Embed Size (px)
Citation preview

MicroRNAs as Prognostic Biomarkers in Prostate Cancer
by
Aida Gordanpour
A thesis submitted in conformity with the requirements for the degree of Doctor of Philosophy
Department of Laboratory Medicine and Pathobiology University of Toronto
© Copyright by Aida Gordanpour 2012

ii
MicroRNAs as Prognostic Biomarkers in Prostate Cancer
Aida Gordanpour
Doctor of Philosophy
Department of Laboratory Medicine and Pathobiology
University of Toronto
2012
ABSTRACT
Prostate cancer, one of the most common cancers among men, can be relatively harmless or
extremely aggressive. The most widely used biomarker for the disease, the PSA test, is not
independently diagnostic or prognostic of prostate cancer. One of the main challenges of prostate
cancer research is to find reliable and effective prognostic biomarkers that will predict cancer
recurrence following surgery, in order to identify clinically significant prostate cancer and
improve management of the disease. In recent years, microRNAs (miRNAs) have been identified
as master regulators of cellular processes, and dysregulated miRNAs have been associated with
cancer development and progression. The intent of my PhD research program was to uncover
novel miRNAs that contribute to prostate cancer pathogenesis in order to assess their potential as
predictors of clinical progression. By analyzing a large cohort of primary prostate cancer
samples, we have discovered that microRNA-221 (miR-221) is associated with metastasis and
biochemical recurrence in prostate cancer, and is downregulated in TMPRSS2:ERG fusion gene-
positive tumors. In addition, we have determined that microRNA-182 (miR-182) is
overexpressed in prostate cancer and is associated with increased metastasis and clinical
progression by targeting a tumors suppressor Forkhead box O1 (FOXO1). Overall, this work
introduces novel candidate miRNA genes and downstream targets that are aberrantly expressed

iii
in more aggressive prostate cancer, and presents a potentially significant role for miRNAs as
prognostic biomarkers that are associated with clinical progression, and perhaps aids in defining
how miRNAs might one day serve as anti-cancer therapeutic agents.

iv
ACKNOWLEDGMENTS
To say I was fortunate to have pursued my PhD studies at Sunnybrook Hospital does not entirely
capture the extent to which I recognize this to be true. The four years I spent there not only
highlight my work as a cancer biologist, but they also represent a period of time that has
profoundly influenced me personally. I am indebted to some of the finest people I have ever met
for inspiring me and making Sunnybrook a positive environment in which I learned and grew.
These individuals set an example of world-class scientists for their passion for cancer research,
mentors who offered their time and expertise so generously, and best of friends that I was truly
lucky to have met. The best way to honor their legacy would be to uphold their high calibre of
professionalism and integrity in my future endeavours.
It is my fortune to gratefully express my appreciation to my supervisor Dr. Arun Seth for the
continuous support of my PhD study and research, and for giving me the opportunities that
opened so many doors for me. I also wish to extend my thanks to the members of my committee,
lab members, and collaborators for their support.
I owe my most sincere gratitude to my closest friends; you know who you are, for bringing out
the best in me, for being a source of friendship as well as inspiration, and mostly for the pure
happiness that there was in times spent with you. The memories will last a lifetime…
Words fail to express my deep sense of appreciation to my family for all their unconditional love
and support. To my parents who raised me with a love of science and education, and to my
encouraging and caring sisters, brother-in-law, and little Audrina, Thank you!
Thank you God for giving me passion in my heart, inspiration in my life, showing me the way,
giving me strength, and for teaching me to always trust in you!
I am now most eager to embark on the next chapter of my life as a medical student…

v
TABLE OF CONTENTS
ABSTRACT ................................................................................................................................... ii
ACKNOWLEDGMENTS ........................................................................................................... iv
TABLE OF CONTENTS ............................................................................................................. v
LIST OF TABLES ....................................................................................................................... ix
LIST OF FIGURES ...................................................................................................................... x
LIST OF ABBREVIATIONS ..................................................................................................... xi
Chapter 1: GENERAL INTRODUCTION ................................................................................ 1
1.1 PROSTATE GLAND ......................................................................................................... 2
1.2 PROSTATE CANCER ....................................................................................................... 4
1.2.1 Epidemiology and Etiology of Prostate Cancer ...................................................... 4
1.2.2 PSA Test ................................................................................................................. 5
1.2.2.1 PSA Controversy ...................................................................................... 6
1.2.3 Pathology of Prostatic Neoplasia ............................................................................ 7
1.2.4 Treatment of Prostate Cancer .................................................................................. 9
1.3 RECURRENT CHROMOSOMAL ABERRATIONS IN PROSTATE CANCER.......... 10
1.3.1 TMPRSS2:ERG as a Biomarker ........................................................................... 14
1.3.2 Gene Fusions and Therapeutics ............................................................................ 15
1.4 NON-CODING RNAs ...................................................................................................... 16
1.4.1 MicroRNAs ........................................................................................................... 17
1.4.1.1 MiRNA History ...................................................................................... 20
1.4.1.2 The Canonical Pathway of MiRNA Processing ..................................... 20
1.4.1.3 MiRNAs in Diseases .............................................................................. 24
1.4.1.4 Putative Targets of MiRNAs .................................................................. 24
1.4.1.5 MiRNAs as Biomarkers in Prostate Cancer Pathogenesis ..................... 26

vi
1.4.1.5.1 MiRNAs as Oncogenes or Tumor Suppressors ...................................... 27
1.4.1.5.2 Metastasis-regulatory MiRNAs .............................................................. 29
1.4.1.5.3 MiRNAs in Preclinical Studies .............................................................. 31
1.4.1.6 MiRNAs as Therapeutic Agents—Breakthroughs and Challenges ........ 34
1.4.1.6.1 MiRNAs in Clinical Trials ..................................................................... 36
1.4.1.6.2 Prostate Cancer-associated MiRNAs Currently in Clinical Trials ......... 37
1.5 Characteristics of Malignant Transformation ................................................................... 40
1.6 PROJECT RATIONAL .................................................................................................... 45
1.7 THESIS OBJECTIVES AND ORGANIZATION ........................................................... 46
2 CHAPTER 2: MIR-221 IS DOWNREGULATED IN TMPRSS2:ERG FUSION-
POSITIVE PROSTATE CANCER ...................................................................................... 47
2.1 ABSTRACT ...................................................................................................................... 48
2.2 INTRODUCTION ............................................................................................................ 49
2.3 MATERIALS AND METHODS ...................................................................................... 52
2.3.1 Study Participants and Prostate Sample Collection .............................................. 52
2.3.2 Patient Follow-up .................................................................................................. 52
2.3.3 Quantitative Real-time Polymerase Chain Reaction (PCR) ................................. 53
2.3.4 RT–PCR and Direct DNA sequencing ................................................................. 53
2.3.5 Statistical Analysis ................................................................................................ 54
2.4 RESULTS ......................................................................................................................... 55
2.4.1 Patient Demographics ........................................................................................... 55
2.4.2 miR-221 is Downregulated in TMPRSS2:ERG Fusion Gene-positive Prostate
Tumours ................................................................................................................ 55
2.4.3 Low MiR-221 Expression is Associated with Metastasis and Biochemical
Recurrence of Prostate Tumour ............................................................................ 58

vii
2.4.4 MiR-221 Levels are Lower in Recurrent/Metastatic TMPRSS2:ERG Fusion-
negative Tumours .................................................................................................. 60
2.5 DISCUSSION ................................................................................................................... 62
3 Chapter 3: ABERRANT MIR-182 EXPRESSION ENHANCES THE
METASTATIC POTENTIAL OF PROSTATE CANCER AND PREDICTS
CLINICAL RECURRENCE................................................................................................. 70
3.1 ABSTRACT ...................................................................................................................... 71
3.2 INTRODUCTION ............................................................................................................ 72
3.3 MATERIALS AND METHODS ...................................................................................... 74
3.3.1 Cell Lines .............................................................................................................. 74
3.3.2 Quantitative Real-Time PCR ................................................................................ 74
3.3.3 Soft Agar Assay .................................................................................................... 74
3.3.4 Wound Healing Assay .......................................................................................... 75
3.3.5 Transwell Migration Assay ................................................................................... 75
3.3.6 Invasion Assay ...................................................................................................... 75
3.3.7 Proliferation Assay ................................................................................................ 76
3.3.8 Western Blotting ................................................................................................... 76
3.3.9 Transfection of SiRNA and MiRNA Inhibitors .................................................... 77
3.3.10 Prostate Sample Collection ................................................................................... 77
3.3.11 Patient Follow-up .................................................................................................. 77
3.3.12 Recurrence Analysis ............................................................................................. 78
3.3.13 Statistical Analysis ................................................................................................ 78
3.4 RESULTS ......................................................................................................................... 79
3.4.1 MiR-182 Expression is Increased in Human Prostate Tumors, and Its
Upregulation is Associated with Clinical Progression in Prostate Cancer. .......... 79
3.4.2 MiR-182 Overexpression Enhances the Tumorigenic Properties of LNCaP
Cells. ..................................................................................................................... 83
3.4.3 miR-182 Enhances the Migratory and Invasive Abilities of LNCaP Cells. ......... 89

viii
3.4.4 MiR-182 Negatively Regulates FOXO1 in LNCaP Cells and Primary Prostate
Tumors. ................................................................................................................. 93
3.4.5 FOXO1 Downregulation is Associated With Biochemical Recurrence in
Prostate Cancer. .................................................................................................. 100
3.5 DISCUSSION ................................................................................................................. 105
4 CHAPTER 4: GENERAL DISCUSSION AND FUTURE DIRECTIONS .................... 108
4.1 SUMMARY AND IMPACT OF MAJOR FINDINGS .................................................. 109
4.2 FUTURE DIRECTIONS ................................................................................................ 111
5 APPENDIX 1: MICRORNA DETECTION IN PROSTATE TUMORS BY
QUANTITATIVE REAL-Time PCR (QRT-PCR) ........................................................... 115
REFERENCES .......................................................................................................................... 130

ix
LIST OF TABLES
1.1- Gene fusions in prostate cancer -13
1.2- MiRNAs associated with primary prostate cancer -28
1.3- MiRNAs associated with metastatic prostate cancer -30
1.4- MiRNAs in prostate cancer preclinical studies -33
1.5- MiRNAs in prostate cancer clinical trials -39
2.1- Clinical demographics of prostate cancer cases used in this study -67
2.2- MiR-221 and TMPRSS2:ERG fusion gene status -68
2.3- MiR-221 as a prognostic factor of prostate cancer recurrence -69
3.1- Clinical demographics of prostate cancer cases used in this study -84

x
LIST OF FIGURES
1.1- Male anatomy -3
1.2- Gleason grades: original drawing -8
1.3- Schematic representation of TMPRSS2:ERG fusion gene -12
1.4- Overview of post-transcriptional gene regulation by miRNAs -18
1.5- The canonical pathway of miRNA processing -23
1.6- The hallmarks of cancer -41
2.1- MiR-221 is suppressed in prostate tumours with TMPRSS2:ERG -57
2.2- MiR-221 is suppressed in prostate tumours with metastasis and/or
biochemical recurrence. -59
2.3- MiR-221 levels are lower in recurrent/metastatic TMPRSS2:ERG fusion-
negative tumours -61
3.1- MiR-182 is overexpressed in human prostate tumors, and its upregulation is
associated with clinical progression -80
3.2- MiR-182 expression in LNCaP cells -85
3.3- MiR-182 overexpression enhances the tumourigenic properties of LNCaP
cells -87
3.4- MiR-182 enhances the migratory and invasive abilities of LNCaP cells -90
3.5- MiR-182 negatively regulates FOXO1 in LNCaP cells -95
3.6- MiR-182 negatively regulates FOXO1 in primary prostate tumors -98
3.7- FOXO1 downregulation is associated with prostate cancer progression -101
A1.1- Steps in qRT-PCR -118
A1.2- Prostate Sample preparation -120
A1.3- Generating a standard curve -124
A1.4- Amplification curve for PCR reaction -127
A1.5- Data analysis for PCR reaction -128

xi
LIST OF ABBREVIATIONS
°C Degree(s) Celsius
% Percent
3’ untranslated region 3’ UTR
Ab Antibody
AGO Argonaute
BPH Benign prostatic hyperplasia
cDNA complementary DNA
C. elegans Caenorhabditis elegans
CO2 Carbon dioxide
DNA Deoxyribonucleic acid
DGCR8 DiGeorge critical region 8
DMEM Dulbecco's modified eagle's medium
ETS E Twenty Six
ERG ETS related gene
Fig Figure
FFPE Formalin fixed paraffin embedded
FOXO1 Forkhead Box O1
FOXO3 Forkhead Box O3
Gb Gigabases
H&E Hematoxylin and Eosin
LnRNA long non-coding RNA
mRNA messenger RNA
g Microgram
l Microliter
mL Milliliter
mm Millimeter
MiRNA MicroRNA
MITF-M Microphthalmia-associated Transcription Factor-M
mAb monoclonal antibody
ncRNA Non-coding RNA
Ng Nanogram
NP-40 Nonidet P40
ORF Open reading frame
PIN Prostatic intraepithelial neoplasia
PSA Prostate specific antigen
PBS Phosphate buffered saline
PCR Polymerase chain reaction
Pre-miRNA Precursor miRNA
Pri-miRNA Primary miRNA
qPCR Quantitative PCR
RFP Red fluorescent protein
RNA Ribonucleic acid
rRNA Ribosomal RNA
RNase Ribonuclease
RISC RNA-induced silencing complex
S Second

xii
SD Standard deviation
SDS Sodium dodecylsulphate
SDS-PAGE Sodium dodecylsulphate-polyacrylamide gel electrophoresis
siRNA Small interfering RNA
SV Seminal vesicle
VEGF Vascular endothelial growth factor
tRNA Transfer RNA
TBS-T Tris buffered saline with tween 20
TCGA The Cancer Genome Atlas

1
Chapter 1: GENERAL INTRODUCTION
A version of this chapter has been published in Prostate Cancer & Prostatic Diseases as
Gordanpour A, Nam RK, Sugar L, Seth A. (2012) doi: 10.1038/pcan.2012.3. [Epub ahead of
print]

2
1.1 PROSTATE GLAND
Prostate is a sex accessory organ which plays a major role in the reproductive process by making
part of the seminal fluid. The prostate is about the size of a large walnut, and is located close to
the rectum, below the bladder at the base of the penis (Fig 1.1). The prostatic epithelium is
composed of two major cell types, stromal cells and epithelial cells. The stromal compartment
consists mainly of connective tissue, smooth muscle cells, and fibroblasts, and they mainly serve
as structural support. Normal prostatic epithelium consists of five cell types: stem cells, basal
epithelial cells, transit-amplifying cells, neuroendocrine cells, and secretory epithelial cells (1).
Prostate specific antigen (PSA) is one of several serine proteases secreted by the prostate cells
into the ejaculate, and although its main function is the regulation of semen coagulation, it has
proved to be an invaluable, yet controversial marker of prostatic diseases.

3
Figure 1.1- Male anatomy. Diagram adapted from medical-look.com

4
1.2 PROSTATE CANCER
Prostate cancer causes substantial clinical, social, and economical burden in the developed
world. Identification of genes that are dysregulated in association with prostate cancer can
provide molecular markers and clues relevant to disease etiology and progression. Prostate
tumors display novel recurrent chromosomal translocations, and aberrant expression of specific
miRNAs which can be used for elucidating prostate cancer biology, and better management of
this disease.
1.2.1 Epidemiology and Etiology of Prostate Cancer
Prostate cancer is the most common cancer in men and the second leading cause of
cancer death in North America, Europe and other developed regions of the world. According to
Canadian Cancer Society, there were 25,500 cases of prostate cancer in Canadian men in 2011,
with 4,100 deaths caused by this disease. On average, Canadian men have a 14% chance of being
diagnosed with prostate cancer at some point in their lives, while their chance of dying from the
disease is only 4%. The 5-year survival rate for prostate cancer is 96%. The prevalence of
prostate cancer increases significantly with age (2), and studies suggest that by the age 80, more
than 50% of men have some cancerous cells in their prostate (3). The introduction of PSA testing
has substantially increased the rate of detection of local-regional disease, whereas the incidence
of metastatic disease has decreased (4). Although the exact causes of prostate cancer
development and progression are not yet known, substantial evidence suggests that both genetics
and environment play a role in the origin and evolution of prostatic neoplasia, and several risk
factors associated with this disease include: familial and genetic influences (5), chronic

5
inflammation and infection (6), sexual activity (7), diet (8), and various molecular biomarkers
such as androgens (9), and vitamin D (10).
1.2.2 PSA Test
Since 1987, men have been undergoing a blood test called the PSA test to detect prostate
cancer at its earliest stages (11). PSA is a protein produced by prostate epithelial cells and the
majority of it is secreted in the semen and functions as a coagulase (12). Typically, a family
doctor will refer a patient to a urologist if his PSA level is above 4 ng/mL, and the patient will
then undergo a prostate biopsy for further evaluation. A small portion of PSA that is found in the
circulation is immediately bound by alpha-1 antichymotrypsin, referred to as complexed-PSA
(12). A small portion also exists as a free form. This unbound PSA may be caused by benign
prostatic hyperplasia (BPH), a non-cancerous enlargement of the prostate, and not prostate
cancer. Therefore the higher a man's percentage of free PSA, the less likely it is that prostate
cancer is to blame. Catalona et al. was the first to show that patients with a low free: total PSA
ratio were associated with a higher risk for prostate cancer (13). However, its ability to improve
the positive predictive value is only when the PSA level is between 4 - 10 ng/mL (14), the grey
zone in which BPH could be the culprit. Readings of greater than 25% free PSA suggest that
much of the elevated PSA is caused by BPH, while a reading of less than 10% indicates that
prostate cancer is most likely causing this elevation.

6
1.2.2.1 PSA Controversy
Despite routine application of PSA assays, PSA screening has been very controversial.
As of October 2011, the United States Preventive Services Task Force (USPSTF) has
recommended against the use of PSA as a screening tool for prostate cancer in asymptomatic
men. Prostate Cancer Canada, however, continues to view PSA as a useful test when applied
appropriately, and recommends PSA testing for the early detection of prostate cancer in Canada.
The controversy around use of PSA as a screening tools stems from the fact that PSA is for the
most part organ-specific and not disease-specific. This widely used test has been criticized for its
high false-positive results, causing over-treatment of cancers that may have never caused harm.
There are several reasons, in addition to prostate cancer, for elevated levels of PSA found in a
man's blood. These include: BPH, prostatitis (an inflammation of the prostate) (15), recent
ejaculation, digital rectal exam, and prostate biopsy. Among men with PSA levels greater than 4
ng/mL, biopsies show that over two-thirds of patients do not have prostate cancer (16).
Meanwhile, there are men with PSAs in the normal range (below 4ng/mL) who have prostate
cancer (17).
Another important limitation of PSA as a biomarker is its inability to identify patients
with aggressive and lethal forms of prostate cancer. Because many forms of prostate cancer are
indolent and do not progress to metastasis and death, it would be important for new biomarkers
to be able to distinguish those from aggressive prostate cancer. Over the last 25 years, no new
blood test, genetic test or medical x-ray have been able to replace PSA. So we are in a midst of a
biomarker crisis as lack of specificity of PSA requires supplementation in order to improve
patient management, and to differentiate cancer from benign diseases of the prostate. Research
has indicated that the therapeutic success rate for prostate cancer can be improved tremendously,

7
if this disease is diagnosed at an earlier stage. Currently, there remains a lack of reliable
biomarkers that can specifically distinguish between patients who need to be treated adequately
to halt the aggressive form of the disease and those who should avoid overtreatment of the
indolent form. Thus, current biomarkers for prostate cancer are not ideal as they fail to replace
PSA.
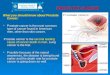
1.2.3 Pathology of Prostatic Neoplasia
Histologic grading is the clinically most useful prognostic factor for prostate cancer (18).
The Gleason grading system is a method used to evaluate prostatic carcinoma based on the
architectural pattern of the tumor as identified at relatively low magnification. This technique
was developed in 1966 by Dr Donald F. Gleason, a pathologist, and members of the Veterans
Administration Cooperative Urological Research Group (19). This grading system assesses the
histologic pattern of cancer cells in Hematoxylin and Eosin (H&E)-stained sections (20). The
primary (predominant) and the secondary (second most prevalent) patterns are identified and
assigned a grade from 1 to 5, with 1 being the most differentiated and 5 being the least
differentiated, and the Gleason score is calculated by the addition of the primary and secondary
grades. The five different grade patterns used to generate a score are illustrated in a drawing by
Dr Gleason (Fig 1.1). Due to different biopsy practices and also the widespread use of PSA test,
diagnosis of prostate cancer today is significantly different from the time that Gleason grading
was first used—when most men had advanced disease. Hence there have been modifications in
how the Gleason grading is utilized, namely to restrict the definition of Gleason pattern 3 and
expand the definition of pattern 4, therefore causing a general trend toward upgrading (18).

8
Figure 1.2- Gleason grades: original drawing. Adapted from Humphrey, Modern
Pathology. (17) 292-306, 2004.

9
1.2.4 Treatment of Prostate Cancer
Because prostate cancer exhibits a wide spectrum of aggressiveness, different methods of
treatment have been developed. Clinically localized prostate cancer is confined to the prostate,
and the treatment options for it include surgery (radical prostatectomy), radiation therapy, and
active surveillance (deferred treatment until necessary) (21). Locally advanced prostate cancer is
defined as disease that shows evidence of spread outside of the prostate capsule, involvement of
the seminal vesicles, or involvement of adjacent organs. Although the widespread use of PSA
screening has caused a decline in proportion of men who are diagnosed with locally advanced
prostate cancer, it remains a considerable fraction which contributes disproportionately to disease
mortality. Hence better treatment options in these cohorts can have a major positive influence on
overall morbidity and mortality of prostate cancer. It has been shown that only 25% of men with
seminal vesicle invasion and none with lymph node metastases are biochemically progression-
free at 10 years after radical prostatectomy (22).
Metastatic disease constitutes cancer that has spread beyond the prostate. The most
frequent sites of metastatic prostate carcinoma are lymph node and bones, followed by lungs,
bladder, liver and adrenal glands (23). Treatment options for metastatic prostate cancer include
chemotherapy and hormonal therapy. In hormone refractory disease, prostate cancer continues to
grow despite the fact that hormonal therapy suppresses male hormones androgen and
testosterone. Patients with hormone refractory prostate cancer are given chemotherapy which
may lengthen survival time and reduce the pain caused by cancer spreading to the bones.

10
1.3 RECURRENT CHROMOSOMAL ABERRATIONS IN PROSTATE CANCER
Chromosomal aberrations in haematological disorders such as leukemia and lymphoma
were identified and characterized years ago, and are now routinely used for diagnosis, patient
stratification, and drug development (24). Genomic aberrations were largely ignored in solid
tumours, mostly due to technological limitations associated with cytogenetics, until recently
recurrent gene fusions were found in epithelial tumours. Six years after the discovery of the first
gene fusion in prostate cancer by Tomlins et al.(25), they are now considered to be an important
and distinct class of mutations that occur in a high percentage of patients and can be powerful
biomarkers of prostate cancer.
The most prevalent form of genetic aberration in prostate cancer is gene fusions. They
can involve a variety of 5’ regulatory elements fused to ETS genes such as ERG, ETV1, ETV4,
and ETV5, which are normally involved in self-renewal-associated proliferation (26).
TMPRSS2:ERG, the most common gene fusion, is present in over 50% of prostate cancers (27),
and is absent in normal and benign prostate tissue (28). It is characterized by fusion of the
promoter region of an androgen-responsive, prostate-specific, trans-membrane serine protease
(TMPRSS2), to v-ets erythroblastosis virus E26 oncogene homolog (ETS related gene; ERG),
resulting in the over-expression of a transcript that encodes a truncated ERG protein product (Fig
1.3). ERG gene is a member of the ETS (E Twenty Six) family of transcription factors. In vitro
studies have shown that overexpression of ERG stimulates cell migration and invasion, while its
knockdown decreases the invasive properties of VCaP cells (29).
TMPRSS2 and ERG are both on the long arm of chromosome 21, and are separated by
about 3 Mbps. In the case of this fusion gene, a deletion removes the intervening sequence.

11
Fusion genes with ETV1, ETV4, and ETV5 are a lot less common, only present in about 10% of
prostate tumours, but display great variability in fusion structure (Table 1.1). It is unclear what
causes the formation of fusion genes, but there is emerging evidence that inherited
polymorphisms can predispose individuals to develop TMPRSS2:ERG (30).

12
Figure 1.3- Schematic representation of TMPRSS2:ERG fusion gene.

13
Table 1.1- Gene fusions in prostate cancer
Fusion gene Fusion type Locus Study
TMPRSS2-ERG Deletion/insertion (21)(q22.2) Tomlins et al. (25) SLC45A3-ERG translocation (1;21)(q32.1;q22.2) Esgueva et al. (31) HERPUDI-ERG translocation (16;21)(q13;q22.2) Maher et al. (32) NDRG1-ERG translocation (8;21)(q24.22;q22.2) Pflueger et al. (33) TMPRSS2-ETV1 translocation (7;21)(p21.2;q22.3) Tomlins et al. (34) HERVK-ETV1 translocation (7;22)(p21.2;q11.23) Hermans et al. (35) C15orf21-ETV1 translocation (7;15)(p21.2;q21) Tomlins et al. (34) HNRPA2B1-ETV1 translocation (7;7)(p21.2;p15) Tomlins et al. (34) SLC45A3-ETV1 translocation (1;7)(q32;p21.2) Han et al. (36) ACSL3-ETV1 translocation (2;7)(q36.1;p21.2) Attard et al. (37) FLJ35294-ETV1 translocation (7;17)(p21.2;p13.1) Han et al. (36) EST14-ETV1 translocation (7;14)(p21.2;q21) Hermans et al. (35) TMPRSS2-ETV4 translocation (17;21)(q21;q22.3) Tomlins et al. (38) KLK2-ETV4 translocation (17;19)(q21;q13) Hermans et al. (39) CANT1-ETV4 inversion (17;17)(q22;q25) Hermans et al. (39) DDX5-ETV4 translocation (17;17)(q21;q21) Han et al. (36) TMPRSS2-ETV5 translocation (3;21)(q27;q22.3) Helgeson et al. (40) SLC45A3-ETV5 translocation (1;3)(q32;q27) Helgeson et al. (40) SLC45A3-BRAF translocation (1;7)(q32.1;q34) Palanisamy et al. (41) SLC45A3-ELK4 unknown 1(q32.1) Rickman et al. (42) ESRP1-RAF1 translocation (3;8)(p25.1;q22.1) Palanisamy et al. (41)

14
1.3.1 TMPRSS2:ERG as a Biomarker
PSA is the current standard of care in prostate cancer screening, but due to its lack of
specificity and low sensitivity, it has become increasingly clear that other biomarkers are needed
to complement PSA. Ever since the discovery of recurrent gene fusions in prostate cancer,
TMPRSS2:ERG fusion gene has been considered a potentially suitable biomarker to supplement
or even replace the PSA test, and many groups are working towards that goal. For example, a
noninvasive, urine-based multiplex prostate cancer assay that uses TMPRSS2:ERG as one of the
four genetic markers of cancer diagnosis was able to outperform the PSA test by achieving a
specificity and positive predictive values of 76.0% and 79.8%, respectively (43). A similar study
has also reported that a combined test for TMPRSS2:ERG and prostate cancer antigen 3 (PCA3)
expression in urine can significantly improve the sensitivity of prostate cancer detection (44). For
quite some time the ERG protein was not detectable due to lack of specific antibodies. But
recently a rabbit anti-ERG monoclonal antibody (Epitomics, Burlingame, CA) has been shown
reliable and sensitive, and can detect ERG-rearrangement positive prostate cancer (45). ERG
protein expression, thus, can potentially be a valuable marker to identify patients’
TMPRSS2:ERG status with a prostate needle biopsy.
Another important aspect of TMPRSS2:ERG as a biomarker is whether it can act as a
reliable prognosticator in prostate cancer. After years of investigation and numerous studies,
there is still no clear consensus on gene fusion status and clinical outcome. While there are many
studies that have linked fusion positive tumours to negative prognostic outcome (46-53), there
are also studies that show no association (54-57), or even improved clinical outcome (58-60).
The inconsistency between these studies highlights the likelihood that TMPRSS2:ERG interacts
with additional factors to affect the progression of prostate cancer. There is much optimism that

15
TMPRSS2:ERG can effectively stratify patients for surveillance or treatment options, and
potentially act as a direct and specific target for molecularly-based drugs.
1.3.2 Gene Fusions and Therapeutics
Gene fusions can be powerful and specific targets for anticancer drugs and some cancers
are classified and treated based on a particular gene fusion status. Chronic myeloid leukemia
patients with BCR–ABL fusion gene are treated with Imatinib mesylate (Gleevec®, STI-571)
(61), and acute promyelocytic leukemia patients with PML–RARα fusion of the retinoic acid
receptor-α are treated with all-trans retinoic acid in combination with chemotherapy (62). A
recent study by our group, summarized in Chapter 2 of this thesis, showed miR-221 is predictive
of recurrence in prostate cancer, and most notably, we also found that miR-221 is significantly
down-regulated in patients with TMPRSS2:ERG fusion gene (63). That study was the first to
show a clinical link between aberrant expression of a miRNA and TMPRSS2:ERG fusion gene
status, and highlights the potential in identification of miRNAs regulating the oncogenic ERG.
Fusion gene status can potentially pave the way for individualized therapy and better treatment
options for men diagnosed with the disease.

16
1.4 NON-CODING RNAs
More than 98% of the human genomic size consists of non-coding sequences (64). Until
recently, these sequences were called ‘junk DNA' based on the fact that no specific function
could be attributed to them. The discovery of non-coding RNAs (ncRNAs) as invaluable
regulators of gene expression has fundamentally changed our understanding of the genome and
gene expression. These RNA molecules play crucial roles in eukaryotic physiological and
pathological processes. NcRNAs vary in size from miRNAs that are short in size, a mature
miRNA is 18-25 nucleotides long, to long non-coding RNAs (lnRNAs) that are thousands of
nucleotides long.
In 1965 the first ncRNA, alanine transfer RNA (tRNA), was discovered in baker’s yeast
(65). tRNAs are adaptor molecules that acts as a bridge between the four-letter genetic code of
mRNA and the twenty-letter code of amino acids in proteins (66). One end of the tRNA contains
the three-nucleotide sequence called the anticodon which forms three base pairs with the
matching codon in mRNA. On its other end, each tRNA is covalently attached to the amino acid
that corresponds to the anticodon sequence. During protein synthesis, a tRNA is delivered to the
ribosome where it attach the amino acid from its 3’ end to a growing polypeptide chain.
Ribosomal RNA (rRNA) is the RNA component of the ribosome that forms 2 subunits that
envelope mRNA during translation and catalyze the formation of a peptide bond between 2
amino acids (67). While tRNA and rRNA are two major house-keeping ncRNAs, there are
several others that also play important roles in the cellular processes. For example, 7SL RNA is a
component of signal recognition particle which binds to the signal peptide emerging from a
ribosome and directs the traffic of the protein to the plasma membrane or the endoplasmic
reticulum membrane for secretion or membrane insertion within the cell (68).

17
1.4.1 MicroRNAs
MiRNAs constitute a large family of naturally-occurring, endogenous, single stranded
RNA molecules that regulate gene expression in metazoans and plants (69). MiRNAS are not
translated into proteins. The canonical mechanism of miRNA-mediated gene regulation is by its
binding to the 3’ UTR of an mRNA and inhibiting protein production (70). Nucleotides 2-7 from
the 5’ end of the miRNA, called seed sequence, are essential to the binding of the miRNA to the
target mRNA, and has to bind perfectly. All the other nucleotides of the miRNA can bind
imperfectly. Initially, miRNAs were thought to regulate gene expression solely by inhibiting
protein translation of a target mRNA (71). Conversely, recent studies have shown that miRNAs
can also degrade their target mRNA by affecting its stability, thereby inhibiting protein synthesis
indirectly (72;73) (Fig 1.4). Although the majority of published data has focused on miRNAs
that act via the canonical pathway, there are no mechanistic requirements that restrict miRNA
action to only the 3’ UTR. In fact, miRNAs have also been shown to use 5’ UTR and open
reading frame (ORF) binding sites to regulate mRNA expression (74;75). In addition, there have
been reports of miRNAs that influence gene expression by directly binding to DNA (76-78), and
some miRNAs can even activate, rather than inhibit gene expression (79). Taken together, these
findings highlight the complexity of gene regulation by the miRNAs.
The field of cancer genetics has rapidly expanded from identifying the first cancer-related
miRNAs (80), to exploring their potential as therapeutic options in less than ten years. Based on
the latest release 18 of miRBase database, the primary online repository for all miRNA
sequences and associated annotations (81), there are 21643 mature miRNA products in 168
species, including 2,154 mature human miRNAs. MiRBase is available online at:
http://www.mirbase.org/. It is estimated that 60% of mRNAs are regulated by miRNAs (82-85).

18
Figure 1.4- Overview of post-transcriptional gene regulation by miRNAs. MiRNAs regulate gene expression by binding to their target
mRNAs, and thereby repress translation or promote degradation of the target mRNA, determined by the amount of complementarity that
exists between the miRNA and its target mRNA.

19
MiRNAs have been associated with almost every cellular process, such as development (86),
differentiation (87), apoptosis (88), and cell cycle regulation (89). Consequently, misregulation
at any of these processes due to abnormal miRNA expression or mutations can potentially lead to
cancer, as the aberrant expression of many miRNAs has been characterized in numerous cancer
subtypes, including prostate cancer.

20
1.4.1.1 MiRNA History
The first miRNA was discovered in Caenorhabditis elegans (C. elegans) in 1993 by Lee
et al., who observed that a non-coding 22-base RNA (lin-4) could bind to the 3’ untranslated
region (3’UTR) of lin-14 messenger RNA (mRNA), and repress its translation (90). At that time,
the scientific community did not know that a new class of RNAs had been discovered that would
revolutionize our understanding of molecular biology. It was not until 7 years later, in 2000, that
a second 21-base RNA let-7 was identified in C. elegans which could also bind to the 3’UTR of
lin-41 mRNA, and repress its protein levels (91). Identification and functional studies of
miRNAs then extended from C. elegans to other species, including humans. Since then, there has
been an exponential growth of both miRNA discoveries as well as number of publication
focusing on miRNAs.
1.4.1.2 The Canonical Pathway of MiRNA Processing
MiRNA biogenesis starts with transcription in the nucleus (Fig 1.5) (92). MiRNA genes
are transcribed by RNA polymerase II or RNA polymerase III to form primary miRNA
transcripts (pri-miRNA) (93;94). Many pri-miRNAs are polyadenylated and have a 5’ 7-
methylguanylate cap (95). Usually, one miRNA gene codes for a single miRNA, but there are
also miRNAs that are clustered together and are transcribed as one transcript (96). A typical
human pri-miRNA consists of one or more hair pin structures, and each hairpin has a stem of 33
base-pairs, a terminal loop, and two single-stranded flanking segments upstream and downstream
of the hairpin stem (97). The pri-miRNA is cleaved by the nuclear microprocessor complex
formed by Drosha, a member of the RNase III family of enzymes, and the DGCR8 (DiGeorge

21
critical region 8) proteins (98). DGCR8 functions as an anchor which recognizes the junction
between the single-stranded flanking regions and the double-stranded stem, and acts as a
molecular ruler that guides Drosha to cleave the RNA molecule 11 base pairs away from the
single-stranded/double-stranded RNA junction at the base of the hairpin stem (99). The product
of this cleavage is the precursor miRNA (pre-miRNA) that has a 2 nucleotide 3’ overhang, and is
exported from the nucleus to the cytoplasm by Exportin-5 (100). Exportin-5 recognizes the pre-
miRNA regardless of sequence or structure of the pre-miRNA, but the 16 base pair minimal
length of the double-stranded stem and single-stranded flanking regions are important for the
proper binding pre-miRNA to Exportin-5 (101).
In the cytosol, RNase III Dicer cleaves off the pre-miRNA loop and generates an
approximately 22-nucleotide miRNA duplex (102). This mature miRNA duplex is double
stranded, and gets separated into two strands by helicases. A functional strand, the strand with
the less stable base pair at its 5’ end, is incorporated into the RNA-induced silencing complex
(RISC) and guides it to bind to the target mRNA, thereby inhibiting translation by degradation of
the mRNA or by translational repression (103). The other strand, the passenger strand, will
subsequently get degraded. Argonaute (AGO) proteins are important factors in the assembly and
function of the RISC complex that mediate mRNA degradation or translational inhibition (104).
The human genome encodes four AGO proteins (hAgo1-4) (105), of which only hAgo2 is
endonucleolytically active (106). An Activated RISC complex consists of an AGO protein and a
single-stranded miRNA, which acts as a guide to target complementary sequences within
mRNAs, in a process which is thought to be driven by diffusion (107). When the seed sequence
of miRNA and its target mRNA have high complementation, the RISC induces mRNA
degradation via the RNase III catalytic domain of AGO proteins, followed by degradation by

22
enzyme XRN1, the exosome, and the SKI complex (108). If partial complementarity is formed
between the two RNA molecules, mRNA degradation can occur by two pathways. Initially, the
poly(A) tail of the mRNA is deadenylated which allows the cytoplasmic exonucleases to degrade
the mRNA from the 3’ end (109). Another mRNA degradation mechanism is by removal of 5’
cap via decapping complex proteins (DCP1 and DCP2) and CAF1-CCR4-NOT complexes (110).
The mRNA is then degraded from 5’ to 3’ end by XRN1 (111). Regardless of which of these
mechanisms occur, there will be a decrease in the amount of protein synthesized by the mRNA.

23
Figure 1.5- The canonical pathway of miRNA processing. Adapted from Winter et al., Nature
Cell Biology, (11) 3:228-234, 2009

24
1.4.1.3 MiRNAs in Diseases
Considering that miRNAs are vastly involved in various physiological and pathological
processes in the cell, it is therefore not surprising that miRNA dysregulation is linked to
numerous diseases including many different cancers. There are several different ways by which
misregulation of miRNAs may lead to cancer. Misregulation by miRNA processing molecules
such as Drosha and Dicer may lead to disruption of miRNAs and cancer (112;113). Changes in
transcription of miRNAs by altered methylation, histone modifications, or transcription factors
that regulates miRNA genes can lead to different regulation of miRNAs (114). Genomic
instabilities such as translocation, deletion, amplification can also affect miRNA regulation.
Approximately half of human miRNA genes are encoded in genomic regions prone to cancer-
associated alterations (115). If a miRNA is located in a genomic region that is amplified, it can
become oncogenic. In contrast, a miRNA that is located in a genomic region that is deleted in
cancer can act as a tumor suppressor. Loss of miRNA binding site in the target mRNA, caused
by a SNP, alternate splicing, or separation of 3’UTR due to a chromosomal translocation may
also lead to misregulation of miRNAs (116). MiRNA expressions may also correlate with the
clinical characteristics of cancer, such as the tumor aggressiveness and its response to therapy,
which will be discussed more thoroughly in subsequent thesis chapters.
1.4.1.4 Putative Targets of MiRNAs
The major functional role of miRNAs is marked by the protein-coding mRNAs that they
target. Due to partial complementarity between a miRNA molecule and its binding sequence,
every miRNA can potentially bind to numerous mRNA targets, and each mRNA could be

25
regulated by many different miRNAs. This makes elucidating the biological function of a
miRNA challenging, as identification of a single target may not adequately reflect the entire
function of a miRNA. Most studies thus far have used bioinformatic tools to predict targets of
different miRNAs, and algorithms on the most frequently used bioinformatic programs often
give hundreds of putative targets for each miRNA. Different computational target prediction
software such as TargetScan, PicTar, miRanda, and miRBase can use varying methods
combining thermodynamics-based models and sequence alignment of miRNA-mRNA molecules
and their conservation in different species to name putative targets of miRNAs (117). While
these algorithms are a good starting point for the prediction of miRNA targets, ultimately
functional experiments must be used to validate a gene target.
Prediction of an miRNA-mRNA interaction is based on several factors (118). A
conserved sequence known as the seed sequence, situated at nucleotides 2-7 from the miRNA 5´-
end, is essential for the binding of the miRNA to the mRNA. Although base pairing between
miRNA and its target mRNA is only partially complementary, the seed sequence has to be
perfectly complementary. The complementarity between the 6 base pairs in the seed sequence is
the minimum for efficient binding, and the probability that a mRNA sequence is a target of a
miRNA is higher if there are more than 6 paired bases. Their specificity is also increased by
cross species binding site conservation, binding site structural accessibility (119), and proximity
of one binding site to another within the same 3' UTR (120).

26
1.4.1.5 MiRNAs as Biomarkers in Prostate Cancer Pathogenesis
The initial studies of miRNA deregulation in prostate cancer were performed by miRNA
microarray profilings, and since then several studies have analyzed prostate cancer-specific
miRNA profiles by using genome-wide screenings and validation by quantitative PCR
technology (121-126). Examination of prostate tumor miRNA expression profiles has revealed
widespread dysregulation of miRNAs in primary prostate cancer compared to normal prostate
tissue. In addition there are miRNAs that are abnormally expressed in different stages of disease
progression and metastasis, and are hence implicated as prognostic markers of clinical
aggressiveness and recurrence. There are some miRNAs that have been found to influence tumor
microenvironment (127). This extensive body of knowledge points to miRNAs as potentially
being novel cancer biomarkers, and as such they have several advantageous features. The small
size and distinctive biochemical structure of miRNAs makes them resistant to endogenous
RNase activity (128). MiRNAs are easily detected in tissue, blood, urine, saliva, and oral mucosa
by the sensitive and easy detection methods such as quantitative Real-Time PCR (qPCR). Small
RNAs such as miRNAs are extremely stable in formalin-fixed tissues, and that is a critical factor
in the detection of a biomarker (129). Although there is a decrease in the level of miRNA
molecules after the Formalin fixed paraffin embedded (FFPE) process, this diminishment is
proportional to, and correlates with the corresponding miRNA levels in cryopreserved specimens
(130;131). miRNAs are also detectable in plasma and serum fluids, as Mitchell et al. were able
to distinguish patients with prostate cancer from healthy controls by measuring miR-141 levels in
serum (128). This remarkable stability allows miRNAs to be readily detectable and makes them
potentially invaluable biomarkers.

27
1.4.1.5.1 MiRNAs as Oncogenes or Tumor Suppressors
MiRNA expression is deregulated in cancer cells compared to the corresponding normal
tissue, and depending on the specific cell type and stage of development, miRNAs can function
as either tumor-suppressors or oncogenes. Together, they have been termed as oncomirs. Table
1.2 shows numerous miRNAs that have been reported to be abnormally expressed in primary
prostate cancer in comparison to normal tissue. Some miRNAs, specifically marked in Table 1,
have shown conflicting results—having been reported to be up-regulated in some studies and
down-regulated in others. Different methods of sample collection, varied study designs, and
diverse specificity of the platforms used, could explain some of these inconsistencies. Aberrant
expressions of some miRNAs such as miR-25, miR-34a, miR-145 and miR-205 have been
reported in several miRNA studies. Oncogenic miRNAs that are up-regulated in cancer may
promote tumorigenesis by negatively regulating tumor suppressor genes inhibiting proliferation,
or by repressing genes associated with apoptosis and differentiation. In this case, inhibition of
miRNA activity can be achieved by antisense synthetic oligonucleotides (antagomirs) or by
drugs that can inhibit the oncogenic activity of the miRNA. Conversely, down-regulation of a
miRNA that functions as a tumor-suppressor can promote neoplastic development by enhancing
proliferation. Restoration of these miRNAs with synthetic mimics or by using viral vectors
encoding the miRNAs could have a therapeutic benefit by halting or even reversing tumor
growth. It is important to note that there is a vast number of miRNAs that have been linked to
prostate cancer in genome-wide screening studies, but there is still no consensus on exactly
which miRNAs are involved in prostate cancer formation and progression. Hence further
experimental validation is critical in better elucidating miRNAs’ biological function and role in
the pathogenesis of prostate cancer.

28
♦Opposite expression of miRNAs in different studies
Table 1.2- MiRNAs associated with primary prostate cancer
Up-regulated in prostate tumors Down-regulated in prostate tumors
let-7a♦(125) miR-106b(122) miR-198(123;126) let-7a♦(123;126) miR-34a♦(122) miR-329(122) let-7d♦(126) miR-122a(125) miR-199(126) let-7b(122;123) miR-92♦(123) miR-331-3p(132) let-7i(122;126) miR-124a(126) miR-200c(122) let-7c(123) miR-99(123) miR-340(122) miR-16♦(126) miR-125a♦(122) miR-202(123) let-7d♦(123) miR-100(123;125) miR-345♦(122) miR-17-5p(126) miR-129(125) miR-203♦(126) let-7f(123) miR-101♦(133) miR-368(124) miR-20a(125;126) miR-130b(124) miR-206(126) let-7g(123) miR-103(123) miR-373♦(134) miR-21(126) miR-135(126) miR-210(123) miR-1-2(122) miR-125a♦(123) miR-410(122) miR-25(122;125;126) miR-141♦(125;128) miR-214(126) miR-7-1(122) miR-125b(123;125) miR-449a(135) miR-26a♦(126) miR-146(126) miR-221♦(136) miR-7-2(122) miR-126(122) miR-487(122) miR-26a-1(122) miR-148(126) miR-222♦(136) miR-15a (137) miR-128a(122;126) miR-490(122) miR-26a-2(122) miR-181a-1(122) miR-223(126) miR-16♦(123;124) miR-133a-1(122) miR-494(122) miR-27a♦(126) miR-181a-2(122) miR-296(123) miR-16-1(137) miR-141♦(123) miR-497(123) miR-29b♦(126) miR-181b♦(126) miR-302(125) miR-19b(123) miR-143(123;125) miR-499(122)
miR-30c♦(126) miR-182(122;138) miR-302c(123) miR-22(123) miR-145(122;123;125)
miR-520c♦(134)
miR-31♦(122) miR-182*(124) miR-320(123) miR-23a(123) miR-148a(123) miR-520h(122) miR-32(122;126) miR-183(124;138) miR-345♦(123) miR-23b(123;125) miR-149(124;126) miR-34a♦(126) miR-184(123;126) miR-370(122;123) miR-24(125;126) miR-181b♦(124) miR-34b(122) miR-187(126) miR-373♦(123) miR-26a♦(123) miR-195♦(123) miR-92♦(126) miR-188(122) miR-375(122;124) miR-26b(123) miR-199a(123) miR-92-1(122) miR-191(126) miR-425(122) miR-27a♦(123) miR-203♦(139)
miR-92-2(122) miR-194-1(122) miR-449(122) miR-27b(123) miR-205(122-125;140)
miR-93(122;126) miR-194-2(122) miR-491(123) miR-29a(123) miR-218(126) miR-95(126) miR-195♦(126) miR-498(123) miR-29b♦(123) miR-218-2(122) miR-96(124;138) miR-196(126) miR-503(123) miR-30a-5p(123) miR220(122) miR-99b(122) miR-196a-1(122) miR-513(123) miR-30b(123) miR-221♦(122-125) miR-101♦(126) miR-196a-2(122) miR-524*(124) miR-30c♦(123) miR-222♦(123-125) miR-106a(126) miR-197(126) miR-634(124) miR-31♦(124) miR-330(141)

29
1.4.1.5.2 Metastasis-regulatory MiRNAs
Cancer metastasis, rather than the primary tumor itself, is the cause of death in prostate
cancer patients. In addition to their role as potential oncogenes and tumour-suppressors, miRNAs
can also be involved in the regulation of metastasis (142). These “metastamirs” could be
regulating various steps of the metastatic cascade such as epithelial to mesenchymal transition
(EMT), migration, invasion, angiogenesis, adhesion, and colonization of distant organs. So far,
only a few studies have investigated miRNAs associated with metastatic prostate cancer. One
recent study by Watahiki et al. has used next generation sequencing to compare the miRNA
profiles of a transplantable metastatic versus a non-metastatic prostate cancer xenograft, and has
identified many miRNAs that are differentially expressed in prostate cancer metastasis (143).
Table 1.3 lists miRNAs that have been linked to metastasis in prostate cancer as either pro-
metastatic or anti-metastatic. Some miRNAs such as miR-21, miR-331-3p, miR-205, and miR-
203 have been associated with prostate cancer metastasis. For instance, miR-21 has been shown
to be up-regulated in the majority of human cancers, including prostate, and has been associated
with increased invasiveness of prostate LNCaP cells (144). MiR-205 has been reported by
Gandellini et al. to inhibit EMT and reduce cell migration and invasion through down-regulation
of protein kinase Cepsilon (140).

30
Table 1.3- MiRNAs associated with metastatic prostate cancer
Pro-metastatic miRNAs Anti-metastatic miRNAs
let-7d(143) miR-128(143) miR-451(143) let-7c(145) miR-143(146;147) miR-361-5p(143) let-7g(143) miR-136(143) miR-486-3p(143) miR-1(148) miR-145(143;147) miR-363(143) let-7g*(143) miR-138(143) miR-486-5p(143) miR-7-1*(143) miR-146a(149;150) miR-424(143) let-7i(143) miR-140-5p(143) miR-520c(151) miR-15a(137) miR-146b-5p(143) miR-425(143) miR-7(143) miR-142-5p(143) miR-542-3p(143) miR-15b(143) miR-185(143) miR-454(143) miR-9(143) miR-144(143) miR-744*(143) miR-16(143;152) miR-186(143) miR-497(143) miR-9*(143) miR-144*(143) miR-1246(143) miR-16-1(137) miR-188-5p(143) miR-503(143) miR-17(143) miR-148b*(143) miR-17-3p(153) miR-191(143) miR-542-5p(143) miR-18a(143) miR-151-3p(143) miR-24(143) miR-193a-3p(143) miR-556-5p(143) miR-18b(143) miR-152(143) miR-24-2*(143) miR-195(143) miR-582-5p(143) miR-20b*(143) miR-181a-2*(143) miR-26b(143) miR-196a(143) miR-590-5p(143) miR-21(144;150) miR-191(150) miR-28-5p(143) miR-200b(154) miR-627(143) miR-27a(143) miR-200a(143) miR-29a(143) miR-200b*(143) miR-651(143) miR-27b(143) miR-210(143) miR-29b(155) miR-200c*(143) miR-652(143)
miR-30a(143) miR-218(143) miR-29c(143) miR-203(139;143;156)
miR-660(143)
miR-30a*(143) miR-223*(143) miR-33b(143) miR-205(140;143;157)
miR-664(143)
miR-31(143) miR-301*(143) miR-34a(143;158) miR-218(145) miR-708(143) miR-34c-5p(143) miR-340*(143) miR-95(143) miR-221(63;159) miR-1180(143) miR-99a(143) miR-373(151) miR-100(145) miR-324-5p(143) miR-1269(143) miR-106a(143) miR-374a(143) miR-101*(143) miR-331-3p(143;160) miR-1287(143) miR-125b(143) miR-379(143) miR-106b(143) miR-335(143) miR-2115*(143)
miR-125b-2*(143) miR-449a(143) miR-126*(143;161)
miR-339-5p(143) miR-3065-5p(143)
miR-126(143) miR-450a(143) miR-133a(148) miR-342-3p(143)

31
1.4.1.5.3 MiRNAs in Preclinical Studies
There is a small but increasing number of preclinical studies focusing on the role of
miRNAs in prostate cancer, and have been reported in Table 1.4. Bonci et al. have shown that
miR-15a and mir-16-1 are down-regulated in cancer cells of advanced prostate cancer, resulting
in BCL2, CCND1, and WNT3A up-regulation, which leads to increased cell survival and
invasion (137). They showed that the loss of function of miR-15a and miR-16 causes the
nontumorigenic prostate cell line RWPE-1 to form tumours in NOD-SCID mice. The authors
propose that miR-15a and miR-16 may have significant therapeutic potential, as single agents or
in combination with chemotherapy, since delivery of miR-15a and miR-16 to prostate cancer
xenografts caused tumour regression. Interestingly, Takeshita et al. reported that systemic
delivery of miR-16 by atelocollagen inhibits the growth of metastatic prostate cancer in bone of
nude mice, and may potentially be used for the treatment of metastatic prostate cancer (152).
Zhang et al. observed that miR-17-3p regulates the expression of vimentin and altered expression
of miR-17-3p decreases the tumourigenic behaviour and reduces tumour size in nude mice (153).
Saini et al. also noted that relative miR-203 expression is lower in prostate cancer cell lines
derived from bone metastasis, and over-expression of miR-203 has a negative effect on the
development of metastases in nude mice (156). Significantly, miR-203 has been shown to repress
bone-specific transcriptional regulators such as Runx2 and Dlx5, in addition to regulating pro-
metastatic genes ZEB2, Bmi, and Survivin. A recent preclinical study by Liu et al. showed
convincing preclinical therapeutic evidence, implicating miR-34a in prostate cancer metastasis
(162). Intratumoral injection of miR-34a into subcutaneous tumours inhibited tumour growth,
and systemic delivery of miR-34a into the tail vein could reduce tumour burden by half. In
orthotopic LAPC9 tumors, they showed that miR-34a can reduce lung metastasis without

32
changing tumour growth, and can increase survival of mice. These results make a strong case for
the effectiveness of miR-34a as a therapeutic agent.

33
Table 1.4- MiRNAs in prostate cancer preclinical studies
miRNA Target References
miR-15a, miR16-1 BCL2, CCND1, WNT3A Bonci et al. (137)
miR16 CDK1, CDK2 Takeshita et al. (152)
miR-17-3p Vimentin Zhang et al. (153)
miR-203 Runx2, Dlx5, ZEB2, Bmi, survivin Saini et al. (156)
miR-34a CD44 Liu et al. (162)
miR-221, miR-222 P27 Mercatelli et al. (136)

34
1.4.1.6 MiRNAs as Therapeutic Agents—Breakthroughs and Challenges
The association between the differential expression of miRNAs and cancer is now so
convincing that there has been considerable attention focused on applying these gene regulators
as therapeutic agents. Although the utilization of miRNA-targeted therapy has not yet been
implemented in clinical practice, miRNA-based therapeutics can potentially be applied as
effective therapeutics by utilizing them as either drugs or drug targets: miRNA molecules that act
as tumor suppressors or antagomirs of oncogenic miRNAs may serve as cancer treatments. This
approach offers a new paradigm for treating human diseases, one that can utilize the entire
human genome for therapeutic purposes. After establishment of a miRNA’s role in cancer
pathogenesis and identification of its targets, expression of the specific transcripts bearing the
complementary sequence of the miRNA can be manipulated by small nucleic acids that mimic or
antagonize miRNAs, using endogenous miRNA machinery.
MiRNAs are naturally occurring molecules, and so they benefit from million years of
evolutionary “fine tuning” of their function, and there are likely distinct advantages in applying
miRNAs as therapeutic agents over the current conventional drugs. Unlike other nucleic acid
therapeutics such as siRNAs, specificity for a single target is not the purpose for miRNA drugs,
since one miRNA can target multiple downstream effectors (163). Consequently miRNA-based
drugs have the advantage that they can target multiple genes in a pathway concurrently by either
inhibiting or mimicking a single miRNA. Also, multiple tumor-suppressive miRNAs can be used
simultaneously on one or a group of target genes to augment the effect of the therapy. For
example, if there is a mutation in the binding sequence of an oncogenic target, one miRNA may
not be able to bind to it, but combination of several miRNAs that target the same gene would

35
reduce the probability of mutation-induced resistance (164). Another advantage of using
miRNAs as drugs is that they are small in size, and are therefore less antigenic than protein-
coding gene replacement therapies (164).
Despite the great potential of miRNAs as anticancer drugs, there are some challenges that
need to be addressed in order to bring miRNAs into the clinic. The development of miRNA-
based therapies requires that miRNAs pertinent to cancer formation and progression be first
identified and validated. As previously mentioned, oligonucleotides or virus-based constructs can
be used to directly introduce miRNAs to the system, or drugs can be used to regulate miRNA
expression and processing (165). Perhaps the main hurdle in using miRNA-based therapy is
delivering the drug to cancer cells, whether locally in the prostate or to metastatic cells
disseminated throughout the body. Several delivering strategies are considered plausible, such as
incorporating the miRNAs within liposomes, or conjugating them to peptides that can penetrate
the plasma membrane (166). For example, a lipid-based system was used to deliver chemically
synthesized miR-34a, resulting in inhibition of tumor growth in mouse models of non-small-cell
lung cancer (167). Also, tandem arrays of miRNA mimics delivered by lentiviral vectors proved
effective against Bcr-Abl lymphoid leukemia (168). Although the multispecific nature of
miRNAs makes them very effective in regulating cellular processes associated with normal cell
function and neoplastic development, this property can also be a weakness of miRNA-based
therapy, as perturbing miRNA levels may affect the expression of unintended mRNA target.
Another major concern would be that the introduction of exogenous miRNAs may overwhelm
the RNA-induced silencing complex and inhibit the processing of other miRNAs that are
involved in normal cellular function (169). An additional challenge for developing miRNA-
based therapies is the issue of toxicity and safety. There is concern that miRNAs would affect

36
off-target genes, delivery with liposomes might be toxic (170), and high concentration of small
RNAs may cause liver damage (171). Ongoing and future clinical trials will be crucial in
assessing the safety of miRNAs as therapeutics.
1.4.1.6.1 MiRNAs in Clinical Trials
There are currently a number of major pharmaceutical companies that have miRNA
therapeutics programs. The most advanced miRNA therapy program is being done in liver,
benefiting from the fact that oligonucleotides administered systematically will largely localize to
the liver. In 2010, Santaris Pharma A/S, a biopharmaceutical company that focuses on the
development of RNA-targeted therapies announced that their drug miravisen (SPC3649), a miR-
122 inhibitor, has been advanced into Phase II studies. This drug is being used to treat Hepatitis
C virus (HCV) infections by sequestering miR-122, and thereby inhibiting the replication of
Hepatitis C virus. Miravirsen was the first miRNA-targeted drug to enter clinical trials. This drug
was developed using Santaris Pharma A/S Locked Nucleic Acid (LNA) Drug Platform. Data
from the drug’s phase I trial on healthy volunteers showed that this miRNA-targeted therapy is
well tolerated. Recently, the FDA gave the miRagen Therapeutics’ compound MGN—4893 that
targets miR-451 orphan drug status, a designation intended for drugs used for treatment of rare
conditions, and clinical trials for this compound are scheduled to begin in 2012. This drug is
being used to treat polycythemia vera, a myeloproliferative disease which is characterized by the
abundance of blood cells and platelets in the body.

37
1.4.1.6.2 Prostate Cancer-associated MiRNAs Currently in Clinical Trials
Recently, Mirna Therapeutics and researchers at the University of Texas MD Anderson
Cancer Center have had success in inhibiting prostate cancer tumour growth, decreasing lung
metastasis, and extending survival in mice by using miRNAs. This represents a very exciting
step towards the clinical use of miRNA-based drugs. As previously mentioned, Liu et al. showed
that miR-34a is under-expressed in prostate cancer stem cells, and systemic delivery of miR-34a
using a liposome-based delivery agent inhibited prostate cancer stem cells from replicating by
suppressing the adhesion molecule CD44 (162). This group hopes to advance miR-34a as a
treatment option for prostate cancer patients. Although impeding cancer in mice might be a lot
easier than in human patients, these results are indeed promising as miRNA-targeted therapeutic
tools that have clinical value.
Currently there are several observational clinical trials that aim to study miRNAs in
prostate cancer (Table 1.5) (clinical trials.gov). ‘Micro-RNA expression profiles in high risk
prostate cancer’, NCT01220427, is being conducted at Wurzburg University hospital. Its main
goal is to examine whether specific miRNA expression profiles are correlated with prostate
cancer outcome. Another trial ‘Molecular correlates of sensitivity and resistance to therapy in
prostate cancer’, NCT01050504, is based at the University of Washington. It aims to study
differences in gene expression patterns of miRNAs, in addition to other genes, in order to
discover new biomarkers and drug targets. Both of these trials are currently recruiting
participants. There are also two clinical trials that briefly study miRNA expression as a
secondary objective of their study. While there are no clinical trials that use miRNAs as a
treatment option for prostate cancer, the emerging field of miRNA-targeted therapeutics is

38
undoubtedly progressing and the vast body of knowledge from basic science and pre-clinical
work is reassuring.

39
Table 1.5- MiRNAs in prostate cancer clinical trials
Trial title Study type Institution Trial Identifier
Micro-RNA expression profiles in high risk prostate cancer Observational Wuerzburg University Hospital, Germany
NCT01220427
Molecular correlates of sensitivity and resistance to therapy in prostate cancer
Observational University of Washington NCT01050504
Trial of vaccine therapy in curative resected prostate cancer patients using autologous dendritic cells loaded with mRNA from primary prostate cancer tissue, hTERT and Survivin
Treatment Rikshospitalet University Hospital, Norway
NCT01197625
Phase II randomized study of combined androgen depripation comprising Bicalutamide and Goserelin or Leuprolide Acetate with versus without Cixutumumab in patients with newly diagnosed hormone-sensitive metastatic prostate cancer
Biomarker/ Laboratory analysis, Treatment
Saint Anthony's Hospital at Saint Anthony's Health Center, Illinois
NCT01120236

40
1.5 Characteristics of Malignant Transformation
Tumorigenesis is a multi-step process caused by genetic changes that result in the
progressive conversion of normal human cells into extremely malignant derivatives (172). This
cellular transformation is depended on an inherent or acquired genetic instability that leads to
numerous mutations, thus allowing the cell to acquire multiple malignant characteristics which
confer uncontrollable proliferative advantage to the malignant cell. These characteristics or
‘acquired capabilities’ that govern the transformation of normal human cells into cancer cells
were outlined by Hanahan and Weinberg in “The Hallmarks of Cancer” (173). There are over
100 different types of cancer, and these capabilities are shared by the majority of these cancers.
These features are: self-sufficiency in proliferative signals, evading anti-growth signals, evading
programmed cell death (apoptosis), increased replicative potential, sustained angiogenesis, and
invasion and metastasis (173) (Figure 1.2). Recently the authors have added four additional
capabilities as hallmarks of cancer. They are: genomic instability, inflammation, deregulation of
energy metabolism, and evading immune destruction (174).
Normal cells require stimulatory signals in order to transition from a resting state into a
proliferative state (173). These signals that are induced by growth factors, extracellular matrix
components and cell-to-cell adhesion molecules are transmitted into the cell by trans-membrane
receptors. Although neoplastic cells divide at a fast rate, they however have significantly less
dependence on external growth signals since they have achieved autonomy from their normal
microenvironment. They are multiple ways in which increased proliferation is sustained. The
growth factor cell-surface receptors with intracellular tyrosine kinase domains that stimulate cell
cycle progression are over-expressed in many cancers (175;176). Also, mutations can cause
structural changes in the receptors that causes sustained signalling independent of ligand

41
Figure 1.5- The Hallmarks of Cancer. The features involved in enabling the conversion of
normal human cells into malignant cancers. These traits are: self-sufficiency in growth signals,
insensitivity to anti-growth signals, evading programmed cell death, limitless replicative
potential, sustained angiogenesis, and tissue invasion and metastasis (Adapted from Hanahan and
Weinberg, 2000(177))

42
stimulation (178). Additionally, changes in the different downstream components of complex
signalling pathways may lead to growth signal autonomy (173). For example, in 25% of human
tumours, Ras proteins are structurally altered so that they can release mitogenic signals into cells
without stimulation from their upstream regulators (179).
Within a normal tissue, antigrowth signals bind to transmembrane cell surface receptors
that are coupled to intracellular signaling circuits to maintain cellular quiescence (173).
Neoplastic cells are insensitive to anti-proliferative signals, mostly through pathways involving
the retinoblastoma protein (pRb). When pRb is hypophosphorylated, it blocks proliferation by
sequestering E2F transcription factors that control the expression of genes associated with
progression from G1 into S phase (180).
The ability of cancer cells to resist programmed cell death—apoptosis—is also a
hallmark of cancer. Evading apoptosis can be acquired by cancer cells by a number of strategies.
One of the most common ways involves mutation of the p53 tumor suppressor gene, resulting in
the inactivation of the p53 protein in more than 50% of human cancers (173). The BCL-2 family
of proteins are also important apoptotic inhibitors that are over-expressed in cancers (181).
In order for a clone of cells to form a macroscopic tumor, cancerous cells need to have
unlimited replicative potential, contrary to normal cells that have a finite cell division cycles
before cell death (173). Neoplastic cells achieve unlimited multiplication by maintaining their
telomeres at a length above a critical threshold (182). Telomeres are region of repetitive
nucleotide sequences that protect the ends of eukaryotic chromosomes and are progressively
shortened during each round of DNA replication. Malignant cells maintain their telomere length
which permits the accumulation of mutant oncogenes and tumour suppressors and this genomic
instability can lead to immortalized cells.

43
Metastasis, the spread of cancer cells from the primary tumor mass to new parts in the
body, is the cause of 90% of human cancer deaths (183). The metastatic process occurs in
successive and multiple steps, starting with local invasion, followed by intravasation of cancer
cells into blood and lymphatic vessels, transportation of cancer cells through such vessels,
breaking out of cancer cells from the vessels into the parenchyma of distant tissues, formation of
micrometastases, and ultimately development of macroscopic tumors (184). Numerous classes of
transcription factors and proteins have been associated with increased invasiveness and
migratory capabilities of cancer cells, such as cell-cell adhesion molecule, cadherin (173).
The cells that make up the body’s tissues require the oxygen and nutrients
supplied by the vasculature in order to proliferate; consequently the processes that control new
blood vessel formation are critical for normal physiological functions, as well as some
pathological conditions including cancer (173). The development of new blood vessels during
embryonic development requires the assembly of angioblasts, immature endothelial cell
precursors, into a de novo capillary network. This process is called vasculogenesis and is
distinguished from angiogenesis, a process by which new blood capillaries are formed from pre-
existing vasculature. It was first reported by Judah Folkman in 1971 that tumors require vascular
networks to grow beyond 1-2 mm in diameter (185). The ability of a malignant cell to influence
its microenviroment to favor the formation of new blood vessels, and the potential of
angiogeneic processes as an effective anti-cancer therapy have been an area of intense interest.
The most well studied of the positive regulators of angiogenesis is vascular endothelial growth
factor (VEGF), which can directly activate mitogenic, survival, and anti-apoptotic properties in
vascular endothelial cells (186), mobilize hematopoetic cells including monocytes and
granulocyte-macrophage progenitor cells (187), which can promote angiogenesis (188). Also,

44
VEGF can cause vascular permeability, potentially resulting the formation of edema and thereby
facilitating the distant spread of metastases (189).
The emerging hallmarks of cancer whose validity needs to be further established include:
the ability to evade destruction by the immune system, and reprogramming of energy metabolism
in cancer cells in order to maintain the uncontrollable cell proliferation and division (184). In
addition, genomic instability and inflammation are considered enabling characteristics of
neoplasia (184). The body’s genomic surveillance system has the ability to identify and repair
mutations in the DNA, therefore spontaneous mutations are typically rare. However, some
mutant genotypes can give selective advantage to the cells harboring them, and cancer cells often
increase the rate of mutations (190). They can do so by increasing the sensitivity of cells to
mutagenic agents, and/or by disrupting genes that identify DNA damage and enable DNA repair
(184). Tumors are heavily infiltrated by the innate and adaptive cell types of the immune system
and thus mimic non-neoplastic inflammatory conditions (184). Inflammation can enhance
hallmark capabilities of tumorigenesis and progression by providing molecules to the tumor
microenvironment that promote proliferation, survival factors that limit cell death, pro-
angiogenic factors, and metastasis signals that lead to activation of EMT (191).

45
1.6 PROJECT RATIONAL
Given the importance of miRNAs in regulating gene expression in cells and their
involvement in tumor development and progression as demonstrated by the aberrant miRNA
expressions in various cancer subtypes, the hypothesis I studied at the onset of this thesis project
was that some miRNAs might play an integral role in the biology of prostate cancer. If such a
miRNA existed, and analysis of the miRNA and its downstream targets could establish this, it
would allow for the further studies into the possible role of the miRNA as a prognostic
biomarker of prostate cancer. While the importance of such a miRNA would be studied to
investigate its role during tumor progression, it would also be important to assess its potential as
a therapeutic agent — either as a drug or drug target. These considerations have significant
implications for current testing of miRNAs as the new class of anti-cancer agents in the clinical
setting.

46
1.7 THESIS OBJECTIVES AND ORGANIZATION
The objectives of this thesis were as follows: a) to establish whether a relationship could
be found between miR-221, prostate cancer prognosis, and TMPRSS2:ERG fusion gene; b) and
to investigate whether miR-182 is associated with enhanced metastasis and clinical progression
in prostate cancer, and to identify its target that could result in phenotypic changes.
Chapter 2 details studies demonstrating for the first time that miR-221 is down-regulated in
TMPRSS2:ERG fusion-positive prostate cancer. This study, published in 2011 (63), was the
first of its kind by providing a link between a miRNA and gene fusion expression, and it also
demonstrated a link between miR-221 and clinical progression in prostate cancer.
Chapter 3 demonstrates studies that validate miR-182 overexpression in prostate cancer, and
analyze its merit as a prognostic biomarker. These results illustrate the effects of miR-182
overexpression on metastatic potential in prostate cancer cells, and identify FOXO1 as a target of
miR-182. This study may be pertinent to further clinical exploitation of miR-182 as a biomarker
or perhaps therapeutic tool for prostate cancer.
Finally, appendix 1 which was recently published in the Journal of visualized experiments(192),
presents quantitative Real-Time PCR methodology as an assay for accurate quantification of
miRNA levels in prostate tumor tissues.

47
2 CHAPTER 2: MIR-221 IS DOWNREGULATED IN TMPRSS2:ERG FUSION-POSITIVE PROSTATE
CANCER
A version of this chapter has been published in Anticancer Research as Gordanpour A,
Stanimirovic A, Nam RK, Moreno CS, Sherman C, Sugar L, Seth A. (2011) 31(2):403-10.

48
2.1 ABSTRACT
Expression profiling studies using microarrays and other methods have shown that
miRNAs are dysregulated in a wide variety of human cancers. The up-regulation of miR-221 has
been reported in carcinomas of the pancreas, breast, and papillary thyroid, as well as in
glioblastoma and chronic lymphocytic leukaemia. In prostate cancer, however, down-regulation
of miR-221 has been repeatedly confirmed in miRNA expression studies. Also unique to prostate
cancer, and found in more than 50% of patients, is the aberrant expression of a known oncogene,
the TMPRSS2:ERG fusion. To date, there has been no published study describing miRNA
associations in prostate tumours that over-express the ERG oncogene from the TMPRSS2:ERG
fusion transcript. Herein we report that in a large and diverse cohort of prostate carcinoma
samples, miR-221 is down-regulated in patients with tumours bearing TMPRSS2:ERG fusion
transcripts, thus providing a link between miRNA and gene fusion expression.

49
2.2 INTRODUCTION
Prostate cancer exists along a biological continuum that ranges from clinically
insignificant to extremely aggressive disease. Although clinically localized prostate cancer is
largely manageable by surgery, with patients rarely developing clinical recurrence, recurrent
disease remains essentially incurable. Because of this difference in treatment options, it is critical
to establish specific biomarkers to differentiate between different stages of disease not only to
determine presence of disease, but also to differentiate between indolent and aggressive cancer.
The most common class of prostate cancer biomarker thus far is gene fusions resulting from
chromosomal rearrangements. Numerous recurrent chromosomal rearrangements have been
identified that are generally characterized by the fusion of various 5’ regulatory elements to E
twenty-six (ETS) transcription factors, leading to high expression of these oncogenic
transcription factors. Transmembrane protease serine 2: ETS-related gene (TMPRSS2:ERG),
present in over 50% of all prostate cancer, is the most commonly identified fusion gene (193).
TMPRSS2 is an androgen-responsive, prostate-specific serine protease of unknown
function, and ERG is a member of the ETS transcription factor family and is rarely detected in
normal prostate tissue (194). The consequences of ERG over-expression, and its correlation to
the progression of prostate cancer remains unclear. What is known is that androgen stimulation
induces the over-expression of an mRNA containing the ERG ORF and 3’ UTR when ERG is
fused to the TMPRSS2 5’ UTR. In vitro studies have shown that over-expression of ERG
stimulates cell migration and invasion, while its knockdown decreases the invasive properties of
VCaP cells (195). In addition, it has been shown by our laboratory and others that
TMPRSS2:ERG fusion mRNAs are present in prostate tumours but seldom in normal prostate
cells. Thus, detection of abnormally high ERG expression could, at least theoretically, be a

50
potential diagnostic and/or prognostic marker for prostate cancer. Although the chromosomal
alterations seem to be important in the development of prostate neoplastic development, they
alone may not be sufficient to induce cancer formation (194).
An active area of prostate cancer research is to find biomarkers that are predictive of
recurrence in patients in order to aid oncologists in treatment or non-treatment decisions.
TMPRSS2:ERG has been repeatedly, but not unanimously, associated with a poorer prognosis in
prostate cancer patients and our laboratory, in collaboration with other groups, has previously
demonstrated that prostate cancer patients with the TMPRSS2:ERG gene fusion have a higher
risk of recurrence (196), while others have reported no association between this chromosomal
rearrangement and clinical outcome (197). The reasons for this lack of congruity between
findings are unclear and the value of TMPRSS2:ERG as an independent prognostic biomarker of
prostate cancer remains contentious; however, it is possible technical discrepancies may be an
important factor, such as institutional inconsistencies in disease staging, or statistical variations,
such as disparate cohort sizes (193). In fact, studies that show an absence of clinical correlation
between fusion and prognosis highlight the importance of finding out how the fusion may be
epigenetically regulated.
MiRNAs have received considerable attention in recent years as possible biomarkers not
only in prostate cancer, but also in various cancer subtypes. They are small, single-stranded,
noncoding RNA molecules that regulate mRNA function by binding to the 3’ UTR of mRNAs to
which they are partially complementary, thereby repressing translation (198). MiRNAs are
known to be involved in almost every cellular function, including early development (86),
differentiation (199), apoptosis (200), and cell cycle regulation (201); as such, it is not surprising
that miRNAs have also been linked to cancer, since misregulation of any of these important

51
cellular functions can lead to cancer (202). Aberrant expression of miRNAs has been found in
prostate cell lines, xenografts, and clinical tissues. Since the majority of cancer deaths are
caused by complications from metastasis, miRNAs that specifically regulate cancer metastasis
(metastamirs) are of particular interest. The study of miRNAs as biomarkers and their exact
involvement in the formation and/or progression of prostate cancer is still at its early stages, and
more research is needed to evaluate the potential use of miRNAs as diagnostic and prognostic
markers of prostate cancer.
Given that miRNAs and fusion genes have been independently linked to prostate disease
and progression, we set out to study the possible connection between miRNA regulation, prostate
cancer recurrence, and TMPRSS2:ERG gene fusion status. We used a large cohort of men with
clinically localized prostate cancer who were treated with radical prostatectomy and had long-
term follow-ups. We assessed whether miR-221, a metastasis-promoting miRNA (203) located
on the X chromosome that is differentially expressed in recurrent prostate cancers, is also
associated with TMPRSS2:ERG fusion gene. Thereby, our study may prove applicable to future
use of these regulators as surrogate biomarkers of prostate cancer.

52
2.3 MATERIALS AND METHODS
2.3.1 Study Participants and Prostate Sample Collection
Tumour samples were obtained after radical prostatectomy from prostate cancer patients
who had surgery at Sunnybrook Health Sciences Centre (Toronto, Canada) between 1998 and
2006. As described by Nam et al., following radical prostatectomy, a midsection of the specimen
was snap-frozen in liquid nitrogen, and stored at −80°C until extraction of RNA (196). Most
tumours were not visible within the prostatectomy specimen, and thus, the samples obtained
from the prostate were considered to be random. The banked slices of specimens were
photocopied, oriented (anterior, posterior, right and left), quadrisected and cut into 5 mm
sections on a cryostat. The sections were stained with hematoxylin and eosin (H&E) and then
reviewed by the pathologist. The areas of tumour were marked on the stained slides and on the
photocopied diagram. The marked areas were used to extract the tissue for total RNA extraction.
All research was conducted with the approval of Sunnybrook Health Sciences Centre Research
Ethics Board.
2.3.2 Patient Follow-up
In 1998, a prostate tumour tissue bank was established at Sunnybrook hospital. Clinical
data and follow-up information were collected prospectively. The medical records were
thoroughly reviewed using standardised data entry forms by trained data abstractors and stored
within a prostate cancer-specific database. Clinical follow up consisted of four assessments in the
year following surgery, two assessments in the second year and one assessment every year
thereafter. At each follow-up, patients had a prostate-specific antigen (PSA) test, and clinical
evaluation. Biochemical recurrence was defined as a rise in blood levels of PSA in prostate

53
cancer patients on two consecutive measurements after radical prostatectomy. Data on the
following characteristics were available for each patient: Age, family history of prostate cancer,
PSA score, Gleason grade, tumour stage, seminal vesicle invasion, surgical margins (categorized
as positive or negative), metastasis (categorized as absence or presence), and TMPRSS2:ERG
translocation (categorized as presence or absence). The clinical demographics of the patients
used in the study are summarized in Table 2.1.
2.3.3 Quantitative Real-time Polymerase Chain Reaction (PCR)
Total RNA from prostate tumours was extracted with TRIzol reagent (Invitrogen
Corporation, Carlsbad, CA, USA) according to the manufacturers’ instructions. One microgram
of total RNA was reverse-transcribed using the
QuantiTect Rev. Transcription Kit (Qiagen GmbH, Hilden, Germany). Quantitative real-time
PCR was performed in triplicate by using QuantiTect SYBR Green PCR Kit (Qiagen GmbH,
Hilden, Germany) on LightCycler Real-time PCR system (Roche Applied Science, Mannheim,
Germany). The miRNA level was normalized by house keeping gene RNU6B. Optimized
miRNA-specific primers for miR-221 as well as for the endogenous control RNU6B are also
commercially available (miScript Primer Assays; Qiagen). The relative amount of miR-221 in
each sample was calculated based on the crossing-point analysis (Relquant, version 1.01).
2.3.4 RT–PCR
Total RNA was extracted from the frozen prostate cancer tissue by homogenization in
Trizol (Invitrogen Corporation) followed by ethanol precipitation. RNA pellets were dissolved in
RNase-free H2O and quality determined using 2100 Bioanalyzer (Agilent Technologies, Inc.,

54
Santa Clara, CA, USA). The presence of TMPRSS2:ERG was assayed using RT-PCR as
previously described (196). All reactions were performed with two primer sets that yield a 125-
bp (F-TAGGCGCGAGCTAAGCAGGAG, R-GTAGGCACACTCAAACAACGACTGG) and
595-bp (F-CAGGAGGCGGAGGCGGA, R-GGCGGTTGTAGCTGGGGGTGAG) product.
2.3.5 Statistical Analysis
Results were statistically analyzed using Prism v4.0 software (GraphPad Software Inc.,
La Jolla, CA, USA). Scatter plots were analyzed using Student’s t-tests which were two-tailed
and unpaired. P-values less than 0.01 were considered statistically significant.

55
2.4 RESULTS
2.4.1 Patient Demographics
RNA was extracted from and TMPRSS2:ERG status analyzed in 170 radical
prostatectomy samples. The distribution of clinical characteristics such as PSA, Gleason grade,
pathological stage, and surgical margin status are described in Table 2.1. Of these 170 patients,
we opted to eliminate 17 from our study because certain clinical information was missing. Table
2.2 summarizes the fusion gene status and complete clinical information of the 153 remaining
patients. The age range at diagnosis of the 153 patients was 31–75 years. The average follow-up
lasted 5.4 years in the fusion-negative group, and 4.69 years in the fusion-positive group.
Among the cohort, 54.2% of the patients had tumours confined to the prostate gland and 69.3%
of tumours were of Gleason score 7.
2.4.2 miR-221 is Downregulated in TMPRSS2:ERG Fusion Gene-positive Prostate Tumours
Due to the high prevalence of TMPRSS2:ERG in prostate cancer and its association with
higher chances of recurrence and poorer prognosis, we hypothesized that miR-221, an miRNA
previously linked to metastasis and recurrence in prostate cancer, may be associated with the
fusion gene. To test our hypothesis, TMPRSS2:ERG status was determined for 153 radical
prostatectomy samples by using RT–PCR and sequencing using random and oligo-dT primers.
Prostate samples from 83 out of 153 (54.2%) patients were found to be positive for transcripts of
TMPRSS2:ERG, while 70 (45.8%) lacked the fusion gene (Table 2.2). TMPRSS2:ERG fusion-
positive samples produced the expected 125 or 595 bp bands depending upon which primer set
was used, as previously described (196). To analyse whether miR-221 expression was associated
with the presence of the fusion gene, quantitative RT-PCR was performed, which showed that

56
the mean expression level of miR-221 was significantly lower (p<0.01) in fusion-positive
(4.52±0.34) compared with fusion-negative (7.70±1.09) tumours (Figure 2.1). Gleason grade
and surgical margin status were not different between fusion-positive and fusion-negative
populations (Table 2.2).

57
Figure 2.1- MiR-221 is suppressed in prostate tumours with TMPRSS2:ERG

58
2.4.3 Low MiR-221 Expression is Associated with Metastasis and Biochemical Recurrence of Prostate Tumour
To further analyse the association of miR-221 levels with the aggressiveness of prostate
cancer, we categorized the tumour samples into two subgroups: those from patients who had
metastasis at the time of radical prostatectomy and/or had biochemical recurrence in follow-up
years and those from patients with non-metastatic and non-recurrent disease. Long-term follow-
up information on all 153 patients was not available, as some had moved to other hospitals or did
not follow-up with their appointments. Of the 153 tumours, 99 were from patients with sufficient
follow-up, more than 5 years on average (Table 2.3). These 99 patients were divided into two
groups: those with recurrent and/or metastatic tumours, and those with non-recurrent and non
metastatic tumours.
Quantitative RT-PCR was carried out in order to analyse the expression levels of miR-
221 in prostate tumours. The qRT-PCR analysis using the 55 recurrent and/or metastatic samples
and 44 non-recurrent and non-metastatic samples confirmed that the mean expression level of
miR-221 was down-regulated (p<0.01) in the tumours with metastasis and/or recurrence
(3.89±0.39) compared to tumours with no metastasis or recurrence (6.32±0.73) (Figure 2.2).

59
Figure 2.2- MiR-221 is suppressed in prostate tumours with metastasis and/or biochemical
recurrence.

60
2.4.4 MiR-221 Levels are Lower in Recurrent/Metastatic TMPRSS2:ERG Fusion-negative Tumours
We wanted to further analyse the differential expression of miR-221 in relation to both
genetic aberrations and clinical parameters: specifically, TMPRSS2:ERG fusion status,
metastasis and recurrence. All 99 tumour samples from patients with long-term follow-ups were
studied. We found that tumours positive for the oncogenic TMPRSS2:ERG had down-regulated
miR-221 levels with or without metastasis and biochemical recurrence (4.20 and 4.22,
respectively) (Figure 2.3). In TMPRSS2:ERG fusion-negative tumours, on the other hand, miR-
221 expression differs with clinical status: miR-221 levels are significantly lower in patients with
recurrence and/or metastasis (3.52) than in those with no recurrence or metastasis (8.62) (Figure
2.3).

61
Figure 2.3- MiR-221 levels are lower in recurrent/metastatic TMPRSS2:ERG
fusion-negative tumours

62
2.5 DISCUSSION
The potential utility of the TMPRSS2:ERG fusion product as an independent prognostic
marker for patients with clinically localized prostate cancer is becoming clearer every year.
Initial studies comparing clinico-pathological parameters (196;204-207) and prognostic
significance (196;204;205;208-210) of this fusion gene showed conflicting results. Some studies
showed no correlation between histological grade (Gleason score) (204), while others found
positive associations (207;208), and yet others demonstrated correlations between fusion status
and tumour stage (205;206). Wang et al. examined 119 patients for fusion status from a case
control approach and found significant correlations with tumour stage, but no associations were
found with early recurrence (48). Furthermore, Lapointe et al. in another case control study
found no correlations with any clinico-pathological parameter and recurrence-free survival (204).
However, two cohort studies of men with clinically localized prostate cancer who did not
undergo treatment (i.e. watchful waiting) showed that men who had TMPRSS2:ERG fusion had
lower prostate cancer-specific survival compared to men without fusion expression (208;209).
Patients managed and selected for watchful waiting from these cohorts have different baseline
distributions in grade, stage and PSA level to patients treated with surgery and may not be
comparable. Initially, it was unclear whether the TMPRSS2:ERG gene fusion is only a surrogate
marker for established prognostic factors of grade and stage, or whether it is an independent
molecular-based marker for disease recurrence with no association with grade or stage,
particularly for patients who are candidates for surgery for clinically localized prostate cancer.
Since that time, other groups, including our own, have found robust associations. More
than two dozen high-impact reports have found clinically significant links to fusions in greater

63
than 50% of over 1500 samples of clinically localized prostate cancer (193). Furthermore, a
recent publication by Carver et al. showed that prostatic intraepithelial neoplasia (PIN) is
induced in host prostates by transgenic over-expression of ERG (211), suggesting that high
frequency of ETS genetic rearrangements and subsequent overexpression of ETS factors may
represent a crucial event in prostate tumourigenesis (212).
In addition to TMPRSS2:ERG, there are increasing reports of miRNAs or specific
miRNA signatures that correlate with a wide range of clinico-pathological features in prostate
cancer. Many miRNAs are found to be predictive of patient clinical outcome and/or response to
treatment suggesting that miRNAs can be used as diagnostic or prognostic/predictive biomarkers
(213). Investigations of miRNA dysregulation in prostate cancer typically compare miRNA
expression profiles in prostate cancer versus normal/benign tissues. Two studies have included
prostate cancer among other human cancer types, finding overall down-regulation of miRNAs in
solid tumours compared with normal tissues (214;215). A study comparing prostate tumours
with benign prostate tissue found 37 down- and 14 up-regulated miRNAs (216). Others have
found either overall down-regulation of miRNAs (217) or general up-regulation of miRNAs
(218). Most significantly for prostate cancer, Spahn et al. assessed miRNA expression profiles in
lymph node metastasis of prostate carcinoma and found that miR-221 down-regulation was a
hallmark of metastasis (219). In a larger patient group, they found that the expression of miR-221
was associated with prostate cancer progression and clinical recurrence.
miR-221 is known to be over-expressed in tumours of the breast (220), pancreas (221),
glioblastoma (222), papillary thyroid carcinoma (223), and chronic lymphocytic leukaemia
(224). Up-regulation of miR-221 in chronic lymphocytic leukaemia was found to be associated
with a poor prognosis. Functional studies have found that miR-221 induces down-regulation of

64
p27 and that inhibition of miR-221 impairs tumour formation in xenografted mice (225). Indeed,
high levels of miR-221 are required in many different cancer types to inhibit the expression of
p27, and stimulate proliferation.
However, none of these studies have looked for miRNA associations in prostate tumours
that overexpress the ERG oncogene from the TMPRSS2:ERG fusion transcript. To that end, we
investigated miR-221 expression in tumours bearing the TMPRSS2:ERG fusion transcript. Using
a large and diverse cohort of prostate carcinoma samples, including patients with or without the
fusion gene, as well as patients with different clinical progressions, we found that miR-221 is
down-regulated in both patients with TMPRSS2:ERG fusion gene, and in patients with more
aggressive tumours.
As a major centre for prostate cancer, we assembled a large database and tissue bank
from patients undergoing a prostate biopsy to determine the presence of prostate cancer and
surgery for clinically local prostate cancer. This resource has been used for study of multiple
genetic and serological markers for prostate cancer diagnosis and prognosis (196;205;226-230).
The resource includes men who underwent a prostate biopsy because of an abnormal PSA or
digital rectal examination and preserves outcome data, along with DNA, plasma, and paraffin-
embedded tumour samples. TMPRSS2:ERG status was determined as presence or absence by
RT-PCR of RNA for samples used in this study and Table I shows the range of clinical and
molecular characteristics.
Almost half of the samples (54.2%) bear TMPRSS2:ERG fusion transcripts (Table 2). We
found that miR-221 expression in fusion-positive tumours is 1.7-fold lower than in fusion-
negative samples, with a tighter distribution (Figure 2.1, Table 2.2). Over-expression of the ERG
transcription factor is a powerful inducer of prostate tumourigenesis, as we and others have

65
reported. Similarly, one would also expect to see down-regulation of tumour suppressors, such as
miR-221 in at least some prostate tumours. It is therefore possible that the ERG transcription
factor or one of its target genes could directly down-regulate miR-221.
A subset of 99 cases from the full cohort had enough clinical follow-up with which to
examine the long-term consequences of miR-221 dysregulation in prostate cancer patients (Table
2.3). Average miR-221 expression was 1.6-fold lower in samples from tumours that recurred or
metastasized after surgery (Figure 2). Previously we found that the TMPRSS2:ERG fusion is also
strongly associated with recurrence. That report and our finding here that miR-221 is reduced in
fusion-positive tumours suggests that they have a cumulative effect on tumour aggressiveness. It
may be that TMPRSS2:ERG fusion has the stronger effect as we found that tumours positive for
the oncogenic TMPRSS2:ERG have lower levels of miR-221 regardless of recurrence status
(Figure 2.3).
In numerous other cancer types, miR-221 is up-regulated and acts to induce proliferation
and tumour formation by down-regulating the p27 tumour suppressor. The role of miR-221 in
prostate cancer appears to be different than in other cancer types, as reported here and by others
(219). Although the Spahn group did not examine the mechanisms, they found that miR-221 is
down-regulated in metastatic tumours (219). We also found down-regulation of miR-221 in
prostate tumours and that such down-regulation can occur in the absence of TMPRSS2:ERG.
Although results shown in Figure 2.1 suggested a link between fusion status and miR-221 status,
direct dependence of miR-221 expression on TMPRSS2:ERG in every tumour seems unlikely
given the independence of miR-221 down-regulation in fusion-negative tumours.
This is the first study to reveal that miR-221 down-regulation in prostate cancer is
associated with the presence of the oncogenic TMPRSS2:ERG fusion transcript. Understanding

66
how this association leads to greater metastasis and biochemical recurrence will facilitate
understanding of prostate cancer biology. More practically, a deeper molecular understanding of
the mechanisms of prostate cancer genesis and maintenance will eventually provide therapeutic
interventions designed to affect the unique characteristics of individual prostate tumours.

67
Table 2.1- Clinical demographics of prostate cancer cases used in this study. Cohort clinical
characteristics for 170 prostate cancer patients initially used in this study are summarized. Data on the
following characteristics were available for each patient: Age, PSA score, Gleason grade, seminal vesicle
invasion, surgical margins (categorized as positive or negative), and biochemical recurrence.
Table 2.1- Clinical demographics of prostate cancer cases used in this study
Total
Cohort size (n) 170
Biochemical recurrence 58
Range of Follow-up (years) 1-11
Range of Age (years) 38-83
Preoperative PSA (ng/ml)
Average 9.2
Range 0.8-43.0
Gleason score
6 31 (18.2%)
7 111 (56.3%)
8-9 14 (8.2%)
Unknown 14 (56.3%)
Pathologic stage
Seminal vesicle invasion 17 (10%)
Positive margins
No 86 (50.6%)
Yes 84 (49.4%)

68
Table 2.2- miR-221 is associated with TMPRSS2:ERG fusion gene status. Cohort
clinical characteristics for 153 prostate cancer patients are listed for TMPRSS2:ERG
fusion positive and negative patients. Age, years of follow-up, Gleason score, surgical
margins, treatments, and number of patients with metastasis in both fusion-positive and -
negative groups, along with miR-221 expression levels are listed.
Table 2.2- MiR-221 and TMPRSS2:ERG fusion gene status
TMPRSS2:ERG
Total Positive Negative
Cohort size (n) 153 83 70
Age at diagnosis, Range (years) 31-75 44-75 31-73
Average follow-up (years)
4.69 5.4
Gleason score
6 33 (21.6%) 19 (22.9%) 14 (20%)
7 106 (69.3%) 58 (69.9%) 48 (68.6%)
8-9 14 (9.1) 6 (7.2%) 8 (11.4%)
Positive margin
No 83 (54.2%) 45 (54.2%) 38 (54.3%)
Yes 70 (45.8%) 38 (45.8%) 32 (45.7%)
Initial treatment
Surgery only 111 (72.5%) 60 (72.3%) 51 (72.9%)
Surgery + adjuvant treatment 42 (27.5%) 23 (27.7%) 19 (27.1%)
Metastasis
7 2
Average miR-221 level
4.52 7.70

69
Table 2.3- miR-221 is predictive of prostate cancer recurrence. Cohort clinical
characteristics for 99 prostate cancer patients are listed for patients with recurrent and/or
metastatic disease and those with non-recurrent and non-metastatic disease. Age, years of
follow-up, PSA, Gleason score, surgical margins, seminal vesicle invasion, and miR-221
expression levels are summarized.
Table 2.3- MiR-221 as a prognostic factor of prostate cancer recurrence
Total
Recurrent and/or
metastatic
Non recurrent and
Non-metastatic
Cohort size (n) 99 54 45
Age at diagnosis, range (years) 44-75 50-75 44-70
Average follow-up (years) 5.33 5.23 5.45
Preoperative PSA (ng/ml)
Average 9.24 10.36 7.73
Range 0.8-42.99 3-42.99 0.8-27
Gleason score
6 17 2 15
7 73 43 30
8-9 9 9 0
Positive margins
Yes 61 47 14
No 38 7 31
Seminal vesicle invasion T3b 16 16 0
Average miR-221 level 3.89 6.32

70
3 Chapter 3: ABERRANT MIR-182 EXPRESSION ENHANCES THE METASTATIC POTENTIAL OF PROSTATE CANCER AND PREDICTS CLINICAL RECURRENCE

71
3.1 ABSTRACT
Despite great advancements in prostate cancer management, finding new biomarkers to
accurately evaluate the aggressiveness of the disease and to determine which prostate cancers are
likely to recur following radical prostatectomy remains a major challenge. Using a large cohort
of primary prostate carcinomas, we determined here that miR-182, which has been previously
linked to increased metastatic potential in other cancer subtypes, is highly expressed in prostate
tumor samples compared to normal prostate tissue, and is its elevation is associated with higher
Gleason grade, seminal vesicle invasion, and biochemical recurrence. Ectopic expression of
miR-182 in prostate cell line LNCaP enhanced their motility and invasiveness, demonstrating its
direct involvement in metastatic progression. The effects of miR-182 are likely mediated, at least
in part, through downregulation of a tumor suppressor Forkhead box O1 (FOXO1), which has
been previously reported to be a direct target of miR-182. Herein, we show that miR-182
negatively regulates FOXO1 in LNCaP cells, and FOXO1 is inversely correlated with miR-182
expression in primary prostate cancer specimens. Our findings suggest a mechanism for
increased aggressiveness and recurrence in prostate cancer by miR-182 which may be valuable
in future therapeutic interventions.

72
3.2 INTRODUCTION
Cancer metastasis is the major cause of morbidity and death in patients with prostate
cancer. The current prognostic indicators for prostate cancer survival, blood PSA level and
Gleason grade are not precise in predicting clinical progression following surgical radical
prostatectomy. It is thus imperative to identify genes or gene products that may be used as
prognostic biomarkers. MicroRNAs (miRNAs) are involved in critical steps of metastatic
progression, such as migration, invasion, and angiogenesis, and their aberrant expressions have
been reported in numerous cancer subtypes—including prostate cancer (231). MiRNAs are
endogenously expressed, ~18-25 nucleotide, single-stranded RNA molecules. They regulate gene
expression at the post-transcriptional level by mainly binding to the 3’-untranslated region
(UTR) of a target messenger RNA (mRNA). MiRNAs can regulate gene expression by inhibiting
protein translation of a target mRNA (71), and also by degrading their target mRNA by affecting
its stability, thereby inhibiting protein synthesis indirectly (72;73). MiRNAs that are
overexpressed in tumors often target tumor suppressors associated with various steps of the
tumor progression, and identifying these miRNAs may be vital in the management of disease.
MiR-182, a member of the miR-96/-182/-183 cluster (232), has been associated with
metastatic progression in various tumor types. In particular, miR-182 has been shown to promote
invasion and migration of melanoma cells, and to enhance their metastatic ability by directly
repressing Forkhead Box O3 (FOXO3) and Microphthalmia-associated Transcription Factor-M
(MITF-M) (233). In breast cancer cells, miR-182 appears to regulate cell proliferation and
survival by directly targeting Forkhead box O1 (FOXO1) (234). Although some genome-wide
profiling studies of miRNA expressions have revealed that miR-182 may be aberrantly expressed

73
in prostate tumors (235;236), these findings are yet to be validated. Moreover, there is yet no
evidence that miR-182 regulates metastasis and affects clinical progression in prostate cancer.
For these reasons, we investigated the possible role of miR-182 in prostate cancer
progression, with the hope of gaining more insights into the mechanisms of prostate cancer
pathogenesis. We analyzed the expression levels of miR-182 in a large cohort of primary
prostate tumors in various clinical states, normal prostate tissues, as well as several prostate cell
lines. We found that miR-182 is overexpressed in human prostate tumors compared to normal
prostate tissue, and miR-182 levels rise with increasing Gleason grades in tumor samples.
Further analysis of human tumor specimens revealed that miR-182 levels are significantly higher
in patients who have seminal vesicle (SV) invasion and biochemical recurrence after
prostatectomy. Furthermore, we demonstrate that ectopic expression of miR-182 enhances the
tumorigenic, migratory, and invasive properties of LNCaP cells. We identified a tumor
suppressor FOXO1, which has been linked to regulation of migration and invasion in prostate
cancer (237), as a miR-182 target in prostate cancer. Indeed, in LNCaP cells miR-182
overexpression reduced FOXO1 protein levels similar to FOXO1-specific siRNA. Furthermore,
in primary prostate carcinoma specimens miR-182 expression has an inverse correlation with
FOXO1 expression. We also show that FOXO1 mRNA levels are lower in recurrent prostate
cancer, and higher FOXO1 levels correlate with increased time of recurrence free survival. These
findings may be applicable to future exploitation of miR-182 as a prognostic biomarker of
prostate cancer, as well as a potential therapeutic target.

74
3.3 MATERIALS AND METHODS
3.3.1 Cell Lines
The nontumorigenic human prostatic epithelial cell line, RWPE-1, and human prostate cancer
PC-3, Du145, LNCaP, and VCaP cell lines were obtained from the American Type Culture
Collection. LNCaP cells were stably transduced with the pLemiR lentiviral vector (Open
Biosystems) using standard techniques. The base medium for RWPE-1 cells is provided by
Invitrogen as Keratinocyte Serum Free Medium (K-SFM; Gibco). This kit is supplied with the
two additives required to grow this cell line: bovine pituitary extract and human recombinant
EGF. PC-3 cells are maintained in F-12K Medium, DU145 in Eagle's Minimum Essential
Medium, VCaP in Dulbecco's Modified Eagle's Medium, and LNCaP and its transduced
derivative cells in RPMI-1640 Medium, all supplemented with 10% fetal bovine serum (FBS)
(Life Technologies-Invitrogen Corp.). Cells were incubated at 37ºC and 5% CO2 in a humidified
incubator.
3.3.2 Quantitative Real-Time PCR
Total RNA was extracted from prostate tissues using TRIzol Reagent (Invitrogen)
according to the manufacturers’ instructions. 1 µg of total RNA was reversed transcribed using
QuantiTect Reverse Transcription Kit (Qiagen). Quantitative Real-Time PCR analysis of miR-
182, miR-93, and miR-98 was performed in triplicate using miScript SYBR Green PCR Kit
(Qiagen) on LightCycler Real-time PCR system (Roche applied science). The miRNA level was
normalized by a reference gene, RNU6B. Primers for each miRNA as well as for RNU6B were
commercially purchased (miScript Primer Assays, Qiagen). For genes other than miRNAs, two
primer sets were used. The Primers used are as follows: FOXO1 (Forward:
AAGAGCGTGCCCTACTTCAA, Reverse: CTGTTGTTGTCCATGGATGC), Beta-actin

75
(Forward: GGAGAATGGCCCAGTCCTC, Reverse: GGGCACGAAGGCTCATCAT). The
relative amount of genes in each sample was calculated based on the crossing-point analysis
(RelQuant, version 1.01). For semiquantitative end point PCR analysis, cDNA was subjected to
40 rounds of amplification, and PCR products were electrophoresed and visualized in ethedium
bomide-stained 1.5% agarose gels.
3.3.3 Soft Agar Assay
The bottom layer was obtained by covering 6-well dishes with 1 ml of 0.8% agarose in
Dulbecco's Modified Eagle Medium 2X (DMEM 2X) containing 10% Fetal bovine serum (FBS).
After an hour, 1.2x104 stably transduced LNCaP cells with either empty vector or with miR-182-
expressing vector were seeded in triplicate in 1 ml DMEM 2X containing 0.5% agarose and 10%
FBS. After two weeks, colonies were photographed and scored.
3.3.4 Wound Healing Assay
LNCaP Cells were seeded in six-well culture plates and grown to confluence forming a
monolayer covering the surface of the entire well. After cells were serum-starved in serum-free
RPMI for 18 h, the wound was created in the center of the cell monolayer by the gentle removal
of the attached cells with a 10 µl pipette tip. Debris was removed by PBS wash, and the cells
received fresh RPMI with 10% FBS and 10 mmol/L of hydroxyurea. Cells were photographed
with a Zeiss Axiovert 200M inverted microscope at the intervals of 0, 24, and 72 hours.
3.3.5 Transwell Migration Assay
A suspension of 1x105
stably transduced LNCaP cells with either empty vector or with
miR-182-expressing vector in 0.5% FBS medium was added to BD cell culture inserts with 8-μm
porous membrane coated with 20% FBS (BD Biosciences, Bedford, MA). Cells were incubated

76
for 2-5 days at 37°C in a 5% CO2 incubator. To quantify migrating cells, cells remaining on the
top-side of the membrane were removed using a cotton-tipped swab, and cells that had migrated
to the underside were visualized with a Zeiss Axiovert 200M inverted microscope and
photographed.
3.3.6 Invasion Assay
A suspension of 1x105 cells in RPMI media (600 µL) containing 1% serum was added to
transwell Boyden chamber containing 8.0 µm pore size filters coated with matrigel (BD
Biosciences, Bedford, MA). The lower chamber was filled with 1.5 mL of media with 10% FBS
as a chemo-attractant. Cells were incubated for 5 days under standard culture conditions. Tumor
cells remaining on the top-side of the membrane were removed and cells that had migrated to the
underside were visualized under a Zeiss phase contrast microscope, and photographed.
3.3.7 Western Blotting
Total cell extracts were prepared from subconfluent cells using Nonidet P-40 lysis buffer
(20 mM HEPES-KOH [pH 7.6], 100 mM KCl, 5 mM MgCl2, 2 mMdithiothreitol, 0.25%
Nonidet P-40, 2 μg/ml leupeptin, 2 μg/ml pepstatin, and 102 μg/ml cycloheximide). Lysates were
mixed with 2X SDS-loading buffer containing 10% β-mercaptoethanol, boiled for 5 min, and
loaded onto 12% SDS-polyacrylamide gel for electrophoretic separation. The proteins were
transferred onto a nitrocellulose membrane (Bio-Rad Laboratories, CA) at 300 miliamps for 40
minutes at room temperature (RT). Membranes were then blocked for nonspecific binding with
5% non-fat dry milk in TBST (TBS with 0.1% Tween 20; pH 7.4) for 1 hour, and incubated with
the primary antibodies overnight at 4ºC. The following monoclonal and polyclonal antibodies
were used: anti-FOXO1 and anti-FOXO3a (Cell Signaling, USA); anti-MiTF (abcam, USA);

77
anti-β-Tubulin (Sigma). After washing with TBST, membranes were incubated with species-
specific secondary antibody conjugated to horseradish peroxidase enzyme for 1 hour at RT, and
washed three times with TBST. Proteins were detected using an enhanced chemiluminescence
detection reagent kit (Amersham Biosciences Inc.) and Kodak X-OMAT-AR film for
autoradiography.
3.3.8 Transfection of SiRNA and MiRNA Inhibitors
1.5 x 105 cells were seeded per well in a 24-well plate. 5 nM of siRNA (control and
FOXO1 siRNA) and 50 nM of miRNA inhibitor (control and anti-miR-182) from Qiagen were
diluted into 100 μl of serum free medium. Subsequently, 3 μl of HiPerFect Transfection Reagent
(Qiagen) was added to the mixture and incubated at room temperature for 10 min. The
transfection complexes formed were then added drop-wise onto cells and incubated under normal
growth conditions. Cells were harvested 48 hours after the transfections.
3.3.9 Prostate Sample Collection
Tumor samples were obtained, with the approval of the research ethics board, from
prostate cancer patients who had surgery at Sunnybrook Health Sciences Centre (Toronto,
Canada) between 1998 and 2006. As described by Gordanpour et al. (238), following radical
prostatectomy, the specimen was oriented using anatomic landmarks and the prostate and
seminal vesicles were painted right side green and left side blue. A random transverse midsection
of the prostate was taken perpendicular to the rectal surface, frozen in liquid nitrogen, and stored
at -80°C. Banked slices of specimens were photocopied, oriented (anterior, posterior, right and
left), quadrisected, and cut on the cryostat. Sections were stained with H&E and reviewed by a
pathologist to delineate tumor versus normal areas on the stained slides and a corresponding

78
image. The marked areas were used as a guide to indicate areas from which to extract the tumor
tissue from which RNA was later extracted. All research was conducted with the approval of
Sunnybrook Health Sciences Centre research ethics board.
3.3.10 Patient Follow-up
The medical records and follow-up information of patients were thoroughly reviewed
using standardised data entry forms by trained data abstractors and stored within a prostate
cancer-specific database. Biochemical recurrence was defined as a rise in blood levels of PSA in
prostate cancer patients on three consecutive measurements after radical prostatectomy. Data on
the following characteristics were available for each patient: age, months of follow-up, PSA
level, Gleason grade, biochemical recurrence status after 60 months of follow-up, and presence
of seminal vesicle invasion. The clinical demographics of the patients used in the study are
summarized in Table 1.
3.3.11 Recurrence Analysis
Patient data and mRNA data from DASL assays were published previously (239). Cox
survival regression analysis was performed using data from 68 patient specimens with recurrence
at <48 months or without recurrence for >48 months.
3.3.12 Statistical Analysis
Results were statistically analyzed using GraphPad Prism v4.0 software. Error bars
represent the standard error of the mean (SEM). Scatter plots were analyzed using student’s t
tests which were two-tailed and unpaired. Nonlinear regression (curve fit) and Pearson product-
moment correlation coefficient were used for correlation analysis of miR-182 and FOXO1.

79
3.4 RESULTS
3.4.1 MiR-182 Expression is Increased in Human Prostate Tumors, and Its Upregulation is Associated with Clinical Progression in Prostate Cancer.
MiR-182 expression was analyzed in 164 primary prostate cancer specimens that were
obtained after radical prostatectomy, and in 5 normal prostate tissue samples by qPCR. As shown
in Fig. 3.1A, miR-182 expression was progressively upregulated with increasing Gleason grades,
and was significantly higher in tumors with Gleason 8 or 9 as compared with normal prostate
tissue.
Based on the aforementioned results, we postulated that dysregulation of miR-182 is
associated with clinicopathological features and clinical progression of prostate cancer. The
clinical demographics of the patients in this study cohort are summarized in Table 3.1. We found
that the mean expression level of miR-182 was significantly higher (p = 0.03) in patients who
developed biochemical recurrence (0.92 ± 0.10) compared to those who remained recurrence-
free five years following prostatectomy (0.63 ± 0.07) (Fig. 3.1B). Moreover, we established that
miR-182 expression was higher in patients with seminal vesicle invasion at the time of radical
prostatectomy (Fig. 3.1C). We also analyzed the expression levels of miR-98 and miR-93 which
were thought to possibly be associated with prostate cancer. Levels of miR-98 and miR-93,
however, did not differ significantly between different Gleason grades (Fig. 3.1A), and in
relation to biochemical recurrence (data not shown).

80
Figure 3.1- MiR-182 is overexpressed in human prostate tumors, and its upregulation is associated with clinical progression. (A), miR-
182 expression was measured using qPCR in various prostate tumor samples representing different Gleason grades compared with normal
prostate tissue. Sample categories were distributed as follows: Normal (n=5), Gleason 5 or 6 (n=35), Gleason 7 (n=111), Gleason 8 or 9
(n=14). All samples were normalized to RNU6B expression. Expression levels of miR-93 and miR-98 were also assessed, but did not
show a significant difference. *, p < 0.05.
[Continued on next page]

81
Figure 3.1- miR-182 is overexpressed in human prostate tumors, and its upregulation is
associated with clinical progression. [Continued from previous page] B) expression levels of
miR-182 were assessed using qPCR in groups of recurrent (n=64) and non-recurrent (n=48)
prostate tumors. All samples were normalized to RNU6B expression. *, significant difference in
the median expression levels (black lines) between subgroups (p < 0.05). p-Values were
calculated with two tailed t-test.
[Continued on next page]

82
Figure 3.1- miR-182 is overexpressed in human prostate tumors, and its upregulation is
associated with clinical progression. [Continued from previous page] C) expression levels of
miR-182 were assessed using qPCR in groups patients with seminal vesicle invasion (n=19) and
no seminal vesicle invasion (n=145) at the time of prostatectomy. All samples were normalized
to RNU6B expression. *, significant difference in the median expression levels (black lines)
between subgroups (p < 0.05). p-Values were calculated with two tailed t-test.

83
3.4.2 MiR-182 Overexpression Enhances the Tumorigenic Properties of LNCaP Cells.
We next assessed the expression levels of miR-182 by semi-quantitative end point PCR analysis
in a series of prostate cell lines (Fig. 3.2A). The expression of miR-182 appears to be highest in
highly aggressive PC3 prostate carcinoma cells, compared to less invasive cell lines, such as DU-
145 and LNCaP. To assess the biological effects of miR-182, we used lentiviral pLemiR vectors
(Figure 3.2B) to increase miR-182 expression in prostate cancer cell line, LNCaP. LNCaP cells
stably transduced with miR-182 were found to robustly upregulate miR-182 levels to express
more than 4-fold higher levels of miR-182 than the empty vector control (Fig. 3.2C). To analyze
tumorigenic potential of these cells, we tested their anchorage-independent growth in soft agar as
a marker of tumorigenicity. As shown in figure 3.3A, miR-182 cells developed more colonies in
soft agar compared to vector control (p = 0.0007), indicating that miR-182 supports anchorage-
independent growth of LNCaP cells. Importantly, miR-182 cells exhibited sharp morphological
differences in culture (Fig. 3.3B). They appear to lose contact inhibition and form aggregates.
The appearance of these irregular spherical structures is dense and individual cells are difficult to
visualize. In contrast, vector control LNCaP cells exhibit a typical LNCaP morphology in bright
field and fluorescent microscopy: growing as loosely adherent cells that do not form a uniform
monolayer. Therefore, miR-182 overexpression enhances the tumorigenic properties of LNCaP
cells.

84
Table 3.1- Clinical demographics of prostate cancer cases used in this study
Cohort clinical characteristics for 164 prostate cancer patients are listed. Data includes: age,
years of follow-up, PSA, Gleason score, biochemical recurrence, and seminal vesicle invasion.
PSA: Prostate Specific Antigen.

85
Figure 3.2- MiR-182 expression in LNCaP cells. (A), Expression levels of miR-182 was analyzed by semi-quantitative end point PCR in
a series of Human Prostate Cell Lines. [Continued on next page]

86
Figure 3.2- MiR-182 expression in LNCaP cells. [Continued from previous page]
(B), Lentiviral pLemiR construct used to deliver miR-182 expression in LNCaP cell line. (C) Relative expression of miR-182 in stably
transduced LNCaP cells with empty vector control (Vector) or with miR-182-expressing vector (miR-182). The level of the mature miR-
182 was detected using qPCR, and the expression level was normalized to the RNU6B control.

87
Figure 3.3- MiR-182 overexpression enhances the tumourigenic properties of LNCaP cells. (A), Growth in soft agar of vector and miR-
182 LNCaP cells. Graph is representative of 3 independent experiments (n=3). (*p<0.05; ** p<0.01, ***p<0.001).
[Continued on next page]

88
Figure 3.3- MiR-182 overexpression enhances the tumourigenic properties of LNCaP cells. [Continued from previous page]
(B) Morphological transformation in culture of LNCaP cells with empty or miR-182-expressing vector. RFP: Red Fluorescent Protein.

89
3.4.3 miR-182 Enhances the Migratory and Invasive Abilities of LNCaP Cells.
Since aggressive and recurrent prostate tumors exhibit higher expression levels of miR-
182, we questioned whether miR-182 expression levels might be involved in increased
metastasis. We therefore investigated the role of miR-182 in cell migration of stably transduced
LNCaP cells. When these cells were analyzed in a trans-well migration assay, the results shown
in figure 3.4A indicate that miR-182 increased the migration of LNCaP cells by more than 10-
fold (p = 0.0012). MiR-182 cells were also more proficient than equivalent vector-transduced
cells at closing an artificial wound created over a confluent monolayer (Fig. 3.4B), further
reaffirming the conclusion that miR-182 enhances the migratory of LNCaP cells.
Invasive potential of miR-182 cells was analyzed using Matrigel-coated cell culture inserts. As
seen in figure 3.4C, miR-182 upregulation dramatically enhanced the invasive capacity of
normally poorly invasive LNCaP cells (p = 0.0006). Therefore, miR-182 upregulation enhances
migratory and invasive properties of LNCaP prostate cells. Together with the observed higher
miR-182 expression levels in aggressive and recurrent prostate tumors, these findings implicate
miR-182 in metastatic progression.

90
Figure 3.4- MiR-182 enhances the migratory and invasive abilities of LNCaP cells. (A), Trans-well migration assay on LNCaP cells
stably transduced with either empty vector (Vector) or miR-182 (miR-182) (n=3). (*p<0.05; ** p<0.01, ***p<0.001).
[Continued on next page]

91
Figure 3.4- MiR-182 enhances the migratory and invasive abilities of LNCaP cells. [Continued from previous page]
(B) Wound-healing assay on LNCaP cells transduced with empty or miR-182 vectors. Pictures were taken upon making the wound and at
day 1, day 3, and day 4 (n=2). [Continued on next page]

92
Figure 3.4- MiR-182 enhances the migratory and invasive abilities of LNCaP cells. [Continued from previous page]
(C), Trans-well invasion assay of LNCaP transduced with empty or miR-182 vectors (n=2). Cells were added to cell culture inserts
containing a filter with pores coated with matrigel. (*p<0.05; ** p<0.01, ***p<0.001).

93
3.4.4 MiR-182 Negatively Regulates FOXO1 in LNCaP Cells and Primary Prostate Tumors.
To identify targets that when inhibited by miR-182, could result in enhanced metastatic
properties that we saw in LNCaP cells, we searched the literature and prediction algorithms such
as miRBase and TargetScan, and found FOXO1, FOXO3, and MITF among the putative gene
targets of miR-182. FOXO1 protein is dramatically reduced in LNCaP-miR-182 cells as
compared to parental and vector alone cells, as shown by Western blot analysis (Fig. 3.5A). We
also found a slight diminishment in MITF protein levels in response to miR-182 overexpression
(Fig. 3.5B). We did not, however, detect any changes in FOXO3 protein levels.
To further confirm that miR-182 blocks FOXO1 expression, LNCaP-miR-182 cells were
transfected with antisense miR-182 or negative control inhibitors. These are single-stranded
RNA molecules which, after transfection, specifically inhibit miRNA function. A non-targeting
inhibitor was used as a negative control. As seen in Fig. 3.5A, inhibition of miR-182 resulted in
elevation of FOXO1 protein levels, while negative control miRNA inhibitors had no effect
(compare lanes 3, 4, and 5). Increased expression of FOXO1 after transfection of miR-182
inhibitor proves that miR-182 is regulating FOXO1 expression. LNCaP-miR-182 cells were also
transfected with FOXO1 siRNA, and miR-182-mediated downregulation of FOXO1 was
comparable to that achieved by FOXO1-specific siRNA (Fig 3.5C, compare lanes 4 and 5).
While over-expression of miR-182 resulted in 76% knockdown of FOXO1, concomitant over-
expression of miR-182 and FOXO1 siRNA increased the FOXO1 knockdown efficiency to 94%.
To compare paired expression levels of FOXO1 and miR-182, we analyzed the expression of
miR-182 and FOXO1 by qPCR in groups of prostate cancer samples, characterized by recurrent
or non-recurrent prostate cancer. As shown in figures 3.6, we find that there is a significant

94
inverse correlation between FOXO1 and miR-182 expression levels (Pearson’s Test; p=0.0054).
Together, these results identify FOXO-1 as a target of miR-182.

95
Figure 3.5- MiR-182 negatively regulates FOXO1 in LNCaP cells. (A), Western blot analysis of FOXO1 protein expression in LNCaP
cells, stably transduced LNCaP cells with empty vector (Vector), and with miR-182-expressing vector (miR-182). FOXO1 levels
inversely correlate with miR-182 expression. LNCaP-miR-182 cells were also transfected with antisense miR-182 or control inhibitors.
Inhibition of miR-182 increases FOXO1 levels. Densitometric quantification of FOXO1 relative to β-Tubulin from three independent
experiments is graphically depicted. [Continued on next page]

96
Figure 3.5- MiR-182 negatively regulates FOXO1 in LNCaP cells. [Continued from previous
page] (B), Western blot analysis of putative targets of miR-182, FOXO3 and MITF, in LNCaP,
Vector, and miR-182 cells. Representative immunoblotting from two independent experiments
are shown. [Continued on next page]

97
Figure 3.5- MiR-182 negatively regulates FOXO1 in LNCaP cells. [Continued from previous page] (C), LNCaP-miR-182 cells
transfected with FOXO1 siRNA shows that miR-182-mediated downregulation of FOXO1 was comparable to that achieved by FOXO1-
specific siRNA. Densitometric quantification of FOXO1 relative to β-Tubulin from three independent experiments is graphically depicted.

98
Figure 3.6- MiR-182 negatively regulates FOXO1 in primary prostate tumors. (A), FOXO1 levels inversely correlate with miR-182
expression in primary prostate tumor tissues, organized in aggressive and non-aggressive subgroups. Relative expression levels of miR-
182 and FOXO1 were measured by qPCR. [Continued on next page]
A

99
Figure 3.6- MiR-182 negatively regulates FOXO1 in primary prostate tumors. [Continued from
previous page] (B) Nonlinear regression (curve fit) and Pearson product-moment correlation
coefficient were used for correlation analysis.
B

100
3.4.5 FOXO1 Downregulation is Associated With Biochemical Recurrence in Prostate Cancer.
Considering that FOXO1 is a tumor suppressor that has been linked to increased metastasis
in prostate cancer cells, we sought to ascertain whether FOXO1 is associated with biochemical
recurrence and clinical progression in prostate cancer. FOXO1 genomic loss has been detected in
prostate cancer by The Cancer Genome Atlas (TCGA) consortium (p = 7.128E-8) (Fig. 3.7A).
Furthermore, analysis of the Oncomine database (240) determined that FOXO1 mRNA levels
were significantly decreased in cancerous versus normal prostate tissue, as reported by Lapointe
(p = 3.34E-10, Fig. 3.7B) (241), LaTulippe (p = 9.22E-6, Fig. 3.7C) (242), and Tomlins (p =
1.98E-4, Fig. 3.7D) (243), among others.
We subsequently used qPCR analysis on primary prostate cancer specimens to assess
whether FOXO1 expression levels are associated with clinicopathological features. We found
that the mean expression level of FOXO1 mRNA was significantly lower (p=0.0014) in tumors
from patients who developed biochemical recurrence (1.95 ± 0.13) compared to tumors from
patients who were recurrence-free five years after their radical prostatectomy (3.00 ± 0.34) (Fig.
3.7E). Furthermore, higher FOXO1 levels were associated with increased time of recurrence-free
survival using Cox Survival Analysis (Fig. 3.7F). These results confirm that FOXO1 acts as a
tumor suppressor in prostate cancer.

101
Figure 3.7- FOXO1 downregulation is associated with prostate cancer progression. [Refer
to next page] (A), box plot of FOXO1 DNA profile analysis in normal prostate tissue and
prostate adenocarcinoma from The Cancer Genome Atlas (TCGA) consortium. Oncomine
(Compendia Bioscience) was used for analysis and visualization. Analysis of FOXO1 mRNA
expressions in normal versus prostate cancer tissue as reported by Lapointe (B), LaTulippe (C),
and Tomlins (D), using the Oncomine database.

102

103
Figure 3.7. FOXO1 down-regulation is associated with prostate cancer progression. [Continued
from previous page] (E), mRNA expression levels of FOXO1 were assessed using qPCR in
groups of patients with biochemical recurrence or patients who were recurrence-free five years
after surgery. All samples were normalized to β-actin expression. *, significant difference in the
median expression levels (black lines) between subgroups (*p<0.05; ** p<0.01, ***p<0.001). P-
values were calculated with two tailed t-test. [Continued on next page]

104
Figure 3.7- FOXO1 down-regulation is associated with prostate cancer progression. [Continued
from previous page] (F), Cox survival regression analysis of FOXO1.

105
3.5 DISCUSSION
MiRNAs have been implicated in the post-transcriptional regulation of gene expression
in diverse cellular processes. Recent studies have evaluated miRNAs as potential biomarkers of
disease formation and progression in numerous cancer subtypes, in part based on the recognition
that some miRNAs are abnormally expressed in tumor versus normal tissue (244), and in many
cases, in metastatic and aggressive disease (245). One of the miRNAs that shows great potential
as a regulator of cancer metastasis is miR-182. Although several studies, including in melanoma
and breast cancer, suggest that miR-182 is an important oncogenic miRNA which is closely
linked to tumor progression and metastasis, however, so far there had been no studies examining
the role of miR-182 in prostate pathogenesis. Herein we show, using a large cohort of primary
prostate carcinoma, that higher expression of miR-182 promotes metastasis and clinical
recurrence in Prostate cancer.
In these studies, we have demonstrated several novel findings. First, miR-182 was shown
to be significantly up-regulated in primary tumors from patients who develop biochemical
recurrence post-surgery. This observation can be utilized in potential future use of miR-182 as a
prognostic biomarker of Prostate cancer. Next, we show that up-regulation of miR-182 causes
prostate cancer cells to be more proficient at anchorage-independent growth and losing contact
inhibition, thus displaying dramatic morphologic changes. These cells also show increased
migratory and invasive abilities. Finally, our studies demonstrate that miR-182 targets FOXO1 in
prostate cells, and down-regulates its mRNA and protein levels. This inverse correlation was
shown in the expression levels of mir-182 and FOXO1 in LNCaP prostate cells, as well as over a
hundred primary prostate tumors.

106
Malignant tumors are associated with altered tumour-host interactions leading to the
ability to invade and metastasize. To identify specific gene targets of miR-182 through which it
can promote invasive behaviour, we searched public target prediction algorithms such as
TargetScan and miRANDA for putative target genes whose down-regulation could mediate the
observed effects of miR-182 over-expression. These computational target prediction programs
point to 3’ UTR of FOXO1 mRNA as a direct target of miR-182, and this has already been
confirmed by several groups using luciferase assay (246-248).
The Forkhead Box O family of transcription factors with three main members (FOXO1,
FOXO3, and FOXO4) are involved in many important cellular processes such as cell cycle
regulation, cell differentiation, glucose metabolism, and other cellular functions (249). FOXO
proteins are mainly located in the nucleus and regulate expression of an extensive array of tumor
suppression genes. FOXO1 is itself a tumor suppressor that negatively regulates the highly
oncogenic phosphatidylinositol 3-kinase (P13K)/AKT signaling pathway (250). It is located on
the q14 band of chromosome 13 (13q14), where chromosomal deletions in cell lines, xenografts,
and clinical specimens of prostate cancer are common (251). Here, we report that FOXO1 is also
post-transcriptionally down-regulated via miRNA regulation. Our qPCR analysis of miR-182
and FOXO1 mRNA levels in primary samples of prostate carcinoma show a direct correlation
between overexpression of miR-182 and reduction of FOXO1 mRNA. This inverse correlation is
especially intriguing when considering the established role of FOXO1 as a tumor suppressor in
prostate cancer. Recent data suggests that FOXO1 binds to and suppresses another transcription
factor, Runx2, which plays a critical role in osteoblast maturation, bone formation, and prostate
cancer cell metastasis (237). Thus overexpression of miR-182, causing degradation of FOXO1
mRNA, may lead to enhanced Runx2 and increased migration and invasion in prostate cancer

107
cells. While our studies validate FOXO1 as a target of miR-182, future studies are needed to
identify and functionally evaluate downstream targets of FOXO1 that are related to prostate
cancer progression and metastasis, and are affected by miR-182 expression levels.
There are indeed promising breakthroughs in the field of miRNA research, such that we
can now envision miRNA-based therapeutics in the near future. In prostate cancer, for example,
systemic delivery of a synthetic mimic of miR-34a has been used to inhibit prostate cancer
metastasis and extend survival of tumor-bearing mice (252). However, more research is needed
in order to elucidate the interplay of various miRNAs involved in prostate cancer development
and progression, and the exact mechanism of their action through their target genes. Our study
presents a model of prostate cancer progression, in which increased miR-182 expression
enhances cancer aggressiveness by promoting oncogenic and invasive characteristics. According
to this model, upregulation of miR-182 promotes metastasis by affecting tumor suppressor
FOXO1. These findings demonstrate that miR-182 and its downstream effectors could prove to
be valuable in future exploitation of miR-182 as a prostate cancer biomarker and therapeutic
agent.

108
4 CHAPTER 4: GENERAL DISCUSSION AND FUTURE DIRECTIONS

109
4.1 SUMMARY AND IMPACT OF MAJOR FINDINGS
The preceding chapters in this thesis introduce miRNAs previously undescribed in prostate
cancer, confirm their aberrant expression in human prostate tumors, and outline possible modes
of action for these miRNAs. In Chapter 2 of this thesis, we described the association of miR-221
downregulation and recurrent prostate fusion gene, TMPRSS2-ERG, and found miR-221 to be
downregulated in recurrent prostate cancer. In Chapter 3, we implicated miR-182 as a novel
prostate cancer oncogene – we showed that miR-182 is overexpressed in the more aggressive
prostate tumors. Evidence is presented demonstrating that the ectopic expression of miR-182
corresponds to increased invasive and metastatic potential in prostate cells. Despite these
important findings, several questions pertaining to the role of miR-221 and miR-182 in the
biology of prostate cancer remain to be elucidated. These questions are discussed in more detail
in the Future Directions section.
One of the major themes of this thesis is the examination of miRNAs as potential
biomarkers for prostate cancer. Biomarkers can be used for purpose of screening, diagnosis,
prognosis, as an imaging marker, in predicting drug efficacy, as well as an indicator of cancer
recurrence. The current biomarker for prostate cancer, the PSA test, while limited in diagnosis
and screening, its rise post-surgery is a very effective indicator that cancer has returned. But we
are still in need of a biomarker that would predict which cancers are likely to recur in order to
improve management of the disease. Aberrant expression of miRNAs in cancer may potentially
be a useful predictor of clinical progression. We have analyzed miR-221 and miR-182 as
biomarkers capable of predictive prognosis and cancer recurrence. Still, many more questions
remain about the potential functional role of miR-221 and miR-182 as biomarkers of prostate

110
cancer progression, and also their potential utility as anticancer therapeutics. Although it remains
unclear exactly how ectopic expression of one miRNA may influence the expression of many
potential targets of that miRNA, and this certainly presents a challenge, it does not diminish the
potential of miRNAs as biomarkers.
One important feature of the studies presented in this thesis deals with use of a large
cohort of primary human prostate tumors to conduct translational research. For investigations
into the mechanisms of gene regulation by miRNAs, it is imperative that a sufficient sample size
is studied. In addition, in our identification and analysis of miRNA targets, we have utilized large
multi-institutional cancer genomics databases such as The Cancer Genome Atlas (TCGA) and
Oncomine database. These sources are particularly valuable for profiling large numbers of
tumours in order to analyze abnormalities in DNA sequence, genomic copy numbers,
chromosomal rearrangements, as well as gene expressions. Results from large-scale collaborative
efforts such as TCGA can offer a more comprehensive characterization of cancer genome than
ever before, and new candidate biomarkers can be discovered. These collaborative approaches
should continue to advance our ability to diagnose, stratify, and treat prostate cancer, ultimately
leading to decreased morbidity and mortality and enhanced quality of life for patients.

111
4.2 FUTURE DIRECTIONS
Many questions remain about the specific nature and potential functional role of miRNAs in
cancer. This includes identification of all the miRNAs that are involved in regulation of cancer
progression, the exact molecular mechanisms involved in various pathways, and feasibility of
employing miRNAs that are specific and sensitive enough to be used as biomarkers and/or
therapeutics. While this lack of information does present a challenge for more detailed
understanding of miRNAs, it does not detract from their potential importance in tumour biology.
In a direct sense miRNAs function as post-transcriptional regulators of the cell’s gene
expression. However, our work and others show that such regulation of transcription factors will
cause altered regulation of transcription as well. It is, therefore, of critical importance to identify
the magnitude of mRNAs targeted by a given miRNA in order to fully understand the influence
of the miRNA on various cellular processes. By validating the mRNA targets of a given miRNA,
the effects of deregulated miRNA expression can be put into context. Given the breadth and
diversity of targets of miRNAs, there are numerous significant consequences that aberrant
miRNA expression could have. Presently there are no confirmed targets of miR-221 and miR-
182 in the context of prostate cancer. MiR-221 has been implicated by Galardi et al to increase
proliferation in prostate cell lines by targeting a tumor suppressor p27kip1 (253). However,
Spahn et al has suggested that miR-221 is a tumor suppressor that negatively regulates the proto-
oncogene c-kit, though they did not find any correlation with p27Kip1 expression levels (159).
As for miR-182, FOXO3 and MITF have been validated as direct targets of miR-182 in other
cancer subtypes (247). In order to better understand the oncogenic effect(s) of aberrant miR-182
expression in prostate cancer, approaches aimed at identifying its mRNA targets in this tumour

112
are warranted. Although it has not yet been fully elucidated how many and which mRNAs are
regulated by miR-182, it is conceivable that miR-182 could influence several targets. Using
overexpression and/or knockdown strategies combined with array-based expression profiling,
bona fide targets of miR-182 in prostate cancer may be discovered, bringing a more
comprehensive insight into miR-182 function in prostate pathogenesis. We have shown that
FOXO1 is a target of miR-182 in prostate cancer. Nonetheless, as promising as FOXO1 appears
as a target of miR-182, it is likely that additional genes and pathways affected by miR-182 are
involved in prostate cancer progression. In this regard, future experiments will require a more
exhaustive look at other genes that may act as potential targets of miR-182.
In our characterization of the miRNAS that are aberrantly expressed in prostate cancer
using array-based profiling of miRNAs as described in Chapter 3, miR-182 and miR-221 were
among the six candidates that emerged. MiR-221 exhibited widespread downregulation and miR-
182 showed upregulation in recurrent prostate cancer compared to non-recurrent cancer – thus
making these two miRNAs a logical choice for follow-up studies. Other than miR-221 and miR-
182, our miRNA microarray identified four additional candidate miRNAs that were aberrantly
expressed in recurrent prostate cancer. MiR-103, miR-339, and miR-183 were also found to be
upregulated and miR-136 was downregulated when comparing recurrent versus non-recurrent
prostate cancer. To best of our knowledge, there are yet no studies that have analyzed the
expression of miR-103, miR-339, and miR-136 in prostate cancer. As previously mentioned,
miR-183, miR-182, and miR-96 are expressed as a cluster. Recently, Mihelich et al described
overexpression of the entire miR-183-96-182 cluster in prostate cancer and reported that their
upregulation suppressed five zinc transporters including hZIP1 (254). Prostate tumors are known
to have lower zinc levels, which are directly correlated with lower expression of the zinc

113
transporter hZIP1. This study suggests that miR-182 and miR-182 that are co-expressed are also
co-regulated. The significance of the differential expression observed for the additional four
miRNAs in our microarray study will require further investigation in future studies. Considering
the widespread role miRNAs have been shown to play in cancer, it is very likely that more
miRNA candidates beyond miR-221 and miR-182, such as those alluded to above, also
contribute to progression of prostate cancer.
With respect to potential future studies to decipher the functional role miR-182, a critical
first step is to confirm our results in other prostate cancer cell lines. The cell model used in this
study was human prostate cancer cell line, LNCaP. An important consideration should be to test
the effects of miR-182 ectopic expression in several other established cell lines such as DU145
and PC-3. Although our work has clearly implicated miR-182 as a novel prostate cancer
oncogene, mouse models would be great means of investigating the effects of overexpressing
miR-182 in an in vivo setting. Future approaches exploring miR-182 function by utilizing
genetically engineered mouse models, including spontaneous and implanted xenograft tumors, as
well as a comparison of both local and metastatic disease, can further elucidate the role of the
miRNA in the progression of prostate cancer. For example, subcutaneous and orthotopic
injection of cell lines overexpressing miR-182 and analyzing the rate of tumor growth and
metastasis to distant sites would be an ideal strategy to further characterize whether increased
miR-182 levels are associated with tumor burden.
In order to gain a full appreciation of the complexity of prostate cancer genome, bona
fide oncogenes and tumour suppressors need to be identified and validated, and the signaling
pathways that they regulate must be characterized. Recent technological advancement has made
examination of the genome more possible than ever before. In recent years, DNA sequencing

114
technology has transformed the field of cancer genomics (255-257). After three decades of
relying on Sanger-based sequencing technologies, next-generation sequencing or “deep”
sequencing now allows for the acquisition of tens of gigabases (Gb) of DNA sequence in one
experiment. Numerous studies have now utilized next-generation sequencing to analyze diverse
aspects of both normal and cancer genomes (258-260). Whole-transcriptome profiling using
next-generation sequencing has also allowed for quantification of transcript expression (mRNA
and miRNA) (261-263). By using this method, there does not need to be a prior knowledge of
the transcriptome under study, and global assessment of the known and unknown transcriptome
would eliminate any bias associated with characterization of transcriptome. As the cost
associated with the using the technique and analyzing the results becomes more affordable every
year, deep sequencing- profiling of prostate cancer genome and transcriptome will definitely
become more widespread. Hence, next-generation sequencing-based strategies for analyzing
cancer genetics can play a decisive role for a more complete characterization prostate cancer and
discovering new biomarkers.

115
5 APPENDIX 1: MICRORNA DETECTION IN PROSTATE TUMORS BY QUANTITATIVE REAL-Time PCR (QRT-PCR)
A version of this appendix has been published in Journal of Visualized Experiments as
Gordanpour A, Nam RK, Sugar L, Bacopulos S, Seth A. 2012 May 16;(63):e3874. doi:
10.3791/3874.

116
A1.1. ABSTRACT
Quantitative Real Time polymerase chain reaction (qPCR) is a rapid and sensitive method to
investigate the expression levels of various miRNA molecules in tumor samples. Using this
method expression of hundreds of different miRNA molecules can be amplified, quantified, and
analyzed from the same cDNA template. Methods used to investigate miRNA expression include
SYBR green I dye- based as well as Taqman-probe based qPCR. If miRNAs are to be effectively
used in the clinical setting, it is imperative that their detection in fresh and/or archived clinical
samples be accurate, reproducible, and specific. qPCR has been widely used for validating
expression of miRNAs in whole genome analyses such as microarray studies (264). The samples
used in this protocol were from patients who underwent radical prostatectomy for clinically
localized prostate cancer; however other tissues and cell lines can be substituted in. Prostate
specimens were snap-frozen in liquid nitrogen after resection. Clinical variables and follow-up
information for each patient were collected for subsequent analysis (238).
The main steps in qPCR analysis of tumors are: Total RNA extraction, cDNA synthesis,
and detection of qPCR products using miRNA-specific primers. Total RNA, which includes
mRNA, miRNA, and other small RNAs were extracted from specimens using TRIzol reagent.
Qiagen’s miScript System was used to synthesize cDNA and perform qPCR (Figure A1.1).
Endogenous miRNAs are not polyadenylated, therefore during the reverse transcription process,
a poly(A) polymerase polyadenylates the miRNA. The miRNA is used as a template to
synthesize cDNA using oligo-dT and Reverse Transcriptase. A universal tag sequence on the 5’
end of oligo-dT primers facilitates the amplification of cDNA in the PCR step. PCR product
amplification is detected by the level of fluorescence emitted by SYBR Green, a dye which

117
intercalates into double stranded DNA. Specific miRNA primers, along with a Universal Primer
that binds to the universal tag sequence will amplify specific miRNA sequences.
The miScript Primer Assays are available for over a thousand human-specific miRNAs,
and hundreds of murine-specific miRNAs. Relative quantification method was used here to
quantify the expression of miRNAs. To correct for variability amongst different samples,
expression levels of a target miRNA is normalized to the expression levels of a reference gene.
The choice of a gene on which to normalize the expression of targets is critical in relative
quantification method of analysis. Examples of reference genes typically used in this capacity are
the small RNAs RNU6B, RNU44, and RNU48 as they are considered to be stably expressed
across most samples. In this protocol, RNU6B is used as the reference gene.

118
Figure A1.1- Steps in qRT-PCR

119
A1.2. PROTOCOL
Prostate sample collection
Collect the prostate samples at the time of prostatectomy. The specimen is oriented using
anatomic landmarks. The prostate and seminal vesicles are painted as follows: right side
green, left side blue.
A random transverse midsection of the prostate is taken perpendicular to the rectal surface,
frozen in liquid nitrogen, and stored at -80°C (265).
Banked slices of specimens are photocopied, oriented (anterior, posterior, right and left),
quadrisected. Sections are cut using the Cryostat.
Sections are stained with H&E and reviewed by a pathologist to determine and delineate
tumor versus normal areas on the stained slides and a corresponding image. The marked
areas are used as a guide to indicate areas from which to extract the tumor tissue from which
RNA will be extracted in the subsequent steps.

120
Figure A1.2- Prostate Sample preparation

121
Isolating total RNA, including miRNA, from samples
Place frozen prostate samples on dry ice and referring to the delineated photocopy, cut out a
small portion of the prostate tumor (between 50 to 100 mg).
Homogenize the prostate tumor tissue in 1 mL of TRIzol Reagent. The quantities in the
following steps are based on use of 1 mL of TRIzol Reagent.
Note: Here we have used TRIzol Reagent for extracting RNA, however other kits that isolate
small RNA-containing total RNA can also be used.
Incubate the homogenized samples for 5 minutes at room temperature.
Add 0.2 mL of chloroform to the samples and shake vigorously for 15 seconds. Incubate
samples for 3 minutes at room temperature, then centrifuge at 12,000 x g for 15 minutes at
4°C.
Transfer the colorless upper aqueous phase to fresh tubes, and add 0.5 mL of isopropyl
alcohol. Incubate samples for 10 minutes at room temperature, then centrifuge at 12,000 x g
for 10 minutes at 4°C.
Carefully aspirate the supernatant without disturbing the pellet containing the RNA. Wash
the RNA pellet with 1 mL of 75% ethanol. Vortex the sample and re-sediment by
centrifugation for 5 minutes at 7,500 x g at 4°C.
Carefully aspirate the supernatant and dry the RNA pellet for 5-10 minutes, making sure the
RNA pellet is not completely dry. Re-dissolve in Nuclease-free water appropriate to pellet
size. Measure the concentration of RNA using the NanoDrop 1000 spectrophotometer
(measure absorbance at 260 nm and 280 nm).
Check the quality and integrity of the RNA samples using Agilent Bioanalyzer.

122
Reverse transcription of RNA
Reverse transcription of RNA was performed using miScript Reverse Transcription Kit
according to manufacturer’s instructions (Qiagen). This kit includes a reverse transcriptase
and a poly(A) polymerase. The miScript RT Buffer includes Mg2+
, dNTPs, oligo-dT primers,
and random primers.
Use between 10 pg and 1 μg of RNA to synthesize cDNA. If using more than 1 μg of RNA,
scale up the reaction linearly to the appropriate volume.
Prepare a master mix that contains 5X miScript RT Buffer (4 μl), miScript Reverse
Transcription Mix (1 μl), and RNase-free water to bring reactions to final volume of 20 μl.
Also include template RNA (up to 1 μg) in the master mix.
Incubate the samples for 60 minutes at 37°C followed immediately by incubation for 5
minutes at 95°C. This step can be performed in a PCR machine, heating block, or water bath.
Thermocyclers are the most suitable and accurate method. Store the cDNA on ice for short
term, and -20 °C for long term storage.
Generating a standard curve
Prior to experiment with target miRNAs, a standard curve is generated by using cDNAs of
known concentrations against their crossing points (CP) (Fig A1.3).
Prepare a series of dilutions of 2-fold, 10-fold, 50-fold, 250-fold, and 1250-fold the original
cDNA of a sample that is known to have a substantial expression of your gene of interest.
Run the PCR as specified in Section 5 “Real-time PCR for detection of miRNA”, with the
modification that cDNA is in serial dilutions not a static 40x dilution.
Perform analysis using RelQuant software (Roche) to generate your standard curve.

123
Note: a new standard curve must be generated for each gene of interest.

124
Figure A1.3- Generating a standard curve

125
Real-time PCR for detection of miRNA
Real time PCR for miRNAs was performed using miScript SYBR Green PCR Kit and
miScript Primer Assay according to manufacturer’s instructions (Qiagen). Prepare a master
mix containing 2x QuantiTect SYBR Green PCR Master Mix, 10x miScript Universal
Primer, 10x miScript Primer Assay, and RNase-free water. Prepare a master mix for a 20 μl
volume reaction.
The Primer Assay is specific to the miRNA of interest. To reconstitute 10x miScript Primer
Assay, centrifuge the vial briefly, and add 550 μl TE buffer, pH 8.0. Vortex the vial briefly to
mix, aliquot primers to smaller volumes, and store at -20°C. Two primers are required:
primers for the target gene and the reference gene. RNU6B is used as the reference gene.
Dilute the cDNA 40x and store extra aliquots at –20°C.
cDNA serves as the template for the PCR. Use 2 μl of 40x diluted cDNA and dispense to the
20 μl light cycler capillaries (Roche).
Add 18 μl of the master mix to each capillary, and centrifuge using a capillary adapter.
Place the capillaries in a capillary based Real-Time cycler, such as LightCycler ® 3.5 Real-
Time PCR System with a 32-capillary carousel format.
Run the PCR cycling program as follows:
To activate HotStarTaq Polymerase that is in the 2x QuantiTect SYBR Green PCR Master Mix,
pre-incubate at 95°C for 15 minutes.
Followed by 50 cycles of:
Denaturation, 15 s, 94°C;
Annealing, 30 s, 55°C;
Extension, 30 s, 70°C.

126
Select a sample to be the calibrator, and set its normalized target amount to 1. Compare the
relative expression of the miRNA in all the other samples to the calibrator.
Note: Within a study, the same calibrating sample should be used to maintain consistency of
results.
Analyzing data
Amplification curves for the PCR reactions are depicted graphically and numerically by
Molecular Biochemicals LightCycler Software version 3.5 (Roche) (Fig A1.4). Quantify
reactions in the “Quantification” tab, and export the data to a text file.
Import the data to the RelQuant analysis software (Roche) to generate quantification results.
Import separate files for target gene, reference gene, and standard curve data.
Specify the position of calibrator for both target and reference gene. Also specify positions of
the samples. Data is expressed as the target to reference ratio of different samples divided by
the target to reference ratio of the calibrator. The standard curve previously generated for a
particular miRNA and housekeeping gene is used as a reference standard for extrapolating
quantitative data for miRNA targets of unknown concentrations.
Three replicates of samples are analyzed as a group and mean concentrations and standard
deviations of the triplicate is calculated. If one of the triplicates is inconsistent with the rest
of the set, it will be excluded by the program (Fig A1.5).

127
Figure A1.4- Amplification curve for PCR reaction

128
Figure A1.5- Data analysis for PCR reaction

129
A1.3. DISCUSSION
Aberrant expressions of some miRNAs have been consistently found in prostate tumors
when compared to normal tissue (235), and some of these miRNAs have been named as potential
novel therapeutic agents against prostate cancer (252). Hence the aberrant expression levels of
miRNAs can be useful diagnostic and/or prognostic biomarkers. The Real-Time qPCR
methodology presented here provides an assay for accurate quantification of miRNA levels in
prostate tumor tissues. The miScript PCR system used can detect single nucleotide differences
between mature miRNAs. The miScript miRNA qPCR Assays, however, are not intended for
detection of stem loop precursor miRNAs, for which different miScript Precursor Assays are
available.
The reliability of this technique depends on the quality of the input RNA, therefore
concentration, integrity, and purity of RNA should be tested prior to Real-Time PCR. Moreover,
ribonucleases are very stable and readily degrade RNA, thus extra caution should be taken in
handling of the RNA. All reactions should be set up on ice to minimize RNA degradation. RNase
inhibitors can also be added to the reaction prior to reverse transcription. Gloves should be
frequently changed, while sterile and disposable plasticware must be used throughout the
procedure. If there is no PCR product or the amplification curve is detected late in Real-Time
PCR, try increasing the number of PCR cycles, and make sure that the cycling program includes
activation of HotStarTaq DNA polymerase for 15 minutes at 95°C. Low amplification might also
be due to inadequate starting cDNA template, therefore try to increase the amount of cDNA.
Late amplification can also represent a false positive.

130
REFERENCES
(1) Peehl DM. Primary cell cultures as models of prostate cancer development. Endocr Relat
Cancer 2005 Mar;12(1):19-47.
(2) Jemal A, Siegel R, Ward E, Murray T, Xu J, Smigal C, et al. Cancer statistics, 2006. CA
Cancer J Clin 2006 Mar;56(2):106-30.
(3) Sakr WA, Ward C, Grignon DJ, Haas GP. Epidemiology and molecular biology of early
prostatic neoplasia. Mol Urol 2000;4(3):109-13.
(4) Newcomer LM, Stanford JL, Blumenstein BA, Brawer MK. Temporal trends in rates of
prostate cancer: declining incidence of advanced stage disease, 1974 to 1994. J Urol 1997
Oct;158(4):1427-30.
(5) Bratt O. Hereditary prostate cancer: clinical aspects. J Urol 2002 Sep;168(3):906-13.
(6) Dennis LK, Dawson DV. Meta-analysis of measures of sexual activity and prostate
cancer. Epidemiology 2002 Jan;13(1):72-9.
(7) Honda GD, Bernstein L, Ross RK, Greenland S, Gerkins V, Henderson BE. Vasectomy,
cigarette smoking, and age at first sexual intercourse as risk factors for prostate cancer in
middle-aged men. Br J Cancer 1988 Mar;57(3):326-31.
(8) Mannisto S, Pietinen P, Virtanen MJ, Salminen I, Albanes D, Giovannucci E, et al. Fatty
acids and risk of prostate cancer in a nested case-control study in male smokers. Cancer
Epidemiol Biomarkers Prev 2003 Dec;12(12):1422-8.
(9) Rebbeck TR, Jaffe JM, Walker AH, Wein AJ, Malkowicz SB. Modification of clinical
presentation of prostate tumors by a novel genetic variant in CYP3A4. J Natl Cancer Inst
1998 Aug 19;90(16):1225-9.
(10) Peehl DM, Feldman D. The role of vitamin D and retinoids in controlling prostate cancer
progression. Endocr Relat Cancer 2003 Jun;10(2):131-40.
(11) Stamey TA, Yang N, Hay AR, McNeal JE, Freiha FS, Redwine E. Prostate-specific
antigen as a serum marker for adenocarcinoma of the prostate. N Engl J Med 1987 Oct
8;317(15):909-16.
(12) Oesterling JE, Jacobsen SJ, Cooner WH. The use of age-specific reference ranges for
serum prostate specific antigen in men 60 years old or older. J Urol 1995
Apr;153(4):1160-3.

131
(13) Catalona WJ, Smith DS, Wolfert RL, Wang TJ, Rittenhouse HG, Ratliff TL, et al.
Evaluation of percentage of free serum prostate-specific antigen to improve specificity of
prostate cancer screening. JAMA 1995 Oct 18;274(15):1214-20.
(14) Catalona WJ, Partin AW, Slawin KM, Brawer MK, Flanigan RC, Patel A, et al. Use of
the percentage of free prostate-specific antigen to enhance differentiation of prostate
cancer from benign prostatic disease: a prospective multicenter clinical trial. JAMA 1998
May 20;279(19):1542-7.
(15) Jung K, Meyer A, Lein M, Rudolph B, Schnorr D, Loening SA. Ratio of free-to-total
prostate specific antigen in serum cannot distinguish patients with prostate cancer from
those with chronic inflammation of the prostate. J Urol 1998 May;159(5):1595-8.
(16) Welch HG, Fisher ES, Gottlieb DJ, Barry MJ. Detection of prostate cancer via biopsy in
the Medicare-SEER population during the PSA era. J Natl Cancer Inst 2007 Sep
19;99(18):1395-400.
(17) Thompson IM, Pauler DK, Goodman PJ, Tangen CM, Lucia MS, Parnes HL, et al.
Prevalence of prostate cancer among men with a prostate-specific antigen level < or =4.0
ng per milliliter. N Engl J Med 2004 May 27;350(22):2239-46.
(18) Egevad L, Mazzucchelli R, Montironi R. Implications of the International Society of
Urological Pathology modified Gleason grading system. Arch Pathol Lab Med 2012
Apr;136(4):426-34.
(19) Gleason DF. Classification of prostatic carcinomas. Cancer Chemother Rep 1966
Mar;50(3):125-8.
(20) Humphrey PA. Gleason grading and prognostic factors in carcinoma of the prostate. Mod
Pathol 2004 Mar;17(3):292-306.
(21) Middleton RG, Thompson IM, Austenfeld MS, Cooner WH, Correa RJ, Gibbons RP, et
al. Prostate Cancer Clinical Guidelines Panel Summary report on the management of
clinically localized prostate cancer. The American Urological Association. J Urol 1995
Dec;154(6):2144-8.
(22) Epstein JI, Pizov G, Walsh PC. Correlation of pathologic findings with progression after
radical retropubic prostatectomy. Cancer 1993 Jun 1;71(11):3582-93.
(23) Saitoh H, Hida M, Shimbo T, Nakamura K, Yamagata J, Satoh T. Metastatic patterns of
prostatic cancer. Correlation between sites and number of organs involved. Cancer 1984
Dec 15;54(12):3078-84.
(24) Chinnaiyan AM, Palanisamy N. Chromosomal aberrations in solid tumors. Prog Mol Biol
Transl Sci 2010;95:55-94.

132
(25) Tomlins SA, Rhodes DR, Perner S, Dhanasekaran SM, Mehra R, Sun XW, et al.
Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer.
Science 2005 Oct 28;310(5748):644-8.
(26) Clark JP, Cooper CS. ETS gene fusions in prostate cancer. Nat Rev Urol 2009
Aug;6(8):429-39.
(27) Kumar-Sinha C, Tomlins SA, Chinnaiyan AM. Recurrent gene fusions in prostate cancer.
Nat Rev Cancer 2008 Jul;8(7):497-511.
(28) Clark J, Merson S, Jhavar S, Flohr P, Edwards S, Foster CS, et al. Diversity of
TMPRSS2-ERG fusion transcripts in the human prostate. Oncogene 2007 Apr
19;26(18):2667-73.
(29) Tomlins SA, Laxman B, Varambally S, Cao X, Yu J, Helgeson BE, et al. Role of the
TMPRSS2-ERG gene fusion in prostate cancer. Neoplasia 2008 Feb;10(2):177-88.
(30) Hofer MD, Kuefer R, Maier C, Herkommer K, Perner S, Demichelis F, et al. Genome-
wide linkage analysis of TMPRSS2-ERG fusion in familial prostate cancer. Cancer Res
2009 Jan 15;69(2):640-6.
(31) Esgueva R, Perner S, LaFargue J, Scheble V, Stephan C, Lein M, et al. Prevalence of
TMPRSS2-ERG and SLC45A3-ERG gene fusions in a large prostatectomy cohort. Mod
Pathol 2010 Apr;23(4):539-46.
(32) Maher CA, Palanisamy N, Brenner JC, Cao X, Kalyana-Sundaram S, Luo S, et al.
Chimeric transcript discovery by paired-end transcriptome sequencing. Proc Natl Acad
Sci U S A 2009 Jul 28;106(30):12353-8.
(33) Pflueger D, Rickman DS, Sboner A, Perner S, LaFargue CJ, Svensson MA, et al. N-myc
downstream regulated gene 1 (NDRG1) is fused to ERG in prostate cancer. Neoplasia
2009 Aug;11(8):804-11.
(34) Tomlins SA, Laxman B, Dhanasekaran SM, Helgeson BE, Cao X, Morris DS, et al.
Distinct classes of chromosomal rearrangements create oncogenic ETS gene fusions in
prostate cancer. Nature 2007 Aug 2;448(7153):595-9.
(35) Hermans KG, van der Korput HA, van MR, van de Wijngaart DJ, Ziel-van der Made A,
Dits NF, et al. Truncated ETV1, fused to novel tissue-specific genes, and full-length
ETV1 in prostate cancer. Cancer Res 2008 Sep 15;68(18):7541-9.
(36) Han B, Mehra R, Dhanasekaran SM, Yu J, Menon A, Lonigro RJ, et al. A fluorescence in
situ hybridization screen for E26 transformation-specific aberrations: identification of
DDX5-ETV4 fusion protein in prostate cancer. Cancer Res 2008 Sep 15;68(18):7629-37.
(37) Attard G, Clark J, Ambroisine L, Mills IG, Fisher G, Flohr P, et al. Heterogeneity and
clinical significance of ETV1 translocations in human prostate cancer. Br J Cancer 2008
Jul 22;99(2):314-20.

133
(38) Tomlins SA, Mehra R, Rhodes DR, Smith LR, Roulston D, Helgeson BE, et al.
TMPRSS2:ETV4 gene fusions define a third molecular subtype of prostate cancer.
Cancer Res 2006 Apr 1;66(7):3396-400.
(39) Hermans KG, Bressers AA, van der Korput HA, Dits NF, Jenster G, Trapman J. Two
unique novel prostate-specific and androgen-regulated fusion partners of ETV4 in
prostate cancer. Cancer Res 2008 May 1;68(9):3094-8.
(40) Helgeson BE, Tomlins SA, Shah N, Laxman B, Cao Q, Prensner JR, et al.
Characterization of TMPRSS2:ETV5 and SLC45A3:ETV5 gene fusions in prostate
cancer. Cancer Res 2008 Jan 1;68(1):73-80.
(41) Palanisamy N, Ateeq B, Kalyana-Sundaram S, Pflueger D, Ramnarayanan K, Shankar S,
et al. Rearrangements of the RAF kinase pathway in prostate cancer, gastric cancer and
melanoma. Nat Med 2010 Jul;16(7):793-8.
(42) Rickman DS, Pflueger D, Moss B, VanDoren VE, Chen CX, de la Taille A, et al.
SLC45A3-ELK4 is a novel and frequent erythroblast transformation-specific fusion
transcript in prostate cancer. Cancer Res 2009 Apr 1;69(7):2734-8.
(43) Laxman B, Morris DS, Yu J, Siddiqui J, Cao J, Mehra R, et al. A first-generation
multiplex biomarker analysis of urine for the early detection of prostate cancer. Cancer
Res 2008 Feb 1;68(3):645-9.
(44) Hessels D, Smit FP, Verhaegh GW, Witjes JA, Cornel EB, Schalken JA. Detection of
TMPRSS2-ERG fusion transcripts and prostate cancer antigen 3 in urinary sediments
may improve diagnosis of prostate cancer. Clin Cancer Res 2007 Sep 1;13(17):5103-8.
(45) Park K, Tomlins SA, Mudaliar KM, Chiu YL, Esgueva R, Mehra R, et al. Antibody-
based detection of ERG rearrangement-positive prostate cancer. Neoplasia 2010
Jul;12(7):590-8.
(46) Demichelis F, Fall K, Perner S, Andren O, Schmidt F, Setlur SR, et al. TMPRSS2:ERG
gene fusion associated with lethal prostate cancer in a watchful waiting cohort. Oncogene
2007 Jul 5;26(31):4596-9.
(47) Mehra R, Tomlins SA, Yu J, Cao X, Wang L, Menon A, et al. Characterization of
TMPRSS2-ETS gene aberrations in androgen-independent metastatic prostate cancer.
Cancer Res 2008 May 15;68(10):3584-90.
(48) Wang J, Cai Y, Ren C, Ittmann M. Expression of variant TMPRSS2/ERG fusion
messenger RNAs is associated with aggressive prostate cancer. Cancer Res 2006 Sep
1;66(17):8347-51.
(49) Nam RK, Sugar L, Yang W, Srivastava S, Klotz LH, Yang LY, et al. Expression of the
TMPRSS2:ERG fusion gene predicts cancer recurrence after surgery for localised
prostate cancer. Br J Cancer 2007 Dec 17;97(12):1690-5.

134
(50) Rajput AB, Miller MA, De LA, Boyd N, Leung S, Hurtado-Coll A, et al. Frequency of
the TMPRSS2:ERG gene fusion is increased in moderate to poorly differentiated prostate
cancers. J Clin Pathol 2007 Nov;60(11):1238-43.
(51) Attard G, Clark J, Ambroisine L, Fisher G, Kovacs G, Flohr P, et al. Duplication of the
fusion of TMPRSS2 to ERG sequences identifies fatal human prostate cancer. Oncogene
2008 Jan 10;27(3):253-63.
(52) Yoshimoto M, Joshua AM, Cunha IW, Coudry RA, Fonseca FP, Ludkovski O, et al.
Absence of TMPRSS2:ERG fusions and PTEN losses in prostate cancer is associated
with a favorable outcome. Mod Pathol 2008 Dec;21(12):1451-60.
(53) Nam RK, Sugar L, Wang Z, Yang W, Kitching R, Klotz LH, et al. Expression of
TMPRSS2:ERG gene fusion in prostate cancer cells is an important prognostic factor for
cancer progression. Cancer Biol Ther 2007 Jan;6(1):40-5.
(54) Lapointe J, Li C, Giacomini CP, Salari K, Huang S, Wang P, et al. Genomic profiling
reveals alternative genetic pathways of prostate tumorigenesis. Cancer Res 2007 Sep
15;67(18):8504-10.
(55) Tu JJ, Rohan S, Kao J, Kitabayashi N, Mathew S, Chen YT. Gene fusions between
TMPRSS2 and ETS family genes in prostate cancer: frequency and transcript variant
analysis by RT-PCR and FISH on paraffin-embedded tissues. Mod Pathol 2007
Sep;20(9):921-8.
(56) Yoshimoto M, Joshua AM, Chilton-Macneill S, Bayani J, Selvarajah S, Evans AJ, et al.
Three-color FISH analysis of TMPRSS2/ERG fusions in prostate cancer indicates that
genomic microdeletion of chromosome 21 is associated with rearrangement. Neoplasia
2006 Jun;8(6):465-9.
(57) Furusato B, Gao CL, Ravindranath L, Chen Y, Cullen J, McLeod DG, et al. Mapping of
TMPRSS2-ERG fusions in the context of multi-focal prostate cancer. Mod Pathol 2008
Feb;21(2):67-75.
(58) Petrovics G, Liu A, Shaheduzzaman S, Furusato B, Sun C, Chen Y, et al. Frequent
overexpression of ETS-related gene-1 (ERG1) in prostate cancer transcriptome.
Oncogene 2005 May 26;24(23):3847-52.
(59) Winnes M, Lissbrant E, Damber JE, Stenman G. Molecular genetic analyses of the
TMPRSS2-ERG and TMPRSS2-ETV1 gene fusions in 50 cases of prostate cancer. Oncol
Rep 2007 May;17(5):1033-6.
(60) Saramaki OR, Harjula AE, Martikainen PM, Vessella RL, Tammela TL, Visakorpi T.
TMPRSS2:ERG fusion identifies a subgroup of prostate cancers with a favorable
prognosis. Clin Cancer Res 2008 Jun 1;14(11):3395-400.

135
(61) Druker BJ, Talpaz M, Resta DJ, Peng B, Buchdunger E, Ford JM, et al. Efficacy and
safety of a specific inhibitor of the BCR-ABL tyrosine kinase in chronic myeloid
leukemia. N Engl J Med 2001 Apr 5;344(14):1031-7.
(62) Licht JD. Acute promyelocytic leukemia--weapons of mass differentiation. N Engl J Med
2009 Feb 26;360(9):928-30.
(63) Gordanpour A, Stanimirovic A, Nam RK, Moreno CS, Sherman C, Sugar L, et al. miR-
221 Is Down-regulated in TMPRSS2:ERG Fusion-positive Prostate Cancer. Anticancer
Res 2011 Feb;31(2):403-10.
(64) Venter JC, Adams MD, Myers EW, Li PW, Mural RJ, Sutton GG, et al. The sequence of
the human genome. Science 2001 Feb 16;291(5507):1304-51.
(65) Holley RW. Structure of an alanine transfer ribonucleic acid. JAMA 1965 Nov
22;194(8):868-71.
(66) HOLLEY RW, APGAR J, EVERETT GA, MADISON JT, MARQUISEE M, MERRILL
SH, et al. STRUCTURE OF A RIBONUCLEIC ACID. Science 1965 Mar
19;147(3664):1462-5.
(67) Eddy SR. Non-coding RNA genes and the modern RNA world. Nat Rev Genet 2001
Dec;2(12):919-29.
(68) Nagai K, Oubridge C, Kuglstatter A, Menichelli E, Isel C, Jovine L. Structure, function
and evolution of the signal recognition particle. EMBO J 2003 Jul 15;22(14):3479-85.
(69) Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004 Jan
23;116(2):281-97.
(70) Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 2004 Jan
23;116(2):281-97.
(71) Olsen PH, Ambros V. The lin-4 regulatory RNA controls developmental timing in
Caenorhabditis elegans by blocking LIN-14 protein synthesis after the initiation of
translation. Dev Biol 1999 Dec 15;216(2):671-80.
(72) Eulalio A, Rehwinkel J, Stricker M, Huntzinger E, Yang SF, Doerks T, et al. Target-
specific requirements for enhancers of decapping in miRNA-mediated gene silencing.
Genes Dev 2007 Oct 15;21(20):2558-70.
(73) Bagga S, Bracht J, Hunter S, Massirer K, Holtz J, Eachus R, et al. Regulation by let-7 and
lin-4 miRNAs results in target mRNA degradation. Cell 2005 Aug 26;122(4):553-63.
(74) Lytle JR, Yario TA, Steitz JA. Target mRNAs are repressed as efficiently by microRNA-
binding sites in the 5' UTR as in the 3' UTR. Proc Natl Acad Sci U S A 2007 Jun
5;104(23):9667-72.

136
(75) Duursma AM, Kedde M, Schrier M, le SC, Agami R. miR-148 targets human DNMT3b
protein coding region. RNA 2008 May;14(5):872-7.
(76) Khraiwesh B, Arif MA, Seumel GI, Ossowski S, Weigel D, Reski R, et al.
Transcriptional control of gene expression by microRNAs. Cell 2010 Jan 8;140(1):111-
22.
(77) Gonzalez S, Pisano DG, Serrano M. Mechanistic principles of chromatin remodeling
guided by siRNAs and miRNAs. Cell Cycle 2008 Aug 15;7(16):2601-8.
(78) Kim DH, Saetrom P, Snove O, Jr., Rossi JJ. MicroRNA-directed transcriptional gene
silencing in mammalian cells. Proc Natl Acad Sci U S A 2008 Oct 21;105(42):16230-5.
(79) Orom UA, Nielsen FC, Lund AH. MicroRNA-10a binds the 5'UTR of ribosomal protein
mRNAs and enhances their translation. Mol Cell 2008 May 23;30(4):460-71.
(80) Calin GA, Dumitru CD, Shimizu M, Bichi R, Zupo S, Noch E, et al. Frequent deletions
and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic
lymphocytic leukemia. Proc Natl Acad Sci U S A 2002 Nov 26;99(24):15524-9.
(81) Kozomara A, Griffiths-Jones S. miRBase: integrating microRNA annotation and deep-
sequencing data. Nucleic Acids Res 2011 Jan;39(Database issue):D152-D157.
(82) Friedman RC, Farh KK, Burge CB, Bartel DP. Most mammalian mRNAs are conserved
targets of microRNAs. Genome Res 2009 Jan;19(1):92-105.
(83) Krek A, Grun D, Poy MN, Wolf R, Rosenberg L, Epstein EJ, et al. Combinatorial
microRNA target predictions. Nat Genet 2005 May;37(5):495-500.
(84) Betel D, Wilson M, Gabow A, Marks DS, Sander C. The microRNA.org resource: targets
and expression. Nucleic Acids Res 2008 Jan;36(Database issue):D149-D153.
(85) Lewis BP, Shih IH, Jones-Rhoades MW, Bartel DP, Burge CB. Prediction of mammalian
microRNA targets. Cell 2003 Dec 26;115(7):787-98.
(86) Reinhart BJ, Weinstein EG, Rhoades MW, Bartel B, Bartel DP. MicroRNAs in plants.
Genes Dev 2002 Jul 1;16(13):1616-26.
(87) Chen CZ, Li L, Lodish HF, Bartel DP. MicroRNAs modulate hematopoietic lineage
differentiation. Science 2004 Jan 2;303(5654):83-6.
(88) Xu P, Vernooy SY, Guo M, Hay BA. The Drosophila microRNA Mir-14 suppresses cell
death and is required for normal fat metabolism. Curr Biol 2003 Apr 29;13(9):790-5.
(89) Bueno MJ, Perez dC, I, Malumbres M. Control of cell proliferation pathways by
microRNAs. Cell Cycle 2008 Oct;7(20):3143-8.

137
(90) Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes
small RNAs with antisense complementarity to lin-14. Cell 1993 Dec 3;75(5):843-54.
(91) Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC, Rougvie AE, et al. The
21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans.
Nature 2000 Feb 24;403(6772):901-6.
(92) Winter J, Jung S, Keller S, Gregory RI, Diederichs S. Many roads to maturity:
microRNA biogenesis pathways and their regulation. Nat Cell Biol 2009 Mar;11(3):228-
34.
(93) Lee Y, Kim M, Han J, Yeom KH, Lee S, Baek SH, et al. MicroRNA genes are
transcribed by RNA polymerase II. EMBO J 2004 Oct 13;23(20):4051-60.
(94) Borchert GM, Lanier W, Davidson BL. RNA polymerase III transcribes human
microRNAs. Nat Struct Mol Biol 2006 Dec;13(12):1097-101.
(95) Cai X, Hagedorn CH, Cullen BR. Human microRNAs are processed from capped,
polyadenylated transcripts that can also function as mRNAs. RNA 2004
Dec;10(12):1957-66.
(96) Saini HK, Griffiths-Jones S, Enright AJ. Genomic analysis of human microRNA
transcripts. Proc Natl Acad Sci U S A 2007 Nov 6;104(45):17719-24.
(97) Han J, Lee Y, Yeom KH, Nam JW, Heo I, Rhee JK, et al. Molecular basis for the
recognition of primary microRNAs by the Drosha-DGCR8 complex. Cell 2006 Jun
2;125(5):887-901.
(98) Lee Y, Ahn C, Han J, Choi H, Kim J, Yim J, et al. The nuclear RNase III Drosha initiates
microRNA processing. Nature 2003 Sep 25;425(6956):415-9.
(99) Han J, Lee Y, Yeom KH, Nam JW, Heo I, Rhee JK, et al. Molecular basis for the
recognition of primary microRNAs by the Drosha-DGCR8 complex. Cell 2006 Jun
2;125(5):887-901.
(100) Yi R, Qin Y, Macara IG, Cullen BR. Exportin-5 mediates the nuclear export of pre-
microRNAs and short hairpin RNAs. Genes Dev 2003 Dec 15;17(24):3011-6.
(101) Zeng Y, Cullen BR. Structural requirements for pre-microRNA binding and nuclear
export by Exportin 5. Nucleic Acids Res 2004;32(16):4776-85.
(102) Winter J, Jung S, Keller S, Gregory RI, Diederichs S. Many roads to maturity:
microRNA biogenesis pathways and their regulation. Nat Cell Biol 2009 Mar;11(3):228-
34.
(103) Khvorova A, Reynolds A, Jayasena SD. Functional siRNAs and miRNAs exhibit strand
bias. Cell 2003 Oct 17;115(2):209-16.

138
(104) Pillai RS, Artus CG, Filipowicz W. Tethering of human Ago proteins to mRNA mimics
the miRNA-mediated repression of protein synthesis. RNA 2004 Oct;10(10):1518-25.
(105) Sasaki T, Shiohama A, Minoshima S, Shimizu N. Identification of eight members of the
Argonaute family in the human genome small star, filled. Genomics 2003 Sep;82(3):323-
30.
(106) Rivas FV, Tolia NH, Song JJ, Aragon JP, Liu J, Hannon GJ, et al. Purified Argonaute2
and an siRNA form recombinant human RISC. Nat Struct Mol Biol 2005 Apr;12(4):340-
9.
(107) Ameres SL, Martinez J, Schroeder R. Molecular basis for target RNA recognition and
cleavage by human RISC. Cell 2007 Jul 13;130(1):101-12.
(108) Orban TI, Izaurralde E. Decay of mRNAs targeted by RISC requires XRN1, the Ski
complex, and the exosome. RNA 2005 Apr;11(4):459-69.
(109) Parker R, Sheth U. P bodies and the control of mRNA translation and degradation. Mol
Cell 2007 Mar 9;25(5):635-46.
(110) Behm-Ansmant I, Rehwinkel J, Doerks T, Stark A, Bork P, Izaurralde E. mRNA
degradation by miRNAs and GW182 requires both CCR4:NOT deadenylase and
DCP1:DCP2 decapping complexes. Genes Dev 2006 Jul 15;20(14):1885-98.
(111) Parker R, Sheth U. P bodies and the control of mRNA translation and degradation. Mol
Cell 2007 Mar 9;25(5):635-46.
(112) Yan M, Huang HY, Wang T, Wan Y, Cui SD, Liu ZZ, et al. Dysregulated Expression of
Dicer and Drosha in Breast Cancer. Pathol Oncol Res 2011 Sep 7.
(113) Kobel M, Gilks CB, Huntsman DG. Dicer and Drosha in ovarian cancer. N Engl J Med
2009 Mar 12;360(11):1150-1.
(114) Gong H, Liu CM, Liu DP, Liang CC. The role of small RNAs in human diseases:
potential troublemaker and therapeutic tools. Med Res Rev 2005 May;25(3):361-81.
(115) Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch E, Yendamuri S, et al. Human
microRNA genes are frequently located at fragile sites and genomic regions involved in
cancers. Proc Natl Acad Sci U S A 2004 Mar 2;101(9):2999-3004.
(116) Iwai N, Naraba H. Polymorphisms in human pre-miRNAs. Biochem Biophys Res
Commun 2005 Jun 17;331(4):1439-44.
(117) Huang Y, Zou Q, Song H, Song F, Wang L, Zhang G, et al. A study of miRNAs targets
prediction and experimental validation. Protein Cell 2010 Nov;1(11):979-86.

139
(118) Maragkakis M, Alexiou P, Papadopoulos GL, Reczko M, Dalamagas T, Giannopoulos G,
et al. Accurate microRNA target prediction correlates with protein repression levels.
BMC Bioinformatics 2009;10:295.
(119) Kertesz M, Iovino N, Unnerstall U, Gaul U, Segal E. The role of site accessibility in
microRNA target recognition. Nat Genet 2007 Oct;39(10):1278-84.
(120) Grimson A, Farh KK, Johnston WK, Garrett-Engele P, Lim LP, Bartel DP. MicroRNA
targeting specificity in mammals: determinants beyond seed pairing. Mol Cell 2007 Jul
6;27(1):91-105.
(121) Martens-Uzunova ES, Jalava SE, Dits NF, van Leenders GJ, Moller S, Trapman J, et al.
Diagnostic and prognostic signatures from the small non-coding RNA transcriptome in
prostate cancer. Oncogene 2011 Jul 18.
(122) Ambs S, Prueitt RL, Yi M, Hudson RS, Howe TM, Petrocca F, et al. Genomic profiling
of microRNA and messenger RNA reveals deregulated microRNA expression in prostate
cancer. Cancer Res 2008 Aug 1;68(15):6162-70.
(123) Porkka KP, Pfeiffer MJ, Waltering KK, Vessella RL, Tammela TL, Visakorpi T.
MicroRNA expression profiling in prostate cancer. Cancer Res 2007 Jul 1;67(13):6130-5.
(124) Schaefer A, Jung M, Mollenkopf HJ, Wagner I, Stephan C, Jentzmik F, et al. Diagnostic
and prognostic implications of microRNA profiling in prostate carcinoma. Int J Cancer
2010 Mar 1;126(5):1166-76.
(125) Tong AW, Fulgham P, Jay C, Chen P, Khalil I, Liu S, et al. MicroRNA profile analysis
of human prostate cancers. Cancer Gene Ther 2009 Mar;16(3):206-16.
(126) Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F, et al. A microRNA
expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad
Sci U S A 2006 Feb 14;103(7):2257-61.
(127) Musumeci M, Coppola V, Addario A, Patrizii M, Maugeri-Sacca M, Memeo L, et al.
Control of tumor and microenvironment cross-talk by miR-15a and miR-16 in prostate
cancer. Oncogene 2011 Oct 13;30(41):4231-42.
(128) Mitchell PS, Parkin RK, Kroh EM, Fritz BR, Wyman SK, Pogosova-Agadjanyan EL, et
al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc Natl
Acad Sci U S A 2008 Jul 29;105(30):10513-8.
(129) Xi Y, Nakajima G, Gavin E, Morris CG, Kudo K, Hayashi K, et al. Systematic analysis
of microRNA expression of RNA extracted from fresh frozen and formalin-fixed
paraffin-embedded samples. RNA 2007 Oct;13(10):1668-74.
(130) Jay C, Nemunaitis J, Chen P, Fulgham P, Tong AW. miRNA profiling for diagnosis and
prognosis of human cancer. DNA Cell Biol 2007 May;26(5):293-300.

140
(131) Nelson PT, Baldwin DA, Scearce LM, Oberholtzer JC, Tobias JW, Mourelatos Z.
Microarray-based, high-throughput gene expression profiling of microRNAs. Nat
Methods 2004 Nov;1(2):155-61.
(132) Epis MR, Giles KM, Barker A, Kendrick TS, Leedman PJ. miR-331-3p regulates ERBB-
2 expression and androgen receptor signaling in prostate cancer. J Biol Chem 2009 Sep
11;284(37):24696-704.
(133) Varambally S, Cao Q, Mani RS, Shankar S, Wang X, Ateeq B, et al. Genomic loss of
microRNA-101 leads to overexpression of histone methyltransferase EZH2 in cancer.
Science 2008 Dec 12;322(5908):1695-9.
(134) Yang K, Handorean AM, Iczkowski KA. MicroRNAs 373 and 520c are downregulated in
prostate cancer, suppress CD44 translation and enhance invasion of prostate cancer cells
in vitro. Int J Clin Exp Pathol 2009;2(4):361-9.
(135) Noonan EJ, Place RF, Pookot D, Basak S, Whitson JM, Hirata H, et al. miR-449a targets
HDAC-1 and induces growth arrest in prostate cancer. Oncogene 2009 Apr
9;28(14):1714-24.
(136) Mercatelli N, Coppola V, Bonci D, Miele F, Costantini A, Guadagnoli M, et al. The
inhibition of the highly expressed miR-221 and miR-222 impairs the growth of prostate
carcinoma xenografts in mice. PLoS One 2008;3(12):e4029.
(137) Bonci D, Coppola V, Musumeci M, Addario A, Giuffrida R, Memeo L, et al. The miR-
15a-miR-16-1 cluster controls prostate cancer by targeting multiple oncogenic activities.
Nat Med 2008 Nov;14(11):1271-7.
(138) Mihelich BL, Khramtsova EA, Arva N, Vaishnav A, Johnson DN, Giangreco AA, et al.
The miR-183-96-182 cluster is overexpressed in prostate tissue and regulates zinc
homeostasis in prostate cells. J Biol Chem 2011 Nov 1.
(139) Viticchie G, Lena AM, Latina A, Formosa A, Gregersen LH, Lund AH, et al. MiR-203
controls proliferation, migration and invasive potential of prostate cancer cell lines. Cell
Cycle 2011 Apr 1;10(7):1121-31.
(140) Gandellini P, Folini M, Longoni N, Pennati M, Binda M, Colecchia M, et al. miR-205
Exerts tumor-suppressive functions in human prostate through down-regulation of protein
kinase Cepsilon. Cancer Res 2009 Mar 15;69(6):2287-95.
(141) Lee KH, Chen YL, Yeh SD, Hsiao M, Lin JT, Goan YG, et al. MicroRNA-330 acts as
tumor suppressor and induces apoptosis of prostate cancer cells through E2F1-mediated
suppression of Akt phosphorylation. Oncogene 2009 Sep 24;28(38):3360-70.
(142) Hurst DR, Edmonds MD, Welch DR. Metastamir: the field of metastasis-regulatory
microRNA is spreading. Cancer Res 2009 Oct 1;69(19):7495-8.

141
(143) Watahiki A, Wang Y, Morris J, Dennis K, O'Dwyer HM, Gleave M, et al. MicroRNAs
associated with metastatic prostate cancer. PLoS One 2011;6(9):e24950.
(144) Schramedei K, Morbt N, Pfeifer G, Lauter J, Rosolowski M, Tomm JM, et al.
MicroRNA-21 targets tumor suppressor genes ANP32A and SMARCA4. Oncogene 2011
Feb 14.
(145) Leite KR, Sousa-Canavez JM, Reis ST, Tomiyama AH, Camara-Lopes LH, Sanudo A, et
al. Change in expression of miR-let7c, miR-100, and miR-218 from high grade localized
prostate cancer to metastasis. Urol Oncol 2009 Apr 15.
(146) Xu B, Niu X, Zhang X, Tao J, Wu D, Wang Z, et al. miR-143 decreases prostate cancer
cells proliferation and migration and enhances their sensitivity to docetaxel through
suppression of KRAS. Mol Cell Biochem 2011 Apr;350(1-2):207-13.
(147) Peng X, Guo W, Liu T, Wang X, Tu X, Xiong D, et al. Identification of miRs-143 and -
145 that is associated with bone metastasis of prostate cancer and involved in the
regulation of EMT. PLoS One 2011;6(5):e20341.
(148) Kojima S, Chiyomaru T, Kawakami K, Yoshino H, Enokida H, Nohata N, et al. Tumour
suppressors miR-1 and miR-133a target the oncogenic function of purine nucleoside
phosphorylase (PNP) in prostate cancer. Br J Cancer 2011 Nov 8.
(149) Lin SL, Chiang A, Chang D, Ying SY. Loss of mir-146a function in hormone-refractory
prostate cancer. RNA 2008 Mar;14(3):417-24.
(150) Leite KR, Tomiyama A, Reis ST, Sousa-Canavez JM, Sanudo A, Camara-Lopes LH, et
al. MicroRNA expression profiles in the progression of prostate cancer-from high-grade
prostate intraepithelial neoplasia to metastasis. Urol Oncol 2011 Aug 29.
(151) Huang Q, Gumireddy K, Schrier M, le SC, Nagel R, Nair S, et al. The microRNAs miR-
373 and miR-520c promote tumour invasion and metastasis. Nat Cell Biol 2008
Feb;10(2):202-10.
(152) Takeshita F, Patrawala L, Osaki M, Takahashi RU, Yamamoto Y, Kosaka N, et al.
Systemic delivery of synthetic microRNA-16 inhibits the growth of metastatic prostate
tumors via downregulation of multiple cell-cycle genes. Mol Ther 2010 Jan;18(1):181-7.
(153) Zhang X, Ladd A, Dragoescu E, Budd WT, Ware JL, Zehner ZE. MicroRNA-17-3p is a
prostate tumor suppressor in vitro and in vivo, and is decreased in high grade prostate
tumors analyzed by laser capture microdissection. Clin Exp Metastasis 2009;26(8):965-
79.
(154) Kong D, Li Y, Wang Z, Banerjee S, Ahmad A, Kim HR, et al. miR-200 regulates PDGF-
D-mediated epithelial-mesenchymal transition, adhesion, and invasion of prostate cancer
cells. Stem Cells 2009 Aug;27(8):1712-21.

142
(155) Steele R, Mott JL, Ray RB. MBP-1 upregulates miR-29b that represses Mcl-1, collagens,
and matrix-metalloproteinase-2 in prostate cancer cells. Genes Cancer 2010 Apr
1;1(4):381-7.
(156) Saini S, Majid S, Yamamura S, Tabatabai ZL, Suh SO, Shahryari V, et al. Regulatory
role of miR-203 in prostate cancer progression and metastasis. Clin Cancer Res 2010 Dec
15.
(157) Majid S, Dar AA, Saini S, Yamamura S, Hirata H, Tanaka Y, et al. MicroRNA-205-
directed transcriptional activation of tumor suppressor genes in prostate cancer. Cancer
2010 Dec 15;116(24):5637-49.
(158) Fujita Y, Kojima K, Hamada N, Ohhashi R, Akao Y, Nozawa Y, et al. Effects of miR-
34a on cell growth and chemoresistance in prostate cancer PC3 cells. Biochem Biophys
Res Commun 2008 Dec 5;377(1):114-9.
(159) Spahn M, Kneitz S, Scholz CJ, Stenger N, Rudiger T, Strobel P, et al. Expression of
microRNA-221 is progressively reduced in aggressive prostate cancer and metastasis and
predicts clinical recurrence. Int J Cancer 2010 Jul 15;127(2):394-403.
(160) Epis MR, Giles KM, Barker A, Kendrick TS, Leedman PJ. miR-331-3p regulates ERBB-
2 expression and androgen receptor signaling in prostate cancer. J Biol Chem 2009 Sep
11;284(37):24696-704.
(161) Musiyenko A, Bitko V, Barik S. Ectopic expression of miR-126*, an intronic product of
the vascular endothelial EGF-like 7 gene, regulates prostein translation and invasiveness
of prostate cancer LNCaP cells. J Mol Med 2008 Mar;86(3):313-22.
(162) Liu C, Kelnar K, Liu B, Chen X, Calhoun-Davis T, Li H, et al. The microRNA miR-34a
inhibits prostate cancer stem cells and metastasis by directly repressing CD44. Nat Med
2011 Feb;17(2):211-5.
(163) Seto AG. The road toward microRNA therapeutics. Int J Biochem Cell Biol 2010
Aug;42(8):1298-305.
(164) Kasinski AL, Slack FJ. MicroRNAs en route to the clinic: progress in validating and
targeting microRNAs for cancer therapy. Nat Rev Cancer 2011;11(12):849-64.
(165) Garzon R, Marcucci G, Croce CM. Targeting microRNAs in cancer: rationale, strategies
and challenges. Nat Rev Drug Discov 2010 Oct;9(10):775-89.
(166) Petrocca F, Lieberman J. Promise and challenge of RNA interference-based therapy for
cancer. J Clin Oncol 2011 Feb 20;29(6):747-54.
(167) Wiggins JF, Ruffino L, Kelnar K, Omotola M, Patrawala L, Brown D, et al. Development
of a lung cancer therapeutic based on the tumor suppressor microRNA-34. Cancer Res
2010 Jul 15;70(14):5923-30.

143
(168) McLaughlin J, Cheng D, Singer O, Lukacs RU, Radu CG, Verma IM, et al. Sustained
suppression of Bcr-Abl-driven lymphoid leukemia by microRNA mimics. Proc Natl
Acad Sci U S A 2007 Dec 18;104(51):20501-6.
(169) Bader AG, Brown D, Winkler M. The promise of microRNA replacement therapy.
Cancer Res 2010 Sep 15;70(18):7027-30.
(170) Zuhorn IS, Engberts JB, Hoekstra D. Gene delivery by cationic lipid vectors: overcoming
cellular barriers. Eur Biophys J 2007 Apr;36(4-5):349-62.
(171) Grimm D, Streetz KL, Jopling CL, Storm TA, Pandey K, Davis CR, et al. Fatality in
mice due to oversaturation of cellular microRNA/short hairpin RNA pathways. Nature
2006 May 25;441(7092):537-41.
(172) Lengauer C, Kinzler KW, Vogelstein B. Genetic instabilities in human cancers. Nature
1998 Dec 17;396(6712):643-9.
(173) Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000 Jan 7;100(1):57-70.
(174) Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell 2011 Mar
4;144(5):646-74.
(175) Yarden Y, Ullrich A. Growth factor receptor tyrosine kinases. Annu Rev Biochem
1988;57:443-78.
(176) Slamon DJ, Clark GM, Wong SG, Levin WJ, Ullrich A, McGuire WL. Human breast
cancer: correlation of relapse and survival with amplification of the HER-2/neu
oncogene. Science 1987 Jan 9;235(4785):177-82.
(177) Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000 Jan 7;100(1):57-70.
(178) Cheng N, Chytil A, Shyr Y, Joly A, Moses HL. Transforming growth factor-beta
signaling-deficient fibroblasts enhance hepatocyte growth factor signaling in mammary
carcinoma cells to promote scattering and invasion. Mol Cancer Res 2008
Oct;6(10):1521-33.
(179) Medema RH, Bos JL. The role of p21ras in receptor tyrosine kinase signaling. Crit Rev
Oncog 1993;4(6):615-61.
(180) Weinberg RA. The retinoblastoma protein and cell cycle control. Cell 1995 May
5;81(3):323-30.
(181) Adams JM, Cory S. The Bcl-2 apoptotic switch in cancer development and therapy.
Oncogene 2007 Feb 26;26(9):1324-37.
(182) Artandi SE, DePinho RA. Telomeres and telomerase in cancer. Carcinogenesis 2010
Jan;31(1):9-18.

144
(183) Sporn MB. The war on cancer. Lancet 1996 May 18;347(9012):1377-81.
(184) Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell 2011 Mar
4;144(5):646-74.
(185) Folkman J. Anti-angiogenesis: new concept for therapy of solid tumors. Ann Surg 1972
Mar;175(3):409-16.
(186) Leung DW, Cachianes G, Kuang WJ, Goeddel DV, Ferrara N. Vascular endothelial
growth factor is a secreted angiogenic mitogen. Science 1989 Dec 8;246(4935):1306-9.
(187) Barleon B, Sozzani S, Zhou D, Weich HA, Mantovani A, Marme D. Migration of human
monocytes in response to vascular endothelial growth factor (VEGF) is mediated via the
VEGF receptor flt-1. Blood 1996 Apr 15;87(8):3336-43.
(188) Rafii S, Lyden D. Therapeutic stem and progenitor cell transplantation for organ
vascularization and regeneration. Nat Med 2003 Jun;9(6):702-12.
(189) Olsson AK, Dimberg A, Kreuger J, Claesson-Welsh L. VEGF receptor signalling - in
control of vascular function. Nat Rev Mol Cell Biol 2006 May;7(5):359-71.
(190) Negrini S, Gorgoulis VG, Halazonetis TD. Genomic instability--an evolving hallmark of
cancer. Nat Rev Mol Cell Biol 2010 Mar;11(3):220-8.
(191) Grivennikov SI, Greten FR, Karin M. Immunity, inflammation, and cancer. Cell 2010
Mar 19;140(6):883-99.
(192) Gordanpour A, Nam RK, Sugar L, Bacopulos S, Seth A. MicroRNA Detection in
Prostate Tumors by Quantitative Real-time PCR (qPCR). J Vis Exp 2012;(63).
(193) Kumar-Sinha C, Tomlins SA, Chinnaiyan AM. Recurrent gene fusions in prostate cancer.
Nat Rev Cancer 2008 Jul;8(7):497-511.
(194) Clark J, Merson S, Jhavar S, Flohr P, Edwards S, Foster CS, et al. Diversity of
TMPRSS2-ERG fusion transcripts in the human prostate. Oncogene 2007 Apr
19;26(18):2667-73.
(195) Tomlins SA, Laxman B, Varambally S, Cao X, Yu J, Helgeson BE, et al. Role of the
TMPRSS2-ERG gene fusion in prostate cancer. Neoplasia 2008 Feb;10(2):177-88.
(196) Nam RK, Sugar L, Yang W, Srivastava S, Klotz LH, Yang LY, et al. Expression of the
TMPRSS2:ERG fusion gene predicts cancer recurrence after surgery for localised
prostate cancer. Br J Cancer 2007 Dec 17;97(12):1690-5.
(197) Gopalan A, Leversha MA, Satagopan JM, Zhou Q, Al-Ahmadie HA, Fine SW, et al.
TMPRSS2-ERG gene fusion is not associated with outcome in patients treated by
prostatectomy. Cancer Res 2009 Feb 15;69(4):1400-6.

145
(198) He L, Hannon GJ. MicroRNAs: small RNAs with a big role in gene regulation. Nat Rev
Genet 2004 Jul;5(7):522-31.
(199) Chen CZ, Li L, Lodish HF, Bartel DP. MicroRNAs modulate hematopoietic lineage
differentiation. Science 2004 Jan 2;303(5654):83-6.
(200) Xu P, Vernooy SY, Guo M, Hay BA. The Drosophila microRNA Mir-14 suppresses cell
death and is required for normal fat metabolism. Curr Biol 2003 Apr 29;13(9):790-5.
(201) Bueno MJ, Perez dC, I, Malumbres M. Control of cell proliferation pathways by
microRNAs. Cell Cycle 2008 Oct;7(20):3143-8.
(202) Calin GA, Croce CM. MicroRNA-cancer connection: the beginning of a new tale. Cancer
Res 2006 Aug 1;66(15):7390-4.
(203) Hurst DR, Edmonds MD, Welch DR. Metastamir: the field of metastasis-regulatory
microRNA is spreading. Cancer Res 2009 Oct 1;69(19):7495-8.
(204) Lapointe J, Kim YH, Miller MA, Li C, Kaygusuz G, van de Rijn M, et al. A variant
TMPRSS2 isoform and ERG fusion product in prostate cancer with implications for
molecular diagnosis. Mod Pathol 2007 Apr;20(4):467-73.
(205) Nam RK, Sugar L, Wang Z, Yang W, Kitching R, Klotz LH, et al. Expression of
TMPRSS2:ERG gene fusion in prostate cancer cells is an important prognostic factor for
cancer progression. Cancer Biol Ther 2007 Jan;6(1):40-5.
(206) Perner S, Demichelis F, Beroukhim R, Schmidt FH, Mosquera JM, Setlur S, et al.
TMPRSS2:ERG fusion-associated deletions provide insight into the heterogeneity of
prostate cancer. Cancer Res 2006 Sep 1;66(17):8337-41.
(207) Rajput AB, Miller MA, De LA, Boyd N, Leung S, Hurtado-Coll A, et al. Frequency of
the TMPRSS2:ERG gene fusion is increased in moderate to poorly differentiated prostate
cancers. J Clin Pathol 2007 Nov;60(11):1238-43.
(208) Attard G, Clark J, Ambroisine L, Fisher G, Kovacs G, Flohr P, et al. Duplication of the
fusion of TMPRSS2 to ERG sequences identifies fatal human prostate cancer. Oncogene
2008 Jan 10;27(3):253-63.
(209) Demichelis F, Fall K, Perner S, Andren O, Schmidt F, Setlur SR, et al. TMPRSS2:ERG
gene fusion associated with lethal prostate cancer in a watchful waiting cohort. Oncogene
2007 Jul 5;26(31):4596-9.
(210) Narod SA, Seth A, Nam R. Fusion in the ETS gene family and prostate cancer. Br J
Cancer 2008 Sep 16;99(6):847-51.
(211) Carver BS, Tran J, Gopalan A, Chen Z, Shaikh S, Carracedo A, et al. Aberrant ERG
expression cooperates with loss of PTEN to promote cancer progression in the prostate.
Nat Genet 2009 May;41(5):619-24.

146
(212) Squire JA. TMPRSS2-ERG and PTEN loss in prostate cancer. Nat Genet 2009
May;41(5):509-10.
(213) Garzon R, Calin GA, Croce CM. MicroRNAs in Cancer. Annu Rev Med 2009;60:167-
79.
(214) Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, et al. MicroRNA
expression profiles classify human cancers. Nature 2005 Jun 9;435(7043):834-8.
(215) Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F, et al. A microRNA
expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad
Sci U S A 2006 Feb 14;103(7):2257-61.
(216) Porkka KP, Pfeiffer MJ, Waltering KK, Vessella RL, Tammela TL, Visakorpi T.
MicroRNA expression profiling in prostate cancer. Cancer Res 2007 Jul 1;67(13):6130-5.
(217) Ozen M, Creighton CJ, Ozdemir M, Ittmann M. Widespread deregulation of microRNA
expression in human prostate cancer. Oncogene 2008 Mar 13;27(12):1788-93.
(218) Ambs S, Prueitt RL, Yi M, Hudson RS, Howe TM, Petrocca F, et al. Genomic profiling
of microRNA and messenger RNA reveals deregulated microRNA expression in prostate
cancer. Cancer Res 2008 Aug 1;68(15):6162-70.
(219) Spahn M, Kneitz S, Scholz CJ, Stenger N, Rudiger T, Strobel P, et al. Expression of
microRNA-221 is progressively reduced in aggressive prostate cancer and metastasis and
predicts clinical recurrence. Int J Cancer 2010 Jul 15;127(2):394-403.
(220) Iorio MV, Ferracin M, Liu CG, Veronese A, Spizzo R, Sabbioni S, et al. MicroRNA gene
expression deregulation in human breast cancer. Cancer Res 2005 Aug 15;65(16):7065-
70.
(221) Lee EJ, Gusev Y, Jiang J, Nuovo GJ, Lerner MR, Frankel WL, et al. Expression profiling
identifies microRNA signature in pancreatic cancer. Int J Cancer 2007 Mar
1;120(5):1046-54.
(222) Ciafre SA, Galardi S, Mangiola A, Ferracin M, Liu CG, Sabatino G, et al. Extensive
modulation of a set of microRNAs in primary glioblastoma. Biochem Biophys Res
Commun 2005 Sep 9;334(4):1351-8.
(223) He H, Jazdzewski K, Li W, Liyanarachchi S, Nagy R, Volinia S, et al. The role of
microRNA genes in papillary thyroid carcinoma. Proc Natl Acad Sci U S A 2005 Dec
27;102(52):19075-80.
(224) Calin GA, Ferracin M, Cimmino A, Di LG, Shimizu M, Wojcik SE, et al. A MicroRNA
signature associated with prognosis and progression in chronic lymphocytic leukemia. N
Engl J Med 2005 Oct 27;353(17):1793-801.

147
(225) Mercatelli N, Coppola V, Bonci D, Miele F, Costantini A, Guadagnoli M, et al. The
inhibition of the highly expressed miR-221 and miR-222 impairs the growth of prostate
carcinoma xenografts in mice. PLoS One 2008;3(12):e4029.
(226) Nam RK, Jewett MA, Krahn MD, Robinette MA, Tsihlias J, Toi A, et al. Delay in
surgical therapy for clinically localized prostate cancer and biochemical recurrence after
radical prostatectomy. Can J Urol 2003 Jun;10(3):1891-8.
(227) Nam RK, Trachtenberg J, Jewett MA, Toi A, Evans A, Emami M, et al. Serum insulin-
like growth factor-I levels and prostatic intraepithelial neoplasia: a clue to the relationship
between IGF-I physiology and prostate cancer risk. Cancer Epidemiol Biomarkers Prev
2005 May;14(5):1270-3.
(228) Nam RK, Zhang WW, Jewett MA, Trachtenberg J, Klotz LH, Emami M, et al. The use of
genetic markers to determine risk for prostate cancer at prostate biopsy. Clin Cancer Res
2005 Dec 1;11(23):8391-7.
(229) Nam RK, Toi A, Klotz LH, Trachtenberg J, Jewett MA, Loblaw A, et al. Nomogram
prediction for prostate cancer and aggressive prostate cancer at time of biopsy: utilizing
all risk factors and tumor markers for prostate cancer. Can J Urol 2006 Apr;13 Suppl 2:2-
10.
(230) Vesprini D, Nam RK, Trachtenberg J, Jewett MA, Tavtigian SV, Emami M, et al. HPC2
variants and screen-detected prostate cancer. Am J Hum Genet 2001 Apr;68(4):912-7.
(231) Hurst DR, Edmonds MD, Welch DR. Metastamir: the field of metastasis-regulatory
microRNA is spreading. Cancer Res 2009 Oct 1;69(19):7495-8.
(232) Xu S, Witmer PD, Lumayag S, Kovacs B, Valle D. MicroRNA (miRNA) transcriptome
of mouse retina and identification of a sensory organ-specific miRNA cluster. J Biol
Chem 2007 Aug 24;282(34):25053-66.
(233) Segura MF, Hanniford D, Menendez S, Reavie L, Zou X, Alvarez-Diaz S, et al. Aberrant
miR-182 expression promotes melanoma metastasis by repressing FOXO3 and
microphthalmia-associated transcription factor. Proc Natl Acad Sci U S A 2009 Feb
10;106(6):1814-9.
(234) Guttilla IK, White BA. Coordinate regulation of FOXO1 by miR-27a, miR-96, and miR-
182 in breast cancer cells. J Biol Chem 2009 Aug 28;284(35):23204-16.
(235) Ambs S, Prueitt RL, Yi M, Hudson RS, Howe TM, Petrocca F, et al. Genomic profiling
of microRNA and messenger RNA reveals deregulated microRNA expression in prostate
cancer. Cancer Res 2008 Aug 1;68(15):6162-70.
(236) Schaefer A, Jung M, Mollenkopf HJ, Wagner I, Stephan C, Jentzmik F, et al. Diagnostic
and prognostic implications of microRNA profiling in prostate carcinoma. Int J Cancer
2010 Mar 1;126(5):1166-76.

148
(237) Zhang H, Pan Y, Zheng L, Choe C, Lindgren B, Jensen ED, et al. FOXO1 inhibits Runx2
transcriptional activity and prostate cancer cell migration and invasion. Cancer Res 2011
May 1;71(9):3257-67.
(238) Gordanpour A, Stanimirovic A, Nam RK, Moreno CS, Sherman C, Sugar L, et al. miR-
221 Is down-regulated in TMPRSS2:ERG fusion-positive prostate cancer. Anticancer
Res 2011 Feb;31(2):403-10.
(239) Long Q, Johnson BA, Osunkoya AO, Lai YH, Zhou W, Abramovitz M, et al. Protein-
coding and microRNA biomarkers of recurrence of prostate cancer following radical
prostatectomy. Am J Pathol 2011 Jul;179(1):46-54.
(240) Rhodes DR, Yu J, Shanker K, Deshpande N, Varambally R, Ghosh D, et al.
ONCOMINE: a cancer microarray database and integrated data-mining platform.
Neoplasia 2004 Jan;6(1):1-6.
(241) Lapointe J, Li C, Higgins JP, van de Rijn M, Bair E, Montgomery K, et al. Gene
expression profiling identifies clinically relevant subtypes of prostate cancer. Proc Natl
Acad Sci U S A 2004 Jan 20;101(3):811-6.
(242) LaTulippe E, Satagopan J, Smith A, Scher H, Scardino P, Reuter V, et al. Comprehensive
gene expression analysis of prostate cancer reveals distinct transcriptional programs
associated with metastatic disease. Cancer Res 2002 Aug 1;62(15):4499-506.
(243) Tomlins SA, Mehra R, Rhodes DR, Cao X, Wang L, Dhanasekaran SM, et al. Integrative
molecular concept modeling of prostate cancer progression. Nat Genet 2007
Jan;39(1):41-51.
(244) Ambs S, Prueitt RL, Yi M, Hudson RS, Howe TM, Petrocca F, et al. Genomic profiling
of microRNA and messenger RNA reveals deregulated microRNA expression in prostate
cancer. Cancer Res 2008 Aug 1;68(15):6162-70.
(245) Gordanpour A, Stanimirovic A, Nam RK, Moreno CS, Sherman C, Sugar L, et al. miR-
221 Is down-regulated in TMPRSS2:ERG fusion-positive prostate cancer. Anticancer
Res 2011 Feb;31(2):403-10.
(246) Stittrich AB, Haftmann C, Sgouroudis E, Kuhl AA, Hegazy AN, Panse I, et al. The
microRNA miR-182 is induced by IL-2 and promotes clonal expansion of activated
helper T lymphocytes. Nat Immunol 2010 Nov;11(11):1057-62.
(247) Segura MF, Hanniford D, Menendez S, Reavie L, Zou X, Alvarez-Diaz S, et al. Aberrant
miR-182 expression promotes melanoma metastasis by repressing FOXO3 and
microphthalmia-associated transcription factor. Proc Natl Acad Sci U S A 2009 Feb
10;106(6):1814-9.
(248) Guttilla IK, White BA. Coordinate regulation of FOXO1 by miR-27a, miR-96, and miR-
182 in breast cancer cells. J Biol Chem 2009 Aug 28;284(35):23204-16.

149
(249) Huang H, Tindall DJ. Dynamic FoxO transcription factors. J Cell Sci 2007 Aug 1;120(Pt
15):2479-87.
(250) Kau TR, Schroeder F, Ramaswamy S, Wojciechowski CL, Zhao JJ, Roberts TM, et al. A
chemical genetic screen identifies inhibitors of regulated nuclear export of a Forkhead
transcription factor in PTEN-deficient tumor cells. Cancer Cell 2003 Dec;4(6):463-76.
(251) Dong XY, Chen C, Sun X, Guo P, Vessella RL, Wang RX, et al. FOXO1A is a candidate
for the 13q14 tumor suppressor gene inhibiting androgen receptor signaling in prostate
cancer. Cancer Res 2006 Jul 15;66(14):6998-7006.
(252) Liu C, Kelnar K, Liu B, Chen X, Calhoun-Davis T, Li H, et al. The microRNA miR-34a
inhibits prostate cancer stem cells and metastasis by directly repressing CD44. Nat Med
2011 Feb;17(2):211-5.
(253) Galardi S, Mercatelli N, Giorda E, Massalini S, Frajese GV, Ciafre SA, et al. miR-221
and miR-222 expression affects the proliferation potential of human prostate carcinoma
cell lines by targeting p27Kip1. J Biol Chem 2007 Aug 10;282(32):23716-24.
(254) Mihelich BL, Khramtsova EA, Arva N, Vaishnav A, Johnson DN, Giangreco AA, et al.
miR-183-96-182 cluster is overexpressed in prostate tissue and regulates zinc
homeostasis in prostate cells. J Biol Chem 2011 Dec 30;286(52):44503-11.
(255) Schuster SC. Next-generation sequencing transforms today's biology. Nat Methods 2008
Jan;5(1):16-8.
(256) Rusk N, Kiermer V. Primer: Sequencing--the next generation. Nat Methods 2008
Jan;5(1):15.
(257) Shendure J, Ji H. Next-generation DNA sequencing. Nat Biotechnol 2008
Oct;26(10):1135-45.
(258) Hodges E, Xuan Z, Balija V, Kramer M, Molla MN, Smith SW, et al. Genome-wide in
situ exon capture for selective resequencing. Nat Genet 2007 Dec;39(12):1522-7.
(259) Ng SB, Turner EH, Robertson PD, Flygare SD, Bigham AW, Lee C, et al. Targeted
capture and massively parallel sequencing of 12 human exomes. Nature 2009 Sep
10;461(7261):272-6.
(260) Wheeler DA, Srinivasan M, Egholm M, Shen Y, Chen L, McGuire A, et al. The complete
genome of an individual by massively parallel DNA sequencing. Nature 2008 Apr
17;452(7189):872-6.
(261) Pan Q, Shai O, Lee LJ, Frey BJ, Blencowe BJ. Deep surveying of alternative splicing
complexity in the human transcriptome by high-throughput sequencing. Nat Genet 2008
Dec;40(12):1413-5.

150
(262) Wilhelm BT, Marguerat S, Watt S, Schubert F, Wood V, Goodhead I, et al. Dynamic
repertoire of a eukaryotic transcriptome surveyed at single-nucleotide resolution. Nature
2008 Jun 26;453(7199):1239-43.
(263) Mortazavi A, Williams BA, McCue K, Schaeffer L, Wold B. Mapping and quantifying
mammalian transcriptomes by RNA-Seq. Nat Methods 2008 Jul;5(7):621-8.
(264) Coppola V, De MR, Bonci D. MicroRNAs and prostate cancer. Endocr Relat Cancer
2010 Mar;17(1):F1-17.
(265) Nam RK, Sugar L, Yang W, Srivastava S, Klotz LH, Yang LY, et al. Expression of the
TMPRSS2:ERG fusion gene predicts cancer recurrence after surgery for localised
prostate cancer. Br J Cancer 2007 Dec 17;97(12):1690-5.