Embed Size (px)
Citation preview

~ AND E. DAVID MORGAN
'EetES
Reference
8Jochemical SystemauC$ 8fId Ecology, Vol. 16. No. 7/8. pp.65!'r659. 1988.Printed in Great Britain.
0305-1978/88 $3.00+0,00Q 1988 Pergamon Press pic.
"K
51415
1514
10
10161010
10§
§
§
1515
Biochemical Identification of Phlebotomus (Larroussius) pediferandPhlebotomous (Larroussius) elegonensis
LUCIE M. ROGO, C. P. M. KHAMALA* and M. J. MUTINGAInternational Centre of Insect Physiology and Ecology. P.O. Box 30m. Nairobi. Kenya;
*Depal1ment of Zoology. University of Nairobi. P.O. Box 30197. Nairobi. Kenya
Kev Wont Index-Phlebotomus pedifer; P. elegonensis; electrophoresis; thin-layer starch gel electrophoresis: isoelectric focusing.
Abstract-Females of the vector of Leishmania aerhiopica, Phlebotomus pedifer. have previously been morphologicallyindistinguishable from the non-vector. P. elegonensis. The present studies have biochemically separated these two species.Differences were observed in the mobility of four enzymes. GPI. HK, leo and PGM on thin.Jayer starch gel electrophoresis.Using the isoelectric focusing technique. the two species could be differentiated by one enzyme. GPI.
and Tyler. R. C, (19m
(1985) Adv. Insect Ph.,..
178) Arthropod venoms'""BCoIogy (Bettini. 5..I.
981) in Chemistry and3nd Rembold, H., eds).
P. J. and Morgan. E. D.)9.
is) J. Insect Physiol. 14.
'.28.136.I Morgan. E. D. (1988)
is. Fac. Sci., U. Aix-
'1J. Entomo/. lB. 87.'v!organ, E. D. J. Chern.
itfield. F. B. (19681 Aust
Naturl. 4OC. 665.'Jfrysio£ ZoOt 44. n.1) Ent. Scand. 3. 225.;hannon. J. S. (1973) J.
Biochem. Syst Eco£ 8.
Bestmann. H. J. and1.17.219.. (1972) J. Chromatogr.
IntroductionDipteran sibling species, comprising morphologically indistinguishable populations that arereproductively isolated, are usually very easilydistinguished by many biochemical differencesRS in Anopheles [1, 2], Aedes [3, 4] andSimulium [5]. In studies on the subfamilyPhlebotominae, biochemical methods have alsobeen applied [6-8].
The present studies set out to use biochemical techniques of thin-layer starch gelelectrophoresis and isoelectric focusing onagarose to differentiate Phlebotomus pedifer, avector of Leishmania aethiop;ca (9), from P.elegonensis (non-vector) whose females aremorphologically identical. Previously the onlyclue to their identity has been small differencesin their male terminalia and the females couldonly be differentiated by rearing them individually in isolation or by collecting them in copula.Since the females of P. pedifer are involved indisease transmission, their identity should beverified.
Results and DiscussionOf the 14 enzymes assayed in thin-layer starchgel electrophoresis, nine; glucose phosphateisomerase (GPI), hexokinase (HK), isocitricdehydrogenase (ICD), malate dehydrogenase(MDH), malic enzyme (ME), mannose phosphate
(Received8 February 19881
isomerase (MPI), phosphoglucomutase (PGM)and L-threonine-3-dehydrogenase (TDH) couldbe detected in individual sandflies with varyingdegrees of intensity. The profiles of six of theseenzymes which were fairly distinct and reproducible are presented (Fig. 1). The frequency ofsuccessful identification is shown in Table 1.Phosphoglucomutase is not presented in Fig. 1because it had two or three different alleliccombinations in both P. pedifer and P. elegonensis during every run. These alleles expressedthemselves in different combinations in each ofthese species at every run. In each PGM run, themost common allele of P. eJegonensis alwayshad a lower mobility than the most commonallele of P. pedifer. TDH was too faint and MPI tooinconsistent and are therefore not presented.Aconitate hydratase (ACON), alcohol dehydrogenase (AOH), glucose-6-phosphate dehydrogenase (G-6-PO), nucleosidase hydralase (NH)and superoxide dismutase (500) could not bedetected in individual sandflies. The enzymesextracted from a male or female of each specieshad identical mobilities.
It is evident that GPI, HK, ICD and PGM candifferentiate P. pedifer from P. eJegonensis. Foreach of these enzymes P. eJegonensis had alower mobility than P. pedifer.
Using isoelectric focusing on agarose, onlyfive of the 14 enzymes assayed could bedetected in individual sandflies. These were GPI,HK, MOH, ME and PGM. With this m~thod. onlyGPI differentiated P. pedifer from P. elegonensis.
6S5

The numerical values designated for each electromorph are arbi1rary. The lowest anodic electromorph for the control species IS. shwetZll isdesignated 100. subsequent etectromorphs are designated 101. 102, 103 etc. Sim~arty the lowest cathodic efec:tromorph lor the control wasdesignated -100. Electromorphs lor each specios being studied are relatiw to thet 01 the control species IS. shwelZlllreler also to Fig. 11.
Enzyme electromotph designation in p818l1\hese$
Number 01 specimens Number and frequency
Enzyme S.shwetzi P.pediIIY P. e/egonensis 01 trials S.shwetzi p.pediIIY P. elegonensis
ACON 6 16 18 4 NU NU Nil
ADH 4 19 16 4 NU Nil Nil
~ 4 8 II 3 Nil N~ Nil
GPI 10 39 '17 10 (100)"" 1100.51'" (100)'.00
(101)'"
(102)'"
HK 10 29 26 5 (1001'" (102)'00 1101)'"
ICO 5 17 16 8 (100)'''' (102)'''' (101)'.00
MOH 6 19 18 9 (1001~'" (101)'.00 1101)''''(-100)'" 1-100)'''' (-99)·"
ME 5 22 19 7 1100)'.00 (100)"" (100)'.00
MPI 1 5 5 3 l100)lUD (1001"" (1001~'"
(101)1UIl (101)'-'" (101)''''(102)·... (102)'''' 11021·...
NH, 2 8 9 2 N~ NU N~
PGM 8 25 27 8 (1001'" (951.... (90)....
(10".... (98)'·" (95)'"(102)·... (100)'" (100I~"
(101)'-" (101)""
PGK 4 10 9 3 (100)'00 (1011''' (101)'00
SOD 2 10 9 2 NU NU NilTOH 7 19 18 4 (100)'.00 1100)'" "001''''
.,f
.i
Ij{
lUOE M. ROGO, C. P. M. KHAMAlA AND M. J. Ml1TINGA
---------... ~
~ P-!IJl!C!I.... A-P.~.. iSII
_"'-- 1lD"",,,*,," IlulllrlW. _F_. [J\8y lD'Il
~~ '1 ME,:-- ,_usPA' SPA"
P. pedffer had only four distinct bands whereas P.elegonensis had six distinct bands for GPI (Fig.2). The frequency of successful identification isshown in Table 2. Hexokinase, MDH, ME andPGM could not differentiate the two specieslAg. 2).
When the two biochemical techniques utilizedduring this study were compared, the thin-layerstarch gel electrophoresis technique was morediagnostic than isoelectric focusing for differentiating P. pedffer from P. elegonensis. With theformer method, the two species could be separated by four enzymes, GPI, HK, ICD and PGM,with an electromorph frequency of tOO for themost common allele, while in the latter methodseparation was possible only by one enzyme,GPI, also with an electromorph frequency of tOOfor the most common allele. It was furtherobserved that the IEF technique revealed moreisoenzyme bands than the thin-layer starch geltechnique for the enzymes, GPI and PGM.
Righetti and Drysdale [10] discussed thepossible reasons for an increased number ofbands revealed by IEF and concluded that theymay be a result of deamination, deacetylation
TABlE 1. RELATIVE MIGRATION OF BANOS FOR THtN-LAVER STAROI GEL ELECTROPHORESIS FROM THE ORIGIN
AG. 1. THIN-LAYER STARCH Gel ElECTROPHORESIS. Enzyme profilesof MOH. HK, ICO, PGK. GPI and ME for P. pedikr and P. ~$is.Sergentomyia sdI_tzi was used as e matlter species.
656
658 LUaE M. ROGO. C. P. M. KHAMAtA AND M. J. MunNGA
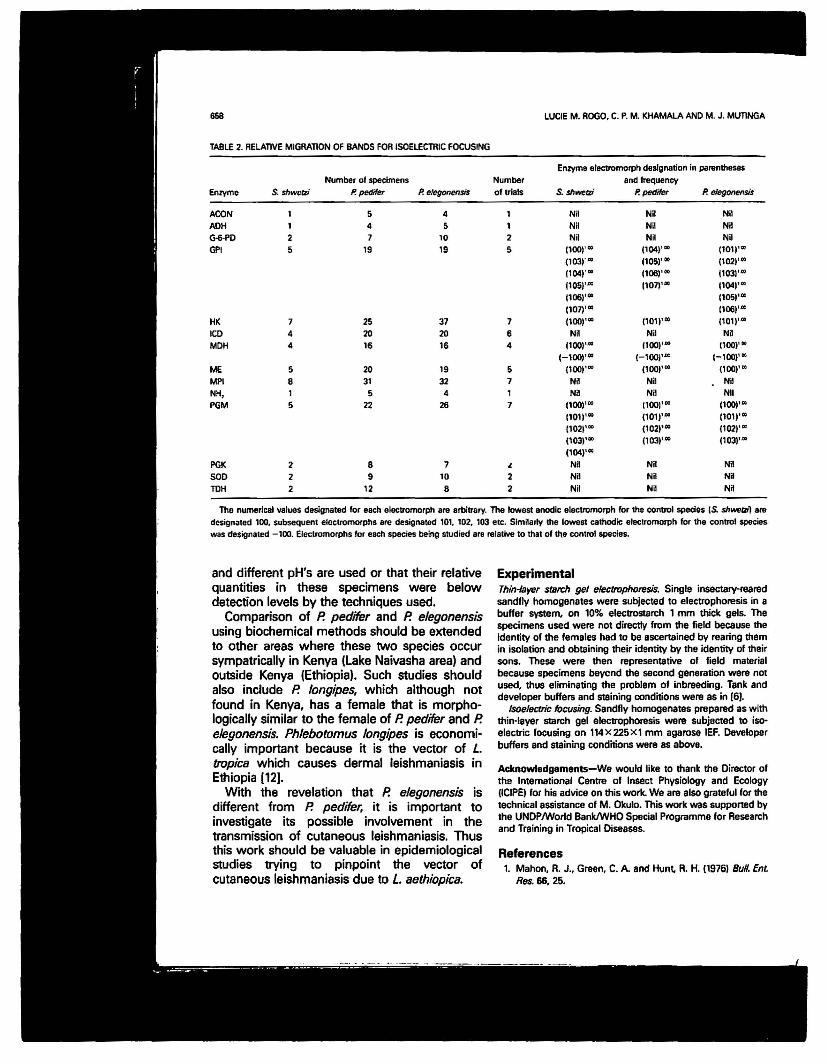
TABLE 2. RELATIVE MIGRATION OF BANDS FOR lSOELEC11lIC FOCUSING
EnZyl119 electromorph designation in pal1ln!hesesNumber ot specimens Number and frequency
Enzyme S. shwrJrzi P. pedifer P. eJegonensis of trials S.shwerzi P. pedifer P. efegonensis
ACON 1 5 4 1 NH Nil NilADH 1 4 5 1 Nil Nil NHG-6.f'D 2 7 10 2 NR NR NRG1'1 5 19 19 5 (1001''' (1041'" (1011''"
(1031''' 11051'" (1021'"(104)''' (1061'" 11031''''11OSl'''' 11071"" 1'041'"(1061'" (1051'"(1071'" (1061'"
HI< 7 25 37 7 (100)'" (lOll'" (101)'''''ICD 4 20 20 6 NR NR NRMDH 4 16 16 4 11(0)'''' (1001'" (1001'"
1-100)'" (-100)''''' 1-1001'"ME 5 20 19 6 (1001'" (1001'" (1001'"MPI 8 31 32 7 NR NR NaNH, 1 5 4 1 NR NR NilPGM 5 22 26 7 11001"" (1001'" 11001'"
11011''''' (1011'''' (1011'"(1021'" (1021''''' (1021'"(1031'" (103)'" (103)'"(1041'"
PGK 2 8 7 ~ NR NH NaSOD 2 9 10 2 NR NR NiTDH 2 12 8 2 NR Nil NR
The numerical values dll6ignaled tor each elec1romorph are arbilrtlry. The lowest anodic eleetromorph tor the control species (S. shwellll aredesignated 100. subsequent electromorphs are designated 10t. 102. 103 etc. SimRarly !he lowest cathodic electromorph for the control species
was designaled - tOO. EleclromOfphs tor each species b1!ing studied are reialive to thaI at II1e control species.
and different pH's are used or that their relativequantities in these specimens were belowdetection levels by the techniques used.
Comparison of P. pedifer and P. elegonensisusing biochemical methods should be extendedto other areas where these two species occursympatrically in Kenya (Lake Naivasha area) andoutside Kenya (Ethiopia). Such studies shouldalso include P. longipes, which although notfound in Kenya, has a female that is morphologically similar to the female of P. pedifer and P.elegonensis. Phlebotomus longipes is economically important because it is the vector of L.tropica which causes dermal leishmaniasis inEthiopia (12).
With the revelation that P. elegonensis isdifferent from P. pedifer, it is important toinvestigate its possible involvement in thetransmission of cutaneous leishmaniasis. Thusthis work should be valuable in epidemiologicalstudies trying to pinpoint the vector ofcutaneous leishmaniasis due to L. aethiopica.
ExperimentalThin-layer stBrch gel electrophoresis. Single insectary-rearedsandfly homogenates were subjected to electrophoresis in abuffer S\'Stem, on 10% e1ectroslarch 1 mm thick gels. Thespecimens used were not directly from the field because theidentity of the females had to be ascertained by rearing lhemin isolation and obtaining their identity by the identity ollheirsons. These were then representative of field materialbecause specimens beycnd the second generation were notused, thus eliminating the problem of inbreeding. Tank anddeveloper buffers and staining conditions were as in 161.
I$f)electric focusing. Sandfly homogenates prepared as withthin-laver stBrch gel electrophoresis were subjected to isoelectric focusing on 114X225x1 mm agarose IEF. Developerbuffers and staining conditions were as above.
Ac:knowledgements-We would like to thank the Director ofthe International Centre of Insect Physiology and Ecology(lCIPEj 101 his advice on this work. We are also grateful for thetechnical assistance of M. Okulo. This work was supponed bythe UNDPIWorid BanklWHO Special Programme for Researchand Training in Tropical Diseases.
References1. Mahon, A. J .• Green. C. A. and Hunt. R. H. (1976) Bull Ent
Res. 66, 25.

But whatever biochemical technique used, P.pedifer could be differentiated from P. elegonensis. These investigations agree with those ofWard et al [7] and Petersen [8] that biochemical methods are reliable and may beused to differentiate between closely relatedspecies of sandfly groups. These biochemicalinvestigations confirm the work of Miles andWard [6] that the main components of theenzymes detected are the same for male andfemale. The demonstration that AOH, G-6-PO,NH and SOD could not be detected in individualsandflies is not necessarily an indication thatthese enzymes were absent It is possible thatthese enzymes may be detected if other buffers
FIG. 2. ISOflECTRIC FOCUSING ON AGAROSE. Enzyme profiles for MOH. HK. PGM. GPI and ME fot P. pcdifer and P. e!egonensis. SergentomyiBsdIwetzi was used as a marfcer species.
and interactions with ligand-like substrates orco-factors after translation. It is most likely thatthe increased number of bands was notrevealed with enzymes MOH, HK and ME whichwere more or less similar to bands resolved bythin-layer starch gel electrophoresis, becausethe different pH ranges used in the presentinvestigation were not the optimum pH's for theenzymes as they gave results contrary to thoseof some workers; for example, the work ofAllsopp and Gibson [11] who compared the twobiochemical techniques using the enzymes, LOHand GPI, from several parasites, but preferredisoelectric focusing because it was more discriminatory.
ItIt~
~
,1iI BlOCHEMICAlIOENTlFICATION OF PHLEBOTOMUS PEDIIER AND P. ELEGONENSIS 6S7
Il,I
I I ., NOH ., HK .1 PGMI
• - • 1011 ;j-• • 103 • • .... ._. : • l8ISI.. •.. - fill
III .. lID
5 P A - 5 P A - S P A
., GPI
~INE
- - IlIIIlI------------ -..--5 P A J 5 P A
S-~. p.e1l!Si!I:!'. A-P.~ _ PrCll1llnl!n band
(1011'.110(lOll'''(lOll'"(-991'"(1001·...(1001'"{lOW"(1021'"
Nil(901'"(9SI""
(100''''{lOll'"(lOll'''
NaPOOl'"
P, eIegonetISis
N~
NilNil
(1001""
arenth8s8s
es (5. shwotzll islhe control was
o Fig. lI.
I. J.MUTINGA
as utilizedthin-layer
was more,r differen-
With the: be separand PGM,00 for theer method3 enzyme,ney of tOOas further·aled morestarch gel
GM.ussed thelumber ofj that theyacetylation
vhareas P.. GPI (Fig.fication is, ME ando species

BIOCHEMICAL IDENllFlCAll0N OF PHLEBOTOMUS PEDIFER AND P. ELEGONENSJS 659
Med. Parasit. 75, 235.8. Petersen, J. L (19821 Proc. Worlcshop Pan American Health
Organization 1980, p. 105. Washington D.C.9. Mutlnga, M. J. 11975) E. Ah: Mer/. J. 52. 340.
10. Righetti, P. G. and Drysdale. J. W. (1976) Isoe/eetricFocusing. North-Holland/American Elsevier, Amsterdam.
11. Allsopp, B. A. and Gibson. W. C. (1983) Ann. TIOp. Mad.Parasit 77, 169.
12. lemma, A.. Foster. W. A.. Gemetehu, T.• Preston. P. M.•Bryceson. A. and Minter. D. M. (1969) Ann. TlOp. Mer/.Parasit 63, 455.
' .. '
\ ,:
",,1
I
I
.,.::;'';-
2. Miles. S. J. (1979) J. Med. EntomoJ. 15. 279.3. Saul. S. H.• Sinsko. M. J., Grimstad. P. R and Craig Jr. G.
B. (1977} J. Med. Ent 13. 705.4. Pasteur. N., Rioux. J. A.• Guilard. E.• Pech-Perillr8s. M. J.
and Verdier. J. M. (1977) Annis. Pamsit. Hum. Comp. !i2,325.
5. Townson, H. T. and Meredith. S. E. O. (1979) Identificationof the Simu5dse In Relstion to Onchoceteiasis p. 145.Blackwells. Oxford. .
6. Mnes, S. J. and Ward. R. D. (1978) Ann. Trop. Med Parasit.12, 398.
7. Ward. R. D.• Pasteur. N. and Rioux. J. (1981) Ann. Tmp.
Director 01nd EcologyleluJ lor theIPPOrled byor Research
~tarrreared
horesis in ak gels. ThelecaUse thedaring themnilY 01 theirId materialm were not). Tank andin [6).
Ired as wilh:ted to iso. Developer
NaNaNa
11011""11021""(103)'''11041""IIOS1'"(1061''''110'1'"
Na11001'"
-100)""
1100)'"NaNil
1'00)'"1101)'"1102)'"(1031'"
·J.MUTINGA
/6) Bull. Ent
.'egonensi$





























