Embed Size (px)
Citation preview

1
Running head: Rice miRNAs in Immunity against the Blast Disease 1
Corresponding author: Wen-Ming Wang 2
Address: Rice Research Institute, Sichuan Agricultural University, Chengdu 611130, China 3
Tel: 86-28-86290949 4
E-mail: [email protected] 5
Research Area: Signaling and Response 6
Plant Physiology Preview. Published on December 13, 2013, as DOI:10.1104/pp.113.230052
Copyright 2013 by the American Society of Plant Biologists
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

2
Multiple Rice miRNAs Are Involved in Immunity against the Blast 7
Fungus Magnaporthe oryzae1 8
9
Yan Li2, Yuan-Gen Lu2, Yi Shi2, Liang Wu, Yong-Ju Xu, Fu Huang, Xiao-Yi 10
Guo, Yong Zhang, Jing Fan, Ji-Qun Zhao, Hong-Yu Zhang, Pei-Zhou Xu, 11
Jian-Min Zhou, Xian-Jun Wu, Ping-Rong Wang and Wen-Ming Wang* 12
Rice Research Institute and College of Agronomy, Sichuan Agricultural University, 13
Chengdu 611130, China (Y.L., Y.-G.L., Y.S., Y.-J.X., F.-H., X.-Y.G., Y.Z., J.F., J.-Q.Z., 14
H.-Y.Z., P.-Z.X., X.-J.W., P.-R.W., W.-M.W.); National Key Facility for Crop Resources 15
and Genetic Improvement, Institute of Crop Science, Chinese Academy of Agricultural 16
Sciences, Beijing 100081, China (L.W.); Institute of Genetics and Developmental Biology, 17
Chinese Academy of Sciences, Beijing 100101, China (J.-M.Z.);Rice and Sorghum 18
Institute, Sichuan Academy of Agricultural Sciences, Deyang 618000, China (X.-Y.G.) 19
20
One-sentence summary 21
Deep sequencing small RNA libraries from susceptible and resistant lines identified 22
multiple miRNAs differentially responsive to the infection of the blast fungus Magnaporthe 23
oryzae and ectopic expression of either miR160a or miR398b confers elevated resistance to 24
the blast disease. 25
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

3
1This work was supported by National Natural Science Foundation of China (grant number 26
31101406 to Y.L.), Sichuan Agricultural University Start-up Fund to W.-M.W., and by the 27
Special Fund for Agro-scientific Research in the Public Interest (201203014 to F.H.). 28
2These authors contributed equally to the paper. 29
* Address correspondence to [email protected] 30
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

4
ABSTRACT 31
MicroRNAs (miRNAs) are indispensable regulators for development and defense in 32
eukaryotes. However, the miRNA species have not been explored for rice immunity against 33
the blast fungus Magnaporthe oryzae, the most devastating fungal pathogen in rice 34
production worldwide. Here, by deep sequencing small RNA (sRNA) libraries from 35
susceptible and resistant lines at normal conditions and upon M. orzyzae infection, we 36
identified a group of known rice miRNAs that were differentially expressed upon M. 37
oryzae infection. They were further classified into three classes based on their expression 38
patterns in the susceptible line LTH (Lijiangxin Tuan Hegu, Oryza sativa L. japonica) and 39
in the resistant line IRBLkm-Ts that contains a single resistance gene locus Pikm within 40
LTH background. RNA-blotting assay on nine of them confirmed sequencing results. 41
Real-time reverse transcription (RT) PCR assay showed the expressions of a part of target 42
genes were negatively correlated with the expressions of miRNAs. Moreover, transgenic 43
rice plants over-expressing miR160a and miR398b displayed enhanced resistance to M. 44
oryzae as demonstrated by decreased fungal growth, increased H2O2 accumulation at the 45
infection site and up-regulated expression of defense-related genes. Taken together, our 46
data indicate that miRNAs are involved in rice immunity against M. oryzae and 47
over-expression of miR160a or miR398b can enhance rice resistance to the disease. 48
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

5
In plant-pathogen co-evolution, plants mount two-layered immune system to protect 49
themselves from being damaged by pathogenic microbes. In the first layer of immunity, 50
plants detect conserved molecular features of microbes, termed pathogen-associated 51
molecular patterns (PAMPs) to trigger immunity (PTI) that is efficient to prevent a large 52
number of potential pathogenic microbes from invasion (Zipfel and Felix, 2005). 53
Successful pathogens deliver effectors into plant cells to suppress PTI and establish 54
parasitism (Boller and He, 2009; Dou and Zhou, 2012). In turn, the second layer of plant 55
immunity, called effector-triggered immunity (ETI), is initiated upon the recognition of 56
effectors by the cognate intracellular immune receptors, i.e. nucleotide-binding-site 57
leucine-rich repeat (NBS-LRR) type proteins. ETI is usually concomitant with 58
hypersensitive response (HR), a programmed cell death at the infection site to inhibit the 59
diffusion of the invading pathogen (Alfano and Collmer, 2004; Chisholm et al., 2006; Jones 60
and Dangl, 2006). In plant-fungal interactions, PTI and ETI equip plant pre- and 61
post-invasive resistance, respectively. Increasing evidence demonstrates that small RNAs 62
are involved in both PTI and ETI signaling and act as key fine-tuning regulators 63
(Padmanabhan et al., 2009; Katiyar-Agarwal and Jin, 2010). Small RNAs are short 64
non-coding RNAs which guide gene silencing either by regulating chromatin 65
methyl-modification or by mRNA degradation and translational repression (Baulcombe, 66
2004; Seo et al., 2013). They are classified into microRNAs (miRNAs) and small 67
interfering RNAs (siRNAs) based on their biogenesis and origin. Both miRNAs and 68
siRNAs are involved in regulation of diverse biological processes including development 69
and responses to biotic and abiotic stresses (Katiyar-Agarwal and Jin, 2010; Chen, 2012; 70
Khraiwesh et al., 2012). In Arabidopsis, a set of miRNAs were identified to be responsive 71
to the PAMP molecule flg22 or to the infection of non-pathogenic strain P. syringae 72
DC3000 (hrcC-) (Fahlgren et al., 2007; Li et al., 2010; Zhang et al., 2011). The first 73
identified PTI-related small RNA is miR393 that can be induced by the PAMP molecule 74
flg22 and positively contributes to PTI by suppressing auxin signaling via silencing auxin 75
receptors (Navarro et al., 2006). In addition, miR160a, miR398b and miR773 are involved 76
in regulation of callose deposition and thus act in PTI signaling (Li et al., 2010). Some 77
miRNAs also exhibit differential induced responses to the infection of pathogenic and 78
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

6
avirulent strains of P. syringae DC3000 and thus are involved in ETI signaling (Zhang et 79
al., 2011). More recently, some miRNAs are found to guide the cleavage of NBS-LRR type 80
disease resistance (R) genes in Solanaceae and Leguminosae species, indicating that these 81
miRNAs are key regulators in ETI (Zhai et al., 2011; Li et al., 2012; Shivaprasad et al., 82
2012). 83
The study of miRNAs in rice has been progressed from computational prediction to 84
experimental identification and functional characterization (Reinhart et al., 2002; Rhoades 85
et al., 2002; Jones-Rhoades and Bartel, 2004; Liu et al., 2005). The first set of rice miRNAs 86
experimentally identified was reported in 2004 (Wang et al., 2004). Later on, more 87
miRNAs either conservative or novel in rice were identified from rice shoot, root, 88
inflorescence, panicle, calli, developing grains, and immature seeds (Liu et al., 2005; 89
Sunkar et al., 2005; Luo et al., 2006; Zhu et al., 2008; Xue et al., 2009). By exposing rice 90
seedlings to drought or salt stress and through high throughput sequencing, Sunkar et al. 91
(2008) identified 23 new miRNAs and 40 candidates (Sunkar et al., 2008). By subject to 92
drought stress from tillering to inflorescence formation stages, 30 miRNAs were identified 93
to be differentially expressed under drought conditions (Zhou et al., 2010). By comparing 94
samples from normal conditions and exposed to oxidative stress, Li et al. (2011) identified 95
seven miRNA families that are differentially responsive to oxidative stress and discovered 96
32 new rice miRNAs (Li et al., 2011). By analyzing 62 small RNA libraries that represent 97
several tissues from control plants and those subjected to different environmental stress 98
treatments, Jeong et al. (2011) re-evaluated ~400 annotated miRNAs and found 76 new 99
miRNAs that are responsive to water stress, nutrient stress, or temperature stress (Jeong et 100
al., 2011). In addition, because miRNA relies on DCL1 for maturation, rice loss of function 101
mutation in OsDCL1 is lethal at seedling stage (Liu et al., 2005), implying that miRNA is 102
essential for development. MiR166 was ectopically expressed in mutants with mutations in 103
SHOOTLESS2 (SHL2), SHL4/SHOOT ORGANIZATION2 (SHO2), and SHO1 that are the 104
orthologues of Arabidopsis RNA-dependent RNA polymerase 6, ARGONAUTE 7, and 105
Dicer-like 4, respectively, leading to the silencing of the miR166 target genes OSHB1 and 106
OSHB2, two HD-ZIPIII genes (Nagasaki et al., 2007). Members of miR156 families target 107
the SQUAMOSA (SQUA) promoter-binding-like (SPL) genes and over-expression of 108
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

7
OsmiR156b/h resulted in increased tillers but reduced plant height and panicle size (Xie et 109
al., 2006). These literatures demonstrate that miRNAs play indispensable roles for 110
development and abiotic stress responses in rice. However, it remains elusive to identify the 111
miRNA species involved in rice immunity against the blast fungus M. oryzae. 112
The rice-rice blast fungus plant-pathogen system has become a model in the study of 113
plant-fungal interactions because rice is one of the most important staple food for the most 114
population of the world and rice blast is the most devastating disease for rice production 115
(Liu et al., 2013). The blast fungus M. oryzae can infect rice plants at any developmental 116
stages. After attaching onto the surface of a rice cell, conidium germinates to form a germ 117
tube at about 2 hours post inoculation (hpi), and then the germ tube differentiates an 118
appressorium at 2-20 hpi, which guides the fungus to penetrate into the underneath 119
epidermal cell at 24-30 hpi (Ribot et al., 2008). After entrance into the host cell, the fungus 120
develops bulbous infectious hyphae without visible damages to the host at first. With the 121
development of colonization, the fungus destroys the infected cells at 2-5 dpi and spreads 122
the disease by sporulation (Ribot et al., 2008). Both PTI and ETI are involved in rice 123
immunity against the blast fungus (Liu et al., 2013). Chitin, a conserved fungal-derived 124
PAMP, can be recognized by rice pattern-recognition receptors CEBiP (chitin 125
oligosaccharide elicitor-binding protein) to trigger innate immunity (Kaku et al., 2006; 126
Kishimoto et al., 2010; Shimizu et al., 2010). Knockdown of CEBiP in rice cell lines results 127
in obvious suppression of chitin-triggered oxidative burst (Kaku et al., 2006). The 128
PTI-related genes, such as OsKS4 and OsNAC4, can be induced by chitin in wild type 129
plants, but suppressed in susceptible transgenic lines (Park et al., 2012). M. oryzae delivers 130
a batch of effectors into rice cells through biotrophic interfacial complex (BIC) or invasive 131
hyphae to enhance its virulence (Mosquera et al., 2009; Khang et al., 2010; Mentlak et al., 132
2012). Some effectors are recognized by intracellular receptors, termed resistance (R) 133
proteins, to trigger race-specific resistance which contributes to the post-invasive resistance. 134
In the past decades, twenty-one rice blast R genes have been characterized by molecular 135
cloning, 19 of which encode NBS-LRR proteins, while Pi-d2 and Pi21 encode a 136
receptor-like kinase (RLK) protein and a Proline-rich protein, respectively (Fukuoka et al., 137
2009; Liu et al., 2010; Liu et al., 2013). Pikm locus contains two NBS-LRR genes, 138
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

8
Pikm1-TS and Pikm2-TS, both are required to recognize the cognate effector Avr-Pikm to 139
trigger effective resistance to the blast disease (Li et al., 2007; Yoshida et al., 2009). The 140
synergistic function of the two adjacent genes confers Pikm-specific resistance strongly 141
against infection of the rice blast fungus (Ashikawa et al., 2008). Pi2 and Piz-t are alleles 142
showing high similarity to Pi9 and they confer broad-spectrum resistance against different 143
sets of M. oryzae isolates (Qu et al., 2006; Zhou et al., 2006). However, up to date, limited 144
literatures imply that small RNAs may be involved in rice immunity against the blast 145
fungus M. oryzae. A recent report assayed the accumulation of miRNAs in rice treated by 146
elicitors extracted from blast fungal mycelia (Campo et al., 2013). A new rice miRNA, 147
osa-miR7695, is identified to be involved in rice resistance by negatively regulating the 148
transcript of OsNramp6 (Natural resistance-associated macrophage protein 6) (Campo et al., 149
2013). Currently, miRBase lists over 700 mature miRNA sequences cloned or predicted in 150
rice (http://www.mirbase.org) (Kozomara and Griffiths-Jones, 2011). Nevertheless, whether 151
and which of the known miRNAs are involved in rice immunity against the blast fungus 152
remain to be identified. 153
To obtain the miRNAs involved in rice immunity against the blast fungus, we 154
performed a systemic screen by comparing the abundance of miRNAs in the susceptible 155
line LTH and the resistance line IRBLkm-Ts that carries the Pikm locus. By deep 156
sequencing six small RNA libraries from samples of LTH and IRBLkm-Ts collected at 0, 157
12, and 24 hpi of M. oryzae, we identified candidate miRNAs that may be involved in rice 158
immunity because of their differential responses to the infection of the blast fungus. 159
Overexpression of miR160a and miR398b in transgenic rice plants enhanced resistance to 160
the blast fungus. Taken together, our data demonstrate that it is feasible to engineering rice 161
resistance by identification and ectopic expression of individual miRNA that is involved in 162
immunity against the blast fungus. 163
164
RESULTS 165
Identification of the Monogenic Resistant Line with Strong Defense Responses 166
To identify miRNAs most possibly involved in rice immunity against the blast disease, we 167
decided to choose an accession harboring an R gene that enables strong defense responses. 168
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

9
Previous study indicates that R genes at two loci render broad-spectrum resistance to the 169
blast disease in rice production (Zhou et al., 2006; Li et al., 2007; Ashikawa et al., 2008). 170
We thus chose rice lines containing R genes at these loci, i.e. IRBLkm-Ts and IRBLz5-CA, 171
in our investigation, and focused on one of them for further investigation. IRBLkm-Ts and 172
IRBLz5-CA are monogenic lines containing the resistance gene Pikm and Pi2 that were 173
introduced into LTH by backcrossing, respectively (Tsumematsu H, 2000). LTH is a 174
japonica variety highly susceptible to over 1300 regional isolates of M. oryzae worldwide 175
and no major R gene is ever identified in it (Lin et al., 2001). To evaluate Pikm- and 176
Pi2-mediated defense responses, three-leaf-old seedlings were inoculated with conidial 177
mixture of 15 M. oryzae strains collected from our rice blast disease nursery in Ya’an, 178
Sichuan province, China. Disease phenotypes were recorded at 10 dpi. Compared to LTH, 179
both IRBLkm-Ts and IRBLz5-CA displayed obvious resistant phenotypes (Fig. 1A). Then 180
by quantitative RT-PCR (qRT-PCR), we examined the expression of five defense-related 181
genes, including OsPR1, OsPR10, and the PTI-related genes OsPBZ1, OsKS4 and OsNAC4 182
(Park et al., 2012; Yamaguchi et al., 2013). The transcripts of all the five genes were 183
significantly increased in IRBLkm-Ts and IRBLz5-CA in comparison with those in LTH 184
(Fig. 1B-F). Moreover, OsRP1, OsPR10, OsPBZ1, and OsKS4 were induced earlier and to 185
greater amplitude in IRBLkm-Ts than in IRBLz5-CA (Fig. 1B-E). These data suggest that 186
Pikm renders defense responses faster and stronger than that does Pi2 against M. oryzae. 187
Because the production of H2O2 in infected leaf cells is a signal that precedes 188
hypersensitive responses (HR) that often culminates the defense responses in ETI (Levine 189
et al., 1994), we examined H2O2 accumulation and fungal growth at 2 and 10 dpi by 190
3,3’-diaminobenzidine and trypan-blue staining, respectively. As anticipated, in contrast to 191
no or trace amount of H2O2 accumulation in leaf cells of LTH, we observed high levels of 192
H2O2 accumulation around the appressoria at 2 dpi in the leaf cells of both IRBLkm-Ts and 193
IRBLz5-CA with stronger in IRBLkm-Ts (Fig. 1G). At 10 dpi, we observed massive aerial 194
mycelia with a great number of conidiophores on leaves of LTH, though H2O2 was also 195
accumulated in some cells (Fig. 1G, Supplemental Fig. S1A). In contrast, little hyphae and 196
no conidiophores were observed on leaves of IRBLkm-Ts and IRBLz5-CA, and high levels 197
of H2O2 were accumulated in the cells attacked by the appressoria (Fig. 1 G, Supplemental 198
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

10
Fig. S1B-C). 199
Taken together, these data suggest that the defense responses in IRBLkm-Ts are 200
stronger than that in IRBLz5-CA. Therefore, we used IRBLkm-Ts in the subsequent 201
screening for miRNAs involved in defense against M. oryzae. 202
203
Deep-Sequencing Analysis on Small RNA Libraries from M. oryzae-Free and Infected 204
Rice Leaves 205
To obtain miRNAs involved in rice defense against M. oryzae, three-week-old seedlings of 206
LTH and IRBLkm-Ts were inoculated with spore mixture of M. oryzae. Infected leaves 207
were collected at 12 and 24 hpi with leaves before inoculation as control. Six small RNA 208
libraries were constructed with the small RNA extracted from the collected leaves and 209
subjected to high-throughput sequencing. In total, 3,791,618, 1,277,195 and 2,559,590 210
reads for LTH and 2,723,456, 1,867,650 and 2,000,003 reads for IRBLkm-Ts from samples 211
collected at 0, 12 and 24 hpi matched to rice genome, respectively (Supplemental Table S1). 212
These reads represent 1,952,485, 765,215 and 1,379,760 unique sRNA sequences for LTH; 213
and 1,686,864, 1,127,034, and 1,139,492 unique sRNA sequences for IRBLkm-Ts at the 214
three time points, respectively (Supplemental Table S1). The size of most small RNAs is 21 215
and 24 nucleotides (nt), while the size of most known miRNAs is 21 nt (Mi et al., 2008; Wu 216
et al., 2009; Zhao et al., 2010). Consistently, small RNAs in our libraries were 217
predominantly 21 and 24 nt (Fig. 2A), while miRNAs were 21 nt (Fig. 2B), indicating that 218
our data is highly reliable. There were 50%-85% of small RNAs matched to rice genome in 219
the six libraries (Supplemental Table S1, Fig. 2C). Five to twenty-five percent of reads in 220
the libraries of inoculated samples (collected at 12 and 24 hpi) matched to M. oryzae 221
genome, while only about 1% reads in the libraries constructed from the control samples 222
matched to M. oryzae genome (Supplemental Table S1, Fig. 2C). These data suggest that 223
the small RNAs identified were of rice and M. oryzae origins. 224
However, there were 20%-30% reads that could not match to the rice or M. oryzae 225
genome in each library (Supplemental Table S1), which is presumably due to the DNA 226
sequence polymorphisms between the rice cultivars / M. oryzae isolates used and the 227
reference genomes. 228
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

11
Then the rice small RNA sequences were classified into different categories based on 229
their match to the rice genome (Supplemental Table S1, Fig. 2D). The largest category of 230
the small RNAs was heterochromatic siRNAs (hc-siRNAs), followed by small RNAs 231
matched to protein-coding genes (Fig. 2D). Over 20 thousand reads from each library that 232
accounted for 10%-20% matched to rice mitochondria and chloroplast genome 233
(Supplemental Table S1, Fig. 2D). There were 2%-3% of reads matched to nat-siRNAs but 234
no ta-siRNAs (Fig. 2D). Reads matched to rice miRNAs accounted for 5%-10%, which is 235
consistent with a previous report (Fig. 2D) (Wu et al., 2009). Compared with control 236
sample at 0 dpi, miRNA reads were decreased significantly at 12 hpi in LTH, whereas 237
increased at 24 hpi, suggesting that the blast fungus suppresses miRNA biogenesis in 238
compatible rice-blast fungus interaction during pre-penetration stage (Supplemental Table 239
S1, Fig. 2D). In contrast, the miRNA reads were increased in IRBLkm-Ts along with the 240
infection of M. oryzae (Supplemental Table S1, Fig. 2D), implying that miRNAs might 241
play roles in rice immunity against M. oryzae. Because of the large volume of total 242
sequence data and our primary focus in this study on learning about how the known 243
miRNAs were involved in rice immunity, novel miRNAs due to M. oryzae infection or the 244
M. oryzae-derived miRNAs and their biological implications will be the focus of future 245
research. 246
247
Candidate MicroRNAs Involved in Rice -M. oryzae Interaction 248
The miRNAs involved in immunity must display differential expression upon M. oryzae 249
infection. According to this speculation, we compared the miRNA accumulation patterns 250
between the libraries from LTH and IRBLkm-Ts upon M. oryzae infection, and the 251
miRNAs with reads changing over-2-fold in any two of the six libraries were selected for 252
further investigation (Supplemental Table S2). Accordingly, these miRNAs were classified 253
into 39 patterns by analysis with Short Time-series Expression Miner (STEM) software 254
(Ernst and Bar-Joseph, 2006) based on their change trends, of which 19 patterns showed 255
significant changes in the accumulation of miRNAs upon M. oryzae infection 256
(Supplemental Table S3). We further classified the miRNAs of the 19 patterns into 3 257
classes based on their change upon M. oryzae infection: 1) no obvious change in LTH, but 258
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

12
significantly increased in IRBLkm-Ts (i.e. patterns #2, #11, #22) or reduced in LTH, but no 259
obvious change or increased in IRBLkm-Ts (i.e. #7); 2) no significant change in LTH, but 260
decreased greatly in IRBLkm-Ts (i.e. #21) or increased in LTH, but no obvious change or 261
decreased in IRBLkm-Ts (i.e. #08, #16, #18, #25, #30, #32, #40, #42 and #45); 3) 262
increased in both LTH and IRBLkm-Ts upon M. oryzae infection, including the patterns 263
#27, #29, #39, #46 and #47. Theoretically, the first two classes of miRNAs should play 264
positive and negative roles in rice resistance, respectively; while the third class may be 265
involved in regulation of basal responses to the M. oryzae infection because of their 266
up-regulation in both susceptible and resistant lines. 267
Next, the miRNAs belonging to the classes mentioned above with more than 100 reads 268
in one of the six libraries were selected for further analysis (Supplemental Table S2). In 269
total, 33 miRNAs were selected with the pattern number listed in the bracket following 270
their name in Table 1. The expressions of nine miRNAs were analyzed by sRNA-blotting to 271
verify the deep sequencing results, including three positive regulators (i.e. miR160a, 272
miR164a and miR168a), three negative regulators (i.e. miR396, miR827 and miR1871), and 273
three basal responsive regulators (i.e. miR169a, miR172a, and miR398b) (Fig. 3A). The 274
sRNA-blotting analysis was basically consistent with the sequencing results. As shown in 275
Fig. 3A, the accumulation of miR160a and miR164a were significantly induced to higher 276
levels upon M. oryzae infection in IRBLkm-Ts. MiR172a and miR398b were increased in 277
both LTH and IRBLkm-Ts, whereas miR1871, miR396d and miR827a were obviously 278
increased in LTH. 279
280
Expression Pattern of The miRNA Target Genes upon M. oryzae Infection 281
Because the regulatory roles of miRNAs in gene expression is to cleave target mRNAs or 282
repress translation, we firstly examined the transcripts of some target genes which were 283
previously validated by degradome assay using qRT-PCR (Supplemental Table S4) (Wu et 284
al., 2009; Li et al., 2010; Zhou et al., 2010). As our anticipation, the expressions of certain 285
targets of miR160, miR164, miR827, miR172 and miR398 exhibited negative correlation to 286
the abundance of miRNAs (Fig. 3B-G), which is consistent with one of the miRNAs’ roles 287
to guide degradation of their target genes. Among the tested target genes, Os04g43910 288
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

13
encodes putative auxin response factor 16 (ARF16) and Os07g46990 encodes superoxide 289
dismutase 2 (SOD2); and they were previously confirmed as the target of miR160 and 290
miR398 by degradome cDNA library assay, respectively (Wu et al., 2009). Os04g48410 291
encoding a copper chaperone for SOD was identified as another target of miR398 (Li et al., 292
2010). In addition, Os12g41680, Os04g48390, and Os05g03040 were confirmed as the 293
targets of miR164, miR827 and miR172 by degradome sequencing, respectively (Wu et al., 294
2009; Li et al., 2010; Zhou et al., 2010). The negative correlation between the abundant of 295
miRNA and its target indicates that they are truly target of the miRNAs and thus may be 296
involved in regulation of rice immunity against M. oryzae. Nevertheless, nine of the 297
confirmed or predicted target genes exhibited partially negative correlation with the 298
miRNA abundance (Supplemental Fig. S3 and Supplemental Table S4), thus whether they 299
are targets of the miRNAs regulated via translational repression needs further experimental 300
verification. 301
302
Enhanced Defense Responses to M. oryzae in The Transgenic Rice Lines 303
Over-Expressing miR160a and miR398b 304
Next, we took transgenic approach to verify representatives of the miRNAs involved in 305
defense responses to M. oryzae. Kasalath is an indica accession that possesses high 306
re-generation rate during tissue culture and is sensitive to most M. oryzae strains (Li et al., 307
2007), thus suitable for employment in transgenic analysis. MiR160a represents positive 308
regulators of resistance; its expression was not much changed in LTH but increased 309
approximately 4-fold upon M. oryzae infection in IRBLkm-Ts (Table 1 and Fig. 3A). Two 310
independent transgenic lines with high miR160a expression were identified by miRNA 311
real-time RT-PCR and RNA-blot analyses in comparison with the control line (Fig. 4A,B). 312
Consistently, the expression of its target genes Os02g41800 and Os04g43910, both of 313
which encode putative auxin response factor 16 (ARF16), were reduced to less than 10% of 314
that in the control line (Fig. 4C). The transcript of Os04g59430, another target of miR160 315
encoding a B3 DNA binding domain containing protein, was also significantly suppressed 316
though not as much as that of Os02g41800 and Os04g43910 (Fig. 4C). However, the 317
expression of Os10g33940 and Os06g47150, both of which also encode putative ARF16, 318
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

14
were up-regulated to levels higher than that in the control line (Supplemental Fig. S4), the 319
simplest explanation is that miR160a may repress their translation or they are under 320
feedback regulation by their products. Next, we examined the transgenic lines by 321
inoculating with mixed M. oryzae strains, and disease phenotypes were recorded at 10 dpi. 322
The number of lesions in the transgenic lines over-expressing miR160a was obviously less 323
than that in the control line (Fig. 4D). In addition, the disease lesions were mainly scored as 324
type 1 - 2 in the miR160a over-expression line #4, which was obviously less severe than the 325
type 3 - 4 in the control line (Fig. 4D). These observations suggested that the resistant level 326
was higher in the transgenic lines. Consistent with the resistant phenotype, the spore 327
number and relative fungal mass in the infected leaves, indicated as the ratio of the M. 328
oryzae Pot2 gene versus the rice ACTIN1 (Berruyer et al., 2006; Park et al., 2012), were 329
significantly reduced in the transgenic lines than that in the control line (Fig. 4E,F). 330
MiR398b represents regulators acting in basal responses because its expression is 331
increased in both LTH and IRBLkm-Ts upon M. oryzae infection. The abundant of 332
miR398b increased approximately 8-fold at 24 hpi in LTH and increased in IRBLkm-Ts 333
over 5-, 8-fold at 12, 24 hpi, respectively (Fig. 3A). We also identified two transgenic lines 334
in which the accumulation of miR398b was significantly increased (Fig. 5A,B). 335
Consistently, its confirmed target genes Os03g22810, Os04g48410, Os07g46990 and 336
predicted target gene Os11g09780 were all suppressed greatly in the two miR398b 337
transgenic lines (Fig. 5C) (Wu et al., 2009; Li et al., 2010; Zhou et al., 2010). Then the 338
miR398b transgenic lines were examined by inoculation with mixed M. oryzae strains and 339
disease phenotypes were recorded at 10 dpi. The number of lesions in the transgenic lines 340
over-expressing miR398b was obviously less than that in the control line (Fig. 5D). In 341
addition, the disease lesions were obviously reduced to type 2 and type 3 in the miR398b 342
over-expression lines #9 and #10, respectively, which were less severe compared to type 3 - 343
4 in the control line (Fig. 5D). These observations suggested that the resistant level was 344
increased in the transgenic lines. Consistent with the resistant phenotype, the spore number 345
and relative fungal biomass in the infected leaves were significantly reduced in transgenic 346
lines than that in the control line (Fig. 5E,F). 347
Resistant phenotypes are usually accompanied with up-regulation of defense-related 348
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

15
genes, thus we examined the transcription of some defense-related genes in one line 349
over-expressing miR160a or miR398b, respectively. As our anticipation, both miR160a and 350
miR398b transgenic lines displayed expression of OsPR1 and OsPR10 significantly higher 351
than that of the control line and the expression level in the miR398-overecpression line was 352
higher than that in the miR160a-over-expression line (Fig. 6A,B). In addition, the 353
expression of PTI-related genes OsKS4 and OsNAC4 was significantly enhanced in the 354
miR398b transgenic line, but was not obviously affected in the miR160a transgenic line 355
(Fig. 6C,D). Furthermore, the transcripts of both OsKS4 and OsNAC4 were significantly 356
enhanced at 1 hour after treated by flg22 and chitin in the miR398b-overexpression line, but 357
were not affected in the miR160a-overexpression line (Fig. 6E,F), suggesting that miR398b 358
is most likely involved in PTI-signaling to regulate defense responses, while miR160a may 359
be in ETI-signaling. 360
To visualize how the transgenic lines over-expressing miR160a and miR398b mount 361
resistance on cellular level, the virulent M. oryzae strain Zhong-8-10-14 expressing 362
enhanced GFP (eGFP) was inoculated on leaf sheath. Then the pathogenesis was observed 363
under Laser Scanning Confocal Microscope (LSCM). The spores were germinated and 364
formed appressoria on sheath of control plants at 12 hpi; and invasive hyphae were formed 365
at 24 hpi and proliferated widely at 36 hpi (Fig. 7A). In contrast, the spores did not 366
germinate and appressoria were not generated until 24 hpi in transgenic plants 367
over-expressing miR160a and 36 hpi over-expressing miR398b, respectively (Fig. 7B-C and 368
F). Furthermore, invasive hyphae were hardly observed on both transgenic plants until 36 369
hpi (Fig. 7B-C and G). 370
Then, the accumulation of H2O2 and fungal growth were examined at 2 and 10 dpi. 371
Microscopic analysis showed that at 2 dpi, H2O2 was accumulated around the 372
appressorium in the leaf cells of miR160a and miR398b over-expression transgenic lines, 373
whereas no or little H2O2 was accumulated in leaf cells in control line (Fig. 7D). At 10 dpi, 374
we observed the well-grown aerial hyphae and massive production of conidiophores in 375
control line, although H2O2 was also accumulated in the cells from where the aerial 376
hyphae were generated (Fig. 7E, Supplemental Fig. S5A). Conversely, in the miR160a and 377
miR398b over-expression lines, we observed high levels of H2O2 accumulation in the leaf 378
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

16
cells beneath the appressoria and few aerial hyphae but no conidiophores were observed 379
(Fig. 7E and Supplemental Fig. S5B-C). These observations indicated that the fungus 380
failed to colonize the leaves. 381
Taken together, the results from transgenic analysis indicate that both miR160a and 382
miR398b contribute to suppression of fungal infection and may act differentially in rice 383
immunity against the blast fungus. 384
385
DISCUSSION 386
Host miRNAs play pivotal roles in plant-microbe interactions. Identification of host 387
miRNAs that are differentially induced by pathogen infection is the first step to investigate 388
their functions in host immunity against the pathogen. Previously, a set of miRNAs were 389
identified to be involved in plant immunity against bacterial and viral pathogens (Navarro 390
et al., 2006; Du et al., 2011; Radwan et al., 2011; Seo et al., 2013). Recently, host miRNAs 391
are also found to be responsive to the infection of a increasing list of filamentous pathogens, 392
such as Cronartium quercuum (Lu et al., 2007), Erysiphe graminis (Xin et al., 2010), 393
Fusarium virguliforme (Radwan et al., 2011), Phytophthora sojae (Guo et al., 2011), and 394
Verticillium dahliae (Yin et al., 2012; Yang et al., 2013). In the present study, small RNA 395
accumulation in rice seedlings of susceptible and resistant lines upon M. oryzae infection 396
was systemically assayed by deep sequencing. The changes in abundance of nine miRNAs 397
were verified by RNA blotting analyses, suggesting that the deep sequencing data are 398
reliable. Totally, we found representatives from 25 known rice miRNA families that were 399
presumably involved in innate immunity against the infection of the blast fungus M. oryzae 400
(Table 1). There seems more known miRNAs that negatively regulate rice immunity than 401
those do positively. Members of four miRNA families, i.e. miR160a/b/c/d/f, miR164a/b/f, 402
miR167a/b/c, and miR168a, were reduced or not much changed in expression in the 403
susceptible line, but significantly increased in the resistant line upon M. oryzae infection, 404
thus they may positively regulate rice immunity against the fungus. In contrast, members 405
from sixteen families were up-regulated in expression in the susceptible line but 406
down-regulated or did not exhibit significant changes in the resistant line after M. oryzae 407
infection, thus they may negatively regulate rice immunity. Members from ten miRNA 408
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

17
families were up-regulated in expression in both the susceptible and resistant lines, thus 409
they may act in fundamental rice responses to M. oryzae infection (Table 1 and Fig. 3). 410
Three miRNA families had members differentially responsive to M. oryzae infection, 411
including miR167, miR169, and miR444. While miR167a/b/c might act as positive regulator 412
for rice immunity, miR167d/e/f/g/h/i/j act as negative regulator. MiR169a/b/c might act in 413
basal responses to M. oryzae infection, whereas miR169f/g/h/i/j/k/l/m might negatively 414
function in rice immunity against the blast fungus. Similarly, miR444b.1/c.1 might function 415
in basal responses to M. oryzae infection, whereas miR444b.2/c.2 might negatively regulate 416
rice immunity against the blast fungus (Table 1 and Fig. 3). Thus the identified known 417
miRNAs involved in rice immunity provides a good starting point to further pursue how 418
rice resistance to the blast fungus is regulated through diverse aspects. 419
In the present investigation, we detected that miR160a was up-regulated, while its 420
target gene Os04g43910 (ARF16) was down-regulated in the resistant line, but both 421
miRNA and its target were not much changed in the susceptible line upon M. oryzae 422
infection, implying that miR160a may positively regulate rice immunity by silencing IAA 423
signaling (Table 1, and Fig. 3). Then we obtained transgenic rice lines over-expressing 424
miR160a that exhibited enhanced resistance to the blast disease. RNA blotting and 425
qRT-PCR analyses verified that miR160a was highly expressed in contrast to the 426
significantly suppressed expression of its three target genes (i.e. Os02g41800, Os04g43910 427
and Os04g59430) (Fig. 4). Nevertheless, the expression of Os10g33940 and Os06g47150, 428
two targets of miR160a, were significantly up-regulated in the transgenic lines 429
over-expressing miR160a (Supplemental Fig. S4). One explanation is that miR160a may 430
repress their translation. This possibility could be easily confirmed if their antibody were 431
available. Alternatively, these two target genes of miR160a may be under feedback 432
regulation by their products as the case for the targets of miR172 in Ariabidopsis (Schwab 433
et al., 2005). It seems that miR160a may act in post-invasive defense responses, because the 434
expression of the PTI-related genes OsKS4 and OsNAC4 were not induced in the 435
miR160a-overexpression line upon M. oryzae infection or after PAMP (flg22 and chitin) 436
treatment (Fig. 6). However, miR160a acts in regulation of PTI and PAMP-triggered 437
defense responses in Arabidopsis (Li et al., 2010). Therefore, the functions of miR160a 438
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

18
seem to be conserved with diversification in plant innate immunity between monocots and 439
dicots. 440
Our data also revealed that the expression of miR398b was induced upon M. oryzae 441
infection in both the susceptible and resistant lines, while its target genes Os07g46990 442
(SOD2) and Os04g48410 (encoding a copper chaperone for SOD) were down-regulated 443
(Table 1, Fig. 3). By transgenic approach, we identified two transgenic lines 444
over-expressing miR398b. Both transgenic lines exhibited resistance to the blast fungus as 445
revealed by weaker disease phenotypes, less fungal mass, up-regulated expression of 446
defense-related genes, and stronger H2O2 production (Fig. 5, 6, 7 and Supplemental Fig. 447
S5). The two PTI-related genes OsKS4 and OsNAC4 as well as OsPR1 and OsPR10 were 448
significantly induced in the miR398b-overexpression transgenic line upon M. oryzae 449
infection and after PAMP treatments, indicating that miR398b might act in positively 450
regulation of both pre- and post-invasive defense responses against the infection of the blast 451
fungus. The cellular responses of the miR398b overexpression line to an eGFP-tagged 452
strain showed that both conidial germination and appressorium formation were largely 453
delayed, thus substantiated the roles of miR398b in positive regulation of pre-invasive 454
defense (Fig. 7). However, from literatures, miR398b seems to negatively regulate PTI and 455
ETI responses against bacterial pathogen, because the accumulation of miR398b is slightly 456
down-regulated after flg22 treatment in Arabidopsis (Li et al., 2010). The abundance of 457
miR398b can also be down-regulated by the infection of the incompatible bacteria Pst 458
DC3000 (avrRpm1 and/or avrRpt2), but not the compatible strain Pst DC3000 459
(Jagadeeswaran et al., 2009). Moreover, transgenic lines overexpressing miR398b leads to 460
repression of flg22- and hrcC--induced callose deposition and hyper-sensitivity to the 461
virulent strain DC3000 and the avirulent strain DC3000 hrcC- (Li et al., 2010). Thus, 462
miR398b may act contrarily between monocots and dicots in the regulation of innate 463
immunity. 464
The findings that miR160a and miR398b positively regulate rice immunity against M. 465
oryzae raise interesting questions concerning the potential roles of their target genes in 466
resistance. Auxin is regarded as a negative regulator for plant resistance (Navarro et al., 467
2006; Fu et al., 2011). ARF proteins are key factors in auxin signal pathway. They bind to 468
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

19
auxin-responsive elements (AuxREs) in promoters to regulate transcription of primary 469
auxin-response genes (Hagen and Guilfoyle, 2002). In Arabidopsis, miR393 specifically 470
targets TIR/AFB transcripts to silence auxin signaling pathway and enhance plant innate 471
immunity against bacterial pathogen (Navarro et al., 2006). Overexpression of miR160a 472
suppresses the expression of ARF16 and enhances flg22- and nonpathogenic bacteria 473
DC3000 (hrcC-)-induced callose deposition (Li et al., 2010). Rice gene GH3-2 encodes an 474
indole-3-acetic acid (IAA)-amido synthetase, which inactivates IAA signaling by 475
catalyzing the formation of an IAA-amino acid conjugation to suppress pathogen-induced 476
IAA accumulation, thus contribute to broad-spectrum resistance against both bacterial and 477
fungal pathogens in rice (Fu et al., 2011). In this study, we demonstrated that miR160a also 478
targets putative ARF16 in rice and contributes to resistance against M. oryzae, suggesting 479
the conservative role of miR160 in plant innate immunity by silencing auxin pathway genes 480
in both monocots and dicots. 481
SODs are a group of metalloenzymes converting superoxide anion (O2−) into hydrogen 482
peroxide and oxygen molecules to protect plant from damages caused by free radical 483
species (Beyer et al., 1991). Overexpressing tobacco Mn-SOD in alfalfa (Medicago sativa 484
L.) enhanced plant tolerance to freezing stress, suggesting a protective role in minimizing 485
oxygen free radical production after freezing stress (McKersie et al., 1993). 486
Down-regulation of miR398 by oxidative stresses leads to higher transcription of CSD1 and 487
CSD2, and enhances oxidative stress tolerance in Arabidopsis (Sunkar et al., 2006). 488
Conversely, miR398 level is up-regulated by sucrose, resulting in decreased CSD1 and 489
CSD2 mRNA and protein accumulation (Dugas and Bartel, 2008). Here, we showed that 490
miR398b accumulation is significantly increased at 24 hpi in the susceptible line LTH and 491
increased at 12 hpi in the resistant line IRBLkm-Ts upon M. oryzae infection (Table 1 and 492
Fig. 3). Three genes, i.e. Os03g22810, Os07g46990 and Os04g48410 that encode SOD, 493
SOD2 and copper chaperone for SOD, respectively, were confirmed as the targets of 494
miR398b by degradome sequencing or RNA ligase-mediated 5’ rapid amplification of 495
cDNA ends (RLM 5’-RACE) (Wu et al., 2009; Li et al., 2010; Zhou et al., 2010). 496
Os11g09780 was also predicted as a target of miR398b and encodes a hypothetical protein 497
(Wu et al., 2009). However, we were not successful in confirming whether the mRNA of 498
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

20
Os11g09780 was cleaved at the predicted target site via RLM 5’-RACE. It is highly 499
feasible that the cleaved mRNA of Os11g09780 is degraded quickly after its cleavage or 500
difficult to be detected because the target site is close to the 3’ end of the cDNA. Consistent 501
with this speculation, Os11g09780 was not detected in previous degradome analyses (Wu et 502
al., 2009; Li et al., 2010; Zhou et al., 2010). Alternatively, Os11g09780 may not be truly the 503
target of miR398b. Nevertheless, consistent with the up-regulated miR398b levels, its target 504
genes were down-regulated, and the transgenic plants overexpressing miR398b displays 505
significantly lower SOD/SOD2 transcripts and higher resistance to M. oryzae, indicating 506
that miR398b may contribute to ROS production by suppressing SOD level in cells to 507
counter-attack the invading blast fungus. It is possible that miR398b guides the degradation 508
of SOD transcripts to keep dynamic balance of superoxide anion and hydrogen peroxide 509
metabolism during M. oryzae infection. Thus future work will focus on functional 510
investigation of the target genes of miRNAs and functional identification of the 511
components of the regulation network. 512
Thus, by deep-sequencing of small RNA libraries constructed from samples of 513
susceptible and resistant lines, we identified a set of known miRNAs that acts either 514
positively or negatively in regulation of rice immunity against the blast fungus. By 515
transgenic approach, we further demonstrated that ectopic expression of a single miRNA is 516
able to establish rice resistance to the blast disease. 517
518
MATERIALS AND METHODS 519
Plant Materials and Growth Conditions 520
Rice susceptible line LTH and its resistant monogenic lines IRBLkm-Ts and IRBLz5-CA 521
were grown in a growth room maintained at 26℃ and 70% relative humidity with a 14/10 522
h of day/night light. The indica accession Kasalath (Oryza sativa ssp indica) was used for 523
transgenic analysis. 524
525
Library Construction, Sequencing and Bioinformatics Analysis of Small RNAs 526
Small RNA library construction and Illumina sequencing were performed as described (Mi 527
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

21
et al., 2008). The small RNA reads with length over 16 were mapped to the O. sativa 528
nuclear, chloroplast, and mitochondrial genomes (http://rice.plantbiology.msu.edu/, version 529
6.0) and M. oryzae genomes (http://www.broadinstitute.org). The perfect genome-matched 530
sRNAs were analyzed following a previous report (Wu et al., 2009). The normalized 531
abundance of small RNAs were calculated as reads per million. 532
533
Small RNA Gel-Blot Analysis 534
RNA-blot analyses for small RNAs from total extracts were performed as previously 535
described (Qi et al., 2005; Mi et al., 2008). Three-leaf-old plants spraying-inoculated with 5 536
×105 spores/mL was used for RNA extraction at 0, 12, 24, 48, and 72 hpi, respectively. 537
MiRNA probes were end labeled with γ-32P-ATP by T4 polynucleotide kinase (NEB). 538
Probes used for RNA-blot were listed in Supplemental Table S4. 539
540
Construction of miRNA Overexpression Transgenic Lines 541
To make over-expression constructs, genomic sequences from 330 bp upstream to 242 bp 542
downstream of miR160a and from 289 bp upstream to 306 bp downstream of miR398b 543
were amplified from Nipponbare (NPB) genomic DNA. PCR products were cloned into the 544
binary vector 35S-pCAMBIA1300 at KpnI/SalI sites. Primers used for making constructs 545
were listed in Supplemental Table S5. Constructs were transformed into Kasalath via 546
Agrobacterium-mediated transformation. Transgenic plants were preliminarily screened by 547
PCR with the forward primer 35S-F and the specific reverse primers (OsmiR160a-R-Sal1 548
for miR160a and OsmiR398b-R-Sal1 for miR398b, respectively) ( Supplemental Table S5). 549
Then, the obtained transformants were subjected to hygromycin-sensitive test as follows. 550
Leaves from 3-4 week old plants were incubated in 0.1 mM 6-BA buffer containing 30 551
mg/L hygromycin for 3-5 days at 26℃ and 70% relative humidity with a 14/10 hours of 552
day/night light. The plants whose leaves showed hygromycin-resistant were selected as 553
positive transgenic plants for further experiments. 554
555
Quantitative RT-PCR 556
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

22
Three-leaf-old plants were inoculated with M. oryzae by spraying spore suspension at the 557
concentration of 5 ×105 spores/mL and samples were collected at 0, 6, 12, 24, and 48 hpi. 558
For PAMP triggered responses, three-leaf-old plants were incubated in water with or 559
without 1 μM of flg22 or 1 μg/ml of chitin for 1 hour, then the treated leaves were 560
subjected to RNA extraction with Trizl reagent (Invitrogen). Reverse transcription of RNA 561
to cDNA was conducted by using the Super-Script first-strand synthesis system 562
(Invitrogen). miRNAs were elongated and reverse transcribed by using NCodeTM miRNA 563
First-strand cDNA module (Invitrogen). SYBR Green Mix (TaKaRa) was used in real-time 564
PCR to determine the abundance of mRNA. Gene expression levels were normalized by 565
using OsActin1 as an internal control. Primers used in real-time RT-PCR were list in 566
Supplemental Table S5. 567
568
Pathogen Infection and Microscopy Analysis 569
M. oryzae strains used in this study (including B9, B15, DJ-15, D35-7, E5, LJ5-4, LJ9-5, 570
LJ15-8, NC-10, NJ-10, OJ-15, U-24, WJ-10, YC-5, and YC-7) were collected from rice 571
fields in Ya’an (E103.0, N29.98, southwest of Sichuan Province, China). The M. oryzae 572
strains were grown on complete medium (CM) with agar for 2 weeks at 28°C with 12/12 h 573
day/night light for sporulation. Inoculum concentration of each strain was adjusted to 5 574
×105 spores/mL, and the same volume of spore suspension of the 15 strains were mixed for 575
spraying inoculation (Qu et al., 2006). Three-week-old seedlings (for small RNA library 576
construction) or eight-week-old plants (for resistant evaluation of transgenic lines) were 577
prepared for spraying inoculation, and disease symptom was observed at 10 dpi. The lesion 578
types were scored from 0 (resistant) to 5 (susceptible) using a standard reference scale 579
(Mackill and Bonman, 1992). The blast sporulation rate on the lesions and the fungal mass 580
in infected rice leaves were measured as described (Park et al., 2012). 581
For cellular response in rice to M. oryzae infection, the eGFP-tagged M. oryzae strain 582
Zhong-8-10-14 was inoculated on 5-cm-long leaf sheath as described (Kankanala et al., 583
2007). The inoculated epidermal layer was excised and analyzed under Laser Scanning 584
Confocal Microscope (Nikon A1, Japan) at 12, 24 and 36 hpi, respectively. 585
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

23
H2O2 accumulation and fungal structure were stained by 3,3’-diaminobenzidine (DAB) 586
and trypan-blue, respectively, referring to a previous report (Xiao et al., 2003). Leaf 587
sections were placed in 1 mg/mL of DAB (Sigma), and incubated at 22℃ for 8 h at 588
illumination. The DAB-stained leaves were double stained with trypan-blue and observed 589
under microscope (Zeiss imager A2, Germany). 590
591
Supplemental data 592
Supplemental Table S1. Category of small RNAs before and after M. oryzae infection. 593
Supplemental Table S2. Normalized reads of rice miRNAs before and after M. oryzae 594
infection. 595
Supplemental Table S3. miRNA profiles assayed by STEM analysis. 596
Supplemental Table S4. List of miRNA targets assayed by RT-PCR. 597
Supplemental Table S5. Primers and probes used in this study. 598
Supplemental Figure S1. Fungal growth on susceptible and resistant lines. Representative 599
leaf section from LTH (A), IRBLkm-Ts (B), and IRBLz5-CA (C), stained by 600
3,3’-diaminobenzidine and trypan blue, respectively, to show the accumulation of H2O2 601
(reddish brown) and fungal mass (blue) at 10 days past inoculation of M. oryzae. Bars, 50 602
μm. 603
Supplemental Figure S2. Patterns of miRNA expression upon M. oryzae infection. The 604
expressions of miRNAs at 0, 12, 24 hpi in LTH and IRBLkm-Ts were categorized into 39 605
patterns according to their changes before and after M. oryzae infection by Short 606
Time-series Expression Miner (STEM) analysis. The colored profiles had a statistically 607
significant number of genes assigned. The number at the top left corner of a box indicates 608
the pattern number. For each pattern, the number of miRNAs assigned to the pattern was 609
showed at the lower left corner of the box. 610
Supplemental Figure S3. Expression of miRNA target genes upon M. oryzae infection. 611
Quantitative RT-PCR analyses of mRNA levels for genes targeted by predicted positive (A), 612
negative (B), and basal responsive (C) miRNAs. Error bars indicate SD. Student’s t test was 613
used to determine the significance of difference between 0 hpi and the indicated time points 614
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

24
in the indicated lines. Asterisks (* and **) indicate significant difference at a P value <0.05 615
and <0.01, respectively. The experiments were repeated two times with similar results. 616
Supplemental Figure S4. Quantitative RT-PCR assay for the indicated two target genes of 617
miR160a in the indicated transgenic lines. 618
Supplemental Figure S5. Fungal growth on transgenic lines. Representative leaf sections 619
stained by 3,3’-diaminobenzidine and trypan blue, respectively, to show the accumulation 620
of H2O2 (reddish brown) and fungal mass (blue) at 10 dpi of M. oryzae on the transgenic 621
plants expressing empty vector (A), 35S:miR160a (B), and 35S:miR398b (C). Bars, 50 μm. 622
623
ACKNOWLEDGMENTS 624
We thank Dr. Cai-Lin Lei (Institute of Crop Science, Chinese Academy of Agricultural 625
Sciences) for providing the monogenic resistant lines IRBLkm-Ts and IRBLz5-CA, Mr. 626
Yang Lei for help with Laser Scanning Confocal Microscopy, Ting-Ting Wang, Jin-Long 627
Sun and Liang Li for technique supports, Professor Li-Huang Zhu for providing the 628
eGFP-tagged strain and suggestions on writing the manuscript, and Drs Xue-Wei Chen, 629
Min He, and Wei-Tao Li for advices during experiments. 630
631
REFERENCES 632
Alfano JR, Collmer A (2004) Type III secretion system effector proteins: double agents in bacterial disease 633
and plant defense. Annu Rev Phytopathol 42: 385-414 634
Ashikawa I, Hayashi N, Yamane H, Kanamori H, Wu J, Matsumoto T, Ono K, Yano M (2008) Two 635
adjacent nucleotide-binding site-leucine-rich repeat class genes are required to confer Pikm-specific 636
rice blast resistance. Genetics 180: 2267-2276 637
Baulcombe D (2004) RNA silencing in plants. Nature 431: 356-363 638
Berruyer R, Poussier S, Kankanala P, Mosquera G, Valent B (2006) Quantitative and qualitative influence 639
of inoculation methods on in planta growth of rice blast fungus. Phytopathology 96: 346-355 640
Beyer W, Imlay J, Fridovich I (1991) Superoxide dismutases. Prog Nucleic Acid Res Mol Biol 40: 221-253 641
Boller T, He SY (2009) Innate immunity in plants: an arms race between pattern recognition receptors in 642
plants and effectors in microbial pathogens. Science 324: 742-744 643
Campo S, Peris-Peris C, Sire C, Moreno AB, Donaire L, Zytnicki M, Notredame C, Llave C, Segundo 644
BS (2013) Identification of a novel microRNA (miRNA) from rice that targets an alternatively 645
spliced transcript of the Nramp6 (Natural resistance-associated macrophage protein 6) gene involved 646
in pathogen resistance. New Phytol 199: 212-217 647
Chen X (2012) Small RNAs in development - insights from plants. Curr Opin Genet Dev 22: 361-367 648
Chisholm ST, Coaker G, Day B, Staskawicz BJ (2006) Host-microbe interactions: shaping the evolution of 649
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

25
the plant immune response. Cell 124: 803-814 650
Dou D, Zhou JM (2012) Phytopathogen effectors subverting host immunity: different foes, similar 651
battleground. Cell Host Microbe 12: 484-495 652
Du P, Wu J, Zhang J, Zhao S, Zheng H, Gao G, Wei L, Li Y (2011) Viral infection induces expression of 653
novel phased microRNAs from conserved cellular microRNA precursors. PLoS Pathog 7: e1002176 654
Dugas DV, Bartel B (2008) Sucrose induction of Arabidopsis miR398 represses two Cu/Zn superoxide 655
dismutases. Plant Mol Biol 67: 403-417 656
Ernst J, Bar-Joseph Z (2006) STEM: a tool for the analysis of short time series gene expression data. BMC 657
Bioinformatics 7: 191 658
Fahlgren N, Howell MD, Kasschau KD, Chapman EJ, Sullivan CM, Cumbie JS, Givan SA, Law TF, 659
Grant SR, Dangl JL, Carrington JC (2007) High-throughput sequencing of Arabidopsis 660
microRNAs: evidence for frequent birth and death of MIRNA genes. PLoS One 2: e219 661
Fu J, Liu H, Li Y, Yu H, Li X, Xiao J, Wang S (2011) Manipulating broad-spectrum disease resistance by 662
suppressing pathogen-induced auxin accumulation in rice. Plant Physiol 155: 589-602 663
Fukuoka S, Saka N, Koga H, Ono K, Shimizu T, Ebana K, Hayashi N, Takahashi A, Hirochika H, 664
Okuno K, Yano M (2009) Loss of function of a proline-containing protein confers durable disease 665
resistance in rice. Science 325: 998-1001 666
Guo N, Ye WW, Wu XL, Shen DY, Wang YC, Xing H, Dou DL (2011) Microarray profiling reveals 667
microRNAs involving soybean resistance to Phytophthora sojae. Genome 54: 954-958 668
Hagen G, Guilfoyle T (2002) Auxin-responsive gene expression: genes, promoters and regulatory factors. 669
Plant Mol Biol 49: 373-385 670
Jagadeeswaran G, Saini A, Sunkar R (2009) Biotic and abiotic stress down-regulate miR398 expression in 671
Arabidopsis. Planta 229: 1009-1014 672
Jeong DH, Park S, Zhai J, Gurazada SG, De Paoli E, Meyers BC, Green PJ (2011) Massive analysis of 673
rice small RNAs: mechanistic implications of regulated microRNAs and variants for differential 674
target RNA cleavage. Plant Cell 23: 4185-4207 675
Jones-Rhoades MW, Bartel DP (2004) Computational identification of plant microRNAs and their targets, 676
including a stress-induced miRNA. Mol Cell 14: 787-799 677
Jones JD, Dangl JL (2006) The plant immune system. Nature 444: 323-329 678
Kaku H, Nishizawa Y, Ishii-Minami N, Akimoto-Tomiyama C, Dohmae N, Takio K, Minami E, Shibuya 679
N (2006) Plant cells recognize chitin fragments for defense signaling through a plasma membrane 680
receptor. Proc Natl Acad Sci U S A 103: 11086-11091 681
Kankanala P, Czymmek K, Valent B (2007) Roles for rice membrane dynamics and plasmodesmata during 682
biotrophic invasion by the blast fungus. Plant Cell 19: 706-724 683
Katiyar-Agarwal S, Jin H (2010) Role of small RNAs in host-microbe interactions. Annu Rev Phytopathol 684
48: 225-246 685
Khang CH, Berruyer R, Giraldo MC, Kankanala P, Park SY, Czymmek K, Kang S, Valent B (2010) 686
Translocation of Magnaporthe oryzae effectors into rice cells and their subsequent cell-to-cell 687
movement. Plant Cell 22: 1388-1403 688
Khraiwesh B, Zhu JK, Zhu J (2012) Role of miRNAs and siRNAs in biotic and abiotic stress responses of 689
plants. Biochim Biophys Acta 1819: 137-148 690
Kishimoto K, Kouzai Y, Kaku H, Shibuya N, Minami E, Nishizawa Y (2010) Perception of the chitin 691
oligosaccharides contributes to disease resistance to blast fungus Magnaporthe oryzae in rice. Plant J 692
64: 343-354 693
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

26
Kozomara A, Griffiths-Jones S (2011) miRBase: integrating microRNA annotation and deep-sequencing 694
data. Nucleic Acids Res 39: D152-157 695
Levine A, Tenhaken R, Dixon R, Lamb C (1994) H2O2 from the oxidative burst orchestrates the plant 696
hypersensitive disease resistance response. Cell 79: 583-593 697
Li F, Pignatta D, Bendix C, Brunkard JO, Cohn MM, Tung J, Sun H, Kumar P, Baker B (2012) 698
MicroRNA regulation of plant innate immune receptors. Proc Natl Acad Sci U S A 109: 1790-1795 699
Li T, Li H, Zhang YX, Liu JY (2011) Identification and analysis of seven H(2)O(2)-responsive miRNAs and 700
32 new miRNAs in the seedlings of rice (Oryza sativa L. ssp. indica). Nucleic Acids Res 39: 701
2821-2833 702
Li Y, Wang L, Jing JX, Li ZQ, Lin F, Huang LF, Pan QH (2007) The Pikm gene, conferring stable 703
resistance to isolates of Magnaporthe oryzae, was finely mapped in a crossover-cold region on rice 704
chromosome 11. Mol Breeding 20: 179-188 705
Li Y, Zhang Q, Zhang J, Wu L, Qi Y, Zhou JM (2010) Identification of microRNAs involved in 706
pathogen-associated molecular pattern-triggered plant innate immunity. Plant Physiol 152: 707
2222-2231 708
Li YF, Zheng Y, Addo-Quaye C, Zhang L, Saini A, Jagadeeswaran G, Axtell MJ, Zhang W, Sunkar R 709
(2010) Transcriptome-wide identification of microRNA targets in rice. Plant J 62: 742-759 710
Lin ZZ, Jiang WW, Wang JL, Lei CL (2001) Research and Utilization of Universally Susceptible Property 711
of Japonica Rice Variety Lijiangxintuanheigu. Scientia Agricultura Sinjca 34: 116-117 712
Liu B, Li P, Li X, Liu C, Cao S, Chu C, Cao X (2005) Loss of function of OsDCL1 affects microRNA 713
accumulation and causes developmental defects in rice. Plant Physiol 139: 296-305 714
Liu J, Wang X, Mitchell T, Hu Y, Liu X, Dai L, Wang GL (2010) Recent progress and understanding of the 715
molecular mechanisms of the rice-Magnaporthe oryzae interaction. Mol Plant Pathol 11: 419-427 716
Liu W, Liu J, Ning Y, Ding B, Wang X, Wang Z, Wang GL (2013) Recent Progress in Understanding 717
PAMP- and Effector-Triggered Immunity Against the Rice Blast Fungus Magnaporthe oryzae. Mol 718
Plant 719
Lu S, Sun YH, Amerson H, Chiang VL (2007) MicroRNAs in loblolly pine (Pinus taeda L.) and their 720
association with fusiform rust gall development. Plant J 51: 1077-1098 721
Luo YC, Zhou H, Li Y, Chen JY, Yang JH, Chen YQ, Qu LH (2006) Rice embryogenic calli express a 722
unique set of microRNAs, suggesting regulatory roles of microRNAs in plant post-embryogenic 723
development. FEBS Lett 580: 5111-5116 724
Mackill DJ, Bonman JB (1992) Inheritance of Blast Resistance in Near-Isogenic Lines of Rice. 725
Phytopathology 82: 746-749 726
McKersie BD, Chen Y, de Beus M, Bowley SR, Bowler C, Inze D, D'Halluin K, Botterman J (1993) 727
Superoxide dismutase enhances tolerance of freezing stress in transgenic alfalfa (Medicago sativa L.). 728
Plant Physiol 103: 1155-1163 729
Mentlak TA, Kombrink A, Shinya T, Ryder LS, Otomo I, Saitoh H, Terauchi R, Nishizawa Y, Shibuya 730
N, Thomma BP, Talbot NJ (2012) Effector-mediated suppression of chitin-triggered immunity by 731
magnaporthe oryzae is necessary for rice blast disease. Plant Cell 24: 322-335 732
Mi S, Cai T, Hu Y, Chen Y, Hodges E, Ni F, Wu L, Li S, Zhou H, Long C, Chen S, Hannon GJ, Qi Y 733
(2008) Sorting of small RNAs into Arabidopsis argonaute complexes is directed by the 5' terminal 734
nucleotide. Cell 133: 116-127 735
Mosquera G, Giraldo MC, Khang CH, Coughlan S, Valent B (2009) Interaction transcriptome analysis 736
identifies Magnaporthe oryzae BAS1-4 as Biotrophy-associated secreted proteins in rice blast 737
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

27
disease. Plant Cell 21: 1273-1290 738
Nagasaki H, Itoh J, Hayashi K, Hibara K, Satoh-Nagasawa N, Nosaka M, Mukouhata M, Ashikari M, 739
Kitano H, Matsuoka M, Nagato Y, Sato Y (2007) The small interfering RNA production pathway 740
is required for shoot meristem initiation in rice. Proc Natl Acad Sci U S A 104: 14867-14871 741
Navarro L, Dunoyer P, Jay F, Arnold B, Dharmasiri N, Estelle M, Voinnet O, Jones JD (2006) A plant 742
miRNA contributes to antibacterial resistance by repressing auxin signaling. Science 312: 436-439 743
Padmanabhan C, Zhang X, Jin H (2009) Host small RNAs are big contributors to plant innate immunity. 744
Curr Opin Plant Biol 12: 465-472 745
Park CH, Chen S, Shirsekar G, Zhou B, Khang CH, Songkumarn P, Afzal AJ, Ning Y, Wang R, Bellizzi 746
M, Valent B, Wang GL (2012) The Magnaporthe oryzae Effector AvrPiz-t Targets the RING E3 747
Ubiquitin Ligase APIP6 to Suppress Pathogen-Associated Molecular Pattern-Triggered Immunity in 748
Rice. Plant Cell 24: 4748-4762 749
Qi Y, Denli AM, Hannon GJ (2005) Biochemical specialization within Arabidopsis RNA silencing pathways. 750
Mol Cell 19: 421-428 751
Qu S, Liu G, Zhou B, Bellizzi M, Zeng L, Dai L, Han B, Wang GL (2006) The broad-spectrum blast 752
resistance gene Pi9 encodes a nucleotide-binding site-leucine-rich repeat protein and is a member of 753
a multigene family in rice. Genetics 172: 1901-1914 754
Radwan O, Liu Y, Clough SJ (2011) Transcriptional analysis of soybean root response to Fusarium 755
virguliforme, the causal agent of sudden death syndrome. Mol Plant Microbe Interact 24: 958-972 756
Reinhart BJ, Weinstein EG, Rhoades MW, Bartel B, Bartel DP (2002) MicroRNAs in plants. Genes Dev 757
16: 1616-1626 758
Rhoades MW, Reinhart BJ, Lim LP, Burge CB, Bartel B, Bartel DP (2002) Prediction of plant microRNA 759
targets. Cell 110: 513-520 760
Ribot C, Hirsch J, Balzergue S, Tharreau D, Notteghem JL, Lebrun MH, Morel JB (2008) Susceptibility 761
of rice to the blast fungus, Magnaporthe grisea. J Plant Physiol 165: 114-124 762
Schwab R, Palatnik JF, Riester M, Schommer C, Schmid M, Weigel D (2005) Specific effects of 763
microRNAs on the plant transcriptome. Dev Cell 8: 517-527 764
Seo JK, Wu J, Lii Y, Li Y, Jin H (2013) Contribution of small RNA pathway components in plant immunity. 765
Mol Plant Microbe Interact 26: 617-625 766
Shimizu T, Nakano T, Takamizawa D, Desaki Y, Ishii-Minami N, Nishizawa Y, Minami E, Okada K, 767
Yamane H, Kaku H, Shibuya N (2010) Two LysM receptor molecules, CEBiP and OsCERK1, 768
cooperatively regulate chitin elicitor signaling in rice. Plant J 64: 204-214 769
Shivaprasad PV, Chen HM, Patel K, Bond DM, Santos BA, Baulcombe DC (2012) A microRNA 770
superfamily regulates nucleotide binding site-leucine-rich repeats and other mRNAs. Plant Cell 24: 771
859-874 772
Sunkar R, Girke T, Jain PK, Zhu JK (2005) Cloning and characterization of microRNAs from rice. Plant 773
Cell 17: 1397-1411 774
Sunkar R, Kapoor A, Zhu JK (2006) Posttranscriptional induction of two Cu/Zn superoxide dismutase 775
genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress 776
tolerance. Plant Cell 18: 2051-2065 777
Sunkar R, Zhou X, Zheng Y, Zhang W, Zhu JK (2008) Identification of novel and candidate miRNAs in 778
rice by high throughput sequencing. BMC Plant Biol 8: 25 779
Tsumematsu H YM, Ebron LA, Hayashi N, Ando I, Kato H, Imbe T, Khush GS (2000) Development of 780
monogenic lines of rice for blast resistance. Breeding science 50: 229-234 781
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

28
Wang JF, Zhou H, Chen YQ, Luo QJ, Qu LH (2004) Identification of 20 microRNAs from Oryza sativa. 782
Nucleic Acids Res 32: 1688-1695 783
Wu L, Zhang Q, Zhou H, Ni F, Wu X, Qi Y (2009) Rice MicroRNA effector complexes and targets. Plant 784
Cell 21: 3421-3435 785
Xiao S, Brown S, Patrick E, Brearley C, Turner JG (2003) Enhanced transcription of the Arabidopsis 786
disease resistance genes RPW8.1 and RPW8.2 via a salicylic acid-dependent amplification circuit is 787
required for hypersensitive cell death. Plant Cell 15: 33-45 788
Xie K, Wu C, Xiong L (2006) Genomic organization, differential expression, and interaction of 789
SQUAMOSA promoter-binding-like transcription factors and microRNA156 in rice. Plant Physiol 790
142: 280-293 791
Xin M, Wang Y, Yao Y, Xie C, Peng H, Ni Z, Sun Q (2010) Diverse set of microRNAs are responsive to 792
powdery mildew infection and heat stress in wheat (Triticum aestivum L.). BMC Plant Biol 10: 123 793
Xue LJ, Zhang JJ, Xue HW (2009) Characterization and expression profiles of miRNAs in rice seeds. 794
Nucleic Acids Res 37: 916-930 795
Yamaguchi K, Yamada K, Ishikawa K, Yoshimura S, Hayashi N, Uchihashi K, Ishihama N, 796
Kishi-Kaboshi M, Takahashi A, Tsuge S, Ochiai H, Tada Y, Shimamoto K, Yoshioka H, 797
Kawasaki T (2013) A Receptor-like Cytoplasmic Kinase Targeted by a Plant Pathogen Effector Is 798
Directly Phosphorylated by the Chitin Receptor and Mediates Rice Immunity. Cell Host Microbe 13: 799
347-357 800
Yang L, Jue D, Li W, Zhang R, Chen M, Yang Q (2013) Identification of MiRNA from Eggplant (Solanum 801
melongena L.) by Small RNA Deep Sequencing and Their Response to Verticillium dahliae Infection. 802
PLoS One 8: e72840 803
Yin Z, Li Y, Han X, Shen F (2012) Genome-wide profiling of miRNAs and other small non-coding RNAs in 804
the Verticillium dahliae-inoculated cotton roots. PLoS One 7: e35765 805
Yoshida K, Saitoh H, Fujisawa S, Kanzaki H, Matsumura H, Tosa Y, Chuma I, Takano Y, Win J, 806
Kamoun S, Terauchi R (2009) Association genetics reveals three novel avirulence genes from the 807
rice blast fungal pathogen Magnaporthe oryzae. Plant Cell 21: 1573-1591 808
Zhai J, Jeong DH, De Paoli E, Park S, Rosen BD, Li Y, Gonzalez AJ, Yan Z, Kitto SL, Grusak MA, 809
Jackson SA, Stacey G, Cook DR, Green PJ, Sherrier DJ, Meyers BC (2011) MicroRNAs as 810
master regulators of the plant NB-LRR defense gene family via the production of phased, 811
trans-acting siRNAs. Genes Dev 25: 2540-2553 812
Zhang W, Gao S, Zhou X, Chellappan P, Chen Z, Zhang X, Fromuth N, Coutino G, Coffey M, Jin H 813
(2011) Bacteria-responsive microRNAs regulate plant innate immunity by modulating plant 814
hormone networks. Plant Mol Biol 75: 93-105 815
Zhao CZ, Xia H, Frazier TP, Yao YY, Bi YP, Li AQ, Li MJ, Li CS, Zhang BH, Wang XJ (2010) Deep 816
sequencing identifies novel and conserved microRNAs in peanuts (Arachis hypogaea L.). BMC 817
Plant Biol 10: 3 818
Zhou B, Qu S, Liu G, Dolan M, Sakai H, Lu G, Bellizzi M, Wang GL (2006) The eight amino-acid 819
differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the 820
resistance specificity to Magnaporthe grisea. Mol Plant Microbe Interact 19: 1216-1228 821
Zhou L, Liu Y, Liu Z, Kong D, Duan M, Luo L (2010) Genome-wide identification and analysis of 822
drought-responsive microRNAs in Oryza sativa. J Exp Bot 61: 4157-4168 823
Zhou M, Gu L, Li P, Song X, Wei L, Chen Z, Cao X (2010) Degradome sequencing reveals endogenous 824
small RNA targets in rice (Oryza sativa L. ssp. indica). Frontiers in Biology 5: 67-90 825
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

29
Zhou M, Gu LF, Li PC, Song XW, Wei LY, Chen ZY, Cao XF (2010) Degradome sequencing reveals 826
endogenous small RNAtargets in rice (Oryza sativa L. ssp. indica). Front Biol 5: 67-90 827
Zhu QH, Spriggs A, Matthew L, Fan L, Kennedy G, Gubler F, Helliwell C (2008) A diverse set of 828
microRNAs and microRNA-like small RNAs in developing rice grains. Genome Res 18: 1456-1465 829
Zipfel C, Felix G (2005) Plants and animals: a different taste for microbes? Curr Opin Plant Biol 8: 353-360 830
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

30
Figure legends 831
Figure 1. Comparison of defense responses among the susceptible line LTH and two 832
monogenic resistant lines. Three-leaf-old seedlings were inoculated with spore suspension 833
(5×105 spores/mL) of M. oryzae and disease phenotypes were recorded at 10 days post 834
inoculation (dpi). A, Representative leaf sections from the indicated lines to show the blast 835
disease phenotypes. B-F, Expressions of the indicated defense-related genes in LTH, 836
IRBLkm-Ts, and IRBLz5-CA upon M. oryzae infection. RNA was extracted at the 837
indicated time points for quantitative RT-PCR analysis. mRNA level was normalized to that 838
in untreated LTH (0 hr). Error bars indicate SD. Student’s t test was carried out to 839
determine the significance of difference between LTH and IRBLkm-Ts or IRBLz5-CA. 840
Asterisks (**) indicate significant difference at a P value <0.01. The experiments were 841
repeated two times with similar results. G, Representative leaf sections from the indicated 842
lines to show fungal growth and H2O2 accumulation at 2 dpi and 10 dpi, respectively. Note 843
that there was no or trace amount of H2O2 accumulation in LTH (arrow), but high levels 844
around the appressoria (arrowheads) at 2 dpi in the leaf cells of both IRBLkm-Ts and 845
IRBLz5-CA. Trypan blue and 3,3’-diaminobenzidine were used to stain the fungal structure 846
and H2O2 accumulation (reddish brown), respectively. Bars, 10 μm. 847
Figure 2. Profiling of small RNAs by deep sequencing of the libraries from different 848
samples. Small RNA libraries were constructed from samples of the indicated lines 849
collected at 0, 12, and 24 hours post inoculation (dpi) of M. oryzae and subjected to 850
deep-sequencing. A, Size distribution of sequenced small RNAs mapped to rice genome. B, 851
Size distribution of miRNAs mapped to miRNA precursor. C, Bar charts summarizing the 852
percentage of small RNA reads matched to rice and M. oryzae genome, respectively. D, Bar 853
charts summarizing the percentage of small RNA reads matched to different rice RNAs. 854
Figure 3. The expression patterns of miRNAs and their target genes upon M. oryzae 855
infection. Three-leaf-old seedlings were inoculated with 5× 105 spores/mL of M. oryzae and 856
total RNA was extracted from leaves of the indicated lines at the indicated time points. A, 857
RNA blotting analysis of miRNAs. Fifteen micrograms of small RNA was loaded. RNA 858
blots were hybridized with DNA oligonucleotide probes complementary to the indicated 859
miRNAs. U6 was used as loading control. Values below each section represent relative 860
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

31
abundance of miRNA normalized to U6. B to G, Quantitative RT-PCR analyses of mRNA 861
levels for genes targeted by miR160 (B, Os04g43910/ARF16), miR164 (C, Os12g41680), 862
miR827 (D, Os04g48390), miR172 (E, Os05g03040), and miR398 (F and G, 863
Os07g46990/SOD2 and Os04g48410). Error bars indicate SD. Student’s t test was used to 864
determine the significance of difference between 0 hpi and the indicated time points in the 865
same line. Asterisks * and ** indicate significant difference at a P value <0.05 and <0.01, 866
respectively. The experiments were repeated two times with similar results. 867
Figure 4. Overexpression of miR160a enhances rice resistance to M. oryzae. A and B, RNA 868
blotting analysis (A) and quantitative RT-PCR (qRT-PCR) assay (B) to examine the 869
accumulation of miR160a in the indicated transgenic lines expressing 35S:miR160a or 870
empty vector (EV). One microgram of total RNA was used for qRT-PCR analysis and 871
fifteen micrograms of small RNA was loaded for RNA-blot analysis. C, qRT-PCR assay for 872
the indicated three target genes of miR160a in the indicated transgenic lines. RNA was 873
extracted from T2 generation of 35S:MIR160a transgenic plants. D, Representative leaf 874
sections from the indicated transgenic lines to show the disease phenotypes. Three-leaf-old 875
seedlings were inoculated with 5×105 spores/mL of M. oryzae and the phenotype was 876
observed at 10 dpi. E and F, Sporulation and relative fungal growth on the inoculated leaves 877
of the indicated transgenic lines. Samples were taken for the assays at 10 dpi. Values were 878
the means of three replications. Error bars indicate SD. Student’s t test was carried out to 879
determine the significance of difference between Kasalath (EV) and 880
miR160a-overexpression transgenic plants. Asterisks * and ** indicate significant 881
difference at a P value <0.05 and <0.01, respectively. The experiments were repeated two 882
times with similar results. 883
Figure 5. Overexpression of miR398b enhances rice resistance to M. oryzae. A and B, RNA 884
blotting analysis (A) and qRT-PCR assay (B) to examine the accumulation of miR398b in 885
the indicated transgenic lines expressing 35S:miR398b or empty vector (EV). One 886
microgram of total RNA was used for qRT-PCR analysis and twenty micrograms of small 887
RNA was loaded for RNA-blot analysis, respectively. C, qRT-PCR assay for the indicated 888
four target genes of miR398b in the indicated transgenic lines. D, Representative leaf 889
sections from the indicated transgenic lines to show the disease phenotypes. 890
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

32
Three-leaves-old transgenic rice seedlings were inoculated with 5×105 spores/mL of M. 891
oryzae and the phenotype was observed at 10 dpi. E and F, Quantitative assay for 892
sporulation (E) and relative fungal growth (F) on the inoculated leaves of the indicated 893
transgenic lines. Samples were collected at 10 dpi. Values were the means of three 894
replications. Error bars indicate SD. Student’s t test was carried out to determine the 895
significance of difference between Kasalath (EV) and miR398b-overexpression transgenic 896
plants. Asterisks * and ** indicate significant difference at a P value <0.05 and <0.01, 897
respectively. The experiments were repeated two times with similar results. 898
Figure 6. MiR160a and miR398b positively and differentially regulate expression of 899
defense-related genes. Three-leaf-old seedlings were inoculated with 5 × 105 spores/mL of 900
M.oryzae and samples were collected at the indicated time points. RNA was extracted for 901
quantitative RT-PCR (qRT-PCR) analysis and mRNA level was normalized to that in 902
untreated control Kasalath (EV). A and B, qRT-PCR assay showing that the late responsive 903
defense-related genes PR1 (A) and PR10 (B) were up-regulated in the miR160a and 904
miR398b over-expression lines. C and D, qRT-PCR assay showing that the PTI-related 905
genes OsKS4 and OsNAC4 were up-regulated in the transgenic line over-expressing 906
miR398b but not in that of miR160a. E and F, qRT-PCR assay showing the expression 907
levels of PTI-related genes OsKS4 and OsNAC4 in the indicated transgenic lines after 908
incubating in water, 1 μg/ml of chitin, or 1 μM of flg22 for 1 hour, respectively. Note that 909
OsKS4 and OsNAC4 were significantly induced by both chitin and flg22 in the transgenic 910
line over-expressing miR398b but not in that of miR160a. Values are the means of three 911
replications. Error bars indicate SD. Student’s t test was carried out to determine the 912
significance of difference between Kasalath (EV) and miRNA overexpression transgenic 913
plants following M. oryzae or PAMP treatment. Asterisks * and ** indicate significant 914
difference at a P value <0.05 and <0.01, respectively. The experiments repeated two times 915
with similar results. 916
Figure 7. Cellular responses to the infection of M. oryzae. A to C, Representative Laser 917
Scanning Confocal Microscopic images to show infection of the eGFP-tagged strain 918
Zhong-8-10-14 on sheath cells of transgenic plants expressing empty vector (A), 919
35S:miR160a (B), and 35S:miR398b (C) at the indicated time points. Note that appressoria 920
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

33
(arrows) were formed at 12 hpi on EV plants, but delayed to 24 hpi on 921
miR160a-overexpression plants and 36 hpi on miR398b-overexpression plants. Invasive 922
hyphae (arrowheads) were formed at 24 hpi and extended to the neighbor cells at 36 hpi on 923
EV plants. Bars, 20 μm. D and E, Representative leaf sections from the indicated transgenic 924
lines stained by 3,3’-diaminobenzidine and trypan blue at 2 dpi (D) and 10 dpi (E), 925
respectively, to show the accumulation of H2O2 (reddish brown) at the infection site where 926
appressoria (arrows) were visualized and the fungal structures. Note that secondary conidia 927
(arrowheads in E) were generated on EV plants at 10 dpi. Bars, 10 μm. F and G, Statistic 928
analyses of conidial germination rate (F) and frequency of invasive hypha (G) at the 929
indicated time points observed on the indicated lines. Inoculated leaves were stained with 930
trypan blue and examined by microscopy. Data are mean±SEM from three independent 931
experiments each of which >200 conidia were evaluated. x2-test was used to test statistical 932
significance between EV and transgenic lines. Asterisks * and ** indicate significant 933
difference at a P value <0.05 and <0.01, respectively. 934
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.
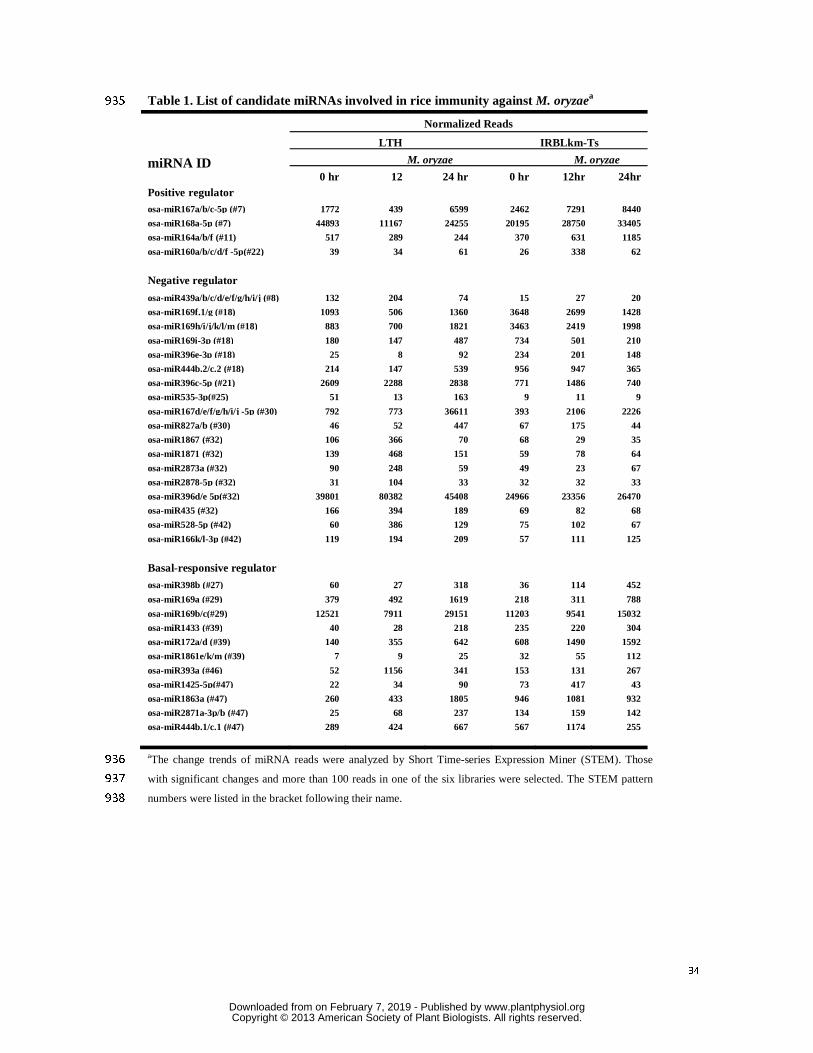
34
Table 1. List of candidate miRNAs involved in rice immunity against M. oryzaea 935
miRNA ID
Normalized Reads
LTH IRBLkm-Ts
M. oryzae M. oryzae
0 hr 12 24 hr 0 hr 12hr 24hr Positive regulator
osa-miR167a/b/c-5p (#7) 1772 439 6599 2462 7291 8440
osa-miR168a-5p (#7) 44893 11167 24255 20195 28750 33405
osa-miR164a/b/f (#11) 517 289 244 370 631 1185
osa-miR160a/b/c/d/f -5p(#22) 39 34 61 26 338 62
Negative regulator
osa-miR439a/b/c/d/e/f/g/h/i/j (#8) 132 204 74 15 27 20
osa-miR169f.1/g (#18) 1093 506 1360 3648 2699 1428
osa-miR169h/i/j/k/l/m (#18) 883 700 1821 3463 2419 1998
osa-miR169i-3p (#18) 180 147 487 734 501 210
osa-miR396e-3p (#18) 25 8 92 234 201 148
osa-miR444b.2/c.2 (#18) 214 147 539 956 947 365
osa-miR396c-5p (#21) 2609 2288 2838 771 1486 740
osa-miR535-3p(#25) 51 13 163 9 11 9
osa-miR167d/e/f/g/h/i/j -5p (#30) 792 773 36611 393 2106 2226
osa-miR827a/b (#30) 46 52 447 67 175 44
osa-miR1867 (#32) 106 366 70 68 29 35
osa-miR1871 (#32) 139 468 151 59 78 64
osa-miR2873a (#32) 90 248 59 49 23 67
osa-miR2878-5p (#32) 31 104 33 32 32 33
osa-miR396d/e 5p(#32) 39801 80382 45408 24966 23356 26470
osa-miR435 (#32) 166 394 189 69 82 68
osa-miR528-5p (#42) 60 386 129 75 102 67
osa-miR166k/l-3p (#42) 119 194 209 57 111 125
Basal-responsive regulator
osa-miR398b (#27) 60 27 318 36 114 452
osa-miR169a (#29) 379 492 1619 218 311 788
osa-miR169b/c(#29) 12521 7911 29151 11203 9541 15032
osa-miR1433 (#39) 40 28 218 235 220 304
osa-miR172a/d (#39) 140 355 642 608 1490 1592
osa-miR1861e/k/m (#39) 7 9 25 32 55 112
osa-miR393a (#46) 52 1156 341 153 131 267
osa-miR1425-5p(#47) 22 34 90 73 417 43
osa-miR1863a (#47) 260 433 1805 946 1081 932
osa-miR2871a-3p/b (#47) 25 68 237 134 159 142
osa-miR444b.1/c.1 (#47) 289 424 667 567 1174 255
aThe change trends of miRNA reads were analyzed by Short Time-series Expression Miner (STEM). Those 936
with significant changes and more than 100 reads in one of the six libraries were selected. The STEM pattern 937
numbers were listed in the bracket following their name. 938
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.
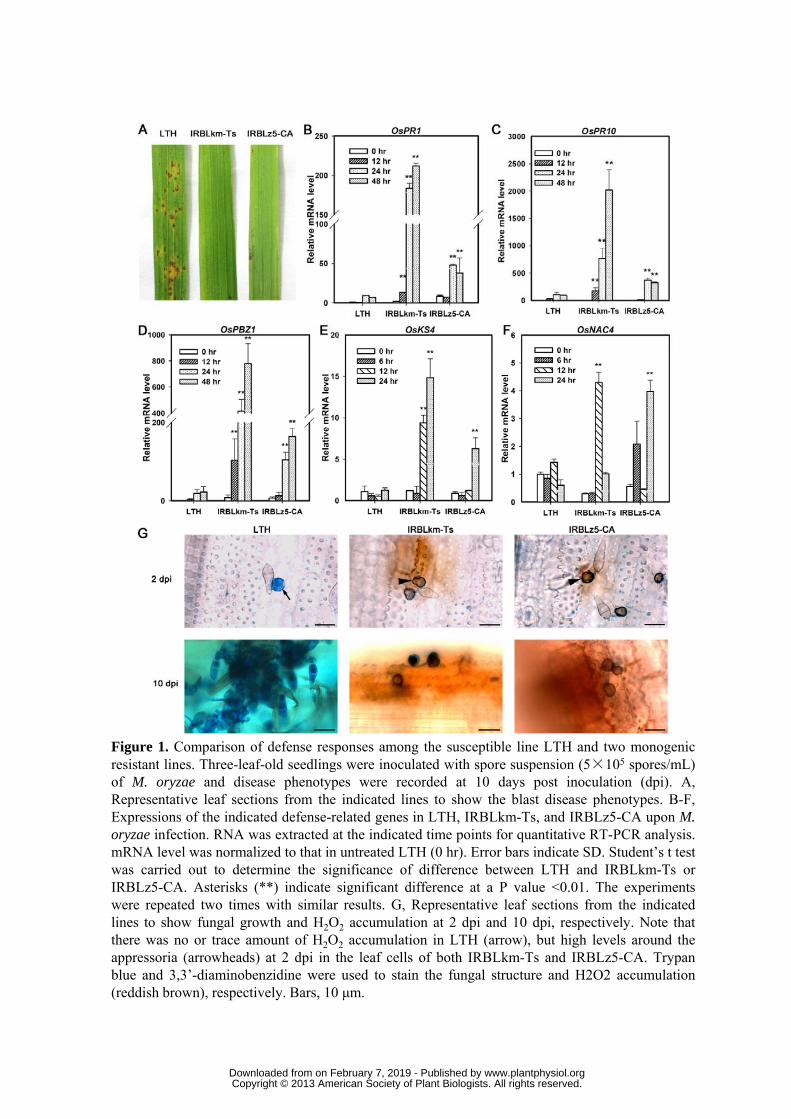
Figure 1. Comparison of defense responses among the susceptible line LTH and two monogenicresistant lines Three leaf old seedlings were inoculated with spore suspension (5×105 spores/mL)resistant lines. Three-leaf-old seedlings were inoculated with spore suspension (5×105 spores/mL)of M. oryzae and disease phenotypes were recorded at 10 days post inoculation (dpi). A,Representative leaf sections from the indicated lines to show the blast disease phenotypes. B-F,Expressions of the indicated defense-related genes in LTH, IRBLkm-Ts, and IRBLz5-CA upon M.oryzae infection. RNA was extracted at the indicated time points for quantitative RT-PCR analysis.mRNA level was normalized to that in untreated LTH (0 hr). Error bars indicate SD. Student’s t testwas carried out to determine the significance of difference between LTH and IRBLkm-Ts orIRBLz5-CA. Asterisks (**) indicate significant difference at a P value <0.01. The experiments
t d t ti ith i il lt G R t ti l f ti f th i di t dwere repeated two times with similar results. G, Representative leaf sections from the indicatedlines to show fungal growth and H2O2 accumulation at 2 dpi and 10 dpi, respectively. Note thatthere was no or trace amount of H2O2 accumulation in LTH (arrow), but high levels around theappressoria (arrowheads) at 2 dpi in the leaf cells of both IRBLkm-Ts and IRBLz5-CA. Trypanblue and 3,3’-diaminobenzidine were used to stain the fungal structure and H2O2 accumulation(reddish brown), respectively. Bars, 10 μm.
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

Figure 2. Profiling of small RNAs by deep sequencing of the libraries from differentsamples. Small RNA libraries were constructed from samples of the indicated linescollected at 0, 12, and 24 hours post inoculation (dpi) ofM. oryzae and subjected to deep-
i A Si di t ib ti f d ll RNA d t i Bsequencing. A, Size distribution of sequenced small RNAs mapped to rice genome. B,Size distribution of miRNAs mapped to miRNA precursor. C, Bar charts summarizingthe percentage of small RNA reads matched to rice and M. oryzae genome, respectively.D, Bar charts summarizing the percentage of small RNA reads matched to different riceRNAs.
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.
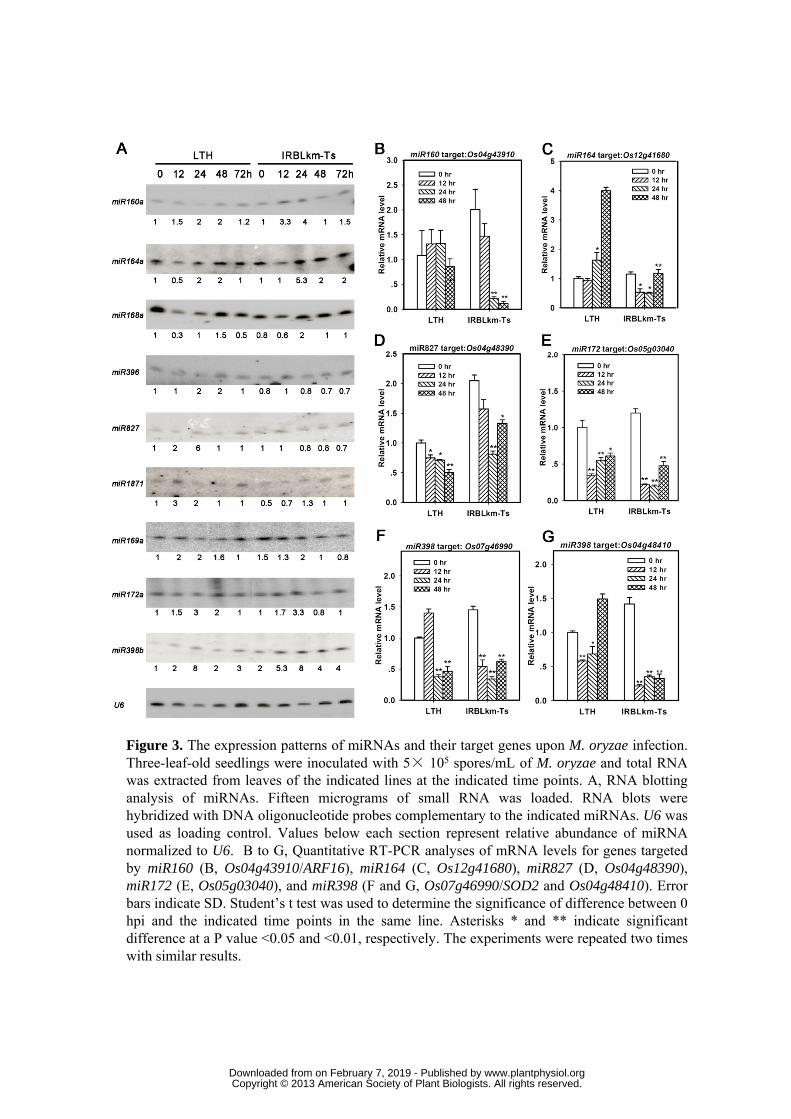
Figure 3. The expression patterns of miRNAs and their target genes upon M. oryzae infection.Three-leaf-old seedlings were inoculated with 5× 105 spores/mL of M oryzae and total RNAThree-leaf-old seedlings were inoculated with 5× 10 spores/mL of M. oryzae and total RNAwas extracted from leaves of the indicated lines at the indicated time points. A, RNA blottinganalysis of miRNAs. Fifteen micrograms of small RNA was loaded. RNA blots werehybridized with DNA oligonucleotide probes complementary to the indicated miRNAs. U6 wasused as loading control. Values below each section represent relative abundance of miRNAnormalized to U6. B to G, Quantitative RT-PCR analyses of mRNA levels for genes targetedby miR160 (B, Os04g43910/ARF16), miR164 (C, Os12g41680), miR827 (D, Os04g48390),miR172 (E, Os05g03040), and miR398 (F and G, Os07g46990/SOD2 and Os04g48410). Errorb i di t SD St d t’ t t t d t d t i th i ifi f diff b t 0bars indicate SD. Student’s t test was used to determine the significance of difference between 0hpi and the indicated time points in the same line. Asterisks * and ** indicate significantdifference at a P value <0.05 and <0.01, respectively. The experiments were repeated two timeswith similar results.
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.
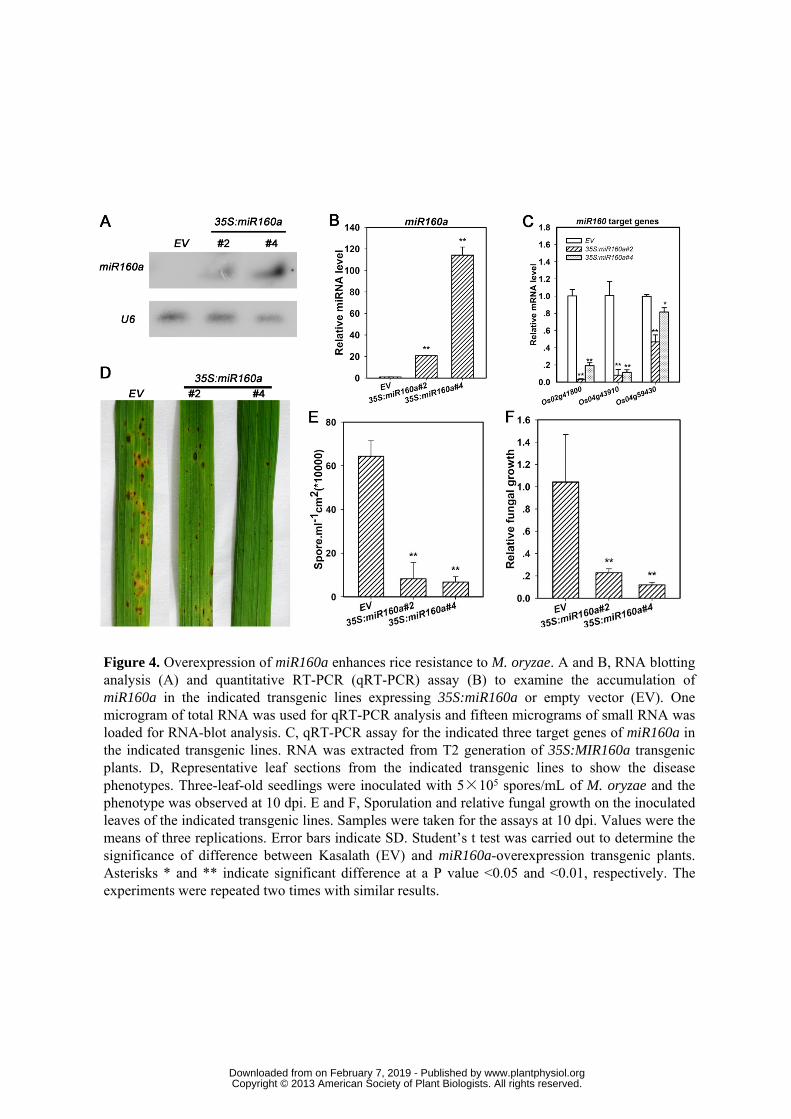
Figure 4. Overexpression of miR160a enhances rice resistance to M. oryzae. A and B, RNA blottinganalysis (A) and quantitative RT-PCR (qRT-PCR) assay (B) to examine the accumulation ofmiR160a in the indicated transgenic lines expressing 35S:miR160a or empty vector (EV). Onemicrogram of total RNA was used for qRT-PCR analysis and fifteen micrograms of small RNA wasloaded for RNA-blot analysis. C, qRT-PCR assay for the indicated three target genes of miR160a inthe indicated transgenic lines. RNA was extracted from T2 generation of 35S:MIR160a transgenicl t D R t ti l f ti f th i di t d t i li t h th diplants. D, Representative leaf sections from the indicated transgenic lines to show the diseasephenotypes. Three-leaf-old seedlings were inoculated with 5×105 spores/mL of M. oryzae and thephenotype was observed at 10 dpi. E and F, Sporulation and relative fungal growth on the inoculatedleaves of the indicated transgenic lines. Samples were taken for the assays at 10 dpi. Values were themeans of three replications. Error bars indicate SD. Student’s t test was carried out to determine thesignificance of difference between Kasalath (EV) and miR160a-overexpression transgenic plants.Asterisks * and ** indicate significant difference at a P value <0.05 and <0.01, respectively. Theexperiments were repeated two times with similar results.
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

Figure 5. Overexpression of miR398b enhances rice resistance to M. oryzae. A and B, RNA blottinganalysis (A) and qRT-PCR assay (B) to examine the accumulation of miR398b in the indicatedtransgenic lines expressing 35S:miR398b or empty vector (EV). One microgram of total RNA wasused for qRT-PCR analysis and twenty micrograms of small RNA was loaded for RNA-blot analysis,respectively C qRT-PCR assay for the indicated four target genes of miR398b in the indicatedrespectively. C, qRT-PCR assay for the indicated four target genes of miR398b in the indicatedtransgenic lines. D, Representative leaf sections from the indicated transgenic lines to show thedisease phenotypes. Three-leaves-old transgenic rice seedlings were inoculated with 5×105spores/mL of M. oryzae and the phenotype was observed at 10 dpi. E and F, Quantitative assay forsporulation (E) and relative fungal growth (F) on the inoculated leaves of the indicated transgeniclines. Samples were collected at 10 dpi. Values were the means of three replications. Error barsindicate SD. Student’s t test was carried out to determine the significance of difference betweenKasalath (EV) and miR398b-overexpression transgenic plants. Asterisks * and ** indicate significantdiff t P l <0 05 d <0 01 ti l Th i t t d t ti ithdifference at a P value <0.05 and <0.01, respectively. The experiments were repeated two times withsimilar results.
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.
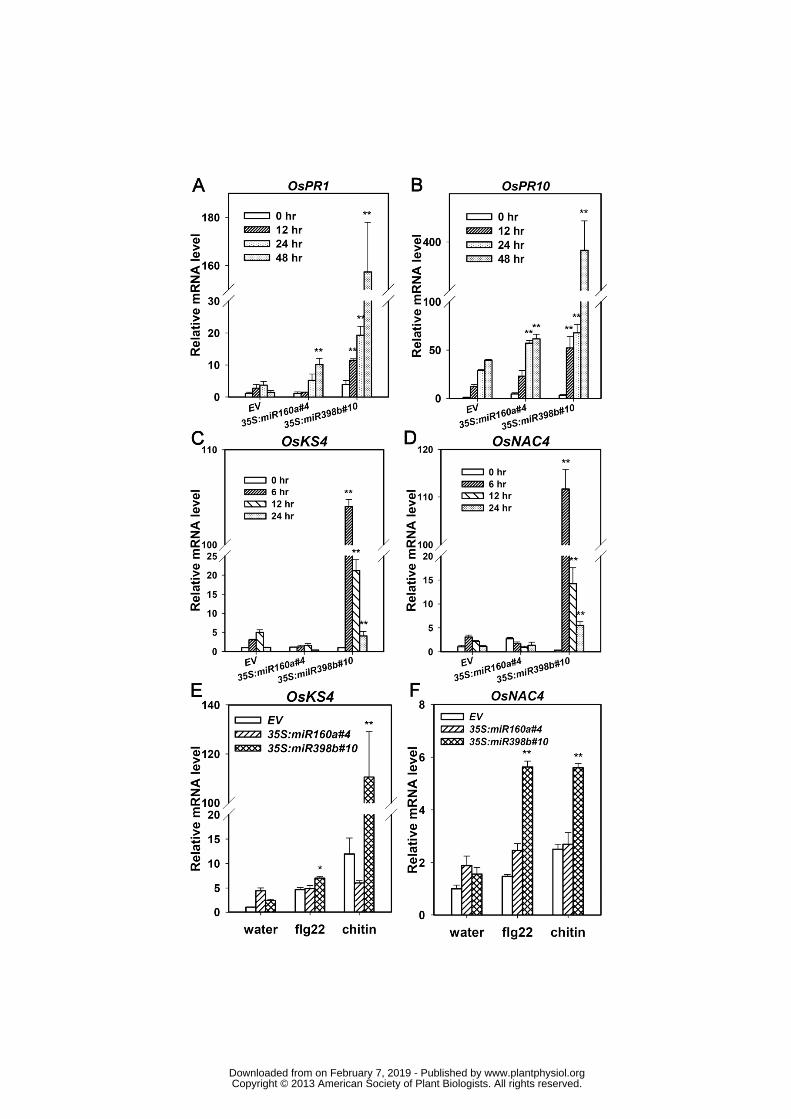
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

Figure 6. MiR160a and miR398b positively and differentially regulate expression ofdefense-related genes. Three-leaf-old seedlings were inoculated with 5 × 105 spores/mLof M oryzae and samples were collected at the indicated time points RNA was extractedof M.oryzae and samples were collected at the indicated time points. RNA was extractedfor quantitative RT-PCR (qRT-PCR) analysis and mRNA level was normalized to that inuntreated control Kasalath (EV). A and B, qRT-PCR assay showing that the lateresponsive defense-related genes PR1 (A) and PR10 (B) were up-regulated in the miR160aand miR398b over-expression lines. C and D, qRT-PCR assay showing that the PTI-related genes OsKS4 and OsNAC4 were up-regulated in the transgenic line over-expressing miR398b but not in that of miR160a. E and F, qRT-PCR assay showing theexpression levels of PTI-related genes OsKS4 and OsNAC4 in the indicated transgenicli ft i b ti i t 1 / l f hiti 1 M f fl 22 f 1 h ti llines after incubating in water, 1 μg/ml of chitin, or 1 μM of flg22 for 1 hour, respectively.Note that OsKS4 and OsNAC4 were significantly induced by both chitin and flg22 in thetransgenic line over-expressing miR398b but not in that of miR160a. Values are the meansof three replications. Error bars indicate SD. Student’s t test was carried out to determinethe significance of difference between Kasalath (EV) and miRNA overexpressiontransgenic plants following M. oryzae or PAMP treatment. Asterisks * and ** indicatesignificant difference at a P value <0.05 and <0.01, respectively. The experiments repeatedtwo times with similar results.
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.
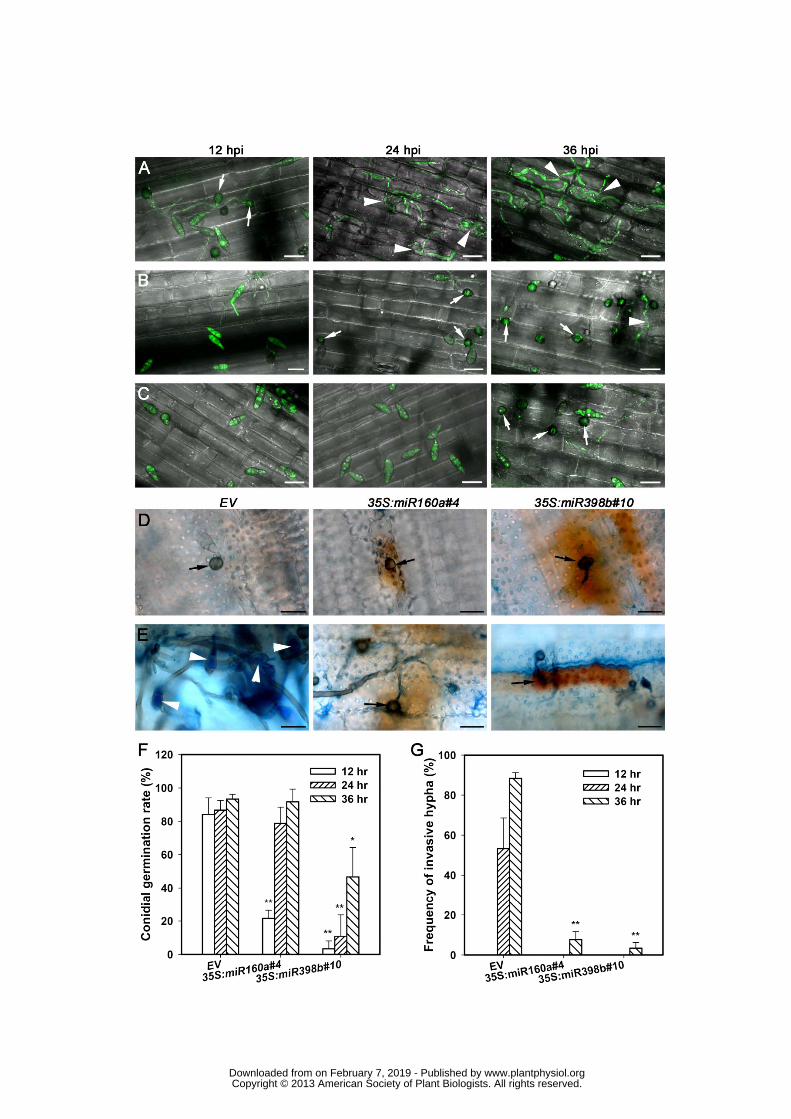
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

Figure 7. Cellular responses to the infection of M. oryzae. A to C, Representative Laser ScanningConfocal Microscopic images to show infection of the eGFP-tagged strain Zhong-8-10-14 on sheathcells of transgenic plants expressing empty vector (A), 35S:miR160a (B), and 35S:miR398b (C) at theindicated time points Note that appressoria (arrows) were formed at 12 hpi on EV plants but delayedindicated time points. Note that appressoria (arrows) were formed at 12 hpi on EV plants, but delayedto 24 hpi on miR160a-overexpression plants and 36 hpi on miR398b-overexpression plants. Invasivehyphae (arrowheads) were formed at 24 hpi and extended to the neighbor cells at 36 hpi on EV plants.Bars, 20 μm. D and E, Representative leaf sections from the indicated transgenic lines stained by 3,3’-diaminobenzidine and trypan blue at 2 dpi (D) and 10 dpi (E), respectively, to show the accumulationof H2O2 (reddish brown) at the infection site where appressoria (arrows) were visualized and the fungalstructures. Note that secondary conidia (arrowheads in E) were generated on EV plants at 10 dpi. Bars,10 μm. F and G, Statistic analyses of conidial germination rate (F) and frequency of invasive hypha (G)t th i di t d ti i t b d th i di t d li I l t d l t i d ith tat the indicated time points observed on the indicated lines. Inoculated leaves were stained with trypanblue and examined by microscopy. Data are mean±SEM from three independent experiments each ofwhich >200 conidia were evaluated. x2-test was used to test statistical significance between EV andtransgenic lines. Asterisks * and ** indicate significant difference at a P value <0.05 and <0.01,respectively.
www.plantphysiol.orgon February 7, 2019 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.






![Semiautomated Feature Extraction from RGB Images for Sorghum Panicle Architecture ... · Semiautomated Feature Extraction from RGB Images for Sorghum Panicle Architecture GWAS1[OPEN]](https://img.pdfslide.us/doc/110x75/5e5e983032be3a67ab7750f7/semiautomated-feature-extraction-from-rgb-images-for-sorghum-panicle-architecture.jpg)






















