Embed Size (px)
Citation preview

Structure, dynamics and function ofnuclear pore complexesMaximiliano A. D’Angelo and Martin W. Hetzer
Molecular and Cell Biology Laboratory, Salk Institute for Biological Studies, La Jolla, CA 92037, USA
Review
Nuclear pore complexes are large aqueous channels thatpenetrate the nuclear envelope, thereby connecting thenuclear interior with the cytoplasm. Until recently, thesemacromolecular complexes were viewed as static struc-tures, the only function of which was to control themolecular trafficking between the two compartments.It has now become evident that this simplistic scenario isinaccurate and that nuclear pore complexes are highlydynamic multiprotein assemblies involved in diversecellular processes ranging from the organization of thecytoskeleton to gene expression. In this review, wediscuss the most recent developments in the nuclear-pore-complex field, focusing on the assembly, disassem-bly, maintenance and function of this macromolecularstructure.
IntroductionA hallmark of eukaryotic cells is the compartmentalizationof the genetic material inside the nucleus. By restrictingthe accessibility of cytoplasmic proteins to DNA with thephysical barrier of the nuclear envelope (NE), eukaryoticcells have achieved a complexity in transcriptional regu-lation not found in prokaryotes. Furthermore, the NEprovides additional levels of regulation of gene expression,such as the selective export of newly synthesized mRNAinto the ribosome-containing cytoplasm and the establish-ment of higher-order levels of organization of the nucleargenome.
The NE comprises two concentric lipid bilayers, theouter and inner nuclear membranes (ONM and INM,respectively) [1]. The ONM is continuous with the endo-plasmic reticulum (ER) and studded with ribosomes,whereas the INM is characterized by a set of integralmembrane proteins [1]. Large multiprotein structuresknown as nuclear pore complexes (NPCs) penetrate theNE at sites at which the INM and the ONM are fused [1].
NPCs function as gatekeepers of the nucleus, perform-ing the essential cellular role of mediating the exchange ofmolecules between the nucleoplasm and the cytoplasm [2].Ions and small metabolites can diffuse through NPCs;however, molecules possessing a mass >40 kDa need tobe actively transported. Nucleocytoplasmic transport is acomplicated process that is carried out by a large family oftransport receptors known as karyopherins in yeastand importins and exportins in mammals, the latternamed depending on their direction of transport [3]. Themolecular mechanism of nuclear transport has recentlybeen the subject of several major reviews and, thus, is not
Corresponding author: Hetzer, M.W. ([email protected]).
456 0962-8924/$ – see front matter � 2008 Elsevier
discussed [2,4]. Here, we focus on the latest advances in ourunderstanding of nuclear pore assembly, disassembly,maintenance and function.
NPC structureOwing to their function as exclusive nucleocytoplasmictransport channels and unique structural features, NPCshave been actively studied since their discovery in the1950s. Although the initial descriptions of the overallNPC structure were performed >40 years ago [5], it wasnot until recently that new techniques, such as cryo-elec-tron tomography, field-emission in-lens scanning electronmicroscopy (FEISEM), 4pi and atomic force microscopyand improved cell-fixation protocols could provide adetailed picture of its three-dimensional organization[6–11]. Overall, the NPC is an eightfold-symmetrical struc-ture comprising a NE-embedded scaffold that surrounds acentral transport channel and two rings – the cytoplasmicand nuclear rings – to which eight filaments are attached(Figure 1a). Whereas the cytoplasmic filaments have looseends, the nuclear filaments are joined in a distal ring,forming a structure known as the nuclear basket. Eventhough the size of the NPC varies between species, itsoverall structure is evolutionarily conserved from yeasts tomammals [12,13].
Despite their high molecular mass of �60–125 MDa inmammals and �40–60 MDa in yeasts, proteomic analyseshave revealed that NPCs contain only �30 differentproteins, known as nucleoporins or nups [13–16](Table 1). Except for three transmembrane proteins thatare believed to anchor the NPC to the NE [17,18], all othernucleoporins are soluble. Owing to the eightfold symmetryof pores, each nucleoporin is present in copies of eight ormultiples of eight, resulting in �500–1000 nups per pore.Remarkably, nucleoporins have a very limited set ofdomains, restricted to b-propellers, a-solenoids, phenyl-alanine-glycine (FG) repeats, coiled–coiled and transmem-brane domains [19,20]. Most of these proteins associate inbiochemically stable subcomplexes that are believed tofunction as the ‘building blocks’ of the NPC (Figure 1b).
Recently, a detailed 3D model for the position andabundance of each nup in the Saccharomyces cerevisiaeNPC structure was proposed based on experimental dataobtained frommolecular, biochemical and structural infor-mation of the NPCs and their components [20,21]. In themodeled structure, the scaffold of theNPC is formed by twomain protein subcomplexes that, through linker proteins,anchor a set of FG-containing nucleoporins [20]. TheFG-rich nucleoporins, which can contain 4–48 GLFG,
Ltd. All rights reserved. doi:10.1016/j.tcb.2008.07.009 Available online 9 September 2008
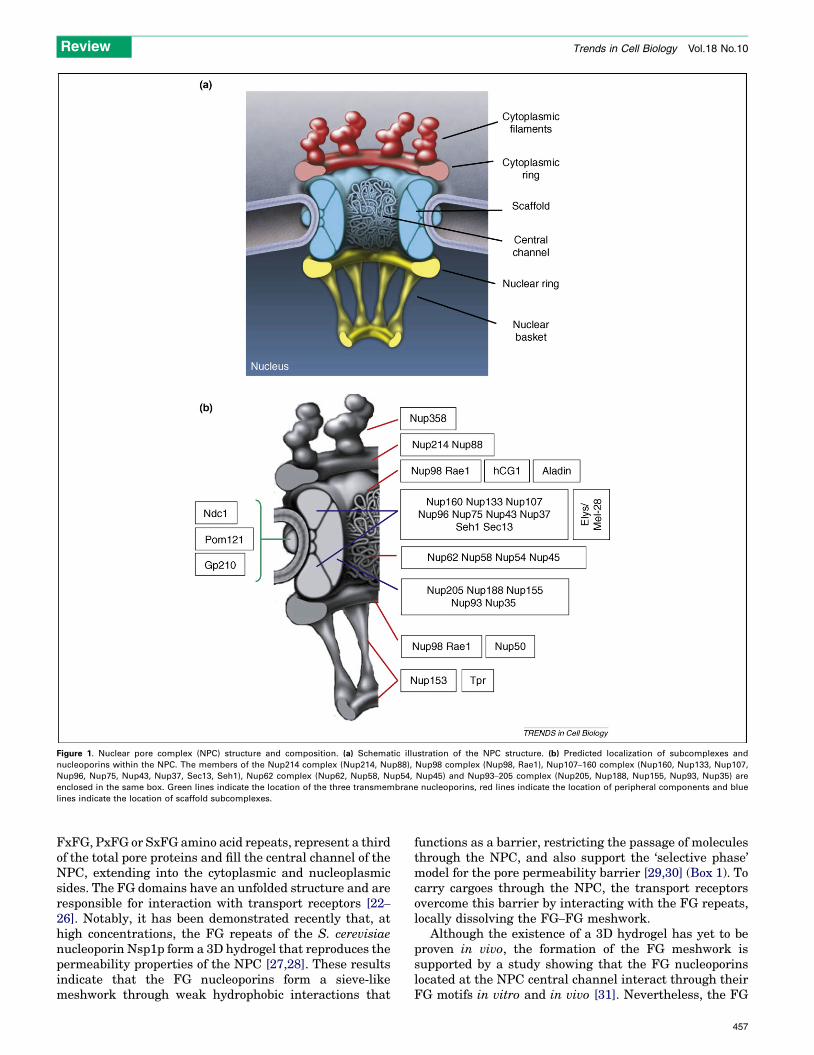
Figure 1. Nuclear pore complex (NPC) structure and composition. (a) Schematic illustration of the NPC structure. (b) Predicted localization of subcomplexes and
nucleoporins within the NPC. The members of the Nup214 complex (Nup214, Nup88), Nup98 complex (Nup98, Rae1), Nup107–160 complex (Nup160, Nup133, Nup107,
Nup96, Nup75, Nup43, Nup37, Sec13, Seh1), Nup62 complex (Nup62, Nup58, Nup54, Nup45) and Nup93–205 complex (Nup205, Nup188, Nup155, Nup93, Nup35) are
enclosed in the same box. Green lines indicate the location of the three transmembrane nucleoporins, red lines indicate the location of peripheral components and blue
lines indicate the location of scaffold subcomplexes.
Review Trends in Cell Biology Vol.18 No.10
FxFG, PxFG or SxFG amino acid repeats, represent a thirdof the total pore proteins and fill the central channel of theNPC, extending into the cytoplasmic and nucleoplasmicsides. The FG domains have an unfolded structure and areresponsible for interaction with transport receptors [22–
26]. Notably, it has been demonstrated recently that, athigh concentrations, the FG repeats of the S. cerevisiaenucleoporin Nsp1p form a 3D hydrogel that reproduces thepermeability properties of the NPC [27,28]. These resultsindicate that the FG nucleoporins form a sieve-likemeshwork through weak hydrophobic interactions that
functions as a barrier, restricting the passage of moleculesthrough the NPC, and also support the ‘selective phase’model for the pore permeability barrier [29,30] (Box 1). Tocarry cargoes through the NPC, the transport receptorsovercome this barrier by interacting with the FG repeats,locally dissolving the FG–FG meshwork.
Although the existence of a 3D hydrogel has yet to beproven in vivo, the formation of the FG meshwork issupported by a study showing that the FG nucleoporinslocated at the NPC central channel interact through theirFG motifs in vitro and in vivo [31]. Nevertheless, the FG
457

Table 1. Mammalian, S. cerevisiae and C. elegans nucleoporinshomologsa,b
Mammalian S. cerevisiae C. elegans
Nup35 Nup53p Npp-19
Nup37 – –
Nup43 – C09G9.2
Nup50 Nup2p Npp-16
Nup54 Nup57p Npp-1
Nup58/45 Nup49p Npp-4
Nup62 Nsp1p Npp-11
Nup75 Nup85p Npp-2
Nup88 Nup82p –
Nup93 Nic96p Npp-13
Nup96 Nup145Cp Npp-10
Nup98 Nup145Np Nup100p Nup116p Npp-10
Nup107 Nup84p Npp-5
Nup133 Nup133p Npp-15
Nup153 Nup1p Nup2p Nup60p Npp-7
Nup155 Nup157p Nup170p Npp-8
Nup160 Nup120p Npp-6
Nup188 Nup188p –
Nup205 Nup192p Npp-3
Nup214 Nup159p Npp-14
Nup358 (RanBP2) – Npp-9
Sec13R Sec13p Npp-20
Seh1 Sehp Npp-18
Pom121 – –
Gp210 – Npp-12
Ndc1 Ndc1p Npp-22
Tpr Mlp1p Mlp2p Npp-21
RAE1 Gle2p Npp-17
ALADIN – –
NLP1(hCG1) Nup42p –
– Nup59p –
– Nup116p –
– Nup100p –
– Pom134p –
– Pom152p –aNote that some nucleoporins, such as Nup98, Nup153, Nup155, Tpr and Nup2p,
share homology with more than one nucleoporin.bA dash (–) indicates that no homologs have yet been described in the organism.
Box 1. Models of NPC selectivity
NPCs can perform 1000 translocations events per second, shuttling
a mass of �100 MDa second�1 [29]. However, these structures
efficiently restrict the passages of large molecules that lack
nuclear-transport signals. Although it has long been known that
the major diffusion barrier of the NPC resides in its central channel,
how nuclear pores achieve their transport selectivity is still a matter
of debate. Several models have been proposed to explain the NPC
selectivity, which take into account that active transport through the
pore occurs by facilitated diffusion and requires the interaction of
transport receptors with the FG nucleoporins.
The ‘virtual gate model’ [33] predicts that the highly dense FG
nucleoporins of the central channel generate an entropic barrier that
prevents the passage of inert molecules. For a molecule that is freely
diffusing in the cytoplasm, entering the overcrowded central
channel of the pore implies an immediate restriction of its liberty
of movement and a drop of its entropy. The entropic price that a
molecule has to pay to move through the pore increases with its
size, thereby lowering its probability of crossing the channel. The
interaction of a molecule with the FG nucleoporins increases the
probability of it entering and moving through the pore. This model
proposes the existence of an energetic barrier and not a physical
barrier controlling NPC permeability.
The ‘spaghetti oil model’ [25] proposes that extended FG repeats
fill and obstruct the pore. The idea is that the FG repeats are
constantly moving at physiological temperature and can be pushed
to one side by translocation complexes. In this model, transport
receptors move through the FG spaghetti by a binding and release
mechanism. Because the FG-transport receptor interactions are
weak, the carriers would bind the FG repeats only transiently and,
once released, they would be free to diffuse through the pore until
they find another nucleoporin to bind. The continuous binding–
release–diffusion cycle would enable the transporter to move
through the pore by a facilitated diffusion.
In contrast to the previous models, the ‘selective phase model’
[29] indicates that the FG nucleoporins from the central channel
form a sieve-like meshwork through weak hydrophobic interactions.
This gel-like network prevents the passage of inert molecules larger
than its pore size. In this model, the transport receptors would
selectively partition into the semi-liquid-pore phase and move
through the central channel by competing with the FG–FG interac-
tions locally dissolving the meshwork. Thus, by interacting with the
FG nucleoporins, the carriers would help molecules carrying
nuclear-transport signals cross the pore by increasing their
solubility in the central channel. This model proposes the existence
of a physical barrier that determines NPC selectivity, whereby the
exclusion size of the pore is defined by the pore size of the sieve-like
meshwork.
Finally, the ‘reduction of dimensionality model’ [26] proposes that
transport receptors function as ferries that can slide on the surface
of an FG surface. In this model, a continuous FG surface extends
from the cytoplasmic filaments to the nucleoplasmic basket and
throughout the walls of the central channel. Transport receptors
bind to FG surface in the cytoplasmic or nucleoplasmic side of the
NPC and move through the pore by a two-dimensional walk. The
model predicts the existence of a selectivity filter in the central
channel, which is also generated by the FG repeats, with an
unobstructed narrow tube in the center that would enable the
diffusion of small molecules. The transport-receptor-cargo com-
plexes would overcome the filter because they can bind and enter
the FG surface through its extremes. However, inert molecules that
are too large to diffuse through the central tube and cannot bind the
FG repeats would not be able to enter the filter and move across
the pore.
Review Trends in Cell Biology Vol.18 No.10
repeats of the nucleoporins that constitute the cytoplasmicand nuclear filaments have not been found to interact withthe FG domains of the nucleoporins from the centralchannel, thereby indicating that the peripheral FG nucleo-porins could function as ‘entropic bristles’ preventing theentry of molecules to the pore. Functioning as ‘entropicbristles’ means that, by movement, the filaments reducethe available space to access the pore channel, thus, gen-erating in this way an entropic barrier that decreases theprobability of the molecules entering the pore withoutphysically repelling them. To access this limited spaceand enter the pore, a molecule would have to reduce itsentropy, which is energetically unfavorable. In support ofthis idea, it has recently been reported that the FGdomains of Nup153, a main component of the nuclearbasket, form a brush-like structure in vitro with entro-pic-repulsion properties [32]. According to this, the fila-ments would help to maintain the permeability barrier, inagreement with the ‘virtual gate’ model [33] (Box 1), whichproposes that the probability of a molecule to overcome theentropic cost of entering a limited space packed with non-interacting nucleoporins decreases with increasing size(Box 1). The interaction of transport receptors with theFG repeats enables largemolecules to overcome the energyloss and move through the entropic barrier.
458
It is worth mentioning that, in the selective-phasemodel, the formation of the sieve-like meshwork wouldalso generate an entropic barrier for those molecules thatcan enter the pores of the FG network. This barrier wouldreduce the probability of the molecules entering the pores

Review Trends in Cell Biology Vol.18 No.10
but would not define the permeability limit of the NPC,which would be determined by the meshwork pore size.
Although these findings point towards the FG networkas the main mechanism for pore selectivity, they are notyet sufficient to completely discard other proposed modelsfor NPC permeability [25,26,29,33] (Box 1). It is possiblethat a combination of these models would be required toexplain all the reported NPC properties.
Dynamic organization of NPCs
A remarkable, yet largely uncharacterized, feature of theNPCs is their dynamic molecular organization. Using asystematic approach, in which 19 GFP-tagged nucleopor-ins were studied by fluorescence recovery after photo-bleaching (FRAP), it was shown that the residence timesof different nups at the NPC varied from a few seconds to>70 h depending on their location or function [34].Although the proteins that form the NPC scaffold, suchas members of the Nup107–160 complex, are stablyembedded in the NE during interphase (having residencetimes longer than the average cell cycle), the more periph-eral components, such as Nup153 and Nup50, are highlydynamic, having residence times of seconds to minutes. Athird class of nucleoporins, which are believed to work aslinkers between the scaffold and the peripheral nucleopor-ins, have intermediate residence times.
The functional importance of ‘dynamic nucleoporins’ isstill unclear. It has been suggested that mobile nucleopor-ins could help to deliver cargo to the NPC. In support ofthis, the mobility of two RNA-binding nucleoporins,Nup153 and Nup98, is transcription dependent, indicatingthat they could assist newly transcribed RNAs to reach thepore and be exported to the cytoplasm [35]. Although atransport-dependent function of dynamic nups seemsplausible, it is also possible that these proteins havepore-independent functions (see later). A dynamic organ-ization of NPCs could also be indicative of changes inprotein composition in response to altered transportrequirements. The presence of tissue and developmen-tal-specific nucleoporins has been reported [36–39]; how-ever, there is no evidence that NPCs of differentcomposition coexist in the same cell.
The dynamic properties of NPCs can also be observed atthe structural level. Early cryo-electron microscopy (EM)studies found that theNPCswere present in either an openor closed conformation, the latter characterized by thepresence of a central plug or transporter [40]. Historically,the central plug was considered to be a structural com-ponent of the NPC, but recent data have indicated that itrepresents translocating cargo. Transmission electronmicroscopy (TEM) and FEISEM have identified differentconformations of theNPCnuclear basket in response to theexport of large ribonucleoprotein particles [41]. Cryo-elec-tron tomography has shown two different structural statesof the cytoplasmic filaments and variability in the positionof the narrowest constriction of the central channel [7].Furthermore, the NPC has been shown to alter itsconformation in response to Ca2+, glucocorticoids andATP [42–47].
The molecular basis of the structural changes in porearchitecture and its physiological role is still a mystery.
One possibility is that the location of nucleoporins isactively reorganized for functional purposes within theNPC structure. Consistent with this, two nucleoporins,Nup153 and Nup214, have been found in different regionsof the NPC in a transport-dependent manner [48,49].Moreover, the yeast NPC has been shown to undergocell-cycle-specific rearrangements [50]. It has beenobserved that, by changing interacting partners, somenucleoporinsmodify their location at the yeast NPC, affect-ing specific nuclear-transport pathways [50]. Anotherpossibility is that structural changes of the NPC are aconsequence of alterations in nucleoporin conformationand not in their location. Recently, the crystal structureof a-helical regions of Nup58/45, a nucleoporin from thepore central channel, was solved. It was found that the a-helical domains formed tetramers through the interactionof two antiparallel dimers [51]. The presence of differenttypes of tetramer conformations that were laterally dis-placed along the axis of the dimer–dimer interface wasfound. Thus, it has been proposed that eight Nup58/45tetramers would be built around the central channel andwould regulate the aperture or diameter of the NPC bysliding along the dimer interface. Also supporting a nucleo-porin conformational change, Nup153 has been reported tocollapse reversibly into a compact conformation when itsFG repeats bind to the transport receptor karyopherin b
(the yeast homolog of importin b) [52].
NPC life cycleAssembly
The biogenesis of nuclear pores is essential for cell survivaland proliferation. There are two phases during themetazoan cell cycle in which NPCs need to be assembled:first, at the end ofmitosis, when theNE reforms around thesegregated chromosomes and second, during interphase inwhich the cells double their number of pores in preparationfor the next round of division. Even though both processeslead to the same final structure, they occur under verydifferent conditions. Mitotic-NPC assembly takes placeconcomitantly with reformation of the nuclear membranearound segregated chromosomes. At this time, NPCs arerebuilt from disassembled subcomplexes that were dis-persed into the cytoplasm during breakdown of the NE(Figure 2). By contrast, NPC assembly during interphaseoccurs in an intactNEusing newly synthesized nups and ina cellular environment in which the nucleus and thecytoplasm are physically separated (Figure 2). Interphaseassembly is the only existing mechanism in organisms,such as yeasts, that undergo closed mitosis (i.e. their NEdoes not break down during cell division). Althoughaccumulating evidence indicates that both types of poreassembly might occur through similar processes, owingto their very different environments it is likely that differ-ences will be uncovered.
Mitotic assembly
As mentioned, NPC assembly during mitosis occurs whenthe NEs form around the chromatin of the two nascentdaughters [53,54]. Data obtained from mammalian cellsand an in vitro nuclear-assembly system based on Xenopusegg extracts have shown that mitotic-NPC assembly is a
459
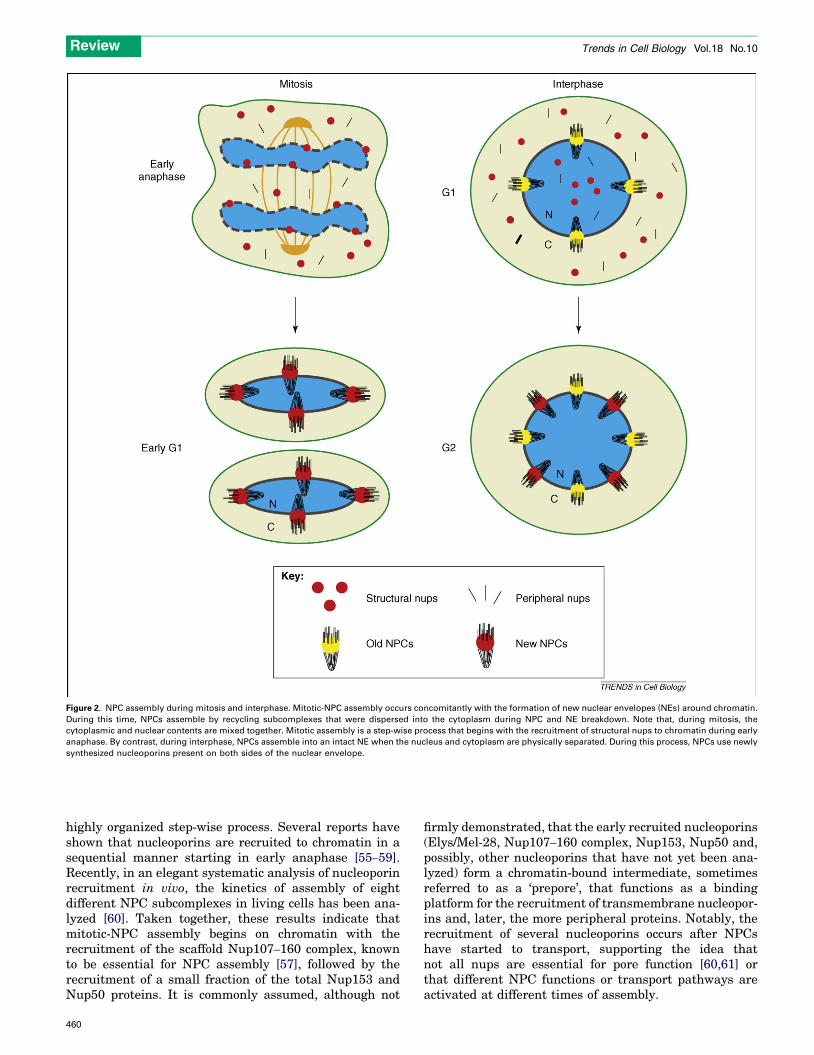
Figure 2. NPC assembly during mitosis and interphase. Mitotic-NPC assembly occurs concomitantly with the formation of new nuclear envelopes (NEs) around chromatin.
During this time, NPCs assemble by recycling subcomplexes that were dispersed into the cytoplasm during NPC and NE breakdown. Note that, during mitosis, the
cytoplasmic and nuclear contents are mixed together. Mitotic assembly is a step-wise process that begins with the recruitment of structural nups to chromatin during early
anaphase. By contrast, during interphase, NPCs assemble into an intact NE when the nucleus and cytoplasm are physically separated. During this process, NPCs use newly
synthesized nucleoporins present on both sides of the nuclear envelope.
Review Trends in Cell Biology Vol.18 No.10
highly organized step-wise process. Several reports haveshown that nucleoporins are recruited to chromatin in asequential manner starting in early anaphase [55–59].Recently, in an elegant systematic analysis of nucleoporinrecruitment in vivo, the kinetics of assembly of eightdifferent NPC subcomplexes in living cells has been ana-lyzed [60]. Taken together, these results indicate thatmitotic-NPC assembly begins on chromatin with therecruitment of the scaffold Nup107–160 complex, knownto be essential for NPC assembly [57], followed by therecruitment of a small fraction of the total Nup153 andNup50 proteins. It is commonly assumed, although not
460
firmly demonstrated, that the early recruited nucleoporins(Elys/Mel-28, Nup107–160 complex, Nup153, Nup50 and,possibly, other nucleoporins that have not yet been ana-lyzed) form a chromatin-bound intermediate, sometimesreferred to as a ‘prepore’, that functions as a bindingplatform for the recruitment of transmembrane nucleopor-ins and, later, the more peripheral proteins. Notably, therecruitment of several nucleoporins occurs after NPCshave started to transport, supporting the idea thatnot all nups are essential for pore function [60,61] orthat different NPC functions or transport pathways areactivated at different times of assembly.
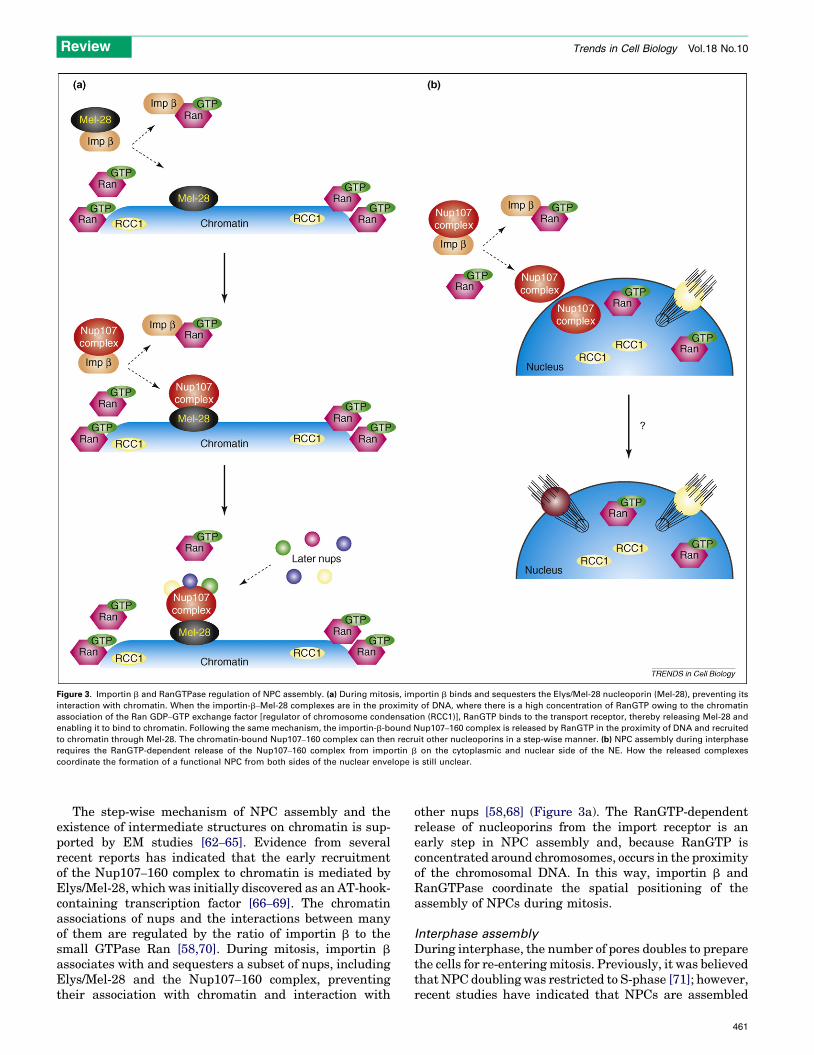
Figure 3. Importin b and RanGTPase regulation of NPC assembly. (a) During mitosis, importin b binds and sequesters the Elys/Mel-28 nucleoporin (Mel-28), preventing its
interaction with chromatin. When the importin-b–Mel-28 complexes are in the proximity of DNA, where there is a high concentration of RanGTP owing to the chromatin
association of the Ran GDP–GTP exchange factor [regulator of chromosome condensation (RCC1)], RanGTP binds to the transport receptor, thereby releasing Mel-28 and
enabling it to bind to chromatin. Following the same mechanism, the importin-b-bound Nup107–160 complex is released by RanGTP in the proximity of DNA and recruited
to chromatin through Mel-28. The chromatin-bound Nup107–160 complex can then recruit other nucleoporins in a step-wise manner. (b) NPC assembly during interphase
requires the RanGTP-dependent release of the Nup107–160 complex from importin b on the cytoplasmic and nuclear side of the NE. How the released complexes
coordinate the formation of a functional NPC from both sides of the nuclear envelope is still unclear.
Review Trends in Cell Biology Vol.18 No.10
The step-wise mechanism of NPC assembly and theexistence of intermediate structures on chromatin is sup-ported by EM studies [62–65]. Evidence from severalrecent reports has indicated that the early recruitmentof the Nup107–160 complex to chromatin is mediated byElys/Mel-28, which was initially discovered as an AT-hook-containing transcription factor [66–69]. The chromatinassociations of nups and the interactions between manyof them are regulated by the ratio of importin b to thesmall GTPase Ran [58,70]. During mitosis, importin b
associates with and sequesters a subset of nups, includingElys/Mel-28 and the Nup107–160 complex, preventingtheir association with chromatin and interaction with
other nups [58,68] (Figure 3a). The RanGTP-dependentrelease of nucleoporins from the import receptor is anearly step in NPC assembly and, because RanGTP isconcentrated around chromosomes, occurs in the proximityof the chromosomal DNA. In this way, importin b andRanGTPase coordinate the spatial positioning of theassembly of NPCs during mitosis.
Interphase assembly
During interphase, the number of pores doubles to preparethe cells for re-entering mitosis. Previously, it was believedthat NPC doubling was restricted to S-phase [71]; however,recent studies have indicated that NPCs are assembled
461

Review Trends in Cell Biology Vol.18 No.10
continuously from G1 to G2 phases [72,73]. Little is knownabout the mechanism of interphase NPC assembly.Genetic studies in S. cerevisiae have identified nucleopor-ins essential for the formation of NPCs [74–76] and arequirement for karyopherin b and RanGTP [77,78]. How-ever, the cellular mechanism used to assemble NPCs intoan intact NE remains unclear. In metazoans, the assemblyof NPCs into the NE also requires importin b and RanGTP[79]. Moreover, RanGTP and the essential Nup107–160complex are required from both sides of the NE, indicatingthat interphase NPC assembly requires the coordination ofnuclear and cytoplasmic events (Figure 3b).
EM studies inDrosophila embryos have shown that newNPCs can form in an intact nuclear envelope that lackspre-existing pores [80]. Consistent with this finding,assembly of NPCs in regions devoid of pores has beenobserved inHeLa nuclei by live imaging [79]. Furthermore,experiments using in vitro-assembled nuclei have indi-cated that NPCs can form de novo, without using subunitsfrom pre-existing pores [79].
Taken together, these results have led to the currentview of interphase NPC assembly, which indicates thatnuclear pore complexes form independently of pre-existingpores and from both sides of the nuclear envelope, follow-ing a mechanism that is at least partially conserved withmitotic-NPC assembly. This raises the question of whetherthe chromatin-bound intermediates observed during mito-sis indeed represent that part of the pore that is insertedinto theNE from the nucleoplasmic side during interphase.If so, the mitotic- and interphase-assembly mechanismswould involve a coordinated interaction between chroma-tin-bound subcomplexes, cytoplasmic subcomplexes andtransmembrane NPC components to assemble the multi-protein NPC into the double membrane of the NE.
Disassembly
NPC disassembly has only been described during mitosisand there is no evidence of pores being dismantled duringinterphase. Similar to the assembly process, NPC disas-sembly takes place through an ordered process. Partiallydisassembled pores have been observed in Drosophilaembryos [80] and in in vitro NE-disassembly studies[81]. In Drosophila embryos, only one kind of disassemblyintermediate was found during prophase, indicative of ahighly synchronized process, the existence of other extre-mely unstable intermediates or the existence of only oneintermediate in this organism [80]. By contrast, a mixedpopulation of different partially disassembled NPCs wasobserved in the same disassembling nuclei using the invitro Xenopus system, indicating the existence of an asyn-chronous mechanism [81].
The step-wise disassembly of NPCs has been followedduringNE breakdown of starfish oocytes. In these cells, theprocess starts with the release of the basket nucleoporinsNup98 and Nup153 and is followed by the release ofNup214 from the cytoplasmic ring. The dissociation ofperipheral nucleoporins into the cytoplasm leads to agradual loss of selective nuclear permeability.
Using a kinetic assay in mammalian cells [60], it hasbeen shown that NPC disassembly occurs synchronously,being faster than assembly and, intriguingly, not following
462
its exact reverse order. For example, nucleoporins that arerecruited to chromatin early during assembly, such asmembers of the Nup107–160 complex, are released beforenucleoporins that are recruited later, such as Nup58.
In support of these results, it has been observed that inDrosophila the scaffold Nup107 protein is also releasedfrom NPCs after the basket components Nup153 and Mtor(the homolog of themammalian Tpr protein) but before thecentral channel nucleoporin Nup58 [82].
In the yeasts S. cerevisiae and Schizosaccharomycespombe, which divide without NE breakdown, no disassem-bly of pores has been reported; however, in other fungiundergoing closed mitosis, such as Aspergillus nidulans, apartial disassembly of pores also occurs [83]. In these cells,the cytoplasmic dispersal of several FG nucleoporins start-ing in prophase leaves partially disassembled pores at theNE, which have an increase in permeability and enablemitotic regulators to access the nucleus.
Although the molecular mechanisms of pore disassem-bly have not been investigated in detail, all the currentevidence points towards mitotic phosphorylation as thetrigger of the disassembly process. Several groups havereported mitotic-specific hyperphosphorylation of nucleo-porins both in vivo and in vitro [61,84–87]. The Ser/Thrkinase CDC2 (Cdk1) seems to be a universal mitotic kinaseinvolved in nucleoporin phosphorylation [83,84,88,89];however, the mammalian protein kinase A (PKA) andprotein kinase C (PKC), the S. cerevisiae casein kinaseand the Aspergillus nidulands Ser/Thr kinase NIMA havealso been shown to phosphorylate NPC components[39,83,88,90]. How mitotic phosphorylation initiatesNPC disassembly is still unclear, but it is possible thatit induces the dissociation of subcomplexes and preventstheir association, because nucleoporins have to be depho-sphorylated to reassemble the pore [58].
Maintenance
The mitotic disassembly–reassembly cycle of NPCs inmetazoans ensures that every nuclear pore gets renewedduring each cell-division cycle. Yet, how NPCs are main-tained during interphase is unknown. As mentioned, sev-eral nucleoporins are dynamic and can come on and off thepore, indicating that they are exchanged or recycled fromNPCs during interphase. By contrast, the NPC scaffoldsare very stable and only exchange once per cell cycle, whenthe NE breaks down. This raises the question of how NPCscaffold proteins are maintained when the cells stop divid-ing (e.g. after differentiation). Are old pores disassembledand replaced by newNPCs? This would imply the existenceof a post-mitotic disassembly mechanism. As mentioned,no evidence of such amechanism has been described so far.Or, do the NPC scaffolds remain embedded in the NE forthe entire life span of the cell? If this is the case, then insome long-lived cells such as neurons, the core of the NPCswould remain in the NE for extensive periods of time.Because of this, the pores could be subject to age-depend-ent damage that could affect their function and possiblycompromise nuclear integrity. In favor of this idea, altera-tions in the nuclear structure and mislocalization ofnuclear proteins have been linked recently to the agingphenomena [91,92].

Review Trends in Cell Biology Vol.18 No.10
Transport-independent functions of nupsAs described, the main function of NPCs is to control thepassage of molecules between the nucleus and the cyto-plasm. A tight regulation of nucleocytoplasmic transport isessential for cell homeostasis. Consequently, alterations inNPC members or the nuclear-transport process have astrong impact on cell growth and survival and, not surpris-ingly, have been associated with several diseases such ascancer [93,94] and the rare autosomal recessive disordertriple A syndrome [95].
Besides functioning as gatekeepers of the nucleus, it hasbecome evident that NPCs and nucleoporins are impli-cated in many other biological functions [1]. NPCs havebeen shown to anchor and modulate the activity of sumoy-lating and desumoylating enzymes [96]. Interestingly,Nup358, the main component of the cytoplasmic filaments,is an active E3 ligase in the sumoylation reaction [97,98].Thus, through the SUMO (small ubiquitin-related modi-fiers) pathway, NPCs are indirectly involved in the regu-lation of numerous cellular processes such as genetranscription, DNA replication, DNA damage and repair,chromosome segregation, genome stability, cell death andsenescence.
Three nucleoporins –Nup153, Nup358 and gp210 – havebeen associated with nuclear-envelope breakdown[1,99,100]. Although Nup153 and Nup358 contribute tothis process by recruiting the coatomer coat protein I(COPI) complex to the NE, the role of gp210 in NE break-down is still unclear. Conversely, several other nucleopor-ins have been reported to have a role in NE assembly.These include the transmembrane nucleoporins Pom121and NDC1, and the soluble nucleoporins Elys/Mel-28,Nup35 and Nup155, the latter two being part of the samesubcomplex [17,99,101–103].
In the past few years, many components of the NPChave been described as having important roles duringmitosis. Nup170p, the yeast homolog of mammalianNup155, was the first nucleoporin linked to kinetochorefunction and chromosome segregation [104]. Subsequently,Nup358 was found anchored to kinetochores and spindlesthrough the export receptor CRM1 (chromosome regionmaintenance 1). Depletion of Nup358 in cells has revealedits essential role in kinetochore assembly and interactionwith microtubules [100,105]. More recently, the Nup107–
160 complex and Elys/Mel-28 were found at kinetochoresduring mitosis [106–108]. The recruitment of the Nup107–
160 complex to these structures was shown to be depend-ent on the NDC80 complex and CENP-F (centromereprotein F) and to occur upstream of Nup358. Cells depletedof several members of the Nup107–160 complex together,or the Seh1 component alone, showed a mitotic delay andfailed to attach microtubules to kinetochores properly,resulting in abnormal chromosome congression. Althoughthese cells showed activation of the spindle checkpoint andlonger spindles in metaphase, no defects in spindle assem-bly were observed [107]. In a different study, the Nup107–
160 complex was shown to be required for assembly of thebipolar spindle in vitro, although no activation of thespindle checkpoint was observed in extracts depleted ofthe complex [109]. The differences observed could beattributable to the fact that the Nup107–160 complex
strongly localizes to spindles assembled in vitro but onlytransiently associates with cellular spindles during pro-metaphase [109]. A role for the Nup107–160 complexes inspindle assembly and function is supported by the findingsthat fission yeasts lacking Nup120p, a homolog of thehuman Nup160 protein, show abnormal spindles [110].Rae1 is a shuttling nucleoporin that has also been associ-ated with spindle formation [14,15,111,112]. It was foundthat this protein is part of a large ribonucleoprotein com-plex that controls microtubule dynamics and has an essen-tial role in spindle assembly [112]. Finally, RNAiexperiments in Caenorhabditis elegans have revealed arole for several nucleoporins in the regulation of spindleorientation [113].
Very recently, a small fraction of the Nup358 nucleo-porin, the main constituent of the NPC cytoplasmic fila-ments, was found in the cytoplasm of cells co-localizingwith microtubules during interphase. In this study,Nup358 was found to interact with and regulate thedynamics of interphasemicrotubules directly, thus, havinga role in organization of the cytoskeleton [114].
Lastly, several reports have associated NPCs with chro-matin organization and the regulation of gene expression(for reviews, see Refs [115,116]), especially in lower eukar-yotes. The nuclear periphery has historically been associ-ated with gene silencing and repression (for a review, seeRef. [117]). The presence of heterochromatin patches, gene-poor chromosomal regions and silent genomic domainsadjacent to the NE support this hypothesis. In agreementwith this idea, NPCs were initially associated with silentchromatin regions [118,119]. In budding yeast, Nup145p isresponsible for tethering silent telomeric chromatin to theNPCs through the myosin-like protein (MLP)1 and MLP2proteins [118], although the role of the MLP proteins intelomere silencing and anchoring has been recently chal-lenged [120]. Opposing the classical view of the nuclearperiphery, there is now increasing evidence indicating thatthis region also functions as a gene-activating domain. In ascreening for proteins with chromatin boundary activity,the Nup2p nucleoporin was found to block the propagationof heterochromatin in specific domains, thus, preventingtheir silencing, and it is thought to do so by recruitingchromatin to pores [121]. These studies indicate that chro-matin activity can be modulated by relocation to the NPC.Chromatin immunoprecipitations (CHIP) studies withantibodies against NPC components have shown thatmany nucleoporins preferentially associate with transcrip-tionally active genes [122]. The results of this technique donot directly connect active genes to the NPC structure andit remains a possibility that the DNA binding of thenucleoporins occurs entirely in the nuclear interior. How-ever, the fact that the NPC-associated protein Sus1 hasbeen described to be a member of the Spt-Ada-Gcn5-acet-yltransferase (SAGA) histone acetylase complex involvedin transcriptional initiation [123] and required for theconfinement of GAL genes to NPCs during transcriptionalactivation in S. cerevisiae [124] directly links nuclear poresto active transcription. The relocation of genes to poresduring activation has been reported for several othergenes, including the HXK1 (encoding hexokinase-1),INO1 (encoding an inositol 1-phosphate synthase) and
463

Review Trends in Cell Biology Vol.18 No.10
several mating-response genes, although there are somecontroversial findings in the nucleoporin requirements forgene tethering (for reviews, see Refs [115,125]). Also sup-porting a role of NPCs in transcriptional activation, exper-iments in which the Nup2p was fused to the microccocalnuclease have revealed that the nuclease cleavage occursmainly in promoter regions, indicating the association ofNup2p with actively transcribed genes [126].
How genes are activated at the NPC is not well under-stood, but the Rap1 transcription factor seems to have animportant role. CHIP analyses have indicated that mostnups preferentially associate with genes containing Rap1-binding sites [122]. Furthermore, the Rap1-dependentactivation in yeast was shown to occur through the Nup84pcomplex (the yeast Nup107–160 complex homolog) at theNPC [127]. This result led to the idea of a ‘reverse recruit-mentmechanism’, indicating that, during activation, genesmove to a platform tethered to the NPC containing thepreassembled transcriptional machinery [127].
In higher eukaryotes, little is known about the connec-tion between the NPC and regulation of gene expression,but the fact that the SAGA-dependent anchorage of genesto the NPC has been reported in Drosophila [128] and thattwo nucleoporins are linked to the transcriptional regula-tion of dosage compensation in this organism [129] indicatethat the mechanism might be conserved. Interestingly, arecent report has shown the association of Nup93, a stablecomponent of the NPC scaffold, with several regions of thehuman genome [130]. But, in this case, the chromosomeregions associatedwithNup93were enriched inmarkers ofheterochromatin, circling back to first findings in buddingyeast indicating the association of NPCs with silent chro-matin. Notably, the chromosome-association of Nup93varied depending on the global histone acetylation stateof chromatin.
The association of chromatin to NPCs, together withthe fact that in higher eukaryotesNPCs are immobile [55],indicates that pores could function as positional markersat the NE for the organization and maintenance of thegenome architecture. The relative location of severalgenes or chromosome regions within the cell nucleushas been shown to vary in response to metabolic changesand during cell differentiation. The findings that theDNA–NPC interactions are dynamic and change depend-ing on the chromatin state indicate that NPCs could beinvolved, at least in part, in regulating these genomicrearrangements. Supporting a role of NPCs in celldifferentiation, Nup133, a member of the scaffoldNup107–160 complex, has recently been described to beimportant for neural differentiation during embryogen-esis, which potentially highlights a new aspect of NPC andnucleoporin function [131].
Concluding remarksOwing to its interesting properties and increasing reper-toire of functions, the biology of NPCs has become an areaof focus for many fields. Despite the substantial progressthat has been made in the understanding of NPC bio-genesis and function in the past decade, there are stilllarge gaps to fill. The actual location of every nucleoporininside the NPC structure, the detailed molecular mechan-
464
isms of assembly, disassembly and translocation throughthe pore, and how NPCs perform their transport-indepen-dent functions are just some of the mysteries that might beclarified in the coming years.
AcknowledgementsWe apologize to all colleagues whose work could not be cited directlyowing to space limitation. We thank members of the Hetzer laboratoryand Marcela Raices for helpful suggestions and critical reading of themanuscript.
References1 D’Angelo, M.A. and Hetzer, M.W. (2006) The role of the nuclear
envelope in cellular organization. Cell. Mol. Life Sci. 63, 316–3322 Cook, A. et al. (2007) Structural biology of nucleocytoplasmic
transport. Annu. Rev. Biochem. 76, 647–6713 Mosammaparast, N. and Pemberton, L.F. (2004) Karyopherins: from
nuclear-transport mediators to nuclear-function regulators. TrendsCell Biol. 14, 547–556
4 Terry, L.J. et al. (2007) Crossing the nuclear envelope: hierarchicalregulation of nucleocytoplasmic transport. Science 318, 1412–1416
5 Gall, J.G. (1967) Octagonal nuclear pores. J. Cell Biol. 32, 391–3996 Huve, J. et al. (2008) 4Pi microscopy of the nuclear pore complex.
Biophys J. 95, 877–8857 Beck, M. et al. (2004) Nuclear pore complex structure and dynamics
revealed by cryoelectron tomography. Science 306, 1387–13908 Beck, M. et al. (2007) Snapshots of nuclear pore complexes in action
captured by cryo-electron tomography. Nature 449, 611–6159 Lim, R.Y. et al. (2008) Towards reconciling structure and function in
the nuclear pore complex. Histochem. Cell Biol. 129, 105–11610 Stoffler, D. et al. (2003) Cryo-electron tomography provides novel
insights into nuclear pore architecture: implications fornucleocytoplasmic transport. J. Mol. Biol. 328, 119–130
11 Maco, B. et al. (2006) Nuclear pore complex structure and plasticityrevealed by electron and atomic force microscopy. Methods Mol. Biol.322, 273–288
12 Suntharalingam,M. andWente, S.R. (2003) Peering through the pore:nuclear pore complex structure, assembly, and function. Dev. Cell 4,775–789
13 Yang, Q. et al. (1998) Three-dimensional architecture of the isolatedyeast nuclear pore complex: functional and evolutionary implications.Mol. Cell 1, 223–234
14 Rout, M.P. et al. (2000) The yeast nuclear pore complex: composition,architecture, and transport mechanism. J. Cell Biol. 148, 635–651
15 Cronshaw, J.M. et al. (2002) Proteomic analysis of the mammaliannuclear pore complex. J. Cell Biol. 158, 915–927
16 Reichelt, R. et al. (1990) Correlation between structure and massdistribution of the nuclear pore complex and of distinct pore complexcomponents. J. Cell Biol. 110, 883–894
17 Mansfeld, J. et al. (2006) The conserved transmembrane nucleoporinNDC1 is required for nuclear pore complex assembly in vertebratecells. Mol. Cell 22, 93–103
18 Stavru, F. et al. (2006) NDC1: a crucial membrane-integralnucleoporin of metazoan nuclear pore complexes. J. Cell Biol. 173,509–519
19 Schwartz, T.U. (2005) Modularity within the architecture of thenuclear pore complex. Curr. Opin. Struct. Biol. 15, 221–226
20 Alber, F. et al. (2007) The molecular architecture of the nuclear porecomplex. Nature 450, 695–701
21 Alber, F. et al. (2007) Determining the architectures ofmacromolecular assemblies. Nature 450, 683–694
22 Isgro, T.A. and Schulten, K. (2007) Cse1p-binding dynamics reveal abinding pattern for FG-repeat nucleoporins on transport receptors.Structure 15, 977–991
23 Isgro, T.A. and Schulten, K. (2007) Association of nuclear poreFG-repeat domains to NTF2 import and export complexes. J. Mol.Biol. 366, 330–345
24 Denning, D.P. et al. (2003) Disorder in the nuclear pore complex: theFG repeat regions of nucleoporins are natively unfolded. Proc. Natl.Acad. Sci. U. S. A. 100, 2450–2455
25 Macara, I.G. (2001) Transport into and out of the nucleus. Microbiol.Mol. Biol. Rev. 65, 570–594

Review Trends in Cell Biology Vol.18 No.10
26 Peters, R. (2005) Translocation through the nuclear pore complex:selectivity and speed by reduction-of-dimensionality. Traffic 6, 421–
42727 Frey, S. and Gorlich, D. (2007) A saturated FG-repeat hydrogel can
reproduce the permeability properties of nuclear pore complexes. Cell130, 512–523
28 Frey, S. et al. (2006) FG-rich repeats of nuclear pore proteins form athree-dimensional meshwork with hydrogel-like properties. Science314, 815–817
29 Ribbeck, K. and Gorlich, D. (2001) Kinetic analysis of translocationthrough nuclear pore complexes. EMBO J. 20, 1320–1330
30 Fried, H. and Kutay, U. (2003) Nucleocytoplasmic transport: takingan inventory. Cell. Mol. Life Sci. 60, 1659–1688
31 Patel, S.S. et al. (2007) Natively unfolded nucleoporins gate proteindiffusion across the nuclear pore complex. Cell 129, 83–96
32 Lim, R.Y. et al. (2006) Flexible phenylalanine-glycine nucleoporins asentropic barriers to nucleocytoplasmic transport. Proc. Natl. Acad.Sci. U. S. A. 103, 9512–9517
33 Rout, M.P. et al. (2003) Virtual gating and nuclear transport: the holepicture. Trends Cell Biol. 13, 622–628
34 Rabut, G. et al. (2004) Mapping the dynamic organization of thenuclear pore complex inside single living cells. Nat. Cell Biol. 6,1114–1121
35 Griffis, E.R. et al. (2004) Distinct functional domains withinnucleoporins Nup153 and Nup98 mediate transcription-dependentmobility. Mol. Biol. Cell 15, 1991–2002
36 Storr, H.L. et al. (2005) Identification of the sites of expression of tripleA syndromemRNA in the rat using in situ hybridisation.Neuroscience131, 113–123
37 Olsson, M. et al. (2004) Limited expression of nuclear pore membraneglycoprotein 210 in cell lines and tissues suggests cell-type specificnuclear pores in metazoans. Exp. Cell Res. 292, 359–370
38 Fan, F. et al. (1997) cDNA cloning and characterization of Npap60: anovel rat nuclear pore-associated protein with an unusual subcellularlocalization during male germ cell differentiation. Genomics 40, 444–
45339 Cai, Y. et al. (2002) Characterization and potential function of a novel
testis-specific nucleoporin BS-63. Mol. Reprod. Dev. 61, 126–13440 Akey, C.W. (1995) Structural plasticity of the nuclear pore complex.
J. Mol. Biol. 248, 273–29341 Kiseleva, E. et al. (1996) RNP export is mediated by structural
reorganization of the nuclear pore basket. J. Mol. Biol. 260, 304–31142 Erickson, E.S. et al. (2006) The role of nuclear envelope calcium in
modifying nuclear pore complex structure. Can. J. Physiol.Pharmacol. 84, 309–318
43 Stoffler, D. et al. (1999) Calcium-mediated structural changes ofnative nuclear pore complexes monitored by time-lapse atomicforce microscopy. J. Mol. Biol. 287, 741–752
44 Rakowska, A. et al. (1998) ATP-induced shape change of nuclear poresvisualized with the atomic force microscope. J. Membr. Biol. 163, 129–
13645 Perez-Terzic, C. et al. (1996) Conformational states of the nuclear pore
complex induced by depletion of nuclear Ca2+ stores. Science 273,1875–1877
46 Shahin, V. et al. (2005) Glucocorticoids remodel nuclear envelopestructure and permeability. J. Cell Sci. 118, 2881–2889
47 Paulillo, S.M. et al. (2006) Changes in nucleoporin domain topology inresponse to chemical effectors. J. Mol. Biol. 363, 39–50
48 Fahrenkrog, B. et al. (2002) Domain-specific antibodies revealmultiple-site topology of Nup153 within the nuclear pore complex.J. Struct. Biol. 140, 254–267
49 Paulillo, S.M. et al. (2005) Nucleoporin domain topology is linked tothe transport status of the nuclear pore complex. J. Mol. Biol. 351,784–798
50 Makhnevych, T. et al. (2003) Cell cycle regulated transport controlledby alterations in the nuclear pore complex. Cell 115, 813–823
51 Melcak, I. et al. (2007) Structure of Nup58/45 suggests flexiblenuclear pore diameter by intermolecular sliding. Science 315,1729–1732
52 Lim, R.Y. et al. (2007) Nanomechanical basis of selective gating by thenuclear pore complex. Science 318, 640–643
53 Maul, G.G. (1977) Nuclear pore complexes. Elimination andreconstruction during mitosis. J. Cell Biol. 74, 492–500
54 Burke, B. and Ellenberg, J. (2002) Remodelling the walls of thenucleus. Nat. Rev. Mol. Cell Biol. 3, 487–497
55 Daigle, N. et al. (2001) Nuclear pore complexes form immobilenetworks and have a very low turnover in live mammalian cells.J. Cell Biol. 154, 71–84
56 Belgareh, N. et al. (2001) An evolutionarily conserved NPCsubcomplex, which redistributes in part to kinetochores inmammalian cells. J. Cell Biol. 154, 1147–1160
57 Walther, T.C. et al. (2003) The conserved Nup107–160 complex iscritical for nuclear pore complex assembly. Cell 113, 195–206
58 Walther, T.C. et al. (2003) RanGTP mediates nuclear pore complexassembly. Nature 424, 689–694
59 Theisen, U. et al. (2008) Dynamic rearrangement of nucleoporinsduring fungal ‘‘open’’ mitosis. Mol. Biol. Cell 19, 1230–1240
60 Dultz, E. et al. (2008) Systematic kinetic analysis of mitotic dis- andreassembly of the nuclear pore in living cells. J. Cell Biol. 180, 857–865
61 Bodoor, K. et al. (1999) Sequential recruitment of NPC proteins to thenuclear periphery at the end of mitosis. J. Cell Sci. 112, 2253–2264
62 Goldberg, M.W. et al. (1997) Dimples, pores, star-rings, and thin ringson growing nuclear envelopes: evidence for structural intermediatesin nuclear pore complex assembly. J. Cell Sci. 110, 409–420
63 Macaulay, C. and Forbes, D.J. (1996) Assembly of the nuclear pore:biochemically distinct steps revealed with NEM, GTP g S, andBAPTA. J. Cell Biol. 132, 5–20
64 Sheehan, M.A. et al. (1988) Steps in the assembly of replication-competent nuclei in a cell-free system from Xenopus eggs. J. CellBiol. 106, 1–12
65 Drummond, S.P. et al. (2006) High resolution analysis of mammaliannuclear structure throughout the cell cycle: implications for nuclearpore complex assembly during interphase and mitosis. Can.J. Physiol. Pharmacol. 84, 423–430
66 Kimura, N. et al. (2002) Identification of a novel transcription factor,ELYS, expressed predominantly in mouse foetal haematopoietictissues. Genes Cells 7, 435–446
67 Rasala, B.A. et al. (2006) ELYS is a dual nucleoporin/kinetochoreprotein required for nuclear pore assembly and proper cell division.Proc. Natl. Acad. Sci. U. S. A. 103, 17801–17806
68 Franz, C. et al. (2007)MEL-28/ELYS is required for the recruitment ofnucleoporins to chromatin and postmitotic nuclear pore complexassembly. EMBO Rep. 8, 165–172
69 Gillespie, P.J. et al. (2007) ELYS/MEL-28 chromatin associationcoordinates nuclear pore complex assembly and replicationlicensing. Curr. Biol. 17, 1657–1662
70 Harel, A. et al. (2003) Importin b negatively regulates nuclearmembrane fusion and nuclear pore complex assembly. Mol. Biol.Cell 14, 4387–4396
71 Maul, G.G. et al. (1972) Time sequence of nuclear pore formation inphytohemagglutinin-stimulated lymphocytes and in HeLa cellsduring the cell cycle. J. Cell Biol. 55, 433–447
72 Maeshima, K. et al. (2006) Cell-cycle-dependent dynamics of nuclearpores: pore-free islands and lamins. J. Cell Sci. 119, 4442–4451
73 Winey, M. et al. (1997) Nuclear pore complex number and distributionthroughout the Saccharomyces cerevisiae cell cycle by three-dimensional reconstruction from electron micrographs of nuclearenvelopes. Mol. Biol. Cell 8, 2119–2132
74 Vasu, S.K. and Forbes, D.J. (2001) Nuclear pores and nuclearassembly. Curr. Opin. Cell Biol. 13, 363–375
75 Lusk, C.P. et al. (2002) Karyopherins in nuclear pore biogenesis: a rolefor Kap121p in the assembly of Nup53p into nuclear pore complexes.J. Cell Biol. 159, 267–278
76 Madrid, A.S. et al. (2006) The role of the integral membranenucleoporins Ndc1p and Pom152p in nuclear pore complexassembly and function. J. Cell Biol. 173, 361–371
77 Ryan, K.J. et al. (2003) The Ran GTPase cycle is required for yeastnuclear pore complex assembly. J. Cell Biol. 160, 1041–1053
78 Ryan, K.J. et al. (2007) The karyopherin Kap95 regulates nuclear porecomplex assembly into intact nuclear envelopes in vivo.Mol. Biol. Cell18, 886–898
79 D’Angelo, M.A. et al. (2006) Nuclear pores form de novo from bothsides of the nuclear envelope. Science 312, 440–443
80 Kiseleva, E. et al. (2001) Steps of nuclear pore complex disassemblyand reassembly during mitosis in early Drosophila embryos. J. CellSci. 114, 3607–3618
465

Q1
Review Trends in Cell Biology Vol.18 No.10
81 Cotter, L. et al. (2007) Nuclear membrane disassembly and rupture.J. Mol. Biol. 369, 683–695
82 Katsani, K.R. et al. (2008) In vivo dynamics of Drosophila nuclearenvelope components. Mol Biol Cell. (in press)
83 De Souza, C.P. et al. (2004) Partial nuclear pore complex disassemblyduring closed mitosis in Aspergillus nidulans. Curr. Biol. 14, 1973–
198484 Macaulay, C. et al. (1995) Differential mitotic phosphorylation
of proteins of the nuclear pore complex. J. Biol. Chem. 270,254–262
85 Favreau, C. et al. (1996) Cell cycle-dependent phosphorylation ofnucleoporins and nuclear pore membrane protein Gp210.Biochemistry 35, 8035–8044
86 Glavy, J.S. et al. (2007) Cell-cycle-dependent phosphorylation of thenuclear pore Nup107–160 subcomplex. Proc. Natl. Acad. Sci. U. S. A.104, 3811–3816
87 Nousiainen, M. et al. (2006) Phosphoproteome analysis of the humanmitotic spindle. Proc. Natl. Acad. Sci. U. S. A. 103, 5391–5396
88 Lusk, C.P. et al. (2007) Nup53p is a target of two mitotic kinases,Cdk1p and Hrr25p. Traffic 8, 647–660
89 Onischenko, E.A. et al. (2005) Cdk1 and okadaic acid-sensitivephosphatases control assembly of nuclear pore complexes inDrosophila embryos. Mol. Biol. Cell 16, 5152–5162
90 Miller, M.W. et al. (1999) Phosphorylation and glycosylation ofnucleoporins. Arch. Biochem. Biophys. 367, 51–60
91 Herndon, L.A. et al. (2002) Stochastic and genetic factors influencetissue-specific decline in ageing C. elegans. Nature 419, 808–814
92 Haithcock, E. et al. (2005) Age-related changes of nuclear architecturein Caenorhabditis elegans. Proc. Natl. Acad. Sci. U. S. A. 102, 16690–
1669593 Kau, T.R. et al. (2004) Nuclear transport and cancer: frommechanism
to intervention. Nat. Rev. Cancer 4, 106–11794 Poon, I.K. and Jans, D.A. (2005) Regulation of nuclear transport:
central role in development and transformation? Traffic 6,173–186
95 Cronshaw, J.M. and Matunis, M.J. (2004) The nuclear pore complex:disease associations and functional correlations. Trends Endocrinol.Metab. 15, 34–39
96 Palancade, B. and Doye, V. (2008) Sumoylating and desumoylatingenzymes at nuclear pores: underpinning their unexpected duties?Trends Cell Biol. 18, 174–183
97 Pichler, A. et al. (2004) The RanBP2 SUMO E3 ligase is neitherHECT- nor RING-type. Nat. Struct. Mol. Biol. 11, 984–991
98 Pichler, A. et al. (2002) The nucleoporin RanBP2 has SUMO1 E3ligase activity. Cell 108, 109–120
99 Galy, V. et al. (2008) A role for gp210 in mitotic nuclear-envelopebreakdown. J. Cell Sci. 121, 317–328
100 Salina, D. et al. (2003) Nup358 integrates nuclear envelopebreakdown with kinetochore assembly. J. Cell Biol. 162, 991–1001
101 Antonin, W. et al. (2005) The integral membrane nucleoporin pom121functionally links nuclear pore complex assembly and nuclearenvelope formation. Mol. Cell 17, 83–92
102 Hawryluk-Gara, L.A. et al. (2008) Nup53 is required for nuclearenvelope and nuclear pore complex assembly. Mol Biol Cell. 19,1753–1762
103 Franz, C. et al. (2005) Nup155 regulates nuclear envelope and nuclearpore complex formation in nematodes and vertebrates. Embo J. 24,3519–3531
104 Kerscher, O. et al. (2001) Novel role for a Saccharomyces cerevisiaenucleoporin, Nup170p, in chromosome segregation. Genetics 157,1543–1553
105 Joseph, J. et al. (2004) The RanGAP1–RanBP2 complex is essentialfor microtubule-kinetochore interactions in vivo. Curr. Biol. 14, 611–
617106 Loiodice, I. et al. (2004) The entire Nup107–160 complex, including
three new members, is targeted as one entity to kinetochores inmitosis. Mol. Biol. Cell 15, 3333–3344
466
107 Zuccolo, M. et al. (2007) The human Nup107–160 nuclear poresubcomplex contributes to proper kinetochore functions. EMBO J.26, 1853–1864
108 Galy, V. et al. (2006) MEL-28, a novel nuclear-envelope andkinetochore protein essential for zygotic nuclear-envelope assemblyin C. elegans. Curr. Biol. 16, 1748–1756
109 Orjalo, A.V. et al. (2006) The Nup107–160 nucleoporin complex isrequired for correct bipolar spindle assembly.Mol. Biol. Cell 17, 3806–
3818110 Bai, S.W. et al. (2004) The fission yeast Nup107–120 complex
functionally interacts with the small GTPase Ran/Spi1 and isrequired for mRNA export, nuclear pore distribution, and propercell division. Mol. Cell. Biol. 24, 6379–6392
111 Pritchard, C.E. et al. (1999) RAE1 is a shuttling mRNA export factorthat binds to a GLEBS-like NUP98 motif at the nuclear pore complexthrough multiple domains. J. Cell Biol. 145, 237–254
112 Blower, M.D. et al. (2005) A Rae1-containing ribonucleoproteincomplex is required for mitotic spindle assembly. Cell 121, 223–234
113 Schetter, A. et al. (2006) Nucleoporins NPP-1, NPP-3, NPP-4, NPP-11and NPP-13 are required for proper spindle orientation in C. elegans.Dev. Biol. 289, 360–371
114 Joseph, J. and Dasso, M. (2008) The nucleoporin Nup358 associateswith and regulates interphasemicrotubules.FEBSLett. 582, 190–196
115 Brown, C.R. and Silver, P.A. (2007) Transcriptional regulation at thenuclear pore complex. Curr. Opin. Genet. Dev. 17, 100–106
116 Kalverda, B. et al. (2008) Chromatin organization in relation to thenuclear periphery. FEBS Lett. 582, 2017–2022
117 Shaklai, S. et al. (2007) Gene silencing at the nuclear periphery.FEBSJ. 274, 1383–1392
118 Galy, V. et al. (2000) Nuclear pore complexes in the organization ofsilent telomeric chromatin. Nature 403, 108–112
119 Feuerbach, F. et al. (2002) Nuclear architecture and spatialpositioning help establish transcriptional states of telomeres inyeast. Nat. Cell Biol. 4, 214–221
120 Hediger, F. et al. (2002) Myosin-like proteins 1 and 2 are not requiredfor silencing or telomere anchoring, but act in the Tel1 pathway oftelomere length control. J. Struct. Biol. 140, 79–91
121 Ishii, K. et al. (2002) Chromatin boundaries in budding yeast: thenuclear pore connection. Cell 109, 551–562
122 Casolari, J.M. et al. (2004) Genome-wide localization of the nucleartransport machinery couples transcriptional status and nuclearorganization. Cell 117, 427–439
123 Rodriguez-Navarro, S. et al. (2004) Sus1, a functional component ofthe SAGA histone acetylase complex and the nuclear pore-associatedmRNA export machinery. Cell 116, 75–86
124 Cabal, G.G. et al. (2006) SAGA interacting factors confine sub-diffusion of transcribed genes to the nuclear envelope. Nature 441,770–773
125 Akhtar, A. and Gasser, S.M. (2007) The nuclear envelope andtranscriptional control. Nat. Rev. Genet. 8, 507–517
126 Schmid, M. et al. (2006) Nup-PI: the nucleopore-promoter interactionof genes in yeast. Mol. Cell 21, 379–391
127 Menon, B.B. et al. (2005) Reverse recruitment: the Nup84 nuclearpore subcomplex mediates Rap1/Gcr1/Gcr2 transcriptionalactivation. Proc. Natl. Acad. Sci. U. S. A. 102, 5749–5754
128 Kurshakova, M.M. et al. (2007) SAGA and a novel Drosophila exportcomplex anchor efficient transcription and mRNA export to NPC.EMBO J. 26, 4956–4965
129 Mendjan, S. et al. (2006) Nuclear pore components are involved in thetranscriptional regulation of dosage compensation inDrosophila.Mol.Cell 21, 811–823
130 Brown, C.R. et al. (2008) Global histone acetylation induces functionalgenomic reorganization atmammalian nuclear pore complexes.GenesDev. 22, 627–639
131 Lupu, F. et al. (2008) Nuclear pore composition regulates neural stem/progenitor cell differentiation in the mouse embryo.Dev. Cell 14, 831–
842





























