Embed Size (px)
Citation preview

Sandflies of the Phlebotomus perniciosus complex:mitochondrial introgression and a new sibling speciesof P. longicuspis in the Moroccan Rif
B. PESSON* , J . S . READY y, I . BENABDENNBI* , J . MARTIN-SANCHEZ z,
S . ESSEGHIR y, § , M. CADI-SOUSSI{, F . MORILLAS-MARQUEZ z and
P . D. READY y
*Laboratoire de Parasitologie, Faculte de Pharmacie, Universite Louis Pasteur – Strasbourg I, Illkirch, France, yDepartment of
Entomology, Natural History Museum, London, U.K., zFacultad de Farmacia, Universidad de Granada, Granada, Spain,§Laboratoire d’Epidemiologie et d’Ecologie Parasitaire, Institut Pasteur de Tunis, Tunisia and {Faculte de Medecine et de
Pharmacie, Universite Mohamed V, Rabat, Morocco
Abstract. The bloodsucking adult females of Phlebotomus perniciosus Newsteadand P. longicuspis Nitzulescu (Diptera: Psychodidae) are important vectors of theprotozoan Leishmania infantum Nicolle (Kinetoplastida: Trypanosomatidae) inwestern Mediterranean countries. The species status of the two phlebotominesandflies was assessed, along with the epidemiological implications. Individualsandflies from three Moroccan Rif populations were characterized morphologic-ally, isoenzymatically (by the isoelectrofocusing of alleles at the polymorphicenzyme loci of HK, GPI and PGM), and by comparative DNA sequence analysisof a fragment of mitochondrial Cytochrome b (mtDNA). By reference to thecharacter profiles of specimens from other locations, including southern Spainand the type-locality countries, the Moroccan flies were placed in three lineages:first, the lineage of P. perniciosus, which contained two mtDNA sublineages, one(pnt) widely distributed and associated with the morphology of the male typesfrom Malta, and the other (pna) associated with a P. longicuspis-like malemorphology; second, the lineage of P. longicuspis sensu stricto, including typicalforms from Tunisia; and third, a new sibling species of P. longicuspis. ThemtDNA sublineage (pnt) of typical P. perniciosus was also found in someP. longicuspis from Morocco, indicating interspecific hybridization. The typicalrace of P. perniciosus occurs in Italy as well as in Malta, Tunisia andMorocco. It isreplaced in southern Spain by the Iberian race (with the pni mtDNA sublineage).The discovery of interspecific gene introgression and a new sibling species meanthat previous records of the two morphospecies do not necessarily reflect their truevectorial roles or geographical and ecological distributions.
Key words. Phlebotomus longicuspis, morphology, Phlebotomus perniciosus com-plex, sibling species, isoenzymes, mitochondrial introgression, Morocco.
Introduction
In this report we assess the species status of various morpho-
logical forms and populations of two closely related taxa in
the Phlebotomus perniciosus species complex (Esseghir et al.,
2000) and discuss the resulting epidemiological implications.
Correspondence: Dr Paul Ready, Molecular Systematics
Laboratory, Department of Entomology, Natural History
Museum, Cromwell Road, London SW7 5BD, U.K. E-mail:
Medical and Veterinary Entomology (2004) 18, 25–37
# 2004 The Royal Entomological Society 25
Phlebotomus perniciosus Newstead and P. longicuspis
Nitzulescu are classified in the subgenus Larroussius
Nitzulescu (Seccombe et al., 1993) and, in the western
Mediterranean, their haematophagous females are vectors
of zoonotic leishmaniasis caused by Leishmania infantum
Nicolle (Protozoa, Trypanosomatidae). The domestic dog
and wild foxes are the reservoir hosts, and cutaneous and
visceral human disease is frequent in rural communities, both
in south-west Europe and in the Maghreb region of north-
west Africa (Killick-Kendrick, 1990; Rioux & Lanotte, 1990).
Described from Malta (Newstead, 1911), P. perniciosus is
widely distributed in the western Mediterranean Basin:
from Portugal to Croatia in Europe, and from Morocco
to Libya in north Africa, where it is found predominantly in
the sub-humid and semi-arid bioclimate zones (Rioux et al.,
1984). Phlebotomus longicuspis was described from Tunisia
(Nitzulescu, 1930), and until 1982 was recorded only from
Libya and other countries of the Maghreb (Algeria and
Morocco), where it is most frequent in the semi-arid, arid
and per-arid bioclimate zones (Rioux et al., 1984). Then
males were reported from southern Spain, in the regions
of Andalucia (Morillas Marquez et al., 1982) and Murcia
(Martınez Ortega et al., 1982).
For many years considered indistinguishable, the females
of P. perniciosus and P. longicuspis were separated by Leger
et al. (1983) according to the form and position of the dilata-
tion on each individual spermathecal duct: spherical and
basal or heart-shaped and subbasal, respectively. As for the
males, they were originally separated by the terminal struc-
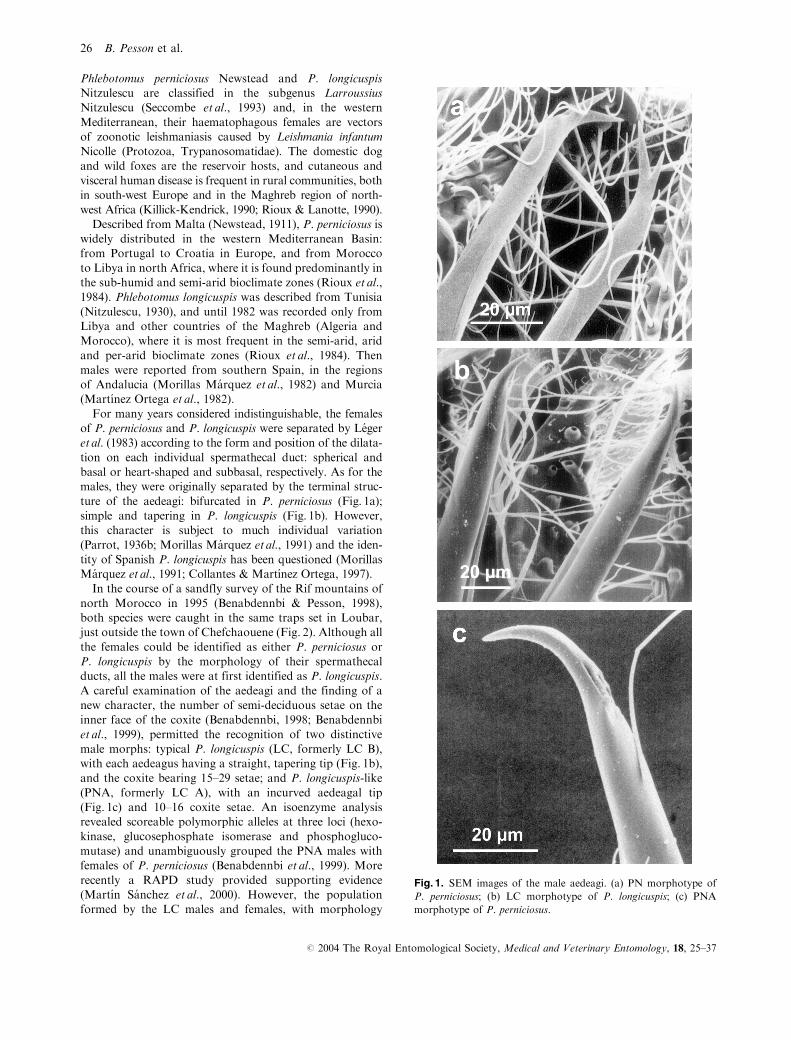
ture of the aedeagi: bifurcated in P. perniciosus (Fig. 1a);
simple and tapering in P. longicuspis (Fig. 1b). However,
this character is subject to much individual variation
(Parrot, 1936b; Morillas Marquez et al., 1991) and the iden-
tity of Spanish P. longicuspis has been questioned (Morillas
Marquez et al., 1991; Collantes & Martınez Ortega, 1997).
In the course of a sandfly survey of the Rif mountains of
north Morocco in 1995 (Benabdennbi & Pesson, 1998),
both species were caught in the same traps set in Loubar,
just outside the town of Chefchaouene (Fig. 2). Although all
the females could be identified as either P. perniciosus or
P. longicuspis by the morphology of their spermathecal
ducts, all the males were at first identified as P. longicuspis.
A careful examination of the aedeagi and the finding of a
new character, the number of semi-deciduous setae on the
inner face of the coxite (Benabdennbi, 1998; Benabdennbi
et al., 1999), permitted the recognition of two distinctive
male morphs: typical P. longicuspis (LC, formerly LC B),
with each aedeagus having a straight, tapering tip (Fig. 1b),
and the coxite bearing 15–29 setae; and P. longicuspis-like
(PNA, formerly LC A), with an incurved aedeagal tip
(Fig. 1c) and 10–16 coxite setae. An isoenzyme analysis
revealed scoreable polymorphic alleles at three loci (hexo-
kinase, glucosephosphate isomerase and phosphogluco-
mutase) and unambiguously grouped the PNA males with
females of P. perniciosus (Benabdennbi et al., 1999). More
recently a RAPD study provided supporting evidence
(Martın Sanchez et al., 2000). However, the population
formed by the LC males and females, with morphology
Fig. 1. SEM images of the male aedeagi. (a) PN morphotype of
P. perniciosus; (b) LC morphotype of P. longicuspis; (c) PNA
morphotype of P. perniciosus.
26 B. Pesson et al.
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

typical of P. longicuspis, showed a deviation from
Hardy–Weinberg equilibrium in its isoenzyme genotype
frequencies, which was significant in the case of phospho-
glucomutase (Benabdennbi et al., 1999), and RAPD pat-
terns remained heterogeneous (Martın Sanchez et al., 2000).
In order to understand the reason for the genetic disequi-
librium that remained within the P. longicuspis analysed
by Benabdennbi et al. (1999), we compared individuals
captured at Chefchaouene with those from two other Rif
localities (Fig. 2), each characterized by a preponderance
of males and females with the morphology typical of one
of the two species: Taounate with 72.5% P. longicuspis, and
Ouezzane with 76.5% P. perniciosus. For this, individuals
were characterized in three ways: morphologically, by
examination of the genitalia; enzymatically, using the
three polymorphic nuclear loci that had permitted the
separation of the two species at Chefchaouene; and with
mitochondrial (mt) DNA haplotyping, by sequencing the 30
end of the Cytochrome b gene, because this marker (nor-
mally maternally inherited) had been used to relate geo-
graphical populations of many species of Larroussius in
the Mediterranean subregion, including P. perniciosus and
P. longicuspis from the countries containing the type local-
ities (Esseghir et al., 1997, 2000; Esseghir, 1998).
With the aim of placing the results from Chefchaouene in
a broader context, we also report variation in allele frequen-
cies at the three polymorphic isoenzyme loci in southern
European populations of P. perniciosus, and we compare
newly characterized mtDNA haplotypes fromMorocco and
Spain with those of the distinctive Iberian sublineage pre-
viously reported from Spain and the typical sublineage
found in Italy, Malta and Tunisia (Esseghir et al., 1997,
2000) (Fig. 2). These additional data help to establish the
stability of the diagnostic characters for P. perniciosus in all
countries except Morocco.
Materials and methods
Sandfly collections
Sandflies were captured overnight in modified miniature
CDC light traps (Madulo-Leblond, 1983) and immediately
conserved in liquid nitrogen. Most of the catches in Morocco
were made 17–25 July 1995 in three localities (Fig. 2). Two
are situated on the southern side of the Rif, at the boundary
of the sub-humid and semi-arid bioclimatic zones: to the
East, Taounate (MT samples; 500 ma.s.l.; 34�360 N,
SPAIN
EH
EG EAEM
MC
MO MT
MOROCCO
TT LC
TM
TS
TUNISIA
GZ PN
MALTA
IP
IMITALY
200 KM
Fig. 2. Geographical locations of the populations studied and type-locality countries of P. perniciosus and P. longicuspis. *¼ localities where
both P. perniciosus and P. longicuspis were collected; *¼ localities where only P. perniciosus was collected; *¼ type localities of P. perniciosus
(PN) and of P. longicuspis (LC). Key to populations (country/region/locality): EH¼Spain/Huelva/Rio Tinto and Higuera de la Sierra;
EG¼ Spain/Granada/Torvizcon, Alfacar and Loja; EA¼Spain/Almerıa/Turre; EM¼Spain/Murcia/Verdolay; IM¼ Italy/Apulia/Monte
Sant’Angelo; IP¼ Italy/Apulia/Parabita; GZ¼Malta/Gozo/Zebbug; TM¼Tunisia/Medjez Al Bab/Goubellat; TS¼Tunisia/Sidi Bouzid/
Felta; TT¼Tunisia/Beja/Tebaba; MO¼Morocco/Rif/Ouezzane; MT¼Morocco/Rif/Taounate; MC¼Morocco/Rif/Chefchaouene.
P. perniciosus complex in Morocco 27
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

4�400 W); and to the West, Ouezzane (MO samples;
350ma.s.l.; 34�520 N, 5�360 W). The latter is slightly more
humid, as is the third locality, Chefchaouene (MC samples;
600ma.s.l.; 35�120 N, 5�150 W) on the Mediterranean side of
the Rif. Locations and codes of additional material, captured
and cryopreserved in the same conditions, are given in Fig. 2.
Morphology
For each individual, the genitalia were dissected and cleared
in Marc-Andre solution (chloral hydrate/acetic acid), in readi-
ness for morphological identification by compound micro-
scopy (� 400–1000). The remainder of the insect was used
either for mtDNA haplotyping or for isoenzyme analysis, or
the head and thorax were separated for isoenzyme analysis,
leaving only the abdominal remains for haplotyping.
The structure of aedeagi (Fig. 1) were photographed from
males prepared as previously described (Pesson et al., 1994)
and observed with a stereoscan Cambridge 100 scanning elec-
tron microscope (LEO Microscopie Electronique, France).
Isoenzyme analysis
Isoelectrofocusing was carried out in ultrathin agarose gels
(PharmaciaTM
multiphor II, Amersham Biosciences, France)
with the ampholyte at pH4.0–6.5, according to the protocols
described by Pesson et al. (1991). The three discriminating
loci used were: hexokinase (HK, E.C.2.7.1.1), glucosepho-
sphate isomerase (GPI, E.C.5.3.1.9) and phosphoglucomu-
tase (PGM, E.C.5.4.2.2). The alleles, represented by coloured
bands on the gels, were numbered chronologically for species
of the subgenus Larroussius, with the more frequent allele
usually having been discovered first: for HK, allele 1¼ pHi
5.60, allele 2¼ pHi 5.49, allele 3¼ pHi 5.36, allele 4¼ pHi
5.63, allele 5¼ pHi 5.58, allele 6¼ pHi 5.33, and allele
7¼ pHi 5.22; for GPI, allele 1¼ pHi 5.30, allele 2¼ pHi
5.64, allele 3¼ pHi 5.00, allele 4¼ pHi 6.19, allele 5¼ pHi
5.09, allele 6¼ pHi 5.43, allele 7¼ pHi 5.69, allele 8¼ pHi
5.60, and allele 9¼ pHi 5.80; for PGM, allele 1¼ pHi 4.93,
allele 2¼ pHi 5.24, allele 3¼ pHi 5.10, allele 4¼ pHi 5.34,
allele 5¼ pHi 5.47, allele 6¼ pHi 5.15, allele 7¼ pHi 5.29,
and allele 8¼ pHi 5.45.
The allele frequencies, tests for deviation from Hardy–
Weinberg equilibrium, and UPGMA phenetic analysis based on
Nei’s genetic distances were all calculated using BIOSYS-2
(Swofford & Selander, 1981). In addition, GENEPOP
(Raymond & Rousset, 1995) was used to test for genotypic
linkage disequilibrium and estimate FST for each pair of
samples.
PCR amplification, sequencing and analysis of mtDNA
The protocols of Esseghir et al. (1997) were followed: to
extract genomic DNA; to amplify by the polymerase chain
reaction (PCR) the 30 end of the Cytochrome b gene (Cyt b)
using primers CB3-PDR and N1N-PDR; to purify the PCR
product by agarose-gel fractionation and binding to glass
milk (Geneclean Kit, BIO 101 Inc, U.S.A.); to sequence the
amplified fragment using each of the PCR primers and the
ABI PRISMTM
BigDye Cycle Sequencing Ready Reaction
kit (PE Applied Biosystems ¼ ABI, U.S.A.); and, to record
the DNA sequences using the ABI 373 or 377 systems and
protocols. DNA sequences were aligned using SeqEd ver-
sion 1.0.3 software (ABI) or Sequencher (Gene Codes
Corp., U.S.A.). Phylogenetic analyses were performed
with PAUP* (Swofford, 2002).
Results
Morphology
According to the aforementioned criteria, fully described by
Benabdennbi et al. (1999), the males characterized from all
countries (Spain, Italy, Malta, Tunisia and Morocco) could
be assigned to three morphotypes: typical P. perniciosus
(PN; Fig. 1a) in all countries; typical P. longicuspis (LC;
Fig. 1b) only in Tunisia and Morocco; and P. longicuspis-like
P. perniciosus (PNA; Fig. 1c) only in northernMorocco. There
were two female morphotypes: typical P. perniciosus (PN) in
all countries; and typical P. longicuspis (LC) only in Tunisia
and Morocco.
Isoenzyme analysis
Allele frequencies at the three polymorphic loci are
reported for nine populations with the PN/PNAmorphology
of P. perniciosus and for three populations with the LC
morphology of P. longicuspis (Table 1). Only some of the
isoenzyme results for the Chefchaouene (MC) flies were
published previously (Benabdennbi et al., 1999). All
P. perniciosus populations, including the PNA morphotypes,
were in Hardy–Weinberg (HW) equilibrium, and there was
no significant linkage disequilibrium except for two popula-
tions: in Ouezzane, Morocco (MO; HK-GPI, P¼ 0.010) and
in Rio Tinto, Spain (EH; GPI-PGM, P¼ 0.046). In contrast
all three populations of P. longicuspis from Morocco were
not in HW equilibrium at the PGM locus.
Nei’s genetic distances (D) and FST values were calculated
among all populations (Table 2). Nei’s D-values suggest two
groups of P. perniciosus populations, as presented graph-
ically in the UPGMA tree (Fig. 3): (1) Italy, Malta and
Morocco, where Chefchaouene (MC) is distinctive; and (2)
Iberia. The FST results indicate the same geographical struc-
ture and confirm the low gene flow observed between
Moroccan and Iberian populations (Pesson et al., 1998).
The UPGMA tree also illustrates the genetic convergence of
Moroccan P. longicuspis with the Iberian populations of
P. perniciosus, although it is necessary to caution that
P. longicuspis in Morocco does not have the genetic char-
acteristics of a single, reproductively isolated species, as
28 B. Pesson et al.
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

manifested by the HW disequilibrium at the PGM locus and
other indicators reported below.
mtDNA haplotypes and lineages
The last 279 nucleotides of Cytochrome b (Cyt b) were
sequenced for 91 individual sandflies from Morocco, a
single sandfly from Tunisia and 35 individual sandflies
from Spain, and the sequences were aligned with those
already obtained from other countries (Esseghir et al.,
1997, 2000). Unique sequences (or haplotypes) were identi-
fied (Table 3) by inspection of the input data matrix and the
distance matrix given by PAUP (Swofford, 2002). The haplo-
types fell into three distinct primary lineages (pern, lcus and
lcx), following a phylogenetic analysis based on genetic
distances (Fig. 4). Haplotypes were coded to denote their
lineage and the morphospecies in which they were usually
found: pern for P. perniciosus, lcus for P. longicuspis, and
lcx for a putative sibling species of P. longicuspis. Pairs of
haplotypes differed only by 1–8 nucleotides (0.4–2.9%)
within each of the three primary lineages, whereas the
range of pairwise differences (and absolute genetic dis-
tances) between these lineages was 12–18 nucleotides
(4.3–6.5%). In Table 4, the distributions of the newly
characterized Cyt b sequences from Morocco are related
to morphotype, sex and Cyt b lineage.
The first of the three primary lineages, pern, has six
diagnostic polymorphisms (at nucleotide positions 27, 49,
58, 168, 171 and 258), and these are fixed in all three of its
Table 1. Allelic frequencies at the three polymorphic isoenzyme loci characterized in nine populationsa of P. perniciosus and three
populations of P. longicuspis
Locusb Phlebotomus perniciosus Phlebotomus longicuspis
No. in sample Spain Italy Malta Morocco Morocco
Allele number EH EG EA EM IM GZc MO MT MCd MO MT MC
HK
No. 25 26 18 21 121 151 167 41 133 42 87 114
1 1.000 0.885 0.889 0.929 0.459 0.467 0.317 0.488 0.023 0.988 0.989 0.996
2 0 0.096 0.111 0.071 0.517 0.526 0.581 0.500 0.970 0 0 0
3 0 0.019 0 0 0 0.003 0.102 0.012 0.008 0.012 0 0.004
4 0 0 0 0 0 0 0 0 0 0 0.006 0
5 0 0 0 0 0.025 0 0 0 0 0 0 0
6 0 0 0 0 0 0.003 0 0 0 0 0 0
7 0 0 0 0 0 0 0 0 0 0 0.006 0
P – 1 1 1 1 1 0.874 0.347 1 1 1 –
GPI
No. 22 26 18 21 126 140 147 40 77 44 80 84
1 0.773 0.808 0.889 0.881 0.988 0.975 0.956 0.975 0.961 0.864 0.756 0.518
2 0.205 0.115 0.111 0.071 0.008 0.011 0.034 0.013 0.026 0.080 0.181 0.327
3 0.023 0.038 0 0.048 0 0.007 0.003 0.013 0.006 0.023 0.044 0.065
4 0 0 0 0 0 0.007 0.007 0 0 0 0 0
5 0 0 0 0 0 0 0 0 0.006 0 0.006 0.012
6 0 0.019 0 0 0 0 0 0 0 0.011 0 0
7 0 0 0 0 0 0 0 0 0 0.023 0.013 0.077
8 0 0.019 0 0 0 0 0 0 0 0 0 0
9 0 0 0 0 0.004 0 0 0 0 0 0 0
P 0.271 0.201 0.166 1 1 1 0.242 1 0.096 0.159 0.770 0.518
PGM
No. 22 26 18 21 124 132 173 39 114 49 88 100
1 0 0 0 0 0.004 0 0 0.013 0 0 0 0
2 0.977 0.981 1.000 0.976 0.980 0.939 0.908 0.923 0.697 0.571 0.756 0.440
3 0 0 0 0 0 0.011 0.064 0.026 0 0 0 0.005
4 0.023 0.019 0 0.024 0.012 0.042 0.020 0.038 0.285 0.184 0.193 0.290
5 0 0 0 0 0 0 0 0 0 0.245 0.051 0.265
6 0 0 0 0 0 0 0.009 0 0 0 0 0
7 0 0 0 0 0 0.008 0 0 0.013 0 0 0
8 0 0 0 0 0.004 0 0 0 0.004 0 0 0
P 1 1 – 1 1 0.383 0.367 1 0.658 < 0.001 < 0.001 < 0.001
aOrigins and their abbreviations are given in Fig. 1.bHK ¼ hexokinase, GPI ¼ glucose phosphate isomerase and PGM ¼ phosphoglucomutase; P ¼ probability of w2 value occurring by chance,
when testing for deviation from Hardy–Weinberg expectations of genotype frequencies.cSome data from Leger et al. (1991). dPNA is the only male morphotype recorded from this locality; some data from Benabdennbi et al. (1999).
P. perniciosus complex in Morocco 29
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37
sublineages, two of which were found in the Moroccan Rif
(Table 3; Fig. 4). Most P. perniciosus from the Rif had the
typical morphology (PN) and the Cyt b haplotype pern01,
which forms with haplotype pern02 the typical sublineage
(pnt) previously isolated only from Italy, Malta and
Tunisia. The typical sublineage of P. perniciosus had been
distinguished from the Iberian sublineage (pni) by two
polymorphisms, at nucleotide positions 147 and 246
(diagnostic) (Esseghir et al., 1997, 2000; Table 3). The
fixation of these polymorphisms in the Iberian sublineage
was confirmed by characterizing the Cyt b sequences of
37 P. perniciosus from four localities in southern Spain
(EH, EG, EA and EM in Fig. 2). Thirty-five flies contained
haplotype pern04 and two flies contained haplotype
pern05, which together form the Iberian sublineage in
Fig. 4. No haplotypes of the Iberian sublineage were found
in the Rif.
Haplotype pern01 of the typical sublineage was asso-
ciated in the Rif not only with the typical male and female
PN morphotypes of P. perniciosus (in Ouezzane and Taou-
nate) but also with the typical male and female LC mor-
photypes of P. longicuspis (in Chefchaouene) (Table 4). This
is consistent with interspecific hybridization and mtDNA
introgression from P. perniciosus into P. longicuspis.
Eleven males of P. perniciosus from Chefchaouene had
the PNA morphology and were characterized for Cyt b: all
had the haplotype pern06, as did two females of P. perni-
ciosus from Chefchaouene. One typical male (PN) of
P. perniciosus from Ouezzane had a similar haplotype,
pern07. The haplotypes pern06 and pern07 are considered
to form the previously unreported pna sublineage (Fig. 4),
which is distinguished from the other two sublineages of
P. perniciosus by three polymorphisms, at nucleotide posi-
tions 147, 183 (diagnostic) and 246 (Table 3). Haplotypes of
the pna sublineage were the only ones found in males with the
PNA genitalia, all from Chefchaouene, and they were not
found in any P. longicuspis sensu lato. However, the pna
sublineage is not diagnostic for males of P. perniciosus with
the PNA morphotype, because it was also found in two
typical male PNmorphotypes: one fromOuezzane,Morocco,
and the second from Tunisia (Table 3).
The second of the three primary Cyt b lineages, lcus, has
six diagnostic, fixed polymorphisms (at nucleotide positions
6, 9, 27, 54, 171 and 252; Table 3), and it had previously
been isolated only from Tunisian P. longicuspis (Esseghir
et al., 2000). It was also found only in typical male and
female LC morphotypes of P. longicuspis from the three
Moroccan localities (Table 4).
The third primary lineage, lcx, is reported here for the
first time, and is characterized by three fixed polymorph-
isms (at nucleotide positions 27, 123 and 171; Table 3). It
was found in the three Moroccan localities and, being asso-
ciated only with typical male and female LC morphotypes
of P. longicuspis, it is a marker for a sibling species.
Combined isoenzyme profiles and mtDNA in Moroccan
sandflies
Forty-nine individual sandflies originating from Ouez-
zane, Taounate and Chefchaouene were fully characterized,
morphologically, isoenzymically (at the HK, GPI and PGM
loci) and by mtDNA sequencing. Allele frequencies were
analysed at each isoenzyme locus for five populations
defined by their unique combinations of morphology and
mtDNA lineage (Table 5).
At the HK locus, the three groups of Moroccan P. long-
icuspis in Table 5 (all LC morphotypes, but each defined by
one of the three primary mtDNA lineages) were almost
monomorphic for allele 1, which also showed high frequen-
cies (0.885–1.000) in all other populations of Moroccan
P. longicuspis (LC morphotypes) as well as in Iberian popula-
tions of P. perniciosus (PN morphotypes) (Table 1). How-
ever, there were discretely lower frequencies of allele 1
Table 2. Nei’s genetic distances (above diagonal) and FST values (below diagonal) calculated from the isoenzyme data at the three
polymorphic loci for nine populations of P. perniciosus and three populations of P. longicuspis
Phlebotomus perniciosus Phlebotomus longicuspis
Populationsa EH EG EA EM IM GZ MO MT MCb MO MT MC
EH – 0.006 0.009 0.008 0.137 0.138 0.201 0.128 0.544 0.055 0.014 0.107
EG 0.0090 – 0.001 0.002 0.088 0.088 0.139 0.080 0.428 0.061 0.023 0.140
EA 0.0283 � 0.0167 – 0.002 0.076 0.077 0.125 0.069 0.403 0.061 0.027 0.152
EM 0.0236 � 0.0115 � 0.0174 – 0.090 0.091 0.145 0.082 0.437 0.052 0.021 0.139
IM 0.3649 0.2506 0.2385 0.2738 – 0.001 0.009 0.001 0.121 0.198 0.169 0.373
GZ 0.3306 0.2266 0.2152 0.2464 � 0.0007 – 0.008 0.000 0.114 0.191 0.164 0.360
MO 0.3623 0.2712 0.2642 0.2938 0.0280 0.0231 – 0.010 0.081 0.265 0.236 0.459
MT 0.3234 0.2035 0. 1987 0.2327 � 0.0027 � 0.0068 0.0210 – 0.123 0.177 0.152 0.342
MCb 0.6506 0.5881 0.5999 0.6160 0.3312 0.2908 0.2066 0.3120 – 0.561 0.548 0.802
MO 0.1415 0.1372 0.1483 0.1330 0.3771 0.3415 0.3741 0.2990 0.5940 – 0.022 0.051
MT 0.0371 0.0551 0.0723 0.0580 0.3476 0.3173 0.3597 0.2833 0.5820 0.0439 – 0.055
MC 0. 1752 0.1982 0.2184 0.2110 0.4368 0.4084 0.4301 0.3480 0.5531 0.0772 0.0952 –
aKey to populations is given in Fig. 2.bPNA is the only male morphotype recorded from this location.
30 B. Pesson et al.
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

(0.000–0.488) in Italian, Maltese and Moroccan popula-
tions of P. perniciosus (PN/PNA morphotypes) (Tables 1
and 5). In contrast, populations of P. perniciosus from the
last three countries were characterized by a high frequency
of allele 2 (0.500–1.000), which was fixed in the Chef-
chaouene population defined by the pna mtDNA subline-
age (Table 5) but absent from all P. longicuspis (Tables 1
and 5).
The GPI locus showed differential polymorphism among
the three primary mtDNA lineages found in P. longicuspis
in Morocco (Table 5). This locus was almost monomorphic
(0.944–1.000) for allele 1 in P. perniciosus (PN/PNA mor-
photypes) from Italy, Malta and northern Morocco, but it
was less frequent in populations of P. perniciosus from
southern Spain (0.773–0.889) and in all Moroccan popula-
tions of P. longicuspis (0.214–0.864) (Tables 1 and 5).
The PGM locus is highly polymorphic in Phlebotomus
(Larroussius) species, and in Chefchaouene it clearly differ-
entiated flies of the lcx mtDNA lineage from all other males
and females with the typical LC morphotype (Table 5).
Allele 5 was found only in the lcx mtDNA lineage, in
which allele 2 was almost absent. All Moroccan P. long-
icuspis populations were not in HW equilibrium when all
individuals were treated as belonging to one species
(Table 1), and this is consistent with the presence of a sibling
species. The frequency of allele 4 was elevated only in the
subpopulation of P. longicuspis with the lcx mtDNA lineage
and in P. perniciosus from Chefchaouene with the pna
mtDNA sublineage (Table 5). This is another characteristic
distinguishing the Chefchaouene population of P. pernicio-
sus from all others (Table 1).
Therefore, based on the PGM locus, mtDNA introgres-
sion is most likely to have occurred by hybridization
between P. perniciosus and P. longicuspis sensu stricto,
not the sibling species. However, the hybrids do have a
uniquely high frequency of allele 2 at the GPI locus
(Table 5).
It is noteworthy that there are no fixed alleles diagnostic
for either P. perniciosus or P. longicuspis from Morocco,
even after removing the sibling species (LC morphotypes,
lcx mtDNA lineage), the P. perniciosus variant (PNA/PN
morphotypes, pna mtDNA sublineage) and the putative
hybrids (LC morphotypes, pern01 haplotype of the pnt
mtDNA sublineage). When present, however, allele 2 of
HK was diagnostic for P. perniciosus and allele 2 of GPI
was diagnostic for P. longicuspis sensu lato (Table 5).
Discussion
Three species of the P. perniciosus complex in the Moroccan
Rif
The current findings indicate the presence of three phy-
logenetic species in the Moroccan Rif. One species is
P. perniciosus, which was characterized by a distinctive iso-
enzyme profile and a discrete Cyt b mtDNA lineage (com-
posed of two sublineages, typical pnt, and pna). This species
includes not only the typical male and female morphotypes
(PN) of P. perniciosus but also a male morphotype (PNA)
that can be confused with P. longicuspis (Benabdennbi et al.,
1999). The pna mtDNA sublineage is not a diagnostic
marker for a sibling species with PNA male morphotypes,
because it was also found in two typical (PN) males of
P. perniciosus fromOuezzane, Morocco, and northern Tunisia
(Tables 3 and 4). However, all the PNA males in
Distance.40 .33 .27 .20 .13 .07 .00
GZ P. perniciosus
MT P. perniciosus
IM P. perniciosus
MO P. perniciosusMC P. perniciosusEH P. perniciosus
EM P. perniciosus
EA P. perniciosus
EG P. perniciosus
MT P. longicuspisMO P. longicuspis
MC P. longicuspis
Fig. 3. UPGMA cluster analysis of the Nei’s genetic distances among nine populations of P. perniciosus and three populations of P. longicuspis,
calculated from allele frequencies at the three polymorphic isoenzyme loci (HK, GPI, PGM).
P. perniciosus complex in Morocco 31
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

Table
3.AlignmentofmitochondrialCytbhaplotypes
(variantcharactersonly)relatedto
thespecim
ens’origins
Character
no.in
alignmenta
No.ofspecim
enswitheach
haplotypein
differentlocalities
b
Cytb
11111111112222
124555777702344667880455
Spain
Italy
Malta
Tunisia
Morocco
Haplotype
69579148235853357581131628
EH
EG
EA
EM
IPGZ
TT
TM
TS
MO
MT
MC
pern01
TAAATATTTATCTTGACATCGAAATG
12
22
18
19
pern02
TAAATAATTATCTTGACATCGAAATG
1
pern04
TAAATATTTATCTTGATATCGAATTG
14
96
4þ2
pern05
TAAATATTTATTTTGATATCGAATTG
2
pern06
TAAATATTTATCTTGACGTCGCATTG
113
pern07
TAAATATTTATCTTGACATCGCATTG
1
lcus01
CGATCACCCATTTTGGTACTGCATCA
42
18
2
lcus02
CGATCACCCATTTTGGTACTGCATCA
1
lcx01
TAGCCATCCACTTCGACACAGAATTA
21
lcx02
TAGCCATCCACTTCGACACAGAATTA
1
lcx03
TAGCCATCCACTTCGACACAGAATTA
1
lcx04
TAGCCATCCACTTCGACACAGAATTA
4
lcx05
TAGCCATCCACTTCGACACAGAATTA
5
lcx06
TAGCCATCCACTTCGACACAGAATTA
2
lcx07
TAGCCATCCACTTCGACACAGAATTA
6
lcx08
TAGCCATCCACTTCGACACAGAATTA
1
lcx09
TAGCCATCCACTTCGACACAGAATTA
1
lcx10
TAGCCATCCACTTCGACACAGAATTA
1
lcx11
TAGCCATCCACTTCGACACAGAATTA
1
Totals
16
96
62
22
34
25
21
45
aUnderlined
nucleotides
¼synapomorphiccharactersdiagnosticforeach
ofthethreemain
lineages
(pern,lcus,lcx),i.e.fixed
polymorphisms.bUnderlined
numbers¼
data
from
Esseghir
etal.(1997,2000).
32 B. Pesson et al.
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

Chefchaouene did have the pna mtDNA sublineage as well as
a characteristic isoenzyme profile (including a high frequency
of allele 4 of PGM), and so there is a suggestion that the
PNA aedeagus might be associated with an imperfect bar-
rier to panmixia and incipient speciation. Direct evidence
for the absence of linkage between mtDNA lineage and
male morphology was provided by finding both PN and
PNA males amongst the laboratory-reared progeny of a
single female from El Jebha, in the Moroccan Rif
(Benabdennbi, 1998). Throughout most of its range outside
Morocco, the patterns of genetic variation of P. perniciosus are
consistent with the isolation of populations in two commonly
proposed Pleistocene Ice-Age refugia (Avise & Walker, 1998).
The isoenzyme analysis (Fig. 3) indicated an Iberian cluster of
populations that is distinct from an Italy–North Africa cluster
(including the type locality in Malta), and the distributions of
mtDNA lineages matched this (Fig. 4; Esseghir et al., 1997,
2000).
The second phylogenetic species is P. longicuspis, which
was also characterized by a distinctive isoenzyme profile
Icus01 Africa LC
Icus02 Rif LC
100
0.005 substitution/sitepern05 Spain PN
pern04 Spain PN
pern07 Rif PNA
pern06 Africa PNA
pern02 Italy PN
pern01 Most PN
79
77
53
99
61
92
5751
Icx09 Rif LC
Icx11 Rif LC
Icx08 Rif LC
Icx05 Rif LC
Icx04 Rif LC
Icx06 Rif LC
Icx10 Rif LC
Icx07 Rif LC
Icx03 Rif LC
Icx02 Rif LC
Icx01 Rif LC
Fig. 4. Phylogenetic relationships among all the haplotypes of mitochondrial Cyt b. Each haplotype is coded by lineage (pern, lcus, lcx) and
number, and it is also labelled with the country or region and the morphotype (PN, PNA, LC) in which it has been found. The tree was
generated by a Neighbour joining search of genetic distances (with ties broken randomly), rooted with the lcus haplotypes as outgroups (PAUP:
Swofford, 2002). Values below some branches are bootstrap support values (100 replicates) given by the 50% majority-rule consensus tree.
A maximum parsimony analysis, using an heuristic search (PAUP: Swofford, 2002), gave a congruent 50% majority-rule consensus tree.
P. perniciosus complex in Morocco 33
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

and a discrete mtDNA lineage (lcus). Genotype frequencies
were in HW equilibrium at all three isoenzyme loci if
specimens with the lcx mtDNA lineage were removed
from the population and treated as a third, sibling species.
Thus, mtDNA characterization permitted the separation of
P. longicuspis sensu stricto from males and females that
shared its known diagnostic morphological characters (LC
morphotypes). Benabdennbi et al. (1999) found a departure
from HW expectations at the PGM locus for P. longicuspis
caught in Chefchaouene and therefore they suspected the
presence of a subpopulation, but they had no independent
marker for separating the two. The males of P. perniciosus
(PN, PNA) and P. longicuspis (LC) were shown to have a
significantly different number of coxite hairs (Benabdennbi
& Pesson, 1998). We can now re-analyse this data, using an
analysis of variance with Scheffe test, to show that the mean
number (m) of coxite hairs in males with the mtDNA line-
age lcx (m¼ 20.3; range: 19–22; n¼ 14) is significantly less
than that in males of P. longicuspis sensu stricto with the
lineage lcus (m¼ 26.4; range: 21–31; n¼ 26).
The relatively large pairwise genetic distances (4.3–6.5%)
between the primary mtDNA lineage characteristic of
P. perniciosus (pern) and the two found only in P. longicuspis
(lcus, lcx) indicate a divergence of these three lineages about
2–3 million years ago (mya), based on a pairwise divergence
rate of 2.3% per million years (Esseghir et al., 1997, 2000).
The divergence of these mtDNA lineages may have accom-
panied allopatric speciation. However, it should be remem-
bered that divergent mtDNA lineages may arise before
speciation, because of maternal inheritance and the absence
of recombination (Avise & Walker, 1998). Therefore, the
presence in P. longicuspis of two highly divergent mtDNA
lineages does not in itself provide strong support for recog-
nizing a sibling species. The necessary, complementary sup-
port for sibling species recognition comes from the coxite
hair counts and especially from the isoenzyme analysis. This
strongly suggests that P. longicuspis sensu stricto and its
sibling species are not just phylogenetic species (character-
ized by discrete isoenzyme profiles and mtDNA lineages) but
are also biological species, with a substantial reproductive
barrier between them. The population genetics analysis,
based on isoenzymes, also indicates that P. perniciosus is a
good biological species.
The discovery of some mtDNA introgression from
P. perniciosus to P. longicuspis does not necessarily invalidate
these conclusions. However, it should be remembered that
even occasional hybridization could lead to the sharing of
epidemiologically important traits, as discussed in the next
section.
Evidence for mitochondrial introgression from P. perniciosusto P. longicuspis and its epidemiological significance
Esseghir et al. (2000) compared mtDNA (Cyt b) and
nuclear gene (EF-a) phylogenies of Larroussius species: thetwo phylogenies were incongruent for the P. perniciosus
complex, and mtDNA introgression between P. perniciosus
and P. longicuspis was suspected. The current findings pro-
vide further, compelling evidence for mtDNA introgression
between wild populations of these two species. In Chef-
chaouene, in the Moroccan Rif, some P. longicuspis mor-
photypes (LC) contained a mtDNA haplotype (pern01) of a
sublineage (pnt) that is usually found only in P. perniciosus
from Morocco, Tunisia, Malta and Italy. The presence of
the same pern01 haplotype in the two species indicates
either that introgression is still occurring or that it has
occurred within the last 125 000 years, assuming the molecu-
lar clock runs no faster than 2.3% pairwise divergence per
million years (Esseghir et al., 2000) and that the sequencing
error is not greater than 1 in 300 nucleotides. Mitochondrial
DNA is usually inherited maternally and the putative
hybrids were morphologically inseparable from, and iso-
enzymatically most similar to, P. longicuspis. Therefore,
Table 4. The number of individual sandflies with each combination of morphotype and Cyt b lineage in three Moroccan localities
Locality
Morphotypes
Cyt b
lineage/sublineageaOuezzane
(MO)
Taounate
(MT)
Chefchaouene
(MC)
LC female lcus/– 1 9 2
lcx/– 3 0 14
pern/pnt 0 0 3
LC male lcus/– 1 10 0
lcx/– 1 1 7
pern/pnt 0 0 6
PN female pern/pnt 9 0 0
pern/pna 0 0 2
PN male pern/pnt 9 1 NFb
pern/pna 1 0 NF
PNA male pern/pna NF NF 11
Total 25 21 45
aOnly the pern lineage has designated sublineages.bNF ¼ morphotype not found in the locality.
34 B. Pesson et al.
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

we conclude that fertile progeny usually result only from
matings between males of P. longicuspis sensu stricto and
the females of P. perniciosus or hybrids.
A comparative sequence analysis of Cyt b also demon-
strated mtDNA introgression between closely related spe-
cies within two subgenera of Lutzomyia (Marcondes et al.,
1997; Testa et al., 2002). Like the P. perniciosus complex,
both of these neotropical sandfly taxa contain many vectors
of Leishmania species, and so it is important to consider the
epidemiological implications of mtDNA introgression in
sandflies.
Mitochondrial introgression usually results from inter-
breeding. Such interspecific hybridization is not uncommon
among related insect species (Arnold, 1997), and the inde-
pendent assortment of many nuclear genes is axiomatic in
Mendelian genetics. Therefore, it should come as no sur-
prise if recently diverged species, such as P. perniciosus and
P. longicuspis, are shown to display reticulate evolution. In
these circumstances, genotypic markers as well as pheno-
typic traits and morphology will not always covary, partly
as a result of recent ancestry (with insufficient time for ‘line-
age sorting’) and partly from continuing hybridization. This
absence of character covariation is not helpful to parasitolo-
gists and applied entomologists who hope that vectorial
traits can be associated with easily identifiable taxonomic
species, which can then be targeted for control measures.
For example, much effort was made to find diagnostic
markers for the major sub-Saharan vectors of malaria
(Hill & Crampton, 1994), even though sibling species of
the Anopheles gambiae Giles complex (Culicidae) are known
to hybridize both in the wild and the laboratory (Black &
Lanzaro, 2001), and mtDNA introgression is great enough to
prevent the separation into two distinct lineages of the most
important vectors,An. gambiae sensu stricto andAn. arabiensis
Patton (Besansky et al., 1994).
Given the evidence for the occurrence of interspecific
introgression and sibling species in the P. perniciosus
complex, it should no longer be assumed that the frequen-
cies of certain vectorial traits can be directly related to
the proportions of the morphospecies previously recorded
in different geographical regions and ecological zones of
north-west Africa. For example, Rioux et al. (1984)
reported that P. longicuspis is more abundant than P.
perniciosus in the drier bioclimates of Morocco, but only
a re-examination of slide-mounted males in various
collections would indicate whether authors had confused
P. longicuspis sensu lato (morphotype LC) with P. perniciosus
(morphotype PNA). Even then, the identification of the
male morphotype LC would still leave unresolved any
ecological replacement of P. longicuspis sensu stricto by
its sibling species. Both P. perniciosus and P. longicuspis have
been found naturally infected with the same strain of Leish-
mania infantum (Esseghir et al., 2000), and now reticulate
evolution makes it less certain that ecological associa-
tions or vectorial roles are going to be linked solely to one
taxon.
Table 5. Allelic frequencies at the three polymorphic isoenzyme loci for Moroccan P. perniciosus and P. longicuspis grouped according to
morphotype (PNþPNA and LC, respectively), mitochondrial Cyt b lineage and localities
Morphotype PN PNA + PN LC LC LC
Cyt b lineage/sublineage pern/pnt pern/pna lcus/- pern/pnt lcx/-
Localities Ouezzane (MO) Chefchaouene (MC) Taounate (MT) Chefchaouene (MC) Chefchaouene (MC)
No. tested (Female þ male) 4þ 4 1þ 8 5þ 5 2þ 5 9þ 6
Locusa and alleles
HK
1 0.375 0 0.950 1.000 1.000
2 0.500 1.000 0 0 0
3 0.125 0 0 0 0
7 0 0 0.050 0 0
P 1 – 1 – –
GPI
1 1.000 0.944 0.750 0.214 0.767
2 0 0 0.200 0.786 0.067
3 0 0 0.050 0 0.067
5 0 0.056 0 0 0.033
7 0 0 0 0 0.067
P – 1 0.822 1 1
PGM
2 0.938 0.667 0.950 0.929 0.067
3 0.063 0 0 0.071 0
4 0 0.333 0.050 0 0.333
5 0 0 0 0 0.600
P 1 0.457 1 1 1
aHK ¼ hexokinase, GPI ¼ glucosephosphate isomerase, and PGM ¼ phosphoglucomutase; P ¼ probability of w2 value occurring by chance,
when testing for deviation from Hardy–Weinberg expectations of genotype frequencies.
P. perniciosus complex in Morocco 35
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

Taxonomic significance of the findings: formal recognition of
the new sibling species?
Those researching leishmaniasis transmission need to be
aware of what information is being communicated by dif-
ferent taxonomic names. Two morphospecies, each with a
Linnean binomial, may be good phylogenetic species and,
despite occasional hybridization, may even behave most of
the time as good biological species (Arnold, 1997). In these
circumstances, it is usual to maintain the taxonomic species
concept, with identifications being based on the morph-
ology of type specimens (Otte & Endler, 1989), not least
because few laboratories have the resources for routine
genetic characterization.
Until more populations have been characterized, and
gene flow between them assessed, we do not favour creating
or synonymizing taxonomic names in the P. perniciosus
complex. Certainly, where there is reticulate evolution, the
reliance on a few characters might well mislead, as is clear
from the history of the complex. Parrot (1936a) reared
in the laboratory a small number of ‘pure strains’ of
P. perniciosus and P. langeroni var. longicuspis and, based
on this evidence for the inheritance of distinctive male
genitalia, raised the latter to full species. Soon afterwards,
however, Parrot (1936b) recorded much individual varia-
tion in the form of the aedeagus of P. perniciosus, but
insisted that the terminal bifurcation of the aedeagus was
diagnostic. Other authors accepted this, along with the later
conclusion of Parrot & Durand-Delacre (1947) that the
morphology of the aedeagus of males collected in the
Sahara (Beni Ounif-de-Figuig and Tamanrasset, Algeria)
was a variant of P. longicuspis. This variant could be the
PNA morphotype, and if so Parrot & Durand-Delacre
(1947) should have identified it as P. perniciosus. Reticulate
evolution within the P. perniciosus complex means that all
its species are unlikely to be identified by unique morpho-
logical characters or vectorial roles.
The results from Chefchaouene, in the Rif, are important
because they show that some sympatric populations of
P. perniciosus and P. longicuspis have the characteristics of
biological species. However, more extensive characteriza-
tion of populations throughout the ranges of these two
morphospecies could indicate that they would be better
treated as three informal incipient species (equivalent to
P. perniciosus, P. longicuspis and the sibling species), with
only a single taxon, P. perniciosus, being formally recog-
nized. This would be appropriate only if the results from
Chefchaouene were shown to be atypical, by demonstrating
ongoing and significant nuclear gene flow between many
sympatric populations of the three species.
Acknowledgements
The authors thank the following colleagues for help with
collecting sandflies: R and M. Killick-Kendrick (Ascot),
N. Leger and H. Ferte (Reims), G. Madulo-Leblond
(Paris), E.Martınez Ortega (Murcia) andM. C. SanchisMarin
(Almerıa). J. Llewellyn-Hughes, W. Edge and J. M. Testa
provided invaluable assistance for the mtDNA sequencing in
theNaturalHistoryMuseum,whereB.P. spentasabbatical.We
acknowledge the support of theMinistere de la Sante Publique
du Maroc (J. Mahjour), Ministere des Affaires Etrangeres
(France), the British Council (Tunis), the EC DG-XII
(Brussels; STD3 contract: TS3*CT93-0253), and the TDR
programme (UNDP/World Bank/W.H.O.).
References
Arnold, M.L. (1997) Natural Hybridization and Evolution. Oxford
University Press, Oxford.
Avise, J.C. & Walker, D. (1998) Pleistocene phylogeographic
effects on avian populations and the speciation process.
Proceedings of the Royal Society of London B, 265, 457–463.
Benabdennbi, I. (1998) Contribution a l’etude de la biodiversite chez
les Larroussius (Diptera: Psychodidae) du bassin mediterraneen.
Apports de l’analyse isoenzymatique. These de Doctorat,
Universite Louis Pasteur – Strasbourg I, France.
Benabdennbi, I. & Pesson, B. (1998) A propos de la variabilite
morphologique de Phlebotomus perniciosus (Diptera: Psychodi-
dae). Bulletin de la Societe Francaise de Parasitologie, 16, 43–50.
Benabdennbi, I., Pesson, B., Cadi-Soussi, M. & Morillas Marquez,
F. (1999) Morphological and isoenzymatic differentiation of
sympatric populations of Phlebotomus perniciosus and Phleboto-
mus longicuspis (Diptera: Psychodidae) in northern Morocco.
Journal of Medical Entomology, 36, 116–120.
Besansky, N.J., Powell, J.R., Caccone, A., Hamm, D.M.,
Scott, J.A., Collins, F.H. (1994) Molecular phylogeny of the
Anopheles gambiae complex suggests genetic introgression
between principal malaria vectors. Proceedings of the National
Academy of Science USA, 91, 6885–6888.
Black, W.C.I.V. & Lanzaro, G.C. (2001) Distribution of genetic
variation among chromosomal forms of Anopheles gambiae s.s.
introgressive hybridization, adaptive inversions, or recent
reproductive isolation? Insect Molecular Biology, 10, 3–7.
Collantes, F. & Martınez Ortega, E. (1997) Sobre la validez
taxonomica de Phlebotomus longicuspis Nitzulescu, 1931
(Diptera: Psychodidae). Boletin de la Asociacion Espanola
de Entomologia, 21, 141–146.
Esseghir, S. (1998) Phylogenie des phlebotomes mediterraneens
vecteurs de Leishmania major et de Leishmania infantum et
marqueurs moleculaires de populations. These de Doctorat,
Faculte des Sciences, Universite de Tunis, Tunisia.
Esseghir, S., Ready, P.D. & Ben-Ismail, R. (2000) Speciation of
Phlebotomus sandflies of the subgenus Larroussius coincided with
the late Miocene-Pliocene aridification of the Mediterranean sub-
region. Biological Journal of the Linnean Society, 70, 189–219.
Esseghir, S., Ready, P.D., Killick-Kendrick, R. & Ben-Ismail, R.
(1997) Mitochondrial haplotypes and phylogeography of Phle-
botomus vectors of Leishmania major. Insect Molecular Biology,
6, 211–225.
Hill, S.M. & Crampton, J.M. (1994) Review: DNA-based methods
for the identification of insect vectors. Annals of Tropical
Medicine and Parasitology, 88, 227–250.
Killick-Kendrick, R. (1990) Phlebotomine vectors of the leishma-
niases: a review. Medical and Veterinary Entomology, 4, 1–24.
36 B. Pesson et al.
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37

Leger, N., Marchais, R., Madulo-Leblond, G., Pesson, B.,
Kristensen, A., Ferte, H., Killick-Kendrick, R. & Killick-
Kendrick, M. (1991) Les phlebotomes implique dans la transmis-
sion des leishmanioses dans l’ıle de Gozo (Malte). Annales de
Parasitologie Humaine et Comparee, 66, 33–41.
Leger, N., Pesson, B., Madulo-Leblond, G. & Abonnenc, E. (1983)
Sur la differentiation des femelles du sous-genre Larroussius
Nitzulescu, 1931 (Diptera, Phlebotomidae) de la region mediterra-
neene.Annales de Parasitologie Humaine et Comparee, 58, 611–623.
Madulo-Leblond, G. (1983) Les Phlebotomes (Diptera, Phleboto-
midae) des Iles Ioniennes. These de Doctorat es Sciences
Pharmaceutiques, Universite de Reims, France.
Marcondes, C.B., Day, J.C. & Ready, P.D. (1997) Introgression
between Lutzomyia intermedia and both Lu. neivai and Lu.
whitmani, and their roles as vectors of Leishmania braziliensis.
Transactions of the Royal Society of Tropical Medicine and
Hygiene, 91, 725–726.
Martın Sanchez, J., Gramiccia, M., Pesson, B. & Morillas
Marquez, F. (2000) Genetic polymorphism in sympatric species
of the genus Phlebotomus, with special reference to Phlebotomus
perniciosus and Phlebotomus longicuspis (Diptera, Phlebotomi-
dae). Parasite, 7, 247–254.
Martınez Ortega, E., Ward, R.D., Martin Luengo, F. & Conesa
Gallego, E. (1982) Nueva distribucion de Phlebotomus (Lar-
roussius) longicuspis Nitzulescu 1930 (Diptera, Phlebotomidae)
en Espana. Revista Iberica de Parasitologia, 42, 283–288.
Morillas Marquez, F., Guevara Benitez, D.C., Gil Collado, J. &
Ubeda Ontiveros, J.M. (1982) Presencia en Espana de Phlebo-
tomus (Larroussius) longicuspis Nitzulescu, 1930. Revista Iberica
de Parasitologia, Volume extra, 191–196.
Morillas Marquez, F., Sanchis Marin, M.C., Martın Sanchez, J. &
Acedo Sanchez, C. (1991) On Phlebotomus perniciosus News-
tead, 1911 (Diptera, Phlebotominae) in the province of Almeria
in south eastern Spain. Parassitologia, 33 (Suppl. 1), 437–444.
Newstead, R. (1911) The papataci flies (Phlebotomus) of the
Maltese Islands. Bulletin of Entomological Research, 2, 47–48.
Nitzulescu, V. (1930) Phlebotomus langeroni n. sp. et P. langeroni
var. longicuspis n. var. de Douar-Shott (Tunisie). Annales de
Parasitologie, 8, 547–553.
Otte, D. & Endler, J.A. (1989) Speciation and its Consequences.
Sinauer Associates Inc, Sunderland, Mass, U.S.A.
Parrot, L. (1936a) Notes sur les phlebotomes XX. – sur
Phlebotomus langeroni var. longicuspisNitzulescu, 1930. Archives
de l’Institut Pasteur d’Algerie, 14, 137–143.
Parrot, L. (1936b) Notes sur les phlebotomes XXI. – sur la valve
copulatrice de Phlebotomus perniciosus Newstead. Archives de
l’Institut Pasteur d’Algerie, 14, 144–147.
Parrot, L. & Durand-Delacre, R. (1947) Notes sur les phlebotomes
LVI. – Phlebotomes de Beni Ounif-de-Figuig. Archives de
l’Institut Pasteur d’Algerie, 25, 77–86.
Pesson, B., Benabdennbi, I., Schuh, C., Alves Pires, C.,
Morillas Marquez, F., Sanchis Marin, M.C., Martinez Ortega, E.,
Lucientes Curdi, J. & Gallego, M. (1998) Genetic variability
of Phlebotomus perniciosus (Diptera: Psychodidae) in the Iberian
peninsula. Acta Parasitologica Portuguesa, 5, 52.
Pesson, B., Madulo-Leblond, G., Killick-Kendrick, M., Tang, Y. &
Killick-Kendrick, R. (1994) The armature in the genital atrium
as a new taxonomic character distinguishing females of
Phlebotomus papatasi and P. duboscqi (Diptera: Psychodidae).
Annals of Tropical Medicine and Parasitology, 88, 539–542.
Pesson, B., Wallon, M., Floer, M.T. & Kristensen, A.R. (1991)
Etude isoenzymatique de populations mediterraneennes de
phlebotomes du sous-genre Larroussius. Parassitologia, 33
(Suppl. 1), 471–476.
Raymond, M. & Rousset, F. (1995) GENEPOP Version 1.2.:
population genetics software for exact test and ecumenicism.
Journal of Heredity, 86, 248–249.
Rioux, J.A. & Lanotte, G. (1990) Leishmania infantum as a cause of
cutaneous leishmaniasis. Transactions of the Royal Society of
Tropical Medicine and Hygiene, 84, 898.
Rioux, J.A., Rispail, P., Lanotte, G. & Lepart, J. (1984) Relations
Phlebotomes – bioclimats en ecologie des leishmanioses.
Corollaires epidemiologiques. L’exemple du Maroc. Bulletin de
la Societe Botanique de France, 131, 549–557.
Seccombe, A.K., Ready, P.D. & Huddleston, L.M. (1993) A
catalogue of Old World phlebotomine sandflies (Diptera:
Psychodidae, Phlebotominae). Occasional Papers on Systematic
Entomology, 8, 1–57.
Swofford, D.L. (2002) PAUP. Phylogenetic Analysis Using Parsi-
mony, version PAUP*4.0. Smithsonian Institution Press,
Washington DC, U.S.A.
Swofford, D.L. & Selander, R.B. (1981) BIOSYS-1: a FORTRAN
program for the comprehensive analysis of electrophoretic data
in population genetics and systematics. Journal of Heredity, 72,
281–283.
Testa, J.M., Montoya-Lerma, J., Cadena, H., Oviedo, M. &
Ready, P.D. (2002) Molecular identification of vectors of
Leishmania in Colombia: mitochondrial introgression in the
Lutzomyia townsendi series. Acta Tropica, 84, 205–218.
Accepted 10 October 2003
P. perniciosus complex in Morocco 37
# 2004 The Royal Entomological Society, Medical and Veterinary Entomology, 18, 25–37





























![Ecology of phlebotomine sandflies and putative reservoir ... · San Carlos, state of Tamaulipas. Several years later, Velasco-Castrejón et al. [88] reported six additional cases](https://img.pdfslide.us/doc/110x75/5f9669209c795f3a6a6c7618/ecology-of-phlebotomine-sandflies-and-putative-reservoir-san-carlos-state-of.jpg)