Embed Size (px)
Citation preview

Review article
Parasitic peptides! The structure and function of neuropeptides inparasitic worms
Tim A. Daya,*, Aaron G. Mauleb
aDepartment of Pharmacology and Toxicology, Michigan State University, East Lansing, MI 48824, USAbParasitology Research Group, The Queen’s University of Belfast, Belfast BT9 7BL, Northern Ireland
Received 15 December 1998, accepted 25 January 1999
Abstract
Parasitic worms come from two very different phyla—Platyhelminthes (flatworms) and Nematoda (roundworms). Although both phylapossess nervous systems with highly developed peptidergic components, there are key differences in the structure and action of nativeneuropeptides in the two groups. For example, the most abundant neuropeptide known in platyhelminths is the pancreatic polypeptide-likeneuropeptide F, whereas the most prevalent neuropeptides in nematodes are FMRFamide-related peptides (FaRPs), which are also presentin platyhelminths. With respect to neuropeptide diversity, platyhelminth species possess only one or two distinct FaRPs, whereas nematodeshave upwards of 50 unique FaRPs. FaRP bioactivity in platyhelminths appears to be restricted to myoexcitation, whereas both excitatoryand inhibitory effects have been reported in nematodes. Recently interest has focused on the peptidergic signaling systems of both phylabecause elucidation of these systems will do much to clarify the basic biology of the worms and because the peptidergic systems hold thepromise of yielding novel targets for a new generation of antiparasitic drugs. © 1999 Elsevier Science Inc. All rights reserved.
Keywords:Invertebrate; Platyhelminth; Nematode; FMRFamide-related peptide; Neuropeptide F
1. Introduction
Morbidity due to helminths in humans is alarming. N.R.Stoll estimated in 1947 [103] that there were 2200 millionhelminth infections (distributed very unevenly) among theexisting world population of 2166 million persons—morethan one infection per human being! Current estimates sug-gest that as many as one in every three humans harbors oneor more species of parasitic helminth ([107]; Table 1).These human helminth infections are focused largely indeveloping countries and, in addition to inflicting immensepersonal suffering, stand as an important barrier to devel-opment in some parts of the world. Although mortality fromthese infections can be calculated reasonably, the chronicand insidious effects of continuous helminth infectionsthroughout childhood are hard to measure, leaving it diffi-cult to assess the overall impact.
The parasitic helminths are a grouping of two very di-verse phyla of worms: flatworms of the phylum Platyhel-
* Corresponding author. Tel.:11-517-432-4075, fax:11-517-353-8915.
E-mail address: [email protected] (T.A. Day)
Table 1Summary of major human helminth infections
Organism Common nameof worm/disease
Infectious(millions)
Platyhelminths (Flatworms)Trematodes (Flukes)Clinorchis sinensis Liver fluke 32
Fasciola hepatica, F. buski Liver fluke, Intestinalfluke
17
Paragonimus westermani Lung fluke 6Schistosomaspp. Blood fluke 276
Cestodes (Tapeworms)Diphyllobothrium latum Fish tapeworm 9Taenia saginata Beef tapeworm 73Taenia solium Pork tapeworm 5
Nematodes (Roundworms)Ascaris lumbricoides Giant roundworm 1129Brugia malayi Filariasis 100Ancylostoma duodenale,
Necator americanusHookworm 877
Onchocerca volvulus River blindness 18Trichinella spiralis Trichinosis 11Trichuris trichiura Whipworm 770Wuchereria bancrofti Filariasis (Elephantiasis) 184
Total estimated infections (millions) 3089
Peptides 20 (1999) 999–1019
0196-9781/99/$ – see front matter © 1999 Elsevier Science Inc. All rights reserved.PII: S0196-9781(99)00093-5

minthes, and roundworms of the phylum Nematoda. Thesetwo phyla have little in common apart from gross wormmorphology and the parasitic lifestyle of some members.
Flatworms are the earliest diverging extant phylum ofbilateral animals. They are the most primitive organismswith anterior cephalization and a centralized nervous sys-tem, features that appear to have accompanied bilateralism.With this neural organization and a greater differentiationand proliferation of muscle, flatworms have more directiveand highly developed motor systems than their radiallysymmetrical ancestors. The phylum consists of threeclasses, two of which are completely parasitic (Trematodaand Cestoda) and one of which is almost exclusively free-living (Turbellaria). Although the relationships within thephylum are not clear, it is not disputed that the parasiticforms have descended from the free-living forms. The flat-worm parasite with the most global impact is, without ques-tion, the schistosome. Schistosomes (primarilySchistosomamansoni, S. mekongi, S. haematobium, and S. japonicum)are blood flukes infecting well over 200 million peopleworldwide, and over 600 million live at risk.
Roundworms are phylogenetically distant from the flat-worms, and their cladistical position is disputed. Compel-ling evidence suggests that nematodes are closely related toarthropods and should be included in a large clade of moult-ing animals [1]. Nematodes are wildly successful organ-isms, occupying almost every conceivable niche. Only asmall percentage of the roundworms are parasitic in man oranimals. Important human infections include the intestinalroundwormAscaris lumbricoides, the hookwormsNecatoramericanusand Ancylostoma duodenale, and the filarialwormsBrugia malayiiandOnchocerca volvulus, etiologicalagents of elephantiasis and onchocerciasis (river blindness),respectively.
Biological dissimilarities between the platyhelminth andnematode parasites are more striking than are their similar-ities. In broad terms, there is a remarkably wide variation ofbody plans within the platyhelminths. In an authoritativepublication on the invertebrates, Libbie Hyman wrote inreference to the flatworms, “The phylum as here understoodis divided into the three classes Turbellaria, Trematoda, andCestoda. These differ so much in anatomy and life historythat a general presentation is not feasible. . . ” [36]. Incontrast, the body plan of nematodes is remarkably consis-tent, although there are considerable size variations. Nem-atodes are pseudocoelomates with internal structures exist-ing in a pressurized, fluid-filled cavity (the pseudocoel)between the outer body wall and the inner digestive tract;platyhelminths are acoelomate, and their cells are packedtightly together with very limited extracellular space. Fur-thermore, nematodes are composed of a rather small numberof typically large cells, and platyhelminths typically consistof a large number of small cells. This generalization extendsto the nervous systems, where there is a marked differencein anatomical complexity. For example, the free-living nem-atodeCaenorhabditis eleganshas 302 neurons (in the her-
maphrodite) and the intestinal pig roundwormAscaris suumhas'300 neurons. Many of the neurons inC. elegansandA. suumhave been extensively characterized in terms ofontogeny, synaptic inter-relationships, and endogenous neu-rotransmitters. Comparatively, platyhelminth nervous sys-tems are much more anatomically complex and much lesswell characterized. There are many more neurons, there areno clearly defined roles for any particular transmitter, andeven the nature of neuromuscular relationships is still un-clear. Despite these important differences between platyhel-minth and nematode parasites, they are often consideredtogether because of the parasitic lifestyle they share. Onemajor drawback of these biological differences betweenflatworm and roundworm parasites is that the spectrum forantiparasitic agents rarely spans both groups.
There are no vaccines against human helminth infectionspresently available, despite continuing efforts [13]. There-fore, chemotherapy continues to bear most of the burden ofefforts to control helminth infections. Disease control strat-egies historically had focused on eradication by means ofintense, time-limited campaigns intended to break the cycleof transmission in specific geographical areas. Today, theemphasis is more often control rather than eradication, withthe practical goal of minimizing helminth-induced morbid-ity. A key component in these efforts to control morbidityhas been population-based chemotherapy, including masschemotherapy of total populations regardless of definitiveindication of infection in each individual [107]. Unfortu-nately, such intense, sometimes indiscriminate use of an-thelmintics is conducive to the development of drug-resis-tant parasites.
Both platyhelminths and nematodes have proven theirability to develop resistance to chemotherapeutics. For ex-ample, drug resistance among nematode parasites of veter-inary importance has crippled the sheep industry in regionsof Australia, South America, and South Africa [31]. Inaddition to the established information on schistosome re-sistance to earlier generations of chemotherapeutics [13],well documented reports of schistosome resistance to prazi-quantel have emerged recently [38], which is frighteningbecause this is the only antischistosomal drug available inmuch of the world. The heavy dependence on chemotherapyand the ability of the targets to develop resistance forceparasitologists into an ongoing search for new drug targets.This requires an improved understanding of the biology ofthe parasites because it is inherent that any potential targetmust be critical to the parasites. The identification of bio-logically relevant targets is essential to the mechanism-based screens central to many current drug developmentstrategies.
In this context, the important role of neuropeptides in thebiology of both platyhelminths and nematodes has capti-vated the interest of many parasitologists. The peptidergicnervous system has emerged as a promising source for newanthelmintic targets. This review will summarize the data
1000 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

that points to the importance of neuropeptides in the lives ofthese parasites.
2. Platyhelminths
Neuropeptide-immunoreactivity (IR) is widespreadthroughout the nervous systems of all flatworms investi-gated to date, with a large number of examples from each ofthe classes (see [33,94]). When these studies began in the1980s, the antisera used were raised against specific neu-ropeptides derived from other animals (usually vertebrates,in some cases insects and mollusks), and the actual identityof the flatworm antigens visualized remained uncertain.These studies have expanded to include over 40 differentflatworm species and antisera to over 30 different peptides.Despite the obscurity of the flatworm antigens involved,patterns emerged from these studies that foreshadowed thepeptides that would later be discovered in flatworms; spe-cifically, antibodies targeting neuropeptide Y (NPY) familymembers and FMRFamide-related peptides (FaRPs) alwaysproduced abundant staining.
3. Neuropeptide F (NPF) in platyhelminths
3.1. Localization
The vertebrate NPY family consists of NPY, peptide YY(PYY), and pancreatic polypeptide (PP), grouped togetherbased on structural similarity and evolutionary relationships[77]. Early reports of IR to vertebrate peptides in flatwormsincluded plentiful examples of immunostaining to each ofthe NPY family members, and antibodies raised againstdifferent NPY family members usually stained the sameneurons. NPY family immunoreactive neurons were identi-fied in extensive regions of both the central and peripheralnervous systems of flatworms. PP-IR was invariably themost extensive among all the neuropeptides tested, andNPY-IR was unreliable and often reported as being lessintense or absent. For example, in the trematodeDiclid-ophora merlangi, which lives clamped to the gills of whit-ing, antibodies to all three NPY family members stained thesame neural structures, but NPY-IR was much less intense[66]. Similarly, PP antisera produced the most intense stain-ing of the nervous system of the fish cestodeTriloculariaacanthiaevulgaris, whereas NPY antisera gave negative re-sults [26]. In these studies and others, the most effectiveantisera were those raised against the C terminus of PP.These data induced the reasonable conclusion that all of theNPY family antisera were binding to the same parasiteneuropeptide, a neuropeptide with a PP-like C terminus.This belief was supported by the chromatographic analysisof ethanolic extracts ofD. merlangishowing a single peakof PP-IR with a molecular weight similar to that of NPYfamily members [74].
These types of immunocytochemical clues facilitated theeventual isolation and structural characterization of a PP-like neuropeptide from the sheep tapewormMoniezia ex-pansa, called NPF ([76]). The structural information en-abled the generation of NPF-specific antibodies that did notcross react with vertebrate NPY, PYY, or PP. InMoniezia,studies with the NPF-specific antibodies revealed identicalstaining patterns to those obtained with NPY family anti-bodies, suggesting that the reported NPY, PYY, and PPstaining were all attributable to the presence of NPF. Ad-dition of excess NPF abolished staining obtained with PPantisera whereas excess PP, PYY, or NPY did not abolishthe NPF-IR, further supporting the attribution of all NPYfamily IR to NPF [72]. Subsequently, antisera raised againstthe C terminus ofM. expansaNPF (mxNPF) have been usedto demonstrate NPF-IR in the nervous systems of almost 30flatworm species and, in each case where the comparisoncan be made, the staining pattern is identical to that obtainedby using NPY family antisera. Important to note, NPF-IRhas been reported as being intense, abundant, and wide-spread throughout the central and peripheral nervous sys-tems of all studied flatworms. In this discussion of thedistribution of NPF-IR, some earlier reports of NPY familyIR are drawn upon and are referred to collectively as NPF-IR.
The flatworm nervous system can be thought of as amelding of a complex brain with a system of more primitivenerve nets. The central nervous system is characterized bythe condensation of anterior elements into paired cerebralganglia, forming a structure that is indeed a brain, not onlyon the basis of gross morphology but also on the basis offunction in controlling and inhibiting signals originating inthe peripheral nervous system [44,45]. Emanating from thebrain are a pair of main longitudinal nerve cords that areconnected by a series of commissures as they progressposteriorly, creating a ladder-like ‘orthogonal’ structure.The peripheral nervous system is primarily comprised of anumber of nerve plexuses that are highly analogous to nervenets of the radially symmetrical cnidaria. The cytology ofthe nervous system is consistent with this depiction of acomplex brain joined to more primitive peripheral nervenets, as the peripheral nervous system consists exclusivelyof multi- and bi-polar cells that are thought to be moreancient, whereas the central nervous system ganglia oftenfeature more specialized unipolar cells. In the central ner-vous system, NPF-IR is present in the paired cerebral gan-glia, the fiber tracts of the longitudinal nerve cords, and theircommissural connections. In the peripheral nervous system,NPF-IR is present throughout the plexuses innervating thereproductive structures, the somatic musculature, the organsof attachment, and the alimentary system.
NPF-IR is remarkable and consistent throughout the re-productive structures of flatworms. In addition to beingpioneers of bilateral symmetry, flatworms are also the ear-liest animals to have established complex reproductivestructures, including gonads and muscle-lined oviducts. Al-
1001T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

though the worms are almost all hermaphroditic, they gen-erally have complicated copulatory structures and repro-duce, when possible, by cross-fertilization. Differencesexist in the NPF-IR of the copulatory structures, likely areflection of the wide variability observed in the structuresthemselves, but staining is abundant in the nerves surround-ing the egg chamber in every flatworm investigated thus far.For example, in the trematodesFasciola hepatica[53,57]and S. mansoni[96,57], NPF-IR is widespread in nervefibers and neural cell bodies surrounding the muscular eggchamber, as well as in specific nerves positioned at theentrances of various ducts. The predominance of NPF-im-munoreactive nerve fibers serving the muscular egg cham-ber ofS. mansoni[57] is of great interest because the mostserious pathology from schistosome infections is attribut-able to their prodigious egg production. A single femaleschistosome can produce thousands of eggs daily, many ofwhich become trapped in the walls of the intestine, the liver,or the spleen, leading to inflammatory responses that resultin the most serious pathologies. The mechanisms of controlof this egg production in platyhelminths remains unknown,but the predominance of NPF-IR in the nerves of the eggchamber and reproductive ducts could reflect a role for thepeptide in controlling the flow of material in and out of thechamber where eggs are produced.
A number of reports demonstrate NPF-IR in the nerveplexuses serving the subtegumental muscle layers, althoughthe functional relationships between the somatic muscula-ture and the nervous system remain unclear. Flatwormshave distinct groupings of somatic muscles that differ some-what among the various classes. Trematodes and turbellar-ians possess a subtegumental layer of circular muscle fibersoverlying a layer of longitudinal muscle and an inner layerof diagonal muscle. In cestodes, the diagonal muscle layersare generally absent [56]. Flatworms are also invested withdorso–ventral muscle fibers believed to maintain their flat-tened shape, and many also possess deeper layers of longi-tudinal muscle fibers; the latter are particularly evident inmonogenean trematodes and cestodes. Ultrastructural evi-dence suggests that the somatic musculature is controlled byboth direct synaptic input and more general, paracrine re-lease from nerve terminals not directly apposed to the mus-cle fibers. It is unclear how the worms exercise differentialcontrol over these distinct muscle groups, whether by dif-ferent pharmacological properties of the muscles or by dis-creet neuromuscular relationships. Nevertheless, examplesof NPF-IR in the peripheral plexuses that serve the somaticmusculature exist in each of the classes of platyhelminths,including the trematodeS. mansoni[96], the cestodeM.expansa[68], and the brackish water turbellarianProcer-odes littoralis[85].
For parasitic platyhelminths, the maintenance of appro-priate position in or on the host is essential, and a widevariety of holdfast organs fulfilling this need have devel-oped. These organs of attachment are also consistently in-nervated with NPF-positive fibers. For example, the primary
holdfast organ of the trematodeD. merlangi is a muscularhaptor, a muscular multi-fingered clamp with which theworm attaches to the gill lamellae of host fish. This haptoris innervated by NPF-positive fibers from the longitudinalcords of the central nervous system and the associatedperipheral plexus [64,66]. Among other trematodes inwhich NPF positive innervation of the holdfast organs hasbeen demonstrated are schistosomes, where the suckerswith which the males attach to the vascular walls of the hostare richly supplied with peptidergic elements [57]. There arealso many examples among the cestodes, including themuscular suckers ofM. expansa[72].
The alimentary systems of platyhelminths also containmany NPF-immunopositive nerves, especially prevalent inthe muscular pharynx or esophagus, which acts to send fooditems into the intestine and to expel waste products. Inturbellarians, which generally forage for either living ordecaying material, the most common plan is a muscularpharynx, an esophagus, and a blind intestine, all of whichare typically innervated with NPF-immunoreactive fibersand cell bodies. The pharynx has proven to be conspicu-ously supplied in many instances, for example the brackishwater turbellarians,Archiloa unipunctata[86] andP. litto-ralis [85]. Many ectoparasitic trematodes have pharynxplans akin to those of the turbellarians, and they have alsoproven to be innervated with NPF-immunopositive nerves,for exampleD. merlangi [55]. Schistosomes are typical ofmany blood-dwelling flatworms in that entry to the esoph-agus is provided by a mouth integral to the oral sucker,which, in schistosomes, is innervated by NPF-positive fibers[56,96].
A consistent observation is that the NPF-IR tends tocorrespond to the cholinergic system, more so than theserotonergic system. This has been noted, for example, inthe cestodeM. expansa[68], the trematodeD. merlangi[66], and the turbellarianMicrostomum lineare[86]. How-ever, conclusive evidence for the co-localization of neu-ropeptides and acetylcholine in flatworm neurons has notbeen presented.
Another consistent observation has been that adult tur-bellarians have a greater quantity of NPF-IR than do theparasitic classes. In the turbellarians examined, there isbetween 2000 and 5000 ng of NPF-IR/g (wet weight) ofworm, with only 20–50 ng/g in adult parasitic forms [40,57]. This may be simply an extension of the observation thatneural elements are, in general, larger and more abundant inthe free-living worms than in adult parasitic forms. Themost accepted hypothesis explaining this disparity is thedifference in the number of motor and sensory capabilitiesrequired to facilitate a free-living as opposed to a parasiticlifestyle. The former would require a higher level of neu-romuscular development to enable worms to find food andto react to environmental cues. Endoparasitism has obviatedthe need for many flatworms to actively search for food.Similarly, the likelihood of requiring an evasive responsehas been greatly diminished by the parasitic habit. It should
1002 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

be noted, however, that parasitic flatworms often have free-living stages that must find the appropriate host organism toensure survival and that these free-living juvenile stages ofthe worms display relatively higher levels of neuromusculardevelopment than the parasitic adults [40].
3.2. Identification
Purification of neuropeptides from flatworm parasiteshas been quite challenging. The largest impediment hasbeen amassing sufficient quantities of tissue because it isoften difficult to obtain a consistent supply of parasites andthe worms are typically quite small. Also, in comparison tomollusks, nematodes, or arthropods, the levels of peptidesin flatworms are generally much lower, and it is even lowerin the parasitic forms than in the free-living turbellarians.Furthermore, it has been generally impossible to enrich forany particular tissue of these flatworms such that wholeworm extracts, which comprise a large proportion of non-neural tissue, represent the starting point for peptide puri-fication processes.
It is these limitations that have dictated the particularflatworms from which neuropeptides have been isolated andstructurally characterized. Specifically, the only parasiticflatworm from which any neuropeptide has been isolated isthe cestodeM. expansa, a remarkably large worm which cangrow to 6 m in the intestine of sheep. All of the otherneuropeptides identified from flatworms have been fromturbellarians, where tissue is more readily available andneuropeptide levels are higher.
In 1991,M. expansayielded the first neuropeptide to bestructurally characterized from any platyhelminth, the 39-amino acid, PP-immunoreactive, C-terminally amidatedpeptide designated NPF [76]; for the purposes of this re-view, it will be referred to as mxNPF. Subsequently, asimilar 36-amino acid peptide was structurally characterized
from the predatory terrestrial turbellarianArtioposthia tri-angulata(atNPF) [18]. Similar attempts were made to de-termine the structure of the PP-immunoreactive peak fromextracts of the liver flukeF. hepatica, but tissue supply (andavailable technology) proved limiting, and only the first 24amino acids from a PP-immunoreactive high performanceliquid chromatography fraction could be determined [54].That this is the N-terminal 24 residues ofF. hepaticaNPFhas yet to be confirmed.
These flatworm NPFs feature some of the residues thatare highly conserved in the NPY family (Table 2). Thehallmark tertiary structure of NPY family members is thePP-fold, which results from two sets of remarkably con-served residues. First is a series of evenly spaced prolylresidues starting at position 2, creating a PxxPxxP structure(the only known exception is sheep PP, which lacks Pro2).The other critical residues are two tyrosyl residues at posi-tions 20 and 27 (the only known exceptions being seven PPsthat have substituted conservatively Tyr20 with Phe20, andeach of these has a Tyr21). Current hypotheses suggest thatthese structurally constant prolines and tyrosines interact tocreate the signature PP-fold; the two tyrosines interdigitateamong the three evenly spaced prolines, bringing their re-spective helices into an anti-parallel alignment, creating astable fold. NPY family members also feature constancynear the C terminus, with perfectly conserved Arg33 andArg35 and either a Tyr36 or Phe36, therefore terminating inRxRY/Famide.
Both flatworm NPFs have five of the six most highlyconserved residues on the C-terminal half of NPY familypeptides. The RxRY/Famide motif is present as RPRFamidein both flatworm peptides [18,76]. Both also have Tyr res-idues at positions corresponding to 20 and 27 in vertebratefamily members (10 and 17 amino acids from the C termi-nus, respectively), residues that are presumably conservedbecause of their role interacting with N-terminal prolines in
Table 2NPF sequences compared with NPY family members
Structure Ref.
atNPF (Artioposthia triangulata) K V V H L R P R S S F S S E D E Y Q I Y L R N V S K Y I Q L Y G R P R F .NH2 [18]
mxNPF (Moniezia expansa) P D K D F I V N P S D L V L D N K A A L R D Y L R Q I N E Y F A I I G R P R F .NH2 [76]
haNPF (Helix aspersa) S T Q M L S P P E R P R E F R H P N E L R Q Y L K E L N E Y Y A I M G R T R F .NH [50]
apNPY/apNPF (Aplysia californica) D N S E M L A P P PR P E E F T S A Q Q L R Q Y L A A L N E Y Y S I M G R P R F .NH2 [84]
NPF homologies –– – – – – – – – – – – – – – – – – – Y L – – – – – Y – – – – G R – R F .NH2
NPY family homologies – P – – P – – P G – – A– – – – – – – Y – – – – – – Y – – – –T R – R – .NH2
NPY conserved residues –P – K P – – P G – – A P – – – – A – Y – – A – R H Y I N L I T R Q R Y .NH2
PYY conserved residues –P – K P – – P G – – A – P E E – – – Y – – – – R – Y – N L – T R Q R Y .NH2 [47,77]
PP conserved residues –P – – P – – P G – – A – – – – – – –Y – – – – – – Y – – – –T R – R F .NH2
The C terminus of NPFs has four key residues (open boxes) present in NPY family members. Proline residues (black boxes) present in the N-terminal halfof NPYs are also present in NPFs, but their positions are not well conserved. Dashes denote variable residues. Abbreviations initalics have either one knownexception or conservative substitutions.
1003T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

the formation of the PP fold. In general, the C terminus ofthe flatworm peptides most resembles the PPs, as was some-what expected because C-terminally directed PP antiseraproduced the most intense immunostaining. The RPRF-amide terminus of the flatworm peptides is identical toamphibian and reptilian PP, and very similar to many otherPPs that terminate RPRYamide. The flatworm peptides arenot as similar to vertebrate NPY family members in theN-terminal half. Most specifically, the highly conservedPro2, Pro5, and Pro8 are not in place in either NPF—mxNPFhas two prolines in the N-terminal half of the protein andatNPF has only one.
In addition to the discovery of NPF in flatworms, similarpeptides have been found in two mollusks, a 39-amino acidpeptide from the gastropodHelix aspersa(haNPF [50]) anda 40-residue peptide from the sea hareAplysia californica(designated apNPY [84]). These molluskan peptides are likethe flatworm NPFs in that they contain the C-terminal NPYfamily consensus features; apNPY ends RTRFamide, andhaNPF terminates with RPRFamide. However, comparedwith the flatworm peptides, the molluskan peptides containmore of the N-terminal similarities, each with four Proresidues in the N-terminal half, two of which occupy posi-tions conserved in NPY family members.
3.3. Function
Nothing is known about the function of NPF in flat-worms. Hypotheses about function based on the distributionof NPF-IR in flatworms include a possible role in the co-ordination of reproductive processes, somatic musculature,organs of attachment, and alimentation. But, to date, none ofthese hypotheses have been tested. This is due largely to alack of bioassays for any of these specific functions inflatworms and is compounded by the difficulty of deliveringthese polar peptides to the interior of the animal (much lessany particular interior part), which is covered by a mem-branous tegument. To date, dissection has not providedmuch relief from these problems, and the limitation remainsexactly how to test these hypotheses of NPF function inflatworms.
For example, the NPF-IR of nerves terminating on theattachment organs of flatworms leads to the hypothesis thatNPF is involved in controlling the muscle of these organs.However, there is currently no assay capable of testing thishypothesis. It is not feasible to isolate the attachment appa-ratus from the rest of the animal, nor is it presently possibleto record electrophysiological events from any individualcells of the organ. The best one could do is measure theability of whole animals to adhere to a biologically relevanttarget and measure the effects of peptides on their perfor-mance. However, a loss of haptor control could be the resultof peptide effects on sensory structures, central neurons, orother sites in systems indirectly affecting the suckers/hap-tors. The problem is that flatworms are small animals, con-structed of small, tightly packed cells that make dissection
or isolation of specific tissue or cells extremely difficult. Asa result, the involvement of NPF in the control of attach-ment in flatworms remains an implication based on immu-nolocalization.
Localization likewise suggests a role for NPF in control-ling reproductive function, and this possibility is supportedby experimentation in the molluskAplysia, in which apNPYproduces a long-term inhibition of neuroendocrine cells thatinitiate egg laying [84]. At present, there are no reproduc-tive system bioassays in flatworms that could test for adirect effect of NPF in flatworm egg production.
3.4. Genes
The only information available on the genomic and pre-cursor information of a flatworm neuropeptide is that ofmxNPF. Like other NPY family genes, the NPF gene ofM.expansais characteristically small—only 445 bp. Althoughit lacks a C-terminal extension peptide (CPON) present inNPY family members, the mxNPF gene possesses an introncharacteristic of the NPY family. All known NPY familygenes have an intron within the codon for the penultimatearginyl residue, and an intron is also present in this exactlocation in the mxNPF gene, providing strong evidence thatthe flatworm NPF is an ancestor of the NPY family peptides(G.R. Mair, D.W. Halton, C. Shaw, A.G. Maule, unpub-lished data).
3.5. Comparative perspectives
Some difficult evolutionary questions are raised by theearly and apparently extensive establishment of NPF inplatyhelminths, the most ancient of bilateral animals. NPYis the most stable member of the NPY family [47]. Forexample, human, guinea pig, rat, rabbit, and alligator NPYhave complete sequence identity, and NPY from the riverlamprey differs from these at only six residues [102]. Intotal, an exceptional 22 of 36 residues are completely iden-tical in all known NPYs, and many of the 14 others arehighly conserved. In contrast, PP, which has a C terminusmost similar to that of NPF, is the least stable member of theNPY family. Whereas human and reptilian NPY are iden-tical, their PPs differ at 16 positions, indeed, only seven ofthe 36 residues are identical in all known PPs. Sequenceanalysis suggests NPY is the most ancient of NPY familymembers, with PP arising much more recently [48]. In thiscontext, the presence of a PP-like peptide in many earlyinvertebrates was unexpected. Initially, it was suggestedthat the PP-like NPFs do not share a common ancestry withthe NPY family, i.e. NPF and NPY arose completely inde-pendently [48]. However, in addition to the C-terminalhomology and similar size of NPFs and NPY family pep-tides, the ancestral ties between NPF and NPY are sup-ported most convincingly by the occurrence of an intra-arginyl intron in the mxNPF gene, a feature of all knownNPY genes.
1004 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

However, if PP emerged late in vertebrate evolution,how is the PP-like NPF related to these NPY family mem-bers? One possibility is that NPF and NPY shared a com-mon ancestor but that NPF and PP convergently arrived atthe RPRFamide C terminus. In this model, NPF and NPYdiverged from a common ancestor in early invertebrates,and then PP later diverged from NPY within vertebrates.This would suggest that the shared characteristics associatedwith the PP fold (specifically, the conserved tyrosines atpositions 10 and 17 relative to the C terminus) have acommon ancestry but that NPF and many PPs indepen-dently evolved RPRFamide endings. The convergent evo-lution of this C terminus is more palatable than the conver-gent evolution of the C terminus and the PP-fold relatedtyrosyl residues. This model would not conflict with se-quence analysis that suggests that PP diverged from NPYlate in vertebrate evolution.
One other possibility is that PPs did not arise from anNPY gene duplication in the vertebrates but were indepen-dently established from a common ancestor earlier than hadbeen suggested. In this case, the striking similarities be-tween NPFs and PPs could be due to NPF being a directancestor of PP.
3.6. Conclusions
NPF is the most abundant peptide in every flatwormexamined, yet its function remains unknown. It is inviting toassign NPF a role proportional to its abundance, but thisawaits evidence of biological activity. Although the distri-bution of NPF suggests a number of important roles, none isyet confirmed, nor is the NPF site of action.
The occurrence of NPF, an NPY family predecessor, inthe earliest bilaterally symmetrical animals and the apparentstability of these related neuropeptides across vast evolu-tionary time periods suggests that important functional roleswere established early in the evolution of this peptide fam-ily. In mammals, receptors for NPY family peptides areclassified as Y receptors, which are divided into the sub-groups Y1, Y2, Y4, Y5, and Y6 receptors. The Y1, Y4, and Y6
are a related subgroup, and they are less similar to the Y2
subgroup than any class of G protein-coupled receptors thatrespond to the same ligand [77], raising the possibility thatthese two receptor groups are not evolutionarily related. TheNPF/NPY family provide a compelling subject for the studyof the co-evolution of peptide ligands and their receptorsacross great time spans.
The potential of exploiting the NPF signaling system asa target for anthelmintic drugs awaits more data, most im-portantly the characterization of helminth NPF receptors.The NPF system only can be considered a valid chemother-apeutic target when the biological significance of the systemis understood and the selectivity of the parasite NPF recep-tors (as opposed to the mammalian Y receptors) is charac-terized.
4. FMRFamide-related peptides (FaRPs) inplatyhelminths
4.1. Localization
Early reports of peptide occurrence in flatworms in-cluded numerous reports of FMRFamide-IR. In these re-ports, the patterns of FMRFamide-IR are similar to those forNPF-IR, as depicted above. That is, there is extensiveFMRFamide-IR throughout the central and peripheral ner-vous system, including the plexuses associated with thereproductive structures, the somatic musculature, the hold-fast organs, and the alimentary system (for review, see[94]).
Interpretation of FaRP immunostaining in flatworms hasbeen confounded by the discovery of NPF and the subse-quent finding that, under noncompetitive immunocytochem-ical conditions, many FMRFamide antisera cross-react withthe RFamide C terminus of NPF [64,72]. Given the abun-dance of NPF in flatworms, this cross-reactivity clouded thepicture of FaRP distribution in flatworms and even calledinto question the occurrence of FaRPs in flatworms. But,FaRPs are present in flatworms, and their eventual isolationallowed for the production of antisera directed toward gen-uine flatworm FaRPs. Unfortunately, some of these antiseraalso cross-react with NPF, leaving the actual distribution ofFaRPs in flatworms unclear [42].
Despite the lack of specificity of FaRP-directed antisera,the presence of FaRPs in flatworms can be confirmed byradioimmunoassay of acid ethanol extracts separated on thebasis of size. For example, extracts ofF. hepaticareveal aFaRP immunoreactive peak that co-elutes with syntheticFaRPs, in addition to a peak that co-elutes with syntheticNPF [57]. These radioimmunoassay studies have provided afairly definitive demonstration of FaRPs in many worms inwhich the structural characterization of the peptides has notbeen possible and also allowed for a quantification of FaRPsin a number of worms. As mentioned previously, free-livingflatworms tend to have greater levels of NPF than do par-asitic worms. Such is also the case with FaRPs. The levelsof FaRPs in free-living flatworms appear to be 100–5000times greater than in trematodes [40,57], as compared witha 100-fold difference for NPF between the classes. As notedwith NPF, this difference could be explained in large part bythe fact that the nervous systems of turbellarians are moresubstantial than those of parasitic flatworms, possibly as aresult of a reduction of the nervous system as the parasiticworms adopted a more sedentary lifestyle [40]. The ampli-fication of this difference in the case of FaRPs would beexplained if they prove to play a primary role in controllingsomatic musculature, as some evidence suggests. Anotherobservation is that NPF is usually more abundant thanFaRPs in any particular worm, e.g.D. merlangipossesses20-fold more NPF that FaRP-IR. But, in most species, thedifference is not as dramatic and two turbellarians—Den-
1005T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

drocoelum lacteumand Polycelis nigra—actually have ahigher concentration of FaRPs than NPF [40].
4.3. Identification
As mentioned previously, tissue availability has limitedthe isolation and structural characterization of neuropep-tides from flatworms. As with NPF, the large cestodeM.expansayielded the first FaRP sequence, and all the otherstructurally identified FaRPs are from the class Turbellaria.
The first FaRP to be isolated and sequenced from flatwormswas GNFFRFamide from the cestodeM. expansa[75]. Un-usual is the Phe occupying the position preceding the C-terminal RFamide, where Met, Leu, or Ile usually is found. Apeptide with this feature has been discovered in the molluskMytilus edulis (ALAGDHFFRFamide [105]) and has beenpredicted by gene analysis in the molluskLymnaea(SDPFFRFamide [52]) and in the nematodeC. elegans(EIVFHQISPIFFRFamide [79]). So, although atypical, the cestodepeptide is not unique in this respect. Continued analysis ofextracts have failed to yield other short RFamides, leavingthe possibility that this rather unusual FaRP is the solerepresentative in this cestode. FaRP structures have not beendetermined from any other cestode, leaving it unclear as towhether the presence of a Phe preceding the C-terminalRFamide (or another aromatic residue) is a feature of allcestode FaRPs or an anomaly ofMoniezia.
Subsequently, all structurally identified FaRPs havecome from turbellarians, and all are more similar to typicalmolluskan FaRPs than to GNFFRFamide (Table 3). Theland planarianA. triangulata, a turbellarian that preys onearthworms, was found to possess RYIRFamide [73]. Sub-sequently, GYIRFamide was characterized from the fresh-water planarianD. tigrina and the ectocommensal turbel-larian found on the horseshoe crabBdelloura candida; thelatter also possessed YIRFamide [41,42]. The common fea-ture of the turbellarian FaRPs is the YIRFamide motif. Ofinterest, three of the species from which FaRPs have beenidentified apparently have only a single FaRP, and the otherhas only two detectable FaRPs. Although these two FaRPswere detected, it is not known if YIRFamide is simplyN-terminally truncated GYIRFamide generated by enzymeaction on the parent peptide.
4.4. Genes
There is no published information regarding the nature ofFaRP-encoding genes of platyhelminths, in contrast to therapidly accumulating data regarding the FaRP-encodinggenes of nematodes. The reasons for this disparity are two-fold. First, theC. elegansgenome sequencing project hasstimulated FaRP gene detection in nematodes and, second,there appears to have been no concentrated efforts to deter-mine flatworm FaRP-gene structure.
4.5. Function
Whereas the earliest discoveries of FaRPs in molluskswere based on their biological activity, flatworm neuropep-tides were immunolocalized and structurally identified be-fore functional characterization. Therefore, the task was todiscover the biological activities of peptides that alreadyhad been localized and identified. With clues about functionbased on localization and the actual peptides in hand, it mayhave seemed an easy task to identify their biological activ-ities. However, physiological assessment has been difficult,mostly because of a lack of definitive cell-, tissue-, or organ-specific bioassays in flatworms. Nevertheless, there is indirectevidence that FaRPs play a role in reproductive function anddirect evidence for a role in somatic muscle control.
There is intense FaRP-IR in the nerve plexuses of themale and female reproductive structures of flatworms. In-nervation of the female structures, including the muscularegg chamber, is predominantly peptidergic. In addition tothis inference by localization, in one species there is atemporal association between the presence of FaRP-IR inreproductive system innervation and reproductive activity.The monogeneanPolystoma nearcticumlives in the urinarybladder of the gray treefrogHyla versicolor, and the parasiteis reproductively active only when the host frog is spawn-ing. The nerves innervating the egg chamber express FaRPsduring the brief periods of egg production, but FaRPs arenot expressed in those same neurons in reproductively qui-escent periods [2]. In contrast, there is apparently no vari-ation in the serotonergic component of the egg chamberinnervation throughout the cycle.
There is direct evidence implicating FaRPs in the controlof flatworm somatic musculature. Flatworm-derived FaRPsare myoexcitatory, not only on nerve-containing muscle-strip preparations but also on nerve-free, individual musclesisolated from a number of flatworms (Table 4). Until re-cently, studies of flatworm muscle physiology were limitedto monitoring tension in preparations of muscle strips orintact worms or some other nerve-containing preparation.These preparations limited interpretation because a changein muscle tension induced by some exogenous agent couldbe attributable to a number of sites of action—on the mus-cle itself or on motor, central, or even sensory neurons. Therecent development of an isolated muscle fiber preparationfrom flatworms has allowed for the testing of putative myo-
Table 3FaRPs isolated from flatworms
Species Structure Ref.
CestodaMoniezia expansa G N F F R F .NH2 [75]
TurbellariaArtioposthia triangulata R Y I R F .NH2 [73]Bdelloura candida G Y I R F .NH2 [42]
Y I R F .NH2
Dugesia tigrina G Y I R F .NH2 [41]
1006 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

active agents directly on muscle itself, without interferencefrom other tissues. The isolated muscle fiber preparationalso relieves difficulties associated with the penetration ofpolar agents through the tegument to the underlying musclelayers. Flatworm-derived FaRPs potently elicit contractionof muscle fibers isolated from the trematodeS. mansoni[22]and the turbellariansB. candida[42] andP. littoralis (Mon-eypenny, personal communication). The potent and specificaction of FaRPs on nerve-free muscle fibers localizes onesite of FaRP action to the muscle itself. Additionally, FaRPshave been shown to increase muscle tension in muscle stripsfrom a number of platyhelminths.
Results from both the muscle strip and isolated musclestudies show that the three YIRFamide-containing FaRPsisolated from turbellarians are all similar in potency on theturbellarian and trematode preparations, but the structurallydissimilar GNFFRFamide from the cestode is much lesspotent. For example, the EC50 for the contraction of isolatedschistosome muscle is between 1 and 7 nm for YIRFamide,RYIRFamide, and GYIRFamide, while the EC50 forGNFFRFamide is 500 nm [22]. In contrast, the converse istrue for the only tapeworm muscle preparation that has beentested—in M. expansamuscle strips, GNFFRFamide ismore potent than the YIRFamide-containing peptides (un-published observations). By using synthetic analogs of flat-worm FaRPs on isolated schistosome muscles, the specific-ity of the response has been demonstrated [23]. The Alasubstitution of any single amino acid of YIRFamide yieldsan inactive peptide. However, conservative substitutions atTyr1 (an aromatic residue) or Ile2 (a hydrophobic residue)produce peptides similar in potency with YIRFamide. Onthe other hand, substitutions in the C-terminal RFamide arenot well tolerated, with most causing a complete loss ofactivity. The potent, specific, and immediate action ofFaRPs on flatworm muscle suggest that they are acting asfast transmitters rather than as modulators (orhormones, anirrelevant term in these animals without a circulatory sys-tem). FaRPs serving as a fast transmitter in these early
animals supports the view that these peptides originated astransmitters and later filled the role of modulators [83].
4.6. Comparative perspectives
The original discovery of FMRFamide in molluskscaused speculation that FMRFamide could be the archetypalFaRP. However, the absence of authentic FMRFamide andthe presence of YIRFamide peptides in flatworms demon-strate that this is not the case. It now appears as thoughxYIRFamide may be the earliest FaRP. It previously hasbeen suggested that FaRPs should be defined by the pres-ence of a C-terminal RFamide, a hydrophobic residue inposition 3 (relative to the C terminus) and an aromaticresidue in position 4 [41]. By that definition, the flatwormpeptides are authentic FaRPs.
The limited structure activity studies suggest that flat-worm FaRP receptors are similar to those of mollusks. Bothflatworm and mollusk receptors share a relative tolerancefor peptides with conservative substitutions in positions 3and 4 from the C terminus but are rather intolerant ofchanges in the C-terminal RFamide, including the amidemoiety.Helix receptors prefer Phe1 over Tyr1 [82], and mostmolluskan tetrapeptides have Phe in this position, whereasthe only flatworm receptor characterized prefers Tyr1 overPhe1 [23], and all of the known turbellarian FaRPs have Tyrin this position.
4.7. Peptide metabolism
There have been no studies directly examining the me-tabolism of FaRPs in flatworms. However, it recently wasdemonstrated that the general metalloprotease inhibitor1,10-phenanthroline induces an immediate excitation ofschistosome muscle strips, in a manner indistinguishablefrom the application of exogenous FaRPs [21]. The proteaseinhibitor induces myoexcitation only in innervated musclestrips and not in nerve-free muscle fibers. One possibleexplanation is that metalloproteases are involved in the
Table 4FaRP activity on flatworm muscle
Species Preparation Order of potency Threshold ofactivity (M)
Ref.
CestodaMoniezia expansa Muscle strips GNFFRFa. GYIRFa 5 YIRFa Unpublished
TrematodaSchistosoma mansoni Isolated muscle cells RYIRFa5 YIRFa 5 GYIRFa .. GNFFRFa 10210 [22]Schistosoma mansoni Muscle strips RYIRFa5 YIRFa 5 GYIRFa .. GNFFRFa 1027 UnpublishedFasciola hepatica Muscle strips RYIRFa5 YIRFa 5 GYIRFa ne: GNFFRFa 1027 [32]Fasciola hepatica Trimmed worms RYIRFa. YIRFa 5 GYIRFa .. GNFFRFa 1029 [58]Diclidophora merlangi Trimmed worms YIRFa. RYIRFa 5 GYIRFa ne: GNFFRFa 1028 [78]
TurbellariaBdelloura candida Isolated muscle cells GYIRFa. YYIRFa 5 RYIRFa 1029 [42]Procerodes littoralis Isolated muscle cells GYIRFa. YIRFa .. GNFFRFa 1029 Unpublished
ne, not effective; a, the amidated C terminus.
1007T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

degradation of endogenous FaRP transmitters, and the in-hibition of these metalloproteases leads to the persistentaction of tonically released myoexcitatory FaRPs. This ex-planation is supported by the observation that 1,10-phenan-throline blocks the degradation of synthetic YIRFamide byschistosome tissue (unpublished observation). Direct stud-ies of FaRP degradation in flatworms are needed to explorethis possibility.
4.8. Conclusions
From the meager data accumulating regarding FaRPsin flatworms, some patterns are emerging. First, eachspecies appears to harbor a small number of unique FaRPs,one or two in each case to date. This stands in stark contrastto the situation in nematodes in which each species appearsto have large numbers of unique peptides. In addition tothis, the flatworms have a lower level of FaRPs than doanimals of other phyla. Furthermore, the parasitic flatwormshave even lower levels of FaRPs than the free-living flat-worms. This could be attributable to the differences inlifestyles and the apparent role of FaRPs in the control ofsomatic muscle.
The localization of FaRPs in flatworms implies that theycould have a function in the control of holdfast organs,reproduction, and somatic muscle control, among otherthings. Of these, there is only direct evidence for a role insomatic muscle control. All of the observed actions ofFaRPs in flatworms are excitatory, different from the situ-ation in nematodes, in which both excitation and inhibitionoccur.
Preliminary evidence suggests that FaRPs in the twomain classes of parasitic platyhelminths might be quitedifferent because the only primary structure from cestodesis somewhat atypical and is not very active in trematodes.However, this apparent difference between the cestodes andtrematodes is based on a single peptide structure from ces-todes and physiological inference in trematodes, wherethere are still no primary structures available.
The critical nature of the functions in which FaRPs areimplicated provokes the interest of parasitologists. In par-ticular, the myoactivity of FaRPs in flatworms demandsattention because the neuromuscular system has been afertile source of anthelmintic targets. Most anti-trematodalsand anti-cestodals have dramatic effects on the somaticmusculature. Also, interference with reproductive functioncould break the life cycle of many parasites, and it couldalso limit the pathology associated with egg production.
5. FMRFamide-related peptides (FaRPs) in nematodes
5.1. Localization
The first reports of peptide-IR in the nervous system ofnematodes occurred in the late 1980s and triggered a num-ber of immunocytochemical studies into peptide occurrence
and distribution (Table 5). Evident from these early studieswas the abundance of FaRP-immunostaining in the nervoussystems of nematodes [3,19,49,51]. Unfortunately, the vastmajority of immunocytochemical studies of the FaRPergicnervous systems of nematodes employ antisera that cannotdiscriminate between members of this peptide family. Ingeneral, the presence of FaRP-IR has confirmed the occur-rence of peptides that terminate in an RFamide motif. Lo-calization studies in the late 1980s were unable to focus onspecific nematode FaRPs because of the absence of primarystructural information. However, even following the avail-ability of such data, most polyclonal antisera could notdistinguish between closely related members of this peptidefamily. This is a particular problem in nematodes because ofthe occurrence of numerous structurally similar FaRPs thatdefy discrimination by using polyclonal antisera and non-competitive immunochemical conditions.
A key feature of the nematode nervous system is itsstructural conformity between species and, unlike platyhel-minths, its relatively simple organization. In this respect,nematodes generally possess in the order of 300 neuronsthat are discretely arranged into ganglia associated with theanterior (brain, circumpharyngeal) and posterior (perianal)nerve rings and longitudinal nerve cords. The motor neuroncell bodies are arranged in the ganglia and along the ventralnerve cord such that the dorsal nerve cord comprises den-dritic branches from the ventral nerve cord. All of theneurons in the free-living nematodeC. eleganshave beenmapped by using electron microscopy [108], and studies onother species have revealed similar neuronal architecture.
Localization studies have demonstrated a wide distribu-tion of FaRP-immunopositive neurons in parasitic and free-living nematodes [3,95]. However, there are conflicting dataon the abundance of FaRP neurons in different nematodespecies, with reports that between 60% [14] and 75% [7] ofthe neurons of the large pig nematodeA. suumare FaRP-immunoreactive whereas only 10% of the neurons in thefree-living nematodesC. elegansandC. vulgarisare FaRP-immunoreactive [92,93]. Although these differences may be
Table 5FaRP immunoreactivity in nematodes
Species Ref.
Ascaris suum [19]Brugia pahangi [106]Caenorhabditis elegans [3]Caenorhabditis vulgaris [92]Cystidicola farionis [109]Dirofilaria immitis [106]Goodeyus ulmi [49]Haemonchus contortus [43]Heterodera glycines [3]Onchocerca volvulus [29]Panagrellus redivivus [3]Romanomermis culicivorax [39]Trichostrongylus colubriformis [29]
1008 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019
genuine, they could simply reflect differential cross-reactivitybetween the antisera used and the native nematode peptides.
The most detailed studies of FaRP distribution in nem-atodes have been carried out onA. suumand have shownclearly that FaRP-IR occurs throughout various componentsof the nervous system, including the longitudinal nervecords (ventral, dorsal, lateral, and sublateral), cephalic pap-illary neurons, circumpharyngeal nerve ring and associatedganglia, retrovesicular ganglion, sensory neurons, tail gan-glia, and vaginal, rectal, and pharyngeal neurons [8–10,14,95].
Recent studies have identified a large number (ref. [18];see below) of FaRP-encoding genes, designatedflp-1 toflp-18, in C. elegans[79]. Polymerase chain reaction tech-niques have confirmed that the first 14 of these genes aretranscribed in the worm [79]. The expression pattern of theC. elegans flp-1 gene was determined by using transgenicanimals that hadlacZ or green fluorescent protein con-structs. Expression was localized to a series of anteriorneurons including: AVK, AVA, AVE, RIG, RMG, AIY,AIA, and M5, all of which had been shown previously to beFaRP-immunopositive [80,92].
All of the major neuronal types in nematodes have beenidentified as FaRP-immunopositive. Indeed, the widespreaddistribution of FaRP-immunoreactive nerves in nematodeshas implicated a range of important roles for these messen-ger molecules, including: central motor function, sensoryperception, pharyngeal pumping, and the regulation of peri-
staltic activity in reproductive ducts. However, direct evi-dence for FaRP involvement in these activities is onlyavailable in a few cases (see below).
5.2. Identification
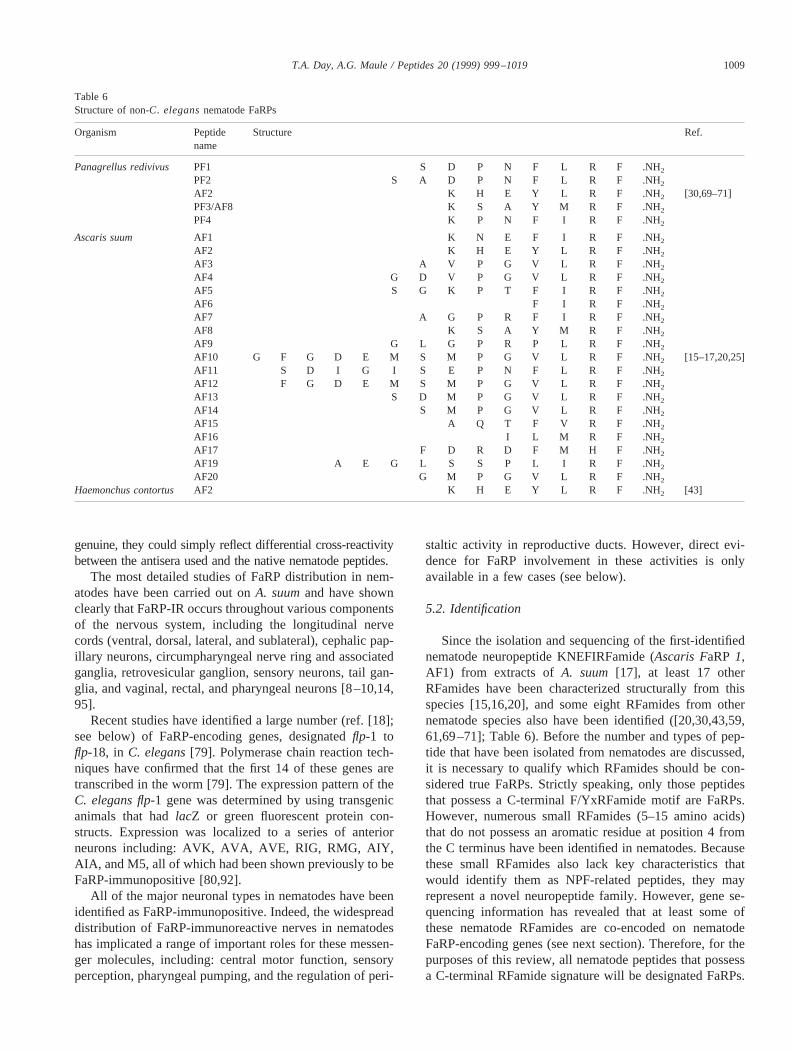
Since the isolation and sequencing of the first-identifiednematode neuropeptide KNEFIRFamide (Ascaris FaRP1,AF1) from extracts ofA. suum [17], at least 17 otherRFamides have been characterized structurally from thisspecies [15,16,20], and some eight RFamides from othernematode species also have been identified ([20,30,43,59,61,69–71]; Table 6). Before the number and types of pep-tide that have been isolated from nematodes are discussed,it is necessary to qualify which RFamides should be con-sidered true FaRPs. Strictly speaking, only those peptidesthat possess a C-terminal F/YxRFamide motif are FaRPs.However, numerous small RFamides (5–15 amino acids)that do not possess an aromatic residue at position 4 fromthe C terminus have been identified in nematodes. Becausethese small RFamides also lack key characteristics thatwould identify them as NPF-related peptides, they mayrepresent a novel neuropeptide family. However, gene se-quencing information has revealed that at least some ofthese nematode RFamides are co-encoded on nematodeFaRP-encoding genes (see next section). Therefore, for thepurposes of this review, all nematode peptides that possessa C-terminal RFamide signature will be designated FaRPs.
Table 6Structure of non-C. elegansnematode FaRPs
Organism Peptidename
Structure Ref.
Panagrellus redivivus PF1 S D P N F L R F .NH2PF2 S A D P N F L R F .NH2AF2 K H E Y L R F .NH2 [30,69–71]PF3/AF8 K S A Y M R F .NH2
PF4 K P N F I R F .NH2
Ascaris suum AF1 K N E F I R F .NH2
AF2 K H E Y L R F .NH2
AF3 A V P G V L R F .NH2
AF4 G D V P G V L R F .NH2
AF5 S G K P T F I R F .NH2AF6 F I R F .NH2
AF7 A G P R F I R F .NH2
AF8 K S A Y M R F .NH2
AF9 G L G P R P L R F .NH2AF10 G F G D E M S M P G V L R F .NH2 [15–17,20,25]AF11 S D I G I S E P N F L R F .NH2AF12 F G D E M S M P G V L R F .NH2AF13 S D M P G V L R F .NH2
AF14 S M P G V L R F .NH2
AF15 A Q T F V R F .NH2
AF16 I L M R F .NH2
AF17 F D R D F M H F .NH2
AF19 A E G L S S P L I R F .NH2AF20 G M P G V L R F .NH2
Haemonchus contortus AF2 K H E Y L R F .NH2 [43]
1009T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

Furthermore, a single peptide that possesses a C-terminalRYamide is co-encoded with RFamides on a singleC.elegansgene (see below); this RYamide will also be con-sidered a FaRP.
All of the 18 RFamides that have been fully structurallycharacterized fromA. suumwere extracted from head and/ortail tissue by using acid ethanol and were purified chromato-graphically (see [20]). A number of other peptides havebeen partially structurally characterized, including FaRPsthat terminate in PNFLRFamide and one peptide that ter-minates in HFamide and is not included in the FaRP cate-gory [20]. It is also known that there are otherA. suumpeptides that terminate in RFamide and that await structuralcharacterization.
A number of peptides have been structurally character-ized from other nematode species, including the free-livingnematodesC. elegans (SDPNFLRFamide [CF1, PF1],SADPNFLRFamide [CF2,PF2], SQPNFLRFamide [CF3],ASGDPNFLRFamide [CF4], AAADPNFLRFamide [CF5],KPNFLRFG [CF62], AGSDPNFLRFamide [CF7], KHEY
LRFamide [CF8, AF2], APEASPFIRFamide [CF9],KSAYMRFamide [CF10,PF3], KPSFVRFamide [CF11],KNEFIRFamide [CF12,AF1], RNKFEFIRFamide [CF13],HFYNFSSESRKPNFLRFamide [CF14] [20,59,61,88] andPanagrellus redivivus(SDPNFLRFamide [PF1], SADPNFLRFamide [PF2], KSAYMRFamide [PF3], KPNFIRFamide [PF4] and KHEYLRFamide [PF5,AF2]) [30,69–71]and the sheep parasite,Haemonchus contortus(KHEYLRFamide [HF1,AF2]) [43]. Although there is remarkablediversity in the neuropeptide complements within and be-tween nematode species, several peptides have been iden-tified in more than one species. These include PF3, whichhas been isolated fromA. suum, P. redivivus, and C. el-egans, and AF2, which has been isolated fromA. suum, C.elegans, H. contortus, andP. redivivus. Of interest, AF2 isthe most abundant peptide identified in acid ethanol/meth-anol extracts of all of these species [15,43,61,69].
5.3. Function
Although over 60 nematode FaRPs have been reported inthe literature, the actions of only a few of them have beenstudied in any detail. The vast majority of data on theactions of nematode peptides have been generated by usingisometric tension recordings from muscle strip preparationsand electrophysiological recordings of body wall muscleand selected motor neurons ofA. suum. The main findingsare outlined below (see Table 7).
1 Most of the nematode peptide abbreviations have been given previ-ously, however, where they were not previously assigned, they have beenappended a number chronologically upon discovery or, if reported simul-taneously, then according to their appearance on an encoding gene. Orig-inal nomenclature appears in bold and will be employed in subsequentcitations.
2 CF6 was not fully structurally characterized and it is unclear if itoccurs as a heptapeptide (KPNFLRFamide) or a hexapeptide (PNFLRFamide).
Table 7Effects of nematode FaRPs onAscarisneuromusculature
Peptide name and sequence Effects on body wall muscle Effects on vagina vera
AF1KNEFIRF.NH2
Biphasic effects ($1 nM)—relaxation followed byincreased contractility; 1–2 mV muscle depolarization;abolishes slow membrane potentials and increasesinput conductances of V1 and D1 inhibitory neurons
Biphasic effects ($10 nM)—profound contractionfollowed by period of relaxation
AF2KHEYLRF.NH2
Similar biphasic effects to AF1 but more potent ($1pM); potentiates ACh-induced contraction andamplitude of excitatory junction potentials; stimulatescAMP levels
Reduces contraction frequency ($0.1 mM)
PF1SDPNFLRF.NH2
Slow Ca21-dependent, nerve cord-independent relaxation($1 nM); decreases amplitude of excitatory junctionpotentials; effects involve nitric oxide; dose-dependentmuscle hyperpolarization (#12 mV)
Induces a transient flaccid paralysis ($3 nM)
PF2SADPNFLRF.NH2
Similar effects to PF1 Transient flaccid paralysis ($1 nM)
PF3 (AF8)KSAYMRF.NH2
Nerve cord-dependent contraction of ventral ($0.1 mM)and relaxation of dorsal muscle strips ($1 mM)
Reduces contraction frequency and amplitude($0.1 mM)
PF4KPNFIRF.NH2
Rapid Cl2-dependent, nerve cord-independent relaxation($1 nM); dose-dependent muscle hyperpolarization(4–6 mV) and increase in input conductance
Induces flaccid paralysis ($10 nM)
AF3AVPGVLRF.NH2
Contraction of dorsal strips (0.1mM); increasessensitivity of muscle to ACh but action is independentof ACh receptor; decreases cAMP levels
Multi-phasic effects ($1 mM)—transient contractionthan relaxation and return of contractile activity;stimulates contractility in inactive preparations
AF4GDVPGVLRF.NH2
Contraction of dorsal strips (0.1mM) Similar effects to AF3 but less potent (3 nM)
Refs. (5,6,15–17,27,34,35,65,81, Fellows personal communication).
1010 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

6. Lys1 heptapeptides
6.1. KNEFIRFamide (AF1) and KHEYLRFamide (AF2)
The actions of these two peptides are very similar and,therefore, will be discussed together. AF1 and AF2 haveexcitatory effects onA. suummuscle strips [5,17,65]. Theiractions comprise an initial transient relaxation phase fol-lowed by an extended period of muscle excitation. Thethreshold concentration for the AF2 effect has been reportedas 1 nM and is several orders of magnitude lower than thatfor AF1 [5]. Of interest, removal of the nerve cords from themuscle strip preparations abolishes the excitatory phase ofthe response such that only the inhibitory phase is observed[65]. This finding suggests that different receptors mediatethe two phases of the AF1 and AF2 response; one is muscle-based and mediates the inhibitory response, the other isneuronally based and mediates the excitatory response.Electrophysiological studies have shown that AF1 inhibitsboth dorsal and ventral inhibitory neurons inA. suumandthat it induces an increase in input conductance consistentwith the opening of channels in the neuronal membranes[17]. Furthermore, AF2 was shown to increase the ampli-tude of excitatory junction potentials induced by acetylcho-line (ACh) [81]. Also, the nematode nicotinic ACh receptorantagonist mecamylamine inhibited the oscillatory activityinduced by AF2. The mechanisms behind the inhibitoryphase and the stimulation of oscillatory activity remainunknown.
Structure–activity studies carried out on AF1 haveshown that both the N and C termini are necessary forobserved biological activity onA. suum[5]. Intriguing tonote, structure-activity studies that used an alanine-scanseries of this peptide identified two analogs (KNAFIRFamide and KNEFARFamide) that were only inhibitory, i.e.they induced the initial inhibitory phase but did not stimu-late oscillatory activity in innervated muscle strip prepara-tions. These findings further support the hypothesis thatthere are at least two different receptors for AF1 (and,therefore, AF2) and that KNAFIRFamide and KNEFARFamide are only biologically active following interactionwith the muscle-based inhibitory receptor. Other studieshave examined the activity of AF1–AF2 chimaeras, includ-ing KHEFIRF amide, KNEYIRFamide, and KNEFLRFamide. These peptides had either a similar or reduced po-tency to that of AF1 such that none was as potent as AF2.Furthermore, although the analog KNEFIAFamide wasfound to antagonize the actions of AF1, it did not antagonizethe actions of AF2. These data provide further support forthe presence of distinct receptors for AF1 and AF2 althoughreceptor binding assays will be required to provide conclu-sive proof.
The effects of both AF1 and AF2 have been examined onthe vagina vera (distal part of the vagina) ofA. suum[27].Unlike the body wall muscle, which comprises only longi-tudinally arranged muscle, the vagina possesses mainly cir-
cular muscle bands. In nematodes, the vagina must coordi-nate the expulsion of eggs (up to 20 000/day inA. suum) andthe uptake of sperm while maintaining the high internalhydrostatic pressure that, if allowed, would force the expul-sion of the internal organs of the worm. AF1 and AF2 hadqualitatively different effects on the vagina vera in thatalthough both inhibited spontaneous rhythmicity (unliketheir effects on body wall muscle), only AF1 had a biphasiceffect that comprised an initial lengthening (the result ofcircular muscle contraction) and a subsequent shortening(circular muscle relaxation) of the vagina [27].
6.2. KSAYMRFamide (PF3)
PF3 has been characterized structurally from bothA.suumandP. redivivus, and inA. suummuscle strips, it hasbeen found to contract ventral muscle and relax dorsalmuscle [65]. This is the first peptide that has been found tohave differential effects on dorsal and ventral muscle stripsof nematodes. The effects on both ventral and dorsal bodywall strips were concentration-dependent, reversible, andnerve-cord-dependent such that denervated strips did notrespond to the peptide. The biology behind the regionaldifferences in response to PF3 is unclear. To date, no analogseries of this peptide has been examined.
Studies on levamisole-resistant strains of the sheep par-asiteHaemonchus contortusfound that there was a similarreduction in sensitivity to levamisole and to PF3 [91]. Le-vamisole is an anthelmintic that is known to act on thenicotinic ACh receptor of nematode musculature [62].Therefore, a reduction in sensitivity to both levamisole andPF3 suggests that PF3 modulates the cholinergic system inthis worm. Whether this reduced sensitivity is due to a directaction of PF3 on the levamisole receptor or to the alterationof ACh release from nerve terminals is unknown. However,as the actions of this peptide are nerve-cord-dependent inA.suum, it is possible that at least part of its activity is throughthe modulation of cholinergic neurons. Other actions of PF3in A. suuminclude the relaxation and inhibiton of vaginavera contractility [27] and an inhibiton of pharyngeal pump-ing [12].
6.3. KPNFIRFamide (PF4)
This peptide has been characterized structurally from thefree-living nematodeP. redivivus[71]. Nevertheless, it hasprofound inhibitory effects on the somatic body wall muscleof A. suum. PF4 induces a rapid, concentration-dependent,nerve-cord-independent, and reversible relaxation of bodywall muscle [65,71]. The relaxation is reversed to a con-traction if muscle strips are bathed in Cl2-free media sug-gesting that Cl2 channels mediate the PF4 effect. The rapidrelaxation is followed by a slow recovery to a plateau level.Electrophysiological studies revealed that PF4 induces amuscle cell hyperpolarization and an increase in input con-ductance [65]. The role of chloride in the PF4-induced
1011T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

relaxations ofA. suumbody wall muscle subsequently wasconfirmed by Holden–Dye et al. [34], who also showed thatthe PF4-induced current has a reversal potential correspond-ing to the equilibrium potential for Cl2, as does theg-ami-nobutyric acid (GABA)-induced current of nematode mus-cle. However, the PF4-induced relaxation is not sensitive tothe nematode GABA channel blocker NCS 281–93 [65],suggesting that PF4 acts on a Cl2 channel other than theGABA-gated channel. Although the evidence indicates thatGABA and PF4 have different sites of action, there may bemultiple GABA receptor subtypes, some of which are in-sensitive to NCS 281-93, and some of these may be mod-ulated by PF4. Only after functional characterization of allof the GABA and PF4 receptors will the possible interactionof PF4 with GABA receptors be understood. Further com-plexity to the PF4 effects is evident from studies on therecovery phase of its action. The recovery phase, whichoccurs after the initial relaxation, is not Cl2-dependent [34],and it remains unknown whether this is a true component ofthe PF4 effect or the result of interaction with other recep-tors in the body wall musculature.
Structure–activity studies on a number of analogs thatdiffered from PF4 in positions 2 and 5 identified peptideswith similar (CF6, KPNFLRFamide) or reduced (KANFIRFamide, KANFLRFamide, KGNFLRFamide, KGNFIRFamide) activity when compared with the parent peptide [46].These studies showed that the replacement of the prolylresidue in position 2 of the parent peptide was detrimental topeptide biological activity.
7. Octapeptides, nonapeptides, and decapeptides
7.1. SDPNFLRFamide (PF1) and SADPNFLRFamide(PF2)
PF1 and PF2 were originally isolated fromP. redivivus[30] and subsequently were identified inC. elegans[87,88].Although, to date, they have not been identified in extractsof A. suum, they have been found to have profound actionson somatic body wall musculature [6,28,35,65]. These pep-tides have similar concentration-dependent and nerve-cord-independent inhibitory actions on muscle strip preparationsof A. suum. Unlike the PF4 effect, the PF1/2 relaxation istypically slow in onset and induces a flaccid paralysis, withno evidence of a recovery phase. The PF1 relaxation hasbeen found to antagonize the contractile actions of ACh, thenematode nicotinic agonist levamisole, and the excitatorypeptides AF1 and AF2. Electrophysiological analysis ofbody wall muscles during treatment with PF1 identified asmall (2–4 mV) hyperpolarization with no significant alter-ation in input resistance.
A series of experiments has implicated nitric oxide (NO)involvement in the PF1-induced relaxation of nematodemusculature [6]. A number of antagonists of nitric oxidesynthase (NOS), the enzyme responsible for the generation
of NO, have been found to blunt the PF1 relaxation. Fur-thermore, the NO donor sodium nitroprusside was found toinduce a relaxation in body wall muscle strips that wasqualitatively similar to that caused by PF1 [6]. Ca21-depen-dent NOS activity also has been identified in the hypoder-mal tissues ofA. suumand in whole body extracts ofP.redivivus. Receptor binding analyses also localized [3H]PF1to hypodermal tissues ofA. suum. The hypodermis of nem-atodes surrounds the somatic body wall muscle layer and isalso intimately associated with the nerve cords. It seemsfeasible that the release of NO from the hypodermal layercould have direct actions on an effector system in the bodywall musculature such that a signaling system, analogous tothat which occurs in the vascular epithelium of mammals,could operate in nematodes. Although the localization ofNADPH diaphorase staining to the worm’s muscle andneuronal tissues [4] does not support the involvement of thehypodermal layer in this signalling pathway, the involve-ment of NO does seem likely.
Examination of truncated analogs of PF1 found that thetwo N-terminal residues (SD) were not essential for activitywhereas the C-terminal hexapeptide-amide (PNFLRFamide) was required [29]. Coupled with an alanine-scanseries of PF1, structure activity studies revealed that the twoaromatic residues (Phe5 and Phe8) as well as the penultimateresidue (Arg7) were essential for biological activity [29].
7.2. AVPGVLRFamide (AF3) and GDVPGVLRFamide(AF4)
AF3 and AF4 have been isolated from extracts ofA.suumand were found to induce a concentration-dependentcontraction of dorsal body wall muscle strips [16]. Electro-physiological analysis of their effects identified a small(2.3–3.7 mV) depolarization that was insensitive to the AChantagonist mecamylamine [104]. It has been reported thatthe other fourA. suumpeptides with C-terminal PGVLRFamide all increase muscle tension [20]. Therefore, the de-polarization induced by AF3 and AF4 is not due to theirinteraction with the muscle nicotinic receptor, and theiractions are clearly distinguishable from the excitatory ac-tions of AF1 and AF2 (see above). These peptides also werefound to induce a slow depolarization in the body wallmuscle of the related parasiteAscaridia galli [104].
7.3. APEASPFIRFamide (CF9)
CF9 was isolated from extracts of the free-living nema-tode C. elegansand has been found to induce slow, con-centration-dependent, and nerve cord-independent relax-ations of both dorsal and ventral body wall musculature ofA. suum[59]. The relaxation was Ca21- and Cl2-indepen-dent but was abolished in high K1 media such that it couldbe distinguished from the actions of PF1, PF2, and PF4 (seeabove). Electrophysiological recordings of its effects onA.suumbody wall musculature have not been published.
1012 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

7.4. RFamides from other phyla
The molluskan peptide FMRFamide was found to inhibitthe motility of P. redivivusas indicated by a sedimentationbioassay [100]. In a few instances, the effects of non-nematode FaRPs onA. suummuscle strips have been tested.For example, the arthropod peptides SDRNFLRFamide(Lobster peptide I), TNRNFLRFamide (Lobster peptide II),and pEDVDHVFLRFamide (Leucomyosuppressin) all haveprofound inhibitory effects on somatic muscle strips ofA.suum (see [63]). The relaxation induced by all three isqualitatively similar to that produced by PF1. Although anumber of flatworm FaRPs (YIRFamide, GYIRFamide,RYIRFamide, and GNFFRFamide) have been tested foractivity on A. suummusculature, none has been found toinduce significant effects at#10mm. Conversely, a numberof nematode peptides have been found to have potent exci-tatory effects on flatworm muscle activity [60]. This sug-gests that nematode FaRP receptors are more discriminatorythan those of flatworms; the reasons why this might be soare discussed below.
8. Genes
The first FaRP-encoding gene to be identified and char-acterized was theFMRFamide-like peptide(flp)-1 genefrom C. elegans[87]. In this study, degenerate oligonucle-otide primers to the amino acid sequence FMRFGK/R wereused to screen a cDNA library ofC. elegans.Subsequently,the entire gene was cloned from a genomic library ofC.elegans. The flp-1 gene was found to encode a series ofrelated FaRPs that terminated in PNFLRFG and a singlepeptide that terminated in PNFMRYG (the G representing aputative amidation site for these products). Two 39 spliceacceptor sites were identified such that 2 transcripts couldbe generated; transcript A encodes eight peptides includingtwo copies of PF2 (SADPNFLRFG) whereas transcript Bencodes single copies of the seven peptides encoded ontranscript A and an extra decapeptide. This gene was char-acterized around the same time that PF1 and PF2 werecharacterized structurally fromP. redivivusand indicatedhomology in the neuropeptides of different nematode spe-cies. At this time, it was generally believed that all of theFaRPs that occurred inC. eleganswere encoded on theflp-1gene. Further studies identified a homologous gene in theclosely related speciesC. vulgaris[93].
Recently, theC. elegansgenome sequencing project cou-pled with polymerase chain reaction strategies have identi-fied at least 17 other RFamide-encoding genes. These geneshave been designatedflps-2 to -18 chronologically upondiscovery [79]. Most of these genes encode peptides withcharacteristic C-terminal signature sequences (see Table 8).C. elegansgenes have been identified that encode some 54different RFamides. Of interest, all of theseC. elegansRFamides possess a nonpolar hydrophobic amino acid (Ile,
Leu, Met, Phe, or Val) in the third position from the Cterminus; six different C-terminal tripeptideamides havebeen identified in nematode FaRPs (FRFamide, IRFamide,LRFamide, MRFamide, VRFamide, and MRYamide). Po-sition 4 from the C terminus appears to be hypervariablewith either a hydrophobic amino acid (Leu, Met, Phe, Pro,or Val) or a polar uncharged residue (Thr or Tyr) in thisposition in all known nematode FaRPs.
To date, a single RFamide-encoding gene has been iden-tified in A. suum[25]. This gene, designatedafp-1, encodessix RFamides that possess a C-terminal PGVLRFamidemotif. All of the six peptides encoded by this gene(AVPGVLRFamide [AF3], GDVPGVLRFamide [AF4],GFGDEMSMPGVLRFamide [AF10], SDMPGVLRFamide [AF13], SMPGVLRFamide [AF14], and GMPGVLRFamide [AF20]) have been characterized structurally [16,20]. Available genetic data from bothA. suumand C.eleganssuggest that RFamides with similar C-terminal re-gions (and variable N-termini) are encoded together onparticular genes [25,79]. This is particularly evident in theC. elegans flp-genes, for example,flp-1 encodes PNFLRFamides,flp-2 encodes PIRFamides, and so on. Althoughmany of the peptides encoded on a single gene have struc-tural similarities and thus may play similar physiologicalroles (PF1 and PF2, which are also co-encoded on theflp-1gene, have qualitatively identical actions on somatic bodywall muscle strips ofA. suum), there is evidence from theC.elegans flp-1 gene of co-encoded peptides that have differ-ent mechanisms of action in nematodes, i.e. SDPNFLRFamide (PF1) and KPNFLRFamide (CF6), which have in-hibitory activities onA. suumbody wall muscle that involveNO and Cl2, respectively (see [63]). Clearly, in some cases,the N-terminal differences of FaRPs have no effect on theirbiological activity whereas in other cases N-terminal differ-ences result in unique biological activities.
Most of the C. elegans flpgenes (flps-1 to -12) areexpressed throughout worm development (egg, four larvalstages, and the adult) [79]. However, althoughflps-8, -9, and-13 were transcribed in egg and larval stages ofC. elegans,they were not transcribed in adult worms, suggesting adevelopmental role for someC. elegansFaRPs. Furthercomplexity is added to the nematode FaRP gene story by thefact that a number of theflp-genes (flps-1, 2 and 11) havebeen shown to be alternatively spliced [79]. Of these,flps-1and -11 produce transcripts that encode different FaRPs andmay provide a mechanism for differential expression.
The recently structurally characterizedC. elegansFaRPHFYNFSESRKPNFLRFamide confirms the lack of under-standing of nematode FaRPergic systems [20]. Proteolyticprocessing of this peptide would be expected to generateKPNFLRFamide (CF6), so this sequence may represent partof a precursor peptide that has not been fully processed. Onthe other hand, this peptide may have biologic activity thatis distinct from that of CF6. Furthermore, enzymatic pro-cessing of theflp-1 precursor was believed to generate anumber of peptides including KPNFLRFamide (CF6). If a
1013T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

similar processing pathway was followed with theflp-1gene product, then a peptide with the sequence SFDNFDRESRKPNFLRFamide would be generated, i.e. a peptidedifferent from that than was structurally characterized.These data indicate that there is anotherflp-1-like peptideencoding gene inC. elegansthat awaits identification.
Recently, it has been reported that the disruption of theflp-1 gene inC. eleganscauses multiple behavioral defectsin the worm [80].Flp-1 gene disruption, which involved thegeneration offlp-1 deletion mutants, displayed a number ofmotor and sensory function defects including: loopy and
uncoordinated movement, hyperactivity and wandering,nose touch insensitivity, absence of osmotic avoidance, and,absence of serotonin-induced movement inhibition [80].Some of theflp-1 knock-out phenotypes were also apparentin C. elegansthat had G protein subunit mutations. A seriesof experiments with double-mutant (flp-1 and G proteinsubunit) worms revealed thatflp-1 peptides act upstream ofselected G protein subunits in movement coordination [80].It is clear that genetic manipulation ofC. eleganswillprovide fundamental insight into the biological activity andmechanisms of action of nematode FaRPs.
Table 8C. elegansFaRPs predicted from FaRP-encoding (flp) genes
flp-1 flp-9S A D P N F L R F G (2 copies) K P S F V R F G
S Q P N F L R F GA S G D P N F L R F G flp-10
S D P N F L R F G Q P K A R S G Y I R F GA A A D P N F L R F G
P N F L R F G flp-11A G S D P N F L R F G A M R N A L V R F G
A S G G M R N A L V R F Gflp-2 N G A P Q P F V R F G
S P R E P I R F GL R G E P I R F G flp-12
flp-3S P L G T M R F G flp-13
T P L G T M R F G S D R P T R A M D S P L I R F G
E A E E P L G T M R F G A A D G A P L I R F GN P L G T M R F G A P E A S P L I R F G
A S E D A L F G T M R F G A S P S A P L I R F GE D G N A P F G T M R F G S P S A V P L I R F G
S A E P F G T M R F G A S S A P L I R F GS A D D D S A P F G T M R F G
N P E N D T P F G T M R F G flp-14(4 copies) K H E Y L R F G
S L L D Y R F GK H E Y L R F G
flp-4 S L L D Y R F GP T F I R F G E I V F H Q I S P I F F R F G
A S P S F I R F Gflp-15
flp-5 G G Q G P L R F GG A K F I R F G G P S G P L R F G
A G A K F I R F GA P K P K F I R F G flp-16
A Q T F V R F Gflp-6 G Q T F V R F G
(6 copies) K S A Y M R F Gflp-17
flp-7 K S A F V R F G(3 copies) S P Q R S S M V R F G K S Q Y I R F G(2 copies) T Q R S S M V R F G
S P E R S A M V R F G flp-18
S P D R S K M V R F G D V P G V L R F GK S V P G V L R F G
flp-8 S E V P G V L R F G(3 copies) K N E F I R F G
Refs. [79,87,88,93].
1014 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

9. Peptide metabolism
To date, only a few studies have examined nematodeneuropeptide metabolism. The first of these studies usedisolated muscle membranes fromA. suumand examined thedegradation of AF1 (KNEFIRFamide) [37,89,90]. In thesestudies, several membrane-bound enzyme actions were re-corded, including endopeptidase (causing Glu3-Phe4 peptidebond cleavage), aminopeptidase (producing N-terminallytruncated metabolites), and deamidase (producing a C-ter-minally carboxylated peptide) activities. Characterization ofthe endopeptidase activity identified a nephrilysin-like en-zyme that was phosphoramidon-sensitive. Other peptidemetabolism studies that investigated the degradation of PF4and the PF4 analogs Gly2PF4 and Ala2PF4 by muscle strippreparations identified only aminopeptidase-catalyzedcleavages thought to be carried out by a soluble enzymereleased by the muscle strips [46]. The absence of thedeamidase and endopeptidase activity in the latter studymay have been due to the fact that these enzymes were onlyexposed to the peptide following the collagenase treatmentstep in muscle membrane generation. It also should be notedthat the proline in position 2 of PF4 rendered this peptideunsusceptible to the aminopeptidase activity.
9.1. Discussion
With such a wide range of structurally related nematodepeptides, the difficulties in delineating the functional role(s)of each one are immediately apparent. Also, because thenumbers of structurally characterized peptides have in-creased rapidly over the last 5 years, the functions and/oreffects of many of the peptides remain unknown. Neverthe-less, physiology studies carried out on nematode tissues byusing nematode RFamides have revealed a surprising com-plexity and diversity in peptide action. This diversity offunction is supported further by the numerous alterations inphenotype induced by FaRP-encoding gene knock-out inC.elegans. Although the complexity in the structure and ac-tions of identified nematode RFamides make it difficult togeneralize about their activities, it is apparent neverthelessthat the activities of the inhibitory peptides investigated todate appear to be postsynaptic in that neurons are not re-quired for their activities. On the other hand, the actions ofall of the excitatory nematode RFamides examined to dateare abolished by nerve cord removal, suggesting that theymay act presynaptically. The physiological effects of allknown nematode peptides need to be examined to establishif this is a general feature of the actions of these peptides.
Although nematode species have a diverse array of RFamides, most of which have distinguishable actions presum-ably mediated by different receptor subtypes, there is clearlycross-species activity, e.g. the effects ofP. redivivus-de-rived PF1 and PF4 onA. suum. It would seem unlikely thatthis cross-species activity is due to the fact that differentnematode species have a full complement of identical pep-
tides and possess specific receptors for each. This wouldmean that there are a large number of RFamides that awaitstructural characterization. The diversity in the RFamidecomplements within particular nematode species might de-mand the occurrence of highly discriminating receptors thatare able to distinguish between these structurally relatedpeptides. If the receptors must be highly discriminatory,interspecies activity would be unexpected. Another mecha-nism that could prevent the interaction of a range of relatedpeptides with single receptors could be restricted expressionpatterns of individual RFamides and/or their receptors. Onlythrough the mapping of gene and/or peptide expressionpatterns can the mechanisms used to integrate this complexsignaling system begin to be understood. Clearly, the firststeps in establishing the expression patterns of selectedFaRP-encoding genes has been taken inC. elegansby usingreporter constructs.
10. Non-FMRFamide-related peptides in nematodes
The occurrence of neuropeptides other than FaRPs innematodes has been indicated by a number of immunocy-tochemical screens (see [8–10,95,97–99]). However, it isunclear how many of the immunoreactivities identified aregenuine and how many are simply due to nonspecific cross-reactivity with, for example, endogenous FaRPs. Examplesof previously identified immunoreactivities that subse-quently were found to be the result of nonspecific cross-reactivity with endogenous FaRPs include pancreaticpolypeptide [7], NPF [67], and SALMFamide [11], each ofwhich were localized to the nervous system ofA. suum.Until all of the nematode neuropeptides have been charac-terized and highly specific antisera generated, immunocy-tochemical screens will remain difficult to interpret andunreliable as indicators of endogenous neuropeptide diver-sity in nematodes.
It should be noted that one other peptide has been struc-turally isolated fromA. suum. This peptide had the primarystructure TKQELE, was designated TE-6, and was isolatedby using an antiserum to KGQELE, a sequence that flanksthe C terminus of pancreastatin in rat chromogranin A[101]. This peptide is believed to represent a novel class ofnematode neuropeptide.
Although NPF has not been characterized from nema-todes, they do possess a NPY receptor homolog [24]. Thenpr-1 gene of C. elegansencodes a G protein coupledreceptor that is homologous to NPY receptors. Loss-of-function mutations in this gene resulted in alterations ofsocial feeding behavior in the worms. Of interest, NPY isknown to influence food intake in higher vertebrates, sug-gesting that there may be functional similarities in the roleof NPY receptors in worms and humans.
With respect to the function of non-RFamide peptides onnematodes, pentagastrin and angiotensin I were found tostimulate the motility ofP. redivivus in a sedimentation
1015T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

assay [100]. However, nothing is known about the func-tional role of other non-FaRP neuropeptides in nematodes.
11. Conclusions
The general pattern of dissimilarity that exists betweenplatyhelminth and nematode parasites extends to the pepti-dergic systems. Although FaRPs play an important role inboth types of worms, there are stark differences between theFaRPergic systems in the two worm phyla. The emergingpattern in flatworms is the presence of a very limited num-ber of FaRPs in a given species (one or two in each exam-ined to date), whereas each species of roundworm has alarge number of distinct FaRPs. This presents an interestingcontrast to the structural differences of the nervous systems,where the flatworms are markedly more complex than thenematodes. A synthetic principle with respect to the pepti-dergic systems is that platyhelminths have structural com-plexity coupled with relative chemical simplicity comparedwith the structural simplicity and chemical complexity innematodes.
There is also an apparent functional difference amongthe FaRP systems in the two phyla. In trematodes, all of theknown actions of FaRPs are excitatory, whereas in nema-todes both exctitatory and inhibitory functions have beendemonstrated. This may be viewed simply as an extensionof the greater chemical complexity of the FaRPergic systemof nematodes, in that the greater array of distinct peptidesallows for a greater range of functions. Further evidence forthis hypothesis resides in the complexity of peptide actionsin nematodes, with differential effects on different musclesystems and multiphasic effects being commonplace.
Despite these differences, the importance of FaRPs inboth types of parasitic worms is enticing to parasitologistsand there are important similarities. For example, the nem-atode-derived FaRPs AF1, AF2, PF1, PF2, PF3/AF8, andPF4 are all excitatory on flatworm muscle [32,60], demon-strating an important degree of similarity between FaRPreceptors in both phyla. FaRP receptors appear to be viablechemotherapeutic targets in both nematodes and flatworms,and the potential that a single phamacological agent couldbe active against parasites of both phyla is very attractive. Inthis vein, the allure of FaRP receptors as broad-spectrumdrug targets is enhanced further by reported cross-phylaactivity between nematode and insect neuropeptides, raisingthe possiblity of anthelminthics with insecticidal activity.
Drug resistance among parasitic helminths is not merelya remote future possibility, it is a real and present problem.Considering the vast burden that chemotherapy currentlybears in controlling helminth-related morbidity, it is incum-bent on us in the present to use existing chemotherapeuticswisely and to endeavor to indentify novel targets that couldbe the basis for new generations of anthelmintics. Parasitepeptidergic nervous systems may prove to be pivotal to thisgoal.
References
[1] Aguinaldo AM, Turbeville JM, Linford LS, Rivera MC, Garey JR,Raff RA, Lake JA. Evidence for a clade of nematodes, arthropodsand other moulting animals. Nature (London) 1997;387:489–93.
[2] Armstrong EP, Halton DW, Tinsley RC, Cable J, Johnston RN,Johnston CF, Shaw C. Immunocytochemical evidence for the in-volvement of an FMRFamide-related peptide in egg production inthe flatworm parasitePolystoma nearcticum. J Comp Neurol 1997;377:41–8.
[3] Atkinson HJ, Isaac RE, Harris PD, Sharpe CM. FMRFamide-im-munoreactivity within the nervous system of the nematodesPan-agrellus redivivus, Caenorhabditis elegansand Heterodera gly-cines. J Zool 1988;216:663–71.
[4] Bascal ZA, Montgomery A, Holden–Dye L, Williams RG, WalkerRJ. Histochemical mapping of NADPH diaphorase in the nervoussystem of the parasitic nematodeAscaris suum. Parasitology 1995;110:625–37.
[5] Bowman JW, Friedman AR, Thompson DP, Ichhpurani AK, Kell-man MF, Marks N, Maule AG, Geary TG. Structure-activity rela-tionships of KNEFIRFamide (AF1), a nematode FMRFamide-re-lated peptide, onAscaris suummuscle. Peptides 1996;17:381–7.
[6] Bowman JW, Winterrowd CA, Friedman AR, Thompson DP, KleinRD, Davis JP, Maule AG, Blair KL, Geary TG. Nitric oxide medi-ates the inhibitory effects of SDPNFLRFamide, a nematode FMRF-related neuropeptide, inAscaris suum. J Neurophysiol 1995;74:1880–8.
[7] Brownlee DJ, Brennan GP, Halton DW, Fairweather I, Shaw C.Ultrastructural localization of pancreatic polypeptide- and FMRF-amide-immunoreactivities within the central nervous system of thenematode,Ascaris suum(Nematoda: Ascaroidea). Parasitology1994;108:587–93.
[8] Brownlee DJ, Fairweather I, Johnston CF. Immunocytochemicaldemonstration of neuropeptides in the peripheral nervous system ofthe roundwormAscaris suum(Nematoda, Ascaroidea). Parasitol Res1993;79:302–8.
[9] Brownlee DJ, Fairweather I, Johnston CF, Shaw C. Immunocyto-chemical demonstration of peptidergic and serotoninergic compo-nents in the enteric nervous system of the roundworm,Ascaris suum(Nematoda, Ascaroidea). Parasitology 1994;108:89–103.
[10] Brownlee DJ, Fairweather I, Johnston CF, Smart D, Shaw C, HaltonDW. Immunocytochemical demonstration of neuropeptides in thecentral nervous system of the roundworm,Ascaris suum(Nematoda:Ascaroidea). Parasitology 1993;106:305–16.
[11] Brownlee DJA, Fairweather I, Thorndyke MC, Johnston CF. Cel-lular and subcellular localization of SALMFamide (S1)-like immu-noreactivity within the central nervous system of the nematodeAscaris suum(Nematoda, Asaroidea). Parasitol Res 1996;82:149–56.
[12] Brownlee DJ, Holden–Dye L, Fairweather I, Walker RJ. The actionof serotonin and the nematode neuropeptide KSAYMRFamide onthe pharyngeal muscle of the parasitic nematode,Ascaris suum.Parasitology 1995;111:379–84.
[13] Cioli D, Pica–Mattoccia L, Archer S. Antischistosomal drugs: past,present. . . and future? Pharmacol Ther 1995;68:35–85.
[14] Cowden C, Sithigorngul P, Brackley P, Guastella J, Stretton AOW.Localization and differential expression of FMRFamide-like immu-noreactivity in the nematodeAscaris suum. J Comp Neurol 1993;333:455–68.
[15] Cowden C, Stretton AOW. AF2, an Ascaris neuropeptide: isolation,sequence, and bioactivity. Peptides 1993;14:423–30.
[16] Cowden C, Stretton AOW. Eight novel FMRFamide-like neuropep-tides isolated from the nematodeAscaris suum. Peptides 1995;16:491–500.
1016 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

[17] Cowden C, Stretton AOW, Davis RE. AF1, a sequenced bioactiveneuropeptide isolated from the nematodeAscaris suum. Neuron1989;2:1465–73.
[18] Curry WJ, Shaw C, Johnston CF, Thim L, Buchanan KD. Neuropep-tide F: primary structure from the tubellarian,Artioposthia triangu-lata. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol1992;101:269–74.
[19] Davenport TRB, Lee DL, Isaac RE. Immunocytochemical demon-stration of a neuropeptide inAscaris suum(Nematoda) using anantiserum to FMRFamide. Parasitology 1988;97:81–8.
[20] Davis RE, Stretton AOW. The motornervous system of Ascaris:electrophysiology and anatomy of the neurons and their control byneuromodulators. Parasitology 1996;113:S99–117.
[21] Day TA, Chen GZ. The metalloprotease inhibitor 1,10-phenanthro-line affectsSchistosoma mansonimotor activity, egg laying andviability. Parasitology 1998;116:319–25.
[22] Day TA, Maule AG, Shaw C, Halton DW, Moore S, Bennett JL, PaxRA. Platyhelminth FMRFamide-related peptides (FaRPs) contractSchistosoma mansoni(Trematoda Digenea) muscle fibres in vitro.Parasitology 1994;109:455–9.
[23] Day TA, Maule AG, Shaw C, Pax RA. Structure-activity relation-ships of FMRFamide-related peptide-induced contractions ofSchis-tosoma mansonimuscle. Peptides 1997;18:917–21.
[24] deBono M, Bargmann CI. Natural variation in a neuropeptide Yreceptor homolog modifies social behavior and food response inC.elegans. Cell 1998;96:679–89.
[25] Edison AS, Messinger LA, Stretton AOW. Afp-1: A gene encodingmultiple transcripts of a new class of FMRFamide-like neuropep-tides in the nematodeAscaris suum. Peptides 1997;18:929–35.
[26] Fairweather I, Mahendrasingam S, Johnston CF, Halton DW, ShawC. Peptidergic nerve elements in three developmental stages of thetetraphyllidean tapewormTrilocularia acanthiaevulgaris. ParasitolRes 1990;76:497–508.
[27] Fellowes RA, Maule AG, Marks NJ, Geary TG, Thompson DP,Shaw C, Halton DW. Modulation of the motility of the vagina veraof Ascaris suum in vitroby FMRFamide-related peptides. Parasi-tology 1998;116:277–87.
[28] Franks CJ, Holden–Dye L, Williams RG, Pang FY, Walker RJ. Anematode FMRFamide-like peptide, SDPNFLRFamide (PF1), re-laxes the dorsal muscle strip preparation ofAscaris suum. Parasi-tology 1994;108:229–336.
[29] Geary TG, Bowman JW, Friedman AR, Maule AG, Davis JP,Winterrowd CA, Klein RD, Thompson DP. The pharmacology ofFMRFamide-related neuropeptides in nematodes: new opportunities forrational anthelmintic discovery? Int J Parasitol 1995;25:1273–80.
[30] Geary, TG, Price DA, Bowman JW, Winterrowd CA, MacKenzieCD, Garrison RD, Williams JF, Friedman AR. Two FMRFamide-like peptides from the free-living nematodePanagrellus redivivus.Peptides 1992;13:209–14.
[31] Geerts S, Coles GC, Gryseels B. Anthelmintic resistance in humanhelminths: learning from the problems with worm control in live-stock. Parasitol Today 1997;13:149–51.
[32] Graham MK, Fairweather I, McGeown JG. The effects of FaRPs onthe motility of isolated muscle strips from the liver fluke,Fasciolahepatica. Parasitology 1997;114:455–65.
[33] Halton DW, Shaw C, Maule AG, Smart D. Regulatory peptides inhelminth parasites. Adv Parasitol 1994;34:163–227.
[34] Holden–Dye L, Brownlee DJA, Walker RJ. The effects of thepeptide KPNFIRFamide (PF4) on the somatic muscle cells of theparasitic nematodeAscaris suum. Br J Pharmacol 1997;120:379–86.
[35] Holden–Dye L, Franks CJ, Williams RG, Walker RJ. The effect ofthe nematode peptides SDPNFLRFamide (PF1) and SADPNFLRFamide (PF2) on synaptic transmission in the parasitic nematodeAscaris suum. Parasitology 1995;110:449–55.
[36] Hyman LH. The invertebrates: Platyhelminthes and Rhynchocoela.The Acoelomate Bilateria, Vol. 2. New York: McGraw-Hill, 1951.pp. 1–550.
[37] Isaac RE, MacGregor D, Coates D. Metabolism and inactivation ofneurotransmitters in nematodes. Parasitology 1996;113:S157–73.
[38] Ismail M, Metwally A, William S, Farghally A, Tao LF, Day TA,Bennett JL. Resistance to praziquantel: direct evidence from para-sites isolated from Egyptian villagers. Am J Trop Med Hyg 1999;60:932–5.
[39] Jagdale GB, Gordon R. Distribution of FMRF-amide-like peptide inthe nervous system of a mermithid nematode,Romanomermis culi-civorax. Parasitol Res 1994;80:467–73.
[40] Johnston RN, Shaw C, Brennan GP, Maule AG, Halton DW. Lo-calisation, quantitation, and characterisation of neuropeptide F- andFMRFamide-immunoreactive peptides in turbellarians and a mono-genean: a comparative study. J Comp Neurol 1995;357:76–84.
[41] Johnston RN, Shaw C, Halton DW, Verhaert P, Baguna J. GYIR-Famide: a novel FMRFamide-related peptide (FaRP) from the tri-clad turbellarian,Dugesia tigrina. Biochem Biophys Res Commun1995;209:689–97.
[42] Johnston RN, Shaw C, Halton DW, Verhaert P, Blair KL, BrennanGP, Price DA, Anderson PAV. Isolation localization, and bioactivityof the FMRFamide-related neuropeptides GYIRFamide and YIRF-amide from the marine turbellarianBdelloura candida. J Neurochem1996;67:814–21.
[43] Keating C, Thorndyke MC, Holden–Dye L, Williams RG, WalkerRJ. The isolation of a FMRFamide-like peptide from the nematodeHaemonchus contortus. Parasitology 1995;111:515–21.
[44] Koopowitz H. Feeding behaviour and the role of the brain in thepolyclad flatworm,Planocerca gilchristi. Anim Behav 1970;18:31–5.
[45] Koopowitz H, Silver D, Rose G. Primitive nervous systems. Controland recovery of feeding behaviour in the polyclad flatworm,Noto-plana acticola. Biol Bull 1976;150:411–25.
[46] Kubiak TM, Maule AG, Marks NJ, Martin RA, Wiest JR. Impor-tance of the proline residue to the functional activity and metabolicstability of the nematode FMRFamide-related peptide, KPNFIRF-amide (PF4). Peptides 1996;17:1267–77.
[47] Larhammar D. Evolution of neuropeptide Y, peptide YY and pan-creatic polypeptide. Regul Pept 1996;62:1–11.
[48] Larhammar D, Blomqvist AG, Soderberg C. Evolution of neuropep-tide Y and its related peptides. Comp Biochem Physiol C PharmacolToxicol Endocrinol 1993;106:743–52.
[49] Leach L, Trudgill DL, Gahan PB. Immunocytochemical localizationof neurosecretory amines and peptides in the free-living nematode,Goodeyus ulmi. Histochem J 1987;19:471–5.
[50] Leung PS, Shaw C, Maule AG, Thim L, Johnston CF, Irvine GB.The primary structure of neuropeptide F (NPF) from the gardensnail,Helix aspersa. Regul Pept 1992;41:71–81.
[51] Li C, Chalfie M. FMRFamide-like immunoreactivity inC. elegans.Soc Neurosci Abstr 1986;12:246.
[52] Linacre A, Kellett E, Saunders S, Bright K, Benjamin PR, Burke JF.Cardioactive neuropeptide Phe-Met-Arg-Phe-NH2 (FMRFamide)and novel related peptides are encoded in multiple copies by a singlegene in the snailLymnaea stagnalis. J Neurosci 1990;10:412–9.
[53] Magee RM, Fairweather I, Johnston CF, Halton DW, Shaw C.Immunocytochemical demonstration of neuropeptides in the ner-vous system of the liver fluke,Fasciola hepatica(Trematoda, Di-genea). Parasitology 1989;98:227–38.
[54] Magee RM, Shaw C, Fairweather I, Thim L, Johnston CF, HaltonDW. Isolation and partial sequencing of a pancreatic polypeptide-likeneuropeptide from the liver fluke,Fasciola hepatica. Comp BiochemPhysiol C Pharmacol Toxicol Endocrinol 1991;100:507–11.
[55] Mair GR, Maule AG, Halton DW, Orr D, Johnston RN, Johnston CF,Shaw C. Comparative analysis of the distribution of bradykinin-, GY-IRFamide-, and neuropeptide F-like immunoreactivities in the mono-genean,Diclidophora merlangi. Parasitology 1997;114:467–73.
[56] Mair GR, Maule AG, Shaw C, Halton DW. Muscling in on parasiticflatworms. Parasitol Today 1998;14:73–6.
[57] Marks NJ, Halton DW, Maule AG, Brennan GP, Shaw C, SouthgateVR, Johnston CF. Comparative analyses of the neuropeptide F
1017T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

(NPF)- and FMRFamide-related peptide (FaRP)- immunoreactivi-ties in Fasciola hepaticaandSchistosoma spp.Parasitology 1995;110:371–81.
[58] Marks NJ, Johnson S, Maule AG, Halton DW, Shaw C, Geary TG,Moore S, Thompson DP. Physiological effects of platyhelminthRFamide peptides on muscle-strip preparations ofFasciola hepatica(Trematoda: Digenea). Parasitology 1996;113:393–401.
[59] Marks NJ, Maule AG, Geary TG, Thompson DP, Davis JP, HaltonDW, Verhaert P, Shaw C. APEASPFIRFamide, a novel FMRF-amide-related decapeptide fromCaenorhabditis elegans: structureand myoactivity. Biochem Biophys Res Commun 1997;231:591–5.
[60] Marks NJ, Maule AG, Halton DW, Geary TG, Shaw C, ThompsonDP. Pharmacological effects of nematode FMRFamide-related pep-tides (FaRPs) on muscle contractility of the trematode,Fasciolahepatica. Parasitology 1997;114:531–9.
[61] Marks NJ, Shaw C, Davis JP, Maule AG, Halton DW, Verhaert P,Geary TG, Thompson DP. Isolation of AF2 (KHEYLRFamide)from Caenorhabditis elegans: evidence for the presence of morethan one FMRFamide encoding gene. Biochem Biophys Res Com-mun 1996;217:845–51.
[62] Martin RJ, Valkanov MA, Dale VME, Robertson A, Murray I.Electrophysiology ofAscarismuscle and antinematodal drug action.Parasitology 1996;113:S137–56.
[63] Maule AG, Bowman JW, Thompson DP, Marks NJ, Friedman AR,Geary TG. FMRFamide-related peptides (FaRPs) in nematodes:occurrence and neuromuscular physiology. Parasitology 1996;133:S119–35.
[64] Maule AG, Brennan GP, Halton DW, Shaw C, Johnston CF, MooreS. Neuropeptide F-immunoreactivity in the monogenean parasiteDiclidophora merlangi. Parasitol Res 1992;78:655–60.
[65] Maule AG, Geary TG, Bowman JW, Marks NJ, Blair KL, HaltonDW, Shaw C, Thompson DP. Inhibitory effects of nematode FMRFamide-related peptides (FaRPs) on muscle strips fromAscarissuum. Invert Neurosci 1995;1:255–65.
[66] Maule AG, Halton DW, Johnston CF, Shaw C, Fairweather I. Theserotoninergic, cholinergic and peptidergic components of the ner-vous system in the monogenean parasite,Diclidophora merlangi: acytochemical study. Parasitology 1990;100:255–73.
[67] Maule AG, Halton DW, Shaw C. Neuropeptide F: a ubiquitousinvertebrate neuromediator? Hydrobiologia 1995;305:297–303.
[68] Maule AG, Halton DW, Shaw C, Johnston CF. The cholinergic,serotoninergic and peptidergic components of the nervous system ofMoniezia expansa(Cestoda, Cyclophyllidea). Parasitology 1993;106:429–40.
[69] Maule AG, Shaw C, Bowman JW, Halton DW, Thompson DP,Geary TG, Thim L. The FMRFamide-like neuropeptide AF2 (As-caris suum) is present in the free-living nematode,Panagrellusredivivus(Nematoda, Rhabditida). Parasitology 1994;109:351–6.
[70] Maule AG, Shaw C, Bowman JW, Halton DW, Thompson DP,Geary TG, Thim L. KSAYMRFamide: a novel FMRFamide-relatedheptapeptide from the free-living nematode,Panagrellus redivivus,which is myoactive in the parasitic nematode,Ascaris suum. Bio-chem Biophys Res Commun 1994;200:973–80.
[71] Maule AG, Shaw C, Bowman JW, Halton DW, Thompson DP,Thim L, Kubiak TM, Martin RJ, Geary TG. Isolation and prelimi-nary biological characterization of KPNFIRFamide, a novel FMR-Famide-related peptide from the free-living nematode,Panagrellusrevividus. Peptides 1995;16:87–93.
[72] Maule AG, Shaw C, Halton DW, Brennan GP, Johnston CF, MooreS. Neuropeptide F (Moniezia expansa): localization and character-ization using specific antisera. Parasitology 1992;105:505–12.
[73] Maule AG, Shaw C, Halton DW, Curry WJ, Thim L. RYIRFamide:a turbellarian FMRFamide-related peptide (FaRP). Regul Pept 1994;50:37–43.
[74] Maule AG, Shaw C, Halton DW, Johnston CF, Fairweather I.Localization, quantification and characterization of pancreaticpolypeptide, immunoreactivity in the parasitic flatwormDiclid-
ophora merlangiand its fish host (Merlangius merlangus). GenComp Endocrinol 1989;74:50–6.
[75] Maule AG, Shaw C, Halton DW, Thim L. GNFFRFamide. A novelFMRFamide-immunoreactive peptide isolated from the sheep tape-worm Moniezia expansa. Biochem Biophys Res Commun 1993;193:1054–60.
[76] Maule AG, Shaw C, Halton DW, Thim L, Johnston CF, FairweatherI, Buchanan KD. Neuropeptide F: a novel parasitic flatworm regu-latory peptide fromMoniezia expansa(Cestoda:Cyclophyllidea).Parasitology 1991;102:309–16.
[77] Michel MC, Beck–Sickinger A, Cox H, Doods HN, Herzog H,Larhammar D, Quirion R, Schwartz T, Westfall T. Internationalunion of pharmacology recommendations for the nomenclature ofneuropeptide Y, peptide YY, and pancreatic polypeptide receptors.Pharmacol Rev 1998;50:143–50.
[78] Moneypenny CG, Maule AG, Shaw C, Day TA, Pax RA, HaltonDW. Physiological effects of platyhelminth FMRFamide-relatedpeptides (FaRPs) on motility of the monogeneanDiclidophora mer-langi. Parasitology 1997;115:281–8.
[79] Nelson LS, Kim KY, Memmott RE, Li C. FMRFamide-related genefamily in the nematode,Caenorhabditis elegans. Mol Brain Res1998;58:103–11.
[80] Nelson LS, Rosoff ML, Li C. Disruption of a neuropeptide gene,flp-1, causes multiple behavioral defects inCaenorhabditis elegans.Science 1998;281:1686–90.
[81] Pang F-Y, Mason J, Holden–Dye L, Franks CJ, Williams RG,Walker RJ. The effects of the nematode peptide KHEYLRFamide(AF2) on the somatic musculature of the parasitic nematodeAscarissuum. Parasitology 1995;110:353–62.
[82] Payza K. FMRFamide receptors inHelix aspersa. Peptides 1987;8:1065–1074.
[83] Price DA. Evolution of a molluskan cardioregulatory neuropeptide.Am Zool 1986;26:1007–15.
[84] Rajpara SM, Garcia PD, Roberts R, Eliassen JC, Owens DF, MaltbyD, Myers RM, Mayeri E. Identification and molecular cloning of aneuropeptide Y homolog that produces prolonged inhibition inAply-sia neurons. Neuron 1992;9:505–13.
[85] Reuter M, Gustafsson MKS, Sahlgren C, Halton DW, Maule AG,Shaw C. The nervous system of Tricladida. I. Neuroanatomy ofProcerodes littoralis (Maricola, Procerodidae): an immunocyto-chemical study. Invert Neurosci 1995;1:113–22.
[86] Reuter M, Maule AG, Halton DW, Gustafsson MKS, Shaw C. Theorganization of the nervous system in Platyhelminthes. The neu-ropeptide F-immunoreactive pattern in Catenulida, Macrostomida,Proseriata. Zoomorph 1995;115:83–97.
[87] Rosoff ML, Burglin TR, Li C. Alternatively spliced transcripts ofthe flp-1 gene encode distinct FMRFamide-like peptides inCaeno-rhabditis elegans. J Neurosci 1992;12:2356–61.
[88] Rosoff ML, Doble KE, Price DA, Li C. Theflp-1 propeptide isprocessed into multiple, highly similar FMRFamide-like peptides inCaenorhabditis elegans. Peptides 1993;14:331–8.
[89] Sajid M, Isaac RE. Identification of a neuropeptide-degrading en-dopeptidase (neprilysin) ofAscaris suummuscle. Parasitology 1995;111:599–608.
[90] Sajid M, Isaac RE. Metabolism of AF1 (KNEFIRF-NH2) in thenematodeAscaris suum. Biochem Soc Trans 1994;22:S293.
[91] Sangster N. Pharmacology of anthelmintic resistance. Parasitology1996;113:S201–16.
[92] Schinkmann K, Li C. Localization of FMRFamide-like peptides inCaenorhabditis elegans. J Comp Neurol 1992;316:251–60.
[93] Schinkmann K, Li C. Comparison of twoCaenorhabditisgenesencoding FMRFamide (Phe-Met-Arg-Phe-NH2)-like peptides. MolBrain Res 1994;24:238–46.
[94] Shaw C, Maule AG, Halton DW. Platyhelminth FMRFamide-re-lated peptides. Int J Parasitol 1996;26:335–45.
1018 T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019

[95] Sithigorngul P, Stretton AOW, Cowden C. Neuropeptide diversityin Ascaris: an immunocytochemical study. J Comp Neurol 1990;294:362–76.
[96] Skuce PJ, Johnston C. F, Fairweather I, Halton DW, Shaw C,Buchanan KD. Immunoreactivity to the pancreatic polypeptide fam-ily in the nervous system of the adult human blood fluke,Schisto-soma mansoni. Cell Tissue Res 1990;261:573–81.
[97] Smart DS, Johnston CF, Curry WJ, Shaw C, Halton DW, Fair-weather I, Buchanan KD. Immunoreactivity to two specific regionsof chromogranin A in the nervous system ofAscaris suum. ParasitolRes 1992;78:329–35.
[98] Smart DS, Johnston CF, Maule AG, Halton DW, Hrckova G, ShawC, Buchanan KD. Localization ofDiploptera punctataallatostatin-like immunoreactivity in helminths: an immunocytochemical study.Parasitology 1995;110:87–96.
[99] Smart D, Johnston CF, Shaw C, Halton DW, Buchanan KD. Use ofspecific antisera for the localisation and quantitation of leucokininimmunoreactivity in the nematode,Ascaris suum. Comp BiochemPhysiol C Pharmacol Toxicol Endocrinol 1993;106:517–22.
[100] Smart D, Preston CM, Lloyd D. The use of a novel motility assay forthe isolation of bioactive peptides from the free-living nematodePanagrellus redivivus. Comp Biochem Physiol B Biochem Mol Biol1990;95:335–9.
[101] Smart D, Shaw C, Curry WJ, Johnston CF, Thim L, Halton DW,Buchanan KD. The primary structure of TE-6: a novel neuropeptidefrom the nematodeAscaris suum. Biochem Biophys Res Commun1992;187:1323–9.
[102] Soderberg C, Pieribone VA, Dahlstrand J, Brodin L, Larhammar D.Role of both peptide YY and neuropeptide Y in vertebrates sug-gested by abundant expression of their mRNAs in a cyclostomebrain. J Neurosci Res 1993;37:633–40.
[103] Stoll NR. This wormy world. J Parasitol 1947;33:1–18.[104] Trim N, Holden–Dye L, Ruddell R, Walker RJ. The effects of
the peptides AF3 (AVPGVLRFamide) and AF4 (GDVPGVLRF-amide) on the somatic muscle of the parasitic nematodesAscaris suumandAscaridia galli. Parasitology 1997;115: 213–22.
[105] Walker RJ. Neuroactive peptides with an RFamide or Famide car-boxyl terminal. Comp Biochem Physiol C Pharmacol Toxicol En-docrinol 1992;102:213–22.
[106] Warbrick EV, Rees HH, Howells RE. Immunocytochemical local-isation of an FMRFamide-like peptide in the filarial nematodesDirofilaria immitis and Brugia pahangi. Parasitol Res 1992;78:252–6.
[107] Warren KS, Bundy DAP, Anderson RM, Davis AR, Henderson DA,Jamison DT, Prescott N, Senft A. Helminth infection. In: JamisonDT, Mosely WH, Measham AR, Bobadilla JL, editors. Diseasecontrol priorities in developing countries. New York: Oxford Uni-versity Press, 1993. pp. 31–60.
[108] White JG, Southgate E, Thompson JN, Brenner S. The structure ofthe nervous system ofCaenorhabditis elegans. Philos Trans R SocLond B Biol Sci 1986;314:1–340.
[109] Wikgren M, Fagerholm HP. Neuropeptides in sensory structures ofnematodes. Acta Biol Hung 1993;44:133–6.
1019T.A. Day, A.G. Maule / Peptides 20 (1999) 999–1019





























