Embed Size (px)
Citation preview

SELECTION OF Zys2 MUTANTS OF THE YEAST SACCHAROMYCES CEREVZSZAE BY THE UTILIZATION OF a-AMINOADIPATE
BHARAT B. CHATTOO1 and FRED SHERMAN
Department of Radiation Biology and Biophysics, Uniuersity of Rochester School of Medicine, Rochester, N e w York 14642
AND
DALIA A. AZUBALIS, THORSTEN A. FJELLSTEDTZ, DAVID MEHNERT and MAURICE OGUR3
Department of Microbiology, Southern Illinois Uniuersity, Carbondale, Illinois 62901
Manuscript received April 12, 1979
ABSTRACT
Normal strains of Saccharomyces cereuisiae do not use a-aminoadipate as a principal nitrogen source. However, a-aminoadipate is utilized as a nitro- gen source by lys2 and lys5 strains having complete or partial deficiencies of a-aminoadipate reductase and, to a limited extent, by heterozygous lys2l + strains. Lys2 mutants were conveniently selected on media containing a-aminoadipate as a nitrogen source, lysine, and other supplements to furnish other possible auxotrophic requirements. The lys2 mutations were obtained in a variety of laboratory strains containing other markers, including other lysine mutations. In addition to the predominant class of Zys2 mutants, low frequencies of lys5 mutants and mutants not having any obvious lysine re- quirement were recovered on a-aminoadipate medium. The mutants not requiring lysine appeared to have mutations at the lys2 locus that caused partial deficiencies of a-aminoadipate reductase. Such partial deficiencies are believed to be sufficiently permissive to allow lysine biosynthesis, but suf- ficiently restrictive to allow for the utilization of a-aminoadipate. Although it is unknown why partial or complete deficiencies of a-aminoadipate reductase cause utilization of a-aminoadipate as a principal nitrogen source, the use of a-aminoadipate medium has considerable utility as a selective medium for lys2 and lys5 mutants.
NUMEKOUS genetic studies are facilitated by convenient methods for select- ing both fonvard and reverse mutations at a given locus. Several selective
systems have been based on physiological properties associated with specific mu- tant genes. Media used for selecting forward and reverse mutations in normal strains of Saccharomyces cereuisiae include chlorolactate medium for obtaining cycl mutants, lactate medium for obtaining cycl revertants (SHERMAN et al.
Present address: Sarabhai Research Centre, Post Box 162, Baroda 390 007, INDIA. 2 Present address: National Institute of Child Health and Human Development, Bethesda,
3 Deceased. Maryland 20014.
Genetics 93: 51-65 September, 1979

52 B. B. CHATTOO et al.
1974), medium containing methyl mercury for obtaining met15 and met2 mu- tants, and medium lacking methionine for obtaining met revertants (SINGH and SHERMAN 1974, 1975). Special strains and special conditions have been devised for selecting and reverting can1 mutants (WHELAN, GOCKE and MANNEY 1979), ural, ura3 and ura5 mutants (BACH and LACROUTE 1972), ade3 through ade8 mutants (ROMAN 1956), and gal mutants (DOUGLAS and HAWTHORNE 1964). Although there are no selection procedures, adel and ade2 mutants, which can be detected by their characteristic red or pink color, have been used in numerous mutational studies (see MORTIMER and MANNEY 1971). In this paper, we describe a method that we believe is more convenient and more effi- cient than any other previously reported for isolating a specific class of nutri- tional mutants; most mutants selected for their ability to use a-aminoadipate as a principal nitrogen source are also auxotrophic for lysine due to mutation at the Zys2 locus.
Normal S. cereuisiae strains differ from some other yeast species in being gen- erally able to utilize glutamate, but not lysine, as a principal nitrogen source for growth (SCHULTZ and POMPER 1948; THORNE 1950; WALTERS and THISEL- TON 1953; MORRIS and EDDY 1957; LARUE and SPENCER 1968). a-Aminoadipate is an intermediate of lysine biosynthesis that might be expected to serve in place of glutamate, since in cell free preparations, unlike lysine, it serves as a nitro- gen donor in a simple transamination of a-ketoglutarate to produce glutamate ( BROQUIST and STIFFEY 1959; DEBOEVER 1963; FJELLSTEDT 1966; PIEDISCALZI, FJELLSTEDT and OGUR 1968; MATSUDA and OGUR 1969a,b). Contrary to expec- tation, however, it was observed that all strains originally tested, except for Zys2 mutants, failed to grow on media containing a-aminoadipate as a principal nit- rogen source (AZUBALIS 1970). This finding was the basis for the method of isolating ZysZ and Zys5 mutants. In this paper, we present evidence that muta- tions at the Zys2 and Zys5 loci that causing complete or partial loss of a-amino- adipate as a principal nitrogen source for growth, and we describe appropriate media that can be used conveniently to isolate these mutants.
MATERIALS A N D METHODS
Yeast strains and genetic procedures: The numerous strains used in this investigation were obtained from the Carbondale, Rochester and Berkeley collections. Most of the Zys2 mutants isolated in this investigation were derived from the ‘hormal” strains AS2-2A (a t r p l - I ) (SINGH and SHERMAN 1975), MO-59-13A (a) (FJELLSTEDT and OGUR 1970) and MO-320- IC (a). Conventional yeast genetic procedures of crossing, sporulation and tetrad analysis were carried out by methods outlined by SHERMAN and LAWRENCE (1974).
Culture media: Routine synthetic and complex nutrient media used in this study were those described by either SHERMAN and LAWRENCE (1974) or FJFLLSTEDT and OGUR (1970). Unless stated otherwise, the synthetic media contained ammonium sulfate as the principal nitrogen source. The following amounts of the other principal nitrogen sources were added to other types of media: glutamate, 100 to 500 mg per 1; L-lysine, 20 to 100 mg per 1; and D, L-a- aminoadipic acid, 1.0 to 2.0 g per 1. The standard solid AA media contained 0.167% Bacto- yeast nitrogen base without acids and without ammonium sulfate, 2% dextrose, 0.2% D, L-a- aminoadipic acid, 30 mg per 1 L-lysine, 2% Bacto-agar, and other standard supplements that were required by the various auxotrophic strains. The a-aminoadipate was added aseptically to

LYSINE MUTANTS O F YEAST 53 the other autoclaved ingredients of the AA media by first preparing a 6% stock solution of a-aminoadipic acid, adjusting the p H to 6.0 with 1 M KOH, filter-sterlizing the solution and thcn adding 33 ml for each liter of AA media.
Growth curves: Freshly grown cells were used to inoculate 50 ml of liquid nutrient medium in a 250 ml Erlenmeyer flask. After 24 hr on a reciprocating shaker at 30", the cells were harvested by centrifugation and washed three times with sterile distilled water. After esti- mating the cell density with a counting chamber, appropriate inocula were introduced into screw cap Klett tubes containing 8 ml of growth medium. Cultures were shaken in a slanted position during incubation at 30". Growth rate was estimated using the Klett-Summerson photoelectric colorimeter equipped with the blue no. 42 filter.
Mutant cultures were checked for revertant frequencies by plating on appropriate media and these frequencies were always less than 6 x 10-7.
Preparation of extracts: Seed cultures, grown for 24 hr in nutrient medium on a shaker, were harvested, washed and reinoculated into 6 to 24 1 of appropriate growth media for a 16 to 18 hr incubation. Cells were harvested, washed, sampled and prepared for sonication. Cells were broken by sonication as previously described (FJELLSTEDT and OGUR 1970), and the homogenates were centrifuged at 20,000 x g for 40 min at 0 to 4" in an RC2B centri- fuge (DuPont-Sorvall, Inc., Norwalk, Connecticut) using a SS34 rotor. After centrifugation, the pellet was discarded. The supernatant fluid was dialyzed 14 to 18 hr against 4 1 of phos- phate-citrate-glutathione buffer, p H 7.0, which contained the following ingredients adjusted to pH 7.0 with KOH: 50 mM K,PO,, 0.4 mM sodium citrate and 1 mM glutathone. The dialyzed homogenate was concentrated to one-half its volume using Lyphogel (Gelman In- strument Co., Ann Arbor, Michigan). Protein content was estimated by the method of WADDELL (1965) and occasionally checked by the method of LOWRY et al. (1951).
Radioactive assay of transaminase activity in homogenates: Glutamate: a-ketoadipate transaminase activity was assayed in the direction from a-aminoadipate to a-ketoadipate (FJELL- STEDT and OGUR 1970). A total of 0.035 ml of the reaction mixture contained the following except where otherwise indicated: 0.7 pmole a-ketoglutarate; 0.7 mg pyridoxalphosphate; 0.1 13 pmole [6-I4C] D,L-a-aminoadipic acid (sp. act. 4.40 PCi per pmole); and 15 ~1 of homogenate. Cofactors and substrates were prepared in 0.5 M phosphate buffer, p H 7.0.
Conditions of incubation, sampling and chromatography were as previously described (PIEDISCALZI, FJELLSTEDT and OGUR 1968; FJELLSTEDT and OGUR 1970). Reference standards of glutamate, glutarate, a-ketoadipate and a-aminoadipate were chromatographed in each of four solvent systems used. Nonradioactive compounds were located by staining procedures. Nin- hydrin-positive spots were obtained by spraying strips with a solution of 0.2% ninhydrin in acetone and, after solvent evaporation, the color was developed by heating gently. a-Keto acids were identified as yellow-green spots, fluorescent under UV (WIELAND and FISCHER 1949) after spraying the strips with a solution of 0.05% o-phenylenediamine in 10% aqueous tri- chloracetic acid and heating for two min at 100".
Radioactive strips were scanned with a Nuclear-Chicago Radiochromatogram Strip Scanner (Actigraph 111). Counting parameters were as follows, unless otherwise indicated: Range 1000 cpm; high voltage, 950 V; scan speed, 60 cm/hr; background suppression, none; time con- stant, 20 sec; collimator slit width, 3.0 mm.
Spectrophotometric assay of a-aminoadipate reductase activity: a-Aminoadipate reductase activity was assayed by following NADPH oxidation at 340 nm (FJELLSTEDT 1969), using a Gilford 2000 recording spectrophotometer (Gilford Instrument Laboratories, Inc.) equipped with auxiliary offset control. A total of 3.0 ml of the reaction mixtures generally contained the following: 1.5 to 10 Pmole D,L-a-aminoadipate; 10 p o l e ATP; 10 pmole Mg++; 1.0 /."le NADPH; 0.1 M Tris buffer, pH 7.5; and 0.3 ml of the concentrated, dialyzed homo- genate preparation.
a-dminoudipate uptake and metabolism by intact cells: The normal, aau (see RESULTS) and lysZ cultures were grown in media that would be expected to induce maximum activity of the enzymes mediating a-aminoadipate utilization. The mu strains were grown in AA

54 B. B. CHATTOO et al. medium lacking lysine. The Zys2 strain (MO-49-14D) was grown in AA medium. The cultures were harvested in early stationary phase and the harvested cells were washed in 0.067 M K,PO, containing 3% glucose and adjusted to comparable cell density. A 0.5 ml suspension of cells of each strain was then incubated with 0.1 ml of [6-14C] D,L-a-aminoadip- ate (50 $3 per ml) for two to six hr in different experiments. The cells were collected by centrifugation, suspended in 1.0 ml of water, and heated in a boiling water bath for 20 min. After centrifugation, 20 Pl of the hot water extract was spotted on paper chromatographic strips. Comparable portions taken from the incubation medium at various incubation times were similarly spotted. Five different solvent systems were used and lysine, saccharopine, a- ketoadipate, a-aminoadipate, and glutarate were used as reference standards.
RESULTS
Growth of strains on AA media: Numerous normal strains and various lysine auxotrophs were tested for the ability to grow on solid synthetic media contain- ing various principal nitrogen sources and various supplements. All of the strains could grow on medium containing ammonium sulfate or glutamate as principal nitrogen source, but none grew on medium containing lysine as a prin- cipal nitrogen source. When the synthetic medium was supplemented with ly- sine, a-aminoadipate could serve as a principal nitrogen source for Zys2 and Zys5 auxotrophs, but not for normal strains or for other lysine auxotrophs blocked at the other loci, Zysl, Zys2,Zys4, and Zys6 through ZyslO. Some Zys5 mutants grew as well as Zys2 mutants, while other Zys5 mutants exhibited only partial growth. Measurements of growth in liquid media indicated that the yield of cells of Zys2 strains increased with increasing amounts of a-aminoadipate up to supplements of approximately 0.9 to 1.0 gm per 1. The Zys4 mutant grew on a medium con- taining a-aminoadipate as a substitute for lysine and containing ammonium sulfate as a principal nitrogen source; however a-aminoadipate did not serve as a principal nitrogen source for the Zys4 mutants. This result is consistent with the finding that the Zys4 mutation blocks homoaconitase, which is before a-ami- noadipate in the metabolic sequence of lysine biosynthesis (Figure 1 ) . Examples of the growth of the normal strain and various lysine auxotrophs on an a-ami- noadipate medium are shown in Figure 2.
a-Aminoadipate served as a principal nitrogen source for a wide range of lys2 mutants containing other nutritional requirements, as long as the media were supplemented for these other requirements. Such media containing a-amino- adipate as the principal nitrogen source and containing lysine (and in some instances other nutritional supplements) are referred to as AA media. The asso- ciation of Zys2 mutations and growth on AA media was demonstrated by testing segregants derived from several crosses heterozygous for Zys2 markers; in all cases, the Zys2 segregants grew on AA media, while the related LYS+ segre- gants did not. As described below, the utilization of a-aminoadipate by Zys2 mu- tants was further demonstrated by the isolation of Zys2 mutants on AA media.
The lysine requirement is recessive and the a-aminoadipate utilization is semidominant: The Zys2 mutations, like most auxotrophic mutations, are re- cessive and all of the LYS2+/Zys2 diploid strains that were tested (more than 20) did not show any significant lysine requirement. In contrast, the ability to
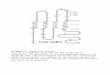
LYSINE MUTANTS OF YEAST 55 CARBON SOURCE
LYSINE
a-AMINOGLUTARATE
a-KETOGLUTARATE SACCHAROPINE L YS5
a-AMINOADIPATE- 6 -SEMIALDEHYDE
REDUCTASE
a-KETOADIPATE 4 TRANSAMINASE
HOMOSUCCINATE (GLUTARATE) a-AMINOADIPATE
(EXOGENOUS)
FIGURE 1 .-Metabolic pathways of a-aminaadipate utilization and lysine biosynthesis and the relationship to LYS genes. Some of the intermediates are not shown. (From BHATTACHAR- JEE and SINHA 1972; BROQUIST 1964; JONES and BROQUIST 1965; Kuo, SAUNDERS and BRO- QUIST 1962, 1964; MITCHELL and HOULAHAM 1948; STRASSMAN, CECI and SILVERMAN 1964; STRASSMAN and WEINHOUSE 1953).
200
100
? 2
h 50 ?I
3 h
Y
-0-0 30
20
10 ' I I I I I I
10 20 30 40 50 60
HOURS
FIGURE 2.-The growth in AA medium of a normal LYS+ strain (MO-394B) and the lysine mutants mu (MO-59-13A-(uAA2), lysZ (MO-49-14D), lys4 (19B), lys9 (MO-49-160), and lysl (SS8C-24).

56 E. E. CHATTOO et al.
use a-aminoadipate as a principal nitrogen source appeared to be a dominant trait. As summarized in Table 1, LYS2+/lys2 heterozygous strains, as well as lys2 haploid strains, grew on AA medium, although the degree of growth of the LYS2'/1ys2 diploid strains was considerably less than the growth of the lys2 haploid strains.
It appears as if growth on AA medium requires a deficiency in the lys2 or lys5 gene products, but the amount of the gene product need not be so low as to impart a lysine requirement. It is expected that the a-aminoadipate reductase activity in heterozygous diploid strains would be approximately 50% of the normal level, and it appears that this level is sufficient to allow growth on lysine- less medium and yet low enough to permit some growth on AA medium. Con- sistent with the view that only partial deficiencies are required for utilization of a-aminoadipate was the growth on AA medium of strains carrying the UAA mutation lys2-I and one or another of several tyrosine-inserting UAA suppres- sors. It was previously demonstrated with the iso-1-cytochrome c system (GIL- MORE, STEWART and SHERMAN 1971; SHERMAN, ONO and STEWART 1979) that these UAA suppressors usually restore from 5 to 15% of the normal activity when acting on typical UAA mutations. Thus, the growth of lys2-I SUP-o strains on lysineless medium and on AA medium (Table 1) suggests that ap- proximately 10% activity is sufficient to supply the lysine requirement, but is not high enough to prevent utilization of a-aminoadipate as a principal nitrogen source.
isolation of lys2 mutants on A A medium: lys2 mutants could be derived from a variety of strains on different types of AA media. An extensive study was undertaken with the normal strain AS2-2A and an AA medium containing ly- sine, tryptophan and leucine, and with the normal strain MO-320-1C and an AA medium containing lysine. Independently derived spontaneous mutants were isolated by subcloning the normal strains on nutrient medium and streak- ing cells from different colonies on separate plates of AA medium. After five to seven days of incubation at 30°, a single colony from each AA plate was sub- cloned and one subclone from each original colony was tested for growth on AA
TABLE 1
Growth of strains on synthetic media either lacking lysine or containing lysine and a-aminoadipate as a principal nitrogen source
Genotype
Synthetic media Ammonium sulfate a-Aminoadipate
(minus lysine) (plus lysine)
LYS2 +
lys2
LYSZf / l y s2
Lys2-I SUP-0
aau mutants
+ - + + +
+ + +

LYSINE MUTANTS O F YEAST 57
medium and on a normal synthetic medium lacking lysine. Nearly all of the colonies retained the ability to grow on AA medium and approximately 90% required lysine. The a-aminoadipate utilizers not requiring lysine are referred to as aau mutants.
A total of 853 spontaneous mutants requiring lysine were crossed to tester strains containing known lysine markers and the resulting diploid strains were tested for growth on medium lacking lysine. Approximately 93% of the lysine mutants failed to complement the Zys2 tester strain, although they comple- mented the other Zys tester strains. The remaining 7% failed to complement the Zys5 strain. The distribution of mutants derived from the AS2-2A and M0-320- 1C strain is presented in Table 2. It is unclear why Zys5 mutants were not un- covered in Experiment 1. The lower recovery of Zys5 mutants is consistent with the findings of BISWAS and BHATTACHARJEE (1974); out of 1587 lysine auxo- trophs that were inducted by a variety of mutagens and that were obtained by replica plating, 353 were Zys2 mutants, but only two were Zys5 mutants.
In addition to demonstrating that Zys2 mutants of spontaneous origin can be readily selected on AA medium, we have shown that this procedure can be used to select Zys2 mutants induced by UV and EMS. The frequencies of colonies appearing on AA m e d i m and the proportion of lysine auxotrophs among these colonies were determined with various doses of W and various lengths of treat- ment with EMS. In a typical experiment with strain AS2-2A and with lo' cells per plate of AA medium, the spontaneous level of approximately col- onies increased to approximately 6 X after a treatment of 25 Jm-2 of UV. Similarly, EMS treatments were shown to produce a more than four-fold in- crease of colonies appearing on AA medium, indicating that this system can be used in studies of induced mutations. Similar to the results with spontaneous mutants, approkimately 90% of the colonies appearing on AA medium after UV or EMS treatments required lysine and most of them carried mutations at the Zys2 locus. Growth of the EMS-treated cells in nutrient medium before being plated on the AA medium did not apepar to increase significantly the final yield or proportion of Zys2 mutants.
TABLE 2
Frequencies of spontaneous mutants selected on AA medium ~~
Parent strain no. AS2-2A MO-320-IC
Experiment?
lys2 mutants
lys5 mutants
aau mutants
Total
1 2
358 28
0 2
89 0
447 30
2
409
56
10
475
?Experiments 1 and 2 were performed in separate laboratories
58 B. B. CHATTOO et al.
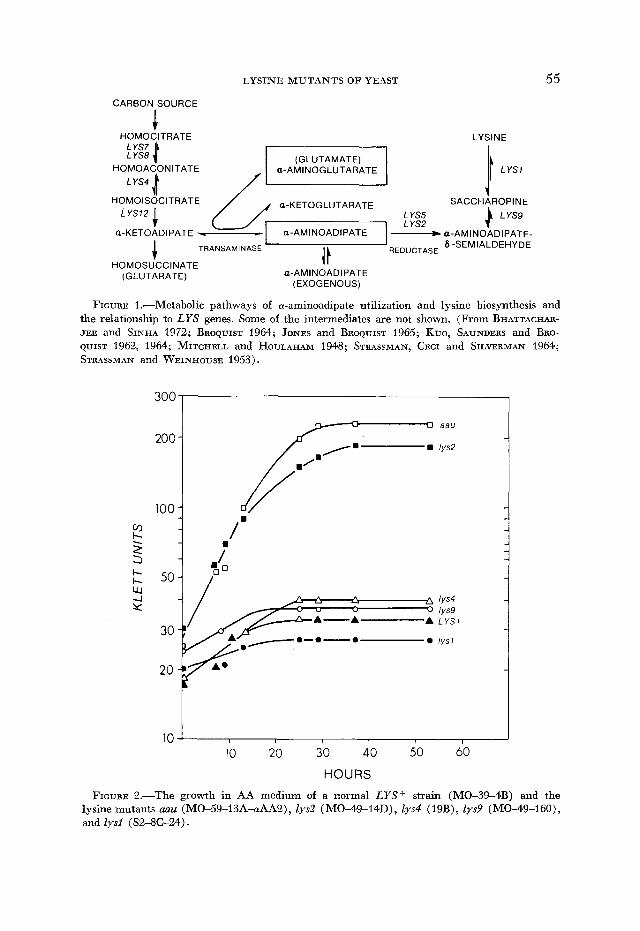
In addition, lys2 mutations were selected in strains containing mutations at other lysine loci, as well as in strains having other nutritional requirements. Strains carrying one or another of the lys4, lys6, lys7, lys8, lys9 or lysl0 muta- tions were plated on AA medium and complementation tests indicated that at least some of the resulting colonies contained lys2 blocks.
LYS2 mutants not requiring lysine: Approximately 10% of the colonies ap- pearing on AA medium did not obviously require lysine, although they still utilized a-aminoadipate as a principal nitrogen source. However examination of growth curves of one of these aau mutants, shown in Figure 3, indicated a partial lysine requirement that was not evident from the growth on solid me- dium lacking lysine. Genetic and biochemical studies described below estab- lished that these aau mutants contain lesions in the lys2 locus and that they contain partial defects in the lys2 gene product, a-aminoadipate reductase, but that the defects were insufficient to produce an obvious lysine requirement.
Twelve spontaneous aau mutants of independent origin were each crossed to either a normal LYS2+ strain or a lys2 strain. The aau/LYS2+ and aau/lys2 diploid strains were sporulated and the sexual progeny were tested for growth on AA medium and on a normal medium lacking lysine. A predominantly 2:2 segregation of the aau/LYS2+ pedigrees on the AA medium established that the aau mutations segregated as single gene mutations; a 2:2 segregation of segre- gants from the auu/lys2 pedigrees on lysineless medium, and the growth of all of these segregants on AA medium indicated that the aau mutations were at or near the LYS2 locus.
I- C-
Y 2
30 I I I I
0 10 20 30 40 50 HOURS
FIGURE 3.-Growth of the normal strain (MO-59-13A) in synthetic minimal medium (W) and in synthetic medium plus lysine (0) and growth of the uuu mutant (MO-59-13A- aAA2) in synthetic minimal medium (0) and in synthetic medium plus lysine (0) .

LYSINE MUT-4NTS O F YEAST 59
Complementation tests on AA medium with the 12 aau mutants, as well as other aau mutants, also indicated that these mutations were at the LYS2 locus. Careful inspection revealed a higher degree of growth of the aau/Zys2 diploid strains compared to the aau/LYS2+ or Zys2/LYS2 + diploid strains, indicating that the aau mutation, although semidominant, does not complement typical Zys2 mutations that cause complete loss of a-aminoadipate reductase.
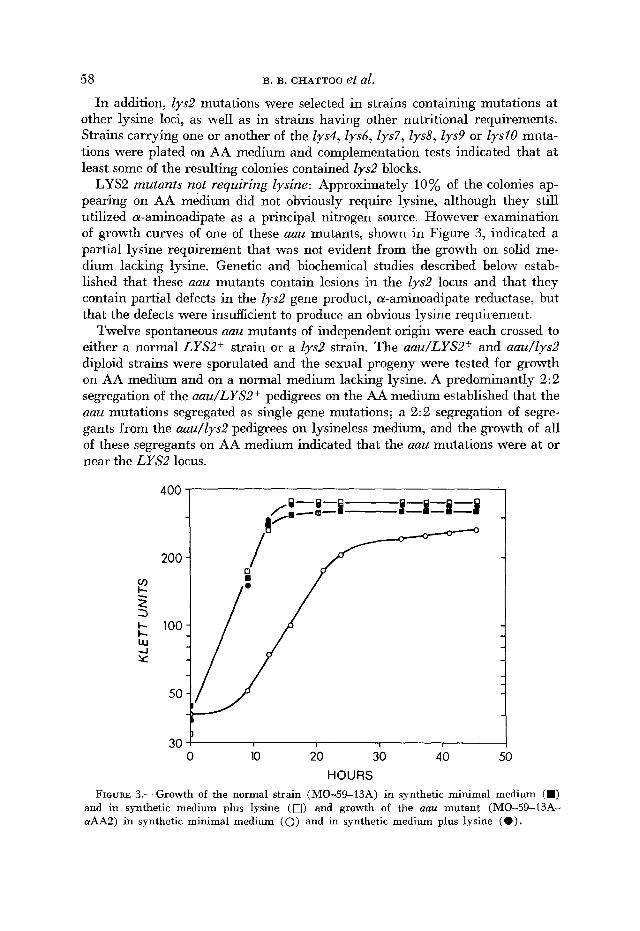
Altered a-aminoadipate reductase: Intact cells of a normal strain (MO-59- 13A), an aau mutant (MO-59-13A-aAA2) and a Zys mutant (MO-49-14D) were incubated with labeled a-aminoadipate as described in MATERIALS and METHODS. At various time intervals, cells were centrifuged and hot water ex- tracts were prepared. Chromatography and subsequent radiochromatogram scanning of both the incubation media and the cell extracts revealed a signifi- cant difference between the normal strain and both the aau and Zys2 mutants. In extracts of the normal strain, essentially all of the [14C] a-aminoadipate was converted to saccharopine and lysinte with no evidence of peaks of a-ketoadipate or glutarate. (As shown in Figure 1, glutarate is a metabolite of the homocitrate pathway formed from a-aminoadipate as a nitrogen source.) The aau mutant, on the other hand, showed diminished conversion of a-aminoadipate to saccharo- pine and lysine and showed peaks of a-ketoadipate and glutarate. Similarly, the Zys2 mutant, completely lacking a-aminoadipate reductase activity, showed no peaks for saccharopine or lysine, but did show peaks for a-ketoadipate and glu- tarate. Thus, the metabolism of a-aminoadipate in intact cells suggests that the aau mutant has reduced activity of a-aminoadipate reductase.
Direct evidence for an altered a-aminoadipate reductase in the aau mutant was provided by differences in the specific activity, the Michaelis constant (K,), and heat stability. Presented in Table 3 are the values for the specific activities and for the K, of the activity of the a-minoadipate reductase from a wild-type bakers' yeast, the normal strain MO-59-13A and the aau mutant, MO-59-1 3A-aAA2, that were estimated by Lineweaver-Burk plot of activity against a-aminoadipate concentrations. The specific activity in the aau mutant was approximately one-half that of the value found in the strain from which the mutant was derived. The K , estimated for the aau mutant was about twice
TABLE 3
Specic activities and Michaelis constants for or-aminoadipate reductase from a wild-type strain, a normal laboratory strain (MO-59-13A) and an aau mutant (MO-59-13A-aAA2)
Strain Growth on Specific K , with AA medium activity * a-aminoadipate
Bakers' yeast - 4.67 0.97 t 0.2 x 10-3 M
Normal strain (MO-59-13A) - 1.05 1.04 t 0.2 x 10-3 M
mu mutant (MO-59-13A-aAA2) + 0.43 2.17 2 0.1 x 10-3 M
'mvmoles NADPH oxidized per min per mg protein.
60 B. B. CHATTOO et al.
100
75
h
> c .- .- ;; 50 U
w C Q
B L
U,
25
a I ' I I I i
40 45 50 55
Temperature ('Cl FIGURE 4.-Comparison of the a-aminoadipate reductase activity from the normal strain
MO-59-13A (e) and the MU mutant MO-59-13A-aAA2 (0) after ten min treatments at various temperatures.
that estimated for the two normal strains, which were comparable to each other and to previous estimates ( SAGISAKA and SHIMUFM 1962; FJELLSTEDT 1969).
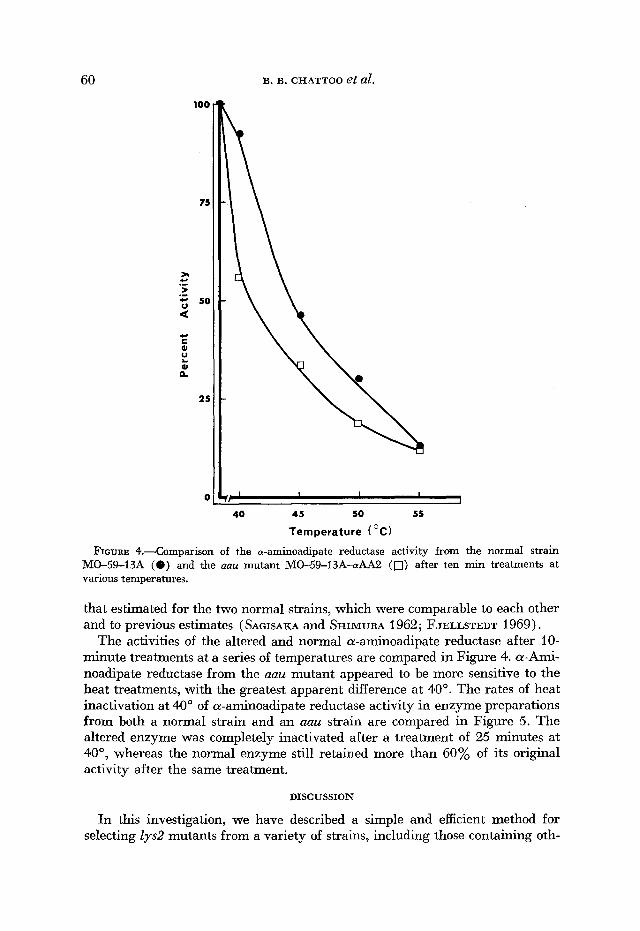
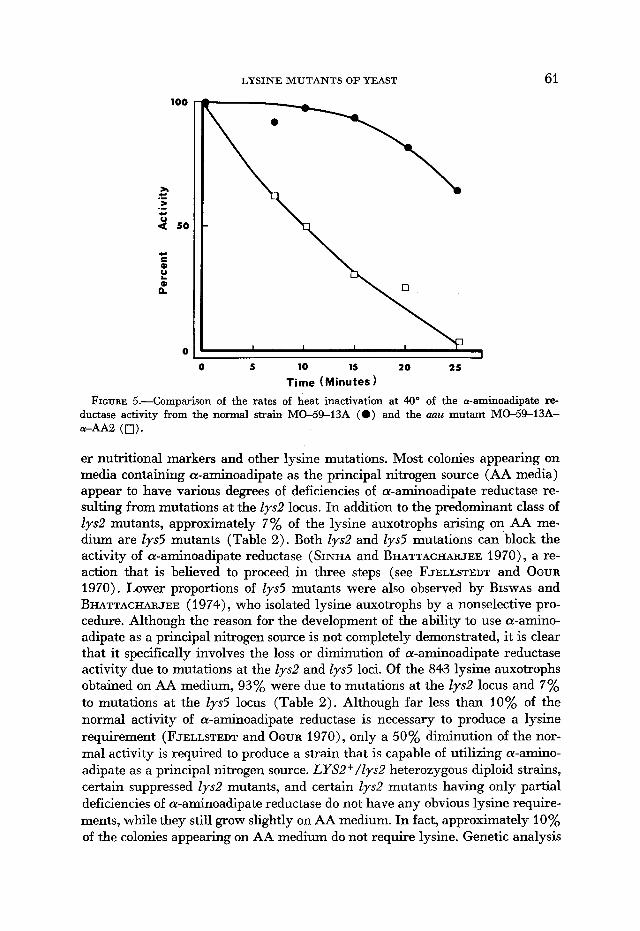
The activities of the altered and normal a-aminoadipate reductase after 10- minute treatments at a series of temperatures are compared in Figure 4. a-Ami- noadipate reductase from the aau mutant appeared to be more sensitive to the heat treatments, with the greatest apparent difference at 40'. The rates of heat inactivation at 40' of a-aminoadipate reductase activity in enzyme preparations from both a normal strain and an aau strain are compared in Figure 5. The altered enzyme was completely inactivated after a treatment of 25 minutes at 40", whereas the normal enzyme still retained more than 60% of its original activity after the same treatment.
DISCUSSION
In this investigation, we have described a simple and efficient method for selecting Zys2 mutants from a variety of strains, including those containing oth-
LYSINE MUTANTS OF YEAST 61 100
c
Y 50
0
0 5 10 15 20 25 Time (Minutes)
FIGURE 5.-Comparison of the rates of heat inactivation at 40" of the a-aminoadipate re- ductase activity from the normal strain MO-59-13A (0) and the mu mutant MO-59-13A- a-AA2 (0).
er nutritional markers and other lysine mutations. Most colonies appearing on media containing a-aminoadipate as the principal nitrogen source (AA media) appear to have various degrees of deficiencies of a-aminoadipate reductase re- sulting from mutations at the Zys2 locus. In addition to the predominant class of Zys2 mutants, approximately 7% of the lysine auxotrophs arising on AA me- dium are Zys5 mutants (Table 2). Both Zys2 and Zys5 mutations can block the activity of a-aminoadipate reductase ( SINHA and BHATTACHARJEE 1970), a re- action that is believed to proceed in three steps (see FJELLSTEDT and OGUR 1970). Lower proportions of Zys5 mutants were also observed by BISWAS and BHATTACHARJEE (1974) , who isolated lysine auxotrophs by a nonselective pro- cedure. Although the reason for the development of the ability to use a-amino- adipate as a principal nitrogen source is not completely demonstrated, it is clear that it specifically involves the loss or diminution of a-aminoadipate reductase activity due to mutations at the Zys2 and Zys5 loci. Of the 843 lysine auxotrophs obtained on AA medium, 93% were due to mutations at the Zys2 locus and 7% to mutations at the Zys5 locus (Table 2). Although far less than 10% of the normal activity of a-aminoadipate reductase is necessary to produce a lysine requirement (FJELLSTEDT and OGUR 1970), only a 50% diminution of the nor- mal activity is required to produce a strain that is capable of utilizing a-amino- adipate as a principal nitrogen source. LYS2+ /Zys2 heterozygous diploid strains, certain suppressed Zys2 mutants, and certain Zys2 mutants having only partial deficiencies of a-aminoadipate reductase do not have any obvious lysine require- ments, while they still grow slightly on AA medium. In fact, approximately 10% of the colonies appearing on AA medium do not require lysine. Genetic analysis

62 B. B. CHATTOO et al.
of representatives of these mutants established that the ability to utilize a-ami- noadipate was due to mutations at the Zys2 locus. Growth in liquid media of one of these partially blocked mutants indicated a slight stimulation of growth by lysine (Figure 3) ; biochemical studies of this mutant indicated that the a-ami- noadipate reductase had one-half the normal specific activity and a Km twice as high as the normal value (Table 3 ) , and that the conversion of a-aminoadipate to saccharopine and lysine was diminished. Thus, the range of Zys2 mutants ob- tained on AA medium extent from those having complete blocks to those with only slight deficiencies that do not produce an obvious lysine requirement.
In addition to providing a method for isolating Zys2 mutants, a medium lack- ing lysine and containing a-aminoadipate as a principal nitrogen source can be used conveniently to select for LYS2+/Zys2 diploid strains from the mating of LYS2+ and Zys2 haploid strains; the diploid strain, but neither of the haploid strains, can grow on such a medium. Thus, a Zys2 and a Zys2 tester strains can be conveniently used to test the mating types of segregants, even those lacking any requirements.
Three explanations can be presented for the association of deficiencies of a-aminoadipate reductase and the utilization of a-aminoadipate as a principal nitrogen source. As shown in Figure 1, the intracellular concentration of a-ami- noadipate is determined by its rate of uptake and by two enzymatic pathways, the reductase and the transaminase. If the a-aminoadipate reductase dominates and if the uptake rate is limiting, a-aminoadipate might be expected to be in- sufficient to serve as a nitrogen donor by the way of the weaker transaminase pathway. In fact, the specific activities of the reductase and the transaminase of the normal strain MO-59-13A were determined by saturating levels of a-amino- adipate (data not presented), and the reductase activity was found to be approxi- mately ten times the transaminase activity. In contrast to normal strains, strains having diminished activity of a-aminoadipate reductase might have more a-ami- noadipate available to serve as a nitrogen donor by way of the transaminase pathway. However, it should bet pointed out that even when it is added at high extracellular concentrations with sufficient lysine, a-aminoadipate does not serve as a nitrogen source for normal strains, nor for lysine auxothrophs blocked be- fore the transaminase or after the reductase pathway.
A second possible explanation for the lack of utilization of a-aminoadipate by normal strains is that the pathway from a-aminoadipate to lysine involves the same key intermediates, NADH and ATP, as in the pathway for a-aminoadi- pate untilization as a principal nitrogen source. It can be suggested that normal strains do not utilize a-aminoadipate as a nitrogen source because certain key intermediates are depleted as a result of lysine production from a-aminoadipate. In contrast, strains lacking a-aminoadipate reductase may have the ability to accumulate sufficiently high levels of the key intermediates to allow utilization of a-aminoadipate as a nitrogen source. However, it would be surprising to find such a specific interaction with only Zys2 and Zys5 mutants since the interme- diates in question (NADH and ATP) have general metabolic functions.

LYSINE MUTANTS O F YEAST 63
A third possible explanation for the lack of utilization of a-aminoadipate by normal strains is that the large concentrations of a-aminoadipate required for a principal nitrogen source lead to the production of an inhibitory level of an in- termediate such as a-aminoadipate-6-semialdehyde or one of its immediate pre- cursors. Complete or partial diminution of a-aminoadipate reductase would be expected to prevent accumulation of inhibitory levels of the intermediate; cer- tain partial blocks of the reductase could allow the formation of a restricted range of the intermediate that would not be inhibitory but that still would serve as a precursor for lysine biosynthesis.
Even though we do not know the exact reason why only mutants defective in a-aminoadipate reductase can use a-aminoadipate as a principal nitrogen source, this observation can be used as a basis for a convenient method to isolate Zys2 mutants.
We acknowledge J. M. WILHELM (University of Rochester Medical School) for useful dis- cussions. This investigation was supported in part by the Public Health Service Research Grants GM12.702 and AM14768, in part by Grant 09-04-08 of the Office of Research and Projects, Graduate School, Southern Illinois University, and in part by the U. S. Department of Energy at the University of Rochester, Department of Radiation Biology and Biophysics. This paper has been designated Report No. UR-3490-1609.
LITERATURE CITED
AZUBALIS, D. A., 1970 Isolation of a mutant with an altered enzyme in a biosynthetic path- way by selective pressure for the reveal of a preceding step. M.S. thesis, Southern Illinois Univ., Carbondale, Illinois.
Direct selective techniques for the isolation of pyri- midine auxotrophs in yeast. Molec. Gen. Genet. 115: 126-130.
Relationship amoung the genes, enzymes, and intermediates of the biosynthetic pathway of lysine in Saccharomyces. Molec. Gen. Genet. 115: 26-30.
Induction and complementation of lysine auxotrophs in Saccharomyces. Antonie van Leewenhoek, J. Microbiol. Serol. 40 : 221-231.
Biosynthesis of lysine from adipic acid precursors by yeast. Federation Proc. 18: 198.
c-N-(L-glutaryl-2) -L-lysine. A novel intermediate in the aminoadipic pathway of lysine biosynthesis. Proc. Sixth Int. Cong. Biochem. 5: 353-364.
Relation to lysine biosynthesis. M.S. thesis, Univ. Illinois, Urbana, Illinois.
genes controlling galactose utilization in Saccharomyces. Genetics 49 : 837-844.
or-Aminoadipic acid utilization for lysine biosynthesis in cell-free preparations of Saccharomyces. M.S. thesis, Southern Illinois Univ., Carbondale, Illinois.
Lysine metabolism in Saccharomyces. Metabolic conversion of or- aminoadipic acid to lysine. Ph.D. dissertation, Southern Illinois Univ., Carbondale, Illi- nois.
Effects of supersuppressor genes on enzymes controll-
BACH, M. L. and F. LACROUTE, 1972
BHATTACHARJEE, J. K. and A. K. SINAH, 1972
BISWAS, G. D. and J. K. BHATTACHARJEE, 1974
BROQUIST, H. P. and H. V. STIFFEY, 1959
BROQUIST, H. P., 1964
DE BOEVER, J. G., 1963 Enzymatic conversion of ketoadipic acid to aminoadipic acid in fungi:
DOUGLAS, H. C. and D. C. HAWTHORNE, 1964
FJELLSTEDT, T. A., 1966
FJELLSTEDT, T. A., 1969
Enzymatic expression and genetic linkage of
FJELLSTEDT, T. A. and M. OGUR, 1970 ing lysine biosynthesis in Saccharomyces. J. Bacteriol. 101: 108-1 17.

64 GILMORE, R. A., J. W. STEWART and F. SHERMAN, 1971
B. B. CHATTOO et al. Amino acid replacements resulting
from super-suppression of nonsense mutants of iso-1-cytochrome c from yeast. J. Mol. Biol. 61: 157-173.
Saccharopine, an intermediate of the aminoadipic acid pathway of lysine biosynthesis. 11. Studies in Saccaromyces cerevisiae. J. Biol. Chem. 240: 2531-2536. -, 1966 Saccharopine, an intermediate of the aminoadipic acid path- way of lysine biosynthesis. 111. Aminoadipic semialdehydeglutamate reductase. J. Biol. Chem. 241 : 343&3434.
Kuo, M., P. SAUNDERS and H. P. BROQUIST, 1962 A metabolite of a-aminoadipic acid in yeast: Relationship to lysine biosynthesis. Federation Proc. 21: 9. -, 19M Lysine biosynthesis in yeast: A new metabolite of a-aminoadipic acid. J. Biol. Chem. 239:
Utilization of nitrogen compounds by yeast of the
Protein measure-
JONES, E. E. and H. P. B R ~ U I S T , 1965
508-515. LARUE, T. A. and J. R. T. SPENCER, 1968
LOWRY, 0. H., N. S. ROSEFJROUGH, A. L. FARR and R. J. RANDALL, 1951
MATSUDA, M. and M. OGUR, 1969a
genus Saccharomyces. Antonie van Leeuyenhoek J. Microbiol. Serol. 34: 153-158.
ment with the folin reagent. J. Biol Chem. 193: 265-275.
Separation and specificity of the yeast glutamate: a- ketoadipate transaminase. J. Biol. Chem. 244: 3352-3358. -, 1969b Enzymatic and physiological properties of the yeast glutamate: a-ketoadipate transaminase. J. Biol. Chem.
MITCHELL, H. K. and M. B. HOULAHAM, 1948 An intermediate in the biosynthesis of lysine in Neurospora. J. Biol. Chem. 174: 883-887.
MORRIS, E. 0. and A. A. EDDY, 1957 Method for the measurement of wild yeast infection in pitching yeast. J. Inst. Brewing 63: 34-35.
MORTIMER, R. K. and T. R. MANNEY, 1971 Mutation induction in yeast. pp. 289-310. In: Chemical Mutagens: Principles and Methods for their Detection. Vol. 1. Edited by A. HOLLAENDER. Plenum Press, New York.
A glutamic: a-ketoadipic trans- aminase in Saccharomyces. Biochem. Biophys. Res. Commun. 32 : 380-384.
A system selective for mutations affecting the synthesis of adenine in yeast. Compt. Rend. Lab. Carlsberg, SQr. Physiol. 26: 299-314.
Studies in lysine biosynthesis I11 enzymatic reduction of a-aminoadipic acid: Isolation and some properties of the enzyme. J. Biochem. (Tokyo) 51: 3 9 8 4 4 -
Amino acids as nitrogen source for growth of yeasts. Arch. Biochem. Biophys. 19: 184-192.
Saccharomyces. pp. 359-393. In: Handbook of Genetics, Vol. I. Edited by R. C. KING. Plenum Press, New York.
of yeast defective in iso-1-cytochrome c. Genetics 77: 255-284.
Use of the iso-1-cytochrome c system for investigating nonsense mutants and suppressors in yeast. pp. 135-153. In: Nonsense Mu- tations and tRNA Suppressors. Edited by J. E. CELIS and J. D. SMITH, Academic Press, London.
Characteristics and relationships of mercury-resistant mu- tants and methionine auxotrophs of yeast. J. Bacteriol. 118: 911-918. -, 1975 Genetic and physiological characterization of met15 mutants of Saccharomyces cerevisiae: A se- lective system for forward and reverse mutations. Genetics 81: 75-97.
244: 5153-5158.
PIEDISCALZI, M. J., T. A. FJELLSTEDT and M. OGUR, 1948
ROMAN, H., 1956
SAGISAKA, S. and K. SHIMURA, 1962
SCHULTZ, A. S. and S. POMPER, 1948
SHERMAN, F. and C. W. LAWRENCE, 1974
SHERMAN, F., J. W. STEWART, M. JACKSON, R. A. GILMORE and J. H. PARKER, 1974
SHERMAN, F., B. ONO, and J. W. STEWART, 1979
Mutants
SINGH, A. and F. SHERMAN, 1974

LYSINE MUTANTS OF YEAST 65
SINHA, A. K. and J. K. BHATTACHARJEE, 1970 Control of a lysine-biosynthetic step by two
STRASSMAN, M., L. N. CECI and B. E. SILVERMAN, 19M Enzymatic conversion of homoiso-
STRASSMAN, M. and S. WEINHOUSE, 1953 Biosynthetic pathways 111. The biosynthesis of
THORNE, R. S. W., 1950 Mechanisms of nitrogen assimilation by yeast and their relation
WADDELL, W., 1956 A simple ultraviolet spectrophotometric method for the determination of
WALTERS, L. S. and M. R. THISELTON, 1953
WHELAN, W. L., E. GOCKE and T. R. MANNEY, 1979 The CAN1 locus of Saccharomyces
WIELAND, T. and E. FISCHER, 1949 Paper chromatography of a-keto acids. Naturwissens-
Corresponding editor: C. SLAYMAN
unlinked genes of Saccharomyces. Biochys. Res. Commun. 39 : 1205-1210.
citric acid into a-ketoadipic acid. Biochem. Biophys. Res. Commun. 14: 268-271.
lysine by Torulopsis utilis. J. Am. Chem. Soc. 75: 1680-1684.
to the problem of yeast growth in Wort Wallerstein Lab. Commun. 13: 319-340.
protein. J. Lab. Clin. Med. 48: 311-314.
59: 401404%
cereuisiae: Fine-structure analysis and forward mutation rates. Genetics 91 : 35-51.
chaften 36: 219.
Utilization of lysine by yeast. J. Inst. Brewing