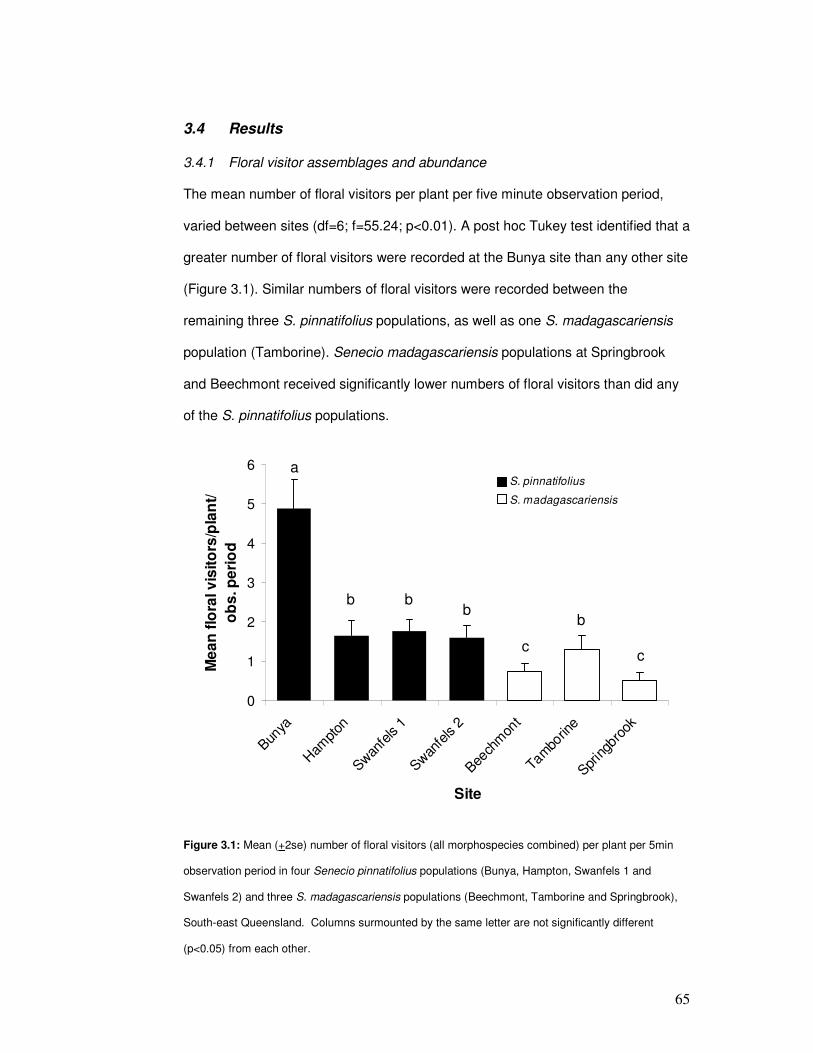
65
3.4 Results
3.4.1 Floral visitor assemblages and abundance
The mean number of floral visitors per plant per five minute observation period,
varied between sites (df=6; f=55.24; p<0.01). A post hoc Tukey test identified that a
greater number of floral visitors were recorded at the Bunya site than any other site
(Figure 3.1). Similar numbers of floral visitors were recorded between the
remaining three S. pinnatifolius populations, as well as one S. madagascariensis
population (Tamborine). Senecio madagascariensis populations at Springbrook
and Beechmont received significantly lower numbers of floral visitors than did any
of the S. pinnatifolius populations.
0
1
2
3
4
5
6
Bunya
Ham
pton
Swan
fels 1
Swan
fels 2
Beech
mon
t
Tambo
rine
Sprin
gbro
ok
Site
Me
an
flo
ral v
isit
ors
/pla
nt/
ob
s. p
eri
od
a
b bb
b
c c
S. pinnatifolius
S. madagascariensis
Figure 3.1: Mean (+2se) number of floral visitors (all morphospecies combined) per plant per 5min
observation period in four Senecio pinnatifolius populations (Bunya, Hampton, Swanfels 1 and
Swanfels 2) and three S. madagascariensis populations (Beechmont, Tamborine and Springbrook),
South-east Queensland. Columns surmounted by the same letter are not significantly different
(p<0.05) from each other.
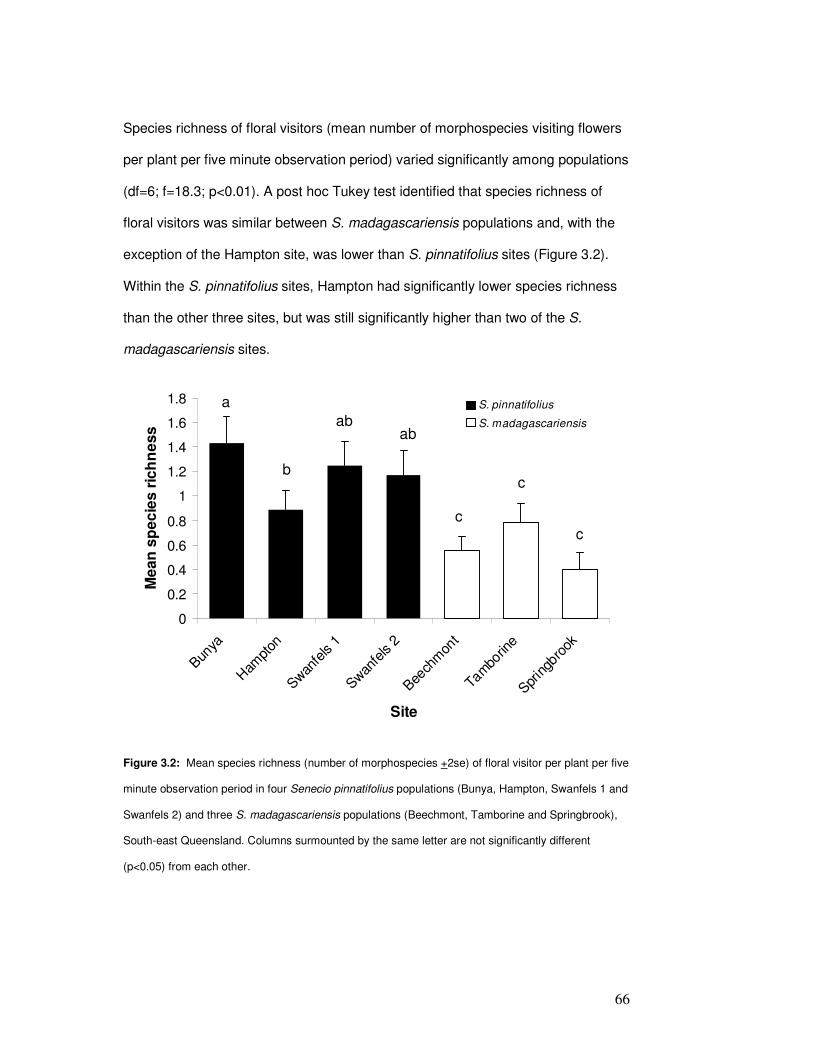
66
Species richness of floral visitors (mean number of morphospecies visiting flowers
per plant per five minute observation period) varied significantly among populations
(df=6; f=18.3; p<0.01). A post hoc Tukey test identified that species richness of
floral visitors was similar between S. madagascariensis populations and, with the
exception of the Hampton site, was lower than S. pinnatifolius sites (Figure 3.2).
Within the S. pinnatifolius sites, Hampton had significantly lower species richness
than the other three sites, but was still significantly higher than two of the S.
madagascariensis sites.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
Bunya
Ham
pton
Swan
fels 1
Swan
fels 2
Beech
mon
t
Tambo
rine
Sprin
gbro
ok
Site
Me
an
sp
ec
ies
ric
hn
ess
a
b
abab
c
c
c
S. pinnatifolius
S. madagascariensis
Figure 3.2: Mean species richness (number of morphospecies +2se) of floral visitor per plant per five
minute observation period in four Senecio pinnatifolius populations (Bunya, Hampton, Swanfels 1 and
Swanfels 2) and three S. madagascariensis populations (Beechmont, Tamborine and Springbrook),
South-east Queensland. Columns surmounted by the same letter are not significantly different
(p<0.05) from each other.

67
The majority of floral visitors at all sites belonged to the orders Diptera,
Hymenoptera and Lepidoptera (see Appendix A). All seven Senecio populations
were similar in the respect that all shared the same two most common floral
visitors: Apis mellifera, which comprised between 43 and 83% of all floral visitors at
each site; and syrphid species, which comprised between 8 and 37% of floral
visitors at each site. Rarer species however, were variable between sites, with
many taxa recorded in very low densities, and often only at a single site.
A hierarchical cluster analysis of presence/absence data showed two large scale
clusters, with the geographically close S. pinnatifolius populations Swanfels 1 and
Swanfels 2 forming one cluster, and the remaining five sites forming the other
(Figure 3.3). Within the larger group, smaller terminal clusters could not be
explained by either geography or plant species. For example, although the
terminal pair of Bunya and Beechmont had very similar faunas (Table 3.1), they
represent different plant species and, of all sites, are the furthest geographically
apart.
Figure 3.3: A hierarchical, average linkage cluster analysis (using Sorensen’s similarity coefficients)
for floral visitor assemblages in four Senecio pinnatifolius populations (Bunya, Hampton, Swanfels 1
and Swanfels 2) and three S. madagascariensis populations (Beechmont, Tamborine and
Springbrook), South-east Queensland.

68
Table 3.1: Sørenson similarity indices (above the diagonal) and species overlap (below the diagonal)
for floral visitor and herbivore assemblages between four populations of Senecio pinnatifolius and
three populations of S. madagascariensis in SE Queensland. The number in bold on the diagonal is
the number of floral visitors, or herbivores, recorded at a particular site.
B
un
ya
Ham
pto
n
Sw
an
fels
1
Sw
an
fels
2
Be
ech
mo
nt
Tam
bori
ne
Sp
ringbro
ok
Bunya 8 0.59 0.40 0.42 0.77 0.46 0.50
Hampton 4 9 0.40 0.42 0.71 0.43 0.46
Swanfels 1 4 3 12 0.52 0.35 0.35 0.38
Swanfels 2 4 4 6 11 0.50 0.38 0.40
Beechmont 5 5 3 4 5 0.60 0.67
Tamborine 3 3 3 3 3 5 0.67
Flo
ral vis
itors
Springbrook 3 3 3 3 3 3 4
Bunya 10 0.35 0.26 0.50 0.43 0.43 0.00
Hampton 3 7 0.20 0.24 0.55 0.55 0.00
Swanfels 1 3 2 13 0.61 0.24 0.24 0.13
Swanfels 2 5 2 7 10 0.29 0.29 0.17
Beechmont 3 3 2 2 4 1.00 0.00
Tamborine 3 3 2 2 4 4 0.00
Herb
ivore
s
Springbrook 0 0 1 1 0 0 2
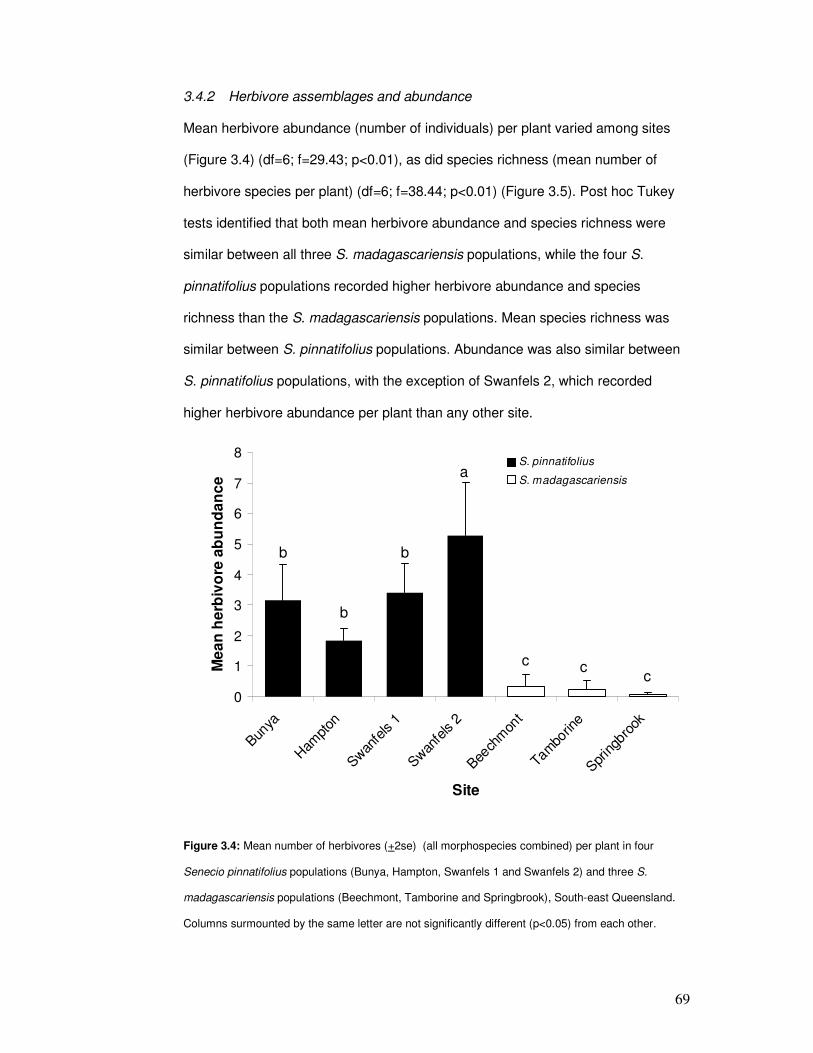
69
3.4.2 Herbivore assemblages and abundance
Mean herbivore abundance (number of individuals) per plant varied among sites
(Figure 3.4) (df=6; f=29.43; p<0.01), as did species richness (mean number of
herbivore species per plant) (df=6; f=38.44; p<0.01) (Figure 3.5). Post hoc Tukey
tests identified that both mean herbivore abundance and species richness were
similar between all three S. madagascariensis populations, while the four S.
pinnatifolius populations recorded higher herbivore abundance and species
richness than the S. madagascariensis populations. Mean species richness was
similar between S. pinnatifolius populations. Abundance was also similar between
S. pinnatifolius populations, with the exception of Swanfels 2, which recorded
higher herbivore abundance per plant than any other site.
0
1
2
3
4
5
6
7
8
Bunya
Ham
pton
Swan
fels 1
Swan
fels 2
Beech
mon
t
Tambo
rine
Sprin
gbro
ok
Site
Me
an
he
rbiv
ore
ab
un
da
nc
e
b b
b
a
c c c
S. pinnatifolius
S. madagascariensis
Figure 3.4: Mean number of herbivores (+2se) (all morphospecies combined) per plant in four
Senecio pinnatifolius populations (Bunya, Hampton, Swanfels 1 and Swanfels 2) and three S.
madagascariensis populations (Beechmont, Tamborine and Springbrook), South-east Queensland.
Columns surmounted by the same letter are not significantly different (p<0.05) from each other.
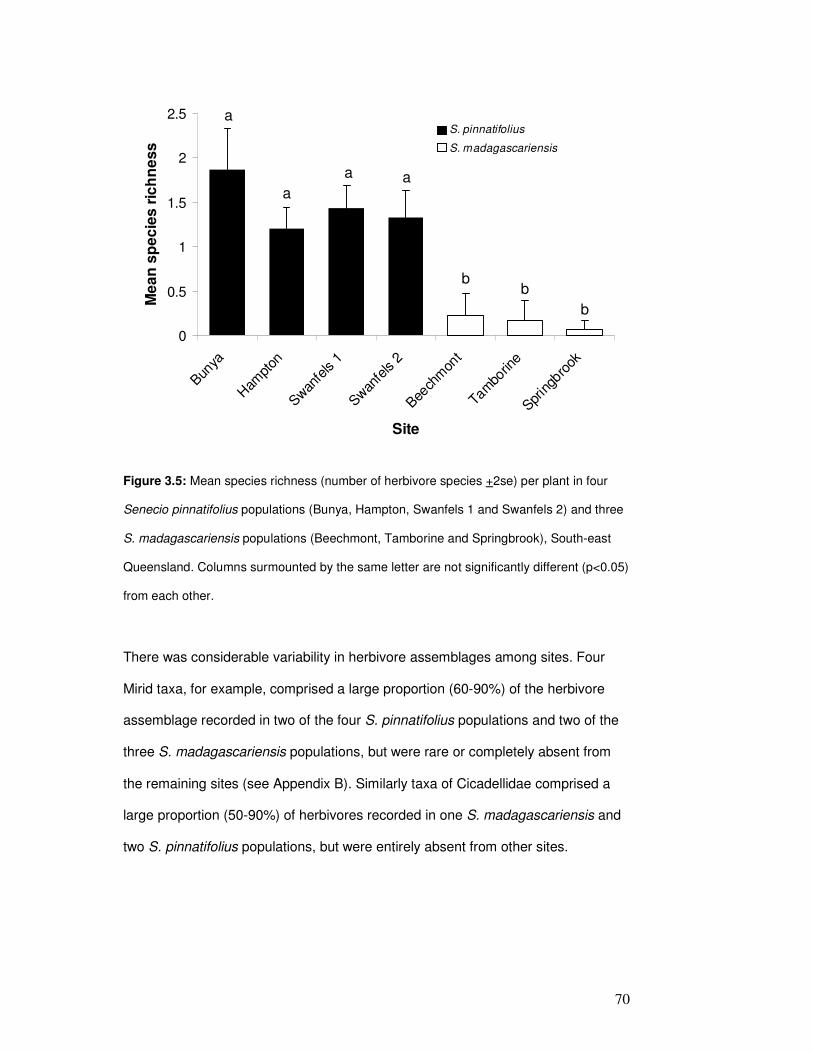
70
0
0.5
1
1.5
2
2.5
Bunya
Ham
pton
Swan
fels 1
Swan
fels 2
Beech
mon
t
Tambo
rine
Sprin
gbro
ok
Site
Me
an
sp
ec
ies
ric
hn
ess
a
a
a a
b b
b
S. pinnatifolius
S. madagascariensis
Figure 3.5: Mean species richness (number of herbivore species +2se) per plant in four
Senecio pinnatifolius populations (Bunya, Hampton, Swanfels 1 and Swanfels 2) and three
S. madagascariensis populations (Beechmont, Tamborine and Springbrook), South-east
Queensland. Columns surmounted by the same letter are not significantly different (p<0.05)
from each other.
There was considerable variability in herbivore assemblages among sites. Four
Mirid taxa, for example, comprised a large proportion (60-90%) of the herbivore
assemblage recorded in two of the four S. pinnatifolius populations and two of the
three S. madagascariensis populations, but were rare or completely absent from
the remaining sites (see Appendix B). Similarly taxa of Cicadellidae comprised a
large proportion (50-90%) of herbivores recorded in one S. madagascariensis and
two S. pinnatifolius populations, but were entirely absent from other sites.

71
Figure 3.6: A hierarchical, average linkage cluster analysis (using Sorensen’s similarity
coefficients) for arthropod herbivore assemblages in four Senecio pinnatifolius populations
(Bunya, Hampton, Swanfels 1 and Swanfels 2) and three S. madagascariensis populations
(Beechmont, Tamborine and Springbrook), South-east Queensland.
Hierarchical cluster analysis of presence/absence data showed grouping of two of
the S. madagascariensis populations, Beechmont and Tamborine (Figure 3.6),
indicating very similar herbivore assemblages (comprised mainly of mirids (see
Appendix B)). However, the third population, Springbrook, was quite distinct from
any other site. This can be explained by the fact that only two morphospecies were
collected from this site, one of which (“Cicadellidae 3”) was also recorded in two of
the S. pinnatifolius populations. Similar herbivore assemblages (comprised largely
of cicadellids) were recorded between the geographically close S. pinnatifolius
populations, Swanfels 1 and Swanfels 2 (Table 3.1). Cicadellids were not recorded
at the other S. pinnatifolius sites, Bunya and Hampton, whose herbivore fauna
comprised mainly of mirids. Folivores, including lepidopteran larvae and
orthopterans, were recorded in low numbers in all S. pinnatifolius populations and
one S. madagascariensis population, the most frequently recorded being arctiidae
larvae (“Arctiidae 1”, which was identified as Nyctemera amica).

72
3.5 Discussion
Although the data showed a trend towards a higher floral visitation rate and higher
floral visitor species richness in S. pinnatifolius populations, this was not consistent
for all sites and the differences observed may be partially explained by factors such
as variability in number of flowers per plant between sites, or between the exotic
and native species. At all sites hoverflies and honeybees were the most common
insect groups recorded. Our data therefore support the prediction that the flowers
of exotic S. madagascariensis would be visited by a group of generalist floral
visitors similar to that which visits native S. pinnatifolius.
Whilst the most common floral visitors were similar between all populations,
distributions of the rare species were highly variable with many taxa recorded in
very low densities and only at single sites. Similarities and differences among
species assemblages at different sites could not be explained entirely by
geographic location, nor did sites consistently cluster based on plant species,
indicating that in this system other factors, perhaps abiotic variables at a landscape
or microsite-level, may play a more important role than intrinsic plant features in
influencing pollinator assemblages. Similar findings were recorded by Herrera
(2005), who found that at a regional level the pollination system of the insect-
pollinated shrub Lavandula latifolia was generalised, but at both the individual and
population levels generalisation was highly variable.
Ornduff (1960) described the pollinations roles of hoverflies and honeybees in the
New Zealand subspecies of S.lautus (closely related to S. pinnatifolius) and it is
probable that these species act as important pollinators for Senecio in Australia.
This is further confirmed by the fact that examination of the bodies of both
hoverflies and honeybees using scanning electron microscopy techniques, shows
that both groups of insects carry large quantities of Senecio pollen (E. White,

73
unpublished data). The fact that S. madagascariensis receives frequent visits by
the same floral visitors as S. pinnatifolius indicates that it is able to utilise local
(albeit introduced, in the case of A. mellifera) generalist pollen vectors. It therefore
seems highly unlikely that the spread and establishment of the invasive Senecio is
restricted by lack of pollen vectors, at least in this part of its range.
Sharing of floral visitors by two sympatric plant species creates the potential for
indirect interactions, which could have a negative or positive impact on one or both
species involved. Documented indirect interactions between invasive and native
plant species that share common pollinators include competition for pollinators
(Ghazoul 2002; Gross and Werner 1983; Lavergne et al. 2005; Moragues and
Travaset 2005), facilitation of pollination (Moragues and Travaset 2005), and
transfer of pollen between species which may result in either reduced seed set due
to foreign pollen interference (Brown and Mitchell 2001; Galen and Gregory 1989;
Grabas and Laverty 1999) or hybridisation (Abbott 1992; Ayres et al. 1999; Bleeker
2003; Huxel 1999; Shibaike et al. 2002). Given that the two Senecio species share
pollinators, overlap in flowering period, sometimes occur sympatrically (Radford
1997), and have the ability to hybridise (Radford 1997), potential exists for any one
of these interactions to take place in this system. We are currently undergoing
further work to determine whether such interactions are occurring.
When considered at the morphospecies level, herbivore assemblages showed
even greater variability than did floral visitor assemblages among sites, with
particular taxa (largely sap-sucking mired and cicadellid morphospecies)
comprising a large proportion (60-90%) of the herbivore assemblage at some sites,
whilst being rare or apparently absent from others. Although our data would
indicate that some taxa are restricted to S. pinnatifolius and absent from S.
madagascariensis populations, the extremely low density of the majority of taxa

74
within-sites, combined with the very high between-site variation in species
assemblages, makes it impossible to identify a clear pattern with regards to the
preferences of individual herbivore taxa for either Senecio species. As with the
floral visitors, the degree of similarity between the herbivore assemblages of
different populations could not be entirely explained by either geographical location
or by plant species.
When herbivores are considered as a group, rather than at morphospecies level, a
clear pattern does become evident however: S. pinnatifolius populations show
consistently higher herbivore abundance and species richness than do the exotic
S. madagascariensis populations. Senecio pinnatifolius has been shown to be
more attractive than S. madagascariensis to ovipositing females and feeding larvae
of the magpie moth, Nyctemera amica (White et al. in press). The current study
offers further evidence of the greater attractiveness of the native species to a range
arthropod herbivores, providing support for the enemy release hypothesis. The
profiles of the most common secondary chemicals found in Senecio species, the
pyrrolizidine alkaloids (Rothschild et al. 1979), vary between the two Senecio
species here (Sims 2004) and may play a role in determining the species’ relative
attractiveness or palatability to phytophagous insects.
3.5.1 Conclusions
At the morphospecies level, arthropod herbivore assemblages are highly variable
within and between populations of exotic and native Senecio species and this
variability cannot be adequately explained either by plant species or by geographic
location of populations. However, as a group, herbivores are more abundant on
the native S. pinnatifolius both in terms of number of taxa and number of
individuals per plant. Like the herbivores, most of the less common floral visitors
are also highly variable among sites, with a tendency towards increased number

75
and richness on the native species. However, the most abundant floral visitors
recorded in S. pinnatifolius populations, hoverflies and honeybees, are also the
most frequent visitors recorded on S. madagascariensis. The success of invasive
S. madagascariensis may be explained in part by the fact that it is unlikely to be
disadvantaged by the absence of positive interactions (namely plant-pollinator
interactions) in its new range, while at the same time (as predicted under the ERH)
having the advantage of being less attractive to native herbivores than is its native
congener S. pinnatifolius.


Chapter 4 - A test of the enemy release
hypothesis: The native magpie moth
prefers a native fireweed (Senecio
pinnatifolius) to its introduced congener
(S. madagascarensis)
The following chapter was recently published as:
White, E.M., Sims, N.M. and Clarke, A.R. (2008) A test of the
enemy release hypothesis: The native magpie moth prefers a
native fireweed (Senecio pinnatifolius) to its introduced congener
(S. madagascarensis). Austral Ecology 33: 110-116.
The roles of co-authors are as follows:
E.M. White: Largely responsible for the experimental design, wholly responsible for
the fieldwork and most of the data analysis and writing.
N.M. Sims: Conducted laboratory studies, and performed associated statistical
analyses, and contributed to first draft of manuscript.
A.R. Clarke: Project supervisor, provided advice about experimental design and
statistical analyses, assisted with addressing reviewers’ comments.


79
Chapter 4 - A test of the enemy release hypothesis: The native magpie moth
prefers a native fireweed (Senecio pinnatifolius) to its introduced congener
(S. madagascarensis).
4.1 Abstract
The enemy release hypothesis (ERH) predicts that native herbivores will either
prefer or cause more damage to native than introduced plant species. We tested
this using preference and performance experiments in the laboratory and surveys
of leaf damage caused by the magpie moth Nyctemera amica on a co-occuring
native and introduced species of fireweed (Senecio) in eastern Australia. In the
laboratory, ovipositing females and feeding larvae preferred the native S.
pinnatifolius over the introduced S. madagascariensis. Larvae performed equally
well on foliage of S. pinnatifolius and S. madagascariensis: pupal weights did not
differ between insects reared on the two species, but growth rates were
significantly faster on S. pinnatifolius. In the field, foliage damage was significantly
greater on native S. pinnatifolius than introduced S. madagascariensis. These
results support the enemy release hypothesis and suggest that the failure of native
consumers to switch to introduced species contributes to their invasive success.
Both plant species experienced reduced, rather than increased, levels of herbivory
when growing in mixed populations, as opposed to pure stands in the field, thus
there was no evidence that apparent competition occurred.

80
4.2 Introduction
The enemy release hypothesis (ERH) (Darwin 1859; Elton 1958; Keane and
Crawley 2002) posits that introduced plant species benefit in their area of
introduction because they have escaped from their natural enemies. This
hypothesis is based on the assumptions that: (1) natural enemies regulate plant
populations, (2) enemies prefer native over introduced species; and (3) plants
introduced to a new area benefit from reduced attack by enemies (Keane and
Crawley 2002). Empirical support for the ERH is equivocal; some native herbivores
prefer or have a greater negative impact on native than co-occurring introduced
plant species (Olckers and Hulley 1991; Schierenbeck et al. 1994; Keane and
Crawley 2002; Lankau et al. 2004), others show no preference (Frenzel and Brandl
2003; Tamayo et al. 2004) or a preference for introduced plants over co-occurring
natives (Agrawal and Kotanen 2003; Parker and Hay 2005; Parker et al. 2006).
Even when herbivores prefer a particular plant species, neighbouring species
might also be impacted by herbivory as a result of apparent competition (Noonburg
and Byers 2005). Apparent competition occurs when a herbivore becomes more
numerous or more efficient at consuming one species in the presence of another
(Holt 1977). The altered spatial or temporal patterns of resource availability
provided by one plant species can result in altered abundance or behaviour of
herbivores, consequently having a greater negative impact on a second plant
species (Holt 1977). Apparent competition has been documented between
sympatric native species (Hämback and Ekerholm 1997; Rand 2003) but is less
commonly reported between introduced and native species (White et al. 2006).
Such an interaction could potentially have an adverse effect on either an invader or
a co-occurring native species, depending on the population dynamics of the plant
and herbivore population.

81
Host plant switches and range expansions by specialist herbivores are more likely
to occur when the native and introduced host plants are closely related (Connor et
al. 1980). Senecio madagascariensis Poir. (fireweed), an introduced Asteraceae, is
closely related to a group of native subspecies belonging to the S. pinnatifolius
Rich. complex. Senecio madagascariensis, an annual or short-lived perennial from
South Africa was first recorded in Australia in 1918 (Radford et al. 1995a) and has
since invaded large areas of arable land and grassland in eastern Australia
(Radford and Cousens 2000). Senecio pinnatifolius is an herbaceous perennial (Ali
1966), generally found in smaller, more scattered populations than the introduced
S. madagascariensis (Radford and Cousens 2000). Populations of the two species
are often found close together or intermixed, with S. pinnatifolius growing along the
borders of disturbed areas or pasture occupied by S. madagascariensis (Radford
1997).
The two species share a number of insect herbivores, one of the most common
folivores being the magpie moth, Nyctemera amica (Holtkamp and Hosking 1993),
a pyrrolizidine alkaloid specialist that is restricted to Senecio species (Common
1993). Female N. amica, oviposit on Senecio leaves, which provide a food source
for the developing larvae (Singh and Mabbett 1976). The close relatedness of the
introduced and native Senecio species and the overlapping geographic range of
the two species make this an ideal system in which to study herbivore preference
and apparent competition between a native and an invasive species.
We test one of the assumptions underpinning the ERH: that native herbivores
exhibit a preference for native plant species over an introduced species. We aim to
determine whether (a) N. amica larvae and adults prefer native Senecio
pinnatifolius, and if so, whether this is reflected in (b) greater larval growth and
survival and (c) higher damage levels in the field. Furthermore, this study aims to

82
determine whether either Senecio species experiences altered damage levels due
to herbivory as a result of growing in mixed populations with the other species.
4.3 Materials and Methods
Adult Nyctemera amica were collected in Lamington National Park (28o08′S,
153o06′E), as well as suburban areas of South East Queensland. The majority of
the adult females had already mated and laid eggs without further access to a male
moth. Eggs are laid in batches of between 2-50 eggs. Larvae from these egg
batches were used in three laboratory experiments, whose methods are
subsequently described. Adults were sexed by their antennal morphology, with
males having more distinctly plumose antennae than females. Fresh stems of S.
pinnatifolius ssp. lanceolatus and S. madagascariensis were collected regularly
from Binna Burra (within Lamington National Park) and Hampton (27o15′S,
152o04′E). Branches were stored in a cool room at approximately 6oC until use. In
areas where S. pinnatifolius and S. madagascariensis co-occur, plants were
identified by leaf morphology and bract number. Senecio pinnatifolius has 12-20
bracts compared to S. madagascariensis’ 19-21 bracts (Radford 1997; Radford
and Cousens 2000). Leaves of S. pinnatifolius tend to be serrated and larger than
those of S. madagascariensis, whilst S. madagascariensis usually has entire leaf
margins.
4.3.1 Larval Feeding and Survival
Neonate larvae (< 24 hrs old) were placed individually into plastic Petri dishes
(85mm diameter). Approximately half of the larvae (n = 37) were reared on S.
pinnatifolius leaves, whilst the remainder (n = 30) were offered S.
madagascariensis: each larva was offered the same wet weight of leaf material
throughout its life. The weight of leaves offered to larvae was doubled each week
after their hatching date, with the weight of leaf first offered to neonates being

83
0.2g/larva. Leaf material was replenished daily and available leaf material was
always in excess of daily consumption. Larvae were kept in a controlled
environment of 25 ±1oC and L12:D12. Pupal weight was recorded 48 h after
pupation and the number of days from hatching till pupation began was also
recorded. Pupal weight and number of days till pupation of larvae reared on each
species were compared using one-way ANOVA. The numbers of surviving larvae
were compared using a chi-square test for association. Larvae which did not
survive through to pupation were excluded from these analyses.
4.3.2 Larval Preference
To determine the feeding preference of larvae throughout their life, choice tests
were conducted in the laboratory. Between five and 10 newly hatched neonate
larvae were placed in round plastic containers (approx. 113 cm2) with an equal
weight of leaves of both S. pinnatifolius and S. madagascariensis. The weight of
leaves provided for larvae varied with each instar, however, there was always
excess of each plant species to ensure apparent preference did not change due to
the lack of a particular species. The number of larvae feeding on each species was
recorded hourly for six hours, for one day in each larval instar. Larvae not on either
plant species at the time of observation were excluded from the analysis.
Independent cohorts of larvae were used for each instar so that learnt preference
or avoidance did not confound results. The mean proportion of larvae feeding on
each species across all instars was analysed using a one-sample t-test, comparing
the proportion to a test value of 50 (which assumes an equal preference for each
plant species). A one-way ANOVA on each host species was used to determine
whether larval preference changed across instars.

84
4.3.3 Oviposition Preference
To determine oviposition preference of N. amica, mated adult females were placed
in groups of three into 30 cm x 30 cm x 30 cm mesh cages. Three moths were
used to ensure that sufficient eggs were produced for each replicate. The moths
were offered similar sized (assessed by visual observation of total leaf area and
height), non-flowering branches of both plant species, as well as a sugar-water
source for feeding. Branches were approximately 20 cm long and all bore young
leaves. The branches were placed in water and positioned at opposite sides of the
cage with the sugar-water in the middle. After a 48 h period, the total number of
eggs on each plant species was recorded. The trial was replicated 16 times, with
fresh branches used for each replicate. Due to high variance in the total number of
eggs laid between replicates, data were analysed as the proportion of eggs laid
(per replicate) on each host plant. A significant preference for either plant species
was judged according to the highest proportion of eggs laid on a particular species,
as analysed by a t-test performed on arcsine-transformed data.
4.3.4 Foliage damage
Leaf damage was assessed in pure and mixed stands of the Senecio species.
Three pure stands of S. pinnatifolius, isolated from S. madagascariensis, were
located near Swanfels (28o 07’S, 152o 23’E and 28o 08’S, 152o 23’E, respectively)
and Hampton (27o 22’S, 152o10’E). Three pure stands of S. madagascariensis
were located near Springbrook National Park (28o11’S, 153o16’E), Mt Tamborine
(27o 58’S, 153o12’E) and Beechmont (28o 07’S, 153o10’E). Three mixed stands
were located just west of Queen Mary Falls (28o 20’S, 152o 21’E), near Killarney
(28o18’S, 152o 21’E) and on private land neighbouring the O’Reilly’s section of
Lamington National Park (28o13’S, 153o 07’E). Senecio pinnatifolius tends to grow
along the edge of open areas, bordering on forest, whilst S. madagascariensis
grows throughout the entire open area (including along the edge). There is

85
therefore considerable mixing of the two species at the interface (and some degree
of mixing throughout).
All sites occur within an approximately 120 km length of the “Border Ranges”, a
group of linked mountain ranges running along the eastern portion of the
Queensland/New South Wales state border. All sites, regardless of location,
occurred within a similar altitudinal range (between 550 m and 700 m ASL), had
similar types of neighbouring vegetation (pasture and moist eucalypt forest or
rainforest), and were surveyed between 25 March and 3 May 2003 when both
species were flowering.
At each site 30 haphazardly selected mature plants were selected to conduct
foliage damage assessments. In sites that contained both species, 30 plants of
each species were used. For ten random stems on each plant we determined the
proportion of damaged leaves per plant by examining the 20 newest leaves and
recording whether or not each leaf had signs of damage consistent with
lepidopteran larval feeding. In 18 months of regular field sampling, Nyctemera
amica was the only folivore regularly collected causing gross leaf damage to
Senecio in the study area (White 2007). Because N. amica populations are patchy
in time and space, accumulated leaf damage is a more consistent measure of
herbivore activity and, because other folivores are rare or absent (White 2007), leaf
damage can be attributed to N. amica with a high level of confidence. The
proportion of leaves damaged per plant was arcsine-transformed before analysis,
using a two-way ANOVA with factors being species and population type (mixed or
pure) and the replicates being site.

86
Simple linear regression analyses were used to determine whether a relationship
existed between plant height (data were log-transformed) and percent damage
(data were arcsine-transformed) for either species.
4.3.5 Plant characteristics
Plant density and height of S. pinnatifolius and S. madagascariensis were
determined by the Point Centred Quarter method (Krebs 1989), using 30 random
plants of each species at each site as ‘centre points’. Height of each of these
plants was also measured.
All statistical analyses were conducted in SPSS 12.0.1 with the exception of chi-
square analyses, which were performed using Microsoft Excel 2003. Data are
presented as mean + 1 standard error.
4.4 Results
4.4.1 Larval feeding and survival
Survival rates of larvae reared individually were greater than 83% and did not differ
significantly between Senecio species (χ2 = 0.13, df = 1, P =0.71). Similarly, mean
pupal weight did not differ between individuals reared on S. pinnatifolius (0.23 ±
0.01 g) and S. madagascariensis (0.22 ± 0.01 g) (F1,45 = 1.54, P = 0.22). However,
larval host plant did affect the time taken to reach pupation. Larvae on S.
pinnatifolius reached pupation on average three days sooner (X̄ = 19.05 + 0.15
days) than larvae on S. madagascariensis (X̄ = 22.10 + 0.20 days) (F1,45 = 58.71,
P<0.001).
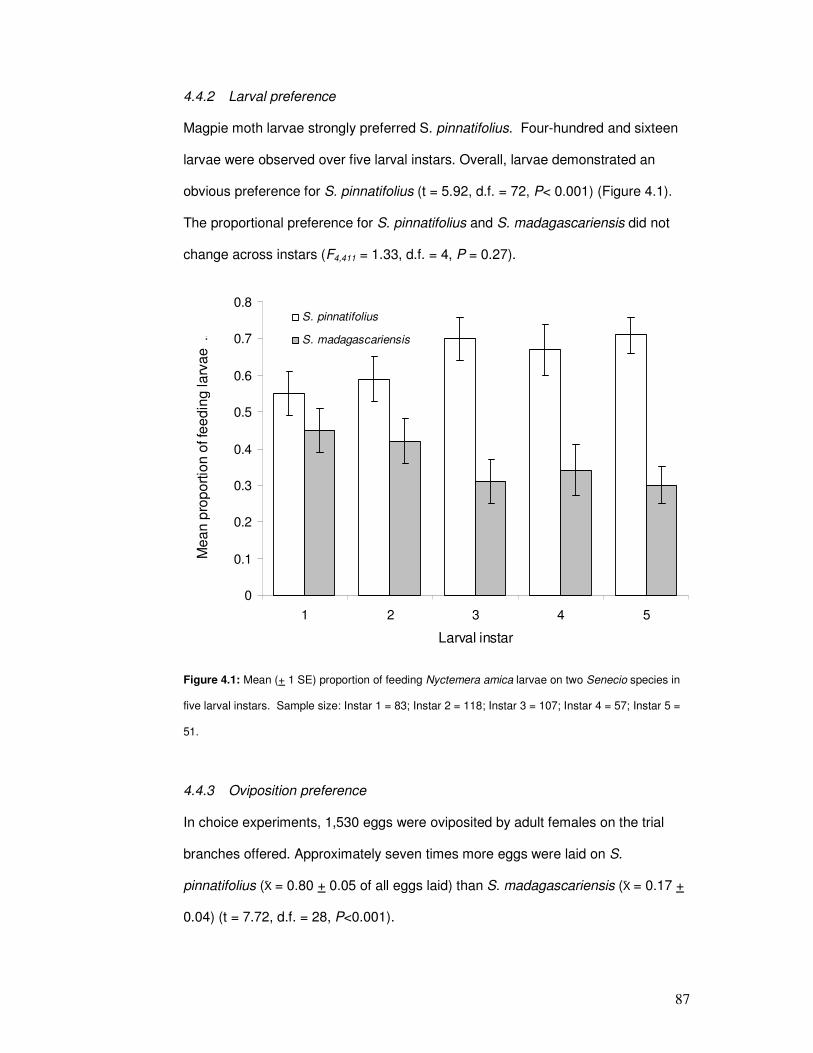
87
4.4.2 Larval preference
Magpie moth larvae strongly preferred S. pinnatifolius. Four-hundred and sixteen
larvae were observed over five larval instars. Overall, larvae demonstrated an
obvious preference for S. pinnatifolius (t = 5.92, d.f. = 72, P< 0.001) (Figure 4.1).
The proportional preference for S. pinnatifolius and S. madagascariensis did not
change across instars (F4,411 = 1.33, d.f. = 4, P = 0.27).
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
1 2 3 4 5
Larval instar
Me
an
pro
po
rtio
n o
f fe
ed
ing
la
rva
e ..
S. pinnatifolius
S. madagascariensis
Figure 4.1: Mean (+ 1 SE) proportion of feeding Nyctemera amica larvae on two Senecio species in
five larval instars. Sample size: Instar 1 = 83; Instar 2 = 118; Instar 3 = 107; Instar 4 = 57; Instar 5 =
51.
4.4.3 Oviposition preference
In choice experiments, 1,530 eggs were oviposited by adult females on the trial
branches offered. Approximately seven times more eggs were laid on S.
pinnatifolius (X̄ = 0.80 + 0.05 of all eggs laid) than S. madagascariensis (X̄ = 0.17 +
0.04) (t = 7.72, d.f. = 28, P<0.001).
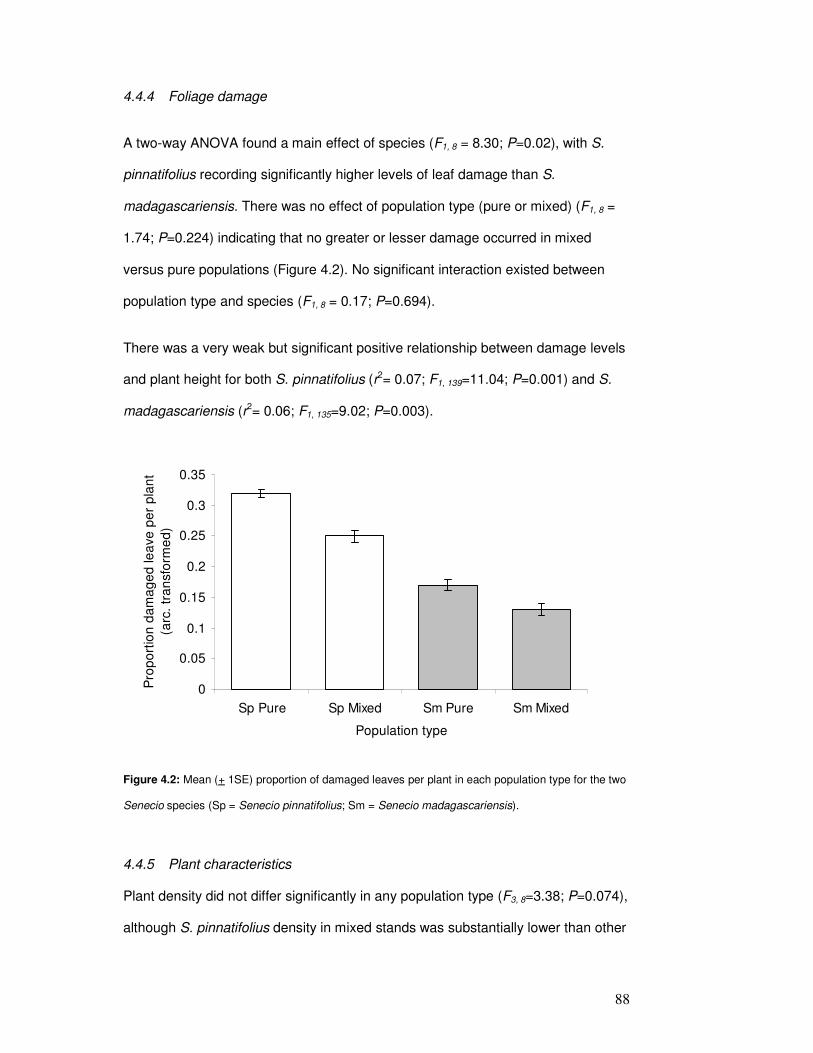
88
4.4.4 Foliage damage
A two-way ANOVA found a main effect of species (F1, 8 = 8.30; P=0.02), with S.
pinnatifolius recording significantly higher levels of leaf damage than S.
madagascariensis. There was no effect of population type (pure or mixed) (F1, 8 =
1.74; P=0.224) indicating that no greater or lesser damage occurred in mixed
versus pure populations (Figure 4.2). No significant interaction existed between
population type and species (F1, 8 = 0.17; P=0.694).
There was a very weak but significant positive relationship between damage levels
and plant height for both S. pinnatifolius (r2= 0.07; F1, 139=11.04; P=0.001) and S.
madagascariensis (r2= 0.06; F1, 135=9.02; P=0.003).
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
Sp Pure Sp Mixed Sm Pure Sm Mixed
Population type
Pro
po
rtio
n d
am
ag
ed
le
ave
pe
r p
lan
t
(arc
. tr
an
sfo
rme
d)
Figure 4.2: Mean (+ 1SE) proportion of damaged leaves per plant in each population type for the two
Senecio species (Sp = Senecio pinnatifolius; Sm = Senecio madagascariensis).
4.4.5 Plant characteristics
Plant density did not differ significantly in any population type (F3, 8=3.38; P=0.074),
although S. pinnatifolius density in mixed stands was substantially lower than other

89
sites and S. pinnatifolius plants were taller than S. madagascariensis plants in both
mixed and pure populations (F3, 8 = 48.53; P<0.001) (Table 4.1).
Table 4.1: Vegetation structure of native S. pinnatifolius and introduced S. madagascariensis
populations in pure and mixed stands (mean + se (n = 3)). Letters in superscript denote groups
(within columns) that are not significantly different from one another (P<0.05).
Stem
density/m2
Height (m)
S. pinnatifolius 0.46+0.20a 0.72+0.07 b Mixed stands
S. madagascariensis 0.11+0.03 a 0.35+0.04 c
S. pinnatifolius 0.73+0.20 a 1.08+0.04 a Pure stands
S. madagascariensis 0.65+0.09 a 0.44+0.03 c
4.5 Discussion
The ERH predicts that native herbivores will exhibit a preference for and/or cause
greater damage to native, compared to introduced plant species. Our results
support this prediction: in the laboratory, both ovipositing adult females and feeding
larvae of the native magpie moth, N. amica, preferred the native S. pinnatifolius
over the introduced S. madagascariensis. Field surveys reflected this preference,
with S. pinnatifolius experiencing significantly higher leaf damage levels associated
with N. amica larval feeding. These findings contrast with those of Parker and Hay
(2005) and Parker et al. (2006), who demonstrated that native plants are better
adapted than introduced plants at repelling generalist herbivores. The ERH is
probably more applicable for specialists like N. amica, than for generalist
herbivores (Parker et al. 2006).

90
Host preference by ovipositing moths can be influenced by plant characteristics
such as height (Nowicki et al. 2005) and stem density (Badenes-Perez et al. 2005).
However given that preference for S. pinnatifolius was evident in laboratory studies
which controlled for these factors, as well as in the field, it is likely that N. amica
preference is determined by other plant characteristics. Host preference is believed
to represent the suitability of hosts for larval survival (Singer 1983; Courtney et al.
1989). Our results suggest that the native Senecio is a more suitable host for
magpie moth larvae than the closely related introduced S. madagascariensis.
Although larvae exhibited similar survival rates and similar mean pupal weight
when reared on the two Senecio species, growth rates were slower, with pupation
being reached later by individuals reared on S. madagascariensis. Retarded
development times in Lepidoptera larvae can be associated with increased risk of
mortality due to parasitism (Benrey and Denno 1997), as well as reduced size
(Leather et al. 1998) and fecundity (Elkington and Liebhold 1990) in adults.
Slower growth rates of larvae reared on the introduced Senecio might be due to
the lower foliar nutrient concentrations of this species (Sims 2004), as low nutrient
levels (in particular N) have been linked to poor larval growth in other Lepidoptera
species (Rausher 1981). Ovipositing female Lepidoptera have also been shown to
demonstrate a preference for plants that are higher in nitrogen (Mattson 1980;
Chen et al. 2004), and plants that are higher in nitrogen may experience higher
rates of folivory by insects (Xiang and Chen 2004). These factors might explain the
female preference for S. pinnatifolius and the higher levels of folivory of this
species in the field. The profiles of the most common secondary chemicals found
in Senecio species, the pyrrolizidine alkaloids (Rothschild et al. 1979), also vary
between the two Senecio species (Sims 2004), and are known to play a role in
host plant selection and larval development (Lill and Marquis 2001).

91
Competitive interactions between introduced and native plant species have been
reported to be altered by the impacts of selective herbivory (Brown 1994;
Schierenbeck et al. 1994; Edwards et al. 2000; Scherber et al. 2003). For instance,
Scherber et al. (2003) investigated the effects of herbivory and competition on
growth, survival and reproduction of Senecio inaquidens, an introduced plant in
Europe, and concluded that populations of this invader gain a competitive
advantage over native species due to selective herbivory of the surrounding native
vegetation by vertebrates. Further research is necessary to determine whether
selective herbivory of S. pinnatifolius has an impact on its competitive interactions
with S. madagascariensis, as knowledge of a plant’s resource acquisition and
allocation is vital to explaining its response to herbivory (Chapin et al. 1987).
However Louda and Potvin (1995) predicted that it is species like S. pinnatifolius –
i.e. short-lived perennials with heavy dependence on current seed production for
regeneration – which will be most negatively affected at the population level by
damage caused by specialist herbivores. Although Senecio species are generally
not killed by defoliation (Obeso and Grubb 1994; Vrieling et al. 1996), damage to
foliage may result in reduced seed production (Crawley and Gillman 1989),
potentially having population-level impacts.
Herbivore populations might be expected to be enhanced in areas inhabited by the
attractive native S. pinnatifolius, resulting in increased herbivory on neighbouring
S. madagascariensis. Apparent competition by such means has been
demonstrated in other species (e.g. Rand 2003). However there was no evidence
that this was occurring in our system, with neither elevated herbivory in mixed
stands or interaction effects between species and stand type. Apparent competition
may never occur if there is very strong herbivore preference for S. pinnatifolius,
providing that abundant native foliage is available. Since we only sampled at one
time of year these results should be interpreted with caution. The situation may

92
differ at different times of year or in a situation in which herbivores are more
abundant (or plants more scarce). Controlled experiments are required to ascertain
under what conditions (if any) apparent competition may occur in this system.
4.5.1 Conclusions
This study provides evidence that specialist native herbivores may be better
adapted to utilise native plants than introduced plants even when an introduced
species is taxonomically and ecologically similar to a native. Herbivore preference
for native species could have implications not only for the control of the introduced
species, but also for competitive interactions between the introduced and native
plant species.

Chapter 5 - Plant-pollinator interactions in
sympatric exotic and native Senecio species:
Is facilitation or competition for pollinators
occurring?
The following chapter is currently in press as:
White, E.M. and Clarke, A.R. (in press) Plant-pollinator
interactions in sympatric exotic and native Senecio species: Is
facilitation or competition for pollinators occurring? Plant
Protection Quarterly.
The roles of co-authors are as follows:
E.M. White: Designed the experiment, conducted data analysis and wrote the
paper.
A.R. Clarke: Project supervisor, provided advice about experimental design and
data analysis and contributed to the writing of the manuscript.


95
Chapter 5 - Plant-pollinator interactions in sympatric exotic and native
Senecio species: Is facilitation or competition for pollinators occurring?
5.1 Abstract
The role of indirect interactions in invasion biology has rarely been addressed.
Indirect interactions between two plant species may be mediated by shared
pollinators: the presence of one plant species can have either a negative impact on
pollination (and seed set) in another by competing for pollinators, or a positive
effect by facilitating pollinator visitation. We investigated whether facilitation or
competition for pollination was occurring between the closely related native
Senecio pinnatifolius (A. Rich) and exotic S. madagascariensis (Poiret) in
Southeast Queensland. Visitation rates by honeybees and syrphid species, as well
as seed set in each Senecio species, were assessed in naturally occurring mixed
and pure stands. The exotic S. madagascariensis did not affect visitation rates to
the native, but seed set of the native species was higher in mixed populations. The
presence of native S. pinnatifolius caused a reduction in honeybee visits and an
increase in syrphid visits to the exotic plant, but altered visitation patterns were not
reflected in a change in seed set in the exotic.
5.2 Introduction
The colonisation of new areas by invasive species is a major conservation issue,
as in many cases it results in alterations to biodiversity and ecosystem function
(Maron and Vila 2001; Agrawal and Kotanen 2003; Scherber et al. 2003). Various
studies have investigated the impacts of invasive species on a system, generally
focussing on direct mechanisms such as predation (Dickman 1996; Wilson et al.
1998; Kinnear et al. 2002) and competition (Cadi and Joly 2003; Kolb and Alpert
2003; Corbin and D'Antonio 2004; Miller and Gorchov 2004), or system-level

96
impacts, which alter abiotic processes (Crooks 2002; Chornesky and Randall
2003). However, an exotic and native species can also affect one another via
indirect interactions, i.e. when changes to interactions between two species occur
as a result of the presence of a third (in this case invasive) species (Strauss 1991;
Wooton 1994; White et al. 2006). The impacts of such interactions might be
positive, negative or neutral, for either or both of the species involved.
Mutualistic interactions, including plant-pollinator relationships, can be important in
shaping natural systems and influencing the outcome of introductions (Richardson
et al. 2000; Bruno et al. 2003). In self-incompatible, animal-pollinated plant
species, plant-pollinator interactions can potentially be altered via indirect effects
caused by the addition to the system of a new, simultaneously flowering plant
species. Such indirect effects might occur via (i) competition - which includes both
(a) competition for pollinators (exploitation competition), and (b) improper pollen
transfer (interference competition) resulting in pollen interference or loss of
conspecific pollen; or (ii) facilitation of pollination (Rathke 1983).
Plant species competing for the services of shared pollinators (exploitation
competition) may or may not be closely related and may have similar or very
different floral structures (Levin 1970; Rathke 1983). Several studies have shown
decreased pollinator visitation rates to natives in the presence of more attractive
exotic species (Chittka and Schurkens 2001; Ghazoul 2004; Moragues and
Travaset 2005) (note though, that reduced visitation rates do not necessarily
translate into reduced seed set, Ghazoul 2004). Alternatively, flowers of an
invasive species may be less attractive to insect pollinators than flowers of native
species, potentially limiting the establishment or spread of the invader. To our
knowledge, no study has directly investigated the impact of the presence of a
native species on pollinator visitation rates or seed set in a sympatric invasive

97
species, although, in an analogous system, studies have shown that native species
can compete with crop species for pollinators (Free 1963; Holm 1966).
Richardson et al. (2000) suggest that pollen limitation is rarely a constraint on the
success of an invader because of the widespread distribution of generalist
pollinators, which visit exotic as well as native plant species. Even in the absence
of pollinator limitation however, the presence of one species can have a negative
impact on another through improper pollen transfer (interference competition). This
can result in reduced seed set either through pollen interference (when
heterospecific pollen on a stigma interferes with fertilisation of the ovules by
conspecific pollen) (Galen and Gregory 1989; Brown and Mitchell 2001), or
conspecific pollen loss (Campbell and Motten 1985; Bell et al. 2005).
Whilst negative impacts are the focus of the majority of studies, the presence of
one plant species may instead have a positive facilitative effect on another by
attracting greater numbers of pollinators to the area (Feldman et al. 2004; Moeller
2004). Facilitation is more likely to occur in plant populations of low density or of a
small size (Rathke 1983) and has been recorded between sympatric native species
(Campbell and Motten 1985; Moeller 2004, 2005). However, facilitation has rarely
been shown to occur between exotic and native species. One exception is a study
which demonstrated that the presence of an invader, Carpobrotus spp., had a
facilitative effect on pollination in two co-occurring native species, Cistus salviifolius
and Anthyllis cytisoides (Moragues and Travaset 2005).
The aim of this study is to determine whether facilitation of, or competition for, visits
by shared pollinators is occurring between two species of Senecio, the native S.
pinnatifolius and the invasive S. madagascariensis, in south-eastern Australia.
Previous studies indicate that both species are self-incompatible, rely on insects for

98
pollination, and share the same common floral visitors (Ali 1966; Radford 1997;
authors’ unpublished data). This creates the possibility for pollinator-mediated
indirect interactions which, if present, may have the potential to affect both the
invasion process and the impacts of the invader on the native species. This study
took place in the middle of the flowering season of the native S. pinnatifolius, which
coincides with the early stages of the S. madagascariensis flowering season. At
this time of year, S. pinnatifolius plants and flowers are likely to occur at a greater
density than S. madagascariensis plants and flowers, so we predict that the
presence of the dominant native S. pinnatifolius is more likely to impact the
invasive S. madagascariensis, than vice versa.
5.3 Materials and Methods
5.3.1 Study species
Senecio madagascariensis, fireweed, is an annual weed from South Africa that
was first recorded in Australia in 1918: it has since invaded large areas of farmland
and grassland in south-eastern Australia (Radford et al. 1995; Radford 1997). In
south-eastern Australia, S. madagascariensis flowers between the months of
March and December (Radford 1997).
Closely related to S. madagascariensis is a group of sub-species belonging to the
Australian native S. pinnatifolius complex. Senecio pinnatifolius is a herbaceous
perennial (Ali 1966) whose geographic range overlaps with that of S.
madagascariensis in Australia, but generally occurs in smaller, more scattered
populations than the exotic (Radford 1997; Radford and Cousens 2000). Senecio
pinnatifolius ssp. lanceolatus, the focus of this study, inhabits disturbed areas and
pasture usually close to the edge of rainforest or moist eucalypt forest and flowers
between January and June in south-eastern Australia (Radford 1997). There is a
four month period of overlap between the flowering periods of the exotic and native

99
Senecio. Previous studies have indicated that both species are self-incompatible
and are likely to rely on insects as pollinators (Ali 1966; Radford 1997). The two
species are morphologically similar; both produce similar-sized yellow capitula
which occur in clusters on the plant, and floral visitors move freely between the two
species when they grow together in the field (E. White, personal observation).
5.3.2 Study sites
This study was conducted using four ‘population types’, each represented by three
replicate populations in south-east Queensland:
(1) Three ‘pure S. pinnatifolius stands’: These were S. pinnatifolius populations
which were at least five km from the nearest known S. madagascariensis
populations. Two sites existed near Swanfels, (located at 28o 07’S, 152o 23’E and
28o 08’S, 152o 23’E respectively) and one was east of Hampton (27o 22’S,
152o10’E);
(2) Three ‘pure S. madagascariensis stands’: These comprised three populations
of S. madagascariensis which were at least five km from the nearest known S.
pinnatifolius populations. One was near Springbrook National Park (28o11’S,
153o16’E), a second at Mt Tamborine (27o 58’S, 153o12’E) and a third, just south of
Beechmont (28o 07’S, 153o10’E);
(3) ‘Mixed S. pinnatifolius stands’: Three populations of S. pinnatifolius existing in
close proximity (within 50 m) to S. madagascariensis populations. These included
one just west of Queen Mary Falls (28o 20’S, 152o 21’E), one near Killarney
(28o18’S, 152o 21’E) and one on private land neighbouring the O’Reilly’s section of
Lamington National Park (28o13’S, 153o 07’E).
(4) ‘Mixed S. madagascariensis stands: Three populations of S. madagascariensis
existing in close proximity (within less than 50m) to S. pinnatifolius populations.

100
These were in the same locations as those described for the mixed S. pinnatifolius
stands.
All sites occur within an approximately 120 km length of the “Border Ranges”, a
group of linked mountain ranges running along the eastern portion of the
Queensland/New South Wales state border. All sites, regardless of location,
occurred within a similar altitudinal range (between 550 m and 700 m ASL), had
similar types of neighbouring vegetation (pasture and moist eucalypt forest or
rainforest), and were surveyed between March and May when both species were
flowering.
5.3.3 Methods
Quantity of pollen on insects:
Honey bees (Apis mellifera) and hoverflies (syrphid species) are the two most
common floral visitors to both S. pinnatifolius and S. madagascariensis at study
sites in southeast Queensland (authors’ unpublished data) and so it seemed likely
that these species play an important role as pollinators. To confirm that these
species were not only visiting flowers, but also carrying pollen, the following
procedures were carried out.
Pollen grains of both S. pinnatifolius and S. madagascariensis were collected from
flowers growing in the field, mounted on stubs, gold coated, examined and
photographed under a scanning electron microscope at 800x magnification. Twelve
specimens of A. mellifera and 13 specimens of syrphid flies found visiting Senecio
flowers were collected from pure and mixed S. pinnatifolius and S.
madagascariensis populations in south east Queensland. Specimens were
mounted individually on stubs (ventral side facing up), gold-coated, and examined
under a scanning electron microscope at 400x magnification. Since it was difficult

101
to determine exact number of pollen grains on an insect (particularly when pollen
grains were extremely abundant and lying one on top of another) we recorded
simply whether an insect was carrying <10; 10-50; 50-100 or >100 Senecio pollen
grains. Body parts on which the pollen grains were found were also noted. Pollen
contained in pollen sacs was visible on bees but was not included in the count
because it was considered unlikely that the majority of these pollen grains would
be transferred between plants.
Pollinator visits
Thirty random plants per population were used for floral visitor observations and
observations were made on sunny days during which the temperature in the shade
ranged between 17 and 23oC and wind gusts did not exceed 15 km/hr. Two
observers monitored individual plants, recording the number of bees and hoverflies
visiting flowers on a plant during a five minute observation period, before moving to
another plant. Since there were often several insects at a plant at any one time it
was not possible to record how many flowers were visited by each insect.
Each observer conducted six, five-minute observations (as described above) per
hour, between the hours of 10am and 3pm (it is during this time period that
hoverflies and honeybees are most active on Senecio at this time of year, E. White,
unpublished data). This procedure was performed by the two observers for one
day per site for each of the six pure stands and two days per site for each of the
mixed populations, in which one observer worked on one plant species and the
second observer worked on neighbouring species simultaneously. Thus for each of
the four population-types (i.e. for each treatment), a total of 13-15 hours of
observations were conducted over a three-day period.

102
For each plant the following data were also recorded: height, number of open
capitula, distance to nearest neighbour of same species, and whether or not the
plant was in sun or shade during the time of observation. Plant density data were
also obtained for each population-type using the PCQ method (Krebs 1989), using
each of the 30 random plants per population-type as centre-points. Number of
open capitula per plant for the four nearest neighbours to each of the 30 random
plants was also recorded.
Seed-set
From each site, six or seven mature capitula (i.e. with shrivelled ray florets,
containing mature seeds which were just about to be released) were collected from
seven random plants, a total of 42-48 capitula per site. This was repeated for both
species in mixed populations. Collections were made approximately two weeks
after the pollinator observations were carried out. Seed set was determined by
counting number of developed seeds per capitula.
Statistical analyses
All analyses were performed in SPSS v. 12.0.1. When variances were unequal,
data were transformed by log10. Because data from individual sites were treated
as a replicate of population-type, the factor “site” was not included in any analysis.
Quantity of pollen on insects: Using categorical data (categories were <10; 10-50;
50-100; and >100 pollen grains) a chi-squared test for association was performed
to determine whether bees and hoverflies carried different amounts of pollen on
their bodies.
Effect of plant characteristics on visitation rate and time spent at plant: A range of
variables can influence the attractiveness of a plant to floral visitors. One-way

103
ANOVAs were used to determine whether population-types differed in regard to
density of plants, number of open capitula and plant height. In order to determine
whether it was necessary to standardise the data to take into account any of these
factors, multiple linear regression analyses (using the stepwise method) were
performed for each of the four population-types separately, and for each of the two
pollinator taxa within each population type. Dependent variables used were (i)
number of visits per plant per five minute observation period (henceforth referred to
as ‘visitation rate’) and (ii) time spent per insect per plant, and independent
variables were: number of open capitula per plant, plant height and distance to
nearest neighbour as independent variables.
Insect activity can also be influenced by micro-environmental variables, including
level of sun or shade (Verma and Rana 1994; Kirchner et al. 2005). In order to test
whether sun/shade was a factor that might explain differences in floral visitor
activity between population-types, Pearson chi-squared tests were used to
determine whether the number of observation periods conducted in a sunny
position differed between population types. Independent-samples t-tests were
performed for each population-type separately to establish whether bee/syrphid
visits were more likely to occur in the sun or shade.
Pollinator visits: One-way ANOVAs, followed by post-hoc Tukey tests, were used to
determine whether (i) bee and (ii) syrphid visits per plant per five minute
observation period varied between population types.
Seed set: A one-way ANOVA was used to determine whether differences existed
in number of seeds set per capitulum between population types.

104
5.4 Results
5.4.1 Quantity of pollen on insects
Pollen grains of the two Senecio species were extremely similar morphologically,
making it difficult to distinguish with any degree of certainty between the two
species. However, both hoverflies and honeybees collected from mixed stands, as
well as pure stands of each Senecio species, carried Senecio pollen on all body
parts including legs, abdomen, thorax, head and mouthparts. Both of these insect
taxa are therefore likely to act as pollinators for both Senecio species. Bees carried
greater quantities of pollen than did hoverflies (df=2; χ2=18.32; p<0.01).
5.4.2 Vegetation structure
Plant density was slightly lower in mixed S. madagascariensis stands than in other
population-types (Table 5.1), but this difference was not significant (df=3; f=3.98;
p=0.05). Number of open capitula per plant and plant height did vary, however,
between population types (df=3; f=205.76; p<0.01 and df=3; f=398.54; p<0.01
respectively), with plants in the S. madagascariensis population-types being
smaller and having fewer open capitula than did S. pinnatifolius plants (Table 5.1).
Table 5.1: Vegetation structure of native S. pinnatifolius and exotic S. madagascariensis populations
in pure and mixed stands (mean + se (n)). Letters in superscript denote groups (within columns) that
are not significantly different from one another (P<0.05).
Stem density/m2 Open capitula/plant Height (cm)
S. pinnatifolius 0.20+0.05 (80)a 19.31+1.69 (317)
a 72.28+2.27 (107)
b Mixed
stands S. madagascariensis 0.03+0.01 (80)
a 3.77+0.26 (307)
b 33.53+0.93 (118)
c
S. pinnatifolius 0.53+0.13 (90) a 25.65+3.11 (360)
a 107.54+2.01 (120)
a Pure
stands S. madagascariensis 0.39+0.10 (90)
a 7.57+0.44 (360)
b 42.65+1.18 (75)
c
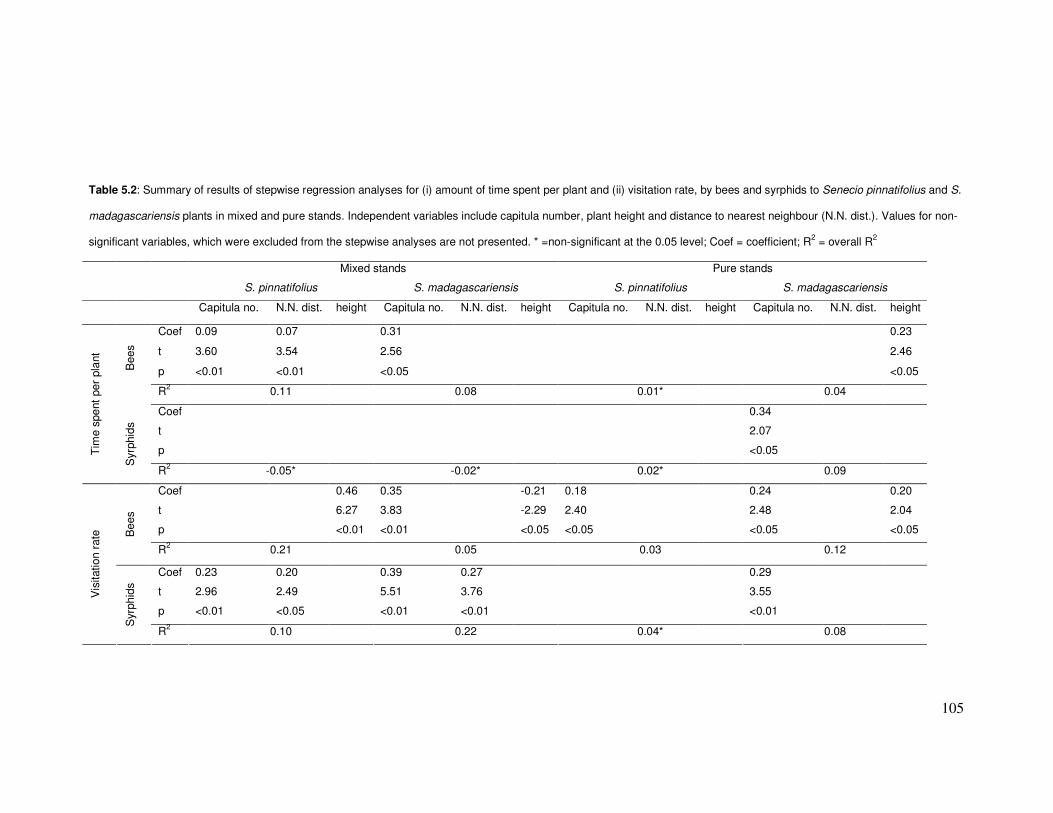
105
Table 5.2: Summary of results of stepwise regression analyses for (i) amount of time spent per plant and (ii) visitation rate, by bees and syrphids to Senecio pinnatifolius and S.
madagascariensis plants in mixed and pure stands. Independent variables include capitula number, plant height and distance to nearest neighbour (N.N. dist.). Values for non-
significant variables, which were excluded from the stepwise analyses are not presented. * =non-significant at the 0.05 level; Coef = coefficient; R2 = overall R
2
Mixed stands Pure stands
S. pinnatifolius S. madagascariensis S. pinnatifolius S. madagascariensis
Capitula no. N.N. dist. height Capitula no. N.N. dist. height Capitula no. N.N. dist. height Capitula no. N.N. dist. height
Coef 0.09 0.07 0.31 0.23
t 3.60 3.54 2.56 2.46
Bees
p <0.01 <0.01 <0.05 <0.05
R2 0.11 0.08 0.01* 0.04
Coef 0.34
t 2.07
p <0.05 Tim
e s
pe
nt
per
pla
nt
Syrp
hid
s
R2 -0.05* -0.02* 0.02* 0.09
Coef 0.46 0.35 -0.21 0.18 0.24 0.20
t 6.27 3.83 -2.29 2.40 2.48 2.04
p <0.01 <0.01 <0.05 <0.05 <0.05 <0.05 Bees
R2 0.21 0.05 0.03 0.12
Coef 0.23 0.20 0.39 0.27 0.29
t 2.96 2.49 5.51 3.76 3.55
p <0.01 <0.05 <0.01 <0.01 <0.01
Vis
itation r
ate
Syrp
hid
s
R2 0.10 0.22 0.04* 0.08

106
5.4.3 Visitation rates and plant characteristics
Significant linear relationships existed between visitation rate and capitula number
and/or plant height for both bees and syrphids. These relationships were weak
and highly variable among population-types, however: overall R2 values were low,
ranging from 0.03 to 0.22, indicating that these variables accounted for only a small
amount of the observed variation (Table 5.2). No significant relationship was
detected between visitation rate and distance to nearest neighbour (for bees) or
visitation rate and plant height (for syrphids) in any population-type. Relationships
between time spent at a plant by bee and syrphid visitors and number of open
capitula, plant height and distance to nearest neighbour were also extremely weak
or non-existent and highly variable among population-types (Table 5.2). Since we
detected only weak and highly variable relationships between the measured
individual plant characteristics and pollinator visitation rate and time spent at plant,
per-plant visitation-rates data were not standardised to take into account any of
these variables in subsequent analyses of results.
5.4.4 Pollinator visits
Bee visitation rate varied between population-types (df=3; f=14.43; p<0.01) (Figure
5.1). A post hoc Tukey test identified that S. pinnatifolius plants experienced a
similar visitation rate regardless of whether or not S. madagascariensis grew
nearby. Therefore, there is no evidence for either a facilitative or competitive effect
of the exotic species on pollinator visits to the native in mixed populations. The
visitation rate to pure S. madagascariensis stands did not differ significantly from
the visitation rate to S. pinnatifolius in mixed stands. However, mixed S.
madagascariensis stands recorded a significantly lower bee visitation rate than did
pure S. madagascariensis stands and recorded a lower visitation rate than that
recorded for S. pinnatifolius plants in either mixed or pure stands. This indicates a
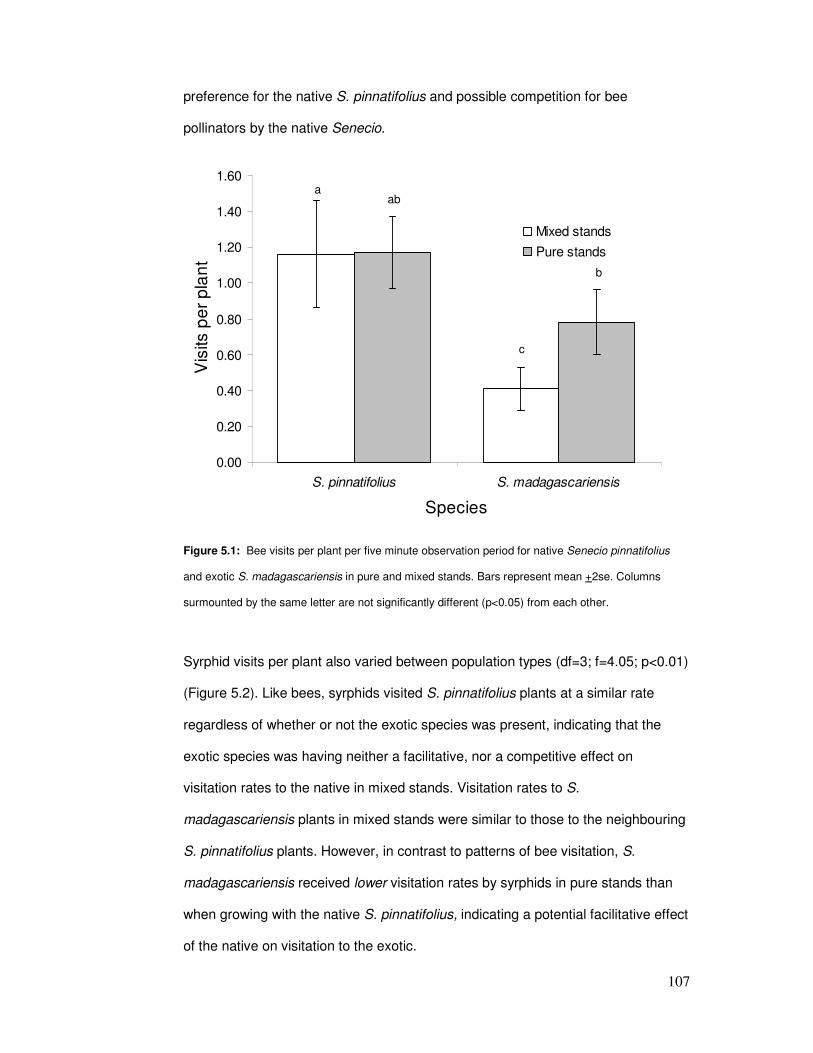
107
preference for the native S. pinnatifolius and possible competition for bee
pollinators by the native Senecio.
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
1.60
S. pinnatifolius S. madagascariensis
Species
Vis
its p
er
pla
nt
Mixed stands
Pure stands
a ab
b
c
Figure 5.1: Bee visits per plant per five minute observation period for native Senecio pinnatifolius
and exotic S. madagascariensis in pure and mixed stands. Bars represent mean +2se. Columns
surmounted by the same letter are not significantly different (p<0.05) from each other.
Syrphid visits per plant also varied between population types (df=3; f=4.05; p<0.01)
(Figure 5.2). Like bees, syrphids visited S. pinnatifolius plants at a similar rate
regardless of whether or not the exotic species was present, indicating that the
exotic species was having neither a facilitative, nor a competitive effect on
visitation rates to the native in mixed stands. Visitation rates to S.
madagascariensis plants in mixed stands were similar to those to the neighbouring
S. pinnatifolius plants. However, in contrast to patterns of bee visitation, S.
madagascariensis received lower visitation rates by syrphids in pure stands than
when growing with the native S. pinnatifolius, indicating a potential facilitative effect
of the native on visitation to the exotic.
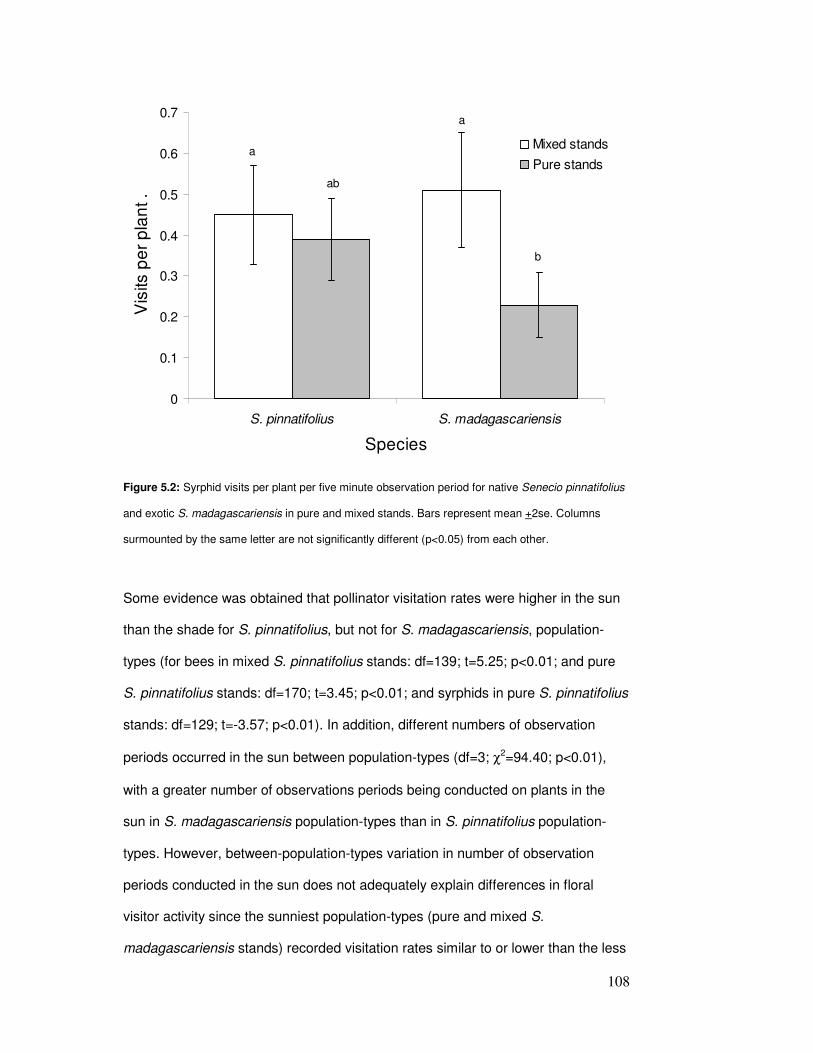
108
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
S. pinnatifolius S. madagascariensis
Species
Vis
its p
er
pla
nt .
Mixed stands
Pure stands
a
a
ab
b
Figure 5.2: Syrphid visits per plant per five minute observation period for native Senecio pinnatifolius
and exotic S. madagascariensis in pure and mixed stands. Bars represent mean +2se. Columns
surmounted by the same letter are not significantly different (p<0.05) from each other.
Some evidence was obtained that pollinator visitation rates were higher in the sun
than the shade for S. pinnatifolius, but not for S. madagascariensis, population-
types (for bees in mixed S. pinnatifolius stands: df=139; t=5.25; p<0.01; and pure
S. pinnatifolius stands: df=170; t=3.45; p<0.01; and syrphids in pure S. pinnatifolius
stands: df=129; t=-3.57; p<0.01). In addition, different numbers of observation
periods occurred in the sun between population-types (df=3; χ2=94.40; p<0.01),
with a greater number of observations periods being conducted on plants in the
sun in S. madagascariensis population-types than in S. pinnatifolius population-
types. However, between-population-types variation in number of observation
periods conducted in the sun does not adequately explain differences in floral
visitor activity since the sunniest population-types (pure and mixed S.
madagascariensis stands) recorded visitation rates similar to or lower than the less
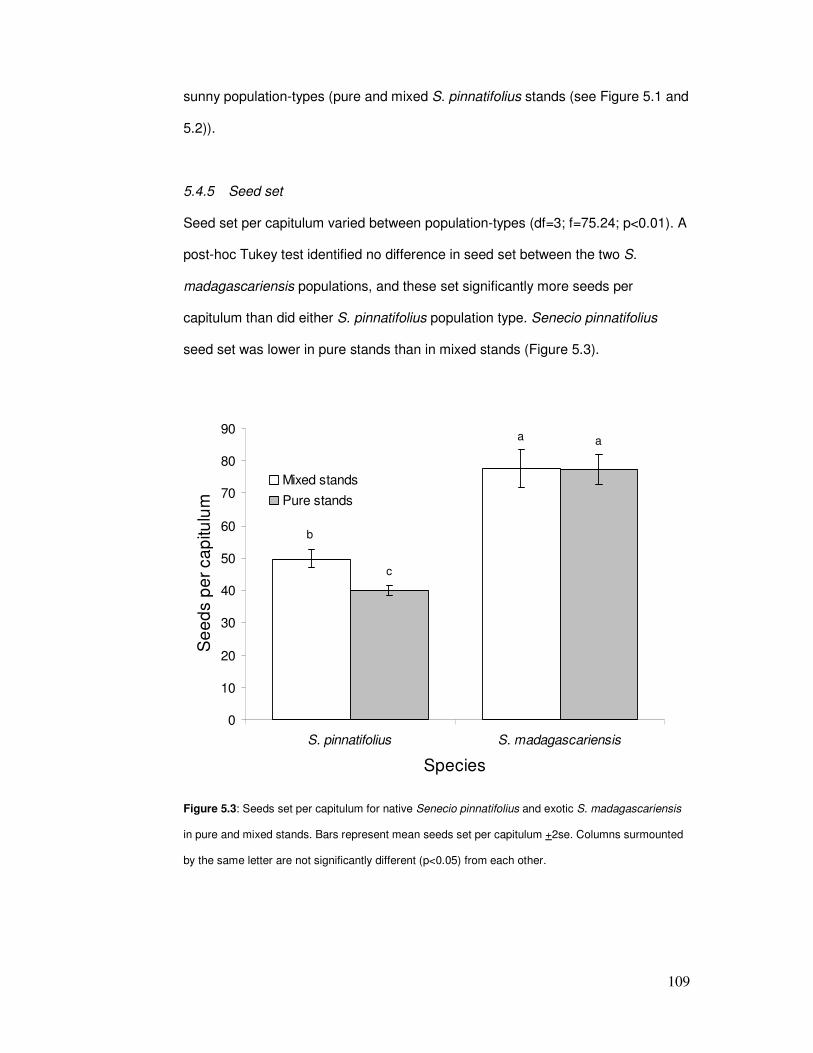
109
sunny population-types (pure and mixed S. pinnatifolius stands (see Figure 5.1 and
5.2)).
5.4.5 Seed set
Seed set per capitulum varied between population-types (df=3; f=75.24; p<0.01). A
post-hoc Tukey test identified no difference in seed set between the two S.
madagascariensis populations, and these set significantly more seeds per
capitulum than did either S. pinnatifolius population type. Senecio pinnatifolius
seed set was lower in pure stands than in mixed stands (Figure 5.3).
0
10
20
30
40
50
60
70
80
90
S. pinnatifolius S. madagascariensis
Species
Se
ed
s p
er
ca
pitu
lum
Mixed stands
Pure stands
a a
b
c
Figure 5.3: Seeds set per capitulum for native Senecio pinnatifolius and exotic S. madagascariensis
in pure and mixed stands. Bars represent mean seeds set per capitulum +2se. Columns surmounted
by the same letter are not significantly different (p<0.05) from each other.

110
5.5 Discussion
We found no evidence therefore, that exotic S. madagascariensis was either
competing for, or facilitating, pollinator visits to the native S. pinnatifolius at this
stage in the flowering season. Native plants received similar numbers of bee and
hoverfly visits regardless of whether or not the exotic species was present in the
area. This most probably is due to the relatively low densities of S.
madagascariensis flowers (compared with S. pinnatifolius flower density) during
the study period.
Surprisingly, seed set in S. pinnatifolius was higher in mixed populations than when
growing in isolation from the alien species. This is clearly not due to facilitation of
pollinator visits by the presence of the alien species, but there are a number of
possible explanations: (1) This may be due to abiotic factors not measured in this
study, (2) Higher levels of herbivory have been recorded in pure populations of S.
pinnatifolius (White et al. in press), which could have consequences for
reproductive success in these plants, (3) seed set may be enhanced indirectly in
mixed populations via hybridisation. Senecio pinnatifolius and S. madagascariensis
are known to hybridise (Radford 1997; Prentis et al. in press). If one species has
greater male fitness (i.e. higher pollen germination rates) than another with which it
is capable of hybridising, seed set can be increased in the latter species when it
receives pollen from the former (Anttila et al. 1998). If S. madagascariensis has
higher male fitness than the native Senecio, seed set could be increased in mixed
populations via this mechanism. Molecular studies show that S. madagascariensis
does in fact, have a hybridisation advantage, siring significantly more progeny to S.
pinnatifolius maternal parents than expected based on proportional representation
of the two species in sympatric populations (Prentis et al. in press). Further genetic
work is currently underway to investigate this in greater detail.

111
Bees are likely to play a more important role than syrphids in pollination of Senecio
species, since they carry significantly greater quantities of pollen and visited
capitula more frequently than did syrphids. The presence of the native S.
pinnatifolius affected pollinator visitation rates to the alien Senecio, having opposite
effects for bees and syrphids: bee visits to S. madagascariensis were significantly
reduced by the presence of S. pinnatifolius, whilst syrphid visits increased. This
may be due simply to differential responses of these two insect taxa to the
presence of the native Senecio, with syrphid visits to S. madagascariensis being
facilitated, whilst competition is occurring for bee visits. It is not uncommon for
response to ecological variables to vary between pollinator species (Mitchell et al.
2004). However, interference competition between syrphids and honeybees might
also be partially responsible for differences in visitation rates. Gross (2001) noted
that Australian native bees were less likely to land on flowers of the shrub Dillwynia
juniperina when honeybees were present. If, like native bees, syrphid activity is
reduced by the presence of honeybees, syrphid visits to S. madagascariensis may
increase in response to lower bee numbers when bees are attracted away from the
alien to the more abundant flowers of the native species. If this were occurring, one
would expect that syrphid visits to S. pinnatifolius plants would be reduced, relative
to those to neighbouring S. madagascariensis plants. This was not the case here
however, so this interaction probably does not fully explain differential syrphid
visitation rates between mixed and pure S. madagascariensis stands.
Changes in pollinator behaviour, such as we have recorded, can have important
consequences for plant reproduction and flowering patterns (Rathke 1983). In this
case, however, altered visitation rates did not affect seed set in the exotic Senecio,
indicating that either the facilitative and competitive effects cancelled each other
out, or simply that S. madagascariensis is not pollen limited at these sites or at this
point in its flowering period. Ghazoul (2004) also reported that although butterfly

112
pollinator activity on the canopy tree species Dipterocarpus obtusifolius was
significantly reduced in disturbed areas, this did not translate into a seed-set effect.
He suggested that visits by other pollinator species probably compensated for
reduced butterfly pollination. Given the generalist nature of plant-pollinator
interactions and the widespread integration of exotic plants into the native plant-
pollinator visitation web (Memmott and Waser 2002), it may be commonplace for
reduced activity by one or two major pollinator taxa to be compensated for by visits
from other generalist pollinators.
Depending on the point in the flowering season for the two Senecio species (e.g. at
the end of the S. pinnatifolius flowering season, when exotic S. madagascariensis
flowers are dominant), different scenarios might be observed than those which we
report. Species such as the exotic and native Senecio, which have staggered
flowering times, may indirectly act as mutualists by jointly maintaining pollinator
populations at high levels over a longer time span than would otherwise be the
case (Waser and Real 1979). Alternatively, competitive interactions may be altered
or reversed at different points in the flowering season, as pollinator preferences
change in response to altered relative abundance of two or more plant species
(Kephart 1983).
5.5.1 Conclusions
The presence of the exotic S. madagascariensis had no effect on pollinator activity
in the native S. pinnatifolius at the stage in the flowering season during which this
study was conducted. This is not surprising considering the relatively low density of
exotic flowers at this time. However, seed set in the native species was higher in
mixed populations. Hybridisation, if it is occurring, might have an impact on seed
set and this issue warrants further investigation. In contrast, the presence of the
native S. pinnatifolius did affect pollinator visitation rates to the exotic species, with

113
bee visits being less frequent, and syrphid visits being more frequent (perhaps as a
result of reduced interference competition with bees), though this did not result in
alterations to seed set. In addition to commonly studied interactions such as
competition, potential indirect interactions between invasive and native plant
species should be taken into account when considering both management
approaches for invasive plants and conservation strategies for native plant species.


Chapter 6 - Can hybridisation cause local
extinction: the case for demographic
swamping of the Australian native,
Senecio pinnatifolius, by the invasive S.
madagascariensis?
The following chapter was recently published as:
Prentis, P.J., White, E.M., Radford, I.J., Lowe, A.J. and Clarke,
A.R. (2007) Can hybridization cause local extinction: the case for
demographic swamping of the Australian native, Senecio
pinnatifolius, by the invasive S. madagascariensis? New
Phytologist 174: 902-912.
Roles of co-authors are as follows:
P.J. Prentis: Conducted molecular laboratory work, part of the data analysis, and
wrote final draft.
E.M. White: Responsible for conceptual basis of project, experimental design, part
of the data analysis and writing of initial drafts.
I.J. Radford: Conducted the reciprocal crossing experiment, contributed data
comprising Appendices C and D, and made comments on manuscript.
A.J. Lowe: Assisted with data analysis and writing.
A.R. Clarke: Project supervisor, assisted with experimental design and writing.


117
Chapter 6 – Can hybridisation cause local extinction: the case for
demographic swamping of the Australian native, Senecio pinnatifolius, by
the invasive S. madagascariensis?
6.1 Abstract
Hybridisation between native and invasive species can have several outcomes,
including; enhanced weediness in hybrid progeny, evolution of new hybrid lineages
and decline of hybridising species. The latter largely depends on the relative
frequencies of parental taxa and viability of hybrid progeny. We investigated
individual and population level consequences of hybridisation between the
Australian native, Senecio pinnatifolius, and the exotic S. madagascariensis, with
AFLP markers and used this information to estimate the annual loss of viable
seeds to hybridisation. A high frequency (range 8.3-75.6 %) of hybrids was
detected in open pollinated seeds of both species, but mature hybrids were absent
from sympatric populations. A hybridisation advantage was observed for S.
madagascariensis, where significantly more progeny than expected were sired
based on proportional representation of the two species in sympatric populations.
Calculations indicated S. pinnatifolius would produce less viable seed than S.
madagascariensis, if hybridisation was frequency dependent and S.
madagascariensis reached a frequency between 10-60 %. For this native-exotic
species pair, prezygotic isolating barriers are weak, but low hybrid viability
maintains a strong postzygotic barrier to introgression. Due to asymmetric
hybridisation, S. pinnatifolius appears under threat if S. madagascariensis
increases numerically in areas of contact.

118
6.2 Introduction
The importance of hybridisation in the evolution and speciation of plants has long
been recognised (Rieseberg et al. 1995; Arnold, 1997; Rieseberg et al. 2003;
Abbott and Lowe 2004; Hegarty and Hiscock 2005; Buggs and Pannell 2006).
Hybridisation can result when divergent lineages, or species formed in allopatry,
change ranges and come into reproductive contact, potentially forming a zone of
secondary contact (Anderson 1949; Lagercrantz and Ryman 1990; Cruzan 2005;
Hoskin et al. 2005). The formation of hybrid zones can be promoted by biological
invasions, if introduced species are sufficiently closely related to native species.
As global trade and passenger travel continues to accelerate (Hanfling and
Kollmann 2002), it seems probable that alien plant invasions will continue at an
alarming rate, leading to increasing contact and hybridisation between previously
allopatric species (Abbott 1992; Abbott and Lowe 2004). In contrast to natural
range changes, biological invasions are more likely to form extensive zones of
contact, potentially accelerating the eventual outcome of hybridisation (Wolf et al.
2001).
Hybridisation between natives and exotics can have several outcomes, including;
enhanced weediness in hybrid offspring (Ellstrand and Schierenbeck 2000; Morrell
et al. 2005; Whitney et al. 2006), evolution of new hybrid lineages (Lowe and
Abbott 2004) and decline or even extinction of hybridising species (Levin et al.
1996). The latter is the most potentially destructive outcome of interspecific
hybridisation, and can occur via two main potential mechanisms (Wolf et al. 2001).
First, introgressive hybridisation, the transfer of genes between species via fertile
or semi-fertile hybrids, may produce hybrid derivatives of superior fitness that
displace one or both pure conspecifics, defined as genetic assimilation (Wolf et al.
2001). Secondly, if hybrids are sterile or display reduced fitness, the population
growth rate of the hybridising taxa may decrease below that required for

119
replacement of one or both parental species, termed demographic swamping (Wolf
et al. 2001).
The potential for introgression is regulated in part by the strength of chromosomal
or genic sterility barriers that prevent the formation of fertile interspecific offspring
(Arnold 1997; Lowe and Abbott 2004; Erickson and Fenster 2006). This can be
particularly true for triploid hybrids resulting from crosses between diploid and
tetraploid species (Lowe and Abbott 2000; Husband 2004). A combination of both
genetic assimilation and demographic swamping may also result in the decline of
hybridising taxa, making it difficult to discern the true causative process. In many
cases molecular methods can be applied to demonstrate the potential for
introgression and distinguish between processes.
Senecio, one of the largest genera of flowering plants, is known worldwide for its
globally important weed species (Holm et al. 1997) and the widespread occurrence
of interspecific hybridisation between native and introduced taxa (Abbott 1992;
Lowe and Abbott 2004; Kadereit et al. 2006). Senecio madagascariensis
(fireweed), a native of southern Africa and Madagascar, was introduced to
Australia more than 80 years ago and is now an aggressive weed in its invasive
range (Radford 1997; Radford et al. 1995a; Sindel et al. 1998). In Australia,
molecular genetic and morphological studies have demonstrated a close affinity
between fireweed and Australian native Senecio species, including S. pinnatifolius
(formerly S. lautus) (Scott et al. 1998). Although S. madagascariensis (2n = 2x =
20) and S. pinnatifolius (2n = 4x = 40) differ in ploidy (Radford et al. 1995b),
empirical and experimental crossing studies have established that both species
can serve as paternal and maternal parents of synthetic hybrids (Radford 1997).
Under greenhouse conditions, synthetic triploid hybrids between the two species
exhibit low viability and are highly sterile (sterile pollen, low pollen production, no

120
stigmatic viability, Radford 1997). Despite low fertility, triploid hybrids can still act
as a genetic bridge between diploid and tetraploid taxa, as demonstrated by Lowe
and Abbott (2000). Thus despite low fertility, F1 triploid hybrids could enable
introgression of S. madagascariensis genes into S. pinnatifolius (or vice-versa), but
this remains untested in the field.
Populations of S. madagascariensis exist in sympatry with populations of the native
S. pinnatifolius across many regions of Australia’s east coast. Within this area, S.
madagascariensis and S. pinnatifolius grow in close physical proximity, have
flowering periods that overlap and are pollinated by the same insect species
(Radford 1997; Radford and Cousens 2000; White 2007). Hybrid formation has
also been observed in sympatric populations of the two species in the field
(Radford 1997; Scott 1994), but may be restricted to certain variants of S.
pinnatifolius, such as the varieties tableland, headland and dune (Radford 1997).
To examine in greater detail the outcome of hybridisation between the native S.
pinnatifolius and the invasive S. madagascariensis, comparisons were made at
population (in sympatric vs allopatric populations) and individual (in sympatric
populations) levels to investigate contemporary and long-term outcomes of
hybridisation. Amplified fragment length polymorphisms (AFLP) were used as
molecular markers in this analysis. Three primary questions are addressed in this
paper. (i) What is the viability of hybrids in the field? - Comprising a comparison of
the frequency of hybrids in open pollinated seed of both species and incidence of
adult stage hybrids in sympatric populations. (ii) Does hybridisation influence the
level of genetic diversity or differentiation within sympatric compared to allopatric
populations of these hybridising species? (iii) Can we estimate the likely outcome
of hybridisation between this native-invasive species pair under a number of

121
hybridisation scenarios and is S. pinnatifolius at risk of genetic assimilation and/or
demographic swamping in sympatric populations?
6.3 Materials and Methods
6.3.1 Study species
Senecio madagascariensis (fireweed), a diploid annual weed from South Africa,
has invaded large areas of farmland and grassland in south-eastern Australia
(Radford et al. 1995a; Radford 1997). Senecio pinnatifolius is an herbaceous
perennial tetraploid (Ornduff 1964; Ali 1966; Radford et al. 1995b, 2004), and
exhibits a similar geographic range to S. madagascariensis in the eastern states,
but generally occurs in smaller, more scattered populations than the exotic
(Radford 1997; Radford and Cousens 2000). Senecio pinnatifolius (var. tableland,
formerly known as Senecio lautus ssp. lanceolatus), the focus of this study,
inhabits disturbed areas and pasture usually close to the edge of rainforest or
moist eucalypt forest and flowers between February and June in south-eastern
Australia (Radford and Cousens 2000, Appendix C). There is a four month period
of overlap between the flowering periods of the native and exotic Senecio, the
latter flowers between the months of March and December in Australia (Radford
and Cousens 2000). Previous studies have indicated that both species are self-
incompatible and insect-pollinated (Ali 1966; Lawrence 1985; Radford 1997). The
two species are superficially morphologically similar (differing in plant size, bract
number and time to senescence), both producing similar-sized yellow capitula
which occur in clusters on the plant: floral visitors move freely between the two
species when they grow together in the field (White 2007).
6.3.2 Study sites and sample collections
To assess genetic diversity in allopatric populations of each Senecio species, leaf
material was collected from approximately 45 (minimum 42) flowering plants from

122
each of three allopatric populations of S. pinnatifolius (var. tableland) and three
allopatric populations of S. madagascariensis.
To determine the number of mature hybrid plants, as well as genetic diversity for
each species when they grow in sympatry, leaf material was collected from
approximately 45 plants (minimum 43, maximum 47) from two sympatric sites
across the morphological range of flowering plants of each species. Plants from
which leaf material was collected were identified as either S. pinnatifolius or S.
madagascariensis using morphological features, including bract number and leaf
morphology, following Ali (1969) and Nelson (1980). Despite repeated searches
over two consecutive flowering seasons, no obvious hybrids (ie. plants with
intermediate morphology) were observed in the field. In addition, ~ 20 seeds per
plant were collected from a random selection of 10 plants of each species from
which leaf material had been collected (a total of ~ 200 seeds per species for each
of the two populations).
All allopatric and sympatric populations sampled occurred within the “Border
Ranges”, a group of linked mountain ranges running along the eastern portion of
the Queensland/New South Wales State border (population locations are indicated
in Table 6.1). All sites, regardless of location, occurred within a similar altitudinal
range (between 550 m and 700 m ASL), had similar types of neighbouring
vegetation (pasture and moist eucalypt forest or rainforest), and were surveyed
during May when both species were flowering. Allopatric populations were
separated by at least five km from the nearest known population of the other
species. In sympatric populations, S. pinnatifolius grew along the rainforest edges,
and in nearby creek beds, while S. madagascariensis inhabited adjacent pasture,
with considerable mixing of the species at the interface.

123
All leaf samples from allopatric and sympatric populations were transported on ice,
then frozen and stored at –80 oC until DNA extractions were performed. Seeds
were germinated on moist filter paper until they reached approximately 20 mm in
height, at which point they were removed, frozen and stored at –80 oC.
Germination percentages for S. pinnatifolius and S. madagascariensis from both
sympatric sites were generally quite low; particularly for S. madagascariensis (< 35
% at both sites), and numbers of resulting progeny for each species for each site
are shown in Table 6.1. Low seed germination was not the result of seed
dormancy, as neither species exhibits dormancy when grown on filter paper
(Radford 1997), but rather due to the collection relatively immature fruiting capitula.
6.3.3 Reciprocal crossing experiments
A reciprocal crossing experiment was undertaken to examine the viability and
number of seed produced from interspecific and intraspecific crosses. Plants were
germinated and grown using the methodology described in Radford and Cousens
(2000). Once plants reached reproductive maturity, inflorescences to be used in
the reciprocal crossing experiment were bagged prior to flowers opening. Once
flowers opened, bags were removed and crosses performed. Hand pollinations
were performed by applying mature anthers from pollen donors to the stigmatic
surface of pollen receivers with forceps. This procedure was repeated for all florets
on an inflorescence. Inflorescences were rebagged until maturation of capitula as
indicated by the exposure of mature pappus. Bags were then removed, and the
number of seed produced for both interspecific and intraspecific crosses was
recorded. To assess the viability of seed produced from crosses, seed were
germinated according to the protocol of Radford and Cousens (2000). ANOVA was
used to determine if differences existed in the number and viability of seed
produced from interspecific and intraspecific crosses.
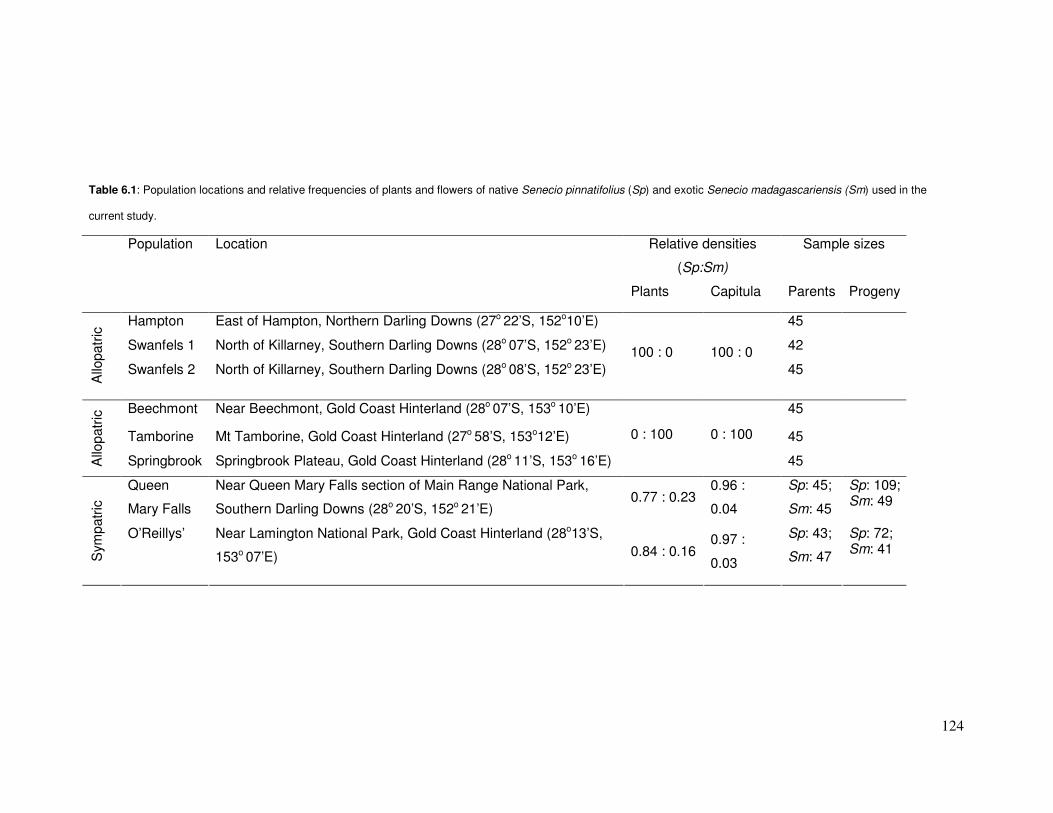
124
Table 6.1: Population locations and relative frequencies of plants and flowers of native Senecio pinnatifolius (Sp) and exotic Senecio madagascariensis (Sm) used in the
current study.
Population Location Relative densities
(Sp:Sm)
Sample sizes
Plants Capitula Parents Progeny
Hampton East of Hampton, Northern Darling Downs (27o 22’S, 152o10’E) 45
Swanfels 1 North of Killarney, Southern Darling Downs (28o 07’S, 152o 23’E) 42
Allo
patr
ic
Swanfels 2 North of Killarney, Southern Darling Downs (28o 08’S, 152o 23’E)
100 : 0 100 : 0
45
Beechmont Near Beechmont, Gold Coast Hinterland (28o 07’S, 153o 10’E) 45
Tamborine Mt Tamborine, Gold Coast Hinterland (27o 58’S, 153o12’E) 45
Allo
patr
ic
Springbrook Springbrook Plateau, Gold Coast Hinterland (28o 11’S, 153o 16’E)
0 : 100 0 : 100
45
Queen
Mary Falls
Near Queen Mary Falls section of Main Range National Park,
Southern Darling Downs (28o 20’S, 152o 21’E) 0.77 : 0.23
0.96 :
0.04
Sp: 45;
Sm: 45
Sp: 109; Sm: 49
Sym
patr
ic
O’Reillys’ Near Lamington National Park, Gold Coast Hinterland (28o13’S,
153o 07’E) 0.84 : 0.16 0.97 :
0.03
Sp: 43;
Sm: 47
Sp: 72; Sm: 41

125
6.3.4 Relative densities of plants and flowers
Relative plant and capitulum densities of each species at each site were
determined using the Point Centred Quarter (PCQ) method (Krebs 1989), using 30
random plants of each species at each site as ‘centre points’. A Chi-square test
was used to determine whether the proportion of hybrids produced in the progeny
of each species was concordant with capitulum densities of each species at each
site.
6.3.5 AFLP profiling
Total cellular DNA was extracted from 0.1 g of plant material per sample according
to the protocol of Doyle and Doyle (1987) with slight modifications. DNA was
quantified visually on ethidium bromide stained agarose gels and samples were
diluted with 0.5 TE buffer to obtain concentrations between 100 and 200 ng/µL.
AFLP restriction/ligation was performed following the protocol of Prentis et al.
(2004). AFLP PCR was performed following the method of Zawko et al. (2001),
using two primer pairs: E-AAG/M-AG and E-AAG/M-GA, where the selective EcoRI
primer was Hex labeled (Geneworks). The fluorescently labeled amplified products
were analysed by gel electrophoresis (5% acrylamide gels), using a Gelscan
GS2000 (Corbet Research) with a TAMRA 500 size standard (Applied
Biosystems). To confirm reproducibility, five adult samples of each Senecio species
were run blindly six times from different extractions for both primer combinations
and loci that were ambiguous were not scored in the full analysis. This information
was also used to produce an error rate of fragment mis-scoring for both primer
combinations. At an individual locus, bands of similar size and intensity were
considered to be homologous, following previous studies of closely related species

126
(Rieseberg 1996; O'Hanlon and Peakall 2000). AFLP profiles were scored for the
presence and absence of bands between 50 and 500 base pairs in size.
6.3.6 Data analysis – population level
Genetic diversity within each population was quantified by calculating Shannon's
index of diversity (Shannon 1948), as this diversity measure has been used
previously to obtain accurate estimates of genetic diversity in polyploid plants with
AFLP markers (Abbott et al. 2007). Shannon’s index was calculated using the
following equation; H = −∑(pi ln pi), where pi is the frequency of a band at a
particular locus, and this value was then averaged over all polymorphic loci. A t-test
was used to compare whether levels of genetic diversity were similar in sympatric
and allopatric populations of both species.
Global FST and pairwise FST, used to characterize the extent of population
differentiation among all population pairs within each species separately, were
estimated in SPAGEDI (Hardy and Vekemans 2002). This program was chosen as
it can estimate F statistics in both diploids and polyploids with dominant marker
data. PHYLIP (Felsenstein 2005) was used to construct a neighbour-joining (NJ)
phenogram in TREEVIEW (Page 1996) from the pairwise FST matrix.
6.3.7 Data analysis – individual level
Principal coordinates analysis (PCOA) was used to examine clustering of individual
S. pinnatifolius and S. madagascariensis genotypes from both sympatric and
allopatric sites using GENALEX (Peakall and Smouse 2006). To assign individuals
to their most likely species of origin, or hybrid status, the assignment method of
Duchesne and Bernatchez (2002) in AFLPOP was used. The assignment method
utilises multilocus AFLP data to test the likelihood that an individual genotype (G) is
a pure species or interspecific hybrid based on population-level allele frequencies.

127
If the frequency of an AFLP fragment was 0, log(0) was replaced by log(ε), where ε
was chosen as 0.001. Individuals are assigned to species or hybrid populations
displaying the highest log-likelihood for G; however allocation of genotypes were
only made if minimal log-likelihood difference (MLD) was ≥ 1 for mature individuals.
This means a genotype is 10 times more likely to originate from a particular
population than any other candidate population. A MLD of 0 was used to allocate
progeny genotypes to parental species or hybrid swarms, as many individuals were
unassigned at higher MLD stringency levels. The MLDs chosen here are similar to
most previous studies (Potvin and Bernatchez 2001; Campbell et al. 2003; He et al.
2004).
To determine the probability of incorrect assignment the AFLPOP simulator was
used. The simulation technique produces 1000 random samples from the source
population file and calculates the proportion of allocations (P) to the second
population. When P is small the incorrect assignment of individuals is highly
unlikely. If P-values for an individual were < 0.001 for both species and all possible
hybrid populations, then the individual could not be assigned.
First generation, F1 parental backcrosses and F2 hybrid populations were
simulated in AFLPOP between all pairs of allopatric populations of S. pinnatifolius
and S. madagascariensis. Mature sympatric individuals of the two species were
then assigned to either allopatric populations or simulated hybrid swarms. Progeny
raised from seed collected from sympatric sites were also allocated to their species
of origin or simulated interspecific hybrid status using the same assignment method
as above.

128
6.3.8 Risk posed by hybridisation
We estimated the number of non-hybrid adults of each species that would be
produced from seeds in a single year under various rates of F1 seed production.
We incorporated data from other studies for the following parameters; monthly
capitulum production for both species in allopatric sites (see Appendix C) to
estimate the proportion of total capitula produced per year during synchronous (Pr
S) and non-synchronous (Pr N) flowering, annual seed production (A), percentage
germination under field conditions (G), survival transition to maturity of both
species in S. pinnatifolius (var. tableland) habitat (E), and hybridisation rate (H)
(see Appendix D for values). Annual viable seed production (AVSP) was then
calculated for both species using the following equation; AVSP = ((Pr S x A) x (1-
H) x G x E) + ((Pr N x A) x G x E). Hybridisation scenarios examined with the
equation were; no hybridisation, maximum hybridisation (all seeds produced during
synchronous flowering were hybrids), fixed level hybridisation (based on actual
levels of hybridisation observed in field-collected progeny in this study), and linear
frequency dependent hybridisation. The hybridisation rate (H) was calculated for
each month, based on flowering synchrony data from field observations (Radford
and Cousens 2000, Appendix C). The proportion of S. madagascariensis (Pm) in a
population is used to estimate the proportion of hybrid seed produced separately
for both S. madagascariensis and S. pinnatifolius using linear frequency dependent
relationships outlined below. Linear density dependent relationships were fitted
based on the assumption that H = (1 – observed H) at Pm = (1 – observed Pm), for
each site and species independently (Equations for lines of best fit, O’Reillys’: S.
madagascariensis y = (-0.101(Pm)) + 0.103, S. pinnatifolius y = (0.739(Pm)) +
0.13; Queen Mary Falls: S. madagascariensis y = (-0.532(Pm)) + 0.766, S.
pinnatifolius y = (0.894(Pm)) + 0.053). Estimates were calculated independently for
each sympatric site based on the actual levels of hybridisation recorded in open

129
pollinated progeny at that site for the fixed rate hybridisation scenario. The principal
simplifying assumptions of our estimates include (1) flowering time in sympatric
populations is similar to allopatric populations (2) rates of hybridisation are
frequency dependent and (3) all hybrids are not viable.
6.4 Results
6.4.1 Reciprocal crossing experiments
Achenes were successfully produced for both interspecific and intraspecific
crosses, regardless of which species was the pollen or seed parent. Although the
mean number of seeds produced from interspecific crosses was lower than that
recorded for for intraspecific crosses, differences in seed production were not
statistically significant (see Table 6.2). Similarly, no statistical difference in
percentage seed germination (viability) was found between the seed produced
from interspecific and intraspecific crosses (Table 6.2).
6.4.2 Relative densities of plants and flowers
The native S. pinnatifolius was the dominant species at both sympatric sites, both
in terms of plant and flower frequency: it had more than three-fold the plant
frequency and approximately 19-fold the flower frequency of S. madagascariensis
(Table 6.1). At both sites the rate of hybrid seed production by S. pinnatifolius was
significantly higher than would be expected if it was occurring proportionally to the
relative frequencies of S. pinnatifolius and S. madagascariensis flowers (O’Reillys’:
χ2 = 5.43, df = 1, p < 0.05; Queen Mary Falls: χ2 = 102.48, df = 1, p < 0.01).
Senecio madagascariensis contributed only five percent of capitula in each of the
sympatric populations, but approximately 15 and 8.5 % of S. pinnatifolius progeny
were identified as F1 hybrids at Queen Mary Falls and O’Reillys’ sites respectively.
In contrast, the rates of hybridisation in S. madagascariensis seed were

130
significantly lower than expected from floral frequency (O’Reillys’: χ2 = 57.76, df =
1, p < 0.05; Queen Mary Falls: χ2 = 1375.14, df = 1, p < 0.01). Senecio pinnatifolius
makes up 95 % of capitula at both sites, but only 10 and 75 % of S.
madagascariensis progeny were recognized as hybrids at the Queen Mary Falls
and O’Reillys’ sites, respectively.
Table 6.2: Seed viability (% germination ± s.e.) and amount of seed produced (mean seed
produced/capitulum ± s.e.) from intra and interspecies reciprocal crosses between Senecio
pinnatifolius and Senecio madagascariensis.
Experimental
crosses
Number of
crosses
(N)
Mean seed
produced/capitulum
(± s.e.)
% Seed
germination
(± s.e.)
Statistical
significance
Seed
produced
Within species 10 46 (± 9.09)
Between
species
13 26 (± 7.19) P = 0.180
Seed viability
Within species 8 70.4 (± 6.07)
Between
species
11 75.2 (± 11.9) P = 0.502
6.4.3 Genetic diversity and population differentiation
The two AFLP primer pair combinations produced 176 fragments for the 718
individuals screened, of which 88% were polymorphic between the two species.
The error rate of mis-scoring estimated from blind running of five individuals of
each Senecio species six times from different extractions was 1.7% and 1.9% for

131
the primer pairs 33-49 and 33-55, respectively. Mean genetic diversity within S.
madagascariensis and S. pinnatifolius populations was H = 0.257 (± 0.007) and H
= 0.277 (± 0.014), respectively. Genetic diversity was similar between allopatric
(0.283 ± 0.013) and sympatric (0.270 ± 0.015) populations for S. pinnatifolius (T3 =
0.648; P > 0.5). However, a significant difference in genetic diversity between
allopatric (0.271 ± 0.003) and sympatric (0.239 ± 0.010) populations of S.
madagascariensis was detected ( T3 = 3.968; P = 0.02).
0.1
B (Sm)A
S (Sm)A
T (Sm)A
O (Sm)S
QM (Sm)S
QM (Sp)S
O (Sp)S
S2 (Sp)A
S1 (Sp)A
H (Sp)A
a)
b)
Figure 6.1: Unrooted neighbour-joining phenogram based on pairwise FST distances among AFLP profiles for a)
Senecio pinnatifolius (Sp) and b) Senecio madagascariensis (Sm) in sympatric (S) and allopatric (
A) sites, i.e.
Hampton (H), Swanfels 1 (S1), Swanfels 2 (S2), Beechmont (B), Tamborine (T), Springbrook (S), Queen Mary
Falls (QM) and O’Reillys’ (O).
Global FST analyses detected pronounced differentiation among populations of both
species, with FST values of 0.271 (P < 0.001) for S. madagascariensis, and 0.162
(P < 0.001) for S. pinnatifolius. The NJ phenograms (Figure 6.1) illustrated that for
each species, sympatric populations were more similar genetically to each other
than they were to allopatric populations (S. pinnatifolius: sympatric – allopatric
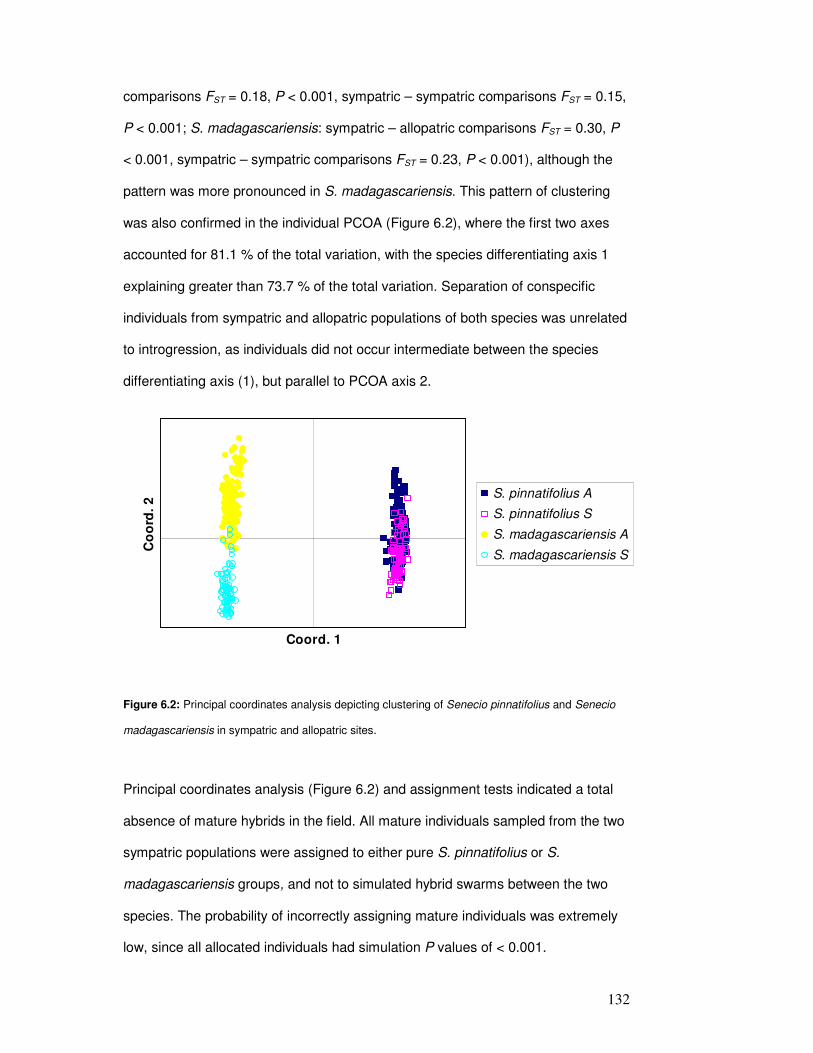
132
comparisons FST = 0.18, P < 0.001, sympatric – sympatric comparisons FST = 0.15,
P < 0.001; S. madagascariensis: sympatric – allopatric comparisons FST = 0.30, P
< 0.001, sympatric – sympatric comparisons FST = 0.23, P < 0.001), although the
pattern was more pronounced in S. madagascariensis. This pattern of clustering
was also confirmed in the individual PCOA (Figure 6.2), where the first two axes
accounted for 81.1 % of the total variation, with the species differentiating axis 1
explaining greater than 73.7 % of the total variation. Separation of conspecific
individuals from sympatric and allopatric populations of both species was unrelated
to introgression, as individuals did not occur intermediate between the species
differentiating axis (1), but parallel to PCOA axis 2.
Coord. 1
Co
ord
. 2
S. pinnatifolius A
S. pinnatifolius S
S. madagascariensis A
S. madagascariensis S
Figure 6.2: Principal coordinates analysis depicting clustering of Senecio pinnatifolius and Senecio
madagascariensis in sympatric and allopatric sites.
Principal coordinates analysis (Figure 6.2) and assignment tests indicated a total
absence of mature hybrids in the field. All mature individuals sampled from the two
sympatric populations were assigned to either pure S. pinnatifolius or S.
madagascariensis groups, and not to simulated hybrid swarms between the two
species. The probability of incorrectly assigning mature individuals was extremely
low, since all allocated individuals had simulation P values of < 0.001.
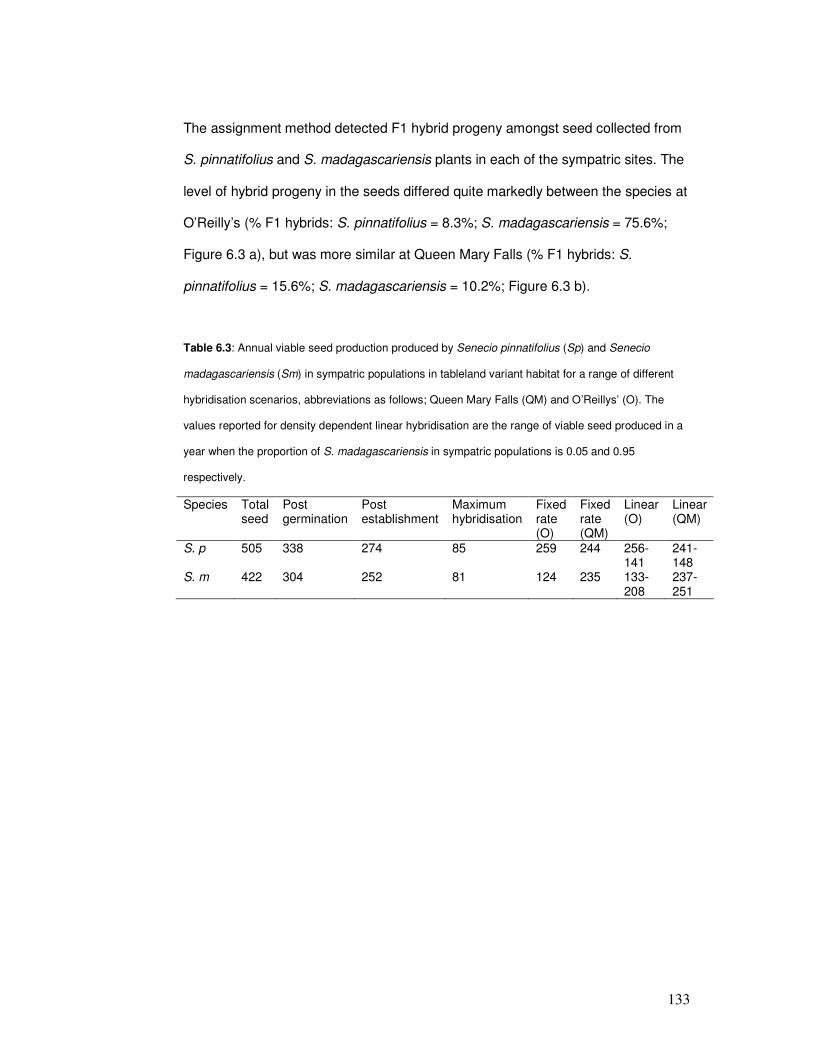
133
The assignment method detected F1 hybrid progeny amongst seed collected from
S. pinnatifolius and S. madagascariensis plants in each of the sympatric sites. The
level of hybrid progeny in the seeds differed quite markedly between the species at
O’Reilly’s (% F1 hybrids: S. pinnatifolius = 8.3%; S. madagascariensis = 75.6%;
Figure 6.3 a), but was more similar at Queen Mary Falls (% F1 hybrids: S.
pinnatifolius = 15.6%; S. madagascariensis = 10.2%; Figure 6.3 b).
Table 6.3: Annual viable seed production produced by Senecio pinnatifolius (Sp) and Senecio
madagascariensis (Sm) in sympatric populations in tableland variant habitat for a range of different
hybridisation scenarios, abbreviations as follows; Queen Mary Falls (QM) and O’Reillys’ (O). The
values reported for density dependent linear hybridisation are the range of viable seed produced in a
year when the proportion of S. madagascariensis in sympatric populations is 0.05 and 0.95
respectively.
Species Total seed
Post germination
Post establishment
Maximum hybridisation
Fixed rate (O)
Fixed rate (QM)
Linear (O)
Linear (QM)
S. p 505 338 274 85 259 244 256-141
241-148
S. m 422 304 252 81 124 235 133-208
237-251
134
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
plants capitula Sp progeny Sm
progeny
hybrid
Sm
Sp
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
plants capitula Sp
progeny
Sm
progeny
hybrid
Sm
Sp
Figure 6.3: Percentage of plants of each species, capitula produced by each species and hybrid and
non-hybrid F1 progeny produced by Senecio pinnatifolius (Sp) and Senecio madagascariensis (Sm)
plants in two sympatric populations; a) Queen Mary Falls and b) O’Reillys’.
6.4.4 Risk posed by hybridisation
Estimates of the annual viable seed production (AVSP) were found to favour the
native S. pinnatifolius under all hybridisation scenarios (Table 6.3), except under
linear frequency dependent relationships where the proportion of S.
(a)
(b) Perc
en
t (%
)

135
madagascariensis in sympatric populations strongly influenced the outcome
(Figure 6.4). The number of seeds to become viable adults of each species in a
year estimated under no hybridisation was greater for S. pinnatifolius (274) than for
S. madagascariensis (252). Senecio pinnatifolius also produced a greater amount
of viable seed than S. madagascariensis, when estimates were based on the fixed
hybridisation rates observed in this study, but this trend was stronger at O’Reillys’
(259:124) than QM Falls (244:235). Estimates of maximum possible hybridisation
also indicated S. pinnatifolius (85) would produce more viable seed than S.
madagascariensis (81), but only by four seeds in a generation. Calculations based
on linear frequency dependent relationships produced estimates for seven different
proportions of S. madagascariensis (5, 10, 25, 50, 75, 90 and 95 %) in both
sympatric sites (see Figure 6.4 a and b). At the QM Falls and O’Reillys’ sites, the
proportion of S. madagascariensis in a mixed population needed to reach 10 %
and ~ 60 % respectively, for S. madagascariensis to produce more viable seed
than S. pinnatifolius in a generation.
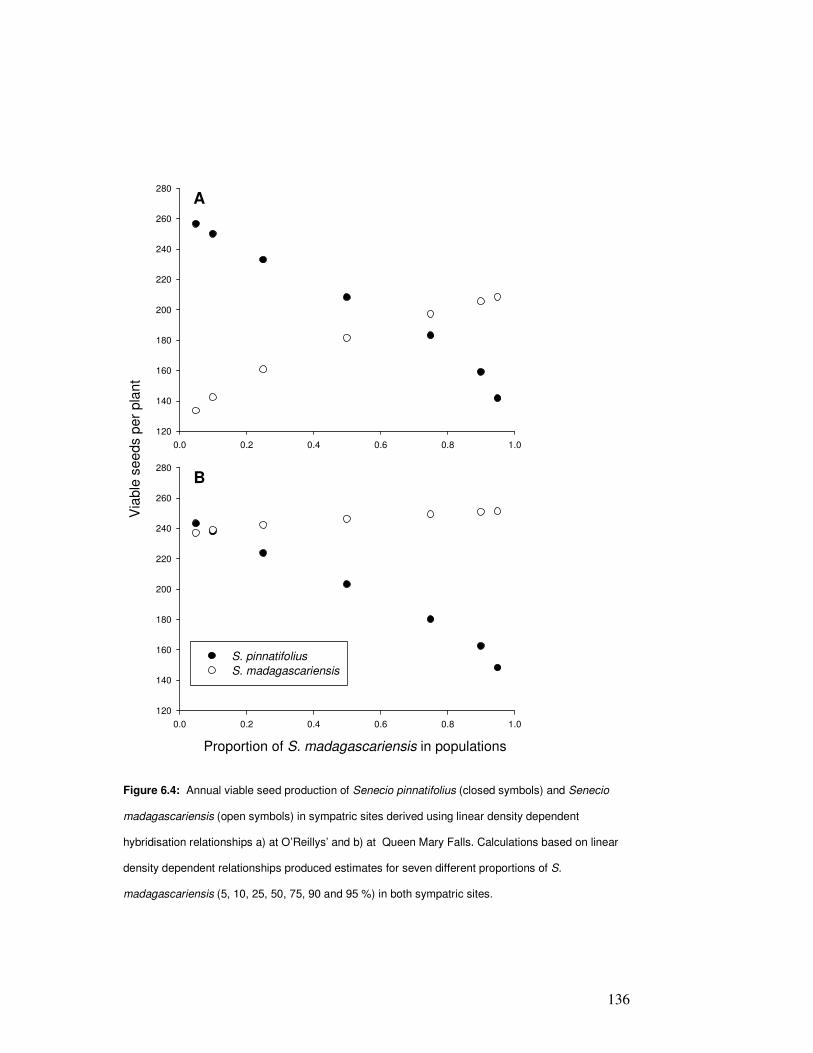
136
A
0.0 0.2 0.4 0.6 0.8 1.0
120
140
160
180
200
220
240
260
280
B
Proportion of S. madagascariensis in populations
0.0 0.2 0.4 0.6 0.8 1.0
Via
ble
se
eds p
er
pla
nt
120
140
160
180
200
220
240
260
280
S. pinnatifolius
S. madagascariensis
A
Figure 6.4: Annual viable seed production of Senecio pinnatifolius (closed symbols) and Senecio
madagascariensis (open symbols) in sympatric sites derived using linear density dependent
hybridisation relationships a) at O’Reillys’ and b) at Queen Mary Falls. Calculations based on linear
density dependent relationships produced estimates for seven different proportions of S.
madagascariensis (5, 10, 25, 50, 75, 90 and 95 %) in both sympatric sites.

137
6.5 Discussion
6.5.1 The incidence of hybridisation and fate of hybrids
Hybridisation between S. madagascariensis and S. pinnatifolius occurs very
frequently in the wild, with a large number of F1 hybrid seed produced by both
species in sympatric sites. Observed levels of hybridisation in this study were in the
same range as those reported previously between S. madagascariensis and S.
pinnatifolius (Radford 1997). In fact, the level of hybridisation recorded in open
pollinated seed is four orders of magnitude greater than that recorded between
another well characterized native-exotic Senecio species pair, S. vulgaris (2n = 4x
= 40) and S. squalidus (2n = 2x = 20) (Marshall and Abbott 1980). It is also an
order of magnitude higher than between S. vulgaris and the recent neo-species S.
eboracensis (2n = 4x = 40). Lowe and Abbott (2004) suggest that the low
frequency of hybridisation between S. eboracensis and S. vulgaris was influenced
by niche separation, differences in flowering phenology and the greater attraction
of S. eboracensis to pollinators. Given that habitat differentiation between S.
madagascariensis and S. pinnatifolius is weak, there is a substantial overlap in
their flowering time, and that they are pollinated by the same insect species, the
high level of hybridisation observed here is not unexpected. The frequency of
hybridisation in our study suggests that prezygotic barriers are weak and do not
prevent gene flow between the species.
Despite the high proportion of hybrid seed collected from both species, mature
hybrids were totally absent from sympatric populations sampled in this study.
These results suggest that there is a very strong postzygotic reproductive barrier
between the study species. Effects of interploidal hybridisation on offspring fitness
can be severe, often resulting in progeny that are highly sterile (Hardy et al. 2001;
Lowe and Abbott 2004; Pannell et al. 2004; Buggs and Pannell 2006). However
sterility cannot be the only consequence of interploidal hybridisation for the study

138
species, since no hybrids, sterile or otherwise, developed to maturity in sampled
populations. A lack of mature hybrids indicates that the viability of interspecific
hybrids must also be much reduced. Given that in this study hybrid seed
germinates at the same percentage as non-hybrid seed, the reduced viability of
hybrids must occur after germination but before maturity. Further study is required
to estimate the exact life history stage at which hybrids are selected against.
Hybrids grown in pots were found to be of low vigor compared to either parental
species (Radford 1997), suggesting out-breeding depression, which may explain
the absence of mature hybrids in the field.
Since the rapid spread of S. madagascariensis, hybrid zones between S.
pinnatifolius and S. madagascariensis have formed in many areas of eastern
Australia (Radford 1997). An absence of mature F1 hybrids in sympatric
populations indicates that contact zones formed between S. pinnatifolius and S.
madagascariensis may represent tension zones. Theoretical tension zone models
assume hybrid fitness is independent of environment and intrinsically low as the
result of genetic incompatibilities, but that low hybrid fitness is balanced by the
continual dispersal of parent types into areas of contact (Barton and Hewitt 1989).
Tension zones may also be maintained by positive frequency-dependent selection
(Buggs and Pannell 2006). Areas of contact between diploid and tetraploid
Centaurea jacea in Belgium (Hardy et al. 2000, 2001), and diploid and hexaploid
Mercurialis annua in northern Spain (Pannell et al. 2004), appear to be other good
examples of tension zones in mixed ploidy plant populations.
6.5.2 Long-term population impacts of hybridisation – genetic diversity and
differentiation
Overall levels of genetic diversity (HE) were higher in the native S. pinnatifolius
compared to the exotic S. madagascariensis. Genetic diversity was significantly

139
lower in allopatric compared to sympatric sites for S. madagascariensis, but no
significant difference was detected for S. pinnatifolius. The level of differentiation
among populations within species was pronounced (S. madagascariensis FST =
0.271, S. pinnatifolius FST = 0.162). Although there were no immediately obvious
impacts of hybridisation on differentiation, populations in areas of sympatry showed
increased differentiation from conspecific allopatric populations and this pattern
was more pronounced in S. madagascariensis.
The pattern of increased differentiation between allopatric and sympatric
populations appears unrelated to introgression, since the PCOA axis of
differentiation was perpendicular to the axis differentiating the two species. A loss
of alleles in non-viable hybrids of early flowering S. madagascariensis, or late
flowering S. pinnatifolius genotypes, might change allele frequencies in sympatric
populations and may be responsible for the observed pattern of differentiation.
Similarly a loss of alleles in non-viable hybrids may also explain lower genetic
diversity of S. madagascariensis at sympatric sites however further work is
warranted on this topic.
6.5.3 What does the future hold for S. pinnatifolius?
In areas of contact between S. pinnatifolius and S. madagascariensis, calculations
demonstrated S. pinnatifolius was not at risk from demographic swamping when no
hybridisation occurred or when levels of hybridisation were constant and not
affected by the proportion of S. madagascariensis. However, S. madagascariensis
displays a hybridisation advantage at both surveyed field sites, where it sires
significantly more progeny than expected based on capitulum frequencies, and S.
pinnatifolius significantly less. Thus hybridisation between the species is
asymmetric, a phenomenon commonly reported in hybrid zones (Rieseberg and
Wendel 1993; Arnold 1997; Burgess et al. 2005). Estimates based on frequency

140
dependent asymmetric hybridisation between the species, indicate that the
proportion of S. madagascariensis need only reach between 10 - 60 % to produce
more viable seeds than S. pinnatifolius in sympatry (Figure 6.4). Under these
circumstances, an invasive species does not necessarily have to outnumber a
native to have an impact on the demography of an interfertile native through
hybridisation. In fact, invasive species may be rare relative to a native plant, but
may nevertheless pose a threat to the native due to superior male fitness (e.g.
production of a greater number of pollen grains), resulting in the invader siring a
disproportionately higher proportion of progeny (Anttila et al. 1998). As a result,
asymmetric hybridisation in favour of an invasive species can contribute to the
decline and extinction of native species (Wolf et al. 2001). Thus, if S.
madagascariensis increases numerically in areas of contact, it may cause the
decline of S. pinnatifolius from east coast areas of Australia.
Three factors may impede the decline of S. pinnatifolius. First, S.
madagascariensis may be driven to local extinction in areas of contact during
colonisation, if it cannot establish within a few generations. Given that S.
madagascariensis can reproduce in the absence of S. pinnatifolius for six months
annually and the O’Reillys’ contact zone has existed for between 14-25
generations (first recorded by Scott 1994), this outcome is unlikely. Second, natural
selection against maladaptive hybridisation may lead to reproductive character
displacement (eg. flowering time divergence) and “avoidance” of the negative
consequences associated with interspecific fertilizations. Reinforcing natural
selection is most likely when contact zones are extensive, exposing a high
proportion of individuals to selection (Pannell et al. 2004; Hoskin et al. 2005). As S.
pinnatifolius and S. madagascariensis form extensive contact zones, reinforcement
may act to impede displacement of S. pinnatifolius. Third, S. pinnatifolius variants
may have physiological and morphological adaptations to specific environments,

141
which allow variants to out perform S. madagascariensis in their native habitat
(Radford and Cousens 2000).
The destructive force of interspecific hybridisation is not uncommon in hybridising
plant species (Wolf et al. 2001; Buggs and Pannell 2006). However, adequate
molecular data from open pollinated progeny and/or mature individuals are often
lacking, meaning the actual level of hybridisation and its impact on native or rare
species are underestimated. Without this information conservation strategies for
the protection of hybridising species cannot be effective. In combination with
ecological approaches, we encourage the use of molecular data to provide a
baseline for comprehensive long-term studies into the consequences of
hybridisation on native species.


Chapter 7 – General Discussion


145
Chapter 7 – General Discussion
The studies described in this thesis focus on some of the more subtle, complex,
and less-frequently-studied biotic interactions that can occur between an invasive
and native species. The two Senecio species investigated here have overlapping
populations, coinciding flowering seasons, and similar suites of insect pollinators
and herbivores, thus they provide an ideal model system in which to investigate the
occurrence of insect-mediated indirect effects. In this system such interactions
could potentially be mediated by either insect pollinators or insect herbivores and,
depending on the nature of the interaction, might have either a positive, negative or
neutral effect on either or both plant species. This discussion focuses on the
pollinator and herbivore mediated indirect effects identified within the thesis, and
highlights the importance of taking this type of interaction into account when
assessing impacts of, and designing management strategies for, invasive species.
7.1 Pollinator-mediated indirect interactions
Invasive species often require mutualistic relationships in order to successfully
invade new environments. For the majority of self-incompatible plant species,
insect-pollination is a key mutualism without which population establishment and
spread could not occur (Hanley and Goulsen 2003). It is hypothesised that, owing
to the generalist nature of many plant-pollinator interactions and the widespread
naturalisation of highly effective generalist pollinators such as the honeybee, Apis
mellifera, plant invasions are rarely limited by lack of pollinators (Richardson et al.
2000).
Empirical studies have also demonstrated the ability of invasive plants to utilise
local native or alien pollinators in their new range (Jesse et al. 2006; Liu et al.
2006). This is apparently the case for S. madagascariensis in its naturalised range

146
in eastern Australia. Senecio madagascariensis receives frequent visits from a
similar suite of generalist insect pollinators - dominated by Apis mellifera and
syrphid species - to those that visit the native S. pinnatifolius. Interaction with this
group of pollinators not only allows successful spread of the invader in its new
range, but also creates the opportunity for pollinator-mediated indirect interactions
to occur between S. madagascariensis and native species such as S. pinnatifolius.
Within their native range, pollinators have been shown to demonstrate a
preference for the pollen of particular species (even among closely related,
morphologically similar plant species) (Hersch and Roy 2007). Pollinators may
demonstrate a preference for species with which they have coevolved and this
association may be maintained when both insects and plants are moved outside
their native range, resulting in a positive synergistic relationship between alien
plants and alien pollinators (Hanley and Goulsen 2003). Conversely native
pollinators may develop a preference for a novel species, choosing attractive
flowers of an alien plant species over less appealing native flowers (Chittka and
Schurkens 2001). Invasive plants have been demonstrated to thus compete with
natives for the services of pollinators (Chittka and Schurkens 2001; Moragues and
Travaset 2005).
Such an effect is not apparent in my Senecio system. Visitation rates to S.
pinnatifolius by the two dominant pollinator groups, honey bees and syrphids, were
not reduced in populations that occurred in sympatry with S. madagascariensis. In
fact, contrary to the hypothesis that alien pollinators prefer the flowers of alien
plants, the results of this study suggest that in areas of sympatry, it is the native
Senecio that competes with its alien congener for bee visits. Syrphid visits to the
invader, on the other hand, increase in sympatric populations. Syrphid visitation
rates are possibly indirectly facilitated by the presence of the native Senecio due to

147
elevated syrphid populations in regions inhabited by the native plant. Alternatively
syrphid visits may increase in response to reduced bee visitation rates (and
subsequent reduction in interference competition from bees) in overlapping
populations.
Altered pollinator activity in sympatric populations apparently has neither a positive
nor negative impact on seed set in S. madagascariensis, suggesting that, like
many other successful invasive plants (Richardson et al. 2000; Jesse et al. 2006),
the spread of this species is probably not constrained by pollen-limitation, at least
in this part of its naturalised range. Pollinator visitation patterns (Hersch and Roy
2007) and pollen limitation (Liu et al. 2006) are known to vary widely depending on
a range of biotic and abiotic variables, so this situation may differ in other regions.
Surprisingly, S. pinnatifolius seed set was higher, rather than lower, in populations
growing in sympatry with S. madagascariensis. There are a number of possible
explanations for this. Firstly, abiotic factors which were not measured in this study
may explain differences in seed set. Secondly, seed set in pure S. pinnatifolius
stands may be lower than in mixed stands owing to the effects of some other biotic
interaction such as plant-herbivore interactions. Herbivory has been shown to
result in both reduced pollen production (Hersch 2006) and seed set (Crawley and
Gilman 1989; Juenger and Bergelson 1997; Hersch 2006) in other species. This is
consistent with the findings presented in Chapters Four and Five of this thesis,
which demonstrated both higher rates of herbivory and reduced seed set in
isolated S. pinnatifolius populations. A third explanation may be that if S.
madagascariensis has higher pollen germination rates and thus higher male fitness
than S. pinnatifolius, seed set may be higher in mixed stands due to interspecific
pollen transfer and hybridisation (see Chapter Six). If this were the case, enhanced

148
seed set will clearly not translate into greater fecundity, since the increase in seed
set is attributable to hybrid seeds, which are unlikely to survive to maturity.
Pollinators can mediate indirect interactions between plant species by transferring
pollen between species, which can have consequences ranging from gametic
wastage (Levin 1995), through to dilution of the native gene pool due to
introgression (Wolf et al. 2001), or production of hybrid offspring that may be
capable of out-competing (Vilà et al. 2003), or dramatically altering indirect
interactions between the parent species (Vilà and D’Antonio 1998; Whitham et al.
1999; Hersch and Roy 2007).
At the time of year during which the study reported in Chapter Six took place, up to
75 percent and 16 percent of seed produced by S. madagascariensis and S.
pinnatifolius maternal parents respectively were hybrids produced as a result of
cross-pollination. Few, if any, of these hybrids are likely to survive to maturity, thus
this represents gametic wastage for both species. The long-term population-level
impacts of this phenomenon in sympatric populations are unknown. However,
given that S. madagascariensis has a six-month window of opportunity in which to
reproduce during which S. pinnatifolius is not flowering it seems unlikely that the
invader will experience any dramatic negative impacts. Due to asymmetric
hybridisation S. pinnatifolius may be under threat if S. madagascariensis increases
numerically in areas of contact. Further work is necessary to investigate the
reasons for the higher-than-expected rates of hybridisation observed in S.
pinnatifolius, to assess variation in rates of hybridisation throughout the flowering
season, and to determine the likely long-term consequences of hybridisation in this
system.

149
7.2 Herbivore-mediated indirect interactions
Plant-herbivore interactions can affect both invasion success and the impacts of an
invader on a system. For example, selective herbivory on a particular plant species
can give other plant species a competitive advantage (Brown 1994; Carson and
Root 2000; Centre et al. 2005). Invasions can be facilitated indirectly when native
species are preferentially grazed upon (Cross 1981; Edwards et al. 2000), or native
plant species may benefit through preferential animal grazing of alien plants
(Zancola et al. 2000). As outlined in Chapter Two, positive indirect effects (indirect
mutualism) in such plant-herbivore systems can occur via host plant switching,
herbivore satiation (Abrams and Masuda 1996), or by a herbivore targeting an
abundant host-plant species, ignoring the less common plant (Abrams 1987), whilst
established herbivore-plant interactions can be negatively impacted through
apparent competition.
Whilst S. madagascariensis receives the benefits of pollinator services in its
naturalised range, it does not appear to experience the potentially negative
consequences associated with insect herbivory. The invader attracted neither the
numbers nor the diversity of insect herbivores hosted by the native Senecio.
Laboratory trials demonstrated that although larvae of a common native arctiid
moth, Nyctemera amica, could successfully survive when reared on foliage of
either Senecio species, both ovipositing females and the larvae show a preference
for the native species. The native Senecio also incurs significantly greater
herbivore damage to foliage than does its alien congener in the field. Results
therefore lend support to the Enemy Release Hypothesis which predicts that
invasive species may be successful, in part, because they escape from their
natural enemies in their new range.

150
Food quality for herbivores is determined by the nutrient and water content of the
plant material, as well as by the concentrations of secondary metabolites (Slansky
and Rodriquez 1987). Specialist herbivores are often unaffected, or even attracted
to higher levels of certain plant defence compounds (Bowers 1984; Leimu et al.
2005). Herbivore preference for S. pinnatifolius could be explained by the higher
nutritional content of the native species, or by differences in levels of secondary
compounds between the two species.
Theory predicts that a less preferred host – in this case S. madagascariensis - may
experience apparent competition if herbivore populations are limited by food
availability or if herbivores aggregate on preferred host patches and spill over onto
secondary hosts nearby (Holt 1977; Abrams and Masuda 1996). In the context of
invasion biology, several studies have demonstrated the occurrence of apparent
competition between an invasive plant species and a native plant species mediated
by an introduced biocontrol agent, which damages the native plant in the vicinity of
its alien host (e.g. Rand and Louda 2004; Russell et al. 2007).
To date there have (to my knowledge) been no published studies demonstrating
such negative indirect impacts of a native plant on a sympatric alien species,
although this scenario could conceivably occur, mediated by a native (or alien)
herbivore. No evidence was found here, however, for the occurrence of apparent
competition between the two Senecio species. In fact both species experienced
lower, rather than higher levels of herbivory in sympatry than when growing in
isolation. If the herbivore population is not food-limited, the reduced damage in
overlapping populations may be due to a dilution-effect of herbivore damage in a
larger mixed population containing both plant species, thus in sympatry the two
species may be having a facilitative effect on one another. Similarly apparent
competition may not occur if the herbivore is not food-limited. Of course the

151
situation may be different at different times of the year or in a situation in which
herbivores are more abundant or foliage of host plants more scarce.
7.3 Implications for management of invasive species
This research, focussing on a model plant-herbivore and plant-pollinator system,
highlights the potential significance of indirect effects in invasion biology, as well as
the importance of understanding the mechanisms behind the observed population
dynamics in invaded systems. Indirect effects can range from the (arguably) trivial
and harmless, such as when the presence of one species alters pollinator visitation
rates to a second species whilst seed set remains unaffected (Chapter Five in this
thesis; Ghazoul 2004), through to the dramatic impacts of the introduction of a new
species precipitating a trophic cascade that alters an entire food web (Flecker and
Townsend 1994, 1996)
These complex interactions should be taken into account not only when assessing
an alien’s ecological impacts, but also when developing control strategies for
invasive species. For instance, Pearson and Callaway (2003) emphasise the
importance of identifying likely non-target indirect effects of biological control
programs, and suggest that interaction strength between a biocontrol agent and its
host is at least as important as host specificity in determining ecological impacts of
the potential agent. Biocontrol agents can have a range of unintended indirect
effects on co-occurring species, as illustrated by Callaway et al. (1999) and
Ridenour et al. (2004). Studies by these workers showed that herbivory by the root-
boring biocontrol moth Agapeta zoegata on the invasive forb Centaurea maculosa
in North America had a negative indirect impact on co-occurring native grass,
Festuca idahoensis. Insect herbivory on C. maculosa failed to reduce biomass of
the alien, instead having the unexpected effect of reducing reproduction in the
native grass. The authors hypothesise that this may be due to a number of possible

152
mechanisms: (1) A strong compensatory growth response to herbivory by C.
maculosa might result in increased resource competition with the native; (2)
Herbivory might stimulate production of harmful root exudates which negatively
impact the native; (3) the negative effect of herbivory on F. idahoensis may be
mediated by complex indirect interactions involving mycorrhizal fungi. These
findings emphasise the importance of looking beyond the most obvious direct biotic
interactions when determining the effect of one species on others, particularly
when dealing with species additions or removals within a system.
Unintended effects on non-target species may occur even without the introduction
of biocontrol agents, simply as a result of the removal of an alien species from a
system. For instance, control of weeds that support large populations of insect
herbivores can result in the migration of the herbivores to the crop species (Barnes
1970; Geddes et al. 1992). Although the study of this phenomenon has been
largely restricted to agricultural systems, it is likely that in some situations removal
of alien species from natural systems will have similarly negative consequences for
native biota.
It is widely accepted that direct biotic interactions, such as herbivory, can be
manipulated to assist with control of invasive species: this principal forms the basis
of biological control theory and practice. However, deliberate manipulation of biotic
indirect interactions by land managers rarely occurs. Exceptions exist in the control
of insect pests in agricultural systems. The establishment of ‘beetle banks’
(overwintering habitats for invertebrate predators of cereal aphids), puts into
practice apparent competition (see Chapter Two), for the purpose of controlling
aphids in crops. Beetle banks allow predator populations to be maintained over
winter, allowing them to more effectively control aphid populations early in the
season (MacLeod et al. 2004). Similarly the use of ‘trap crops’ (plant stands grown

153
to attract herbivorous insects away from target crops), takes advantage of indirect
facilitation to control herbivores in agricultural systems (Shelton and Badenes-
Perez 2006). The intentional use of indirect interactions is however rarely, if ever,
used as part of an integrated management strategy for invasive plants, nor is there
documented use of such techniques in natural systems. This is probably due in
part to our thus-far limited understanding of these complex interactions, as well as
the lack of an economic incentive to develop novel management strategies for
invasive plant species in natural systems.
With an improved understanding of indirect interactions, perhaps they could be
used to our benefit when designing integrated weed management strategies.
Apparent competition might be used to our advantage through the planting of a
native species that provides shelter or an alternative food source for a herbivore,
thereby allowing it to more effectively attack a neighbouring invasive species. Such
a strategy might be used as part of an integrated management approach, as an
adjunct to a biocontrol program or other control methods. Clearly an in-depth
knowledge of the interactions occurring in the system would be required in order to
utilise such a strategy.
Pollination by insects is essential for the successful invasion of many alien plant
species (Hanley and Goulsen 2003). The study described in Chapter Five provides
evidence that the native S. pinnatifolius competes for bee visits with the alien S.
madagascariensis in areas where the two species coexist. In this system, seed set
in the alien is not reduced as a consequence of competition for pollinators. Under
particular circumstances, however, pollen limitation may be a key factor in
determining rates of spread of invasive species (Parker and Haubensak 2002).
With a greater knowledge of multispecies interactions, it may be possible to
manipulate these mutualisms by planting a native species which is highly attractive

154
to pollinators and effectively competes for their services. Before such a strategy is
possible, it would be necessary to acquire some predictive ability of under what
circumstances competition for pollinators is likely to occur. It would also be
necessary to determine whether the alien species has the potential to be pollen-
limited and if so in what situations? Since pollen-limitation is more likely to occur in
small populations (Lamont et al. 1993; Agren 1996), manipulation of plant-
pollinator interactions is more likely to be effective in satellite populations of an
invader or recently colonised areas.
Although the vast majority of studies that investigate interactions between alien
and native species focus on the negative impacts of alien species on natives,
rather than vice versa, indirect interactions with natives also have the potential to
negatively impact invasive species. The deliberate manipulation of indirect effects
to reduce the impact of pest species has been clearly demonstrated in agricultural
systems for the management of insect herbivores. It is therefore conceivable that
such interactions might be similarly manipulated to assist in the control of invasive
flora (and fauna) in natural systems. Due to the complex nature of indirect
interactions, the whole suite of interactions occurring in a system may never be
easily predicted and it is likely to be very difficult to make generalisations that apply
across species, regions and ecosystems. As such, at least until we have a deeper
understanding of indirect interactions, this kind of approach may have to be
designed on a system-by-system basis.
7.4 Conclusions
The studies outlined in this thesis provide evidence that indirect interactions
mediated by insects (specifically competition for visits from bee pollinators,
facilitation of syrphid visits, and interspecific pollen transfer resulting in the
production of sterile hybrid seeds) are occurring between the invasive S.

155
madagascariensis and the native S. pinnatifolius. However, there is no evidence
that either species, as a consequence of these interactions, is experiencing either
negative or positive population-level impacts at this point in time, although
theoretical modelling suggests this could happen under some scenarios.
Increasingly, empirical evidence is pointing towards the importance of indirect
effects in influencing the outcome of invasions and the impacts of an alien invader
on native species. It is likely that in some cases observed impacts that are
attributed to more obvious interactions (such as interspecific competition) might in
fact be caused by other indirect effects (e.g. apparent competition). Further
investigation of indirect effects will provide us with a better understanding and
predictive ability of the range of potential impacts of an alien species and might
assist us in designing management strategies both for alien and native species.
Furthermore, from a theoretical viewpoint, a system that has experienced a species
addition can provide the ideal opportunity to investigate the importance of indirect
effects in structuring ecological systems.

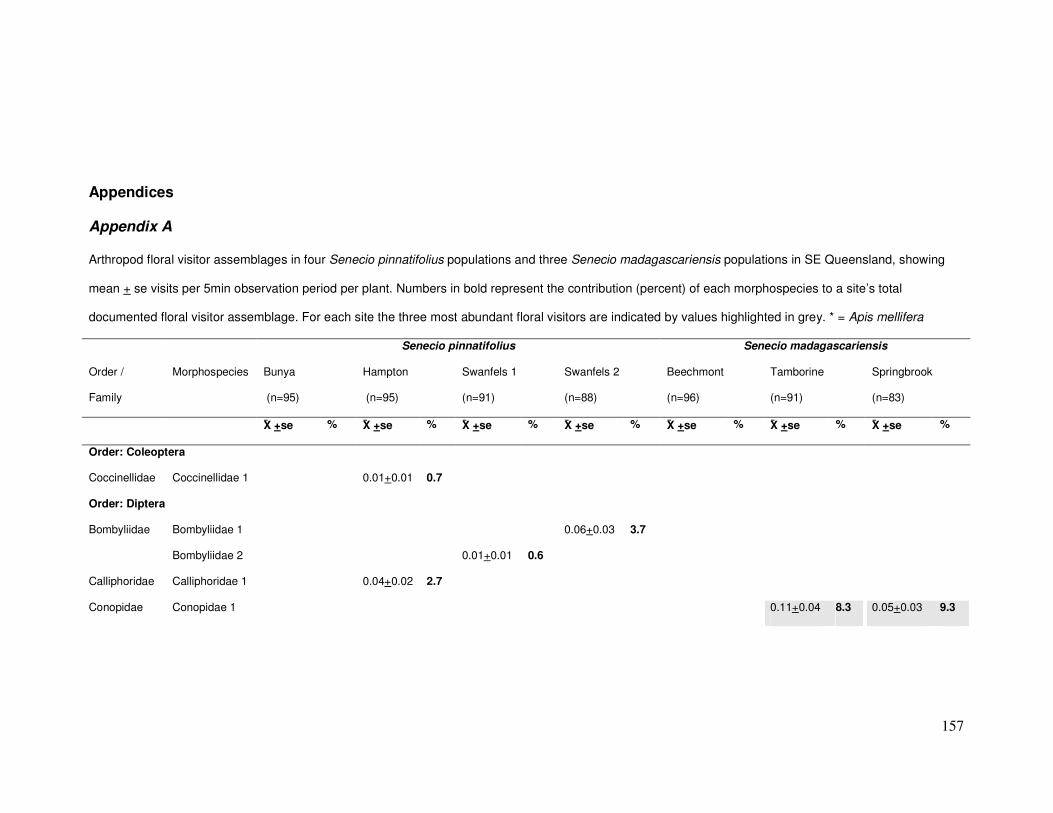
157
Appendices
Appendix A
Arthropod floral visitor assemblages in four Senecio pinnatifolius populations and three Senecio madagascariensis populations in SE Queensland, showing
mean + se visits per 5min observation period per plant. Numbers in bold represent the contribution (percent) of each morphospecies to a site’s total
documented floral visitor assemblage. For each site the three most abundant floral visitors are indicated by values highlighted in grey. * = Apis mellifera
Senecio pinnatifolius Senecio madagascariensis
Order /
Family
Morphospecies Bunya
(n=95)
Hampton
(n=95)
Swanfels 1
(n=91)
Swanfels 2
(n=88)
Beechmont
(n=96)
Tamborine
(n=91)
Springbrook
(n=83)
X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se %
Order: Coleoptera
Coccinellidae Coccinellidae 1 0.01+0.01 0.7
Order: Diptera
Bombyliidae 1 0.06+0.03 3.7 Bombyliidae
Bombyliidae 2 0.01+0.01 0.6
Calliphoridae Calliphoridae 1 0.04+0.02 2.7
Conopidae Conopidae 1 0.11+0.04 8.3 0.05+0.03 9.3
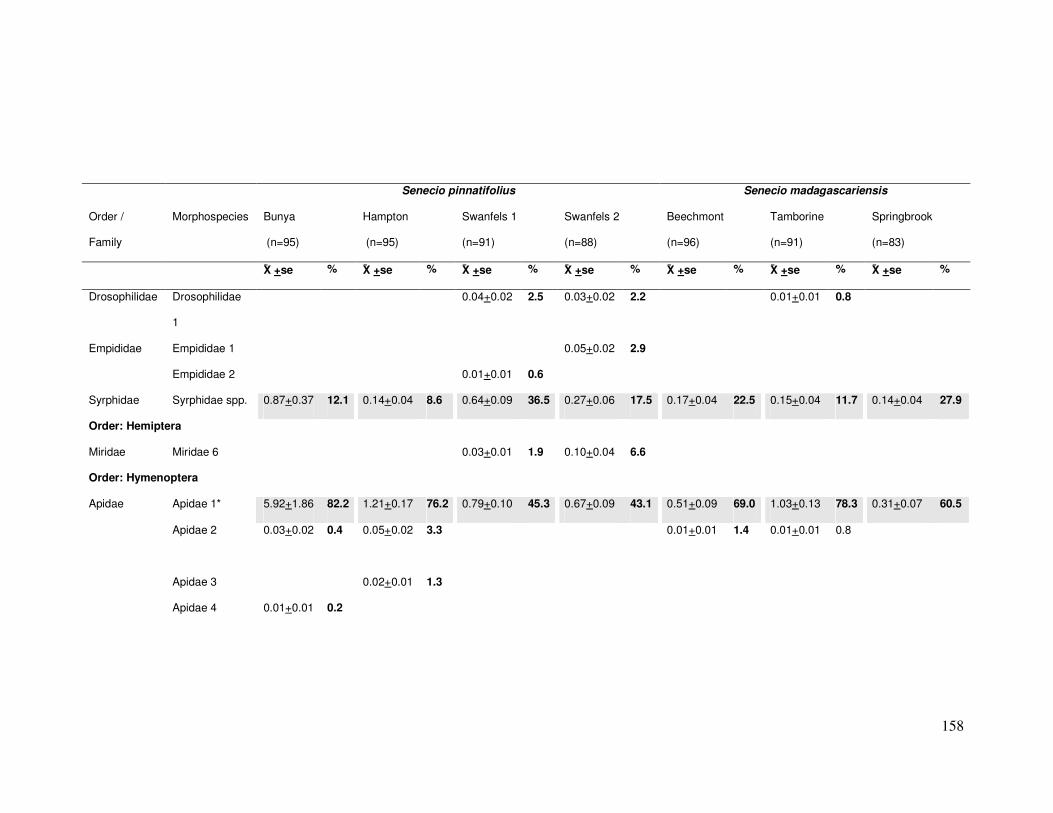
158
Senecio pinnatifolius Senecio madagascariensis
Order /
Family
Morphospecies Bunya
(n=95)
Hampton
(n=95)
Swanfels 1
(n=91)
Swanfels 2
(n=88)
Beechmont
(n=96)
Tamborine
(n=91)
Springbrook
(n=83)
X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se %
Drosophilidae Drosophilidae
1
0.04+0.02 2.5 0.03+0.02 2.2 0.01+0.01 0.8
Empididae 1 0.05+0.02 2.9 Empididae
Empididae 2 0.01+0.01 0.6
Syrphidae Syrphidae spp. 0.87+0.37 12.1 0.14+0.04 8.6 0.64+0.09 36.5 0.27+0.06 17.5 0.17+0.04 22.5 0.15+0.04 11.7 0.14+0.04 27.9
Order: Hemiptera
Miridae Miridae 6 0.03+0.01 1.9 0.10+0.04 6.6
Order: Hymenoptera
Apidae Apidae 1* 5.92+1.86 82.2 1.21+0.17 76.2 0.79+0.10 45.3 0.67+0.09 43.1 0.51+0.09 69.0 1.03+0.13 78.3 0.31+0.07 60.5
Apidae 2 0.03+0.02 0.4 0.05+0.02 3.3 0.01+0.01 1.4 0.01+0.01 0.8
Apidae 3 0.02+0.01 1.3
Apidae 4 0.01+0.01 0.2
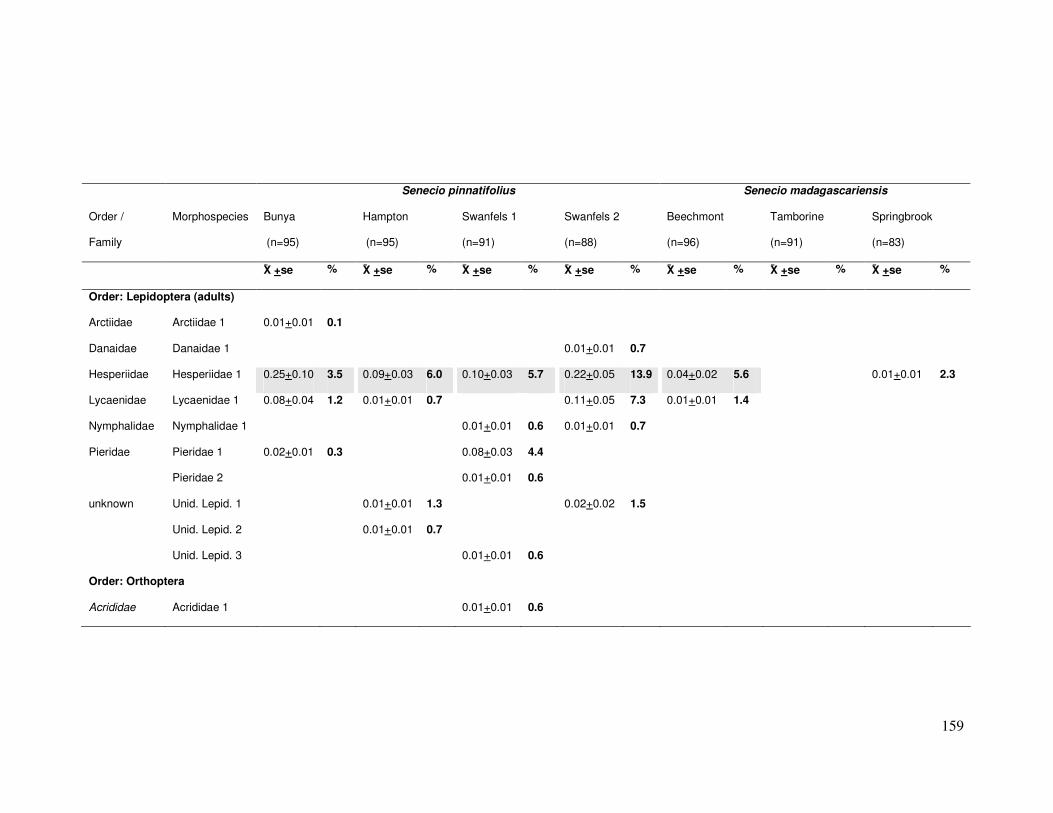
159
Senecio pinnatifolius Senecio madagascariensis
Order /
Family
Morphospecies Bunya
(n=95)
Hampton
(n=95)
Swanfels 1
(n=91)
Swanfels 2
(n=88)
Beechmont
(n=96)
Tamborine
(n=91)
Springbrook
(n=83)
X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se %
Order: Lepidoptera (adults)
Arctiidae Arctiidae 1 0.01+0.01 0.1
Danaidae Danaidae 1 0.01+0.01 0.7
Hesperiidae Hesperiidae 1 0.25+0.10 3.5 0.09+0.03 6.0 0.10+0.03 5.7 0.22+0.05 13.9 0.04+0.02 5.6 0.01+0.01 2.3
Lycaenidae Lycaenidae 1 0.08+0.04 1.2 0.01+0.01 0.7 0.11+0.05 7.3 0.01+0.01 1.4
Nymphalidae Nymphalidae 1 0.01+0.01 0.6 0.01+0.01 0.7
Pieridae 1 0.02+0.01 0.3 0.08+0.03 4.4 Pieridae
Pieridae 2 0.01+0.01 0.6
Unid. Lepid. 1 0.01+0.01 1.3 0.02+0.02 1.5
Unid. Lepid. 2 0.01+0.01 0.7
unknown
Unid. Lepid. 3 0.01+0.01 0.6
Order: Orthoptera
Acrididae Acrididae 1 0.01+0.01 0.6

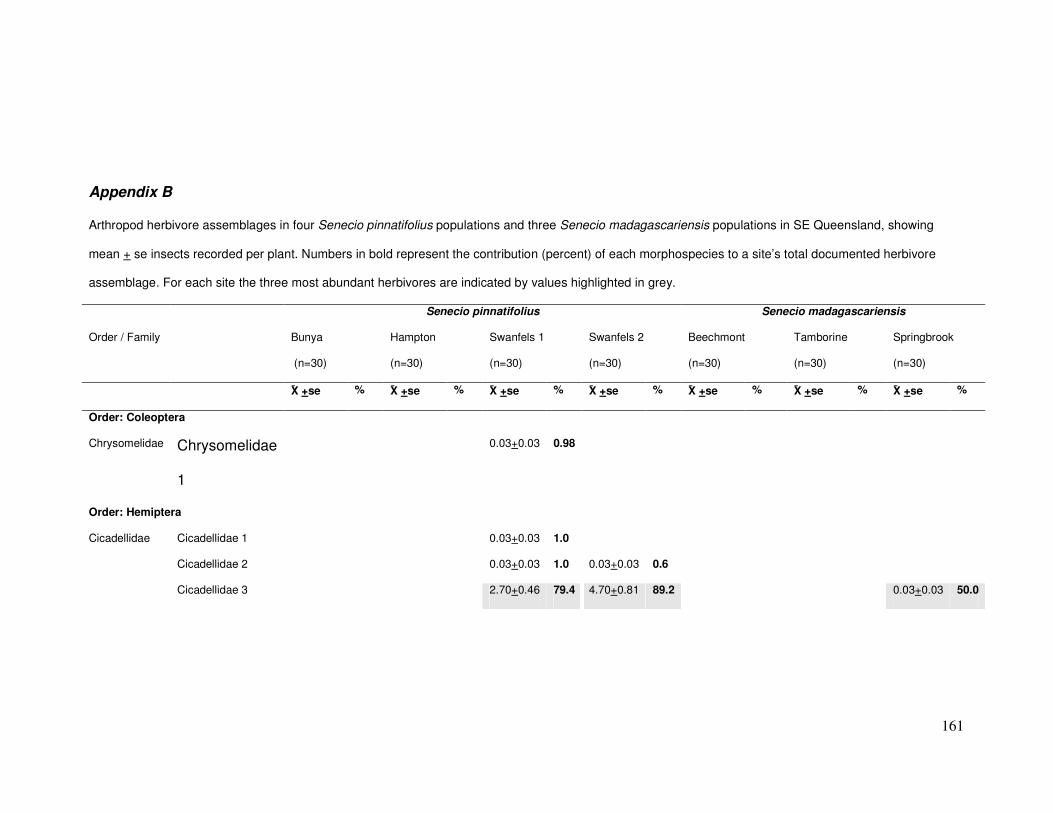
161
Appendix B
Arthropod herbivore assemblages in four Senecio pinnatifolius populations and three Senecio madagascariensis populations in SE Queensland, showing
mean + se insects recorded per plant. Numbers in bold represent the contribution (percent) of each morphospecies to a site’s total documented herbivore
assemblage. For each site the three most abundant herbivores are indicated by values highlighted in grey.
Senecio pinnatifolius Senecio madagascariensis
Order / Family Bunya
(n=30)
Hampton
(n=30)
Swanfels 1
(n=30)
Swanfels 2
(n=30)
Beechmont
(n=30)
Tamborine
(n=30)
Springbrook
(n=30)
X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se %
Order: Coleoptera
Chrysomelidae Chrysomelidae
1
0.03+0.03 0.98
Order: Hemiptera
Cicadellidae 1 0.03+0.03 1.0
Cicadellidae 2 0.03+0.03 1.0 0.03+0.03 0.6
Cicadellidae
Cicadellidae 3 2.70+0.46 79.4 4.70+0.81 89.2 0.03+0.03 50.0
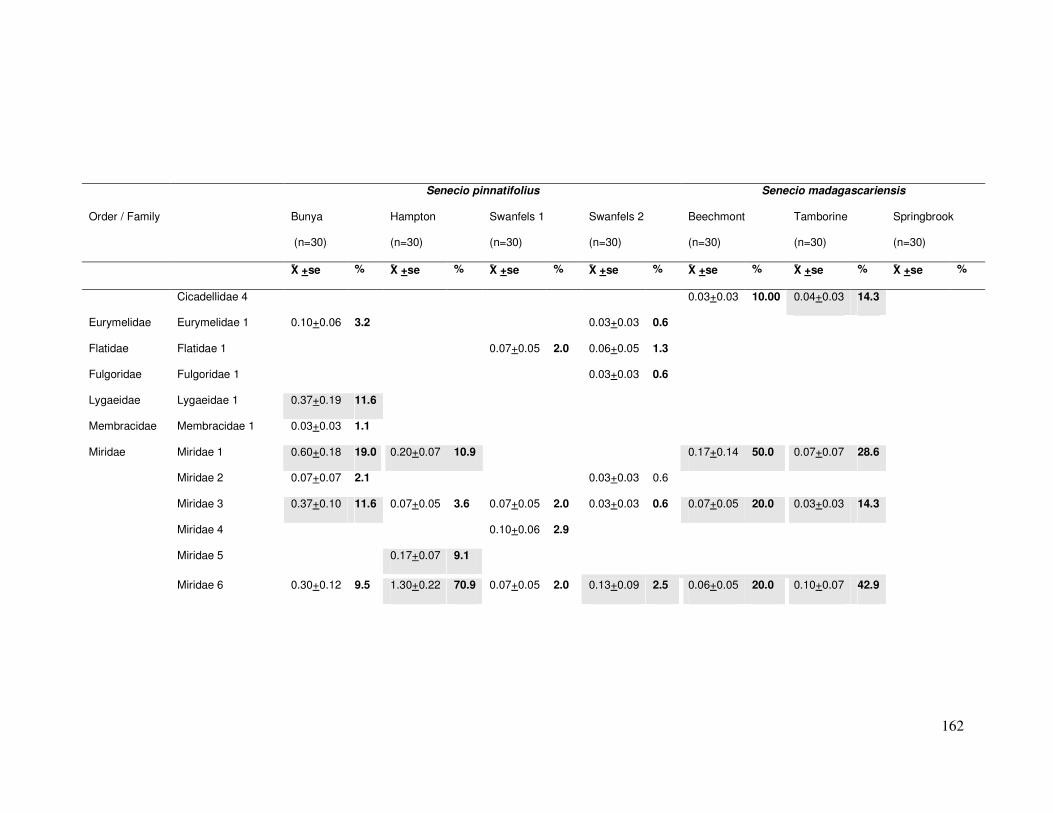
162
Senecio pinnatifolius Senecio madagascariensis
Order / Family Bunya
(n=30)
Hampton
(n=30)
Swanfels 1
(n=30)
Swanfels 2
(n=30)
Beechmont
(n=30)
Tamborine
(n=30)
Springbrook
(n=30)
X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se %
Cicadellidae 4 0.03+0.03 10.00 0.04+0.03 14.3
Eurymelidae Eurymelidae 1 0.10+0.06 3.2 0.03+0.03 0.6
Flatidae Flatidae 1 0.07+0.05 2.0 0.06+0.05 1.3
Fulgoridae Fulgoridae 1 0.03+0.03 0.6
Lygaeidae Lygaeidae 1 0.37+0.19 11.6
Membracidae Membracidae 1 0.03+0.03 1.1
Miridae Miridae 1 0.60+0.18 19.0 0.20+0.07 10.9 0.17+0.14 50.0 0.07+0.07 28.6
Miridae 2 0.07+0.07 2.1 0.03+0.03 0.6
Miridae 3 0.37+0.10 11.6 0.07+0.05 3.6 0.07+0.05 2.0 0.03+0.03 0.6 0.07+0.05 20.0 0.03+0.03 14.3
Miridae 4 0.10+0.06 2.9
Miridae 5 0.17+0.07 9.1
Miridae 6 0.30+0.12 9.5 1.30+0.22 70.9 0.07+0.05 2.0 0.13+0.09 2.5 0.06+0.05 20.0 0.10+0.07 42.9
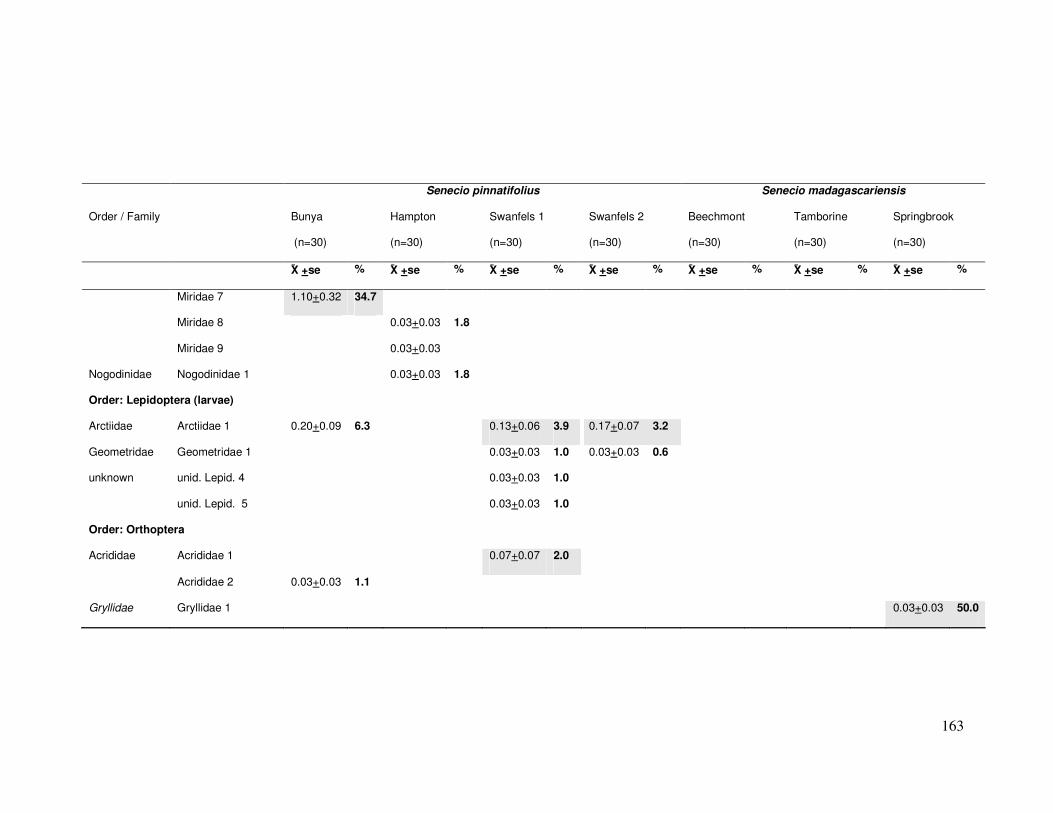
163
Senecio pinnatifolius Senecio madagascariensis
Order / Family Bunya
(n=30)
Hampton
(n=30)
Swanfels 1
(n=30)
Swanfels 2
(n=30)
Beechmont
(n=30)
Tamborine
(n=30)
Springbrook
(n=30)
X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se % X̄ +se %
Miridae 7 1.10+0.32 34.7
Miridae 8 0.03+0.03 1.8
Miridae 9 0.03+0.03
Nogodinidae Nogodinidae 1 0.03+0.03 1.8
Order: Lepidoptera (larvae)
Arctiidae Arctiidae 1 0.20+0.09 6.3 0.13+0.06 3.9 0.17+0.07 3.2
Geometridae Geometridae 1 0.03+0.03 1.0 0.03+0.03 0.6
unid. Lepid. 4 0.03+0.03 1.0 unknown
unid. Lepid. 5 0.03+0.03 1.0
Order: Orthoptera
Acrididae 1 0.07+0.07 2.0 Acrididae
Acrididae 2 0.03+0.03 1.1
Gryllidae Gryllidae 1 0.03+0.03 50.0

164
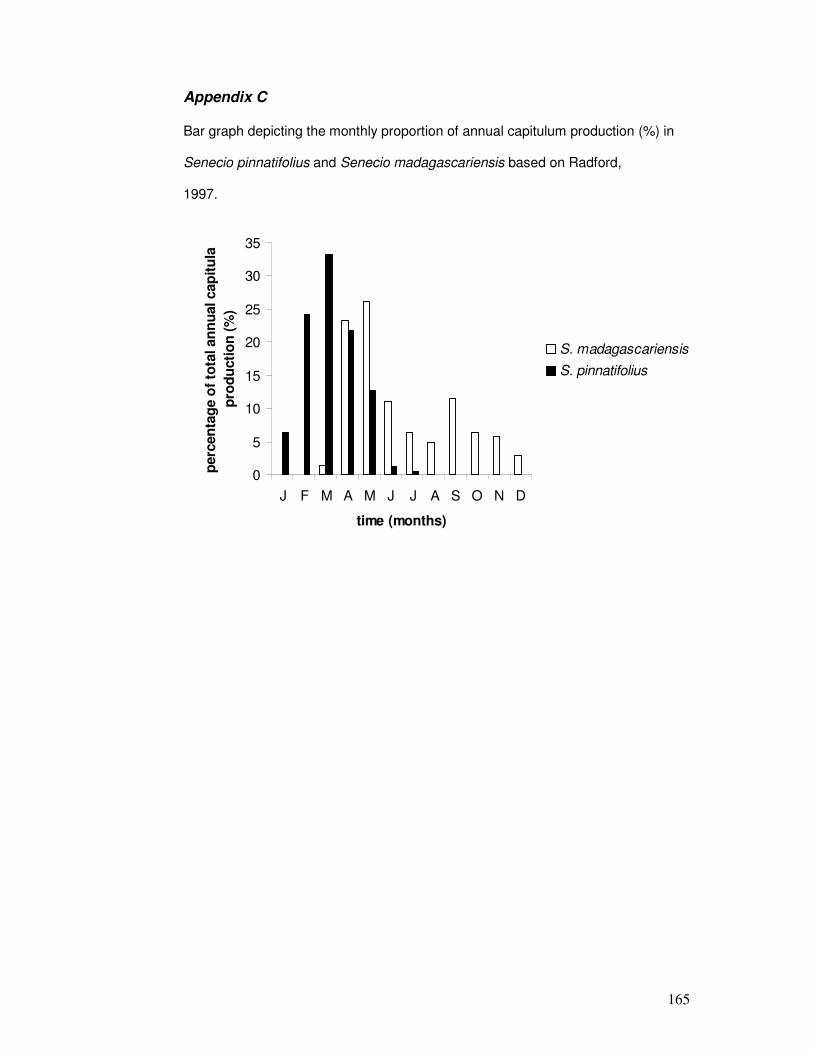
165
Appendix C
Bar graph depicting the monthly proportion of annual capitulum production (%) in
Senecio pinnatifolius and Senecio madagascariensis based on Radford,
1997.
0
5
10
15
20
25
30
35
J F M A M J J A S O N D
time (months)
perc
en
tag
e o
f to
tal an
nu
al cap
itu
la
pro
du
cti
on
(%
)
S. madagascariensis
S. pinnatifolius

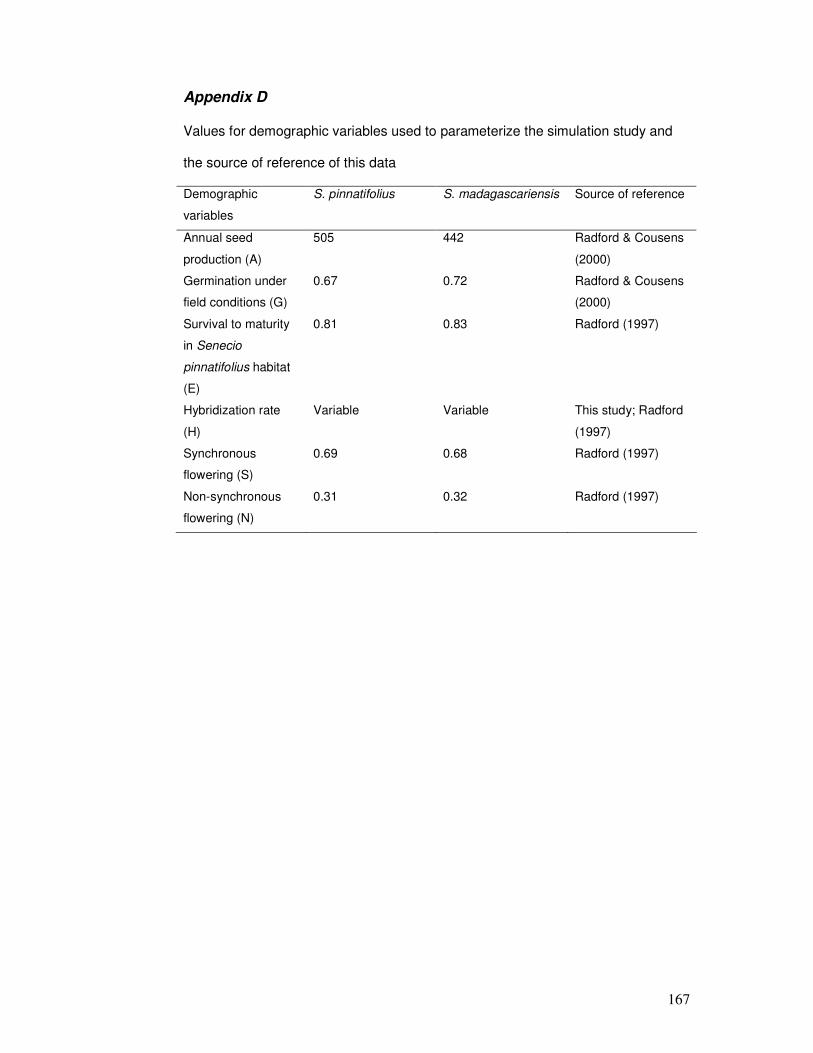
167
Appendix D
Values for demographic variables used to parameterize the simulation study and
the source of reference of this data
Demographic
variables
S. pinnatifolius S. madagascariensis Source of reference
Annual seed
production (A)
505 442 Radford & Cousens
(2000)
Germination under
field conditions (G)
0.67 0.72 Radford & Cousens
(2000)
Survival to maturity
in Senecio
pinnatifolius habitat
(E)
0.81 0.83 Radford (1997)
Hybridization rate
(H)
Variable Variable This study; Radford
(1997)
Synchronous
flowering (S)
0.69 0.68 Radford (1997)
Non-synchronous
flowering (N)
0.31 0.32 Radford (1997)


169
References
Abbott, R.J. (1992) Plant Invasions, interspecific hybridization and the evolution of
new plant taxa. Trends in Ecology and Evolution 7: 401-405.
Abbott, R.J., Ireland, H.E. and Rogers, H.J. (2007) Population decline despite high
genetic diversity in the new allopolyploid species Senecio cambrensis
(Asteraceae). Molecular Ecology 16: 1023–1033.
Abbott, R.J. and Lowe, A.J. (2004) Origins, establishment and evolution of new
polyploid species: Senecio cambrensis and S. eboracensis in the British
Isles. Biological Journal of the Linnean Society 82: 467-474.
Abrams, P.A. (1987) Indirect interactions between species that share a predator:
varieties of indirect effects. In: A. Sih (ed) Predation: direct and indirect
impacts on aquatic communities, pp. 38-54. University Press of New
England, Hanover.
Abrams, P.A. and Masuda, H. (1996) Positive indirect effects between prey species
that share predators. Ecology 77: 610-616.
Adams, M.J., Pearl, C.A. and Bury, R.B. (2003) Indirect facilitation of an anuran
invasion by non-native fishes. Ecology Letters 6: 343-351.
Adler, L.S., Karban, R. and Strauss, S.Y. (2001) Direct and indirect effect of
alkaloids on plant fitness via herbivory and pollination. Ecology 82: 2032-
2044.
Agrawal, A.A. and Kotanen, P.M. (2003) Herbivores and the success of exotic
plants: a phylogenetically controlled experiment. Ecology Letters 6: 712-715.
Agren, J. (1996) Population size, pollinator limitation, and seed set in the self-
incompatible herb Lythrum salicaria. Ecology 77: 1779-1790.
Ali, S.I. (1966) Senecio lautus complex in Australia. III. The genetic system.
Australian Journal of Botany 14: 317-327.

170
Ali (1969) Senecio lautus complex in Australia. V. Taxanomic interpretations.
Australian Journal of Botany 17: 161-176.
Anderson, D. and Panetta, F.D. (1995) Fireweed response to boomspray
applications of different herbicides and adjuvants. Plant Protection Quarterly
10: 152-153.
Anderson, E. (1949) Introgressive Hybridization. John Wiley and Sons, New York.
Anttila, C.K., Daehler, C.C., Rank, N.E. and Strong, D.R. (1998) Greater male
fitness of a rare invader (Spartina alterniflora, Poaceae) threatens a common
native Spartina foliosa) with hybridization. American Journal of Botany 85:
1597-1601.
Arnold, M.L. (1997) Natural Hybridization and Evolution. Oxford University Press,
New York.
Ayres, D.R., Garcia-Rossi, D., Davis, H.G., and Strong, D.R. (1999) Extent and
degree of hybridization between exotic (Spartina alterniflora) and native (S.
floiosa) cordgrass (Poaceae) in California, USA determined by random
amplified polymorphic DNA (RAPDs). Molecular Ecology 8: 1179-1186.
Badenes-Perez F.R., Nault, B.A. and Shelton, A.M. (2005) Manipulating the
attractiveness and suitability of hosts for diamondback moth (Lepidoptera :
Plutellidae). Journal of Economic Entomology 98: 836-844.
Barnes, M.M. (1970) Genesis of a pest: Nysius raphanus and Sisymbrium irio in
vineyards. Journal of Economic Entomology 63: 1462-1463.
Barton, N.H. and Hewitt, G.M. (1989) Adaptation, speciation and hybrid zones.
Nature 341: 497-503.
Bascompte, J., Jordano, P., Melian, C.J. and Olensen, J.M. (2003) The nested
assembly of plant-animal mutualistic networks. Proceedings of the National
Academy of Sciences 100: 9383-9387.

171
Bell, J.M., Karron, J.D. and Mitchell, R.J. (2005) Interspecific competition for
pollination lowers seed production and outcrossing in Mimulus ringens.
Ecology 86: 762-771.
Benrey, B. and Denno, R.F. (1997) The slow-growth-high-mortality hypothesis: a
test using the cabbage butterfly. Ecology 78: 987-999.
Benson, J., Van Drieshe, R.G., Pasquale, A. and Elkington, J. (2003) Introduced
braconid parasitoids and range reduction of a native butterfly in New
England. Biological Control 28: 197-213.
Bleeker, W. (2003) Hybridization and Rorippa austriaca (Brassicaceae) invasion in
Germany. Molecular Ecology 12: 1831-1841.
Blicker, P. S., Olson, B. E. and Wraith, J. M. (2003) Water use and water-use
efficiency of the invasive Centaurea maculosa and three native grasses.
Plant and Soil 254: 371-381.
Bøhn, T. and Amundsen, P. (2001) The competitive edge of an invading specialist.
Ecology 82: 2150-2163.
Bonsall, M.B. and Hassell, M.P. (1997) Apparent competition structures ecological
assemblages. Nature 388: 371-373.
Bowers, M.D. (1984) Iridoid glycosides and host-plant specificity in larvae of the
buckeye butterfly, Junonia coenia (Nymphalidae). Journal of Chemical
Ecology 10: 1567-1577.
Brown, B.J., Mitchell, R.J. and Graham, S.A. (2002) Competition for pollination
between an invasive species (purple loosestrife) and a native congener.
Ecology 83: 2328-2336.
Brown, B.J. and Mitchell, R.J. (2001) Competition for pollination: Effects of pollen of
an invasive plant on seed set of a native congener. Oecologia 129: 43-49.
Brown, D.G. (1994) Beetle folivory increases resource availability and alters plant
invasion in monocultures of goldenrod. Ecology 75: 1673-1683.

172
Brown, J.M., Abrahamson, W.G., Packer, R.A. and Way, P.A. (1995) The role of
natural-enemy escape in a gallmaker host-plant shift. Oecologia 104: 52-60.
Bruno, J.F., Stachowicz, J.J. and Bertness, M.D. (2003) Inclusion of facilitation into
ecological theory. Trends in Ecology and Evolution 18: 119-125.
Buggs, R.J.A. and Pannell, J.R. (2006) Rapid displacement of a monoecious plant
lineage is due to pollen swamping by a dioecious relative. Current Biology 16:
996-1000.
Bryce, J., Johnson, P.J. and Macdonald, D.W. (2002) Can niche use in red and
grey squirrels offer clues for their apparent coexistence? Journal of Applied
Ecology 39: 875-887.
Burgess, K.S., Morgan, M., DeVerno, L. and Husband, B.C. (2005) Asymmetrical
introgression between two Morus species (M. alba, M. rubra) that differ in
abundance. Molecular Ecology 14: 3471-3483.
Burki, C. and Nentwig, W. (1997) Comparison of herbivore insect communities of
Heracleum sphondylium and H. mantegazzianum in Switzerland
(Spermophyta: Apiaceae). Entomologia Generalis 22: 147-155.
Byers, J.E. (2000a) Competition between two estuarine snails: implications for
invasions of exotic species. Ecology 81: 1225-1239.
Byers, J.E. (2000b) Effects of body size and resource availability on dispersal in a
native and non-native estuarine snail. Journal of Experimental Marine Biology
and Ecology 248:133-150.
Byers, J.E., Reichard, S., Randall, J.M., Parker, I.M., Smith, C.S., Lonsdale, W.M.,
Atkinson, I.A.E., Seastedt, T.R., Williamson, M., Chornesky, E., and Hayes,
D. (2002) Directing research to reduce the impacts of nonindigenous species.
Conservation Biology 16: 630-640.
Cadi, A. and Joly, P. (2003) Competition for basking places between the
endangered European pond turtle (Emys orbicularis galloitalica) and the

173
introduced red-eared slider (Trachemys scripta elegans). Canadian Journal
of Zoology 81: 1392-1398.
Callaway, R.M. and Aschehoug, E.T. (2000) Invasive plants versus their new and
old neighbours: a mechanism for exotic invasion. Science 290: 521-523.
Callaway, R.M., DeLuca, T.H. and Belliveau, W.M. (1999) Biological control
herbivores may increase competitive ability of the noxious weed Centaura
maculosa. Ecology 80: 1196-1201.
Callaway, R.M. and Pennings, S.C. (2000) Facilitation may buffer competitive
effects: indirect and diffuse interactions among salt marsh plants. The
American Naturalist 156: 416-424.
Campbell, D., Duchesne, P. and Bernatchez, L. (2003) AFLP utility for population
assignment studies: analytical investigation and empirical comparison with
microsatellites. Molecular Ecology 12: 1979-1991.
Campbell, D.R. (1985) Pollinator sharing and seed set of Stellaria pubera:
Competition for pollination. Ecology 66: 544-553.
Campbell, D.R. and Motten, A.F. (1985) The mechanism of competition for
pollination between two forest herbs. Ecology 66: 554-563.
Carson, W.P. and Root, R.B. (2000) Herbivory and plant species coexistence:
community regulation by an outbreaking phytophagous insect. Ecological
Monographs 70: 73-99.
Centre, T.D., Van, T.K., Dray Jr., F.A., Franks, S.J., Rebelo, T., Pratt, P.D., and
Rayamajhi, M.B. (2005) Herbivory alters competitive interactions between
two invasive aquatic plants. Biological Control 35: 115-123.
Chapin, F.S. III, Bloom, A.J., Field, C.B. and Waring, R.H. (1987) Plant responses
to multiple environmental factors. Bioscience 37: 49-57.
Chen, Y., Lin, L., Wang, C., Yeh, C. and Hwang, S. (2004) Response of two Pieris
(Lepidoptera: Pieridae) species to fertilization of a host plant. Zoological
Studies 43: 778-786.

174
Cheng, X. and Xu, R. (2003) Perspectives on apparent competition in insects. Acta
Entomologica Sinica 46: 237-243.
Chittka, L. and Schurkens, S. (2001) Successful invasion of a floral market. Nature
411: 653.
Chornesky, E.A. and Randall, J.M. (2003) The threat of invasive alien species to
biological diversity: setting a future course. Annals of the Missouri Botanical
Garden 90: 67-76.
Colautti, R.I., Ricciardi, A., Grigorovich, I.A., and MacIsaac, H.J. (2004) Is invasion
success explained by the enemy release hypothesis? Ecology Letters 7: 721-
733.
Common, I.F.B (1993) Moths of Australia, Melbourne University Press, Melbourne.
Connor, E.F., Faeth, S.H., Simberloff, D. and Opler, P.A. (1980) Taxonomic
isolation and the accumulation of herbivorous insects: a comparison of
introduced and native trees. Ecological Entomology 5: 205-211.
Constible, J.M., Sweitzer, R.A., Van Vuren, D.H., Schuyler, P.T. and Knapp, D.A.
(2005) Dispersal of non-native plants by introduced bison in an island
ecosystem. Biological Invasions 7: 699-709.
Corbin, J. D. and D'Antonio, C.M. (2004) Competition between native perennial
and exotic annual grasses: Implications for an historical invasion. Ecology 85:
1273-1283.
Courchamp, F., Langlais, M., and Sugihara, G. (2000) Rabbits killing birds:
modelling the hyperpredation process. Journal of Animal Ecology 69: 154-
164.
Courtney, S.P., Chen, G.K. and Gardner, A. (1989) A general model for individual
host selection. Oikos 55: 55-65.
Crawley, M.J. (1987). What makes a community invasible? In: A.J. Gray, M.J.
Crawley and P.J. Edwards (eds) Colonization, Succession and Stability. The
26th Symposium of The British Ecological Society held jointly with The

175
Linnean Society of London, pp 429-453. Blackwell Scientific Publications,
Oxford.
Crawley, M.J. and Gillman, M.P. (1989) Population dynamics of cinnabar moth and
ragwort in grassland. Journal of Animal Ecology 58: 1035-1050.
Crooks, J.A. (2002) Characterizing ecosystem-level consequences of biological
invasions: the role of ecosystem engineers. Oikos 97: 153-166.
Cross, J.R. (1981) The establishment of Rhododendron ponticum in the Killarney
oakwoods, S.W. Ireland. Journal of Ecology 69: 807-824.
Cruzan, M.B. (2005) Patterns of introgression across an expanding hybrid zone:
analysing historical patterns of gene flow using nonequilibrium approaches.
New Phytologist 167: 267-278.
Csurhes, S.M. (1995). List of declared and non-declared plant species considered
to pose a serious threat to native bushland and/or wetlands in Queensland.
Internal report, Land Protection Branch, Queensland Department of Natural
Resources, Brisbane.
Cunningham, J.P. and West, S.A. (2001) Host selection in phytophagous insects: a
new explanation for learning in adults. Oikos 95: 537-543.
Darwin, C. (1859) The Origin of Species by Means of Natural Selection. John
Murray, London.
Davis, M.A. and Thompson, K. (2000) Eight ways to be a colonizer; two ways to be
an invader: A proposed nomenclature scheme for invasion ecology. Bulletin
of the Ecological Society of America 81: 226-230.
Davis, M.A. and Thompson, K. (2001) Invasion terminology: should ecologists
define their terms differently than others? No, not if we want to be of any
help! Bulletin of the Ecological Society of America 82:206.
Dickman, C. (1996) Overview of the impacts of feral cats on Australian native
fauna. Australian Nature Conservation Agency, Canberra.

176
Dietz, H., Wirth, L.R., and Buschmann, H. (2004) Variation in herbivore damage to
invasive and native woody plant species in open forest vegetation on Mahe,
Seychelles. Biological Invasions 6: 511-521.
Doyle, J.J. and Doyle, J.L. (1987) A rapid DNA isolation procedure for small
quantities of fresh leaf tissue. Phytochemical Bulletin 19: 11-15.
Duchesne, P. and Bernatchez, L. (2002) AFLPOP: a computer program for
simulated and real population allocation, based on AFLP data. Molecular
Ecology Notes 2: 380-383.
Edwards, G.R., Bourdot, G.W. and Crawley, M.J. (2000) Influence of herbivory,
competition and soil fertility on the abundance of Cirsium arvense in acid
grassland. Journal of Applied Ecology 37: 321-334.
Eguchi, K. and Amano, H.E. (1999) Naturalisation of exotic birds in Japan.
Japanese Journal of Ornithology 47: 97-114.
Elkington, J.S. and Liebhold, A.M. (1990) Population dynamics of gypsy moth in
North America. Annual Review of Entomology 35: 571-596.
Ellstrand, N.C. and Schierenbeck, K.A. (2000) Hybridisation as a stimulus for the
evolution of invasiveness in plants? Proceedings of the National Academy of
Sciences USA 97: 7043-7050.
Elton, C.S. (1958) The Ecology of Invasion by Plants and Animals. Chapman and
Hall, London.
Erickson, D.L. and Fenster, C.B. (2006) Intraspecific hybridization and the recovery
of fitness in the native legume Chamaechrista fascaculata. Evolution 60:
225-233.
Fehmi, J.S., Rice, K.J. and Laca, E.A. (2004) Radial dispersion of neighbours and
the small-scale competitive impact of two annual grasses on a native
perennial grass. Restoration Ecology 12: 63-69.
Feldman, T.S., Morris, W.F. and Wilson, W.G. (2004) When can two plant species
facilitate each other's pollination? Oikos 105:197-207.

177
Felsenstein, J. (2005) PHYLIP (Phylogeny Inference Package) version 3.6.
Distributed by the author. Department of Genome Sciences, University of
Washington, Seattle.
Fernández, O.N. and Verona, C.A. (1984) Caracteristicas reproductivas de
Senecio madagascariensis Poiret (Compositae). Revista de la Facultad de
Agronomia de la Universidad de Buenos Aires 5: 125-137.
Figueredo, C.C. and Giani, A. (2005) Ecological interactions between Nile tilapia
(Oreochromis niloticus, L.) and the phytoplanktonic community of the Furnas
Reservoir (Brazil). Freshwater Biology 50: 1391-1403.
Flecker, A.S. and Townsend, C.R. (1994) Community-wide consequences of trout
introduction in New Zealand streams. Ecological Applications 4: 798-807.
Flecker, A.S. and Townsend, C.R. (1996) Interactions between fish, grazing
invertebrates and algae in a New Zealand stream: a trophic cascade
mediated by fish-induced changes to grazer behaviour? Oecologia 108: 174-
181.
Fogarty, G. and Facelli, J.M. (1999) Growth and competition of Cytisus scoparius,
an invasive shrub, and Australian native shrubs. Plant Ecology 144: 27-35.
Foss, L.K. and Rieske, L.K. (2003) Species-specific differences in oak foliage affect
preference and performance of gypsy moth caterpillars. Entomologia
Experimentalis et Applicata 108: 87-93.
Fowler, J. and Cohen, L. (1990) Practical Statistics for Field Biology. John Wiley
and Sons, Chichester.
Free, J.B. (1963) The flower constancy of honeybees. Journal of Animal Ecology
32: 119-131.
Frenzel, M. and Brandl, R. (2003) Diversity and abundance patterns of
phytophagous insect communities on alien and native host plants in the
Brassicaceae. Ecography 26: 723-730.

178
Galen, C. and Gregory, T. (1989) Interspecific pollen transfer as a mechanism of
competition: consequences of foreign pollen contamination for seed set in the
alpine wildflower, Polemonium viscosum. Oecologia 81: 120-123.
Gamboa, G.J., Noble, M.A., Thom, M.C., Togal, J.L., Srinivasan, R. and Murphy,
B.D. (2004) The comparative biology of two sympatric paper wasps in
Michigan, the native Polistes fuscatus and the invasive Polistes dominulus
(Hymenoptera, Vespidae). Insectos Sociales 51: 153-157.
Garcia-Serrano, H., Escarre, J. and Sans, F.X. (2004) Factors that limit the
emergence and establishment of the related Senecio inaequidens and
Senecio pterophorus and the native Senecio malacitanus in Mediterranean
climate. Canadian Journal of Botany 82: 1346-1355.
Garcia-Serrano, H., Escarre, J., Garnier, E. and Sans, F.X. (2005) A comparative
growth analysis between alien invader and native Senecio species with
distinct distribution ranges. Ecoscience 12: 35-43.
Garcia-Serrano, H., Sans, F.X. and Escarre, J. (2007) Interspecific competition
between alien and native congeneric species. Acta Oecologica 31: 69-78.
Geddes, P.S., Le Blanc, J.P.R., Yule, W.N. (1992) Abiotic and biotic factors
affecting Rhagoletis mendax [Diptera: Tephritidae] populations in eastern
Canadian lowbush blueberry fields. Phytoprotection 73: 73-78.
Ghazoul, J. (2002) Flowers at the frontline of invasion? Ecological Entomology 27:
638-640.
Ghazoul, J. (2004) Alien abduction: disruption of native plant-pollinator interactions
by invasive species. Biotropica 36: 156-164.
Goulson, D. and Derwent, L.C. (2004) Synergistic interactions between an exotic
honeybee and an exotic weed: pollination of Lantana camara in Australia.
Weed Research 44: 195-202.

179
Grabas, G.P. and Laverty, T.M. (1999) The effect of purple loosestrife (Lythrum
salicaria L.; Lythraceae) on the pollination and reproductive success of
sympatric co-flowering wetland plants. Ecoscience 6: 230-242.
Grosholz, E.D. (2005) Recent biological invasion may hasten invasional meltdown
by accelerating historical introductions. Proceedings of the National Academy
of Sciences of the United States of America 102: 1088-1091.
Grosholz, E.D., Ruiz, G.M., Dean, C.A., Shirley, K.A., Maron, J.L. and Connors, P.G.
(2000) The impacts of a nonindigenous marine predator in a California bay.
Ecology 81: 1206-1224.
Gross, C.L. (2001) The effect of introduced honeybees on native bee visitation and
fruit-set in Dillwynia juniperina (Fabaceae) in a fragmented ecosystem.
Biological Conservation 102: 89-95.
Gross, K.L. and Werner, P.A. (1983) Relationships among flowering phenology,
insect visitors, and seed set of individuals: Experimental studies on four co-
occurring species of goldenrod (Solidago: Compositae). Ecological
Monographs 53: 95-117.
Hämback, P.A. and Ekerholm, P. (1997) Mechanisms of apparent competition in
seasonal environments: an example with vole herbivory. Oikos 80: 276-288.
Hanfling, B. and Kollmann, J. (2002) An evolutionary perspective of biological
invasions. Trends in Ecology and Evolution 17: 545-557.
Hanley, M.E. and Goulson, D. (2003) Introduced weeds pollinated by introduced
bees: Cause or effect? Weed Biology and Management 3: 204-212.
Hardy, O.J., De Loose, M., Vekemans, X. and Meerts, P. (2001) Allozyme
segregation and inter–cytotype reproductive barriers in the polyploid
complex Centaurea jacea. Heredity 87: 136-145.
Hardy, O.J., Vanderhoeven, S., De Loose, M. and Meerts, P. (2000) Ecological,
morphological and allozymic differentiation between diploid and tetraploid

180
knapweeds (Centaurea jacea) from a contact zone in the Belgian Ardennes.
New Phytologist 146: 281-290.
Hardy, O.J. and Vekemans, X. (2002) SPAGEDi: a versatile computer program to
analyse spatial genetic structure at the individual or population levels.
Molecular Ecology Notes 2: 618-620.
He, T.H., Krauss, S.L., Lamont, B.B., Miller, B.P. and Enright, N.J. (2004) Long-
distance seed dispersal in a metapopulation of Banksia hookeriana inferred
from a population allocation analysis of amplified fragment length
polymorphism data. Molecular Ecology 13: 1099-1109.
Hegarty, M.J. and Hiscock, S. (2005) Hybrid speciation in plants: new insights from
molecular studies. New Phytologist 165: 411–423.
Herrera, C.M. (2005) Plant generalization on pollinators: species property or local
phenomenon? American Journal of Botany 92: 13-20.
Hersch, E.I. (2006) Foliar damage to parental plants interacts to influence mating
success of Ipomoea purpurea. Ecology 87:2026–2036.
Hersch, E.I. and Roy, B.A. (2007) Context-dependent pollinator behavior: an
explanation for patterns of hybridization among three species of indian
paintbrush. Evolution: 111-124.
Hill, A.M. and Lodge, D.M. (1999) Replacement of resident crayfishes by an exotic
crayfish: the roles of competition and predation. Ecological Applications 9:
678-690.
Holm, L., Doll, J., Holm, E., Pancho, J. and Herberger, J. (1997) World Weeds.
Natural Histories and Distribution. Wiley, New York.
Holm, S.N. (1966) The utilization and management of bumble bees for red clover
and alfalfa seed production. Annual Review of Entomology 11: 155-182.
Holt, R.D. (1977) Predation, apparent competition, and the structure of prey
communities. Theoretical Population Biology 12: 197-229.

181
Holt, R.D. (1984) Spatial heterogeneity, indirect interactions, and the coexistence
of prey species. The American Naturalist 124: 377-406.
Holt, R.D. and Barfield, M. (2003) Impacts of temporal variation on apparent
competition and coexistence in open ecosystems. Oikos 101: 49-58.
Holt, R.D. and Kotler, B.P. (1987) Short-term apparent competition. The American
Naturalist 130: 412-430.
Holtkamp, R.H. and Hosking, J.R. (1993) Insects and diseases of fireweed,
Senecio madagascariensis, and the closely related Senecio lautus complex.
In: J.T. Swarbrick, C.W.L. Henderson, R.J. Jettnre, L. Streit and S.R. Walker
(eds) Tenth Australian and 14th Asian Pacific Weed Science Society
Conference, pp. 130-132. Weed Society of Queensland, Brisbane.
Hoogendoorn, M. and Heimpel, G.E. (2002) Indirect interactions between an
introduced and a native ladybird species mediated by a shared parasitoid.
Biological Control 25: 224-230.
Hoskin, C.J., Higgie, M., McDonald, K.R. and Moritz, C. (2005) Reinforcement
drives rapid allopatric speciation. Nature 437: 1553-1556.
Hrabik, T.R., Carey, M.P. and Webster, M.S. (2001) Interactions between young-
of-the-year exotic rainbow smelt and native yellow perch in a northern
temperate lake. Transactions of the American Fisheries Society 130: 568-
582.
Husband, B.C. (2004) The role of triploid hybrids in the evolutionary dynamics of
mixed ploidy populations. Biological Journal of the Linnean Society 82: 537-
546.
Huxel, G.R. (1999) Rapid displacement of native species by invasive species:
effects of hybridization. Biological Conservation 89: 143-152.
Jensen, G.C., McDonald, P. S. and Armstrong, D.A. (2002) East meets west:
Competitive interactions between green crab Carcinus maenas, and native

182
and introduced shore crab Hemigrapsus spp. Marine Ecology Progress
Series, 225, 251-262.
Jesse, L.C., Moloney, K.A. and Obrycki, J.J. (2006) Insect pollinators of the
invasive plant, Rosa multiflora (Rosaceae), in Iowa, USA. Weed Biology and
Management 6: 235-240.
Juenger, T. and Bergelson, J. (1997) Pollen and resource limitation of
compensation to herbivory in Scarlet Gilia, Ipomopsis aggregata. Ecology 78:
1684-1695.
Kadereit, J.W., Uribe-Convers, S., Westberg, E. and Comes, H.P. (2006)
Reciprocal hybridization at different times between Senecio flavus and
Senecio glaucus gave rise to two polyploid species in north Africa and
south-west Asia. New Phytologist 169: 431-441.
Kane, D.D., Haas, E.M. and Culver, D.A. (2003) The characteristics and potential
ecological effects of the exotic crustacean zooplankter Cercopagis pengoi
(Cladocera: Cercopagidae), a recent invader of Lake Erie. Ohio Journal of
Science 103: 79-83.
Karban, R. (1997) Neighbourhood affects a plant’s risk of herbivory and
subsequent success. Ecological Entomology 22: 433-439.
Keane, R.M. and Crawley, M.J. (2002) Exotic plant invasions and the enemy
release hypothesis. Trends in Ecology and Evolution 17: 164-170.
Kephart, S.R. (1983) The partitioning of pollinators among three species of
Asclepias. Ecology 64: 120-133.
Khan, T.A., Wilson, M.E. and Khan, M.T. (2003) Evidence for invasive carp
mediated trophic cascade in shallow lakes of western Victoria, Australia.
Hydrobiologia, 506-509: 465-472.
Kido, M. H., Heacock, D.E. and Asquith, A. (1999) Alien rainbow trout
(Oncorhynchus mykiss) (Salmoniformes: Salmonidae) diet in Hawaiian
streams. Pacific Science 53: 242-251.

183
Kiesecker, J.M., Blaustein, A.R., and Miller, C.L. (2001) Transfer of a pathogen
from fish to amphibians. Conservation Biology 15: 1064-1070.
Kinnear, J. E., Sumner, N. R. and Onus, M. L. (2002) The red fox in Australia: An
exotic predator turned biocontrol agent. Biological Conservation 108: 335-
359.
Kinzler, W. and Maier, G. (2003) Asymmetry in mutual predation: Possible reason
for the replacement of native gammarids by invasives. Archiv fuer
Hydrobiologie, 157: 473-481.
Kirchner, F., Luijten, S.H., Imbert, E., Riba, M., Mayol, M., Gonzalez-Martinez,
S.C., Mignot, A. and Colas, B. (2005) Effects of local density on insect
visitation and fertilization success in the narrow-endemic Centaurea
corymbosa (Asteraceae). Oikos 111: 130-142.
Kolb, A. and Alpert, P. (2003) Effects of nitrogen and salinity on growth and
competition between a native grass and an invasive congener. Biological
Invasions 5: 229-238.
Krebs, C.J. (1989) Ecological Methodology. Harper Collins, New York.
Ladner, D.T. and Altizer, S. (2005) Oviposition preference and larval performance
of North American monarch butterflies on four Asclepias species.
Entomologia Experimentalis et Applicata 116: 9-20.
Lagercrantz, U. and Ryman, N. (1990) Genetic structure of Norway spruce (Picea
abies): concordance of morphological and allozymic variation. Evolution 44:
38-53.
Lamont, B.B., Klinkhamer, P.G.L. and Witkowski, E.T.F. (1993) Population
fragmentation may reduce fertility to zero in Banksia goodie – a
demonstration of the Allee effect. Oecologia 94: 446-450.
Lancau, R.A., Rogers, W.E. and Siemann, E. (2004) Constraints on the utilisation
of the invasive Chinese tallow tree Sapium sebiferum by generalist native
herbivores in coastal prairies. Ecological Entomology 29: 66-75.

184
Landwer, A.J. and Ferguson, G. W. (2002) Long-term structural habitat use of male
individuals of two native and one introduced Anolis (Iguanidae) species on
the north coast of Jamaica. Texas Journal of Science 54: 51-58.
Larson, K.C., Fowler, S.P. and Walker, J.C. (2002) Lack of pollinators limits fruit set
in the exotic Lonicera japonica. American Midland Naturalist 148: 54-60.
Lau, J.A. and Strauss, S.Y. (2005) Insect herbivores drive important indirect effects
of exotic plants on native communities. Ecology 86: 2990-2997.
Lavergne, S., Debussche, M., and Thompson, J.D. (2005) Limitations on
reproductive success in endemic Aquiegia viscosa (Ranunculaceae) relative
to its widespread congener Aquilegia vulgaris: the interplay of herbivory and
pollination. Oecologia 142: 212-220.
Lavorel, S., Prieur-Richard, A.H. and Grigulis, K. (1999) Invasibility and diversity of
plant communities: From patterns to processes. Diversity and Distributions 5:
41-49.
Lawrence, M.E. (1985) Senecio L. (Asteraceae) in Australia: reproductive biology
of a genus found primarily in unstable environments. Australian Journal of
Botany 33: 197-208.
Laxson, C.L., McPhedran, K.N., Makarewicz, J.C., Telesh, I.V. and MacIsaac, H.J.
(2003) Effects of the non-indigenous cladoceran Cercopagis pengoi on the
lower food web of Lake Ontario. Freshwater Biology 48: 2094-2106.
Leather, S.R., Beare, J.A., Cooke, R.C.A. and Fellowes, M.D.E. (1998) Are
differences in life history parameters of the pine beauty moth Panolis
flammea modified by host plant quality or gender? Entomologia
Experimentalis et Applicata 87: 237-243.
Leimu, R., Riipi, M. and Staerk, D. (2005) Food preference and performance of the
larvae of a specialist herbivore: variation among and within host-plant
populations. Acta Oecologica 28: 325-330.

185
Lenz, L. and Taylor, J.A. (2001) The influence of an invasive tree species (Myrica
faya) on the abundance of an alien insect (Sophonia rufofascia) in Hawai’i
Volcanoes National Park. Biological Conservation 102: 301-307.
Lenz, T.I., Moyle-Croft, J.L. and Facelli, J.M. (2003) Direct and indirect effects of
exotic annual grasses on species composition of a South Australian
grassland. Austral Ecology 28: 23-32.
Le Roux, J.J., Wieczorek, A.M., Ramadan, M.M. and Tran, C.T. (2006) Resolving
the native provenance of invasive fireweed (Senecio madagascariensis Poir.)
in the Hawaiian Islands as inferred from phylogenetic analysis. Diversity and
Distributions 12: 694-702.
Levin, D.A. (1995) Metapopulations: An arena for local speciation. Journal of
Evolutionary Biology 8: 635-644.
Levin, D.A. and Anderson, W.W. (1970) Competition for pollinators between
simultaneously flowering plant species. The American Naturalist 104: 455-
467.
Levin, D.A., Francisco-Ortega, J. and Jansen, R.K. (1996) Hybridization and the
extinction of rare plant species. Conservation Biology 10: 10-16.
Levin, P.S., Coyer, J.A., Petrik, R. and Good, T.P. (2002) Community-wide effects
of nonindigenous species on temperate rocky reefs. Ecology 83: 3182-3193.
Levine, J.M. (1999) Indirect facilitation: evidence and predictions from a riparian
community. Ecology 80: 1762-1769.
Levine, J.M., Adler, P.B., and Yelenik, S.G. (2004) A meta-analysis of biotic
resistance to exotic plant invasions. Ecology Letters 7: 975-989.
Levine, J.M., Vilà, M., D’Antonio, C.M., Dukes, J.S., Grigulis, K. and Lavorel, S.
(2003) Mechanisms underlying the impacts of exotic plant invasions.
Proceedings of the Royal Society of London B 270: 775-781.
Levine, S. (1980) Indirect mutualism: variations on a theme. The American
Naturalist 116: 441-448.

186
Lill, J.T. and Marquis, RJ. (2001) The effects of leaf quality on herbivore
performance and attack from natural enemies. Oecologia 126: 418-428.
Liu, H, Pemberton, R.W. and Stiling, P. (2006) Native and introduced pollinators
promote a self-incompatible invasive woody vine (Paederia foetida L) in
Florida. Journal of the Torrey Botanical Society 133: 304-311.
LoGuidice, K. (2003) Trophically transmitted parasites and the conservation of
small populations: Raccoon roundworm and the imperilled allegheny
woodrat. Conservation Biology 17: 258-266.
Lombadero, M.J., Ayres, M.P., Hofstetter, R.W., Moser, J.C. and Lepzig, K.D.
(2003) Strong indirect interactions of Tarsonemus mites (Acarina:
Tarsonemidae) and Dendroctonus frontalis (Coleoptera: Scolytidae). Oikos
102: 243-252.
Lorenzoni, M., Corboli, M., Dorr, A. J. M., Giovinazzo, G., Selvi, S. and Mearelli, M.
(2002) Diets of Micropterus salmoides Lac. and Esox lucius L. in Lake
Trasimeno (Umbria, Italy) and their diet overlap. Bulletin Francais de la
Peche et de la Pisciculture, 365-366: 537-547.
Lortie, C.J., Brooker, R.W., Choler, P., Kikvidze, Z., Michalet, R., Pugnaire, F.I. and
Callaway, R.M. (2004) Rethinking plant community theory. Oikos 107: 433-
438.
Louda, S.M. and Potvin, M.A. (1995) Effect of inflorescence-feeding insects on the
demography and lifetime fitness of a native plant. Ecology 76: 229-45.
Louda, S.M., Rand, T.A., Arnett, A.E., McClay, A.S., Shea, K., and McEachern,
A.K. (2005) Evaluation-of ecological risk to populations of a threatened plant
from an invasive biocontrol insect. Ecological Applications 15: 234-249.
Lowe, A.J. and Abbott, R.J. (2000) Routes of origin of two recently evolved hybrid
taxa: Senecio vulgaris var. hybernicus and York radiate groundsel
(Asteraceae) American Journal of Botany 87: 1159-1167.

187
Lowe, A.J. and Abbott, R.J. (2004) Reproductive isolation of a new hybrid species,
Senecio eboracensis Abbott and Lowe (Asteraceae) Heredity 92: 386-395.
Mack, R.N. (1996) Biotic barriers to plant naturalisation. In: V.C. Moran and J.H.
Hoffman (eds) Proceedings of the IX International Symposium on Biological
Control of Weeds 19-26 January 1996, Stellenbosch, South Africa, pp. 39-46.
University of Cape Town, Cape Town.
MacLeod, A., Wratten, S.D., Sotherton, N.W. and Thomas, M.B. (2004) ‘Beetle
banks’ as refuges for beneficial arthropods in farmland: long-term changes in
predator communities and habitat. Agricultural and Forest Entomology 6:
147-154.
MacNeil, C., Dick, J.T.A., Hatcher, M.J., Terry, R.S., Smith, J.E. and Dunn, A.M.
(2003) Parasite-mediated predation between native and invasive amphipods.
Proceedings of the Royal Society of London B 270: 1309-1314.
Maezono, Y., and Miyashita, T. (2003) Community-level impacts induced by
introduced largemouth bass and bluegill in farm ponds in Japan. Biological
Conservation 109: 111-121.
Maezono, Y., Kobayashi, R., Kusahara, M. and Miyashita, T. (2005) Direct and
indirect effects of exotic bass and bluegill on exotic and native organisms in
farm ponds. Ecological Applications 15: 638-650.
Malmstrom, C.M., Hughes, C.C., Newton, L.A. and Stoner, C.J. (2005a) Virus
infection in remnant native bunchgrasses from invaded California grasslands.
New Phytologist 168: 217-230.
Malmstrom, C.M., McCullough, A.J., Johnson, H.A., Newton, L.A. and Borer, E.T.
(2005b) Invasive annual grasses indirectly increase virus incidence in
California native perennial bunchgrasses. Oecologia 145: 153-164.
Marler, M.J., Zabinski, C.A., and Callaway, R.M. (1999) Mycorrhizae indirectly
enhance competitive effects of an invasive forb on a native bunchgrass.
Ecology 80: 1180-1186.

188
Marohasy, J.J. (1989) A survey of fireweed (Senecio madagascariensis Poir) and
its natural enemies in Madagascar with a view to biological control in
Australia. Plant Protection Quarterly 4: 139-140.
Maron, J.L. and Vila, M. (2001) When do herbivores affect plant invasion?
Evidence for the natural enemies and biotic resistance hypothesis. Oikos
95: 361-373.
Marshall, D.F. and Abbott, R.J. (1980) On the frequency of introgression of the
radiate (Tr) allele from Senecio squalidus L. into Senecio vulgaris. Heredity
45:133-135.
Mattson, W.J. (1980) Herbivory in relation to plant nitrogen content. Annual Review
of Ecology and Systematics 11: 119-161.
McDowall, R. M. (2003) Impacts of introduced salmonids on native galaxiids in
New Zealand upland streams: A new look at an old problem. Transactions of
the American Fisheries Society 132: 229-238.
Memmott, J. and Waser, N.M. (2002) Integration of alien plants into a native flower-
pollinator visitation web. Proceedings of the Royal Society of London B 269:
2395-2399.
Meng, L. and Orsi, J.J. (1991) Selective predation by larval striped bass on native
and introduced copepods. Transactions of the American Fisheries Society
120: 187-192.
Miller, K.E. and Gorchov, D.L. (2004) The invasive shrub, Lonicera maackii,
reduces growth and fecundity of perennial forest herbs. Oecologia 139: 359-
375.
Miller, T.E. (1994) Direct and indirect species interactions in an early old-field plant
community. The American Naturalist 6: 1007-1025.
Mistri, M., Rossi, R. and Fano, E. A. (2004) The spread of an alien bivalve
(Musculista senhousia) in the Sacca di Goro Lagoon (Adriatic Sea, Italy).
Journal of Molluscan Studies 70: 257-261.

189
Mitchell, R.J., Karron, J.D., Holmquist, K.G. and Bell, J.M. (2004) The influence of
Mimulus ringens floral display size on pollinator visitation patterns. Functional
Ecology 18: 116-124.
Moeller, D.A. (2004) Facilitative interactions among plants via shared pollinators.
Ecology 85: 3289-3301.
Moeller, D.A. (2005) Pollinator community structure and sources of spatial variation
in plant-pollinator interactions in Clarkia xantiana ssp. xantiana. Oecologia
142: 28-37.
Mooney, H.A. and Cleland, E.E. (2001) The evolutionary impact of invasive
species. Proceedings of the National Academy of Sciences of the United
States of America 98: 5446-5415.
Moragues, E. and Travaset, A. (2005) Effect of Carpobrotus spp. On the pollination
success of native plant species of the Balearic Islands. Biological
Conservation 122: 611-619.
Morrell, P.L., Williams-Coplin, T.D., Lattu, A.L., Bowers, J.E., Chandler, J.M. and
Paterson, A.H. (2005) Crop-to-weed introgression has impacted allelic
composition of johnsongrass populations with and without recent exposure
to cultivated sorghum. Molecular Ecology 14: 2143-2154.
Morris, R.J. (2002) The role of indirect interactions in structuring tropical insect
communities. Oikos 97: 308-311.
Morris, R.J., Lewis, O.T. and Godfray, H.C.J. (2004) Experimental evidence for
apparent competition in a tropical forest food web. Nature 428: 310-313.
Mothershead, K. and Marquis, R.J. (2000) Fitness impacts of herbivory through
indirect effects on plant-pollinator interactions in Oenothera macrocarpa.
Ecology 81: 30-40.
Naumann, I.D., Carne, P.B., Lawrence, J.F., Nielsen, E.S., Spradbery, J.P., Taylor,
R.W., Whitten, M.J., and Littlejohn, M.J. (1991) The Insects of Australia. A

190
Textbook for Students and Research Workers. 2 Vols. Melbourne University
Press, Melbourne.
Nelson, N.R. (1980) The germination and growth characteristics of fireweed
(Senecio madagascariensis). B.Sc.Agr. Thesis, University of Sydney.
Noble, I. (1989) Attributes of invaders and the invading process: terrestrial and
vascular plants. In: J.A. Drake, H.A. Mooney, F. di Castri, R.H. Groves, F.J.
Kruger, M. Rejmánek and M. Williamson (eds) Biological invasions. a global
perspective, pp. 301-313. John Wiley, Chichester.
Noonburg, E.G. and Byers, J.E. (2005) More harm than good: when invader
vulnerability to predators enhances impact on native species. Ecology 86:
2555-2560.
Norbury, G. (2001) Conserving dryland lizards by reducing predator-mediated
apparent competition and direct competition with introduced rabbits. Journal
of Applied Ecology 38: 1350-1361.
Novotny, V., Miller, S.E., Cizek, L., Leps, J., Janda, M., Basset, Y., Weiblen, G.D.,
and Darrow, K. (2003) Colonising aliens: caterpillars (Lepidoptera) feeding on
Piper aduncum and P. umbellatum in rainforests of Papua New Guinea.
Ecological Entomology 28: 704-716.
Nowicki P., Witek M., Skorka P. and Woyciechowski M. (2005) Oviposition patterns
in the myrmecophilous butterfly Maculinea alcon Denis and Schiffermuller
(Lepidoptera: Lycaenidae) in relation to characteristics of foodplants and
presence of ant hosts. Polish Journal of Ecology 53: 409-417.
Nyström, P., Svensson, O., Lardner, B., Brönmark and Granéli, W. (2001) The
influence of multiple introduced predators on a littoral pond community.
Ecology 82: 1023-1039.
Obeso, J.S. and Grubb, P.J. (1994) Interactive effects of extent and timing of
defoliation and nutrient supply on reproduction in a chemically protected
annual Senecio vulgaris. Oikos 71: 506-514.

191
O’Dowd, D.J., Green, P.T. and Lake, P.S. (2003) Invasional ‘meltdown’ on an
oceanic island. Ecology Letters 6: 812-817.
O'Hanlon, P.C. and Peakall, R. (2000) A simple method for the detection of size
homoplasy among amplified fragment length polymorphism fragments.
Molecular Ecology 9: 815-816.
Olckers, T. and Hulley, P.E. (1991) Impoverished insect herbivore faunas on the
exotic bugweed Solanum mauritianum Scop. relative to indigenous Solanum
species in Natal/KwaZulu and the Transkei. Journal of the Entomological
Society of South Africa 34: 39-50.
Ornduff, R. (1960) An interpretation of the Senecio lautus complex in New Zealand.
Transactions of the Royal Society of New Zealand 88: 63-77.
Ornduff, R. (1964) Evolutionary pathways of the Senecio lautus alliance in New
Zealand and Australia. Evolution 18: 349-360.
Page, R.D.M. (1996) TREEVIEW: An application to display phylogenetic trees on
personal computers. Computer Applications in the Biosciences 12: 357-
358.
Pannell, J.R., Obbard, D.J. and Buggs, R.A. (2004) Polyploidy and the sexual
system: what can we learn from Mercurialis annua? Biological Journal of
the Linnean Society 82: 547-560.
Parker, I.M. (1997) Pollinator limitation of Cytisus scoparius (Scotch broom), an
invasive exotic shrub. Ecology 78: 1457-1470.
Parker, I.M. and Haubensak, K.A. (2002) Comparative pollinator limitation of two
non-native shrubs: do mutualisms influence invasions? Oecologia 130: 250-
258.
Parker J.D., Burkepile D.E. and Hay M.E. (2006) Opposing effects of native and
exotic herbivores on plant invasions. Science 311: 1459-1461.
Parker J.D and Hay M.E. (2005) Biotic resistance to plant invasions? Native
herbivores prefer non-native plants. Ecology Letters 8: 959-967.

192
Parker, M.A. (2001) Mutualism as a constraint on invasion success for legumes
and rhizobia. Diversity and Distributions 7: 125-136.
Peakall, R. and Smouse, P.E. (2006) GENALEX 6: genetic analysis in Excel.
Population genetic software for teaching and research. Molecular Ecology
Notes 6: 288-295.
Pearson, D.E. and Callaway, R.M. (2003) Indirect effects of host-specific biological
control agents. Trends in Ecology and Evolution 18: 456-460.
Pelser, P.B., Nordenstam, B., Kadereit, J.W. and Watson, L.E. (2006) An ITS
phylogeny of Tribe Senecioneae (Asteraceae) and a new delimitation of
Senecio. Botany 2006, Chico, USA.
Petren, K. and Case, T.J. (1996) An experimental demonstration of exploitation
competition in an ongoing invasion. Ecology 77: 118-132.
Potvin, C. and Bernatchez, L. (2001) Lacustrine spatial distribution of landlocked
Atlantic salmon populations assessed across generations by multilocus
individual assignment and mixed-stock analyses. Molecular Ecology 10:
2375-2388.
Prentis, P.J., Vesey, A., Meyers, N.M. and Mather, P.B. (2004) Genetic structuring
of the stream lily Helmholtzia glaberrima (Philydraceae) within Toolona
Creek, south-eastern Queensland. Australian Journal of Botany 52: 201-207.
Pyšek, P., Richardson, D.M., Rejmánek, M., Webster, G.L., Williamson, M. and
Kirschner, J. (2004) Alien plants in checklists and floras: towards better
communication between taxonomists and ecologists. Taxon 53: 131-143.
Radford, I.J. (1997) Impact assessment for the biological control of Senecio
madagascariensis Poir. (fireweed). PhD Thesis, The University of Sydney,
N.S.W. Australia, Sydney.
Radford, I.J. and Cousens, R.D. (2000) Invasiveness and comparative life-history
traits of exotic and indigenous Senecio species in Australia. Oecologia 125:
531-42.

193
Radford, I.J., King, D., and Cousens, R.D. (1995a) A survey of Senecio
madagascariensis Poir. (fireweed) density in pastures of coastal New South
Wales. Plant Protection Quarterly 10: 107-111.
Radford, I.J., Liu, Q. and Michael, P.W. (1995b). Chromosome counts for the
Australian weed known as Senecio madagascariensis (Asteraceae).
Australian Systematic Botany 8: 1029-1033.
Raikow, D.F. (2004) Food web interactions between larval bluegill (Lepomis
macrochirus) and exotic zebra mussels (Dreissena polymorpha) Canadian
Journal of Fisheries and Aquatic Sciences 61: 497-504.
Rand, T.A. (2003) Herbivore-mediated apparent competition between two salt
marsh forbs. Ecology 84: 1517-1526.
Rand, T.A. and Louda, S.M. (2004) Exotic weed invasion increases the
susceptibility of native plants to attack by a biocontrol herbivore. Ecology 85:
1548-1554.
Rathcke, B. (1983) Competition and facilitation among plants for pollination. In: L.
Real (ed) Pollination Biology, pp. 305-329. Academic Press, New York.
Rausher, M.D. (1981) Host plant selection by Battus Philenor Butterflies: the roles
of predation, nutrition, and plant chemistry. Ecological Monographs 51: 1-20.
Rejmánek, M. (1998) Invasive plant species and invasible ecosystems. In: O.T.
Sandlund, P.J. Schei, and A. Viken (eds) Invasive Species and Biodiversity
Management, pp. 79-102. Kluwer Academic Publishers, Dordrecht.
Ricciardi, A. (2003) Facilitative interactions among aquatic invaders: is an
“invasional meltdown” occurring in the Great Lakes? Canadian Journal of
Fisheries and Aquatic Sciences 58: 2513-2528.
Ricciardi, A. and Atkinson, S.K. (2004) Distinctiveness magnifies the impact of
biological invaders in aquatic ecosystems. Ecology Letters 7: 781-784.

194
Richardson, D.M., Allsopp, N., D’Antonio, C.M., Milton, S.J. and Rejmánek, M.
(2000) Plant invasions – the role of mutualisms. Biological Reviews 75: 65-
93.
Ridenour, W.L. and Callaway, R.M. (2003) Root herbivores, pathogenic fungi, and
competition between Cenaurea maculosa and Festuca idahoensis. Plant
Ecology 169: 161-170.
Rieseberg, L.H. (1996) Homology among RAPD fragments in interspecific
comparisons. Molecular Ecology 5: 99-105.
Rieseberg, L.H., van Fossen, C. and Desrochers, A. (1995) Hybrid speciation
accompanied by genomic reorganization in wild sunflowers. Nature 375:
313-316.
Rieseberg, L.H., Raymond, O., Rosenthal, D.M., Lai, Z., Livingstone, K., Nakazato,
T., Durphy, J.L., Schwarzbach, A.E., Donovan, L.A. and Lexer, C. (2003)
Major ecological transitions in wild sunflowers facilitated by hybridization.
Science 301: 1211-1216.
Rieseberg, L.H. and Wendel, J.F. (1993) Introgression and its consequences in
plants. Pp. 70-109 in: Harrison, R.G. (ed.) Hybrid zones and the
evolutionary process. Oxford University Press, New York.
Roane, M.K., Griffin, G.J. and Elkins, J.R. (1986) Chestnut blight, and other
Endothia diseases, and the genus Endothia. American Phytopathological
Society, St. Paul.
Roemer, G.W., Donlan, C.J. and Courchamp, F. (2002) Golden eagles, feral pigs,
and insular carnivores: How exotic species turn native predators into prey.
Proceedings of the National Academy of Sciences 99: 791-796.
Rogers, W.E. and Siemann, E. (2002) Effects of simulated herbivory and resource
availability on native and invasive exotic tree seedlings. Basic and Applied
Ecology 3: 297-307.

195
Rooney, T.P. and Waller, D.M. (2003) Direct and indirect effects of white-tailed
deer in forest ecosystems. Forest Ecology and Management 181: 165-176.
Rothschild, M., Aplin, R.T., Cockrum, P.A., Edgar, J.A., Fairweather, P. and Lees,
R. (1979) Pyrrolizidine alkaloids in Arctiid moths (Lep.) with a discussion on
host plant relationships and the role of these secondary plant substances in
the Arctiidae. Biological Journal of the Linnean Society 12: 305-326.
Russell, F.L. and Louda, S.M. (2005) Indirect interaction between two native
thistles mediated by an invasive exotic floral herbivore. Oecologia 146: 373-
384.
Russell, L.F., Louda, S.M., Rand, T.A. and Kachman, S.D. (2007) Variation in
herbivore-mediated indirect effects of an invasive plant on a native plant.
Ecology 88: 413-423.
Sans, F.X., Garcia-Serrano, H. and Afan, I. (2004) Life-history traits of alien and
native Senecio species in the Mediterranean region. Acta Oecologica 26:
167-178.
Savidge, J. (1987) Extinction of an island forest avifauna by an introduced snake.
Ecology 68: 660-668.
Scherber, C., Crawley, M.J. and Porembski, S. (2003) The effects of herbivory and
competition on the invasive alien plant Senecio inaequidens (Asteraceae).
Diversity and Distributions 9: 415-426.
Schierenbeck, K.A., Mack, R.N. and Sharitz, R.R. (1994) Effects of herbivory on
growth and biomass allocation in native and introduced species of Lonicera.
Ecology, 75: 1661-1672.
Schoener, T.W. (1993) On the relative importance of direct versus indirect effects
in ecological communities. In: H. Kawanabe, J.E. Cohen and K. Iwasaki (eds)
Mutualism and community organisation, pp. 365-415. Oxford University
Press, Oxford.

196
Scott, L.J., Congdon, B.C. and Playford, J. (1997) Molecular evidence that fireweed
(Senecio madagascariensis, Asteraceae) is of South African origin. Plant
Systematics and Evolution 213: 251-257.
Sessions, L. and Kelly, D. (2002) Predator-mediated apparent competition between
an introduced grass, Agrostis capillaries and a native fern, Botrychium
australe (Ophioglossaceae), in New Zealand. Oikos 96: 102-109.
Settle, W.H. and Wilson, L.T. (1990) Invasion by the variegated leafhopper and
biotic interactions: parasitism, competition and apparent competition. Ecology
71: 1461-1470.
Shannon, C.E. (1948) A mathematical theory of communication. Bell Systems
Technical Journal 27: 379–423.
Shelton, A.M. and Badenes-Perez, F.R. (2006) Concepts and applications of trap
cropping in pest management. Annual Review of Entomology 51: 285-308.
Shibaike, H., Akiyama, H., Satoshi, U., Kasai, K., and Morita, T. (2002)
Hybridization between European and Asian dandelions (Taraxacum section
Ruderalia and section Mongolica) 2. Natural hybrids in Japan detected by
chloroplast DNA marker. Journal of Plant Research 115: 321-328.
Shurin, J.B., Borer, E.T., Seabloom, E.W., Anderson, K., Blanchette, C.A.,
Briotman, B., Cooper, S.D. and Halpern, B.S. (2002) A cross-ecosystem
comparison of the strength of trophic cascades. Ecology Letters 5: 785-791.
Siemann, E. and Rogers, W.E. (2003) Herbivory, disease, recruitment limitation,
and success of alien and native tree species. Ecology 84: 1489-1505.
Simberloff, D. (1986) Introduced insects: a biogeographic and systematic
perspective. In: H.A. Mooney and J.A. Drake (eds) Ecology of Biological
Invasions of North America and Hawaii, pp. 3-26, Springer-Verlag, New York.
Simberloff, D. and Von Holle, B. (1999) Positive interactions of nonindigenous
species: invasional meltdown? Biological Invasions 1: 21-32.

197
Simon, K.S. and Townsend, C.R. (2003) Impacts of freshwater invaders at different
levels of ecological organisation, with emphasis on salmonids and ecosystem
consequences. Freshwater Biology 48: 982-994.
Sims, N. (2004) Preference and performance of Nyctemera amica on a native and
introduced species of Senecio: linking herbivory studies with invasion biology.
Unpublished B. Appl. Sc. (Hons) thesis, Queensland University of
Technology, Brisbane.
Sindel, B.M. and Michael, P.W. (1992) Growth and competitiveness of Senecio
madagascariensis Poir. (fireweed) in relation to fertilizer use and increases in
soil fertility. Weed Research 32: 399-406.
Sindel, B.M. and Michael, P.W. (1996) Seedling emergence and longevity of
Senecio madagascariensis Poir. (fireweed) in coastal south-eastern
Australia. Plant Protection Quarterly 11: 14-19.
Sindel, B.M., Radford, I.J., Holtkamp, R.H. and Michael, P.W. (1998) The biology of
Australian weeds 33. Senecio madagascariensis Poir. Plant Protection
Quarterly 13: 2-15.
Singer, M.C. (1983) Quantification of host preferences by manipulation of
oviposition behaviour in the butterfly Euphydryas editha. Oecologia 52: 230-
35.
Singh, P. and Mabbett, F.E. (1976) Note on the life history of the magpie moth,
Nyctemera amica (Lepidoptera: Arctiidae). New Zealand Journal of Ecology
3: 277-278.
Slansky Jr, F. and Rodriquez, J.G. (1987) Nutritional Ecology of Insects, Mites,
Spiders and Related Invertebrates. Wiley Inter-Science, New York.
Smith, A.P. and Quin, D.G. (1996) Patterns and causes of extinction and decline in
Australian conilurine rodents. Biological Conservation 77: 243-267.

198
Standish, R.J., Robertson, A.W. and Williams, P. A. (2001) The impact of an
invasive weed Tradescantia fluminensis on native forest regeneration.
Journal of Applied Ecology 38: 1253-1263.
Stastny, M., Schaffner, U. and Elle, E. (2005) Do vigour of introduced populations
and escape from specialist herbivores contribute to invasiveness? Journal of
Ecology 93: 27-37.
Stone, L. and Roberts, A. (1991) Conditions for a species to gain advantage from
the presence of competitors. Ecology 72: 1964-1972.
Stout, J.C., Kells, A.R. and Goulson, D. (2002) Pollination of the invasive exotic
shrub Lupinus arboreus (Fabaceae) by introduced bees in Tasmania.
Biological Conservation 106: 425-434.
Strauss, S.Y. (1991) Indirect effects in community ecology: Their definition, study
and importance. Trends in Ecology and Evolution 6: 206-210.
Strong, D.R. (1992) Are trophic cascades all wet? Differentiation and donor-control
in speciose ecosystems. Ecology 73: 747-754.
Talman, S.G. and Keough, M.J. (2001) Impact of an exotic clam, Corbula gibba, on
the commercial scallop Pecten fumatus in Port Phillip Bay, south-east
Australia: Evidence of resource-restricted growth in a subtidal environment.
Marine Ecology Progress Series 221: 135-143.
Tamayo, M., Grue, C.E. and Hamel, K. (2004) Densities of the milfoil weevil
(Euhrychiopsis lecontei) on native and exotic watermilfoils. Journal of
Freshwater Ecology 19: 203-211.
Taylor, R.H. (1979) How the Macquarie Island parakeet became extinct. New
Zealand Journal of Ecology 2: 42-45.
Thomas, C.D. (1986) Butterfly larvae reduce host plant survival in vicinity of
alternative host species. Oecologia 70: 113-117.
Thompson, I.R. (2006) A taxanomic treatment of tribe Senecioneae (Asteraceae) in
Australia. Muelleria 24: 51-110.

199
Tompkins, D.M., Sainsbury, A.W., Nettleton, P., Buxton, D. and Gurnell, J. (2002)
Parapoxvirus causes a deleterious disease in red squirrels associated with
UK population declines. Proceedings of the Royal Society of London B 269:
529-533.
Torrusio, S., Cigliano, M.M. and de Wysiecki, M.L. (2002) Grasshopper
(Orthoptera: Acridoidea) and plant community relationships in the Argentine
pampas. Journal of Biogeography 29: 221-229.
Townsend, C.R. (1996) Invasion biology and ecological impacts of brown trout
Salmo trutta in New Zealand. Biological Conservation 78: 13-22.
Turner, D. and Conran, J.G. (2004) The reproductive ecology of two naturalised
Erica species (Ericaceae) in the Adelaide Hills: The rise and fall of two
"would-be" weeds? Transactions of the Royal Society of South Australia 128:
23-31.
van der Putten, W.H., Yeates, G.W., Duyts, H., Schreck Reis, C. and Karssen, G.
(2005) Invasive plants and their escape from root herbivory: a worldwide
comparison of the root-feeding nematode communities of the dune grass
Ammophila arenaria in natural and introduced ranges. Biological Invasions 7:
733-746.
van Riper, C., van Riper, S.G., Goff, M.L. and Laird, M. (1986) The epizootiology
and ecological significance of malaria in Hawaiian land birds. Ecological
Monographs 56: 327-344.
Vazquez, D.P. and Aizen, M.A. (2004) Asymmetric specialization: A pervasive
feature of plant-pollinator interactions. Ecology 5: 1251-1257.
Veech, J.A. (2000) Predator-mediated interactions among the seeds of desert
plants. Oecologia 124: 402-407.
Verma, L.R. and Rana, R.S. (1994) Further studies on the behavior of Apis cerana
and Apis mellifera foraging on apple flowers. Journal of Apiculture Research
33: 175-179.

200
Vilà, M. and D’Antonio, C.M. (1998) Fitness of invasive Carpobrotus (Aizoaceae)
hybrids in coastal California. Ecoscience 5: 191-199.
Vilà, M., Gomez, A. and Maron, J.L. (2003) Are alien plants more competitive than
their native conspecifics? A test using Hypericum perforatum L. Oecologia
137: 211-215.
Waser, N.M. and Real, L.A. (1979) Effective mutualism between sequentially
flowering plant species. Nature 281: 670-672.
Wauters, L.A., Gurnell, J., Martinoli, A. and Tosi, G. (2002) Interspecific
competition between native Eurasian red squirrels and alien grey squirrels:
Does resource partitioning occur? Behavioral Ecology and Sociobiology 52:
332-341.
Webster, M.S. and Almany, G.R. (2002) Positive indirect effects in a coral reef fish
community. Ecology Letters 5: 549-557.
White, E.M. (2007) Insect-mediated indirect interactions between an exotic and
native Senecio species. Thesis, Queensland University of Technology,
Brisbane.
White, E.M., Sims, N.M. and Clarke, A.R. (in press) A test of the enemy release
hypothesis: The native magpie moth prefers a native fireweed (Senecio
pinnatifolius) to its introduced congener (S. madagascarensis). Austral
Ecology.
White, E.M., Wilson, J.C. and Clarke, A.R. (2006) Biotic indirect effects: A
neglected concept in invasion biology. Diversity and Distributions 12: 443-
455.
Whitham, T.G., Martinsen, G.D., Floate, K.D., Dungey, H.S., Potts, B.M. and Keim,
P. (1999) Plant hybrid zones affect biodiversity: Tools for a genetic-based
understanding of community structure. Ecology 80: 416-428.

201
Whitney, K.D., Randall, R.A. and Rieseberg, L.H. (2006) Adaptive Introgression of
Herbivore Resistance Traits in the Weedy Sunflower Helianthus annuus.
American Naturalist 167: 794-807.
Wilson, D.S. (1986) Adaptive indirect effects. In Diamond, J.D. and Case, T.J.
(eds) Community ecology, pp. 437-444. Harper and Row, New York.
Wilson, P. R., Karl, B. J., Toft, R. J., Beggs, J. R. and Taylor, R. H. (1998) The role
of introduced predators and competitors in the decline of kaka (Nestor
meridionalis) populations in New Zealand. Biological Conservation 83: 175-
185.
Wolf, D.E., Takebayashi, N. and Rieseberg, L.H. (2001) Predicting the Risk of
Extinction through Hybridization. Conservation Biology 15: 1039-1053.
Wolf, J.J., Beatty, S.W. and Seastedt, T.R. (2004) Soil characteristics of Rocky
Mountain National Park grasslands invaded by Melilotus officinalis and M.
alba. Journal of Biogeography 31: 415-424.
Wolfe, L.M. (2002) Why alien invaders succeed: support for the escape-from-
enemy hypothesis. The American Naturalist 160: 705-711.
Wootton, J.T. (1994). The nature and consequences of indirect effects in ecological
communities. Annual Review of Ecology and Systematics 25: 443-466.
Wootton, J.T. (2002) Indirect effects in complex ecosystems: recent progress and
future challenges. Journal of Sea Research 48: 157-172.
Xiang, H. and Chen, J. (2004) Interspecific Variation of Plant Traits Associated with
Resistance to Herbivory Among Four Species of Ficus (Moraceae). Annals of
Botany 94: 377-384.
Yelenik, S.G., Stock, W.D. and Richardson, D.M. (2004) Ecosystem level impacts
of invasive Acacia saligna in the South African fynbos. Restoration Ecology
12: 44-51.

202
Zancola, B.J., Wild, C. and Hero, J.M. (2000) Inhibition of Ageratina riparia
(Asteraceae) by native Australian flora and fauna. Austral Ecology 25: 563-
569.
Zawko, G., Krauss, S.L., Dixon, K.W. and Sivasithamparam, K. (2001)
Conservation genetics of the rare and endangered Leucopogon obtectus
(Ericaceae). Molecular Ecology 10: 2389-2396.





























