Embed Size (px)
Citation preview

Selective differentiation of CD4+ T helper cell subsets
Phillip Scott
School of Veterinary Medicine, University of Pennsylvania, Philadelphia, USA
During the past year significant advances have been made in our
understanding of the factors contributing to the differentiation of CD4+ T helper cell subsets. These have been driven, in part, by the realization
that signals (cytokines) from the innate immune response are critical in T cell subset differentiation. In addition, several studies have also elucidated
how the antigen-presenting cell, the antigenic epitope and the antigen concentration may contribute to the selective development of a particular
T helper cell subset.
Current Opinion in immunology 1993, 5:391-397
Introduction
A major advance in the field of immunology was the es- tablishment of a simple paradigm that provides a frame- work for analping why certain immune responses domi- nate following exposure to a particular pathogen or anti- gen. The essence of this paradigm is that CD4+ T cells can be separated into subsets based upon the repertoire of cytokines produced following stimulation, and that the distinct cytokine profile observed in these subsets determines the cell’s function [l-lo]. Although proba- bly more complex than initially envisioned, this model includes two major CD4+ T helper (Th) cell subsets, Thl and Th2, that mediate distinct immune responses, namely cell-mediated and humoral immunity. As these immune responses are frequently mutually exclusive, it was logical to assume that CD4 + subsets would have cross-regulatory activity. In fact, this turns out to be the case, and several cytokines appear to be important in this regard [6,9,10,11*]. However, an unresolved question in this field has been what mechanisms operate to initiate Thl- or Th2-cell development from a naive T cell popula- tion. This review will specifically focus on advances made over the last year in addressing this particular question.
Models of CD4+ T-cell differentiation
The discovery that murine T-cell clones often produce distinct repertoires of cytokines was rapidly followed by studies directed at identifying similar cells in viva. Interestingly, the clearest examples of the existence of such cells has been a variety of parasitisms [ 12.1. This
is probably not accidental, but more likely reflects the evolution of distinct immune responses to eliminate di- verse pathogens. It has been argued that the recognition of pathogens by innate host defenses [macrophages, natural killer (NK) cells, $5 T cells and the alternative complement pathway] signals the adaptive immune re- sponse of the presence of an infection [13]. An equally compelling hypothesis is that signals from the immune system also direct the differentiation of the CD4+ T cell [14], and that in their absence a clear bias in the im- mune response towards either Thl or Th2 cells may not occur. This might explain the existence of CD4+ T-cell clones that do not fit into either the Thl/Th2 paradigm. It follows then that studies with infectious disease models might be most appropriate for identifying these signals. On the other hand, in vivo models are obviously com- plex. Thus, several in vitro systems have been established to identify the minimum requirements for Th-cell differ- entiation. These models involve isolation and stimulation of naive CD4+ cells in the presence of various antigen- presenting cells (ARCS), cytokines or anti-cytokine anti- bodies, followed by measuring the cytokines produced upon restimulation. Initial studies used polyclonal acti- vators (e.g. antiCD3, mitogens) as a stimulus of naive CD4+ T cells, as the frequency of cells responding to a conventional antigen is low [15,16]. During the last year, however, a more elegant approach was used involv- ing CD4+ cells from TCR transgenic mice [ 17**-20**]. Because of the high percentage of CD4+ cells bearing the transgenic TCR, this model permits stimulation with a conventional antigen, presumably more accurately re- flecting in vivo events. The combination of in vivo and
in vitro approaches has been quite fruitful in elucidating mechanisms of CD4+ subset differentiation.
Abbreviations
APC-antigen-presenting cell; HSA-heat stable antigen; IF+interferon; IL-interleukin; mA&monoclonal antibody; NK-natural killer; NKSF-natural killer cell stimulatory factor; WItsevere combined immunodeficient;
TCF-transforming growth factor; Th-T helper.
@ Current Biology Ltd ISSN 0952-7915 391

392 Lymphocyte activation and effector functions
The role of cytokines in CD4+ T-cell
differentiation
Studies in both in vitro and in vivo systems have shown that the cytokines present during primary stimulation are the most important influence on naive Th-cell differen- tiation. The in vivo system where this has been most clearly shown is with the protozoan parasite Letima- niu major. Depending upon the inbred mouse strain infected, this parasite causes either a progressive and eventually fatal infection or a self-healing lesion. Studies from several laboratories have shown that the outcome of infection is dependent upon which CD4+ T-cell subset is stimulated: Thl-cell development leads to resistance, whereas Th2 cells confer susceptibility [2I]. Endoge- nous cytokine levels control which subset predominates [9]. Thus, treatment of resistant mice with anti-interferon (IFN)-y monoclonal antibody (mAb) ablates Thl-cell de- velopment and promotes Th2cell differentiation [22*]. In contrast, treatment of susceptible mice with anti-inter- let&in (IL)-4 mAb inhibits Th2-cell expansion, leading to Thl-cell development [23,24*]. Similar results with anti- IL-4 mAb treatment have recently been obtained with the fungus Candida [25], and even infections that are pn- marily associated with cell-mediated immunity (e.g. Lis terh) exhibit enhanced resistance following elimination of IL-4 [ 261. The importance of IL-4 in Th2-cell differen- tiation is supported by data from the in vitro models as well. For example, in the TCR transgenic systems incuba- tion of CD4+ cells with IL-4 during the primary stimula- tion leads to the development of Th2 cells, whereas addi- tion of anti-IL-4 augmented the development of cells that produce IFN-y [ 17**,18**]. Furthermore, the differentia- tion of T-cell clones exhibiting a ThO cytokine pattern towards a Th2 phenotype was shown to be dependent upon the presence of IL-4 [27-l. In combination, these data provide a powerful argument that IL-4 is critical in Th2cell development.
Implicit in the studies described above is that IL-4 or IFN-y production is induced soon after infection to direct the immune response. In fact, 72 hours after L. major infection, enhanced IL-4 production is observed in the lymph nodes draining the infection site of susceptible (Th2) mice and IFN-y is present in the lymph nodes taken from resistant (Thl) mice, suggesting that the pro- duction of these cytokines is, indeed, part of the innate immune response [22*]. NK cells appear to be a major source of IFN-Y in this system ([28-l ; TM Scharton, P Scott, unpublished data). Thus, in resistant C3H/HeN mice infected with L major, NK cells are activated within the lymph node, while BALB/c mice fail to exhibit this re- sponse [ 28.1. Similarly, NK cells have been implicated as the source of IFNy that drives the differentiation of naive Th cells towards a Thl phenotype following transfer to severe acquired immunodeficient (SCID) mice and infec- tion with L. major [ 29’1. In contrast, the cells producing IL-4 during the initial stages of infection have not been identified. Possible sources include mast cells and/or ba- sophils [30*]. Alternatively, IL-4 may be secreted by the T cells themselves. For example, it was suggested that
recent thymic emigrants provide the IL-4 that drives Th2- cell differentiation [8]. Furthermore, footpad injection of eggs from Schistosomu mansoni induces a rapid cytokine response in the lymph node characterized by elevated levels of IL-4 and IL-5 [31*], and in this system CD44*O T cells produce IL-4 [32*]. Another potential source of cytokines during the early stages of immunity are y6 T cells. In Listeria these cells have been reported to produce IFN-y within three days of infection [33]. In- terestingly, y6 T cells are also present in lesions from patients with Letimaniu [34], and increased numbers are found in both resistant and susceptible strains of mice infected with L, major 135’1. However, whether they promote susceptibility or resistance remains unclear. $3 T cells appear at somewhat later stages of infection with murine influenza virus, and it has been suggested that in this system they participate in resolution of the disease ]361.
In contrast to in vivo studies, a role for IFN-y in Th-cell subset differentiation in vitro has been more diihcult to demonstrate. IFN-y-augments Thl-cell expansion in vitro, possibly through inhibition of Th2cell prolifer- ation [11*,37]. However, when added to naive CD4+ cells a strong bias towards Thl-cell development was not observed, suggesting that other cytokines might be required [ 17**,18*-] . The recently identified cytokine NK cell stimulator-y factor (NKSF) or IL-12 is a likely candi- date as it is produced by macrophages and B cells and stimulates the production of IFN-y from T cells and NK cells [38*,39]. Originally described in the human [40,41], the murine equivalent has now been cloned [42]. IL-12 inhibits Th2-cell expansion in cell lines taken from atopic patients [43-l. Most recently it has been found that the presence of IL-12 during primary stimulation of CD4+ cells from TCR transgenic mice skews the response in the direction of Thl -cell differentiation [20**]. These are ex- tremely important results which suggest that IL-12 may be the IL-4 equivalent for differentiation of Thl cells. One of the appealing aspects of this hypothesis is that it directly links the induction of a Thl response with the nature of the invading pathogen and the innate immune response. Those pathogens associated with Thl-cell development, including intracellular protozoa and bacteria, are the same pathogens shown to induce IL-12 production. For example, the bacterium Staphylococcus aureeus induces IL-12 production by murine and human macrophages [38-l (P Scott, unpublished data). Similarly, the ability of the protozoan Toxoplusmu to stimulate production of IFN-y by NK cells is dependent upon a soluble factor secreted by macrophages that has recently been identi- fied as IL-12 [44*] (A Sher, personal communication). In the leishmanial system, IL-12 production correlates with Thl-cell development. Furthermore, IFN-y production by cells from resistant mice was inhibited when IL-12 was neutralized with mAbs (P Scott, M Wysocka, TM Schar- ton, G Trinchieri, abstract, J Immunol 1333, 150:86A). It is likely that over the ensuing years a series of studies on IL-12 will be reported that will further demonstrate a central role for this cytokine in linking the innate and adaptive immune response, and in directing CD4+ T-cell development.

Selective differentiation of CD4+ T helper cell subsets Scott 393
Several other cytokines may also participate in Th cell subset differentiation. For example, it was demonstrated that in vivo depletion of transforming growth factor (TGF)-p promotes resistance in mice normally sus- ceptible to Leishmania, and that this resistance is asso- ciated with enhanced IFN-y and decreased IL-4 produc- tion [ 45.1. While these results suggest that TGF-l3 may augment Th2-cell development, conflicting data suggest that the presence of TGF-p during in vitro stimulation of either human or murine T cells enhances IFN-y produc- tion, and suppresses the development of IL-4 producing cells [46.,47*]. This discrepancy may relate to the pres- ence or absence of NK cells, as TGF-P has been shown to inhibit NK-cell activity [48-l. Other cytokines that may participate indirectly in the expansion of different Th sub- sets include IL-1 and IL-IO. IL-1 receptors are expressed only after primary stimulation, and thus IL-1 is unlikely to influence naive Th cell activation [49-l. However, high levels of IL-1 might lead to preferential expansion of Th2 cells [49’], particularly as IL-4 up-regulates the IL-1 receptor on Th2 cells [50]. IL-10 may also play a modulatory role, as IL-10 inhibits the APC function of macrophages/dendritic cells, and in vitro neutraliza~
tion of IL-IO, when splenic cells were utilized as APCs, led to enhanced Thl-cell differential [I?*].
Antigen-presenting cells
Studies in the TCR transgenic model indicate that the most efficient APCs for stimulating naive CD4+ T cells are dendritic cells, although other cell types may substi- tute for dendritic cells [ 17**-19**]. These studies suggest that the nature of the APC does not influence which Th subset predominates following primaty stimulation. How- ever, it remains to be determined whether interaction with different co-stimulatory molecules on the APC in- fluences T cell subset differentiation. Several important advances have been made over the last year that will facilitate investigations into this question, These include the demonstration that both B7 and heat stable antigen (HSA) function as co-stimulatory molecules in the murine system [ 510-54.1. Particularly relevant to the stimulation of naive Th cells were studies showing that the presenta
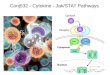
Innate immune sptem Thl cell
L-7 Bacteria
Protozoa
Viruses
Helminths -
Macrophage N IL-12
ends
Naive T helper cell
I!
Th2 cell
.itic cell
Fig. 1. Cytokines generated by cells of the innate immune response drive CD4 f Th cell subset differentiation. Pathogens and their prod- ucts interact with several components of the innate immune system, including macrophages, NK cells, y6 T cells and/or basophils/mast cells. Some pathogens (quite frequently bacteria, protozoa and viruses) interact with macrophages, which stimulates productron of IL-12 and enhances EN-y production by NK cells and/or y6 T cells and promotes differentiation of naive Th cells towards the Thl-cell phe- notype. Other pathogens (frequently helminths) stimulate cells to produce IL-4 that drives Th2-cell differentiation. The cells producing IL-4 during the early stages of infection have not been defined but could include basophils or mast cells, y6 T cells, or an undefined cell population. All of the in vitro models suggest that the most efficient APCs are dendritic cells.

394 lymphocyte activation and effector functions
tion of the TCR ligand and the co-stimulatory molecules needed to be on the same cell for efficient T-cell activa- tion [55-l.
Influence of antigen on differentiation
The relationship between different pathogens and the in- duction of distinct Th-cell subsets suggests that distinct epitopes might induce the development of a particular Thcell subset. However, most evidence indicates that differences at the epitope level are not a major fac- tor in determining which Th-cell subset predominates. For example, following stimulation with anti-CD3 mAb isolated T-cell clones could differentiate in either di- rection, depending upon culture conditions [ 56*]. Sim- ilarly, in the TCR transgenic models either Thl or Th2 cells can develop [ 17**,18**]. In addition, in the leishma- nial system Thl- and Th2-cell clones bearing the same TCR have been derived from infected mice [57**]. The paramount importance of the cytokine environment at the time of primary stimulation is further demonstrated by the switch of the m-cell response to sperm whale myoglobin from a Thl- to a Th2-cell response when animals are infected with S. munsoni [58*]. Notwith- standing these observations, recent data obtained using a peptide derived from human Type IV collagen suggest that certain characteristics of the immunogenic peptide may influence subset selection [4,59*]. Thus, when H-2s mice are immunized with collagen peptide a Thl-cell re- sponse develops, whereas H-2b and H-2d mice develop a Th2-cell response. The density of the ligand on the APC, which is a function of the affinity between the processed peptide and class II molecules, may mediate these results [4,59’]. If this is the case it suggests that in certain in- stances the epitope may indeed influence which subset predominates by dictating the tinity of the interaction between class II and itself. In this regard, the contin- ued presence in culture of a anti-V@3 mAb specifically enhanced IL-4 production in the Vp8 T-cell population, suggesting that continuous ligation of the TCR might influence subset development [bO*] Finally, one of the oldest factors known to inkluence the development of Th-cell subsets is antigen dose. This has been reinforced by the recent demonstration that low dose challenge in the murine leishmanial system induces a cell mediated immune response that leads to resistance to reinfection [61-l. This is an important observation that may have a significant influence on the development of vaccines [62]. Although the mechanism mediating this effect re- mains undefined, it may relate to some of the in vitro observations described above.
Conclusion
Efforts to define the factors responsible for the induction of distinct Th-cell subsets have accelerated over the last
year. One of the major advances was the recognition that cytokines generated by cells belonging to the innate im- mune system are the principle signals for Th-cell subset differentiation (Fig. 1). However, several issues have yet to be addressed. These include determining the source of IL-4 during the innate immune response, providing a model encompassing the roles of IFN-)I and IL-12 in vivo and in vitro, further defining the role of other cytokines (e.g. IL-l, IL-IO, TGF-p) in Th-cell development, and de- termining at the molecular level how particular antigen characteristics, including antigen concentration and epi- tope affinity for MHC class II molecules, influence Th cell subset selection. Nevertheless, the rapid advances in this field suggest that in the not too distant future application of our knowledge of T cell subset selection will dramati- cally improve immunotherapy and vaccine development.
Acknowledgements
The author wishes to thank: C-S Hsieh, SE Macatonia, CS Tripp, SF Wolf, A O’Garra, KM Murphy and A Sher for providing data from submitted manuscripts for inclusion in this raiew; K Varkila, R Chate- lain, LMCC meal, RL Coffman, R Manetti, P Parronchi, MD Guidizi, MP Piccini, E Ma@, G Trinchieri, S Romagnani, A Sher, 1 Oswald, S Heiny and RT Gazzinelli for providing copies of manuscripts in press; and Drs J Farrell, L Vieira and T Scharton for reading the manuscript. The author is supported by NIH grant AI30073.
References and recommended reading
Papers of particular interest, published within the annual period of review, have been highlighted as: . . . 1.
2.
3.
4.
5.
6.
7.
8.
of special interest of outstanding interest
ABBAS AK, WUAMS ME, BURSI’EIN HJ, CHANG T~L, Bossu P, LICHTMAN AH: Activation and Functions of CD4+ T-cell Sub- sets. Immunol Rev 1‘991, 123:5-22.
PELTL G: A Role for CD4+ T-cell Subsets Producing a Selec- tive Pattern of Lymphokines in the Pathogenesis of Human Chronic Itiammatory and Allergic Diseases. Immunol Rev 1991, 123:23-35.
FOWELL D, MCKNKXT AJ, POWRIE F, DVKE R, MAsON D: Subsets of CD4+ T Cells and Their Roles in the Induction and Prevention of Autoimmunity. Immunol Rev 1991, 123:376%
PFEIFFER C, MURRAY J, MADR~ J, BOTTOMLY K: Selective Acti- vation of Thl- and ThZ-like Cells in Viva-Response to Human Collagen N. Immunol Rev 1991, 123:65-84.
KEISO A, TR0Ut-r AB, MARksKOVSKv E, GOLJGH NM, MORRIS L, PECH MH, THOMSON JA: Heterogeneity in Lymphokine Profiles of CD4+ and CD8+ T Cells and Clones Activated in Vivo and in Vitro. Immunol Rev 1991, 123:8%114.
SWAIN SL, BRADLEY LM, CROFT M, TONKONOCY S, ATKINS G, WEINBERG AD, DUNCAN DD, HEDRICK SM, DU~ON RW, HUSTON G: Helper T-cell Subsets: Phenotype, Function and the Role of Lymphokines in Regulating Their Development. Immunol Rev 1991, 123:115144.
HAYAKAWA K, HARDY RR: Murine CD4+ T-cell Subsets. Im- munol Rev 1991, 123:145-l&3.
BENDE~AC A, SCHWARTZ RH: ThO Cells in the Thymus: The Question of T-helper Lineages. Immunol Rev 1991, 123:16F188.

Selective differentiation of CD4+ T helper cell subsets Scott 395
9. COFFMAN RI, VAIWLA K, SCOTT P, CHATEWN R: Role of Cy- tokmes in the Differentiation of CD4+ T-cell Subsets in Vito. Immunol Rev 1991, 123:189207.
10. MOSMANN TR, SCHUMACHER JH, STREET NF, BUDD R, O’GARRA A, FONG TAT, BOND MW, MOORE KWM, SHER A, FIORENTINO DF: Diversity of Cytoklne Synthesis and Function of Mouse CD4+ T Cells. Immunol Rev 1991, 123:203-229.
11. MAC&I E, PARRONCHI P, MANETTI R, SIMONELLI C, PICCINNI Mu . P, RUGIU FS, DE CAFXI M, RICCI M, ROMAGNANI S: Reciprocal
Regulatory Effects of IFN-y and IL-4 on the in Vitro Devel- opment of Human Tbl and Tb2 Clones. J Immunol 1992, l&:2142-2147.
It is demonstrated that IFN-Y inhibits the expansion of Th2-cell clones, whereas IL-4 promotes the expansion of T-cell clones capable of pro- ducing IL-4.
12. SHER A, COFFMAN RL: Regulation of Immunity to Parasites by . T Cells and T Cell-derived Cytokines. Annu Rev Immunol
1992, 10:385X)9. This review provides an excellent overview of the interactions between pathogens and the immune response.
13. JANWAY CA JR: The Immune System Evolved to Discriminate Infectious Nonself from Noninfectious Self. Immunol Today 1992, 13:llp16.
14. ROMAGNANI S: Induction of Thl and Th2 Responses: A Key Role for the ‘Natural’ Immune Response? Immunol Today 1992, 133379383.
15. SWAIN SL, WELNBERG AD, ENGUSH M, HUSTON G: IL-4 Directs the Development of TbZ-Uke Helper Effecters. J Immunol 1990, 145:379&3806.
16. LE GROS G, BEN-SASSON SZ, SEDER R, FINKELMAN FD, PAUL WE: Generation of Interleukin 4 (IL-4) Producing CeUs in Vivo and in Vitro IL-2 and IL-4 are Required for in Vitm Gener- ation of IL-4-producing Cells. J Exp Med 1990, 172:921-929.
17. HSIEH CS, HELMBERGER AB, GOLD J, O’GARRA A, MUWHY K: Dif- . . feremial Regulation of T Helper Phenotype Development
by Interleukins 4 and 10 in an up T Cell-receptor Trans- genie System. Proc Natl Acad Sci USA 1992, 89:6065-6069.
This, together with [ 180*], demonstrates the critical role IL-4 plays dur~ ing activation of naive CD4+ T cells with conventional antigens.
18. SEDER RA, PAUL WE, DAVIS Mh4, FAZEKAS DE ST GROTH B: . . Tbe Presence of Interleukin 4 During in Vitro Priming
Determines the Lymphokine-producing Potential of CD4 + T Cells from T CeU Receptor Transgenic Mice. J Exp Med 1992, 176:1091-1098.
see [17**1.
19. . .
CROF? M, DUNCAN DD, SWAIN SL: Response of Naive Antigen-specific CD4+ T Cells in Vitro Characteristics and Antigen-presenting CeU Requirements. J Exp Med 1992, 176:1431-1437.
This paper, in combination with [ 17**,18°*], demonstrates the super rior antigen-presenting capacity of dendritic cells for primary stimula~ tion of CD4 + cells.
20. HSIEH CS, MACATON~A SE, TRIPP CS, WOLF SF, O’GARRA A, . . MURPHY KM: Listeria-induced Thl Development in up-TCR
Transgenic CD4+ T Cells Occurs Through Macrophage Production of IL-12. Science 1993, in press.
It is shown that ListerM-infected macrophages drive the differentiation of naive T cells towards the Thl phenotype in an in vitro TCR trans- genie system. This effect is demonstrated to be due to the production of IL-12 by macrophages following infection with Listerziz
21. &cKSL!ZY RM, Sco’rr P: Helper T-cell Subsets in Mouse Leish- manlasii: Induction, Expansion and Effector Function. Irn~ munol Today 1991, 12A5sA61.
22. SCOT P: IFN-y Modulates the Early Development of Thl . and Th2 Responses in a Murlne Model of Cutaneous Leish-
maniasis. J Immunol 1991, 147:31493155. This paper demonstrates the critical role of IFN-1 in Thl-cell develop- ment in experimental leishmaniasis.
23. HEINZEL FP, SADICK MD, HOLADAY BJ, COFFMAN RL, IXKSL!ZY RM: Reciprocal Expression of Interferon y or Interleukin 4 During the Resolution or Progression of Murlne Lelsluna- rdasis. J Exp Med 1989, 169:59.
24. CHATEWN R, VARIKILA K, COFFMAN RL: IL-4 Induces a Th2 . Response in Leishmania major-infected Mice. J Immunol
1992, 148:1182-1187. The authors demonstrate that IL-4 is required for Th2-cell develop- ment in experimental leishmaniasis, and found that injections of IL-4 promoted Th2-cell development in viva.
25. ROMANI L, MENCACCI A, GROHMANN U, MOCCI S, Moscl P, PUCCET~ P, BISTONI F: Neutralizing Antibody to Interleukin 4 Induces Systemic Protection and T Helper Type l-asso- ciated Immunity in Murine Candid&is. J Exp Med 1992, 176:1%25.
26. HAAK-FREDNSCHO M, BROWN JF, IIZAWA Y, WAGNER RD, CZLJPRYNSKI CJ: Administration of Anti-IL-4 MonoclonaI An- tibody llBl1 Increases the Resistance of Mice to Listeria monocytogenes Infection. J Immunol 1992, 148:397%3985.
27. ABEHSIRA-AMAR 0, GIBERT M, JOLN M, THEZE J, JANKOVIC DL: . IL-4 Plays a Dominant Role in the Differential Develop-
ment of ThO into Thl and Th2 Cells. J Immunol 1992, 148:382c3829.
ThO clones are generated in vitro they can be driven towards a Th2 phenotype in the presence of IL-4, or towards a Thl phenotype in the presence of IFN-Y and anti-IL-4 antibody.
28. SCHARTON TM, SCOTT P: Immediate IFN-1 Production in . C3H/HeN Mice Infected with Leisbmaniu major. In
Cytokines and Resistance to Nonviral Pathogenic Infections Edited by Sonnenfeld G, Czamlecki C, Nacy C, Byrne G, Degre M. Augusta GA: The Biomedical Press; 1992:103-110.
This study demonstrates that NK cells are activated fo produce IFN~y 2 days after infection of resistant mice with Letimania.
29. VARK~~A K, CHATEWN R, LEAL LMCC, COFFMAN RL: Reconsti- . tution of CB-17 scid Mice with BALB/c T Cells Initiates a
T Helper Type-l Response and Renders them Capable of Healing Leishmania major Infection. Eur J Immunoll993, in press.
In this study it was shown that naive CD4+ T cells transferred to SCID mice differentiate into Thl cells following infection with Leishmania. However, treatment of the recipients with anti-IFN y-antibody led to ThZ-cell development. This, together with [22-j demonstrates the criti- cal role IFN-y plays in the development of Thl cells following infection with Leisbmania.
30. SEDER RA, PAUL WE, DVORAK AM, SCHARKJS SJ, KAGF~X~OBOTKA . A, NIV J, FINKELMAN FD, BARBIERI SA, GALU SJ, P~AUT M: Mouse
Splenic and Bone Marrow CeU Populations that Express High Affinity Fca Receptors and Produce Interleukin 4 are Highly Enriched in Basopbils. Proc Nat1 Acad Sci USA 1991, 88:2835.
It is shown that an Fc&R+ cell population that produces large amounts of IL-4 is enriched for basophils. These data implicate basophils as an important source of IL-4 (other than T cells) that then promote Th2-cell dominance.
31. VEUA AT, PEARCE EJ: CD4+ Th2 Response Induced by Scbis- . tosoma mansoni Eggs Develops Rapidly, Through an Early,
Transient, ThO-like Stage. J Immunol 1992, 148:2283-2290. See [32-l.
32. VELLA AT, HUISEBOSCH MD, PEARCE EJ: Schistosoma mansoni . Eggs Induce Antigen-responsive cD44hi T Helper 2 Cells
and IL-4-secreting CD4’0 Cells. Potential for T Helper 2 Subset Differentiation is Evident at the Precursor Level. J Immunol 1992, 149:1714-1722.
This paper, together with [31*], characterizes the initial cytokine re- sponse to S. mansoni eggs, and this study indicates that CD44’o cells may be a source of IL-4 driving Th2~cell differentiation.
33. HIROMA’I’SU K, YOSHIKAI Y, MATSU~XKI G, OHGA S, MURAMORI K, MATSUMOTO K, BLUESTONE JA, NOMOTO K: A Protective Role of y6 T Cells in Primary Infection with Listeriu monocy togenes in Mice. J Exp Med 1992, 175:4!+56.

3% Lymphocyte activation and effector functions
34. U~EMURA K, ~TZ J, PIRMEZ C, OHMEN J, WANG X-H, Ho C, HOFFMAN WL, MODUN RL: Microanatomic Clonality of y6 T Cells in Human Leishmaniasis Lesions. J Immunol 1’992, 148:120>1211.
35. ROSAT J-P, MACDONALD HR, LOUIS JA: A Role for y6 T Cells . During Experimental Infection of Mice with Leishmania
major J Immunol 1993, 150:55&555. This is the first study demonstrating that y6 T cells are activated dur- ing experimental leishmaniasis, although whether these cells promote resistance or susceptibility remains to be determined.
36. CARDING SR, ALIAN W, KVES S, HAYDAY A, BOT~OMLY K, DOHERTY PC: Late Dominance of the Intkimmatory Pro- cess in Murine Influenza by y6+ T Cells. J Exp Med 1990, 172:122>1231.
37. GAJEWXI TF, FITCH FW: Anti-proliferative Effect of IFN-y in Immune Regulation. I. IFN-y Inhibits the Proliferation of Th2 but not Thl Murine Helper T Lymphocyte Clones. J Immunol 1988, 140:4245.
38. D’ANDREA A, RENGARAJU M, VAUANTE NM, CHEMHIMI J, KUBIN . M, AXE M, CHAN SH, KOBAYASHI M, YOUNG D, NICKBARG E,
ET AL.: Production of Natural Killer Cell Stimulatory Factor (NKSF) by Peripheral Blood Mononuclear Cells. J Exp Med 1992, 176:1387-1398.
The authors show that both adherent (monocytes) and non-adherent cells produce IL-12 in reponse to stimulation with S. aureus and that IL-12 is one of the major factors mediating production of IFN-y by cells from the peripheral blood.
39. PERUSSIA B, CHAN SH, D’ANDREA A, TSUJI K, SANTOU D, POSPISIL M, YOUNG D, WOLF SF, TRINCHIERI G: Natural Killer (NK) Stimulatory Factor or IL-12 has Differential Effects on the Proliferation of TCR-up+, TCRyG+ T Lymphocytes, and NK Cells. J Immunol 1992, 149:34993502.
40. KOBAYASHI M, Fm L, RVAN M, HEW~CK RM, Cwuc SC, CHAN S, L~UD~N F, SHERMAN B, PERUSSIA B, TR~NCHIERI G: Identi- fication and Purification of Natural Killer CeU Stimulatory Factor (NKSF), a Cytokine with Multiple Biologic Effects on Human Lymphocytes. J E3cp Med 1989, 170:827-820.
41. STERN AS, PODIAXI FJ, HULMES JD, PAN YE, QUINN PM, WOLITZKV AG, FAMILLE’I-I’I PC, STREMLO DL, TRUITI T, CHJZZONITE R, ET AL: Purification to Homogeneity and Par- tial Characterization of Cytotoxic Lymphocyte Maturation Factor from Human B-Lymphoblastoid Cells. PYOC Nutl Acud Sci USA 1990, 8768086800.
42. SCHOENHALJT DS, CHUA AO, WouTwy AG, QUINN PM, DWVER CM, MCCOMA~ W, FAMILLET-II PC, GATELY MK, GUBLER U: Cloning and Expression of Murine IL-12. J Immunol 1992, 148:3433-3440.
43. MANET~ R, PARRONCHI P, GIUDIZI MG, PICCINNI M-P, MAGGI E, . . TRINCHIEIU G, ROMAGNANI S: Natural Killer CeU Stimulatory
Factor (NKSF/IL-12) Induces Thl-type Specific Immune Re- sponses and Inhibits the Development of IL-4 Producing Th Cells. J Exp Med 1993, 177:119’+1204.
This is the first study to demonstrate a role for IL-12 in the regulation of T-cell subsets.
44. SHER A, OSWALD I, HIENY S, GAWNELU RT: Toxoplasma . gondii Induces a T-independent IFN-y Response in NK
Cells Which Require Both Adherent Accessory Cells and TNF-a. J Immunol 1993, in press.
This paper establishes the link between macrophages and NK cells for the production of IFN-y foUowing stimulation with the proto- zoan Tox@hsma. The study describes a soluble factor derived from the macrophages that drives NK-cell stimulation,
45. BARRAL-NE’ITO M, BARRAL A, BROWNELL CE, SKEIKV YAW, . ELLINGSWORTH LR, T~ARDZIK DR, REED SG: Transforming
Growth Factor-p in Leishmanial Infection: A Parasite Es- cape Mechanism. Science 1992, 257:545-548.
The authors demonstrate that TGF-P augments susceptibility to Le&b mania, establishing TGF-p as a cytokine that may either drive Th2-cell differentiation, or inhibit Thl-cell development, in uivo.
46. SWAIN SL, HUSTON G, TONKONCGY S, WEINBERG A Transform- . ing Growth Factor-p and IL-4 Cause Helper T CeU Pre-
cursors to Develop into Distinct Effector Helper Cells that Dither in Lymphokine Secretion Pattern and Cell Surface Phenotype. J Immunol 1991, 147:299-3000
47. FARGEA~ C, Wu CY, NAKAJIMA T, Cox D, NLITMAN T, DELESPESSE . G: DilTerential Effect of Transforming Growth Factor p on
the Synthesis of Thl- and ThZ-like Lymphokines by Human T Lymphocytes. Eur J Immund 1992, 222173-2176.
These studies indicate that TGF-fl inhibits IL-4 and IL-5 synthesis by human T cells and has no effect on IFN-1 production. This contradicts the in viva results reported in [45*]. The differences in these results may relate to the presence of NK cells in the in vi00 model.
48. Su HC, LEITE-MORRIS KA, BRAUN L, BIRON CA: A Role for Trans- . forming Growth Factor-p 1 in Regulating Natural Killer Cell
and T Lymphocyte Proliferative Responses During Acute Infection with Lymphocytic Choriomeningitis Virus. J Im- munol 1991, 147~2717-2727.
These authors demonstrate that the down-regulation of the NKceU re- sponse in viral infections is due to the expansion of a TGF-p producing T~cell population,
49. LUQMAN M, GREENBAUM L, Lu D, BO~TOMLY K: DBerentiaI . Effect of Interleukin 1 on Naive and Memory CD4+ T
Cells. Eur J Immunol 1992, 22:95-100. This study shows that IL-1 has no effect on the response of naive CL%+ (cD45@i) T cells, a result which is compatible with the authors find- ing that naive CD4+ T cells fail to express IL-l receptors.
50. KOCH K-C, Cwu< BD, DINARELLO CA: Interleukin4 (IL)4 Up- regulates Gene and Surface IL-1 Receptor ‘Qpe 1 in Murine T Helper Type 2 Cells. Eur J Immunol 192, 22:153_157.
51. LIU Y, JONES B, ARLJFFO A, SULWXN KM, L~NSIEY PS, JANEWAY . C JR: Heat-stable Antigen is a Costimulatory Molecule for
CD4 T Cell Growth. J EJCP Med 1992, 175:437-445. This reference, together with [52*-54-l, identifies HSA and B7 as im- portant co-stimulatory molecules for murine T~cell activation.
52. Lru YU, JONES B, BRADY W, JANEWAY CA JR, LINLFY PS: Co-sthn- . ulatin of Murine CD4 T Growth: Cooperation Between B7
and Heat-Stable Antigen. Eur J Immunoll992, 22:2855-2859. See [51*].
53. RAZ-WOLF Z, FREEMAN GJ, GALVIN F, BENACERRAF B, NADLER . L, REISER H: Expression and Function of the Murine B7
Antigen, the Major Costimulatory Molecule Expressed by Peritoneal Exudate Cells. Proc Nat1 Acad Sci USA 1992, 89:42104214.
See [51*].
54. LARSON CP, R~TCHIE SC, PEARSON TC, LINSLF~ PS, I~WRY . RP: Functional Expression of the Costimulatory Molecule,
B7/BBl, on Murine Dendritic CeU Populations. J Exp Med 1992, 176:1215-1220.
See [51*].
55. LIU Y, JANEWAY CA JR: CeUs that Present Both Specific Ligand . and Costimulatory Activity are the Most Efficient Inducers
of Clonal Expansion of Normal CD4 T Cells. Proc Nat1 Acud Sci IJSA 1992, 89:3%5-3849.
In a series of studies using three distinct experimental approaches, the authors demonstrate that activation of naive CD4+ T cells is most efficient when both the T-cell receptor l&and and the costimuatory molecule are presented on the same antigen-presenting cell. Minimal activation of CD4+ T cells occurs when these molecules are presented on separate cells.
56. R~CKEN M, SAURAT J-H, HAUSER C: A Common Precursor for . CD4+ T Cells Producing IL-2 and IL-4. J Immunol 1992,
148:1031-1036. Primary CD4+ T-cell clones were split into two cultures soon after in- fection Continuous ligation of the TCR with anti-CD3 antibody led to the development of T cells with a Th2 phenotype, whereas cells stim- ulated with IL-2 only became Thl-like (see also [bo.]).
57. REINER SL, WANG Z-E, HATAM F, SCOTT P, LOCKsLF( RM: COm- . . mon Lineage of Thl and Th2 Subsets in Leishmaniasis.
Science 1993, 259:1457-1460.

Selective differentiation of CD4+ T helper cell subsets Scott 397
It is shown that following L major infection or immunization, there is 60. ROCKEN M, MULLER KM, SAURAT JH, MULIER I, Louts JA,
preferential expansion of T cells bearing a particular TCR. T-cell clones . CEROTFINI J-C, HAUSER C: Central Role for TCRKD3 Ligation with a Thl or Th2 profile, using the identical TCR could be identified. in the Differentiation of CD4+ T CeUs Toward a Thl or These studies demonstrate that the antigen may be less important than Th2 Functional Phenotype. J Immunoi 1992, 148~47-54. the manner of presentation in determining the subsequent Th-cell phe- This study reports that continuous ligation of the TCR during in llitro notype. culture can influence subset differentiation.
58. KULLBERG MC, PEARCE EJ, HIENY SE, SHER A, BERZOFSKY JA:
. Infection with Schis#osoma mansoni Alters Tbl/TbZ Cy- tokine Responses to a Non-parasite Antigen. J Immunol 1W2, 148:3264-3270.
61. . .
BRETSCHER PA, WEI G, MENON JN, BIELEFELDT-OHMANN H: Es- tabhshment of Stable, Cell-mediated Immunity that Makes ‘Susceptible’ Mice Resistant to Leishmania major. Science 1992, 257:53?542.
This paper demonstrates that the strong ThZ-cell response associated with S. mansoniinfection can influence Th-cell subset differentiation to an unrelated antigen.
‘The authors demonstrate that low doses of Leishmurziu can induce pro- tective immunity (Thkeil responses) in animals that are normally sus ceptible and develop a Th2~cell response.
59. MURRAY JS, PFEIFFER C, MADIU J, BOTOMLY K: Major Histocom- . patibiliry Complex (MHC) Control of CD4+ T CeII Subset
Activation. II. A Single Peptide Induces Either Humoral or Cell-mediated Responses in Mice of Distinct MHC Geno- type. Eur J Immunol 1992, 22:55?565.
This paper extends previous studies (see [4]) on the ability of human Type IV collagen to induce Thl~ or Th2~cell responses depending on the haplotype of the immunized mice. Here the authors show that they can repeat these observations with a single peptide.
62. RRETSCHER PA A Strategy to Improve the Efficacy of Vac- cination Against Tuberculosis and Leprosy Imn~zrnol 7b&,lj 1992, 13:342-345.
P Scott, Department of Pathobiology, School of Veterinary Medicine, 3800 Spruce Street, Philadelphia, Philadelphia 19104, USA.














![Stochastic Cytokine Expression Induces Mixed T Helper Cell ...web.mit.edu › biophysics › papers › PLOS2013.pdf · PU.1 versus Gata1 [2]; naive CD4 T cells differentiate into](https://img.pdfslide.us/doc/110x75/5f2618f0ce8dba3e521981af/stochastic-cytokine-expression-induces-mixed-t-helper-cell-webmitedu-a-biophysics.jpg)













