Embed Size (px)
Citation preview

J . Zool., Lond. (1987) 213, 545-568
Scale epidermis of Nutvix piscutor during its sloughing cycle- structural organization and protein
histochemistry (Reptilia: Colubridae)
A J A Y K U M A R MITTAL A N D J A I P R A K A S H NARAIN S l N G H '
Skin Physiology Laboratory, Department ofzoology, Banaras Hindu Uniuersity, Varanasi 22I00.5, India
(Accepted 14 October 1986)
(With 2 plates and 1 figure in the text)
The structure and histochemistry of proteinaceous constituents in cellular components of various layers of the epidermis of the chequered water snake, Nutrix piscurur, throughout the sloughing cycle have been described.
Histologically. six consecutive stages are recognized during one complete sloughing cycle. In Stage I , immediately following sloughing, the epidermis consists of an inner stratum germinati- vum, one or two layers of undifferentiated epidermal cells and the new outer epidermal generation represented by the Oberhautchen. /]-layer, mesos layer and an r-layer which continues to differentiate and mature throughout Stage I and is observed even in Stage 2. The presence of several layers of undifferentiated epidermal cells over the stratum germinativum, a characteristic feature of Stage 2, results from the temporary slowing of their differentiation into the cells of various epidermal layers. Stage 3 includes differentiated lacunar tissue and the clear layer of the outer epidermal generation, and the presumptive Oberhautchen and 8-layer of the inner epidermal generation. In addition to marked cytological and histochemical changes in the lacunar tissue and the maturation of various keratinized layers, Stages 4 and 5 include the presumptive mesos and a-layers, respectively. Stage 6 is characterized by total disintegration of the lacunar tissue and maturation of various keratinized layers, except the presumptive a-layer, of the inner epidermal generation, which is still incomplete. The degeneration of the lacunar tissue is discussed in relation to the cessation of the supply of nutrients and other body fluids from underlying epidermal cells.
Histochemically. the Oberhautchen resembles the mesos and r-layers in being strongly positive for -SH and -NH2 groups. bound calcium and basic proteins. However, i t shows more similarity with the a-layer in its moderate reaction for tyrosine and tryptophan, which is more intense in the mesos layer; and shows more similarity with the mesos layer with its very strong reactions for -SS groups. unlike the r-layer which remains unstained.
The functional value of tyrosine- and tryptophan-rich proteins in the keratinized layers of the epidermis is not immediately apparent.
In addition to the high content of -SS groups. providing durability and mechanical strength, and protein-bound calcium, adding hardness, the vertical striations in the Oberhautchen of the scales probably reinforce the epidermal superficial layer to guard against wear and tear by abrasion.
Contents Page
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 546 Materials and methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . 547
'Supported by University Grants Commission, Govt of India. as a teacher fellow; present address: Department of Zoology, R.H.S. Degree College, Singramau, Jaunpur, India
0022-5460/87/01 I545 + 23 $03'00 545
hz 1987 The Zoological Society of London

546 A. K . MITTAL AND J . P. N . SINGH
Observations . . . . . . . . . . . . . . Stage 1 . . . . . . . . . . . . . . . . Stage 2 . . . . . . . . . . . . . . . . Stage 3 . . . . . . . . . . . . . . . . Stage 4 . . . . . . . . . . . . . . . . Stage 5 . . . . . . . . . . . . . . . .
Discussion . . . . . . . . . . . . . . . . References . . . . . . . . . . . . . . . . Abbreviations used in the figure and the plates
Stage 6 . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . 552
. . . . . . . . . . . . . . . . . . 552
. . . . . . . . . . . . . . . . . . 555
. . . . . . . . . . . . . . . . . . 551
. . . . . . . . . . . . . . . . . . 558
. . . . . . . . . . . . . . . . . . 559
. . . . . . . . . . . . . . . . . . 559
. . . . . . . . . . . . . . . . . . 560
. . . . . . . . . . . . . . . . . . 566
. . . . . . . . . . . . . . . . . . 568
Introduction
Maderson (1 965) first systematically studied the epidermis of snakes during the sloughing cycle; his descriptions are based mainly on Elaphe taeniura. Six arbitrary stages were described, each characterized by the specific histological appearance of the epidermis during one complete cycle. These stages were described as ‘the resting phase’ (Stage I ) , ‘proliferation-renewal phase’ (Stages 2-5) and ‘pre-shedding phase’ (Stage 6) (Banerjee & Mittal, 1980). The period of time from one slough to the next is termed ‘the inter-slough period’. In addition to these six stages, Maderson, Chiu & Phillips (1970) described one more stage-Stage 0-in Ptyas korros. Surveying vertebrate epithelia in relation to keratinization, Spearman (1966, 1973) and Parakkal & Alexander (1972) described cyclic changes in the epidermis of squamates. Landmann (1975) gave a brief account of the skin structure of snakes in general, and Roth & Jones (1970) and Landmann (1979) reported details of the ultrastructure of the epidermis of Constrictor constrictor and Natrix natrix, respectively, in different stages of the sloughing cycle. Other information on the epidermis of snakes is concerned with the sub-microscopic structure of the stratum corneum of Constrictor constrictor amarali, Eunectes murinus, Epicrates cenchria crassus and Boa hortulana (Hoge & Santos, 1953); the nature of the Oberhautchen surface in Natrix natrix (Horstmann, 1964); the types of fibrous protein in freshly-shed skin of Constrictor constrictor (Baden, Roth & Bonar, 1966); the ultrastructure of the epidermis of Constrictor constrictor during the resting phase (Roth & Jones, 1967); the fine structure of the basal layer (Miscalencu & Ionescu, 1972), intermediate layer (Miscalencu, 1974) and horny cells (Ionescu & Miscalencu, 1972) in the epidermis of Natrix natrix; the ultrastructure of the epidermis and dermal chromatophores in Vipera ammodytes (Miscalencu & Ionescu, 1973) and sites of the permeability barrier in the epidermis of Natrix natrix during the resting stage (Landmann, Stolinski & Martin, 1981).
Knowledge ofcellular localization of protein constituents in the skin of snakes at different stages of the sloughing cycle is very limited. Roth & Baden (1967) described the site of protein synthesis in the epidermis of the indigo snake, Drymarchon corais couperi using 3H leucine, and Downing & Roth (1974) showed the derivation of cells of the epidermal strata of Constrictor constrictor using 3H thymidine. Banerjee & Mittal (1978a, b) reported proteins and carbohydrates in various cellular components of the epidermis of Natrixpiscator in a stage corresponding to the late third- early fourth stage of Maderson’s (1965) description. Goslar (1958, 1964) extensively studied the structural organization of the skin of Natrix natrix and its histochemistry with regard to carbohydrates, lipids, nucleic acids, distribution of cysteine-bound sulphydryl groups and disulphide bonds of cystine and localization of several enzymes. These findings, however, need further clarification since Goslar (1 958, 1964) assumed continuous epidermal keratinization and

SCALE EPIDERMIS O F N A T R I X P I S C A T O R 547
STAGE 1 STAGE2 , STAGE 3 STAGE 4 STAGE 5 STAGE 6
I I I I
FIG. 1. Diagrammatic representation of the epidermis of Nuirixpiscator at the outer scale surface in different stages of the sloughing cycle. For explanation of abbreviations, see p. 567.
his descriptions of the sloughing cycle are incomplete and quite different from those described by Maderson (1969, Maderson, Chiu et ul. (1970) and Landmann (1979).
The skin of Nutrix piscator, the chequered keel-back water snake, includes a series of small discrete scales, each having an outer and inner surface. The scales join each other by folds of the inter-scale hinge region. The histological appearance of the epidermis at the inter-scale hinge region resembles the inner surface of the scale, but differs from that of the outer scale surface. The present study is concerned with the structural organization and histochemical characterization of proteinaceous constituents in different cellular components of the epidermis at the outer surface of scales of Nutrix piscator in different stages of sloughing. The results obtained are interpreted in terms of their possible functional significance.
Materials and methods
Natrixpiscator ( = Tropidonotuspiscator, Gharpurey, 1962; Xenochrophispiscator, Whitaker, 1978) is non- poisonous and commonly found in and near freshwater ponds, tanks, lakes, flooded paddy fields and rivers in India. It belongs to the subfamily Colubrinae, family Colubridae, sub-order Serpentes and order Squamata (Smith, 1961).
Live specimens of Nafrix piscafor (approx. 75-90 cm in length) were collected from the vicinity of local ponds at Varanasi and maintained in the laboratory. They were kept in separate cages with an adequate supply of fresh water and fed once a week with live frogs and small fishes.
Specimens at different stages of the sloughing cycle were anaesthetized with chloroform/ether vapour. Skin pieces (approx. 10 x I5 mm) excised from the back and belly were fixed in 10% neutral formalin (Lillie, 1954), Helly’s fluid (Helly, I903), aqueous Bouin’s fluid (Bouin, 1897) and 70% ethyl alcohol. Paraffin-embedded material was sectioned at 5 pm.
Ehrlich’s haematoxylin/eosin (H/E), Verhoeffs haematoxylin eosin (VHE) (Lillie, 1954), Mallory’s triple stain (MTS) (Mallory, 1936) and Papanicolaou’s stain (PS) (Gurr, 1958) were used as general histological stains and the Feulgen reaction (Feulgen & Rossenbeck, 1924) for DNA.
Skin sections were stained with Ninhydrin/Schiff technique to demonstrate protein-bound NH2 groups (Yasuma & Itchikawa, 1953); solochrome cyanin R technique for basic proteins (Pearse, 1968); Millon reaction for tyrosine (Baker, 1956); dimethylaminobenzaldehyde-nitrite (DMAB-nitrite) for tryptophan

548 A. K. MITTAL AND J. P. N . S l N G H
TABLE I A summary of protein histochemistry o/scole epidermis o/ Narrix
Outer epidermal generation __ ~
Chemical Histochemical Oberha- 8- Mesos constituents techniques utchen layer layer
Lacunar Clear a-layer tissue layer
Stages Stages 1-6 1 4
Stages Stages 1-2 Stages Stages Stage Stages I4 3 4 3 4 3
Outer compact
part
Inner swollen
cells
Protein-
bound -NH2 groups
Cysteine- bound sulphydryl (-SH) groups
Ninhydrin/Schiff +Pa
Deaminationl -
Ninhydrin/Schiff
dinaphthyl- disulphide (DDD)
Dihydroxy- **a
+ + P -
+ +PUR
-
+ + B L
-
+ + + B L
-
+ + + R -
+ + P
-
-
+ + S
+ +P -
f BR
-
-
- + + P U R - -
Iodine oxidation or ethyl- maleimide oxidation/DDD
Performic acid alcian blue
-
+ + +BLa
-
+ BLa
-
+ RB -
++P'
-
-
+ +S'
Cystine- bound disulphide (4s) groups
Thioglycollate reduction/ Performic acid alcian blue
Dimethyl amino benzaldehyde (DMAB)-nitrite
Iodinat ion/ DMAB-nitrite
Millon reaction
lodination/ Millon reaction
Solochrome cyanin R
Sakaguchi reaction Permanganate haematoxylin Alizarin red S
Tryptophan + BL - + B L - -
Tyrosine +R -
Basic protein
Arginine
Keratohyaline
Bound calcium
+ + P - + + P - -
-
+ + S
Symbols: 'vertical striations; areas between the striations stain In much weaker shade; narrow apical zone; strong at narrow apical zone and vertical striations; cell boundaries; *, ' f P in Stage 4 and ' + P in Stages 5 and 6; **, ' + + PUR' in Stages I and 2, * + PUR' in Stages 3 and4and'+PUR'inStagesSand6;***,'-'inStage4and'+PURinStages5and6; # , '+PURinS tage3and '+ +PUR'inStage4; #.

SCALE EPIDERMIS OF N A T R I X PISCATOR 549
pisearor in differmi siuges of ihe sloughing cycle
Inner epidermal generation
Presumptive Presumptive Oberhautchen ,%layer
~~~~~
Stratum germinativum
and undifferen-
Presumptive tiated epi- z-layer dermal cells
Presumptive mesos layer
Stage Stage Stages Stages Stages Stage Stage Stage Stage Sedge 6 Stages 3 4 5-6 3-4 5 4 4 5 6 5 1 - 6
Outer Inner compact swollen
part cells
+PURb +PURC + + f P U R a # +PUR *PUR +PUR + + P U R - 5 B R - -
* B L + B L d + BL + + + B L - + B L - + B L -
+ + + R -
+ P + + p a + P + P + P - + + P - + P -
- - -
+sc + +sa + S
' + + S' in Stage 5 and ? S' in Sedge 6; BL, blue; BR, brownish red; P, pink; PUR. purple: R. red; S , scarlet; - , negative; * , weak; +. moderate; + +, strong; + + + , very strong reactions.

550 A . K . MITTAL A N D J . P. N . SINGH
TABLE I1 A summary offluorescence hisrochemislry for keratin in scale epidermis of
Outer epidermal generation -
Oberhautchen, p-layer and Mesos layer a-layer
- Hislochemical Stages
techniques I 6
Congo red- Titan yellowl Thioflavin T
DNase/RNase/ + + +GBL Congo red- Titan yellowl Thioflavin T
Rhodamine B + + +SR
+ + + GBL
DNase/RNase/ + + +SR Rhodamine B
Thioflavin T + + + YG
DNase/RNase/ + + + Y G Thioflavin T Titan yellow + + +YG
DNase/RNase/ + + +YG Titan yellow
Stages 1-2
Lacunar Clear Presumptive tissue layer Oberhautchen
Stages Stages Stage 3 4 3 6 3
Outer Inner compact swollen
part cells
+ G B L ~ + +GBL + +GBLa. + + +GBL + +GBL',
~ G B L ~
++GBL -
+ + S R ++SRa, *SRb
+ + S R -
+ + Y G ++YGa, *YGb
+ + Y G -
+ + Y G ++YGB, +YGb
+ + Y G -
+ + + G B L -
+ + + S R ++SRB, +SRb
+ + + S R -
+ + + Y G ++YGB *YGb
+ + + Y G -
+ + + Y G + + Y @ *YGb
+ + + Y G -
+GBLa, fGBLC
*GBLC
+ SR', + SRC
+ SRC
+YG', *YGC
*YGC
+YG', *YGC
*YGC
Stages Stage Stage Stages &6 3 4 5 4
+GBL + G B L ~
~ G B L + G B L ~
*SR +SRd
+SR +SRd
*YG +YGd
+YG +YGd
+YG +YGd
*YG +YGd
+ + G B L + + + G B L
+ t G B L + + + G B L
++SR + t + S R
+ + S R + + + S R
+ + Y G + + + Y G
+ + Y G + + + Y G
+ + Y G + + + Y G
+ + Y G + + + Y G
Symbols: ', nucleus; ', cytoplasm; ', cell boundary; ', narrow apical zone; GBL, greenish blue; SR, salmon red; YG, yellowish green; -,

SCALE EPIDERMIS OF NATRIX PISCATOR 55 I
Natrix piseator in different siuges of /he sloughing c.ycle
Inner epidermal generation -. .~ .. .
Stratum germinativum
and undifferen-
Presumptive Presumptive Presumptive tiated epi- 8-layer mesos layer u-layer dermal cells
Stage Stage Stages Stage Stage Stage Stage 3 4 5-6 ' 4 5 6 5
~ ~~~~~
+GEL + + G E L + + + G B L +GEL", + + G E L + + + G E L ++GEL", ~ G B L ~ + G B L ~ ,
+GELC
+GEL + + G E L + + + G B L +GEL' + + G E L + + + G E L +GELc
+ SR
+SR
+ YG
+ YG
+YG
+YG
+ + S R
+ +SR
+ +YG
+ +YG
+ +YG
+ +YG
+ + + S R
+ + +SR
+ + +YG
+ + + Y G
+ + + Y G
+ + + Y G
+SR", + SRC
f SR'
+YG'. t Y G C
f YGC
+YGa. + YG=
fYGC
+ + S R
+ +SR
+ +YG
+ +YG
+ +YG
+ +YG
+ + +SR
+ + +SR
+ + + Y G
+ + + Y G
+ + + Y G
+ + + Y G
+ +SRa, + SRb. + SRC + SR'
+ +YGa. +YGb. + YGC + YGC
+ + YGa, +YGb, +YGC + YGC
Stage Stages 6 1-5
Outer Inner compact swollen
part cells
+ + G E L + +GELa. + +GELa, ~ G B L ~ + G B L ~
+ + G E L - -
+ + S R
+ +SR
+ +YG
+ + Y G
+ +YG
+ +YG
+ + SR", + + S R b
-
+ +YGa, k Y G b
-
+ + YG", +YGb
-
+ + SR", + SRb
-
+ +YGa, +YGb
-
+ +YGa, +YGb
-
negative; f , weak; +, moderate; + + , strong; + + +, very strong reactions

552 A . K. MITTAL A N D J . P . N . SlNGH
(Adams, 1957); dihydroxy-dinaphthyl-disulphide (DDD) for cysteine-bound sulphydryl groups (Barrnett & Seligman, 1952); performic acid alcian blue for cystine-bound disulphide groups (Adams & Sloper, 1955); Sakaguchi reaction for arginine (Baker, 1947); permanganate haematoxylin for keratohyaline (Gurr, 1958) and alizarin red S technique for bound calcium (Lillie, 1954). Diazo-deazo procedure for blocking-NH? groups, iodination for tyrosine and tryptophan, iodine oxidation and ethyl maleimide oxidation for -SH groups, and thioglycollate reduction for -SS groups were used as controls (see Pearse, 1968).
For keratin fluorescence studies, paraffin sections of skin fixed in 70?4 ethyl alcohol were stained with rhodamine B, titan yellow, thioflavin T and congo red-titan yellow/thioflavin T, with and without prior DNase and RNase treatments (Jarrett, Spearman & Hardy, 1959) and were examined with a Fluoval Carl Zeiss (Jena) microscope using BG 12/4G excitation and OGI barrier filters.
Snakes were not re-used after biopsy because operated snakes invariably shed surface layers of epidermis as flakes, unlike intact snakes which slough the epidermis as a complete sheet. Operated snakcs stopped feeding and ultimately died in a few days.
Observations
The epidermis of Natrix piscator undergoes cyclic changes in its organization associated with periodic sloughing. Based on histological organization, six arbitrary stages are identified during each inter-slough period. (Fig. 1; Tables I and 11).
Stage I
The epidermis at this stage, immediately following sloughing, consists of (a) an incompletely formed outer epidermal generation, (b) a layer of undifferentiated epidermal cells and (c) the stratum germinativum.
The outer epidermal generation in this stage is represented by the Oberhautchen, /%layer, mesos layer and an a-layer which is still undergoing differentiation and maturation.
The most superficial Oberhautchen, of dead cornified cells devoid of nuclei and without distinct cell boundaries, is intimately associated with the underlying p-layer. It is characterized by vertical striations traversing its whole thickness (Plate Ia). The vertical striations and inner margin of the Oberhautchen are moderately eosinophilic in H/E and PS; stain purplish blue in VHE and red in MTS; stain very strongly for cystine-bound disulphide (4s) groups (Plate Ib); stain strongly for cysteine-bound sulphydryl (-SH) groups (Plate Ic), basic proteins and bound calcium; stain moderately for protein-bound amino (-NH*) groups, tyrosine and tryptophan. The areas between the vertical striations, in general, stain in relatively much weaker shades. In most histological preparations the thin free margins of this layer are brownish and refractile in nature.
The P-layer is relatively thick, composed of compactly arranged dead cornified cells, invariably impregnated with granular melanin inclusions, and without apparent cell boundaries and nuclei (Plate la). It is weakly eosinophilic, stains light orange in MTS, grey in VHE and displays very strong reactions for tyrosine and tryptophan; there are weak reactions for -SS groups, -SH groups, basic proteins and bound calcium.
The mesos layer is thin, with no differentiation of cellular boundaries and nuclei and is firmly attached to the inner boundary of the 8-layer. The mesos layer is frequently represented as a strand between the /I-layer and underlying cc-layer; probably this is a histological artefact. It is strongly eosinophilic in H/E and PS, stains purplish blue in VHE and red in MTS and gives strong reactions for -NH2 and -SS groups (Plate Ib), -SH groups (Plate Ic), bound calcium and basic proteins. With

S C A L E E P I D E R M I S OF NATRIX PISCATOR 553
histochemical preparations for tyrosine and tryptophan, the mesos layer is indistinguishable from the a-layer for both layers react strongly for thesc protein end groups.
The Oberhautchen, /)- and mesos layers show very strong DNase- and RNase-resistant fluorescence in Congo red-titan yellow/thioflavin T, rhodamine B, thioflavin T and titan yellow preparations, and they are not delineated from each other.
At Stage 1 the a-layer is represented by an outer compact part of flattened cells, often having distinct cell boundaries and flat pycnotic nuclei (Plate Ia). Its staining reaction is similar to that of the mesos layer in H/E, PS, VHE and MTS and it gives strong reactions for basic proteins, -NH2 groups, bound calcium and with various fluorochromes for keratin, and moderate reactions for tyrosine and tryptophan. I t stains brownish red with DDD reaction, indicating the presence of a lower concentration of-SH groups (Plate Ic) and remains unstained for -SS groups. Immediately underlying the compact part are swollen, roughly rectangular cells arranged in one or two layers (Plate la), destined to be incorporated into the compact part on completion of their maturation. Each cell has a dilated spherical or oval nucleus with distinct nucleolus and chromatin material moderately stained for DNA. The cytoplasm is finely granular, weakly eosinophilic in H/E and PS, stains purple with VHE and remains unstained with MTS. I t does not give positive reactions for various protein end groups except a weak reaction for protein-bound -NH2 groups and bound
~ __ ~- -~ ~. ~. ~~ ~~ ~~
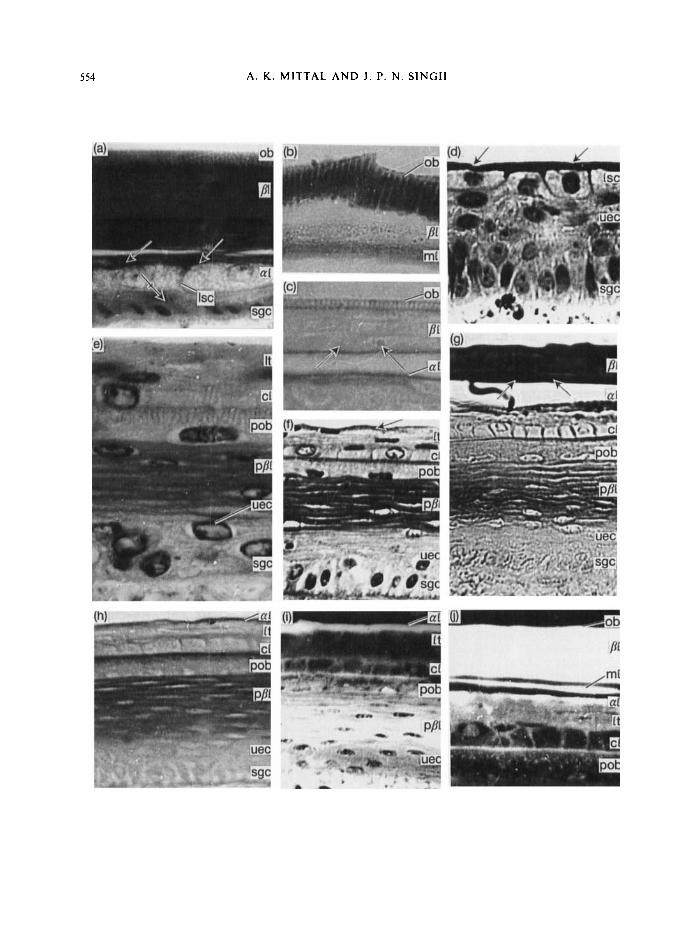
PLATE I . (at(j) Photomicrographs of cross-sections of the skin of Nurrixpiscufor showing the epidermis a t the outer surface of scales in different stages of the sloughing cycle. For explanation of abbreviations, see p. 568.
(a) Showing the ob having vertical striations (purplish blue in the original) and a thick []I (grey in original). The ml is indistinguishable from the /I1 owing to similar staining behaviour. The rl is represented by a layer of flattened cells (arrows) and a layer of large swollen cells each having a dilated spherical nucleus and granular cytoplasm. The gap between the al and ml is an histological artefact. Note an uec (barred arrow) between the Isc of the a1 and the sgc. (Belly skin, Stage 1. VHE. x 2800.)
(b) A part o f the epidermis showing a very strong reaction for disulphide bonds of cystine in vertical striations of oh (deep blue in original). strong reaction in the ml (blue in original) and weak reaction in the /I1 (light blue in original). (Belly skin, Stage 1. Performic acid alcian blue. x 3500.)
(c) A part of the epidermis showing a strong reaction for -SH groups in vertical striations and at the basal margin of the oh and ml (arrows) (purple in original). The compact part of 21 stains brownish red in the original. indicating a lower concentration of-SH groups. (Back skin. Stage I . DDD reaction. x 3100.)
(d) A part of the epidermis showing the rl represented by an outer compact part (arrows) (deep pink in original) and a layer of large swollen cells. Note the uec are arranged i n 4 -5 layers and the sgc have acquired a columnar shape. (Belly skin, Stage 2. H / E . x 1600.)
(e) A part of the epidermis showing the I t and cl of the outer epidermal generation and various layers of the inner epidermal generation. Note the presence of vertical striations (deep pink in original) in the apical region of the pob. (Belly skin. Stage 3. H/E. x 3500.)
(f) A part of the epidermis showing a thin compact layer at the outer boundary of the It representing the mature al (arrows). Note that the p/lI flattened cells. and the boundary between the cl and the pob. are darkly stained (deep pink in original. Back skin, Stage 3. HIE. x 1600.)
(g) Strong reactions for tyrosine in the P I and the ml (arrows) (dcep red in original). Note moderate reactions (light red in original) in the X I . cI cell boundaries. the p/jI and the pob. (Belly skin. Stage 3 . Millon reaction. x 2250.)
(h) A part of the epidermis showing positive reaction (purple in original) for -SH groups in the al. narrow apical zone of the pob and the ppl. (Belly skin. Stage 3. DDD reaction. x 1800.)
( i ) A part o f the epidermis showing Huorescence (salmon red in original) in the rl, the cl cell boundaries and the p/II. (Back skin, Stage 3. Rhodaniine B. x 1300.)
(j) A part of the epidermis showing very strong Huorescence (salmon red in original) in the ob. /A and ml, indistinguishable from each other. Fluorescence in the rl is less strong. Note weak fluorescence in the I t and cl cell boundaries and moderate reaction in the boundary between the cl and pob. (Belly skin. Stage 3. Rhodaininc B. x 1460.)
554 A. K . MITTAL AND J . P. N . SINGH

SCALE EPlDERMIS OF NATRIX PISCATOR
calcium. With various fluorochromes the cytoplasm of these cells shows faint and the nuclei very strong fluorescence that is blocked by pretreatment of sections with RNase and DNase, respectively. The outer margins and apical half of lateral margins of these cells stain similarly to the outer compact part of the a-layer.
The undifferentiated layer of the epidermis consists of vertically compressed, spindle-shaped cells arranged in one or two layers, each having a central, apparently viable spherical or oval nucleus (Plate Ia) with distinct chromatin and nucleolus, stained moderately for DNA.
The stratum germinativum of closely approximated, almost cuboidal cells, each having a central spherical nucleus (Plate Ia) with distinct chromatin material and nucleolus, stained strongly for DNA. The cells lie on a thin non-cellular basement membrane, which is eosinophilic and stains dark blue in MTS. It is thrown into minute papillae-like projections interdigitating with corresponding infoldings at the inner surface of stratum germinativum cells. Pigment cells occur between the stratum germinativum cells, in the dorsal body epidermis (Mittal & Singh, In prep.).
The cytoplasmic content of undifferentiated epidermal cells and cells of the stratum germinativum in this and in subsequent stages of the sloughing cycle are slightly eosinophilic in H/ E, PS and VHE; they stain purplish in MTS and show staining similar to that of the large swollen cells of the a-layer for various protein end groups.
5 5 5
Stage 2
In contrast to Stage 1, the epidermis of Stage 2 has several layers of undifferentiated epidermal cells between the a-layer and the stratum germinativum (Plate Id), whose cells have now acquired an elongated columnar shape. The a-layer is relatively thick, compact and mature, devoid of nuclei and cell boundaries, and there is also a layer of large, swollen, almost rectangular cells which represent its immature region (Plate Id).
PLATE 11. (aHh) Photomicrographs of cross-sections of the skin of Nutrixpiscator, showing the epidermis at the outer surface of scales in different stages of the sloughing cycle. For explanation of abbreviations, see p. 568.
(a) A part of the epidermis showing vacuoles (barred arrow) in the It, flat pycnotic nuclei in pob and PSI, vertical striations in the full thickness of pob and very elongated cells with healthy nuclei immediately underlying the ppl representing pml. A layer or uec (arrow) is seen between the pml and sgc. Compare with Plate Ie. (Belly skin, Stage 4. H/ E. x 3200.)
(b) A part of the epidermis showing positive reactions (purple in original) for -SH groups in the al (arrows), the pob and the ppl. (Belly skin, Stage 4. DDD reaction. x 1760.)
(c) Epidermis showing a mature cl (arrows) (deep pink in original) at the inner boundary of the It having vacuolated cells, mature pob (light pink in original), mature pfll (pink in original) and a pml, the outer part of which is mature and compact (deep pink in original). (Belly skin, Stage 5. H/E. x 1280.)
(d) A part of the epidermis showing the ml, the outer part of which is mature and compact (arrows) (deep purple in original) and the pal represented by large swollen cells having central dilated nuclei. (Belly skin, Stage 5. VHE. x 2500.)
(e) Strong reactions (scarlet in original) for bound calcium in various keratinized layers except the 81, staining weakly (pink in original), and thecl and pml, stainingmoderately(1ight red in original). (Back skin, Stage 5 . Alizarin red S. x 1400.)
(f) Strong fluorescence in various keratinized layers and at the boundaries of the pml cells. (Back skin, Stage 5 . Rhodamine B. x 310.)
(9) A part ofthe epidermis showing strong reaction (deep blue in original) for -SS groups in the pob and pml and weak reaction (light blue in original) in the pfll. (Belly skin, Stage 6. Performic acid alcian blue. x 1750.)
(h) A part of the epidermis showing the inner epidermal generation having mature pob, p/?I and pml (arrows). Note a layer of uec between large swollen pal cells and the sgc. (Belly skin, Stage 6. H/E. x 2950.)

556 A . K. MlTTAL A N D J. P. N . SINGH

SCALE EPIDERMIS OF N A T R I X P I S C A T O R 557
The staining properties of the cellular components of different layers of the epidermis are similar to those of Stage I .
Stage 3
The epidermis in Stage 3 is characterized by the complete differentiation of the outer epidermal generation, for in addition to the Oberhautchen, 8- and mesos layers there is maturation of the a- layer and formation of the lacunar tissue and clear layer. Furthermore, the presumptive Oberhautchen and /?-layer of the inner epidermal generation are also differentiated (Plate Ie, f).
The Oberhautchen, B- and mesos layers and the a-layer, now represented by a thin, mature, compact layer devoid of nuclei and cell boundaries, in general show no significant differences in staining properties from those of Stages 1 and 2. The intensity of reactions for -SH groups in the vertical striations and inner margin of the Oberhautchen is, however, relatively weak. The purple reaction with the DDD technique indicates a relatively higher concentration of -SH groups in the a-layer.
The lacunar tissue, immediately below the u-layer, consists of several layers of vertically compressed, almost rectangular spindle-shaped cells, each with a flat, darkly stained nucleus (Plate Ie, f) and staining strongly for DNA. Their cytoplasm is slightly eosinophilic in H/E, PS and VHE, stains blue in MTS and gives only weak reactions for -NH2 groups and bound calcium. The cytoplasm gives weak RNase-labile fluorescence and the nuclei strong DNase-labile fluorescence with various fluorochromes.
The clear layer is composed of a single layer of rectangular cells limiting the inner boundary of the lacunar tissue. The cells of the clear layer have distinct cell boundaries and a prominent, central, spherical nucleus with distinct chromatin and nucleolus (Plate If) that stains moderately for DNA. The cytoplasm is homogeneous, does not stain with H/E, PS, VHE and MTS preparations and shows weak reactions for -NH2 groups and bound calcium, similar to that for lacunar tissue. The cell boundaries, however, are positive for tyrosine (Plate Ig), -SH groups (Plate Ih) and bound calcium and give weak RNase-resistant fluorescence with various fluorochromes (Plate Ii, j).
The presumptive Oberhautchen is a single layer of cells between the outer clear layer and inner presumptive 8-layer (Plate Ie, f). The lateral boundaries of these cells are indistinct. The nuclei, often flat and with compact chromatin material, are basal, enclosed within a vacuole (Plate re) and stain moderately for DNA. In general, the cytoplasm is homogeneous but apically it is striated vertically, imparting saw-like serrations to the margins (Plate Ie). The area between the striations and the rest of the cytoplasm of the presumptive Oberhautchen cells stains with a relatively weaker shade; the vertical striations are strongly eosinophilic, stain bluish purple in VHE and red in MTS. The boundary between the clear layer and the presumptive Oberhautchen, in addition to having staining properties similar to those of the vertical striations, shows moderate reactions for -SH groups (Plate Ih), basic proteins, -NH2 groups and bound calcium and with various fluorochromes (Plate Ti, j ) that are DNase- and RNase-resistant, and weak reactions for -SS groups, tyrosine (Plate Ig) and tryptophan.
The presumptive 8-layer consists of several layers of compactly arranged spindle-shaped cells (Plate Ie, f), each with a flattened nucleus. They show moderate reaction for DNA and weak DNase-labile fluorescence. The nuclei are relatively large and prominent in the lower region of the [&layer and relatively less conspicuous in the more compact outer layers. The cytoplasm is homogeneous and shows eosin colour in H/E and PS, purplish blue in VHE and red in MTS. It

558 A . K. MITTAL A N D J . P. N . SINGH
stains moderately for tyrosine (Plate Ig), tryptophan, -SH groups (Plate Ih), -NH2 groups, basic proteins and bound calcium; reacts weakly for -SS groups and gives DNase- and RNase- resistant fluorescence of moderate intensity (Plate Ii). A narrow zone of cytoplasm around the nuclei remains unstained (Plate Ie). The limiting membranes of these cells are thick, prominent and stain more strongly, In dorsal scale epidermis between cells of the presumptive P-layer a few pigment granules are discernible.
Underlying the presumptive P-layer there are undifferentiated epidermal cells arranged in two to three layers. Further proximally the stratum germinativum is represented by a layer of columnar cells.
Though the epidermis undergoes significant changes in its organization in the subsequent stages, the Oberhautchen and the P-, mesos and a-layers of the outer epidermal generation do not show any special changes in their organization and staining properties.
Stage 4
At this stage the lacunar tissue begins to degenerate, a feature indicated by vacuolization in the cell cytoplasm (Plate IIa) and pycnotic nuclei. A few eosinophilic granular cells appear, spherical, oval or irregular in outline, filled with coarse granular contents strongly eosinophilic in H/E and PS, stained bluish black in VHE and red in MTS preparations.
The relatively flat clear layer cells acquire slight affinity for eosin, stain purplish with VHE and bluish with MTS. In general, the nuclei appear healthy but a few cells have flat pycnotic nuclei.
The staining properties of the various layers of the outer epidermal generation do not differ from those of the previous stage, for various protein end groups.
In the presumptive Oberhautchen cells the flatter nuclei (Plate IIa) strongly react for DNA. The vertical striations now traverse the entire cell (Plate IIa) but still stain in the same way as in Stage 3. The cytoplasmic contents show increased affinity for eosin, stain light orange in MTS and bluish grey in VHE, stain moderately for -SH groups (Plate IIb), bound calcium, -NH2 groups and basic proteins and weakly for tryptophan and tyrosine. The intensity of colour reaction for -SH groups (Plate IIb), -NHz groups and bound calcium is, however, stronger at the narrow apical zone and at the vertical striations. This layer as a whole gives strong reaction for -SS groups and fluoresces strongly with various fluorochromes that are DNase- and RNase-resistant.
The compactly arranged presumptive P-layer cells have less distinct cell boundaries and flat pycnotic nuclei (Plate Ha); they stain strongly for DNA. The cells overall show an increased intensity of reaction for H/E, PS, VHE, and MTS and stain for various protein end groups, very much as in Stage 3. The intensity of reaction for -SH groups and with various DNase- and RNase- resistant fluorochromes is, however, relatively stronger.
The presumptive mesos layer consists of several layers of flattened cells each with a spherical nucleus and distinct nucleolus (Plate Ha); chromatin stained moderately for DNA and there are thick eosinophilic cell boundaries. The cytoplasm is weakly eosinophilic, stains bluish purple in VHE and slightly reddish in MTS, gives weak reactions for -NH2 groups, basic proteins, -SH groups, bound calcium and tyrosine, and remains unstained for - S S groups and tryptophan. The boundaries of these cells give RNase-resistant fluorescence. Between the presumptive mesos layer and the stratum germinativum there are one or two layers of undifferentiated epidermal cells.

SCALE EPIDERMIS OF N A T R I X PISCATOR
Stage 5
559
The lacunar tissue cells are now characterized by an overall increase in vacuolization (Plate IIc). They are infiltrated by eosinophilic granular cells as in Stage 4.
The clear layer is a thin band limiting the inner margin of the lacunar tissue, often separated from the underlying presumptive Oberhautchen. It is eosinophilic in VHE, H/E and PS, stains red in MTS, gives moderate reactions for -SH groups, bound calcium and -NH2 groups and weak fluorescence with various fluorochromes used.
Staining properties of other layers of the outer epidermal generation are like those found in Stage 4, except that the Oberhautchen now shows a weak reaction for -SH groups.
Cells of the presumptive Oberhautchen are anucleate. The cytoplasm in H/E, VHE, MTS and PS preparations stains similarly to that of the presumptive /?-layer. The presumptive Oberhaut- chen is distinguished from the latter by the vertical striations which stain red in MTS preparation. It is more eosinophilic in H/E and PS and purplish blue in VHE, stains very strongly for -SH groups and -SS groups, strongly for basic proteins and bound calcium (Plate IIe) and moderately for -NH2 groups, tryptophan and tyrosine. The areas between the vertical striations stain more weakly.
The presumptive /?-layer is more compact than in the previous stage, having cells with indistinct boundaries (Plate IIc, d). It is weakly eosinophilic, stains grey in VHE and orange in MTS and gives very strong reactions for tyrosine, tryptophan and bound calcium (Plate IIe), and a weak reaction for -SH groups, -SS groups and basic proteins.
The presumptive Oberhautchen and the presumptive /?-layer show very strong DNase- and RNase-resistant fluorescence (Plate 110.
The part of the presumptive mesos layer immediately below the presumptive 8-layer consists of compactly arranged flattened cells with flat, darkly stained pycnotic nuclei (Plate IIc, d). The remaining major, less mature part differs from that of Stage 4 in that cell boundaries are relatively thick and more distinct (Plate IIc, d). The outer compact region of the presumptive mesos layer and its underlying cell boundaries overall show an increase in intensity of various reactions compared with those of the previous stage. They stain moderately for -SH groups, -NH2 groups, tyrosine, basic proteins and bound calcium. In addition there is a weak reaction for -SS groups and tryptophan. The outer, mature part of the presumptive mesos layer and the cell membranes of the inner, immature part strongly fluoresce and thus in this regard are indistinguishable from the presumptive 8-layer (Plate 110. The nuclei of the immature cells also show moderate fluorescence which is DNase-labile.
The most characteristic feature of the epidermis in Stage 5 is the differentiation of the presumptive a-layer beneath the presumptive mesos layer. It comprises two to three layers of swollen, roughly rectangular cells with staining properties similar to those described in Stage 1 (Plate Ild).
Stage 6
In this stage the outer epidermal generation invariably is detached from the underlying inner
The lacunar tissue is a thin layer of shrunken vacuolated cells with pycnotic nuclei. Eosinophilic
The clear layer is a thin chromophilic band limiting the inner boundary of the lacunar tissue.
epidermal generation.
granular cells are relatively few compared with their number in Stages 4 and 5 .

560 A . K . MITTAL A N D J . P . N . SINGH
The staining properties of different layers of the outer epidermal generation and of the presumptive Oberhautchen and presumptive P-layer of the inner epidermal generation do not differ significantly from those of Stage 5 except that the presumptive P-layer loses affinity for bound calcium and stains light orange in MTS preparation.
The presumptive mesos layer is a thin, compact layer devoid of nuclei, limiting the inner boundary of the P-layer from which it is differentiated. It stains red in MTS, purplish blue in VHE, blue in performic acid alcian blue for -SS groups (Plate IIg) and purple in DDD technique for -SH groups. It also shows increased intensity of reactions for other protein end groups compared with those of Stage 5.
The presumptive a-layer, still in the process of differentiation, consists of an outer compact part of flattened cells and an inner part of two to three layers of swollen, roughly rectangular cells (Plate IIh). The cytological details and the staining properties of these cells are similar to those described in Stage 1.
The entire epidermis in all the six stages of the sloughing cycle shows a negative reaction for arginine and keratohyaline.
At all stages the structure of the epidermis at the outer surface of the scales on the back and belly of the body is similar. The back epidermis, however, is relatively thin compared with that of the ventral surface. Furthermore, the mid-line region of each scale is elevated into a keel that is absent on the ventral surface.
Discussion
Periodic changes in the histological organization of snake epidermis associated with sloughing are well known and were first described by Maurer (1892, 1895). Schmidt (1914), Zimmermann & Pope (1948), Klauber (1956), Bechtel (1957) and Goslar (1958) also described changes in the epidermis but, by and large, their descriptions are incomplete and their terminology for various skin layers differs. Maderson (1965) gave a systematic account of the histological events throughout the sloughing cycle in Elaphe taeniura in an attempt to redefine the terminology and homologize his own with that of previous workers. Nevertheless, the terminology of the different epidermal layers proposed by Maderson (1965) and accepted by various research workers could well be unsatisfactory (see below).
Based on histological organization, six stages are distinguished for the epidermis of Narrix piscator during one complete inter-slough period, similar to that described in Elaphe taeniura by Maderson (1 965), Constrictor constrictor by Roth & Jones (1 970) and Natrix natrix by Landmann (1979). Maderson, Chiu et al. ( 1 970) also described an additional stage-Stage 0 in Ptyas korros, characterized by a poorly developed a-layer of the new outer epidermal generation and three to five layers of the presumptive a-cells. Their Stage 1 consists of a well-developed a-layer and the subjacent immature presumptive a-layer cells are fewer than in Stage 0.
In the present work, the epidermis of Narrixpiscator, immediately after sloughing (Stage l), is represented by an incompletely formed and new outer epidermal generation, consisting of the Oberhautchen, P-, mesos and a-layers, a layer of undifferentiated epidermal cells and the stratum germinativum. The a-layer comprises an outer, thin, compact and partially mature region and an inner region consisting of one or two layers of large, swollen, somewhat rectangular cells with finely granular cytoplasm. Cytologically and histochemically these cells resemble the presumptive a-layer cells immediately below the presumptive mesos layer in Stage 5, and also those immediately below the thin, partially mature, compact part of the a-layer in Stage 6. The underlying and lateral

SCALE EPIDERMIS O F NATRIX PISCATOR 56 I
boundaries of the large swollen cells of the a-layer are thick and have staining properties similar to the overlying compact part of the same a-layer, suggesting that these underlying cells are maturing and ultimately incorporated into the outer compact part of the a-layer. As the cells i,i the outer region of the a-layer mature, they are replaced by new generations of presumptive a-layer cells differentiated from the underlying epidermal cells. Thus, in Narrix piscator, differentiation and maturation of the a-layer is a continuous process throughout Stage 1. Increase in thickness of the compact part of the a-layer by the end of Stage 1 supports this view. Again, Downing & Roth (1974) showed that 3H thymidine-labelled cells gradually move through viable strata into the a- layer throughout the resting phase. During the subsequent phases of Stage 1, incorporation of new epidermal cells into the poorly developed a-layer seen immediately after sloughing has also been reported by Roth & Jones ( I 967), Roth & Baden (1967) and Landmann ( I 979). The ‘presumptive lacunar tissue’ consists of a single layer of cells in the outer epidermal generation of Elaphe taeniura (Maderson, 1965). In Stage 1 of the sloughing cycle, its components are incorporated into the overlying a-layer. The clear layer cells of Roth & Jones (1967) in the epidermis of Constrictor constrictor at Stage 1 lie intermittently in the most superficial region of the ‘spinous layer’. Ultrastructurally they resemble the stellate cells in lower layers of the underlying a-layer and cells described by Maderson, Chiu et al. (1970) in the Stage 1 epidermis of Ptyas korros, homologous to the clear cells of Roth &Jones (1967). All these cells could be in the process of maturation leading to their incorporation in the a-layer. Reports describing the appearance of presumptive clear layer cells immediately below the a-layer (Maderson, 1965; Roth & Jones, 1967; Maderson, Chiu et al., 1970) are untenable and cast doubt on the origin of the lacunar tissue cells which differentiate between the a-layer and the clear layer in the subsequent stages. Indeed, the possibility of obtaining lacunar tissue cells by cell proliferation between the presumptive a-layer and presumptive clear layer is excluded as the cells in these layers are in various stages of keratinization and ultimate degeneration.
Since maturation of the a-layer is a continuous process throughout Stage 1, the term Stage 0 proposed by Maderson, Chiu et al. ( 1 970) for the condition immediately after sloughing should be discarded in order to avoid further confusion.
In the present study, lacunar tissue, the clear layer and the presumptive Oberhautchen could not be distinguished in the epidermis of Natrixpiscator throughout Stages 1 and 2. In the epidermis of Natrix narrix at the end of Stage 1, Landmann (1 979) reported the differentiation of lacunar tissue, which consisted of one to four layers of flattened cells indistinguishable ultrastructurally from the overlying immature a-layer cells, and the clear layer below. In Stage 2, he described the differentiation of the presumptive Oberhautchen, the peripheral plasma membrane of which regularly protrudes into the overlying clear layer. Maderson (1965) described the formation of several layers of cells of the presumptive inner epidermal generation cells, in Stage 2. He regarded the cells in the outermost layer, though not clearly distinguishable from rest of the underlying cells, as presumptive Oberhautchen.
The proliferation of stratum germinativum cells, which results in the formation of undifferen- tiated epidermal cells, is a continuous process. These undifferentiated epidermal cells transform into specialized cells characteristic of various layers of the epidermis. Only one or two layers of undifferentiated epidermal cells overlie the stratum germinativum in most stages of the sloughing cycle. The presence of several layers of undifferentiated epidermal cells between the stratum germinativum and the a-layer in Stage 2 of Natrix piscator may thus be a result of temporary slowing down of differentiation of the undifferentiated epidermal cells. Maderson (1965) likewise observed several layers of cells overlying the stratum germinativum and, in contrast, thought that they were due to active proliferation of stratum germinativum cells.

562 A. K . MITTAL A N D J . P. N. SINGH
The lacunar tissue and clear layer of the outer epidermal generation and the presumptive Oberhautchen and presumptive P-layer of the inner epidermal generation are clearly differentiated in the epidermis of Natrix piscutor in Stage 3 . Various histochemical tests reveal that throughout the sloughing cycle lacunar tissue cells do not undergo keratinization. However, these cells become vertically compressed in Stage 3, show vacuolization and signs of disintegration in Stage 4 and finally become very much shrunken and vacuolated, with pycnotic nuclei, in Stage 6 when lacunar tissue is represented by a thin layer. These changes appear to be initiated by the reduction or cessation of the supply of nutrients and other body fluids from the inter-cellular spaces of the inner epidermal generation. This view is supported by evidence that tight junctions have formed between the lateral plasma membranes of the clear layer in Stage 4 of Natrix natrix (Landmann, 1979). Roth & Jones (1970) and Landmann (1979), in Constrictor constrictor and Natrix natrix, respectively, also reported that lacunar tissue cells do not keratinize and remain immature until shedding occurs. In contrast, Jackson & Sharawy (1978) reported that lacunar tissue cells in skin of the rat snake, Eluphe obsoleta, gradually mature, keratinize and contribute to the thickness of the a-layer.
Flaxman, Maderson, Szab6 & Roth (1968) reported that lacunar tissue cells in Anolis carolinensis, in vitro, gradually mature into a-cells. However, studies on the epidermis of lizards have shown that, in general, lacunar tissue cells do not keratinize even up to the time of the shedding of the outer epidermal generation (Maderson, 1966; Maderson & Licht, 1967), except in certain specialized regions, e.g. around the climbing setae of Anolis carolinensis, where some indications of patchy keratinization are seen just before sloughing. However, in Dipsosaurus, Uma and Sphenodon, after a brief period of true lacunar tissue formation, the entire innermost portion of the outer epidermal generation keratinizes, producing tissue essentially similar to the overlying a-layer (Maderson, Mayhew & Sprague, 1970).
Maderson’s ( 1965) description regarding lacunar tissue in the snake is somewhat confusing and has not been confirmed. According to Maderson, lacunar tissue forms in Stage 4 as a result of the disintegration of the clear layer cells and the presence of a layer of nuclear remnants associated with the inner margins of the a-layer. Here it consists of a series of protoplasmic strands with large lacunae between them and this is followed by breakdown in Stage 5 . Maderson, Chiu et al. (1970) also reported similar observations in Ptyas korros, that the epidermis in Stages 2 4 resembles that in similar stages described for Eluphe taeniura by Maderson (1965). However, they did not refer to lacunar tissue in Stages 5 and 6. Nor did Goslar (1958, 1964) refer to lacunar tissue in his descriptions.
Positive reactions for tyrosine, bound calcium, and -SH groups, and weak RNase-resistant fluorescence at the boundaries of clear layer cells (the intensity slightly increasing in subsequent stages as the cells flatten and become enucleated) suggest that as the epidermis nears sloughing clear cells gradually transform into weakly keratinized cells. This view is supported by results of Roth & Jones ( I 970), who reported clear layer cells, initially indistinguishable ultrastructurally from those of the lacunar tissue, to keratinize gradually and resemble a-layer cells at the time of sloughing. Landmann (1975) described the clear layer as a chromophilic band at the lower margin of the outer epidermal generation; in 1979 he also reported ultrastructural observations similar to those of Roth & Jones (1970), and further found that keratinization in the clear layer proceeds basally towards the apical region and is completed by late Stage 5. Maderson (1965) described a distinct clear layer of cells, unstained in Stage 3 of Elaphe taeniura, which disappears in Stage 4 to give rise to part of the lacunar tissue. He did not refer to this layer in Stages 5 and 6. However, studies on the epidermis of lizards (Maderson, 1966; Maderson & Licht, 1967; Flaxman et al.,

SCALE EPIDERMIS OF N A T R I X P I S C A T O R 563
1968; Maderson, Mayhew et al., 1970) reveal that the clear layer does not disintegrate but remains attached to the outer epidermal generation at the time of shedding. Maderson’s (1965) interpretation of the origin and maturation of the lacunar tissue and clear layer in the epidermis of snakes is therefore open to doubt or applicable only to the epidermis of EIaphe iaeniura.
The Oberhautchen in the epidermis of Natrix piscator is separate and distinct from the underlying /?-layer though they are intimately associated in all stages of the sloughing cycle. There are significant differences in staining properties of various proteins of the Oberhautchen and the /?-layer, for in contrast to the highly chromophilic nature of the Oberhautchen, the mature /?-layer is either unstained or stains weakly for most protein reactions (Table I). Nevertheless, in histochemical preparations for the amino acids, tyrosine and tryptophan, the mature 8-layer stains very strongly compared to the moderate reaction of the Oberhautchen.
As the Oberhautchen matures it gradually increases the intensity of its reactions for -SH groups and -SS groups. In contrast, in the 8-layer, the intensity of reactions for -SH groups, though strong initially, gradually declines in subsequent stages. The intensity of reaction for -SS groups, however, remains weak in the 8-layer throughout its existence. The strong reactions for -SS groups at the surface of the epidermis of Natrixpiscator (Banerjee & Mittal, I978a), in the late third/early fourth stage of the sloughing cycle, are from the Oberhautchen and not from the superficial part of the b-layer, which was earlier misinterpreted. Staining with silver (See Bailey, 1965 cited in Maderson, 1966) or with barium permanganate (Ruibal & Ernst, 1965) also distinguished the Oberhautchen as a separate layer. Others, however, considered the Oberhautchen as a superficial part of the 8-layer (Maderson, 1965, 1967; Ernst & Ruibal, 1966; Roth & Jones, 1967, 1970; Maderson & Licht, 1967; Lillywhite & Maderson, 1968).
As yet there are no histochemical methods available to specify a- or 8-keratins. 8-keratin fibrils contain more cystine than a-keratin and this disulphide-containing amino acid does not fit easily into the a-helix (Fraser, MacRae & Rogers, 1972; Spearman, 1973, 1977). The mature /?-layer in Natrix piscator, which in other squamates contains 2-3 nm 8-keratin filaments (Alexander, 1970; Alexander & Parakkal, 1969; Parakkal & Alexander, 1972; Maderson, Flaxman, Roth & Szabo, 1972; Landmann, 1979), shows a weak reaction for -SS groups indicating the presence of only small amounts of cystine. Goslar (1958) reported that highly polymeric -SS compounds are traceable only in the outer layers. Further investigations are needed to clarify this point.
The present investigation reveals that the mature Oberhautchen has most of its reactions restricted mainly to the vertical striations. In many respects they resemble the mature a- and mesos layers. The three layers are eosinophilic, stain red in MTS and purplish blue in VHE, give strong reactions for -SH groups, -NH2 groups, bound calcium and basic proteins. Nevertheless, the mature Oberhautchen is unique. On one hand it resembles the a-layer in showing moderate reactions for tyrosine and tryptophan; these are very strong in the mesos layer. On the other hand i t resembles the mesos layer in showing very strong reactions for -SS groups; the a-layer does not stain for the -SS groups.
The mature Oberhautchen in the outer and inner epidermal generations contains regularly spaced vertical striations traversing its full thickness. In the initial stages of the differentiation these vertical striations are restricted apically but subsequently they gradually invade the deeper regions. The intra-cellular oblique structures in the presumptive Oberhautchen that subsequently split the individual cell (Maderson, 1965) and the bristle-like structures in the limiting zone between the so called ‘old loose keratinized layer’ and the ‘young Oberhautchen’ (Goslar, 1958, 1964), may be regarded as the homologues of these vertical striations. Other workers, without referring to the vertical striations of the Oberhautchen, have described serrations, sculptured

564
patterns, micro-ornamentation, spines, ridges, setae, teeth and bristles on the surface of the squamate epidermis (Maderson, 1966; Ernst & Ruibal, 1966; Maderson & Licht, 1967; Flaxman et al., 1968; Lillywhite & Maderson, 1968; Maderson, Mayhew et al., 1970). Roth & Jones (1970), Maderson et al. (1972) and Landmann (1979) made no reference to vertical striations in the epidermis of Constrictor constrictor, Anolis carolinensis and Natrix natrix, respectively. However, they reported small finger-like outgrowths protruding from the presumptive Oberhautchen into the overlying clear layer and they considered that these projections represent the primordia seen on the surface of the mature P-layer.
The vertical striations in the Oberhautchen may reinforce this most superficial layer of the epidermis, and act as a protection against the wear and tear by abrasion to which snakes are subjected during movements on the ground. Furthermore, the large amounts of -SS groups in the Oberhautchen, which when associated with hard fibrous keratin (Matoltsy, 1962; Barrnett & Sognnaes, 1962) confer strength and durability on this fibrous protein (Spearman & Riley, 1969), may also be responsible for giving this layer the hardness, durability and mechanical strength that its function requires. The calcium in the Oberhautchen, which according to Jarrett, Spearman, Riley & Cane (1965) is linked to phospholipids possibly attached to keratin side-chains, may provide additional hardness to this layer, to assist in crawling movements, as in the insect cuticle (see Wigglesworth, 1973).
The presence of high concentrations of tyrosine in the mature p- and mesos layers and the moderate amounts in the a-layer and Oberhautchen of the epidermis of Natrixpiscator (Table I ) is of interest. Small quantities of proteins, distinguishable by their high content of glycyl and aromatic residues, especially tyrosine, occur in wool (Gillespie, 1960; Harrap & Gillespie, 1963; DeDeunvaerder, Dobb & Sweetman, 1964; O’Donnell & Thompson, 1964; Zahn & Biela, 1968). Fraser, Gillespie & Macrae ( 1 973), who studied keratinized tissues using spectrophotometer and X-ray diffraction techniques, confirmed the presence of tyrosine- and glycine-rich protein in mammalian hard keratins; they are particularly abundant in porcupine and echidna quills. Information available on proteins of hard and soft keratins from reptiles and birds is scanty. Fraser et al. (1 973) reported tyrosine-rich proteins in the beak of Dacelo gigas and in the claw of Varanus uarius, representing avian and reptilian hard keratins, respectively. De Deurwaerder et al. (1964), Andrews, Inglis & Williams (1966) and Bradbury, Chapman & King (1965) suggested that these high-tyrosine proteins originate from inter-cellular layers and thus, by inference, do not form part of the intra-cellular micro-fibril-matrix texture. However, Fraser et a f . (1973) reported such proteins to occur in the matrix and concluded that, though the special properties which glycine- tyrosine-rich proteins might confer upon the keratinized tissues are not immediately apparent, it seems likely that they play specific structural roles in mammalian hard keratins. These tyrosine- rich proteins may perform a similar function in snake keratins.
Tryptophan is present in various keratinized epidermal layers of Natrix piscator. Unfortuna- tely, no data are available on the presence of tryptophan in reptilian epidermis. However, small amounts of tryptophan occur in the rachis of the bird feather (Harrap & Woods, 1964). No reference to tryptophan is made in various reports on the amino acid analysis of the protein fraction of a wide variety of mammalian hard keratinized tissues, e.g. rhinoceros horn and guinea- pig hair (Crewther, Gillespie, Harrap & Inglis, 1966), human hair and nail (Crewther et al., 1966), human epidermis stratum corneum (Baden & Bonar, 1968), cow lip epidermis (O’Donnell, 1971). and wool (Thompson & O’Donnell, 1965; Zahn & Biela, 1968; Gillespie & Darskus, 1971).
The presumptive u-layer cells of Natrix piscator first appear in Stage 5 of the sloughing cycle. They are distinguished from the overlying mesos layer cells and the underlying undifferentiated
A. K . MITTAL A N D J . P. N. SlNGH

SCALE E P I D E R M I S OF NATRIX PISCATOR 565
epidermal cells by their swollen, somewhat rectangular shape, finely granular, almost chromopho- bic, cytoplasm and spherical or oval, centrally placed nuclei. Maderson, Chiu ef al. (1970) and Roth & Jones (1970) illustrated the presumptive a-layer cells in Stage 5 (see their figs 3 and 4, respectively), the former workers reporting that these cells appear and mature at Stage 6 in Ptyas korros, and the latter workers (who did not describe the stage of the sloughing cycle) that in Constrictor constrictor the a-layer arises during the proliferative phase immediately after the formation of the mesos layer. Landmann (1979) reported the emergence and keratinization of the a-layer in Nutrix nutrix as the characteristic feature of Stage 6. Maderson (1965) described cells underlying the presumptive /%layer in Stage 3 to represent the presumptive a-layer cells of the inner epidermal generation. A distinct presumptive a-layer in late Stage 3--early Stage 4 of Natrix piscator could not be distinguished by Banerjee & Mittal(l978a) and, following Maderson (1963, they concluded that cells of the inner epidermal generation below the presumptive /I-layer may form this layer. The results of Maderson (1965) and Banerjee & Mittal(1978a) were not confirmed in the present study and it seems that undifferentiated epidermal cells immediately below the presumptive /I-layer were misinterpreted as presumptive a-layer cells.
The presumptive mesos layer first appears immediately below the presumptive /I-layer in Stage 4. It gradually matures and in Stage 6 is a thin, compact, highly chromophilic layer. Roth & Jones (1970), Maderson, Chiu et al. (1970) and Landmann (1979) likewise described a presumptive mesos layer to first appear in Stage 4. Maderson (1965) described a layer in Stage 4 Elaphe taeniura (see his plate IIa) of cells similar to those described in the same stage of the present study, but he assumed these to be of the presumptive a-layer.
Cells in the mesos layer probably contain a- and /I-keratin and represent a gradual transition from the /I- to a-pattern corresponding to the adjacent /I-and a-layers (Alexander, 1970). Roth & Jones (1 970), however, reported only a-keratin in cells of the mesos layer. According to Landmann (1979), the mesos layer, like the clear layer, shows no keratin pattern. The keratin-like contents of these layers contain poorly visible 10-15 nm filaments but no matrix. According to Matoltsy (1976) such keratin must be relatively flexible and elastic.
Shedding of the outer epidermal generation of squamates appears to be a purely mechanical process. Ultrastructural studies on the epidermis of Nutrix natrix reveal that desmosomes between the presumptive Oberhautchen and the clear layer begin to degenerate in Stage 4 and they have degenerated completely in Stage 5 (Landmann, 1979). The loss of desmosomal attachments results in the separation of the clear layer and presumptive Oberhautchen, though peripheral plasma membranes of the presumptive Oberhautchen extending into the clear layer (Roth & Jones, 1970; Landmann, 1979) probably help to keep the outer and inner epidermal generations together for considerably longer periods. This feature may protect the animal during this period of the sloughing cycle, when the inner epidermal generation is represented by incompletely keratinized layers. When sloughing occurs the clear layer is gradually separated from the presumptive Oberhautchen by a force exerted through gradual shrinkage, effected by the simultaneous disintegration of the cytoplasmic contents of the cells of the lacunar tissue still firmly attached to the mature a-layer on one side and the clear layer on the other.
The authors wish to thank Dr H . Fox, University College London, for critically reading the manuscript and for many helpful suggestions.

566 A. K. MITTAL A N D J . P. N. SINGH
REFERENCES
Adams, C. W. M. (1957). A p-dimethylaminobenzaldehyde nitrite method for the histochemical demonstration of
Adams, C. W. M. & Sloper, J. C. (1955). Technique for demonstrating neurosecretory material in the human
Alexander, N. J. (1970). Comparison of a and j keratin in reptiles. Z. Zellforsch. mikrosk. Anat. 110: 153-165. Alexander, N. J. & Parakkal, P. F. (1969). Formation of a- and j-type keratin in lizard epidermis during the molting cycle.
Andrews, M. W., Inglis, A. S. & Williams, V. A. (1966). Chemicalchanges in thecuticle of oxidized wool. T e x f . Res. J . 36:
Baden, H. P. & Bonar, L. (1968). The a-fibrous proteins of epidermis (Humen). J. inuesf. Derm. 51: 478483. Baden, H. P., Roth, S. 1. & Bonar, L. C. (1966). Fibrous proteins of snake scale. Nature, Lond. 212 498499. Baker, J. R. (1947). The histochemical recognition of certain guanidine derivatives. Q. JI microsc. Sci. 88: 115-122. Baker, J. R. (1956). The histochemical recognition of phenols, especially tyrosine. Q. JI microsc. Sci. 97: 161-164. Banerjee, T. K. & Mittal, A. K. (19780). Histochemistry of the epidermis of the Chequered water snake Natrixpiscator
Banerjee, T. K. & Mittal, A. K. (19786). Epidermal mucous cells in the chequered water snake Natrixpiscator (Colubridae,
Banerjee, T. K. & Mittal, A. K. (1980). Histochemistry of snake epidermis. Linn. Soc. Symp. Ser. No. 9: 23-32. Barrnett, R. J. & Seligman, A. M. (1952). Histochemical demonstration of protein-bound sulphydryl groups. Science,
Barrnett, R. J. & Sognnaes, R. F. (1962). Article in Fundamentals ofkerafinization. Bucher, E. 0. & Sognnaes, R. F. (Eds).
Bechtel, H. B. (1957). Some microscopic aspects of ecdysis in snakes. Herpetologica 13: 177-181. Bouin, P. (1 897). Phenomknes cytologiques anormaux dans I’ histogendse. Nancy. Bradbury, J. H., Chapman, G. V. &King, N. L. P. (1965). The chemical composition of wool-Ill. Analysis ofcuticle, skin
flakes and cell membrane material. Proc. int. Wool Text. Res. Conf. 3(1): 359-364. Crewther, W. G., Gillespie, J. M., Harrap, B. S. & Inglis, A. S. (1966). Low sulphur proteins from a-keratins. Inter-
relationship between their amino acid compositions, a-helix contents, and the supercontraction of the parent keratin. Biopolymers 4 905-916.
DeDeurwaerder, R. A., Dobb, M. G. & Sweetman, B. J. (1964). Selective extraction of a protein fraction from wool keratin. Nature, Lond. 203 4849 .
Downing, S. W. & Roth, S. I. (1974). The derivation of the cells of the epidermal strata of the boa constrictor (Constrictor constrictor). J . invest. Derm. 6 2 450-457.
Ernst, V. & Ruibal, R. (1966). The structure and development of the digital lamellae of lizards. J. Morph. 120 233-266. Feulgen, R. & Rossenbeck, H. (1924). Mikroskopisch-chemischer Nachweis einen Nucleinsaure von Typus der
Flaxman, B. A., Maderson, P. F. A., Szab6, G. 8i Roth, S. I. (1968). Control of cell differentiation in lizard epidermis in
Fraser, R. D. B., Gillespie, J. M. & MacRae, T. P. (1973). Tyrosine-rich proteins in keratins. Comp. Biochem. Physiol. 44:
Fraser, R. D. B., MacRae, T. P. & Rogers, G. E. A. (1972). Keratins. their composition. structure and biosynthesis.
Gharpurey, K. G. (1962). Snakes of India and Pakistan. Bombay: Popular Prakashan. Gillespie, J. M. (1960). The isolation and properties of some soluble proteins from wool. I . The isolation of a low-sulphur
protein. Aust. J. biol. Sci. 1 3 81-102. Gillespie, J. M. & Darskus, R. L. (1971). The relation between tyrosine content of various wools and their content of a class
of proteins rich in tyrosine and glycine. Aust. J. biol. Sci. 2 4 1189-1 197. Goslar, H. G. (1958). Beitrage 2um Hautungsvorgang der Schlangen. I . Mitteilung. Histologische und topochemische
Untersuchungen an der Haut von Natrix natrix L. wahrend der Phasen des normalen Hautungszyklus. Acra histochem. 5: 182-212.
Goslar, H. G. (1964). Beitrage zum Hautungsvorgang der schlangen. 2. Mitteilung. Studien zur Fermenttopochemie der Keratogeneses und Keratolyse am Modell der Reptilien haut. Acra histochem. 17: 1-60,
tryptophan and related compounds. J. elin. Path. 1 0 56.
hypothalamus. Lancet 1955(1): 651-652.
Z . ZellJorsch. mikrosk. Anat. 101: 72-87.
407412.
(Colubridae, Squamata). J. Zool.. Lond. 185: 415435.
Squamata). A cytochemical investigation. 2001. J. Linn. Soc. 63: 289-293.
Wash. 116 323-327.
Pubb Am. Ass. Adumt Sci. No. 70: 2745.
Thymonudeinsaure etc. Z. physik. Chem. 135: 203.
vitro. Devl Biol. 18: 354-374.
943-947. *
Springfield, Illinois: Charles C. Thomas.

SCALE E P I D E R M I S OF N A T R I X P I S C A T O R 567
Gurr. E. (1958). Methods of analytical histology and histochernistry. London: Leonard Hill. Harrdp, B. S. & Gillespie, J. M. (1963). A further study on the extraction of the reduced proteins from wool. Ausr. J. biol.
Harrap, B. S. & Woods, E. F. (1964). Soluble derivatives of feather keratins. 1. Isolation, fractionation and amino acid
Helly, K. (1903). Eine Modification der Zenkerschen Fixirungsflussigkeit. Z. wiss. Mikrosk. 20: 41 3. Hoge, A. R. & Santos, P. S. (1953). Submicroscopic structure of ‘stratum corneum’ of snakes. Science, N. Y. 118 4 1 W 1 I , Horstmann, E. (1964). Elektronenmikroskopische Untersuchungen an der Epidermis von Reptilien. Anaf. Anz. 113: 87-
Ionescu, M. D. & Miscalencu, D. (1972). Fine structure of the horny cells in the Nafrix narrix (L.) snake epidermis. Anal.
Jackson, M. K.&Sharawy, M. (1978). Lipidsandcholesterolcleftsin thelacunarcellsofsnakeskin. Anaf. Rec. 190:4146. Jarrett, A,, Spearman, R. I. C. & Hardy, J. A. (1959). The histochemistry of keratinization. Br. J . Derm. 71: 277-295. Jarrett, A., Spearman, R. 1. C., Riley, P. A. & Cane, A. K. (1965). The distribution of epidermal phospholipids and their
Klauber, L. M. (1956). Rarrlesnakes. Their habits, lilp histories. and influence on mankind I , 2. Berkeley, Los Angeles:
Landmann, L. (1975). The sense organs in the skin of the head of Squamata (Reptilia). Israel J. 2001. 24: 99-135. Landmann, L. (1979). Keratin formation and barrier mechanisms in the epidermis of Natrix natrix (Reptilia: Serpentes):
Landmann, L., Stolinski, C. & Martin, B. (1981). The permeability barrier in the epidermis of the grass snake during the
Lillie, R. D. (1954). Histoparhologic lechniques andpractical hisfochemistry. New York: The Blakistan Division, McGraw-
Lillywhite, H. & Maderson, P. F. A. (1968). Histological changes in the epidermis of the subdigital lamellae of Anolis
Maderson, P. F. A. (1965). Histological changes in the epidermis of snakes during the sloughing cycle. J. Zool., Lond. 146:
Maderson, P. F. A. (1966). Histological changes in the epidermis of Tokey (Gekko gecko) during the sloughing cycle. J . Morph. 1 1 9 39-50.
Maderson, P. F. A. (1967). The histology ofthe escutcheon scales of Gonarodes (Gekkonidae) with a comment on squamate sloughing cycle. Copeia 1967: 743-752.
Maderson, P. F. A,. Chiu, K . W. & Phillips, J . G . (1970). Changes in the epidermal histology during the sloughingcycle in the rat snake Plyas korros Schlegel, with correlated observations on the thyroid gland. Eiol. Bull. mar. biol. Lab. Woods Hole. 139 30&312.
Maderson, P. F. A,, Flaxman, B. A,, Roth, S. 1. & Szabo, G . (1972). Ultrastructural contributions to the identification of cell types in the lizard epidermal generation. J. Morph. 136 191-210.
Maderson. P. F. A. & Licht, P. (1967). Epidermal morphology and sloughing frequency in normal and prolactin treated Anolis carolinensis (Iguanidae, Lacertilia). J . Morph. 123 157-171.
Maderson, P. F. A,. Mayhew, W. W. & Sprague. G. (1970). Observations on the epidermis of desert living Iguanids. J . Morph. 130: 25-36.
Mallory, F. B. (1936). The aniline-blue stain. Slain Tech. 11: 101-102. Matoltsy, A. G . (1962). Article in Fundamenrals of kerarinizarion. Butcher, E. 0. & Sognnaes, R. F. (Eds). Publs Am Ass.
Matoltsy, A. G . (1976). Keratinization. J . invest. Derm. 67: 2Ck25. Maurer, F. ( I 892). Haut-Sinnesorgane, Feder- und Haaranlagen und deren gegenseitige Bezichungen etc. Morph. Jb. 18:
Maurer, F. (1895). Die Epidermis und ihre Abkommlinge. Leipzig. Miscalencu, D. ( 1974). Ultrastructural modifications in the keratinization process of the intermediate-layer cells in the
Miscalencu, D. & Ionescu, M. D. (1972). Fine structure of the basal layer of the Nafrix nafrir (L.) epidermis. Anaf. Anz.
Miscalencu, D. & Ionescu. M. D. (1973). The fine structure of the epidermis and dermal chrornatophores in Vipera
Sci. 1 6 542-556.
composition. Biochem. J . 9 2 8-18.
94.
Anz. 132 376-38 I .
relation to alkaline phosphatase activity in the granular layer. J. invesf. Derm. 44: 31 1-319.
University of California Press.
an ultrastructural study. J. Morph. 162 93-126.
resting stage of the sloughing cycle. Cell Tissue Res. 215: 369-382.
Hill Book Co.
carolinensis during the shedding cycle. J. Morph. 125: 379402.
98-1 13.
Advmi Sci No. 70: 1-27.
7 17-804.
epidermis of Narrix narrix L. Acfa anal. 87: 375-384.
131: 97-104.
umrnodyres (L.). Acfa anal. 8 6 I 1 1-122.

568 A. K. MITTAL A N D J . P. N. SINGH
ODonnell, 1. J. (1971). The search for a simple keratin-the precursor keratins from cow's lip epidermis. Aus/. J. hiol. sci .
O'Donnell, 1. J. &Thompson, E. 0. P. (1964). Studies on reduced wool. IV. The isolation of a major component. Aust. J.
Parakkal, P. F. & Alexander, N. J. (1972). Keratinization. A surcey of verrebrure epirhelia. New York, London: Academic
Pears, A. G. E. (1968). Hisrochemistry, theoretical and analytical 1. London: J. & A. Churchill Ltd. Roth, S. I. & Baden, H. P. (1967). An autoradiographic study of the sites of protein synthesis in the epidermis of the Indigo
Roth, S. I. &Jones, W. A. (1967). The ultrastructure and enzymatic activity of the boa constrictor (Constrictor con.s/rictor)
Roth, S . I. &Jones, W. A. (1970). The ultrastructure of epidermal maturation in the skin of boa constrictor (Cons/rictor
Ruibal, R. & Ernst, V. (1965). The structure of digital setae of lizards. J. Morph. 117: 271-294. Schmidt, W. J. (1914). Studien am Integument der Reptilien. 5. Anguiden. Zool. Jb. (Anar.) 38: 1-102. Smith, M. A. (1961). Reptilia-Amphibia 111-Serpentes. In The fauna of British India. Ceylon and Burma. including the
whole of /he Indo-Chinese sub-region. Sewell, R. B. S. (Ed.). London: Taylor & Francis Reprinted, Dehradun; Survey of India Press.
24: 1219-1234.
bid . sci. 17: 973-989.
Press.
snake (Drymarchon corais couperi). J . exp. Zool. 165 345-354.
skin during the resting stage. J. Lllrrastruct. Res. 1 8 304-323.
constricror). J. Ultrasrruct. Res. 32: 69-93.
Spearman, R. I. C. (1966). The keratinization of epidermal scales, feathers and hairs. Biol. Reo. 41: 59 66. Spearman, R. 1. C. (1973). The integument. A text hook of skin biology. London: Cambridge University Press. Spearman, R. 1. C. (1977). Keratins and keratinization. Symp. zool. Soc. Lond. No. 39: 335 352. Spearman, R. I. C. & Riley, P. A. (1969). Acomparison of the epidermis and pigment cells of the crocodile with those in two
Thompson. E. 0. P. & ODonnell, I. J. (1965). Studies on reduced wool. V. A comparison of the two major components.
Whitaker, R. (1978). Common Indian snakes. Afieldguide. Delhi: The Macmillan Company of India Ltd. Wigglesworth, V. B. (1973). The principles of insect physiology. 7th edn. London: Chapman & Hall Ltd. Yasuma, A. & Itchikawa, T. (1953). The Ninhydrin-Schiff and Alloxan-Schiff staining method: A new histochemical
Zahn, H. & Biela, M. (1968). Uber die lsolierung tyrosinreicher Proteine aus Wolle. Tex/.-Pra.x. 2 3 103-106. Zimmermann, A. A. & Pope, C. H. (1948). Development and growth of the rattlesnake. Fieldiana (Zool.) 3 2 357413.
lizard species. Zool. J. Linn. Soc 48: 453466.
Ausr. J. biol. Sci. 18: 1207-1225.
method for protein. J. Lab. d i n . Med. 41: 296-306.
Abbreviations used in the figure and the plates
a-layer /?-layer basement membrane clear layer large swollen cells lacunar tissue mesos layer Oberhautchen outer compact part of the presumptive a-layer presumptive a-layer presumptive /?-layer presumptive mesos layer presumptive Oberhautchen stratum germinativum cells undifferentiated epidermal cells





























