Embed Size (px)
Citation preview

1
for Vitis 1 Braunschweig, Germany 2 3
Oviposition behaviour and life-history performance of Epiphyas postvittana 4
(Lepidoptera: Tortricidae) on the leaves of Vitis vinifera (Vitales: Vitaceae) 5
infected with Botrytis cinerea (Helotiales: Sclerotiniaceae) 6 7
S. Z. M. RIZVI and A. RAMAN* 8
Charles Sturt University & Graham Centre for Agricultural Innovation, PO Box 883, Orange, NSW 2800, 9 Australia 10
11 12
Summary 13 14 In three-way interaction systems involving an insect and a plant-pathogenic fungus, 15
both occurring on the same plant, the insects generally gain in terms of their growth and 16
metabolism. In this study we have tested how the infection by Botrytis cinerea on the leaves 17
of Vitis vinifera influences the life-history performance of larvae and the oviposition 18
behaviour of Epiphyas postvittana. We conducted free-choice and two-choice experiments to 19
test the oviposition behaviour of gravid E. postvittana. We also characterized the effects of B. 20
cinerea-infected leaves of V. vinifera on the growth and development of E. postvittana. We 21
found that the oviposition preference of E. postvittana was strongly influenced by the 22
olfactory and tactile cues. Volatiles from B. cinerea-infected plants significantly deterred 23
oviposition and in consequence, adult females laid fewer eggs on B. cinerea-infected leaves of 24
V. vinifera compared with uninfected leaves. The mortality rate of larvae fed on B. cinerea-25
infected leaves were not significantly different from the larvae fed on uninfected leaves of V. 26
vinifera. Whereas, the larvae of E. postvittana fed on B. cinerea-infected leaves had 27
significantly shorter developmental period, attained heavier pupal mass, and on becoming 28
adults they laid more numbers of eggs than the larvae that were enabled to feed on 29
uninfected leaves of V. vinifera. We also reared the larvae of E. postvittana on exclusive-30
fungus diet but all larvae died before pupation indicating that for a better larval performance 31
and adult reproductive output of E. postvittana, the V. vinifera—B. cinerea interacting system 32
is but imperative. 33
34 35 Key words: grapevine, larval development, light-brown apple moth, oviposition deterrence, 36
three-way interaction. 37 38
Running t it le: Effect of grey mould on grapevine moth 39 ___________________________________________________________________________________ 40 *Correspondence to: Anantanarayanan Raman, Charles Sturt University, PO Box 883, Orange, NSW 2800, 41 Australia. Tel. and FAX: +61 2 63657833. E-mail: [email protected] 42

2
Introduction 43
Infection by pathogenic fungi alters the chemistry of host plants and influences 44
oviposition preference and larval performance in plant-feeding insects (MONDY and CORIO-COSTET 45
2000; RIZVI et al. 2014a). Fungal infections can interfere with the biosynthetic pathways in the 46
plant in various ways. Some lead to a decrease in the production of insect-attracting volatiles 47
(DÖTTERL et al. 2009), whereas others produce several behaviour-modifying volatiles (TASIN et al. 48
2011). Fungal infection of a plant can suppress its direct defence capability against plant-feeding 49
insects by modifying the plant’s secondary metabolism (THALER et al. 1999) and the nutritional 50
quality of the plant, thus rendering the plant susceptible for insect colonization (CARDOZA et al. 51
2003). On the other hand, fungal infection can deteriorate the nutritional quality of the plant and 52
influence the plant-feeding insect’s development negatively (TASIN et al. 2012). 53
Olfactory cues are critical for searching a suitable host from a distance for oviposition but 54
once the insect lands on the target plant, the post-landing cues, viz., contact, together with the 55
volatiles can influence the insect for an assessment of the plant and can either deter or stimulate 56
oviposition (RENWICK and CHEW 1994; FOSTER et al. 1997; TASIN et al. 2011). Based on the sensory 57
cues of a plant-feeding insect, judgment on the appropriateness of a host plant is essential for the 58
success of progeny performance (GRIPENBERG et al. 2010). Optimization theory of host searching 59
suggests that the gravid female should choose those plants, which maximizes the fitness of her 60
progeny (JAENIKE, 1978), although evidences negating Jaenike (1978) also exist (LARSSON and EKBOM 61
1995; LEYVA et al. 2000). The choice made by juveniles of insects for food can substantially differ 62
from that made by adults, particularly when the larvae and adults feed on different plant parts 63
(MAYHEW 1997). In the natural environment, several organisms attack plants; among these, insects 64
and fungi are associated with a majority of plants, establishing a three-way interacting system. In 65
such contexts, the fungi influence the interaction between the insect and the plant, which can be 66
either mutualistic or antagonistic, and, occasionally, neutral as well (HATCHER 1995). The fungi, in 67
such interacting systems, can induce multiple variations in plant chemistry that can be 68
recognized by plant-feeding insects by their olfactory capacity, which, in turn, can modify the 69
preference behaviour in insects (NAJAR-RODRIGUEZ et al. 2010) 70
Nevertheless, in three-way interacting systems involving an insect and a plant-pathogenic 71
fungus, both developing on the same plant, the insects generally gain in terms of their growth 72
and metabolism. Leaf tissues of Arachis hypogaea (Fabales: Fabaceae)-infected with Sclerotium 73
rolfsii (Atheliales: Atheliaceae) showed significantly greater levels of soluble sugars but lower 74
starch content and total soluble phenolics compared with uninfected leaves of A. hypogaea. 75

3
Spodoptera exigua (Lepidoptera: Noctuidae) showed significantly greater survival rates, produced 76
significantly heavier pupae, and had shorter time span between the last larval instar and 77
pupation, when fed on the foliage of A. hypogaea infected with S. rolfsii (CARDOZA et al. 2003). For 78
instance, in the Botrytis cinerea—Lobesia botrana (Lepidoptera: Tortricidae)—V. vinifera 79
interacting system, L. botrana have been demonstrated to gain from B. cinerea by acquiring 80
sterols, which enhanced their ability to metamorphose rapidly gaining greater biomass (MONDY 81
and CORIO-COSTET 2000). The nutrient composition and levels of fungal infection in plants 82
influence the fitness and reproductive performance of an insect (BIERE and TACK 2013). These 83
findings indicate that the Lepidoptera gain from fungi associated with plants, as demonstrated by 84
accelerated growth rate, although in essence the insect can feed only on plant tissue and still 85
perform. 86
Keeping the above in view, we used a three-way interacting system of Epiphyas 87
postvittana—Vitis vinifera—Botrytis cinerea and proposed that that E. postvittana co-occurs with V. 88
vinifera infected by B. cinerea, and that for a better larval performance and oviposition behaviour 89
of E. postvittana, V. vinifera—B. cinerea interacting system is imperative. Epiphyas postvittana, an 90
Australian native Tortricidae, damages leaves and fruits of V. vinifera grown in Australasia 91
extensively. This species was accidentally introduced into England, Ireland, the Netherlands, 92
Sweden, besides Hawaii and California of USA (SUCKLING and BROCKERHOFF 2010). Field evidences 93
and laboratory studies indicate that the larvae of E. postvittana transmit the conidia of B. cinerea 94
(RIZVI et al. 2014b). Botrytis cinerea is an opportunistic necrotrophic pathogen that induces grey-95
mould disease on the leaves and fruits of V. vinifera (FOURNIER et al. 2013). 96
In this study, we sought answers to the following questions: (1) whether the females of E. 97
postvittana prefer ovipositing on V. vinifera infected by B. cinerea, (2) whether B. cinerea-infected 98
leaves positively influence larval performance of E. postvittana, and (3) whether the larvae of E. 99
postvittana can survive and develop feeding exclusively on B. cinerea. To secure answers, we 100
tested the oviposition preference of E. postvittana using B. cinerea-infected and uninfected leaves 101
of V. vinifera To test the larval performance with or without B. cinerea, we raised the larvae of E. 102
postvittana on uninfected and B. cinerea-infected V. vinifera leaves, using standard plant-nutrient 103
solution (e.g., Knop’s solution) as well as on standard synthetic culture media (viz., potato-104
dextrose agar [PDA], Murashige and Skoog medium [M−S medium]). To verify the survival and 105
development rate of the larvae of E. postvittana, we reared the larvae on B. cinerea cultured on 106
PDA and M−S medium, without V. vinifera. 107
108

4
Material and Methods 109 110 Insect culture 111 112 The larvae of E. postvittana were cultured on a Phaseolus lunatus-based semi-synthetic 113
diet (CUNNINGHAM 2007) and maintained at 21±1°C, 60―80% RH, and 16L: 8D regimen. On 114
emergence, an adult female was placed with a similar-aged adult male to facilitate mating as 115
described previously (RIZVI et al. 2014b). All eggs (5—6 days old) were surface-sterilized using 5% 116
formaldehyde solution and then washed with sterilized water following RAMAN and BEIDERBECK 117
(1992). 118
119 Fungus culture and preparation of conidial suspension 120 121 Botrytis cinerea isolated from infected berries of V. vinifera cv. Chardonnay from the 122
CSU—O’s Chardonnay vineyard in February 2013 was used. Stock culture was maintained on PDA 123
(Fisher Scientific, Inc., Scoresby, Victoria, Australia) at 22°C and 12L: 12D. The conidial suspension 124
was prepared in sterile water following RIZVI et al. (2014a) and adjusted to 106
conidia/ml. 125
126 Inoculation of V. vinifera leaves with B. cinerea 127 128 Sixty plants of V. vinifera cv. Chardonnay (hereafter V. vinifera) were raised in 129
polypropylene pots (17 cm tall, 10 cm Ø) containing commercial potting mix (Osmocote, Plus 130
Organics Vegetable & Herb Mix, Sydney, Australia) in an insect-proof glasshouse at 23±2°C, 60% 131
RH, and 16L:8D. Six-week old plants were used in the trials. The conidial suspension was sprayed 132
using a hand-held atomizer to infect the leaves of V. vinifera. Uninfected (control) leaves were 133
sprayed with sterile water identically. The inoculated and uninoculated plants were covered with 134
a polyethylene bag to enhance humidity and placed in an environmental chamber (Thermoline 135
L+M, Thermoline Scientific, NSW, Australia) (23°C, 70% RH, and 16L: 8D). Those plants which 136
manifested disease symptoms were used in the experiments. The infection level of B. cinerea- was 137
measured in percentage, from the photographs made with a digital camera (D―60, Canon, Tokyo, 138
Japan) and calculating the area of infection using ‘ImageJ’, an open-source image processing 139
software (ABRAMOFF et al. 2004). Those plants which showed 10—30% infection level were used in 140
the oviposition bioassays. 141
142 Oviposition behaviour 143 144 Free-choice experiment 145 146 Three uninfected and three B. cinerea-infected plants were placed in an aluminium-mesh 147
cage (200x80x80 cm3
). The plants were placed 10 cm away from the edge of the cage and were 148

5
distributed in two rows, separated by 40 cm between plants. Uninfected and infected plants were 149
placed alternately (Figure 1). Ten gravid females of E. postvittana were released at the midpoint of 150
the cage allowing the insects to choose either B. cinerea-infected or uninfected plants for 151
oviposition. The setup was left undisturbed. After 72 h, the plants were verified for eggs and the 152
number of eggs was counted under a stereo-binocular microscope (S–20, AIS Instrument Services, 153
Croydon, Victoria, Australia). This experiment was repeated six times. In each replicate new plants 154
were used and the position of the infected and uninfected plants were randomized. 155
156 Two-choice experiment 157 158
One B. cinerea-infected and one uninfected plants were individually placed in two glass 159
containers (60x30x30 cm3
), which had tiny holes on the one side to facilitate the passage of air. 160
The two containers were connected by a horizontal glass tube (30 cm long, 3 cm Ø). At the mid-161
point of the horizontal glass tube, a 2-cm wide circular port was cut to introduce the moths 162
(Figure 2). Five gravid individuals of E. postvittana were introduced through the port, enabling 163
them to choose either B. cinerea-infected or uninfected plant for oviposition. Soon after 164
introduction, the port was closed using a Parafilm® strip. After 72 h, the insects were removed. 165
The choices made by the insects were recorded and the numbers of eggs laid on B. cinerea-166
infected and uninfected V. vinifera were counted under a stereo-binocular microscope. This 167
experiment was repeated six times. In each replicate new plants were used and the position of the 168
infected and uninfected plants were randomized. 169
170
Larval development 171 172 On B. c inerea cultured on synthetic media 173 174 Thirty Petrie plates (9 cm Ø) of PDA were inoculated using a 4 mm
2
PDA block from stock 175
culture of B. cinerea. These plates were incubated at 21°C, 12L: 12D. Thirty sterile PDA plates were 176
used as control (Figure 3). After seven days, ensuring that B. cinerea was growing in the newly 177
introduced plates, the 4 mm2
PDA block was removed. Two neonate larvae (<4 h after emergence) 178
from sterile source were placed in every inoculated and sterile PDA plates, which were sealed 179
immediately using Parafilm. The plates were then incubated at 21°C, 12L: 12D. An identical 180
procedure was applied to raise the larvae of E. postvittana on aseptically cultured B. cinerea on 181
M−S medium (MURASHIGE and SKOOG 1972). All procedures were conducted in the clean-air 182
environment of the horizontal laminar-airflow bench (HWS120, Clyde–Apac, Sydney, Australia). 183
184

6
On B. cinerea - infected and uninfected leaves of V. vinifera maintained on 185 nutrient medium and on synthetic media 186
187 From freshly obtained branches of glasshouse-raised V. vinifera, several 20-cm stem 188
segments bearing 5―6 fully unfurled leaves were excised for use in this experiment. The stem 189
segments were surface-sterilized using 1% NaClO solution for 5 min and rinsed in sterile water 190
(3x). The leaves on these excised stem segments were then sprayed with the spore suspension in a 191
laminar-airflow cabinet. The leaves on stem segments used as control were sprayed with sterile 192
water. The basal tip of each segment was fixed on an Oasis® floral foam (4 cm3
) soaked in Knop’s 193
solution (RYCHTER and MIKULSKA 1990). The inoculated and uninoculated leaf-bearing stem 194
segments were placed in a zip-lock plastic bag (35x40 cm2
), which included a damp-filter paper. 195
The set up was maintained at 22°C and 12L: 12D for 4―5 d. 196
Only those leaves bearing around 10% infection level were used in this experiment. The 197
stem segments bearing either infected or uninfected leaves were placed separately in glass jars 198
(22 cm tall, 15 cm Ø) (Figure 4). Two neonate larvae (<4 h after emergence) were placed on 199
infected and uninfected leaves and maintained at 21±1°C, 60−70% RH, and 16L: 8D. After every 200
four days, the stem segments were changed to maintain the infection level under 30% of leaf area 201
and frass was removed from the plate. Fifty larvae were used for each treatment. Data pertaining 202
to the rate of survival of the larvae were collected every four days. The dates of pupation, number 203
of pupae, and pupal mass were recorded. Pupal mass was used as an indicator of adult-body 204
mass, because adult body mass cannot be obtained without killing them. Each pupa was 205
quarantined to a stoppered glass vial (4 cm tall, 2 cm Ø) until emergence. Adult emergence and sex 206
ratio (%) were recorded. The adults were sexed and paired in a corrugated-wall Dixie cup enabling 207
mating and oviposition. The adults enabled to mate and oviposit were subsequently measured for 208
their performance: fecundity rate, possible delays in oviposition (measured in days), viability 209
(measured as the number of emerged larvae). The eggs were incubated at 21±1°C, 60−70% RH, and 210
16L: 8D for 15 d enabling larval emergence. The number of days from the date of emergence of 211
adults to death was also recorded. 212
Life-history performance of E. postvittana on B. cinerea-infected or uninfected leaf was 213
further verified through using two aseptic nutrient media. Excised surface-sterilized leaf 214
(petioles+laminae) of V. vinifera was fixed onto either the PDA or M−S medium in a test tube (10 215
cm long, 1.5 cm ø, sealed with Parafilm) by pushing the petiole through the solid medium (Figure 216
5). Leaves were sprayed with spore suspension, whereas sterile water sprays were used in control. 217
All work was done on laminar-airflow bench, maintained at 22°C and 12L: 12D. 218

7
Leaves with 10% infection were used for rearing the larvae of E. postvittana. Two larvae 219
were released on each leaf. Every four days, the survival larvae were noted and larvae were 220
shifted onto either a new infected or a new uninfected leaf set up in the PDA or M−S medium as 221
described above. The larvae were allowed to pupate and the life-history traits were noted. Fifty 222
larvae were used for each treatment. Aseptic culture system has provided an environment to 223
experimental verify the life-history performance of many plant feeding insects. One of the 224
offshoots of such trials is raising the plant-feeding arthropods on their host plants in control 225
environment (e.g., FORNECK et al. 1998; FORNECK et al. 1999). 226
Statistical analysis 227 228 In the oviposition bioassay, the numbers of eggs laid on B. cinerea-infected and 229
uninfected leaves in free-choice experiment and two-choice experiment were analyzed using 230
paired sample ‘t’ test. Linear regression was applied to discriminate the significant difference in 231
the mortality rate of larvae applying ‘y=a+bx’, where x is the day and y is the mortality. A 232
contingency table (2
) was used to analyze female sex ratio, survival percentage of larvae from egg 233
hatching to pupae, and survival percentage of pupae from pupation to adult emergence. 234
Independent sample t-test was used to analyse all other life-history traits. Analyses were made 235
with SPSS statistic 17.0 (1993―2007) and GenStat (VSN International 2012). Graphs were 236
generated in MS Excel 2013 and GenStat. 237
Results 238 239
Oviposition behaviour 240 241 Free-choice experiment 242 243 Epiphyas postvittana females deposited significantly more number of eggs on uninfected 244
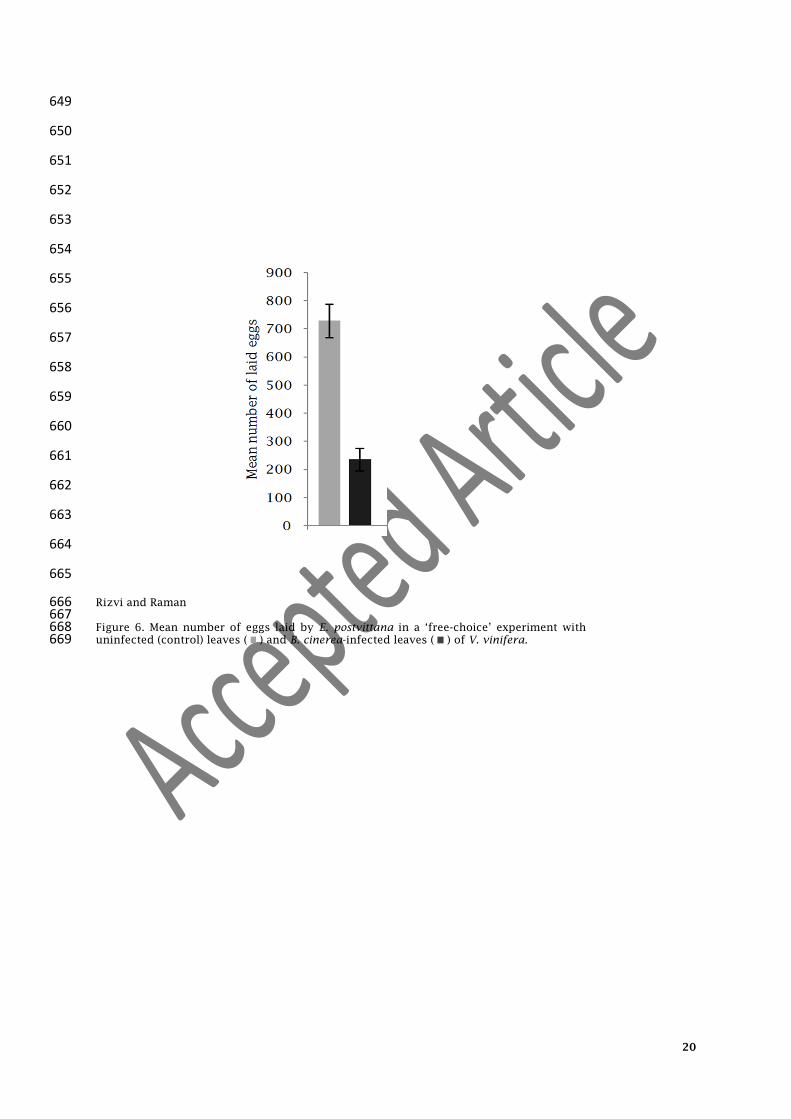
leaves than on B. cinerea-infected leaves (uninfected vs infected, 732.6±59.44 vs 236±40.5, t= 245
13.91, p<0.001, Figure 6). 246
247 Two-choice Experiment 248 249 Gravid females of E. postvittana showed a significant preference for uninfected leaves 250
(73.7%) as against the infected leaves (26.3%, n=30, χ2
=6.53, p<0.05, Figure 7). The B. cinerea-251
infected leaves significantly inhibited oviposition, compared with the uninfected leaves 252
(uninfected vs infected, 399±64vs 82±34, t= 4.36, p<0.01, Figure 8). 253
254 Larval development 255 256 On B. c inerea cultured on potato-dextrose agar 257 258

8
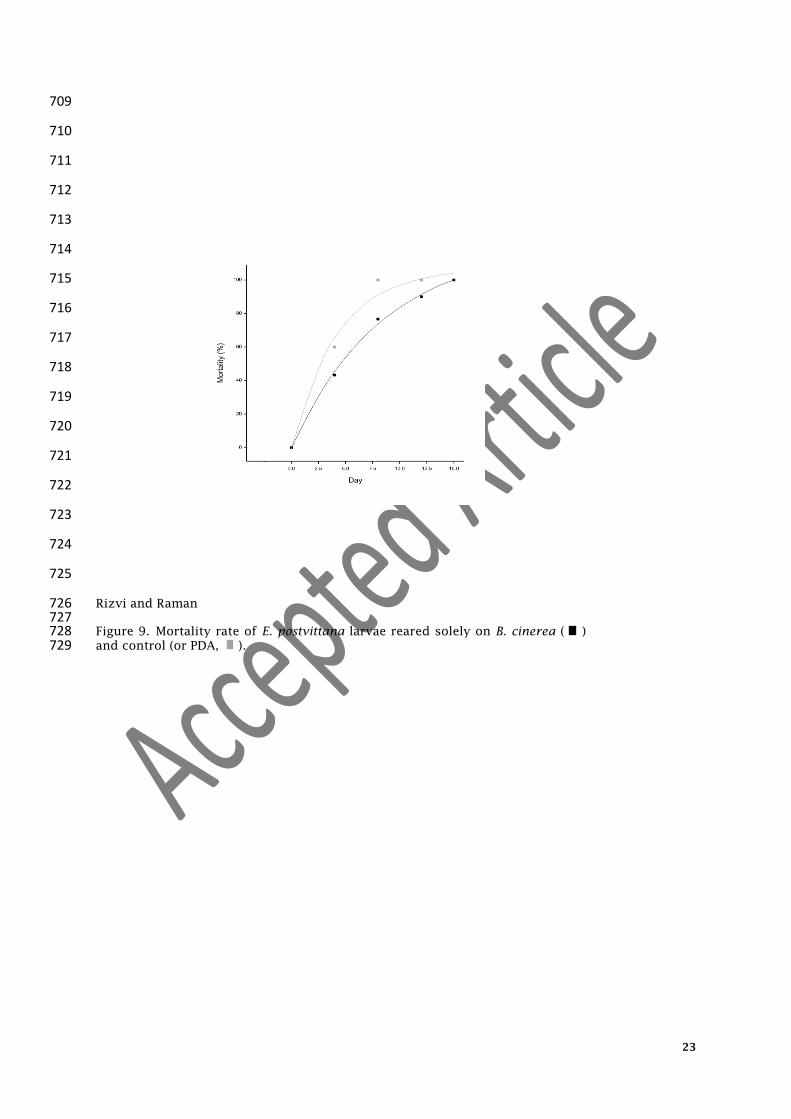
The mortality rate of the larvae of E. postvittana fed on B. cinerea on PDA 259
(y=120.6−120.4*0.89^
x) was significantly (p=0.039) lesser than the larvae that were fed only on 260
PDA (y=106.61-107.57*0.78^
x, where x represented time (days) and y represented the mortality 261
rate (Figure 9). All larvae were dead at conclusion (maximum15 d). 262
263 On B. c inerea cultured on Murashige and Skoog medium 264 265 The mortality rate of the larvae of E. postvittana fed on B. cinerea cultured (y=119.6-266
119.5*0.87^
x) was significantly (p= 0.027) fewer than the larvae fed on only M−S medium 267
(y=102.69-102.94*0.71^
x, where x = day and y = mortality rate (Figure 10). All larvae were dead at 268
conclusion (15 d). 269
On B. cinerea - infected and uninfected leaves of V. vinifera maintained on 270 Knop’s solution 271
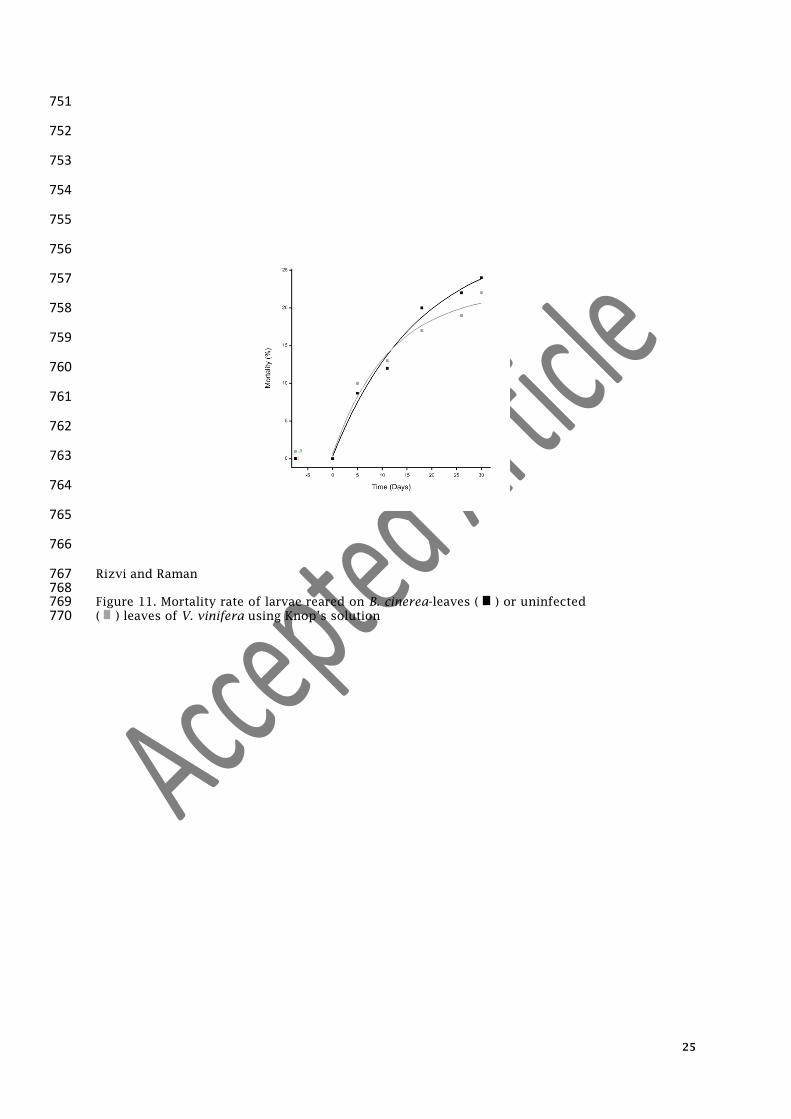
272 Larvae reared on B. cinerea-infected leaves developed faster than larvae reared on 273
uninfected leaves of V. vinifera (Table 1). The mortality rate of larvae fed on B. cinerea-infected 274
leaves (y=29.68-30.55*0.92^
x) was not significantly different than those reared on uninfected 275
leaves of V. vinifera (y=19.65-19.65*0.86^
x), where x = day and y = mortality rate (Figure 11). Pupal 276
masses of female moths reared on B. cinerea-infected leaves of V. vinifera were significantly 277
greater (p<0.05) than larvae reared on uninfected leaves (Table2). Botrytis cinerea infection did 278
not affect pupation and adult emergence percentage, sex ratio of adult and the length of pre-279
oviposition period (Tables3). Botrytis cinerea-infected diet significantly increased the fecundity 280
rates (p<0.05) and viability of eggs (p<0.001) compared with uninfected leaves (Table 3). 281
We found similar effects on larval development and adult performance of E. postvittana, 282
when raised on uninfected or B. cinerea-infected leaves of V. vinifera using PDA and M–S media. 283
284
Discussion 285 286
Gravid E. postvittana prefer to oviposit on uninfected leaves as against that of B. cinerea-infected 287
leaves of V. vinifera. The larvae of E. postvittana which feed on B. cinerea-infected leaves develop 288
quicker, attain a heavier pupal mass particularly in those developing as females, and the adults 289
lay more numbers of eggs than those raised on uninfected leaves of V. vinifera. The viability rate 290
of eggs hatching to neonates too is greater in those adults reared on B. cinerea-infected leaves. 291
292 Females of E. postvittana do not prefer ovipositing on B. cinerea -293
infected leaves of V. vinifera 294 295
In the present study, the gravid individuals of E. postvittana were deterred during oviposition by 296
the olfactory cues arising from V. vinifera leaves infected by B. cinerea. In free-choice experiment 297

9
of this study, when the gravid females of E. postvittana had to choose sites for oviposition among 298
B. cinerea-infected and uninfected leaves of V. vinifera, thus mimicking the natural condition, 299
they deposited significantly more number of eggs on uninfected leaves compared with B. cinerea-300
infected leaves. This observation concurs with our previous finding (RIZVI et al. 2014 a, b), 301
wherein gravid E. postvittana avoided oviposition on the infected leaves and berries of V. vinifera, 302
but laid significantly greater number of eggs on the uninfected leaves and berries of V. vinifera. 303
Preference for oviposition in different species of Lepidoptera has been shown to be regulated 304
generally by either stimulatory or inhibitory cues arising from host plants (RENWICK 1989). Fungus 305
infection can modify the volatiles arising from a plant by interfering with the plant’s biosynthetic 306
pathways. Volatiles, arising during pathogenic interactions between plants and fungi, act either as 307
attractants (COSSÉ et al.1994; CARDOZA et al. 2003) or deterrents (DÖTTERL et al. 2009; Tasin et al. 308
2011) for insects in three-way, three-component interacting systems. Infection by B. cinerea on V. 309
vinifera alters its volatile complexion either by changing their quantity or by producing new 310
compounds, such as alcohols, terpenes, and benzoates (TASIN et al. 2012). Botrytis cinerea-infected 311
berries of V. vinifera include several new alcohols (TASIN et al. 2012), which deter L. botrana. 312
Receptors on the dendritic membrane of sensory neurons of E. postvittana and their specific 313
receptivity reiterate that E. postvittana is capable of recognizing a range of volatiles (e.g., 314
alcohols, terpenoids and benzoates) and volatile-stress signals such as methyl salicylate (JORDAN et 315
al. 2009). 316
A gravid Lepidoptera usually prefers, for oviposition, those sites which maximize its 317
opportunities for a better performance of her progeny (JAENIKE 1978). This builds on the premise 318
that juvenile-life stages have a limited capacity to move on host surfaces and therefore the gravid 319
female chooses the best possible site for oviposition guaranteeing reasonable nutrition to her 320
offspring. In contrast, several arguments and explanations prevail indicating that adult nutrition 321
influences oviposition decision (MAYHEW 1997; SCHEIRS and DE BRUYN 2002). For example, 322
Chromatomyia nigra (Diptera: Agromyzidae) and Altica carduorum (Coleoptera: Chrysomelidae) 323
have been demonstrated to select plants for oviposition that maximize their own fitness rather 324
than that of their progeny, indicating that optimum foraging determines oviposition decisions 325
(SCHEIR et al. 2000; SCHEIRS and De BRUYN 2002). In E. postvittana, the oviposition preference is 326
essentially governed by plant-surface cues, since it does not lay eggs on rough and hairy surfaces 327
(TOMKINS et al. 1991; FOSTER and HOWARD 1999). Incidence of fungal mycelia on the leaf surfaces of 328
V. vinifera, mimicking hairiness precludes oviposition (RIZVI et al. 2014a, b). The lepidopteran 329
larvae are highly mobile and voracious feeders and, therefore, do not connect strongly with 330

10
JAENIKE’s (1978) hypothesis that explains mother’s choice of site for oviposition benefitting the 331
offspring (ROITBERG and MANGEL 1993; SUCKLING and BROCKERHOFF 2010). Under experimental 332
conditions, the larvae of E. postvittana have been established as highly active feeders on plants of 333
various taxa and gravid females lay eggs on those ranges of plants (FOSTER and HOWARD 1999). 334
Botrytis cinerea -infected leaves positively influence larval performance 335 of E. postvittana 336
337 Insect and fungus association ranges from non-specific polyphagy to obligate mutualism (JONSELL 338
and NORDLANDER 2004). In the present study, oviposition choices made by adults of E. postvittana 339
do not synchronize with the best performance of the larvae. This mismatch could be because 340
either the larvae and adults of E. postvittana have differing host requirements or these life forms 341
feed on different organs of different plants. The larvae of E. postvittana feed on leaves and fruits 342
of V. vinifera, M. domestica, and different species of Citrus; whereas their adults feed on floral 343
nectar. Our results suggest a mutualistic relationship between the larvae of E. postvittana and B. 344
cinerea. The larvae develop more rapidly, attain greater pupal mass (particularly in the females), 345
and lay more eggs when fed on B. cinerea-infected leaves. Larval diet plays an important role in 346
determining the size and reproductive performance in the Lepidoptera (MONDY and CORIO-347
COSTET 2000), where the adult females utilize the energy stored by the larvae for egg production 348
(BOGGS, 1997). Botrytis cinerea plays an essential role in the larval diet and the larvae of L. botrana 349
have been demonstrated to disperse the conidia of B. cinerea to adjacent berries of V. vinifera 350
(FERMAUD and LEMENN 1992). Further, the injury caused by the larvae of L. botrana while feeding 351
on V. vinifera tissues facilitates the colonization and establishment of B. cinerea (FERMAUD and 352
LEMENN 1992). CARDOZA et al. (2002) found that leaves of A. hypogaea infected with a necrotrophic 353
fungus S. rolfsii preferably fed by S. exigua and had a positive effect on key life-history traits 354
when fed on S. rolfsii-infected A. hypogaea. 355
Botrytis cinerea is an opportunistic necrotrophic fungus, which, under favourable 356
conditions can kill its hosts. The level and nature of infection by fungi can influence the 357
oviposition behaviour of insects (BIERE and TACK 2013). It is, therefore, possible that over time, the 358
effect of B. cinerea can alter the behaviour of E. postvittana. Nonetheless, the caveat here is that 359
the levels of fungal infection used in the present study have indeed been low inducing a sub-360
lethal infection on V. vinifera leaves. In such a context, the choice of gravid females in avoiding B. 361
cinerea-infected leaves of V. vinifera could be seen as an adaptive strategy because the difference 362
between the time of oviposition and that of egg hatch, which potentially enhances the infection 363
levels on V. vinifera leaves. Therefore, the larval-food quality could have deteriorated with 364

11
accelerated level of infection or the production of a defensive compound could have reached up 365
to the lethal level (TASIN 2012). This could explain why B. cinerea-infected leaves are unattractive 366
for the gravid females for oviposition. Plant-pathogenic fungal infection may influence on the egg 367
survival and its hatchability ratio in insects (VAN BAEL et al. 2009; BITTLESTON et al. 2011). Only 368
further studies can shed light on the survival rate of eggs E. postvittana and its hatchability ratio 369
at different levels of infection by B. cinerea on V. vinifera determining the role played by B. 370
cinerea. 371
372
The larvae of E. postvittana cannot survive and develop by feeding 373 only on B. cinerea 374 375 Lepidopteran larvae require high nutritional levels to match their rapid growth before pupation 376
(SLANSKY and SCRIBER 1985). For example, the host-plant quality has been shown to have a 377
significant, indirect but positive effect on pupation in Manduca sexta (Lepidoptera: Sphingidae) 378
through accelerated growth during its larval stages (DIAMOND and KINGSOLVER 2011). Physiological 379
changes in the larvae in preparation for pupation indicate that later instars switch from protein-380
based diets to lipid-based diets to accumulate more of membrane-bound energy (STOCKHOFF 1993; 381
OJEDA−AVILA et al. 2003). Insects are generally shown to lack the capacity to synthesize sterols that 382
are necessary as precursors to steroids which act as growth regulators. The insects, therefore, 383
acquire sterols from their host plants and/or from associated symbionts (NES et al. 1997; BEHMER 384
and NES 2003). Fungi are an excellent source of sterols and vitamins, which are vital in insect 385
development (MONDY and CORIO-COSTET 2000). Lipids, as principal sources of stored energy and as 386
precursors of steroids are key for insects particularly during metamorphosis (ARRESE and SOULAGES 387
2010). Nevertheless, high levels of lipids and low levels of other essential nutrients appear to 388
stress insects by affecting metabolism and inducing an inability to maintain homeostasis (LORD, 389
2010). One possible reason for the death of larvae reared on exclusive fungus diet in our study 390
could be that the larvae of E. postvittana did not get sufficient energy-containing nutrients, such 391
as carbohydrates and proteins from B. cinerea, although they may have had a rich supply of lipids 392
from B. cinerea. A second possibility is that the concentrated dose of lipidic materials may have 393
induced a dietary imbalance as shown in the larvae of M. sexta parasitized by Cotesia congregata 394
(Hymenoptera: Braconidae) (THOMPSON and RADAK 2010). 395
Conclusion 396 397
Gravid females of E. postvittana use olfactory and volatile cues to ‘evaluate’ infection levels on V. 398
vinifera caused by B. cinerea to oviposit. Larvae of E. postvittana show a mutualistic relationship 399

12
with B. cinerea. On B. cinerea–infected V. vinifera leaves, the larvae have a shorter larval duration, 400
attain heavier pupal mass and the emergent adults lay more eggs than the larvae fed on 401
uninfected V. vinifera leaves. The larvae reared on exclusive-fungus diet died in maximum 15 d 402
indicating that for a better larval performance and oviposition rate of E. postvittana, the V. 403
vinifera—B. cinerea interacting system is but imperative although further chemical-ecological 404
verifications are necessary for confirming this. 405
406 407
Acknowledgment 408 409 We thank Helen Nicol for advice on statistical analysis. 410 411 412 413
References 414 415
ABRAMOFF, M. D.; MAGALHAES, P. J.; RAM, S. J.; 2004. Image processing with ImageJ. Int. J. 416 Biophotonics 11, 36–42. 417
BAILEY, P. T.; FERGUSON, K. L.; MCMAHON, R.; WICKS, T. J.; 1997: Transmission of Botrytis cinerea by 418 light brown apple moth larvae on grapes. Aust. J. Grape Wine R. 3, 90–94. 419
BIERE, A.; TACK, A. J.; 2013: Evolutionary adaptation in three‐way interactions between plants, 420 microbes and arthropods. Funct. Ecol. 27, 646–660. 421
BITTLESTON, L. S.; BROCKMANN, F.; WCISLO, W.; VAN BAEL, S. A.; 2011: Endophytic fungi reduce leaf-422 cutting ant damage to seedlings. Biol. Lett 7, 30–32. 423
BOGGS, C. L.; 1997: Dynamics of reproductive allocation from juvenile and adult feeding: 424 radiotracer studies. Ecology 78, 192–202. 425
CARDOZA, Y. J.; LAIT C. G.; SCHMELZ, E. A.; HUANG, J.; TUMLINSON, J. H.; 2003: Fungus-induced 426 biochemical changes in peanut plants and their effect on development of beet armyworm, 427 Spodoptera exigua Hübner (Lepidoptera: Noctuidae) larvae. Environ. Entomol. 32, 220–228. 428
COSSÉ, A. A.; ENDRIS, J. J.; MILLAR, J. G.; BAKER, T. C.; 1994: Identification of volatile compounds from 429 fungus-infected date fruit that stimulate upwind flight in female Ectomyelois 430 ceratoniae. Entomol. Exp. Appl. 72, 233–238. 431
CUNNINGHAM, N.; 2007: Light brown apple moth (LBAM) culture: Epiphyas postvittana. South 432 Australian Research and Development Institute (S.A.R.D.I.). 433
DIAMOND, S. E.; Kingsolver, J. G.; 2011: Host plant quality, selection history and trade-offs shape 434 the immune responses of Manduca sexta. P. Roy. Soc. B-Biol. Sci. 278, 289–297. 435
DÖTTERL, S.; JURGENS, A.; WOLFE, L.; BIERE, A.; 2009: Disease status and population origin effects on 436 floral scent: potential consequences for oviposition and fruit predation in a complex 437 interaction between a plant, fungus, and noctuid moth. J. Chem. Ecol. 35, 307–319. 438
FERMAUD, M.; MENN, R. L.; Transmission of Botrytis cinerea to grapes by grape berry moth 439 larvae. Phytopathol. 82, 1393–1398. 440
FORNECK, A.; MERKT, N.; BLAICH, R.; 1998: A tripartite aseptic culture system for grape (Vitis species), 441 phylloxera (Daktulosphaira vitifoliae) and mites (Tarsanemus sp.). Vitis 37, 95−96. 442
FORNECK, A.; WALKER, M. A.; MERKT, N.; 1999: A review of aseptic dual culture of grape (Vitis species) 443 and grape phylloxera (Daktulosphaira vitifoliae Fitch; Homoptera: Phylloxeridae). Inter. 444 Jour. Ecol. Environ. Sci. 25, 221−227. 445
FOSTER, S. P.; HOWARD, A. J.; 1999: Adult female and neonate larval plant preferences of the 446 generalist herbivore, Epiphyas postvittana. Entomol. Exp. Appl. 92, 53–62. 447
FOSTER, S. P.; HOWARD, A. J.; HARRIS, M. O.; 1997: The influence of tactile and other non-chemical 448 factors on the ovipositional responses of the generalist herbivore Epiphyas postvittana. 449 Entomol. Exp. Appl. 83, 147–159. 450
FOURNIER, E.; GLADIEUX, P.; GIRAUD, T.; 2013: The ‘Dr Jekyll and Mr Hyde fungus’: noble rot versus 451 gray mold symptoms of Botrytis cinerea on grapes. Evol. Appl. 6, 960–969. 452
GRIPENBERG, S.; MAYHEW, P. J.; PARNELL, M.; ROSLIN, T.; 2010: A meta‐analysis of preference― 453 performance relationships in phytophagous insects. Ecol. Lett. 13, 383–393. 454
HATCHER, P. E.; 1995: Three-way interactions between plant pathogenic fungi, herbivorous insects 455 and their host plants. Biol Rev. 70, 639―694. 456

13
JAENIKE J.; 1978: On optimal oviposition behaviour in phytophagous insects. Theor. Popul. Biol. 14, 457 350–356. 458
JORDAN, M. D.; ANDERSON, A.; BEGUM, D.; CARRAHER, C.; AUTHIER, A.; MARSHALL, S. D. G.; KIELY, A.; 459 GATEHOUSE, L. N.; GREENWOOD, D. R.; CHRISTIE, D. L.; KRALICEK, A. V.; TROWELL, S. C.; NEWCOMB, R. 460 D.; 2009: Odorant receptors from the light brown apple moth (Epiphyas postvittana) 461 recognize important volatile compounds produced by plants. Chem. Senses 34, 383–394. 462
JONSELL, M.; NORDLANDER, G.; 2004. Host selection patterns in insects breeding in bracket fungi. 463 Ecol. Entomol. 29, 697–705. 464
LARSSON, S.; EKBOM, B.; 1995: Oviposition mistakes in herbivorous insects: confusion or a step 465 towards a new host plant? Oikos 72, 155–160. 466
LEYVA, K. J.; CLANCY, K. M.; PRICE, P. W.; 2000: Oviposition preference and larval performance of the 467 western spruce budworm (Lepidoptera: Tortricidae). Environ. Entomol. 29, 281–289. 468
LORDS, J. C.; 2010: Dietary Stress Increases the Susceptibility of Tribolium castaneum to Beauveria 469 bassiana. J. Econ. Entomol. 103, 1542–1546. 470
MAYHEW, P. J.; 1997: Adaptive patterns of host–plant selection by phytophagous insects. Oikos 79, 471 417–428. 472
MONDY, N.; CORIO-COSTET, M. F.; 2000: The response of the grape berry moth (Lobesia botrana) to a 473 dietary phytopathogenic fungus (Botrytis cinerea): the significance of fungus sterols. J. 474 Insect Physiol. 46, 1557–1564. 475
MURASHIGE, T.; SKOOG, F.; 1972: A revised medium for rapid growth and bioassay with tobacco tissue 476 culture. Physiol. Plantarum 15, 473−497. 477
NAJAR-RODRIGUEZ, A. J.; GALIZIA, C. G.; STIERLE, J.; DORN, S.; 2010: Behavioral and neurophysiological 478 responses of an insect to changing ratios of constituents in host plant-derived volatile 479 mixtures. J Exp Biol. 213, 3388―3397. 480
NES, W. D.; LOPEZ, M.; ZHOU, W.; GUO, D. A.; DOWD, P. F.; NORTON, R. A.; 1997: Sterol utilization and 481 metabolism by Heliothis zea. Lipids 32, 1317–1323. 482
OJEDA-AVILA, T.; WOODS, H. A.; RAGUSO, R. A.; 2003: Effects of dietary variation on growth, 483 composition, and maturation of Manduca sexta (Sphingidae: Lepidoptera). J. Insect Physiol. 484 49, 293–306. 485
QUINLAN, R. J.; CHERRETT, J. M.; 2008: The role of fungus in the diet of the leaf-cutting ant Atta 486 cephalotes (L.). Ecol. Entomol. 4, 151–160. 487
RAMAN, A.; BEIDERBECK, R.; 1992: Aseptic dual culture of the greenhouse whitefly, Trialeurodes 488 vaporariorum Westwood (Hom., Aleyrodidae) and its host, Stellaria media (L.) Vill. 489 (Caryophyllaceae). J. Appl. Entomol. 113, 252–257. 490
REAVEY, D.; 1993: Why body size matters to caterpillars. In: Caterpillars: Ecological and 491 Evolutionary Constraints on Foraging (Eds. Stamp, N. E.; Casey, T. M.;). Chapman and Hall, 492 New York, pp. 248–279. 493
RENWICK, J. A. A.; 1989: Chemical ecology of oviposition in phytophagous insects. Experientia 45, 494 223–228. 495
RENWICK, J. A. A.; CHEW, F. S.; 1994: Oviposition in Lepidoptera. Annu. Rev. Entomol. 39, 377–400. 496 RIZVI, S. Z. M.; RAMAN, A.; WHEATLEY, W.; COOK, G., NICOL, H.; 2014a: Influence of Botrytis cinerea 497
(Helotiales: Sclerotiniaceae) infected leaves of Vitis vinifera (Vitales: Vitaceae) on the 498 preference of Epiphyas postvittana (Lepidoptera: Tortricidae). Austral Entomol. doi: 10. 499 1111/aen.12093. 500
RIZVI, S. Z. M.; RAMAN, A.; WHEATLEY, W.; COOK, G.; 2014b: Oviposition preference and larval 501 performance of Epiphyas postvittana (Lepidoptera: Tortricidae) on Botrytis cinerea 502 (Helotiales: Sclerotiniaceae) infected berries of Vitis vinifera (Vitales: Vitaceae). Insect Sci. 503 In press. 504
RYCHTER, A. M.; MIKULSKA, M.; 1990: The relationship between phosphate status and cyanide-505 resistant respiration in bean roots. Physiol. Plantarum 79, 663–667. 506
SCHEIRS, J.; DE BRUYN, L.; 2002: Integrating optimal foraging and optimal oviposition theory in 507 plant—insect research. Oikos 96, 187–191. 508
SCHEIRS, J.; ZOEBISCH, T. G.; SCHUSTER, D. J.; BRUYN, L. D.; 2004: Optimal foraging shapes host 509 preference of a polyphagous leafminer. Ecol. Entomol. 29, 375–379. 510
Slansky, F. Jr.; Scriber, J. M.; 1985: Food consumption and utilization. In: Comprehensive Insect 511 Physiology, Biochemistry and Pharmacology (Eds. KERKUT, G. A.; GILBERT L. I.;). Pergamon 512 Press, Oxford. Vol. 4, pp. 87-163. 513
STOCKHOFF, B. A.; 1993: Ontogenetic change in dietary selection for protein and lipid by gypsy 514 moth larvae. J. Insect Physiol. 39, 677–686. 515
SUCKLING, D. M.; BROCKERHOFF, E. G.; 2010: Invasion biology, ecology, and management of the light 516 brown apple moth (Tortricidae). Annu. Rev. Entomol. 55, 285–306. 517
TASIN, M.; BETTA, E.; CARLIN, S.; GASPERI, F.; MATTIVI, F.; PERTOT, I.; 2011: Volatiles that encode host-518 plant quality in the grapevine moth. Phytochem. 72, 1999–2005. 519
TASIN, M.; KNUDSEN, G. K.; PERTOT, I.; 2012. Smelling a diseased host: grapevine moth responses to 520 healthy and fungus-infected grapes. Anim Behav. 83, 555–562. 521

14
THALER, J. S.; FIDANTSEF, A. L.; DUFFEY, S. S.; BOSTOCK. R. M.; 1999. Tradeoffs in plant defense against 522 pathogens and herbivores? J. Chem. Ecol. 25, 1597–1609. 523
THOMPSON, S. N.; REDAK, R. A.; 2008: Parasitism of an insect Manduca sexta L. alters feeding 524 behaviour and nutrient utilization to influence developmental success of a parasitoid. J. 525 Comp. Physiol., B, 178, 515–527. 526
TOMKINS, A. R.; PENMAN, D. R.; CHAPMAN, R. B.; 1991: Leafroller oviposition on larval host plants. New 527 Zeal. Entomol. 14, 37–41. 528
VAN BAEL, S. A.; VALENCIA, M. C.; ROJAS, E. I.; GOMEZ, N.; WINDSOR, D. M.; HERRE, E. A.; 2009: Effects of 529 foliar endophytic fungi on the preference and performance of the leaf beetle Chelymorpha 530 alternans in Panama. Biotropica 41, 221–225. 531

15
532 533 534 535 536 537 538 539 540 541 542 543 544 545 546 547 548 549 550 551 552 553 554 555 Rizvi and Raman 556 557 Figure 1. Custom-made aluminium cage made for the free-choice experiment of adults of 558 Epiphyas postvittana (not to scale) (am: aluminium mesh, i: infected plant, u: uninfected 559 plant). 560 561 562 563 564 565 566 567 568 569
570
571
572
573
574
575
576
577
578
579
580
581
582
am

16
583
584
585
586
587
588
589
590
591
592
Rizvi and Raman 593 594 Figure 2. Custom-made glass device for the two-choice experiment using adults of 595 Epiphyas postvittana (not to scale) (cp: circular port, gj: glass jar, hgt: horizontal glass 596 tube, ip: infected plant, m: moth, p: Parafilm, up: uninfected plant) 597 598 599
gj m
m
cp
m
hgt
i u

17
600
601
602
603
604
605
606
607
608
609
Rizvi and Raman 610
611 Figure 3. Petri-dish pair for the development of the larvae of E. postvittana on B. cinerea. 612 (not to scale) (l: larva, pd: Petri dish, l: Petri dish lid) 613 614
615
616
pd
l

18
617
618
619
620
621
622
623
624
625
626
627
Rizvi and Raman 628 Figure 4. Custom-made glass device for the rearing of the larvae of Epiphyas postvittana 629 (not to scale) (ff: floral foam, il: infected leaf, l: larva, ps: parafilm seal, ul: uninfected 630 leaf) 631

19
632
633
634
635
636
637
638
639
640
641
642
643
644
Rizvi and Raman 645
Figure 5. Custom-made glass device for the rearing of the larvae of Epiphyas postvittana 646 (not to scale) (l: larva, gt: glass tube, il: infected leaf, ms: medium slant, ps: Parafilm seal, 647 ul: uninfected leaf) 648
20
649
650
651
652
653
654
655
656
657
658
659
660
661
662
663
664
665
Rizvi and Raman 666 667
Figure 6. Mean number of eggs laid by E. postvittana in a ‘free-choice’ experiment with 668 uninfected (control) leaves ( ) and B. cinerea-infected leaves ( ) of V. vinifera. 669

21
670
671
672
673
674
675
676
677
678
679
680
681
682
683
684 Rizvi and Raman 685 686 Figure 7. Response of females E. postvittana in a ‘two-choice’ experiment towards 687 uninfected (control) leaves ( ) and B. cinerea-infected leaves ( ) of V. vinifera. 688

22
689
690
691
692
693
694
695
696
697
698
699
700
701
702
703
704
Rizvi and Raman 705 706 Figure 8. Mean number of eggs laid by E. postvittana in a ‘two-choice’ experiment with 707 uninfected (control) leaves ( ) and B. cinerea-infected leaves ( ) of V. vinifera. 708
23
709
710
711
712
713
714
715
716
717
718
719
720
721
722
723
724
725
Rizvi and Raman 726 727 Figure 9. Mortality rate of E. postvittana larvae reared solely on B. cinerea ( ) 728 and control (or PDA, ). 729

24
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744 745
Rizvi and Raman 746 747 Figure 10. Mortality rate of larvae reared on B. cinerea ( ) and control (or M−S 748 medium, ). 749 750
25
751
752
753
754
755
756
757
758
759
760
761
762
763
764
765
766
Rizvi and Raman 767 768 Figure 11. Mortality rate of larvae reared on B. cinerea-leaves ( ) or uninfected 769 ( ) leaves of V. vinifera using Knop’s solution 770
26
771
772
773
774
Rizvi and Raman 775
776
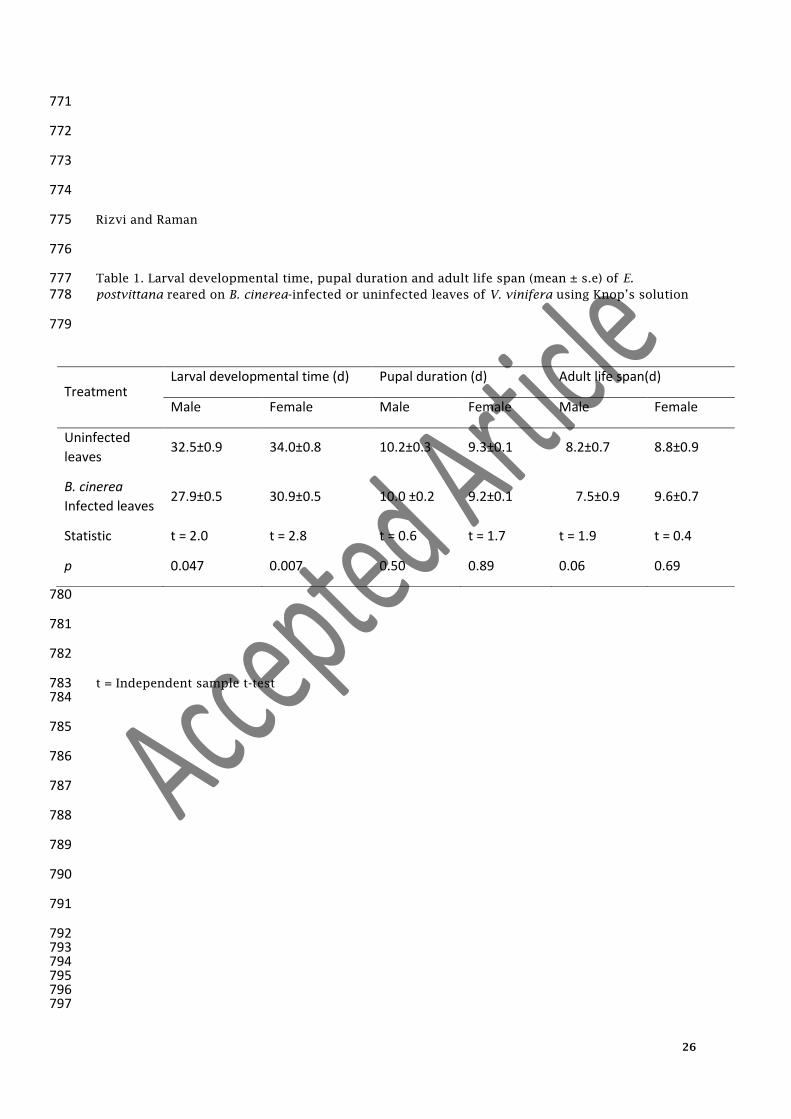
Table 1. Larval developmental time, pupal duration and adult life span (mean ± s.e) of E. 777 postvittana reared on B. cinerea-infected or uninfected leaves of V. vinifera using Knop’s solution 778
779
780
781
782
t = Independent sample t-test 783 784
785
786
787
788
789
790
791
792 793 794 795 796 797
Treatment Larval developmental time (d) Pupal duration (d) Adult life span(d)
Male Female Male Female Male Female
Uninfected
leaves 32.5±0.9 34.0±0.8 10.2±0.3 9.3±0.1 8.2±0.7 8.8±0.9
B. cinerea
Infected leaves 27.9±0.5 30.9±0.5 10.0 ±0.2 9.2±0.1 7.5±0.9 9.6±0.7
Statistic t = 2.0 t = 2.8 t = 0.6 t = 1.7 t = 1.9 t = 0.4
p 0.047 0.007 0.50 0.89 0.06 0.69
27
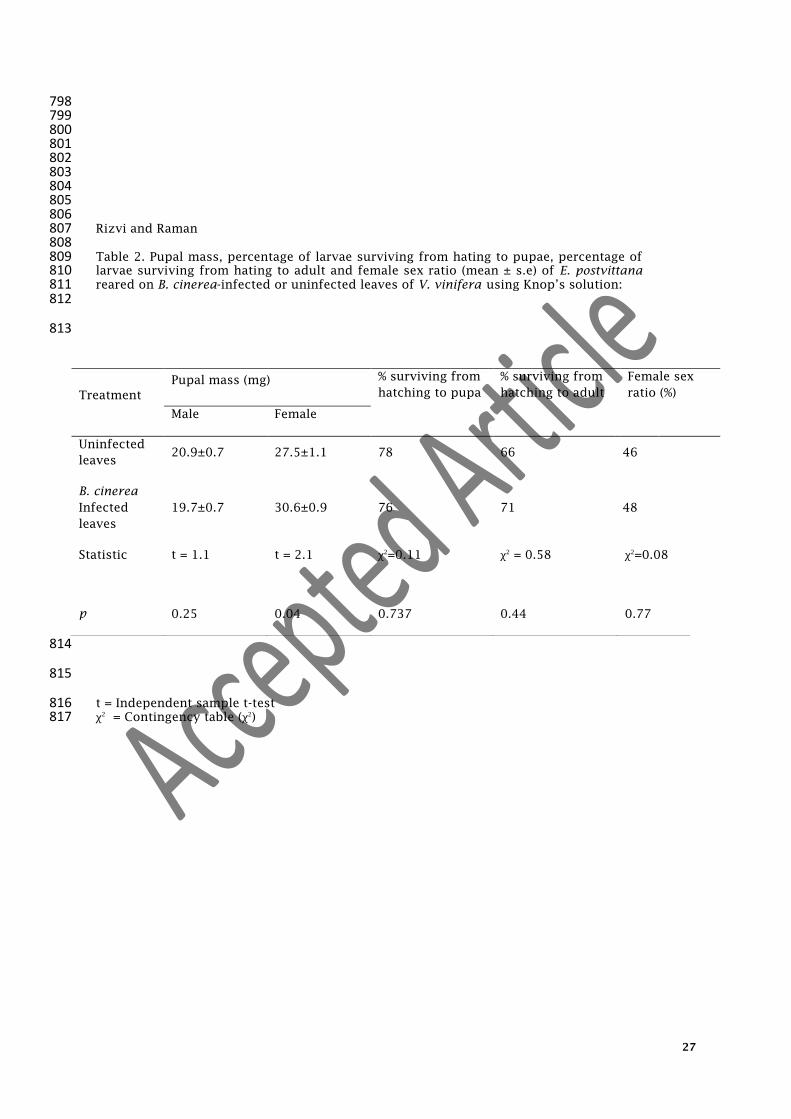
798 799 800 801 802 803 804 805 806 Rizvi and Raman 807 808 Table 2. Pupal mass, percentage of larvae surviving from hating to pupae, percentage of 809 larvae surviving from hating to adult and female sex ratio (mean ± s.e) of E. postvittana 810 reared on B. cinerea-infected or uninfected leaves of V. vinifera using Knop’s solution: 811 812
813
814
815
t = Independent sample t-test 816 χ2
= Contingency table (χ2
) 817
Treatment
Pupal mass (mg) % surviving from
hatching to pupa
% surviving from
hatching to adult
Female sex
ratio (%)
Male Female
Uninfected
leaves 20.9±0.7 27.5±1.1 78 66 46
B. cinerea
Infected
leaves
19.7±0.7 30.6±0.9 76 71 48
Statistic
t = 1.1 t = 2.1 χ2
=0.11 χ2
= 0.58 χ2
=0.08
p 0.25 0.04 0.737 0.44 0.77

28
818 819 820 821 822 823 824 825 826 827 828 829 830 831 Rizvi and Raman 832 833 834 Table 3. Adult performance (mean ± s.e) of E. postvittana reared on B. cinerea-infected 835 and uninfected leaves of V. vinifera using Knop’s solution: 836 837
838 839
t = Independent sample t-test 840 χ2
= Contingency table (χ2
) 841 842
Treatment
Delay in egg laying
(days)
Fecundity
(no. eggs per female)
No. of larvae emerged
per female
Uninfected
leaves 2.3±0.1 80.0±10.9 67.2±5.8
B. cinerea
Infected leaves 2.1±0.1 115.1±13.2 106.7±12.3
Statistic
t = 0.98 t = 2.5 t = 2.98
p 0.86 0.01 0.006


![University of Mumbai [1] RIZVI EDUCATION SOCIETY'S RIZVI .... Sem IV- Regular.… · PROGRAMME: BACHELOR OF COMMERCE- ACCOUNTING & FINANCE (B.COM.- A. & F.) SEMESTER- IV [4] RIZVI](https://img.pdfslide.us/doc/110x75/600174755bcc726418541603/university-of-mumbai-1-rizvi-education-societys-rizvi-sem-iv-regular.jpg)


























