Embed Size (px)
Citation preview

Semenov Institute of Chemical Physics, RAS
New results in applications of p-adic pseudo-differential equations to the protein dynamics
Vladik Avetisov Albert BikulovSergey KozyrevVladimir Osipov
Alexander Zubarev in cooperation with
Viktor Ivanov and Alexander Chertovich

What is a protein?

“Protein? It is very simple” - biologist will say. - Protein is a well folded polymeric chain of a few hundreds amino aside residues.
Proteins looks like “nono-pocket devices” constructed from helixes and sheets. They are fabricated in a cell to provide all biochemical reactions including the protein fabrication too.”

“Nothing of the kind!” – physicist will rejoin.- “Protein looks like an amorphous “nono-drop” consisting of a few thousands closely interacting atoms. There is no symmetry here. O-o-o…! Protein is too complex to be described by a simple way.”
- C carbon atoms, - O oxygen atoms, - N nitrogen
atoms, - hydrogen atoms
are not shown

Figure 5. Hierarchical arrangement of the conformational substates in myoglobin. (a) Schematized energy landscape. (b) Tree diagram. G is Gibbs’ energy of the protein, CC(1-4) are conformational coordinates.
Hans Frauenfelder, in Protein Structure (N-Y.:Springer Verlag, 1987)
p.258.
“The results sketched so far suggest two significant properties of substates and motions of proteins, nonergodicity and ultrametricity.”
Hans Frauenfelder, in Protein Structure (N-Y.:Springer Verlag, 1987)
p.258.
Hans Frauenfelder was first who drown ultrametric tree for the protein states to underline the protein complexity.

«In <…> proteins, for example, where individual
states are usually clustered in “basins”, the
interesting kinetics involves basin-to-basin
transitions. The internal distribution within a basin
is expected to approach equilibrium on a relatively
short time scale, while the slower basin-to-basin
kinetics, which involves the crossing of higher
barriers, governs the intermediate and long time
behavior of the system.”
O.M.Becker and M.Karplus.
J.Chem.Phys. 106, 1495 (1997)
“What does ultrametricity mean physically in protein dynamics?”
This means that the protein dynamics is characterized by a hierarchy of time scales.

Given such picture, we will take an interest to Frauenfelder’s
question,
“Are proteins ultrametric?”
p-Adic mathematics gives us natural tools to try to find an answer.
1w
2w3w
4w
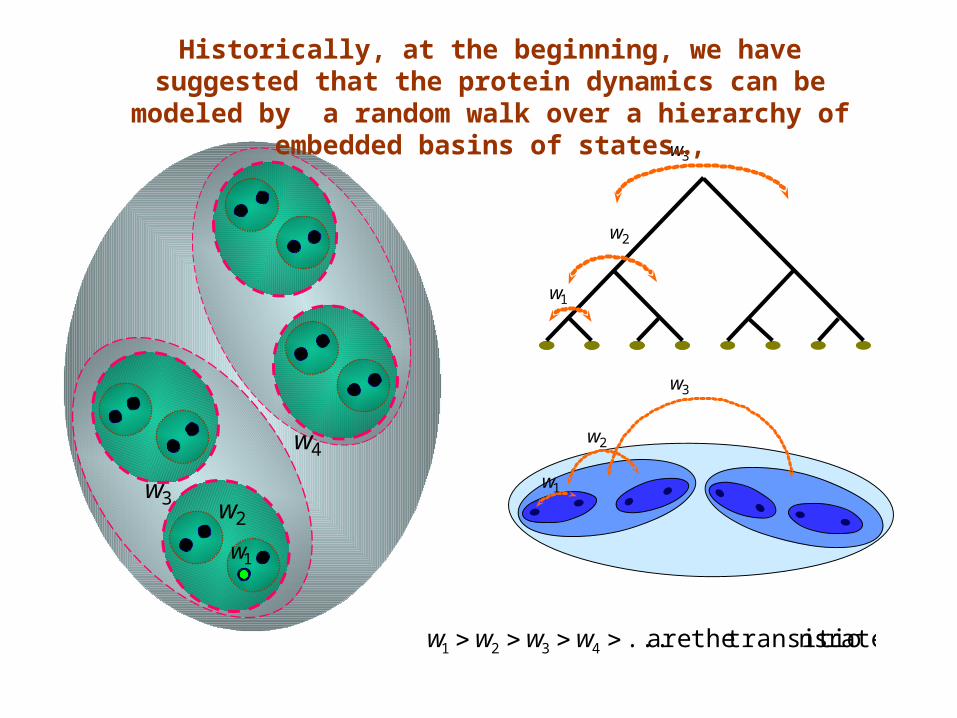
rates ns transitio theare ...4321 wwww
1w
2w
1w
2w
3w
3w
Historically, at the beginning, we have suggested that the protein dynamics can be
modeled by a random walk over a hierarchy of embedded basins of states…,

and so, the protein dynamics can be described by the р-adic pseudo-differential equation of ultrametric
diffusion
p
ydtxftyfyxAt
txfp
Q
)(),(),(||),( 1
.landscape)enargy on the barriers
activation of masuringfor unit scale a is e,temperatur-( /~
; and statesbetween raten transitioa is||
;instant at state ain density population a is ,
states;protein theof space cultrametrian is
00
1
TTTT
yxyxA
txtxf
Q
p
p
In protein dynamic applications, this equation is interpreted as the muster
equation for the transitions between the protein states.

Assuming all this, we have tried to show that our approach is relevant to
observable features of the protein dynamics.
<If you said “A”, do not be “B”>.
Vitalii Goldanskii

h
experiment: kinetics of CO rebinding to myoglobine
(H. Frauenfelder group, since the 1970s)
Mb-COCO
Mb
Measured quantity :.The total concentration of the Mb unbounded to CO.
Mb-CO
rebinding CO to Mb
CObreaking
of chemical
bound Mb-CO
Mb*
strained confor-
mational state
Conformational rearrangements of the Mb
Mb1
active confor-
mational state
laser pulse

p-adic model of CO rebinding kinetics
xdtxftSrB
,
measured quantity:
Zp
reaction sink
Initial distribution
Br
Protein “diffuses” over unbounded
states
ultrametric space of the protein
undounded states
ultrametric diffusion
reaction sink due to CO binding to Mb
protein leaves unbounded
states

experiment and theory
T1>T2>T3
O O
Good agreement between ultrametric model and experiment certainly supports an idea
that protein dynamics possesses ultrametricity (!)

This was a pioneer experience in applications of p-adic pseudo-differential equation to the protein dynamics: it was presented on the First Conference on p-
Adic Mathematical Physic (Moscow, 2003)

Now, I will present new experience in applications of the same p-adic equation to drastically different phenomenon related to the protein dynamics.
This is the spectral diffusion in proteins.

Spectral diffusion in proteins
a chromophore marker
protein
1. A chromophore marker is injected into a protein, then the protein is frozen up to a few degrees of Kelvin, and the adsorption spectrum is measured.
At low temperature this spectrum is very wide, due to different arrangements of the protein atoms around a chromophore in individual protein molecules.
2. Then, a set of chromophore markers are burned using short light impulse at a particular absorption frequency, and a narrow hole is arisen in the absorption spectrum.
3. Then, the hole wide is monitored during the time. Because proteins with unburned markers “diffuse” over the protein state space, the hole is broadening and covering with time.
Thus, spectral diffusion phenomenon is directly coupled with the protein dynamics.

Spectral diffusion features
~ 27,02/1
22www ttt
“Weighting time” experiments:
The “weighting time”, tw , starts immediately after the burning of a hole, i.e. it is the current time for spectral diffusion:
“Aging time” experiments:
The “aging time”, tag , is the time interval between protein freezing and hole burning.
07.04 ~min10, agwag ttt
Spectral diffusion broadening obeys the power law with an exponent drastically smaller then in familiar diffusion.
The aging time, tag, grows, spectral
diffusion broadening becomes slower .
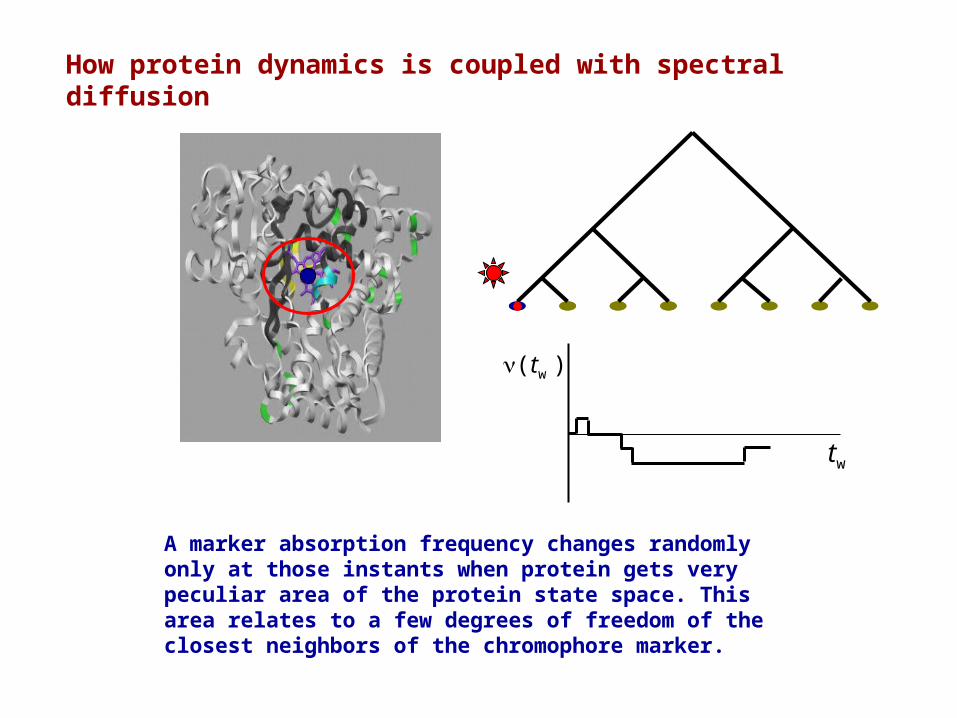
How protein dynamics is coupled with spectral diffusion
(tw )
tw
A marker absorption frequency changes randomly only at those instants when protein gets very peculiar area of the protein state space. This area relates to a few degrees of freedom of the closest neighbors of the chromophore marker.

Description of spectral diffusion in proteins
1. Calculation of the distribution of the first passage (first reaching) time P() for ultrametric diffusion
1,~,121
agag ttP
ydyx
txftyf
t
txf
pQ p
1||
,,,
2. Construction of the random walk on a “frequency line”, which has random delays with the distribution P()
1,, 2
1122
1
wagwag tttt
note, that the first reaching time depends on the aging time tag
p-adic equation for the protein dynamics

Spectral diffusion broadening:(analytical description and computer simulation)
27,0~ ww tt
2,2
ydyx
txftyf
t
txf
pQ p
1||
,,,

0,2
w
Spectral diffusion aging:(analytical description and computer simulation)
2,2
07,0 agag tt
The exponents in the power laws of spectral diffusion broadening and aging are determined only by a degree of Vladimirov’s pseudo-differential operators (2) (!)

ydyx
txftyf
t
txf
pQ p
1||
,,,
Protein is a complex object.
However, the protein dynamics is described by simple p-adic equation.
Conclusion:

Probably, p-adic pseudo-differential equations will be as much important to keep paradoxical union of order and randomness in the “biological mechanics”, as Newton’s equations are in classical mechanics.





























