Embed Size (px)
Citation preview

www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, P
Late Miocene climate and life on land in Oregon
within a context of Neogene global change
Gregory J. Retallack*
Department of Geological Sciences University of Oregon, Eugene, OR 97403-1272, United States
Received 13 November 2003; received in revised form 2 July 2004; accepted 20 July 2004
Abstract
Clarendonian (12 Ma) fossil soils, plants, molluscs, fish, and mammals of eastern Oregon allow reconstruction of late
Miocene paleogeography, paleoclimate, and paleoecology on land between the global thermal maximum of the middle Miocene
(16 Ma) and global cooling and drying of the late Miocene (7 Ma). Six different pedotypes of paleosols recognized near Unity
and Juntura allow reinterpretation of local mammalian paleoecology. Fossil beavers dominated gleyed Entisols of riparian
forest. Abundant camels and common hipparionine horses dominated Alfisols of wooded grassland and grassy woodland.
Diatomites overlying mammal-bearing beds have bullhead catfish [Ictalurus (Ameiurus) vespertinus], as well as fossil leaves
dominated by live oak (Quercus pollardiana). Fossil plants and soils of Unity and Juntura are most like those of grassy live oak
woodland and savanna on the western slopes of the Sierra Nevada in northern California today. Fossil plants and soils indicate
mean annual temperature of 12.9 (7.7–17.7) 8C and mean annual precipitation of 879 (604–1098) mm. Miocene paleoclimatic
changes in eastern Oregon show no relationship to changes in oxygen isotopic composition of marine foraminifera, usually
taken as an index of global paleoclimatic change. Mismatch between land and sea paleoclimatic records is most likely an
artefact of global ice volume perturbation of oxygen isotopic values. Instead, Miocene paleoclimatic change in eastern Oregon
parallels changes in carbon isotopic composition of marine foraminifera, presumably through fluctuations in greenhouse gases.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Miocene; Paleosols; Plants; Mammals; Oregon
1. Introduction
Because Miocene soils, plants, and animals were
close to our own time and modern in many respects,
the surprisingly large amplitude of Miocene paleo-
climatic change is of interest for understanding
0031-0182/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.palaeo.2004.07.024
* Tel.: +1 541 3464558; fax: +1 541 3464692.
E-mail address: [email protected].
future global change. Evidence of global middle
Miocene (16 Ma) warmth includes lateritic paleosols
as far north and south as South Australia, Japan,
Oregon, and Germany (Schwarz, 1997). Middle
Miocene thermophilic trees such as sweetgum
(Liquidambar pachyphylla) grew as far north as
Alaska (Wolfe and Tanai, 1980), and coconuts
(Cocos zeylanica) dispersed as far south as New
Zealand (Fleming, 1975). Middle Miocene thermo-
alaeoecology 214 (2004) 97–123

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–12398
philic large foraminifera, corals, and molluscs
extended as far north as Oregon, Alaska, Japan,
and Kamchatkta (Moore, 1963; Itoigawa and Yama-
noi, 1990; Oleinik and Marincovich, 2001), and as
far south as South Australia and New Zealand (Li
and McGowran, 2000; Fleming, 1975). These
indicators of warmth disappeared from such high
latitudes by later middle Miocene (15 Ma), when
cold water minerals (glendonite pseudomorphs of
ikaite) formed in marine rocks of Oregon (Boggs,
1972). Benthic foraminiferal oxygen isotopic values
also indicate a global middle Miocene marine
thermal maximum, followed by late Miocene global
cooling (Zachos et al., 2001).
Fig. 1. Late Miocene fossil localities of Juntura, Ironside, and Unity, east
Unity (Thayer, 1957; Thayer and Brown, 1973; Reef, 1983; Walker and M
Tables 4–5; unnumbered localities have not yet yielded identifiable specim
These global trends also were felt in eastern
Oregon, where fossil leaves indicate marked cooling
and drying between about 16 and 15 Ma (Wolfe et al.,
1997; Graham, 1999), and a second cooling and
drying to a modern flora by latest Miocene (7 Ma;
Chaney, 1948). Paleosols of eastern Oregon show
similar changes between middle Miocene mesic
forests and late Miocene sagebrush steppe (Bestland
and Krull, 1997; Retallack et al., 2002a; Sheldon,
2003). Associated mammal faunas include a transition
from three-toed, mixed-feeding horses of the middle
Miocene (15 Ma, Barstovian) to modern monodactyl
grazing horses by late Miocene (7 Ma, Hemphillian;
Downs, 1956; Shotwell, 1970).
ern Oregon, with detailed geological map and cross-section around
acLeod, 1991). Numbered fossil localities and fossils are listed in
ens.

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 99
This study examines paleoclimate and paleo-
ecology of the intervening part of the late
Miocene within the context of Miocene global
paleoclimatic extremes, using evidence from fossil
soils, plants, molluscs, fish, and mammals in the
eastern Oregon. Previous study of mammalian
communities of this age from near Juntura
(Shotwell, 1963; Shotwell and Russell, 1963),
Ironside (Merriam, 1916), and Baker City (Downs,
1952) are here supplemented with additional
collections from near Unity, including reappraisal
of a locality for a previously collected gompho-
there skull and jaws (Orr and Orr, 1999). New
Fig. 2. Measured section of the Ironside Formation in Windlass Gulch, 3
and their development, calcareousness, and hue (using conventions of Re
collections of fossil plants and fish are also
reported, as well as new mapping of the area
around Unity (Fig. 1) and an assessment of the
paleoenvironmental significance of paleosols asso-
ciated with these fossils (Figs. 2 and 3). The
record of Neogene paleoclimatic fluctuation from
these and other paleosols in eastern Oregon and
Washington (Retallack, 1997b) does not correlate
with oxygen isotopic records from deep-sea
foraminifera but is in phase with foraminiferal
variations in carbon isotopic composition (Zachos
et al., 2001). This may indicate a closer relation-
ship with the carbon cycle and greenhouse gases,
miles northeast of Unity, Baker County, Oregon, showing paleosols
tallack, 1997a, 2001b).

Fig. 3. Geological sections of fossil quarries excavated by Shotwell
(1963) near Juntura, Oregon, showing paleosols and their develop-
ment, calcareousness, and hue (using conventions of Retallack,
1997a, 2001b).
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123100
such as CO2 and CH4 (Retallack, 2002), than with
deep-sea temperature or polar ice volume tracked
by oxygen isotopic composition.
2. Materials and methods
Mapping and measurement of two stratigraphic
sections near Unity, Baker County, Oregon (Figs.
2 and 4) involved collection of 95 rock specimens
and 62 fossils, which have been catalogued in
collections of John Day Fossil Beds National
Monument, Kimberly, Oregon (Scott Foss curator:
catalog on line at http://www.museum.nps.gov/).
Paleocurrents were sighted along trough cross bed
axes using a Brunton compass. Two stratigraphic
sections also were measured near Juntura, Mail-
heur County, Oregon (Fig. 3) at fossil quarries of
Shotwell (1963). Petrographic thin sections were
cut of all the collected rocks in order to support
field identifications of lithologies, and 36 thin
sections were point-counted to determine paleosol
textures and mineral composition (following the
methods of Retallack, 1997a, 2001b). Clays and
evaporite minerals were identified by X-ray
diffraction using a Rigaku computer-automated
goniometer.
Six distinct kinds of paleosols, or pedotypes, were
recognized (Table 1) throughout the mapped area
(Figs. 1 and 2) and also near Juntura (Fig. 3) and
Ironside. Each pedotype represents a distinct ancient
environment (Table 2). The pedotype names are from
the Sahaptin Native American language (Rigsby,
1965; Delancey et al., 1988) and are part of a wider
classification of Oregon Cenozoic paleosols (Retal-
lack et al., 2000). Descriptive and analytical data for
these paleosols are presented in Fig. 5 and Table 1.
Chemical analyses of 21 paleosol specimens by
Bondar Clegg of Vancouver, Canada, used X-ray
fluorescence of a borate-fused bead for major ele-
ments, titration for ferrous iron, and gravimetry for
loss on ignition. Bulk density for each specimen was
determined using the clod method. Of 109 paleosols
logged near Unity and 42 near Juntura, 45 had
pedogenic carbonate, and 5 were chemically analyzed
for paleoclimatic interpretation. These data are added
to 754 pedogenic carbonate measurements and 92
chemical analyses of other paleosols from Oregon and
Washington ranging back in age to 28.7 Ma (Gus-
tafson, 1978; Bestland and Krull, 1997; Tate, 1998;
Retallack, 2004), in order to assess local Neogene
paleoclimate and its correspondence with global
paleoclimatic change (Zachos et al., 2001).
3. Geological background
Only rocks of the Clarendonian North Ameri-
can Land Mammal bAgeQ around Unity are
described and mapped in detail here (Figs. 1
and 2). Few fossils have been recovered from
poor exposures of the Ironside Formation near
Ironside (Merriam, 1916) and Baker City, Oregon
(Downs, 1952). The Unity, Ironside, and Baker
City sequences and local faunas are similar to

Fig. 4. Badlands of the late Miocene Ironside Formation in central Windlass Gulch (95–135 m of measured section Fig. 2), northeast of Unity. A
gray airfall tuff is visible to the lower right and two thin white tuffs in the middle of these badlands.
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 101
better-known sequences and faunas of the Juntura
Formation near Juntura, Malheur County (Fig. 3;
Bowen et al., 1963; Greene, 1973; Fiebelkorn et
al., 1982; Johnson et al., 1996).
3.1. John Day Formation (late Eocene–early
Miocene)
The oldest rocks exposed near Unity are 300 m of
rhyodacitic tuff breccias, interpreted as lahars by Reef
(1983). He considered them Eocene in age, based on
fossil leaves (Rhamnidium chaneyi) and permineral-
Table 1
Paleosols of the Ironside and Juntura Formations and their classification
Pedotype Sahaptin
meaning
Diagnosis
Abiaxi Bitterroot Near-mollic surface (A) horizon over
weakly weathered subsurface (Bw)
Cmti New Shale and siltstone with root traces
Monana Underneath Thin impure lignite (O) on hale and
siltstone (A) with shallow root traces
Skaw Scare Thin dark gray to black spotted siltstone
with root traces (A) over bedded siltstone
Tutanik Hair Near-mollic brown thick surface (A) with
deep calcareous and siliceous
rhizoconcretions and nodules (Bk)
Xaus Root Sandstone with root traces (A) often
ferruginized over bedded sandstone
ized palm wood (Palmoxylon sp. indet.). However,
Walker (1990) reports K–Ar ages from hornblende and
plagioclase of 19.6F0.8 and 19.5F0.6 Ma, respec-
tively (early Miocene), and regarded these rocks as
outliers of the late Eocene to early Miocene John Day
Formation. The laharic tuff breccia is overlain by a 100-
m-thick rhyodacitic flow of similar petrographic and
geochemical composition to the tuff breccia, additional
tuff breccia, and local basalt flows (Reef, 1983;Walker,
1990). These volcanic rocks are overlain by about 20 m
of red claystone, including numerous paleosols of the
Luca pedotype, which are late middle Eocene to early
Australian
classification
(Stace et al., 1968)
F.A.O.
World Map
(F.A.O., 1974)
U.S. Soil
Taxonomy
(Soil Survey
Staff, 1999)
Brown clay Eutric Cambisol Haplosalid
Alluvial soil Eutric Fluvisol Fluvent
Humic gley Eutric Histosol Fibrist
Wiesenboden Eutric Gleysol Mollic
Endoaquent
Brown earth Chromic Luvisol Typic
Natrixeralf
Alluvial soil Eutric Fluvisol Psamment

Table 2
Paleosols of the Ironside and Juntura Formations and their interpretation
Pedotype Paleoclimate Past vegetation Past animal life Paleotopographic
setting
Parent
material
Time of
formation
Abiaxi No indication Early successional
wooded grassland
cf. Cormohipparion
sphenodus, Prosthennops,
Gomphotherium osborni
Riparian fringe
and low terraces
of floodplain
Volcaniclastic
silt
1000–2000
years
Cmti No indication Riparian early
successional
woodland
Proboscidea indet.,
Merycodus
Streamside levee
deposits
Volcaniclastic
silt
5–100
years
Monana No indication Swamp woodland None found Lake margin and
floodplain swamp
Volcaniclastic
silt and
diatomite
100–1000
years
Skaw No indication Riparian floodway
woodland:
Gramineae,
Equisetum,
Populus, Salix
Meterix, Scapanus,
Hypolagus, Spermophilus
junturensis, Diprionomys,
Eucastor malheurensis,
Peromyscus, Vulpes,
Mammut furlongi,
Prosthennops, Merychyus
Seasonally
waterlogged
streamside swale
Volcaniclastic
silt
50–1000
years
Tutanik Subhumid
(800–1000 mm
mean annual
precipitation),
summer-dry
Grassy woodland Mylagaulus, Mammut
furlongi, cf.
Cormohipparion
sphenodus, Aphelops,
Procamelus grandis,
Megatylopus
Well-drained
floodplain
Volcaniclastic
silt
2000–5000
years
Xaus No indication Riparian early
successional
woodland
Merycodus Streamside sand
bars
Volcaniclastic
sand
5–100
years
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123102
Oligocene (Uintan to Orellan) in age near Clarno and
the Painted Hills, Oregon (Bestland et al., 1999;
Retallack et al., 2000). Comparable red paleosols also
are found in the upper John Day Formation east of
Kimberly, Oregon, where they formed on well-drained
uplands (Retallack et al., 2000).
3.2. Strawberry and Dooley Volcanics (middle
Miocene)
Platy, 2-pyroxene, basaltic andesites are character-
istic of middle Miocene (radiometric ages 15–10 Ma),
Strawberry Volcanics, with smaller amounts of basalt,
rhyolite, rhyolitic tuff, vent breccia, and micronorite
(Robyn, 1977). The Strawberry Volcanics are calcalka-
line and were not comagmatic with voluminous
tholeiitic basalts of the Columbia River Basalt Group,
which also are middle Miocene (radiometric age 17–15
Ma; Sheldon, 2003). Calcalkaline volcanism is pro-
duced by melting above subduction zones, but Straw-
berry Volcanics and other coeval andesitic volcanoes
(Walker, 1990) were far inland from subduction of the
Farallon Plate during the middle Miocene (Orr et al.,
1992). Middle Miocene Strawberry stratovolcanoes
rose at least 1000 m above valley floors of the
Paleozoic and Mesozoic basement (Robyn, 1977).
Dooley Volcanics near Unity are 150 m thick and
include three distinct units of rhyolitic, welded, ash
flow tuffs (Reef, 1983). The ash flow tuffs are light
gray with abundant pumice, rock fragments, and
phenocrysts of plagioclase, biotite, and quartz.
Dooley Volcanics reach a thickness of 2400 m
around volcanic centers only a few kilometers to the
north and northwest of the mapped area. One of the
last eruptive events of the Dooley Volcanics was a
rhyolite dome dated by K–Ar at 14.7F0.4 Ma
(Evans, 1992).
3.3. Ironside Formation (late Miocene)
The Ironside Formation is at least 250 m of
conglomerates, siltstones, diatomites and lignites

Fig. 5. Petrographic and chemical data for newly described paleosols of Tutanik and Xaus paleosols from 4 to 6 m, Tutanik, Cmti, Xaus, Skaw,
and Abiaxi paleosols from 41 to 46 m, and Abiaxi, Cmti and Skaw paleosols from 229 to 232 m in reference section of Ironside Formation in
Windlass Gulch (Fig. 2).
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 103

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123104
named for the small village of Ironside, 15 km east
on U.S. highway 26 from Unity (Thayer, 1957;
Thayer and Brown, 1973). Sediments and paleosols
along the highway east of Ironside are identical with
those near Unity and particularly with the lower part
of a measured section in Windlass Gulch (0–7 m in
Fig. 2). Diatomites like those in the upper part of
the Ironside Formation near Unity, also crop out in
the hills north of Ironside. A fossil locality southeast
of Baker City also exposes tuffs and conglomerates
like those of the lower Ironside Formation (95–100
m in Fig. 2) in Windlass Gulch (Downs, 1952;
Evans, 1992).
These three areas are linked by late Miocene
fossil mammals of the Clarendonian North American
Land Mammal "Age" (8.4–12.3 Ma of Prothero,
Fig. 6. Late Miocene (Clarendonian) fossil mammals from Unity: (A)–(B
and 9017, respectively); (C)–(D) Gomphotherium osborni, skull and mandi
indet., molar with chipped crown (JODA9000). Scale bars as indicated.
1998), although nomenclatural difficulties remain.
For example, a fossil proboscidean skull from Unity
on display at the Oregon Museum of Science and
Industry in Portland (Fig. 6C–D) has remained
undescribed beyond a photograph and account of
its collection (Baldwin, 1964; Orr and Orr, 1999).
The excavation is still visible within an Abiaxi
paleosol on a low knoll southeast of a prominent
bluff, east of Unity (locality 8 of Table 4; Walker,
1990, figs. 5 and 9). George Gaylord Simpson
examined the specimen in 1953 and suggested it was
bMiomastodon merriamiQ (Orr and Orr, 1999). A more
appropriate name is bGomphotherium cingulatumQof Downs (1952), based on a lower jaw discovered
by Albert Werner at a locality 15 km east of Baker
City and about 80 km northeast of Unity. This and
) Hipparionini gen. indet., molar fragments (JODA specimens 9004
ble (Oregon Museum of Science and Industry); (E) Prosthennops sp.

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 105
other validly established taxa were not recognized
by Lambert and Shoshani (1998), who considered
the genus oversplit. Tobien (1972) went so far as to
include all North American Gomphotherium in
bGomphotherium productumQ. Here, I follow Mad-
den and Storer (1985) in using their broadly defined
Gomphotherium osborni (Barbour) for the Unity
skull. Another gomphothere tooth, probably con-
specific, was found by Elmer Molthan in the
Ironside Formation 400 m south of Ironside Post
Office, which is only 15 km east of Unity
(bTetrabelodonQ sp. of Merriam, 1916). Gomphothe-
Fig. 7. Fossil plants and fish from the Ironside Formation near Unity, Orego
box elder, Acer negundoides, winged seed (JODA9409); (C) white o
pollardiana leaf (JODA9058a); (E) shagbark hickory, Carya bendirei, l
(JODA9050a); (G) bullhead catfish, Ictalurus (Ameiurus) vespertinus, sku
rium and two additional proboscidean species are
recognized from the Clarendonian Juntura Formation
to the south in Malheur County (Shotwell and
Russell, 1963): a two-tusker bMammut furlongiQ (a
species not recognized by Lambert and Shoshani,
1998), and a shovel-tusker similar to Platybelodon
barnumbrowni (but regarded as a new genus by
Lambert and Shoshani, 1998).
There are also nomenclatural problems with
Clarendonian horse teeth from Unity and Juntura,
because among the numerous teeth and jaws, there is
no skull with facial fossae or other diagnostic
n: (A) sycamore, Platanus dissecta, leaf (specimen JODA9408); (B)
ak, Quercus prelobata leaf (JODA9053); (D) live oak, Quercus
eaf (JODA9055); (F) swamp ash, Fraxinus dayana, winged seed
ll and partial skeleton (JODA9043a). Scale bars all 1 cm.

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123106
features. The names given to these teeth (bHipparionanthonyiQ of Merriam, 1916 and bHipparion con-
doniQ of Shotwell and Russell, 1963) are considered
by MacFadden (1984) likely junior synonyms of
Cormohipparion occidentale or Cormohipparion
sphenodus, the latter most likely on the basis of
size. Shotwell and Russell (1963) considered that
the large collection of teeth and jaws from
Juntura represented only one species.
Late Miocene geological age is also indicated by
fossil plants from near Unity and Juntura. A new
fossil plant assemblage in Juniper Gulch near Unity
(Fig. 7; locality 6 of Table 4, at 160 m in Fig. 2) is
here designated the Unity flora. Entire to weakly
dentate oak leaves like those dominating this
assemblage have been identified as bQuercus
hannibaliQ, but this name has been included within
Quercus pollardiana (Knowlton) by Axelrod (1995)
or identified with the similar, but more dentate,
living Quercus chrysolepis (Wolfe for Leopold and
Wright, 1985). The Unity flora is taxonomically
most like the late Miocene Stinkingwater flora in the
Juntura Formation near Juntura (Chaney and Axel-
rod, 1959) and the Weiser flora from the Poison
Creek Formation of Idaho (Dorf, 1936; Leopold and
Wright, 1985).
The thick gray volcanic ash of the Ironside
Formation at 171 m in Windlass Gulch is minera-
logically most similar to the 11.31 Ma CPTXI Ash of
the Great Basin (Perkins et al., 1998), and another
gray tuff at 101 m is most like the 11.59 Ma CPTIX
Ash (Figs. 2 and 3). A geological age of 11–12 Ma is
similar to the age of the Juntura Formation, 78 km to
the south in Malheur County. A whole-rock K–Ar
date for 12-m-thick basalt 152 m stratigraphically
below a fossil mammal quarry (OU locality 2337 of
Fig. 3) and 85 m above the Stinkingwater flora
(Chaney and Axelrod, 1959) near Juntura is 12.4 Ma
(Evernden and James, 1964; corrected using Dal-
rymple, 1979). A tuff 76 m stratigraphically above
the mammal quarry has been dated by K–Ar on
shards at 11.5F0.6 Ma (Fiebelkorn et al., 1982), and
the Devine Canyon ashflow tuff at the base of the
Drewsey Formation, 107 m above the quarry, has
been dated by 39Ar/40Ar of sanidine as 9.7 Ma
(Johnson et al., 1996; five similar K–Ar dates are
tabulated by Greene, 1973). Also comparable in age
and lithology are volcanic ashes, conglomerates, and
diatomites of the Coal Valley Formation of Nevada
dated by the K–Ar technique at 11.4–9.4 Ma (Golia
and Stewart, 1984).
4. Late Miocene paleogeography of eastern Oregon
4.1. Sedimentological evidence
The depositional basin of the Ironside Forma-
tion was bounded by volcanic terranes in the
Strawberry and Dooley Volcanics to the north and
west, and fault blocks of Paleozoic and Mesozoic
basement to the north and southwest. There was
erosional paleorelief on the underlying Dooley
Volcanics and Clarno Formation, as indicated by
several inliers cropping out south of Juniper
Gulch. The stepped system of northeast-extensional
faults (Fig. 1) did not exist during deposition,
when this broad alluvial and lacustrine basin
extended southeast from highlands to the north
and west (Fig. 8).
Basin-marginal boulder breccias and conglomer-
ates interfinger with diatomites and tuffaceous
siltstones of the Ironside Formation in road cuts
over Dooley Mountain and north of Whited
Reservoir (Fig. 1). Paleocurrent directions measured
from trough cross bedding in conglomeratic paleo-
channels in the Windlass Gulch section (Fig. 9)
indicate that streams flowed southeast. This early
phase of river channels and floodplains was
followed by extensive oligotrophic lakes, which
eventually accumulated diatomites out to the hilly
margins of the depositional basin. A final deposi-
tional phase of diminished lake area is recorded by
fluvial gravels and lignites at the top of the exposed
sequence near Unity (Fig. 2).
4.2. Paleosol evidence
Various paleosols represent different sedimentary
environments and communities within these river
valleys and lake shores. Weakly developed paleosols
(Abiaxi, Cmti, Xaus) represent ephemeral commun-
ities on the landscape, such as vegetation colonizing
stream and lake margins after flooding. Weakly
developed carbonaceous paleosols (Skaw) represent
riparian or lake-margin marsh, and moderately

Fig. 8. Reconstructed late Miocene (Clarendonian) landscape ecology of the area near Unity, eastern Oregon.
Fig. 9. Paleocurrents measured from trough cross bedding in
conglomerates at 123–131 m (left) and 33–39 m (right) in the
measured section in Windlass Gulch (Fig. 2).
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 107
developed peaty paleosols (Monana) represent
swamp woodlands, where decay of organic debris
was suppressed by seasonally high water table.
In contrast, Tutanik paleosols represent stable
floodplains and alluvial terraces. Tutanik paleosols
are thick, brown, and have little relict bedding due to
bioturbation (including siliceous rhizoconcretions;
Fig. 10A–B) and to development of soil structure
(fine subangular blocky peds) and microfabric (skel-
masepic porphyroskelic plasmic fabric of Brewer,
1976). They are also more aluminous and less rich in

Fig. 10. Chalcedony nodules and rhizoconcretions from Tutanik paleosols of the Ironside Formation near Unity, Oregon: (A) petrographic thin
section under crossed nicols of three rhizoconcretions (black holes with bright banded haloes) in oblique (lower left) and cross-sections
(specimen JODA9225); (B) rhizoconcretion (JODA9311); (C) near-spherical nodule (JODA9310a); (D) lobed nodule (JODA9310b); (E) cigar-
shaped nodule (JODA9309a); (F) tear-shaped nodule (JODA9310c); (G) cross-section of septarian nodule (JODA9310d); (H)–(I) petrographic
thin sections under crossed nicols of dendritic chalcedony and void-filling dolomite (JODA9310d). Scale bars as indicated.
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123108
alkalis and alkaline earths than associated paleosols
(Fig. 5). Although volcanic ash was an important part
of Tutanik paleosol parent material, few recognizable
shards have escaped weathering. This degree of
weathering is found after 8–27 ka in tropical highland
soils of New Guinea (Ruxton, 1968). Tutanik
paleosols have common clay skins in thin section
and a higher proportion of fine pedogenic clay than
associated paleosols (Fig. 5). The degree of enrich-
ment of clay is intermediate between that seen in
upper Modesto soils dated at 10 Ka and lower
Modesto soils dated at 40 Ka in alluvium of the
Merced River, San Joaquin Valley, California
(Harden, 1982, 1990).
4.3. Paleontological evidence
Diatomites near Unity have not been studied in
detail, but of 108 diatom taxa described from coeval
diatomites of Juntura, only two are found in estuaries
and the sea: bCoscinodiscus miocenicusQ and
bCoscinodiscus subtilisQ of Van Landingham (1967;
perhaps better referred to Actinocyclus according to
Bradbury et al., 1985). Melosira granulata, Melosira
distans, and Fragilaria construens dominate individ-
ual collections (58–88%) and indicate fresh water of
low alkalinity, low in dissolved solids, neutral to
slightly acidic in pH, and low in nutrient levels
(Bradbury et al., 1985). Significant (8%) amounts of
C. subtilis in one sample (Van Landingham, 1967)
may indicate connection to the sea by way of low-
gradient streams (Bradbury et al., 1985). Fossil
aquatic molluscs from Juntura and Unity (Tables 4–
5; Taylor, 1985) are evidence for a Miocene course of
the Snake River southwest into the Pit River of
northern California, before Basin and Range faulting
and northward capture by the Columbia River (Van
TassellL et al., 2001). Lakes were also needed by
many of the fossil birds (Brodkorb, 1961): cormor-
ants, coots, mergansers, teals, and extinct straight-
billed flamingos (Table 5).
5. Late Miocene paleoclimate of eastern Oregon
5.1. Evidence from paleosols
Only Tutanik paleosols are useful paleoclimatic
proxies, because other pedotypes were too weakly
developed to show a paleoclimatic signature. Differ-
ences between Tutanik and other comparably devel-
oped Cenozoic paleosols from Oregon may reflect
changing Neogene paleoclimate. Tutanik paleosols
are generally similar in their brown color and clayey
subsurface (argillic horizons) to Skwiskwi paleosols
of the Oligocene John Day and latest Miocene

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 109
Rattlesnake Formations (Retallack et al., 2000,
2002a), and to early Miocene Tima paleosols of
the John Day Formation (Retallack, 2004). Tutanik
paleosols lack thick silcretes of Tima paleosols and
have silica–micrite rhizoconcretions (Fig. 8A–B) and
chalcedony septarian nodules (Fig. 8C–I) unknown
in Skwiskwi paleosols. The large amount of silt
(never less than 27% by volume) and sodium (soda/
potash ratiosN1) also ally Tutanik paleosols with
Tima rather than Skwiskwi paleosols. Tutanik
paleosols also differ from paleosols of the late
Miocene (7 Ma) Rattlesnake Formation, such as
the Tatas pedotype, which was crumb-structured,
moderately leached, and calcareous (Retallack et al.,
2002a). Tutanik paleosols thus show greater affin-
ities with interpreted summer-dry (xeric) paleosols,
such as Tima, than summer-wet (ustic) paleosols,
such as Swkiskwi and Tatas.
Modern soils like Tutanik paleosols are found
near Redding, northern California (map unit Lc3-2a
with duric phase of F.A.O., 1975), where mean
annual temperature is 17.7 8C, and mean annual
precipitation is 1040 mm, January and July temper-
atures are 8 and 29 8C, respectively, and precip-
itation is 216 mm in January, but only 5 mm in July
(Ruffner, 1985). In contrast, for Baker City near
Unity, mean annual temperature is 7.6 8C, mean
annual rainfall is 270 mm, temperature is more
seasonal (�4 8C in January, 19 8C in July), but
precipitation is less seasonal (35 mm in January, 12
mm in July; Ruffner, 1985).
Independent estimates of former precipitation can
be gained from chemical composition of clayey
horizons and from carbonate in rhizoconcretions of
Tutanik paleosols at depths of 79–91 cm. This
depth (D in cm) can be used to estimate mean
annual rainfall (P in mmF156) using a transfer
function derived from study of modern soils
(Retallack, 1994), once allowance is made for burial
compaction (Sheldon and Retallack, 2001) due to at
least 60 m of overlying Pleistocene fanglomerate
(Evans, 1992). A large database of chemical
analyses of North American soil B horizons has
been used to derive yet other transfer functions for
mean annual precipitation (P in mmF182) and
mean annual temperature (T in 8CF4.4) from two
chemical measures: S, the molar ratio Na2O+K2O/
Al2O3; and C, molar Al2O3/(Al2O3+CaO+Na2O)
times 100 (following Sheldon et al., 2002), as
follows:
P ¼ 139:6þ 6:388D� 0:01303D2
P ¼ 221e0:02C
T ¼ � 18:516 Sð Þ þ 17:298:
Paleoclimatic results of these calculations (Table 3)
are 450–800 mm mean annual precipitation from
depth to carbonate and 600–1100 mm from chemical
composition of clayey horizons. Because the carbo-
nate-depth transfer-function was designed for use with
nodules (Retallack, 1994, 2000), rather than rhizo-
concretions, these estimates are less likely to be
reliable than the subhumid regime estimated from
chemical composition of B horizons. Geochemical
composition of paleosol parent rhyodacitic and
andesitic airfall ash shows insignificant change
through time compared with paleosol modification
(Perkins et al., 1998; Retallack et al., 2000).
A highly seasonal climate is indicated by concen-
trically banded rhizoconcretions with silica and
micrite in Tutanik paleosols (Fig. 10A–B). Season-
ality is also indicated by the unusually abundant silt
and soda in Tutanik paleosols, especially considering
their likely mean annual precipitation (Table 3). In
such rainy climates, there would not be so much salt
or dust unless summers were very dry, and most rain
and snow were during winter and spring. Furthermore,
the scarce carbonate and distinctive silica–micritic
rhizoconcretions (Fig. 10A–B) are like those found in
modern soils of Mediterranean (summer-dry) climates
(Chadwick et al., 1987, 1995). Siliceous rhizoconcre-
tions from a few Tutanik paleosols (45, 59, and 69 m
in Fig. 2) grade insensibly through cigar- and tear-
shaped forms into ellipsoidal septarian nodules of a
form not previously reported from paleosols (Fig.
10C–F). Some of these contain fossil wood or lumpy
root traces morphologically comparable to rhizobially
nodulated roots. Others are hollow and cracked, with
void-filling dolomite rhombs (Fig. 10G,I). Micro-
fabric of the nodules is radially oriented, dendritic
chalcedony, creating an external fabric reminiscent of
cone-in-cone structure. The ellipsoidal nodules are
superficially similar to chalcedony nodules of the
bbutton bedsQ, a lacustrine facies of the middle

Table 3
Mean annual temperature and precipitation for Tutanik paleosols of the Ironside Formation
Estimated geological
age (Ma in Fig. 5)
Stratigraphic level
(m in Fig. 3)
Mean annual precipitation
(mm) from Bk
Mean annual precipitation
(mm) from chemical
composition
Mean annual temperature
(8C) from chemical
composition
11.59 100.5 644F156 (503–785)
11.60 Unity Reservoir 657F156 (516–798)
11.69 83 588F156 (447–729) 916F182 (734–1098) 13.3F4.4 (8.9–17.7)
11.83 45.1 601F156 (460–742) 934F182 (752–1116) 13.2F4.4 (8.8–17.6)
11.95 16.6 643F156 (502–784)
11.96 14 617F156 (476–758)
11.97 12.5 626F156 (485–767)
11.00 Ironside 640F156 (499–781)
12.00 5.2 627F156 (486–768) 786F182 (604–968) 12.1F4.4 (7.7–16.5)
12.01 3.3 636F156 (495–777)
12.02 1.7 649F156 (508–790)
12.10 Denny Flat 571F156 (430–712)
12.11 Denny Flat 594F156 (453–735)
Means All listed 623F156 879F182 12.9F4.4
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123110
Miocene Barstow Formation of California (Palmer,
1957). My own examination of the Barstow nodules
found conspicuous relict bedding and microcrystalline
textures, unlike the septarian and radial-dendritic
crystal form of the Unity nodules. Nevertheless,
highly alkaline late-summer aridity could have played
a role in the formation of both kinds of siliceous
nodules.
5.2. Evidence from fossil plants
Close floristic composition of Unity (Table 5)
and Stinkingwater floras (Chaney and Axelrod,
1959) with the modern flora of California supports
evidence from associated paleosols for extension of
Mediterranean climate into eastern Oregon during
the late Miocene. The paleoclimatic affinities of
these floras are quite distinct from more humid and
warm climates evident during the middle Miocene
thermal maximum, represented locally by the
Austin, Tipton, Baker, and Mascall floras (Oliver,
1934; Brown for Gilluly, 1937; Chaney and
Axelrod, 1959), and before the advent of modern
sagebrush and riparian communities during the
Pliocene (Chaney, 1948; Ashwill, 1983, Retallack
et al., 2002a). Mean annual precipitation of around
900–1000 mm and mean annual temperature of 13
(8–18) 8C have been estimated from other late
Miocene fossil floras of the Great Basin (Wolfe,
1994; Wolfe et al., 1997; Graham, 1999). A
summer-dry paleoclimatic regime for the Unity flora
is also compatible with frequent wildfires indicated
by charcoal associated with middle to late Miocene
fossil floras of southeastern Oregon (Taggart et al.,
1982; Taggart and Cross, 1990).
5.3. Evidence from fossil animals
Abundance of bullhead catfish in siltstones with
the Unity flora (Fig. 7; Table 4) can be contrasted
with the overlying diatomites, which have a sparse
fish fauna of other kinds of fish. The leaves and
catfish probably represent a nearshore lacustrine
habitat distinct from that of the open lake. Bullhead
catfish lived in western North America from the
Eocene to Pliocene, but after that time became
restricted to eastern North America. They have been
artificially reintroduced to Oregon and Idaho within
the last 120 years. Today, they live in low-gradient
streams and lakes at elevations less than 1000 m in
warm temperate climates with at least 230 frost-free
days and at least 400 mm mean annual precipitation
(Van Tassell et al., 2001).
Brodkorb (1961) invokes Bergmann’s rule (Brown
and Lomolino, 1998) to argue for a warmer than
present climate from the small size of Juntura
cormorants, coots, and teals compared with living
relatives. Oxygen isotopic studies of fossil mammal
teeth from Juntura, Oregon, indicate mean annual
temperatures of about 18 8C, and a mean annual range

Table 4
Late Miocene (Clarendonian) megafossils of the Ironside Formation, near Unity, Oregon
Fossil taxon Similar living taxon Common name Loc.
Equisetum sp. indet. Equisetum arvense Common horsetail 1–2
Gramineae sp. indet. Gramineae Grass 1–2
Sagittaria sp. indet. Sagittaria cuneata Arum-leaved arrowhead 3
Salix sp. indet. Salix scouleriana Scouler’s willow 3
Populus sp. indet. Populus balsamifera Black cottonwood 3
Trapa americana Knowlton (1898) Trapa natans Water chestnut 3–5
Platanus dissecta Lesquereux (1878) Platanus occidentalis Sycamore 6
Quercus pollardiana (Knowlton) Axelrod (1995) Quercus chrysolepis Maul oak 6
Quercus prelobata Condit (1944) Quercus garryana Oregon white oak 6
Quercus simulata Knowlton (1898) Quercus myrsinaefolia Chinese oak 6
Cercocarpus ovatifolius Axelrod (1985) Cercocarpus betuloides
var. blancheae
Birchleaf mountain
mahogany
6
Acer negundoides MacGinitie (1934) Acer negundo Box elder 6
Fraxinus dayana Chaney and Axelrod (1959) Fraxinus caroliniana Swamp ash 6
Carya bendirei (Lesquereux) Chaney and Axelrod (1959) Carya ovata Shagbark hickory 6
Ptelea miocenica Berry (1931) Ptelea trifoliata Carolina hop tree 7
Radix junturae Taylor (1963) Radix auriculata Aquatic big-ear snail 7
Ictalurus (Ameiurus) vespertinus Miller and Smith (1967) Ictalurus melas Black bullhead catfish 6
Gomphotherium osborni (Barbour) Madden and Storer (1985) None Extinct four-tusker 8
Prosthennops sp. indet. None Extinct large peccary 8
Rhinocerotidae sp. indet. None Large extinct rhino 9
cf. Cormohipparion sphenodus (Cope)
MacFadden and Skinner (1977)
None Extinct cursorial
three-toed horse
10
cf. Megatylopus gigas Matthew and Cook (1909) None Extinct large camel 11
cf. Merycodus sp. Antilocapra americana Pronghorn 11–12
Note: these are identifcations of newly collected material in collections of John Day Fossil Beds National Monument. Legal and NAD27 UTM
coordinates to localities are 1—Windlass Gulch section 60 m NWH NEH SEH SEH S33 T12S R38E 11T 0416253 4925216; 2—Windlass
Gulch section 44 m NWH NEH SEH SEH S33 T12S R38E 11T 0416262 4925216; 3—Windlass Gulch section 61 m NWH NEH SEH SEH
S33 T12S R38E 11T 0416262 4925216; 4—Denny Flat southeast SEH SEH SEH SWH S2 T13S R37E 11T 0409723 492339; 5—Denny flat
southeast NEH NEH NEH NEH S2 T13S R37E 11T 0409761 492353; 6—Windlass Gulch section 158 m NWH NWH SWH SWH S34 T12S
R38E 11T 0416526 495228; 7—Windlass Gulch section 225 m NEH NWH SWH S34 T12S R38E 11T 0416665 4925404; 8—east of Unity
SEH NEH SEH NWH S11 T13S R37E 11T 0408804 4922516; 9—Windlass Gulch section 100 m NEH SWH SWH SWH S34 T12S R38E
11T 0416596 4925185; 10—Windlass Gulch section 5 m SEH NWH SEH SEH S33 T12S R38E 11T 0416227 4925153; 11—Denny Flat
northwest NWH SWH SWH SWH S35 T12S R37E 11T 0408416 4925224; 12—Windlass Gulch section 89 m NEH NEH SEH SEH S33 T12S
R38M 11T 0416315 4925249.
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 111
of temperature of some 25 8C (Kohn et al., 2002),
compared with 7.6 and 23 8C, respectively, for BakerCity today (Ruffner, 1985).
Indications of a late Miocene climate at Juntura,
wetter than at present, come from the sewellel
(Tardontia in Table 5). Similar aplodontids are
now restricted to Oregon and Washington, west of
the Cascades (Shotwell, 1958). The western Amer-
ican mole (Scapanus) is also now found well west
of Juntura and Unity (Hutchison, 1968). On the
other hand, subhumid to semiarid rangeland con-
ditions are indicated by ground squirrels, pocket
gophers, and extinct mylagaulines (Shotwell, 1958,
1970).
6. Late Miocene palaeoecology of eastern Oregon
6.1. Evidence from paleosols
A detailed local mosaic of vegetation (Fig. 8)
can be inferred from the relationship of paleosols to
local sedimentary facies and from plant-induced
features of the paleosols (Figs. 2 and 3; Table 3).
Tutanik paleosols are finely structured (granular
peds about 1 cm across) with abundant fine root
traces as well as scattered rhizoconcretions, as in
soils supporting grass with scattered woody shrubs
or trees (Retallack, 1997b, 2001b). The paleosols
lack fine crumb peds found in sod-forming grass-

Table 5
Late Miocene (Clarendonian) megafossils of the Juntura Formation
near Juntura
Fossil taxon Common name
Sphaerium sp. cf. S. lavernense
Herrington (Herrington and
Taylor, 1958)
Aquatic lake orb
mussel
Pisidium sp. cf. P. clessini
Neumayr and Paul (1875)
Aquatic pea clam
Pisidium sp. indet. Aquatic pea clam
Fluminicola junturae Taylor, 1963 Aquatic pebble snail
Hydrobiidae indet. Small aquatic snail
Viviparus turneri Hannibal, 1912 River snail
Radix junturae Taylor, 1963 Aquatic big-ear snail
Carinifex shotwelli Taylor, 1963 Keeled ramshorn snail
Promenetus sp. indet. Aquatic ramshorn snail
Ictalurus peregrinus Lundberg, 1975 Catfish
Cyprinidae gen. indet. Squawfish
Catastomidae gen. indet. Sucker
Phalacrocorax leptopus
Brodkorb, 1961
Small cormorant
Ciconiidae gen. indet. Extinct stork
Megapaloelodus opsigonus
Brodkorb, 1961
Straight-billed
early flamingo
Eremochen russelli
Brodkorb, 1961
Small goose
Querquedula pullulans
Brodkorb, 1961
Small teal
Ocyplonessa shotwelli
Brodkorb, 1961
Extinct merganser
Neophrontops dakotensis
Compton, 1935
Extinct Old
World vulture
Fulica infelix Brodkorb, 1961 Small coot
Meterix sp. indet. Extinct large shrew
Anouroneomys minimus Hutchison
& Bown (Bown, 1980)
Extinct small shrew
Mystipterus sp indet. Extinct nonburrowing
mole
Scalopoides sp. indet. A Extinct weakly
burrowing mole
Scalopoides sp. indet. A Extinct weakly
burrowing mole
Scapanus sp. cf. S. shultzi
Tedford, 1961
Western American
mole
Chiroptera gen et sp. indet. Extinct bat, like big
brown bat
Hesperolagomys sp. cf. H. galbreathi
Dawson (Clark et al., 1964)
Extinct pika
Hypolagus sp. cf. H. fontinalis
Dawson, 1958
Extinct rabbit
Tardontia sp. cf. T occidentale
(Macdonald) Shotwell, 1958
Extinct sewellel
Mylagaulus sp. indet. Extinct horned rodent
Epigaulus minor Hibbard and
Phillis (1945)
Extinct horned rodent
Fossil taxon Common name
Ammospermophilus junturensis
(Shotwell & Russell) Korth, 1994
Ground squirrel
Ammospermophilus sp. indet. Ground squirrel
Eutamias sp. indet. Extinct chipmunk
Hystricops sp. indet. Extinct large beaver
Eucastor malheurensis Shotwell
and Russell (1963)
Extinct beaver
Microtoscoptes sp indet. Extinct vole
Copemys dentalis (Hall)
Lindsay, 1972
Extinct small field
mouse
Copemys sp. cf. C. esmeraldensis
(Wood) Lindsay, 1972
Extinct large field
mouse
Copemys sp indet. Extinct large field
mouse
Leptodontomys sp. indet. Extinct pocket mouse
Perognathus sp. indet. Pocket mouse
Diprionomys sp. indet. Extinct pocket mouse
Pliosaccomys sp. indet. Extinct pocket gopher
Macrognathomys nanus Hall, 1930 Extinct birch mouse
Borophagus sp. indet. Short-face bone-
crushing dog
Aelurodon sp. indet. Long-face bone-
crushing dog
Vulpes sp. indet. Fox
Leptarctus sp. indet. Extinct badgerlike
mustelid
Hoplictis sp. indet. Extinct like honey
badger
Sthenictis junturensis Shotwell and
Russell (1963)
Extinct otterlike
musteline
Pseudailurus sp. indet. Extinct small cat
Gomphotherium sp. indet. Extinct four-tusk
proboscidean
Mammut furlongi Shotwell and
Russell (1963)
Extinct two-tusk
mastodon
cf. Platybelodon barnumbrowni
(Barbour) Barbour, 1932
Extinct shovel-tusker
cf. Cormohipparion sphenodus (Cope)
MacFadden and Skinner (1977)
Extinct three-toed
horse
Tapiridae gen. indet. Tapir
Aphelops sp. indet. Extinct hornless
rhinoceros
Prosthennops sp. indet. Extinct large peccary
Merychyus major Leidy, 1858 Extinct piglike oreodon
Procamelus grandis Gregory, 1942 Extinct camel
Megatylopus sp. indet. Extinct large camel
Note: this is a compilation of the Black Butte local fauna of
Shotwell (1963, 1967, 1970), Hutchison (1968), and Taylor (1963)
as revised by Honey et al. (1998), Lambert and Shoshani (1998),
Baskin (1998), Martin (1998), Lander (1998), and Wright (1998).
Table 5 (continued)
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123112
land soils (Mollisols, Phaeozems, Chernozems;
Retallack et al., 2002a), so that grasses were most
likely bunch grasses.

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 113
Other paleosols with prominent relict bedding
and large woody root traces supported plant
formations early in ecological succession after
disturbance. For example, Xaus paleosols of in-
channel sandy bars probably supported riparian
woodland, and Cmti and Abiaxi paleosols of levee
siltstones probably supported riparian gallery wood-
land. Cmti paleosols are thinner, less sodic, and
have more relict bedding than Abiaxi paleosols (Fig.
7), indicating a shorter period of vegetation growth
and successional development. Early successional
marsh ecosystems are represented by carbonaceous
clayey paleosols with small root traces and relict
bedding (Skaw pedotype). Swamp ecosystems are
represented by lignitic paleosols with large woody
root traces (Monana pedotype). The swamp paleo-
sols have impure coals and charcoal and were
probably seasonally dry, but there is no comparable
evidence of seasonal exposure of marsh paleosols,
which may have been associated with perennial
watercourses.
6.2. Evidence from fossil plants
Only one kind of paleosol contained preserved
fossil plants: weakly developed paleosols of the
Skaw pedotype (localities 1–3 of Table 4). The best
preserved fossil plants were found within laminated
diatomites and siltstones (localities 4–7 of Table 4).
These are interpreted as leaf litter of marsh soils
(Skaw pedotype) and leaves blown into an oligo-
trophic lake, respectively. In all cases, there is a bias
toward aquatic plants, such as water chestnut,
arrowhead, and horsetail.
Despite these taphonomic limitations, the Stin-
kingwater, Weiser, and Unity floras include locally
abundant fossil grasses and live oak comparable to
those of summer-dry parts of California today. The
Unity flora is most similar to interior live oak
woodland of Barbour and Major (1988) and canyon
live oak woodland of Sawyer and Keeler-Wolf
(1995). These oak woodlands are at moderate
elevation below the zone of abundant conifers on
the southwestern slopes of the Sierra Nevada
flanking the northern California Great Valley. The
oak trees are largely evergreen and up to 15 m high
in both closed and open formation. Their grassy
understorey is brown and dry for most of the
summer. Mesic elements in the Unity flora (Carya,
Ptelea) are not found in modern Californian live oak
woodlands. These middle Miocene holdover taxa are
evidence that late Miocene dry woodlands of Unity
were not exactly like Californian vegetation today.
No taxodiaceous conifers were found near Unity,
where they would be expected considering charcoal
and woody root traces in Monana paleosols. These
swamp conifers are common in the coeval Stinking-
water flora of Juntura (Chaney and Axelrod, 1959)
and the Weiser flora of Idaho (Dorf, 1936).
Fossil grasses are represented by sheathing leaves
near Unity (Table 4) and by abundant pollen in coeval
rocks of Idaho (Leopold and Wright, 1985). Miocene
grasses of the Great Plains and Oregon are unlikely to
have been floristically similar, given differences
between fossil dicot floras of Oregon and the Great
Plains (Thomasson, 1979; Graham, 1999; Strfmberg,
2002) and differences between sodic–silicic Miocene
paleosols of Oregon (Retallack, 2004) and calcareous
Miocene paleosols of the Great Plains (Retallack,
1997b, 2001b). A comparable modern vegetation
distinction is the western wheat grass (Agropyron
spicatum) rangeland province of northern California
and the Great Basin, and blue grama (Bouteloua
gracilis) grasslands of the western Great Plains
(Leopold and Denton, 1987).
6.3. Evidence from fossil animals
Quarry collections from Juntura formed the basis
for Shotwell’s (1963) pioneering study of Clarendo-
nian mammalian paleocommunities (Fig. 11), and
these quarries were revisited in order to examine
their paleosols (Fig. 3). Shotwell’s (1963) bpondbank communityQ came from Skaw paleosols (the
only pedotype found), whereas his bsavannacommunityQ came largely from sandstones immedi-
ately overlying a Tutanik paleosol. His bsavannacommunityQ was mainly camels, hipparionine
horses, two-tusker mastodon, and rhinoceros. The
bpond bank communityQ, which from paleosol and
sedimentological evidence appears more likely to
have been riparian woodland, was mainly fish and
turtles, but also had many beavers, as well as rabbits
and a variety of rodents and insectivores (Table 5).
The weak development and carbonaceous and
manganiferous composition of Skaw paleosols are

Fig. 11. Specimen numbers of fossil mammals of Clarendonian
bsavannaQ and bpond bankQ communities by Shotwell (1963), from
quarries of Fig. 3. Numerous fish and turtle bones from locality
2337 are not included in these figures.
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123114
evidence of seasonally inundated streamside swales
(Table 2), where fish were trapped and devoured by
predators.
Contrary to Shotwell (1963), Bernor et al. (1988)
concluded that coeval late Miocene (11 Ma)
hipparionine horses of Austria were forest browsers,
because forest leaves were found in associated lake
beds. This view is not supported by the abundance
of hipparionines in Tutanik paleosols and rarity in
Skaw paleosols of Juntura and Unity (Fig. 11).
Later studies of hipparionine tooth wear (Hayek et
al., 1992) and associated mammalian fauna (Webb,
1983) also support Shotwell’s (1963) conclusion that
hipparionines were grassland and grassy woodland
mixed feeders. Open grassy terrane was also
required by an extinct species of Old World vulture
from Juntura (Table 5).
Proboscideans are now associated with mosaics of
grassy and woody vegetation, which they maintain
by destructive feeding (Owen-Smith, 1988). A
diversity of proboscidean habitats may be expected
from the high diversity of Clarendonian probosci-
deans, including eight genera in North America and
three in the Unity and Juntura faunas (Tables 4–5).
Shovel-tuskers (Platybelodon) are commonly por-
trayed as aquatic shovelers, although tusk wear
patterns indicate scraping against bark, twigs, peb-
bles, or other hard debris (Lambert, 1992). Four-
tuskers (Gomphotherium) also show tusk wear from
bark stripping and digging, and two-tusk mastodons
(Mammut) are known from Pleistocene stomach
contents to have eaten conifer needles, cones, and
grass (Lambert and Shoshani, 1998). Associated
paleosols are Abiaxi for Gomphotherium, Tutanik
for Mammut, and probably Skaw for cf. Platybelo-
don (uncertainty comes from poor exposure and
inexact location). The inferred paleoenvironments of
these paleosols (Table 3) cannot be claimed to
represent habitat preferences based on only one or
two specimens of each species.
Paleosols also support the idea that late Miocene
rangelands of the Pacific Northwest were distinct from
those of the Great Plains (Leopold and Denton, 1987).
Late Miocene summer-dry siliceous alfisol and aridisol
paleosols of Oregon (Retallack et al., 2002a) can be
contrasted with summer-wet Mollisols of Nebraska
and Montana (Retallack, 1997b, 2001b). The hippar-
ionine province of the Pacific Northwest was distinct
from a Pliohippus province of southern California and
the Great Plains during the Clarendonian (Shotwell,
1963). Late Miocene (7 Ma) grazing Pliohippus in
Oregon appeared in calcareous Mollisol paleosols
reflecting an incursion of summer-wet Gulf Coast air
masses into eastern Oregon at that time (7.3 Ma;
Retallack et al., 2002a). The tridactyl mixed-feeding
hipparionines may reflect rigors of summer-dry range-
land vegetation botanically and structurally distinct
from summer-wet rangeland of monodactyl grazing
Pliohippus. Comparable differences in vegetation and
seasonality may also explain the numerical dominance
of foregut-fermenting ruminant camels over hindgut-
fermenting horses at Juntura (Fig. 11) and other
western Clarendonian faunas (Tedford and Barghoorn,
1993), whereas horses dominated Clarendonian
assemblages of Nebraska and Texas (Webb, 1983).
Muzzle morphology of the camels Procamelus and
Megatylopus has been taken to indicate that they were
browsers, perhaps mixed feeders, but not grazers
(Dompierre and Churcher, 1996). Similar dominance
of ruminants over perissodactyls is found in Kenyan
(Retallack et al., 2002b) and Greek (Solounias, 1981)
Miocene faunas. By reprocessing their cud, ruminants
are sustained by less-nutritious forage than perisso-
dactyls of comparable body sizes (Janis et al., 1994).
Summer-dry seasons in Kenya, Greece, and western
G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 115
North America selected for ruminant-dominated fau-
nas able to cope with seasonal shortages of forage,
whereas equid-dominated faunas of the Great Plains
and Gulf Coast were supported by year-round forage.
7. Implications for global change
7.1. Neogene paleoclimate in eastern Oregon
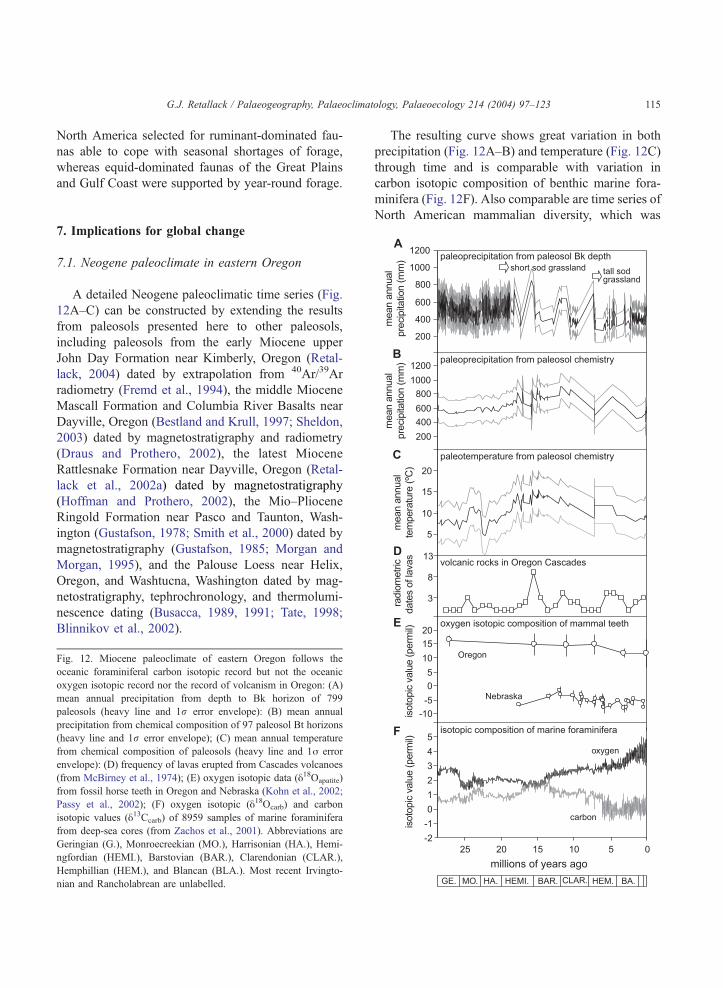
A detailed Neogene paleoclimatic time series (Fig.
12A–C) can be constructed by extending the results
from paleosols presented here to other paleosols,
including paleosols from the early Miocene upper
John Day Formation near Kimberly, Oregon (Retal-
lack, 2004) dated by extrapolation from 40Ar/39Ar
radiometry (Fremd et al., 1994), the middle Miocene
Mascall Formation and Columbia River Basalts near
Dayville, Oregon (Bestland and Krull, 1997; Sheldon,
2003) dated by magnetostratigraphy and radiometry
(Draus and Prothero, 2002), the latest Miocene
Rattlesnake Formation near Dayville, Oregon (Retal-
lack et al., 2002a) dated by magnetostratigraphy
(Hoffman and Prothero, 2002), the Mio–Pliocene
Ringold Formation near Pasco and Taunton, Wash-
ington (Gustafson, 1978; Smith et al., 2000) dated by
magnetostratigraphy (Gustafson, 1985; Morgan and
Morgan, 1995), and the Palouse Loess near Helix,
Oregon, and Washtucna, Washington dated by mag-
netostratigraphy, tephrochronology, and thermolumi-
nescence dating (Busacca, 1989, 1991; Tate, 1998;
Blinnikov et al., 2002).
Fig. 12. Miocene paleoclimate of eastern Oregon follows the
oceanic foraminiferal carbon isotopic record but not the oceanic
oxygen isotopic record nor the record of volcanism in Oregon: (A)
mean annual precipitation from depth to Bk horizon of 799
paleosols (heavy line and 1r error envelope): (B) mean annual
precipitation from chemical composition of 97 paleosol Bt horizons
(heavy line and 1r error envelope); (C) mean annual temperature
from chemical composition of paleosols (heavy line and 1j error
envelope): (D) frequency of lavas erupted from Cascades volcanoes
(from McBirney et al., 1974); (E) oxygen isotopic data (y18Oapatite)
from fossil horse teeth in Oregon and Nebraska (Kohn et al., 2002;
Passy et al., 2002); (F) oxygen isotopic (y18Ocarb) and carbon
isotopic values (y13Ccarb) of 8959 samples of marine foraminifera
from deep-sea cores (from Zachos et al., 2001). Abbreviations are
Geringian (G.), Monroecreekian (MO.), Harrisonian (HA.), Hemi-
ngfordian (HEMI.), Barstovian (BAR.), Clarendonian (CLAR.),
Hemphillian (HEM.), and Blancan (BLA.). Most recent Irvingto-
nian and Rancholabrean are unlabelled.
The resulting curve shows great variation in both
precipitation (Fig. 12A–B) and temperature (Fig. 12C)
through time and is comparable with variation in
carbon isotopic composition of benthic marine fora-
minifera (Fig. 12F). Also comparable are time series of
North American mammalian diversity, which was

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123116
higher during warmer and wetter times than during
cooler and drier times (Alroy et al., 2000; Janis, 2002).
7.2. Mismatch of Neogene terrestrial and marine
oxygen isotopic records
Neither paleoclimatic fluctuations from paleosols
(Fig. 12A–C) nor oxygen isotopic fluctuations from
mammal teeth (Fig. 12E) correspond with fluctua-
tions in oxygen isotopic composition of benthic
marine foraminifera (Fig. 12F). Marine and terrestrial
carbonate oxygen isotopic records do not correspond
during the late Paleocene (Koch et al., 2003) or late
Permian (MacLeod et al., 2000) either. Although
marine oxygen isotopic records have been regarded
as a global paleoclimatic standard (Zachos et al.,
2001), their mismatch with continental records is a
fundamental problem for global change studies.
The mismatch between North American mamma-
lian evolutionary dynamics and marine oxygen iso-
topic records has been discussed at length by Alroy et
al. (2000), who attribute the mismatch to biotic
insensitivity to climatic forcing. This is contrary to
the observation of iterative evolution of mammal
ecomorphs within North American land mammal
bagesQ which average 2.3 Myr in duration (Martin,
1994; Meehan and Martin, 1994; Meehan, 1996,
1999). The sediments of each mammal age in the
Great Plains are bounded at the base by erosional
downcutting, interpreted as evidence of a wet climatic
phase, and terminated by shallow calcic paleosols or a
caprock caliche, interpreted as evidence of a dry
climatic phase (Schultz and Stout, 1980). I have made
comparable observations in Montana and Oregon
(Retallack, 1997b, 2001a; Retallack et al., 2002a).
Late Oligocene mammalian communities of Oregon
also show alternation between sagebrush steppe with
Hypertragulus and wooded grassland communities
with Nanotragulus on Milankovitch time scales (41–
100 ka) within paleosols with differing depth to calcic
horizons reflecting alternating semiarid and subhumid
paleoclimates (Retallack, 2004; Retallack et al., 2004).
Finally, there is annual variation in oxygen isotopic
composition of growth bands within Miocene mam-
malian tooth enamel (Fox, 2001; Kohn et al., 2002).
North American mammals show clear sensitivity to
climate on time scales ranging from evolutionary to
ecological contrary to Alroy et al. (2000).
A possible reason for mismatch of marine oxygen
and continental paleoclimatic records is the Cenozoic
evolution of C4 photosynthetic pathways in land
plants. The C4 (Hatch–Slack) pathways of tropical
grasses result in isotopically heavier plant carbon than
C3 (Calvin–Benson) pathways (Cerling et al., 1997),
and this pathway increases oxygen isotopic values of
plant as well because operating on CO2 (Farquhar et
al., 1993). The antiquity of the C4 pathway has been
estimated from phylogenetic analyses at 25 Ma
(Kellogg, 1999), from isotopic analyses of paleosols
at 15 Ma (Kingston et al., 1994; Morgan et al., 1994),
and from isotopic and anatomical studies of permin-
eralized fossil grasses at 12.5 Ma (Nambudiri et al.,
1978; age revised by Whistler and Burbank, 1992).
Isotopic studies of paleosol carbonate and mammalian
teeth show that C4 grasses became widespread in the
southern Great Plains after 6.6 Ma but were never
common in Oregon or the northern Great Plains
(Cerling et al., 1997; Passy et al., 2002; Fox and
Koch, 2003). C4 grasses avoid cool and summer-dry
climates today (Sage et al., 1999). The spread of C4
grasses after 6.6 Ma is too late to explain the earlier
divergences of marine carbon and oxygen isotopic
records (Fig. 12F), mismatch of marine and terrestrial
oxygen isotopic records (Fig. 12A–C,F), and the
generally upward trend of marine oxygen isotopic
values (Fig. 12F).
A more likely explanation for mismatch of the
oceanic oxygen isotopic and continental paleocli-
matic proxies is isotopic depletion as rain shadows
spread and intensified with uplift of the Cascade
volcanic and other western mountain ranges (Kohn
et al., 2002). The idea of Cenozoic drying by means
of an orographic rain shadow has been a traditional
explanation for pronounced Neogene aridity evident
from fossil plants (Chaney, 1948; Ashwill, 1983).
The growth of the Oregon Cascades can be inferred
from numerous radiometrically dated eruptions (Fig.
12D; McBirney et al., 1974; Priest, 1990), but this
does not match either marine or continental records
(Fig. 12A–F). The Cascade and Klamath Mountain
rain shadow has been important in creating generally
dry long-term climate in eastern Oregon since the
late Oligocene (30 Ma; Retallack et al., 2000).
Rocky Mountain barriers isolated eastern Oregon
from Gulf Coast cyclonic circulation since at least
early Miocene (19 Ma), as indicated by the

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 117
appearance of sodic–silicic rather than calcic paleo-
sols (Retallack, 2004) and very different oxygen
isotopic composition of fossil horse teeth in Oregon
and Nebraska (Fig. 12E). Fossil floras from the
northern Great Basin indicate high altitudes since at
least the early Miocene, for example, 2100 m for the
middle Miocene 49 Camp flora of northwestern
Nevada (Wolfe et al., 1997). Eastern Oregon was
elevated during Miocene initial eruptions of the
Yellowstone hot spot (Humphreys et al., 2000).
The most important reason for a mismatch of
oceanic and continental records is isotopic enrichment
of oceanic oxygen by growth of isotopically depleted
polar ice caps, which has long been recognized to
compromise direct paleotemperature interpretation of
marine oxygen isotopic values (Zachos et al., 2001).
This factor is not unrelated to the growth of mountains
with their expanding rain shadows, because montane
ice caps enrich the isotopic composition of atmos-
pheric oxygen as well. Ice cap fluctuation in volume
introduces a highly variable bias into paleotemper-
ature interpreted from the oxygen isotopic record of
marine benthic foraminifera. This bias can be cor-
rected using independent estimates of paleotemper-
ature from Mg/Ca ratios of foraminifera (Lear et al.,
2000), but this is compromised by dissolution and
other forms of diagenesis (de Villiers, 2003), and by
crustal recycling and other long-term influences on
ocean chemistry (Veizer et al., 2000).
7.3. Neogene greenhouse–icehouse fluctuation and its
causes
Greenhouse mechanisms for Neogene paleocli-
matic variation are suggested by correspondence of
the Oregon paleosol sequence (Fig. 12A–C) with the
marine signal of carbon sequestration inferred from
benthic foraminiferal y13C decrease and carbon oxi-
dation inferred from benthic foraminiferal y13Cincrease (Fig. 12F). High carbon isotopic values in
the middle Miocene ocean correspond to warm-wet
conditions in Oregon, whereas low carbon isotopic
values in the Plio–Pleistocene ocean corresponded to
cool-dry conditions in Oregon. High carbon isotopic
values in marine foraminifera reflect diminished burial
of chemically reduced carbon and higher atmospheric
CO2 levels due to some combination of oceanic
respiration, volcanic emission, or impact destruction
of biomass. A middle Miocene high in atmospheric
CO2 has been confirmed by stomatal index estimates
of atmospheric CO2 (Kurschner et al., 1996; Retallack,
2001c, 2002). Oceanic proxies of atmospheric CO2 do
not show such variation (Pagani et al., 1999; Pearson
and Palmer, 2000) but may be compromised by
volatility of isotopic composition and runoff (Retal-
lack, 2002). Despite these problems, a useful working
hypothesis is control of paleoclimate by a varying
greenhouse effect from changing atmospheric con-
centrations of CO2.
Plausible mechanisms for transient climatic warm-
ing include impact, volcanism (Coutillot, 2002) and
methane clathrate release (Kennett et al., 2002). There
are numerous large (N10 km diameter) Neogene
craters: Haughton, Canada (24 km, 23F1 Ma), Ries,
Germany (24 km, 15F0.1 Ma), Karla, Russia (10 km,
5F1 Ma), ElTgygytygn, Russia (18 km, 3.5F0.5 Ma),
Bosumtwi, Ghana (10.5 km, 1.03F0.02 Ma), and
Zhamanshin, Kazachstan (14 km, 0.9F0.1 Ma;
Dressler and Reimold, 2001; website http://
www.unb.ca/passc/impactdatabase accessed Jan. 10,
2003). Time series of volcanic eruptions are also
strongly episodic (McBirney et al., 1974), with some
exceptionally large eruptions, such as the middle
Miocene Columbia River flood basalts of Oregon
(Wignall, 2001). Columbia River Basalts and Ries
Crater impact may have been involved in the middle
Miocene peak of global warmth, but why was there
not a comparable warm spike after the 23-Ma
Haughton Crater or a better correlation between
North American paleoclimate and Cordilleran volcan-
ism (Fig. 12D). Because of methane’s distinctively
low carbon isotopic values, such methane outbursts
are suspected when carbon isotopic compositions fall
by more than 4x (Jahren et al., 2001). There are no
negative excursions of this magnitude in Neogene
carbon isotopic records (Fig. 12F), thus methane is
unlikely to be the whole explanation either.
A counterpoint to transient carbon-oxidizing events
was progressive long-term Neogene global cooling,
perhaps related to changing oceanic current config-
uration (Broecker, 1997), mountain uplift (Raymo and
Ruddiman, 1992), or biotic intensification of weath-
ering with sod–grassland coevolution (Retallack,
2001a). Both mountain uplift and increased oceanic
thermohaline circulation create oligotrophic alpine and
polar marine communities, with less carbon sequestra-

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123118
tion potential, but grasslands provided more organic
and nutrient-rich runoff to oceanic phytoplankton
(Retallack, 2001b). Grasslands in addition had greater
amounts of soil carbon, higher albedo, and lower rates
of transpiration than the woodlands and shrublands
they replaced in subhumid to semiarid regions of the
world (Nepstad et al., 1994; Jackson et al., 2002), and
this also could have contributed to global drying and
cooling (Retallack, 2001b). This mechanism of global
change is biological, because it is driven by coevolu-
tion of grasses and grazers, with crumb-textured,
organic, fertile soils nourishing hypsodont ungulates
capable of withstanding abrasion from dusty, siliceous
grasses (Retallack, 2001a). The progressive evolution
and spread of crumb-textured soils, siliceous grasses,
and hypsodont ungulates proceeded against a backdrop
of volatile climatic change, and both short-sod and tall-
sod grassland paleosols appearing at times of relatively
high temperature and humidity (Fig. 12A). Grasslands
did not merely fill in dry times and places (Kohn et al.,
2002) but were a biological force for climatic cooling
and drying in their own right (Retallack, 2001b).
Paleoclimatic warm spikes due to volcanism, bolide
impact, and methane clathrate release may also have
been exacerbated by newly coevolved Cenozoic
grasslands, because ruminants liberate CO2 and CH4.
The middle Miocene thermal optimum was a time of
unusually high diversity of North American plants and
mammals, in part due to evolution within high-
productivity grassland–woodland mosaics in a warm-
wet CO2 greenhouse (Janis, 2002), and in part due to
immigration from Asia through high-latitude land
bridges (Wolfe and Tanai, 1980; Kohno, 1997;
Lambert and Shoshani, 1998; Webb, 1998). There
were more grazing mammal species then ever before,
and also more browsers than before or after (Janis et
al., 2000). The middle Miocene greenhouse peak was
preceded and followed by several other episodes of
greenhouse paleoclimate of lesser magnitude, includ-
ing early Clarendonian communities described here.
Warm-wet conditions of greenhouse transients, even if
initiated by tectonic or impact forcing, would be
enhanced by increased mammalian diversity and
cropping of vegetation. Cold-dry conditions follow
carbon and water sequestration by grassland soils, their
high albedo and burial of carbon-rich grassland soil
crumbs. Paleoclimatic cycles on million-year time
scales could reflect alternating evolutionary advances
by plants and animals. Such reciprocating coevolution
is apparent from current estimates of the times for
evolution of bunch grassland paleosols (33 Ma), short-
sod grassland paleosols (19 Ma), and tall-sod grass-
land paleosols (7.3 Ma; Retallack, 1997b, 2001b,
2004; Retallack et al., 2002a), which alternate with
evolution of horse tridactly (36 Ma), cursoriality (25
Ma), hypsodonty and springing gait (17 Ma), mono-
dactyly, wide muzzles, and knee-locking (12 Ma), and
large size and passive-stay shoulder (5 Ma; MacFad-
den, 1992; Janis and Wilheim, 1993; Hermanson and
MacFadden, 1992, 1996). This schedule is compatible
with a coevolutionary process long envisioned for
grassland ecosystems (Jacobs et al., 1999). Global
change involves more than just impact or volcanic
forcing, but a complex interplay of factors for which
detailed proxies are now becoming available (Fig. 12).
These various records will have to become even more
detailed before answers become clear.
Acknowledgements
For introduction to the fossils of this area, I
thank Ted Fremd and his collecting crew, Scott
Foss, Lia Vella, Delda Findieson, Alois Pajak,
Daniella Gould, Matthew, and La Rue Rogers. Kent
Nelson allowed access to his land near Hereford.
Mary Oman of the Baker City Bureau of Land
Management helped with geological reconnaissance,
funding, and research clearance. Megan Smith, Ben
Cook, and Russell Hunt helped with fieldwork and
fossil collection. Alicia Duncan prepared numerous
difficult petrographic thin sections, and Nathan
Sheldon provided bulk density measurements.
References
Alroy, J., Koch, P.L., Zachos, J.C., 2000. Global climate change and
North American mammalian evolution. Paleobiol. Suppl. 26,
259–288.
Ashwill, M., 1983. Seven fossil floras in the rain shadow of the
Cascades Mountains. Or. Geol. 45, 107–117.
Axelrod, D.I., 1985. Miocene floras from the Middlegate Basin,
west-central Nevada. Univ. Calif. Publ. Geol. Sci. 129 (279 pp.).
Axelrod, D.I., 1995. The Miocene Purple Mountain Flora of
western Nevada. Univ. Calif. Publ. Geol. Sci. 139 (62 pp.).
Baldwin, E.M., 1964. Geology of Oregon. Cooperative Book Store,
Eugene. 165 pp.

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 119
Barbour, E.H., 1932. The mandible of Platybelodon barnumbrowni.
Bull. Nebr. State Mus. 1, 155–158.
Barbour, M.G., Major, J. (Eds.), Terrestrial Vegetation of California.
Spec. Publ. Calif. Native Plant Soc. Sacram, vol. 9, 1030 pp.
Baskin, J.A., 1998. Procyonidae. In: Janis, C.M., Scott, K.M., Jacobs,
L.L. (Eds.), Evolution of Tertiary Mammals of North America,
Terrestrial Carnivores, Ungulates and Ungulatelike mammals,
vol. 1. Cambridge University Press, Cambridge, pp. 144–173.
Bernor, R.L., Kovar-Eder, J., Lipscomb, P., Rfgl, F., Sen, S.,
Tobien, H., 1988. Systematics, stratigraphic, and paleoenvir-
onmental contexts of first-appearing Hipparion in the Vienna
Basin, Austria. J. Vertebr. Paleontol. 8, 427–452.
Berry, E.W., 1931. A Miocene flora from Grand Coulee, Wash-
ington. U. S. Geol. Surv. Prof. Pap. 170-C, 31–42.
Bestland, E.A., Krull, E.S., 1997. Mid-Miocene climatic optimum
recorded in paleosols from the Mascall Formation (Oregon).
Abstr. Geol. Soc. Amer. 29 (6), 5.
Bestland, E.A., Hammond, P.E., Blackwell, D.L.S., Kays, M.A.,
Retallack, G.J., Stimac, J., 1999. Geologic framework of the
Clarno Unit, John Day Fossil Beds National Monument, central
Oregon. Or. Geol. 61, 3–19.
Blinnikov, M., Busacca, A., Whitlock, C., 2002. Reconstruction of
the late Pleistocene grassland of the Columbia Basin, Wash-
ington, USA, based on phytolith records in loess. Palaeogeogr.
Palaeoclimatol. Palaeoecol. 177, 77–101.
Boggs, S., 1972. Petrography and geochemistry of rhombic, calcite
pseudomorphs from mid-Tertiary mudstones. Sedimentology
19, 204–219.
Bowen, R.G., Gray, W.L., Gregory, D.C., 1963. General geology of
the northern Juntura Basin. Trans. Am. Philos. Soc. 53, 22–34.
Bown, T.M., 1980. The fossil insectivora of Lemoyne Quarry (Ash
Hollow Formation, Hemphillian), Keith County, Nebraska.
Trans. Nebr. Acad. Sci. 8, 99–121.
Bradbury, J.P., Dieterich, K.V., Williams, J.L., 1985. Diatom flora of
the Miocene lake beds near Clarkia in northern Idaho. In:
Smiley, C.J. (Ed.), Late Cenozoic History of the Pacific
Northwest. American Association for the Advancement of
Science, pp. 33–59.
Brewer, R., 1976. Fabric and Mineral Analysis of Soils. Krieger,
New York. 482 pp.
Brodkorb, P., 1961. Birds from the Pliocene of Juntura, Oregon. Fla.
Sci. 24, 169–184.
Broecker, W.S., 1997. Thermohaline circulation, the Achilles heel
of our climate system: will man-made CO2 upset the current
balance? Science 2878, 1582–1588.
Brown, J.H., Lomolino, M.V., 1998. Biogeography. Sinauer,
Sunderland, Massachussetts. 691 pp.
Busacca, A.J., 1989. Long Quaternary record in eastern Wash-
ington, USA, interpreted from multiple paleosols in loess.
Geoderma 45, 105–122.
Busacca, A.J., 1991. Loess deposits and soils of the Palouse and
vicinity. In: Baker, V.R., Bjornstad, B.N., Busacca, A.J., Fecht,
K.R., Kiver, E.P., Moody, U.L., Rigby, J.G., Stradling, D.F.,
Tallman, A.M. (Eds.), The Columbia Plateau;
In: Morrison, R.B. (Ed.), Quaternary Geology of the United
States. Geological Society of America, Boulder, pp. 216–228.
v. K2.
Cerling, T.E., Harris, J.M., MacFadden, B.J., Leaker, M.G., Quade,
J., Eisenmann, V., Ehleringer, J.R., 1997. Global vegetation
change through the Miocene/Pliocene boundary. Nature 389,
153–158.
Chadwick, O.A., Hendricks, D.M., Nettleton, W.D., 1987. Silica in
duric soils: I. A depositional model. Soil Sci. Soc. Am. J. 51,
975–982.
Chadwick, O.A., Nettleton, W.D., Staidle, G.J., 1995. Soil poly-
genesis as a function of Quaternary climate change, northern
Great Basin, USA. Geoderma 68, 10–26.
Chaney, R.W., 1948. The Ancient Forests of Oregon. University of
Oregon, Eugene. 56 pp.
Chaney, R.W., Axelrod, D.I., 1959. Miocene floras of the Columbia
Plateau, vol. 617. Carnegie Inst., Washington Publ. 237 pp.
Clark, J.M., Dawson, M.R., Wood, A.E., 1964. Fossil mammals
from the lower Pliocene of Fish Lake Valley. Nev. Bull. Mus.
Comp. Zool. 131, 29–63.
Compton, L.V., 1935. Two avian fossils from the lower Pliocene of
South Dakota. Am. J. Sci. 30, 343–348.
Condit, C., 1944. The Remington Hill flora. Carnegie Institute.
Washington Publ. 553, 21-55.
Coutillot, V., 2002. Evolutionary Catastrophism. Cambridge Uni-
versity Press, Cambridge.
Dalrymple, G.B., 1979. Critical tables for conversion of K–Ar ages
from old to new constants. Geology 7, 558–560.
Dawson, M.R., 1958. Later Tertiary Leporidae of North
America. Univ. Kans. Paleontol. Contrib., Pap. 22 (Vertebrata
6, 75 pp.).
Delancey, S., Genetti, C., Rude, N., 1988. Some Sahaptian–
Klamath–Tsimshianic lexical sets. In: Shipley, W. (Ed.), In
Honor of Mary Haas. Mouton de Gruyter, Berlin, pp. 195–224.
de Villiers, S., 2003. Dissolution effects on foraminiferal Mg/Ca
records of sea surface temperature in the western equatorial
Pacific. Paleoceanography 18, 1070.
Dorf, E., 1936. A late Tertiary flora from southwestern Idaho.
Carnegie Inst. Wash. Contr. Paleontol. 426, 185–216.
Dompierre, H., Churcher, C.S., 1996. Premaxillary shape as an
indicator of the diet of seven late Cenozoic New World camels.
J. Vertebr. Paleontol. 16, 141–148.
Downs, T., 1952. A new mastodont from the Miocene of Oregon.
Univ. Calif. Publ. Geol. Sci. 29, 1–20.
Downs, T., 1956. The Mascall fauna from the Miocene of Oregon.
Univ. Calif. Publ. Geol. Sci. 31, 199–354.
Draus, E., Prothero, D.R., 2002. Magnetic stratigraphy of the
middle Miocene (early Barstovian) Mascall Formation, central
Oregon. Abstr. Geol. Soc. Amer. 36 (6), 135–136.
Dressler, B.D., Reimold, W.J., 2001. Terrestrial impact melt rocks
and glasses. Earth Sci. Rev. 56, 205–284.
Evans, J.G., 1992. Geologic map of the Dooley Mountain 7OVQuadrangle, Baker County, Oregon. U.S. Geol. Surv. Geol.
Quad. (Map GQ-1694, 9 pp.).
Evernden, J.F., James, G.T., 1964. Potassium–argon dates and the
Tertiary floras of North America. Am. J. Sci. 262, 945–974.
F.A.O. (Food and Agriculture Organization), 1974. Soil Map of the
World. Legend, vol. I. U.N.E.S.C.O, Paris. 59 pp.
F.A.O. (Food and Agriculture Organization), 1975. Soil Map of the
World. North America, vol. II. U.N.E.S.C.O, Paris. 210 pp.

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123120
Farquhar, G.D., Lloyd, D.J., Taylor, J.A., Flanagan, L.B., Syvert-
son, J.P., Hubick, K.T., Wong, C.S., Ehleringer, J.R., 1993.
Vegetation effects on the isotopic composition of oxygen in
atmospheric CO2. Nature 363, 439–443.
Fiebelkorn, R.B., Walker, G.W., MacLeod, N.S., McKee, E.H.,
Smith, J.G., 1982. Index to K–Ar determinations for the State of
Oregon. Isochron-West, vol. 37. 60 pp.
Fleming, C.A., 1975. Biogeography and ecology in New Zealand.
In: Kuschel, G. (Ed.), The Geological History of New Zealand
and its Biota. W. Junk, The Hague, pp. 1–86.
Fox, D.L., 2001. Growth increments in Gomphotherium tusks
and implications for late Miocene climate change in
North America. Palaeogeogr. Palaeoclimatol. Palaeoecol. 156,
327–348.
Fox, D.L., Koch, P.L., 2003. Tertiary history of C4 biomass in the
Great Plains, USA. Geology 31, 809–812.
Fremd, T., Bestland, E.A., Retallack, G.J., 1994. John Day Basin
Fieldtrip Guide and Road Log for the Society of Vertebrate
Paleontology Annual Meeting. Northwest Interpretive Associ-
ation, Seattle. 90 pp.
Gilluly, J., 1937. Geology and mineral resources of the Baker
Quadrangle. U.S. Geol. Surv. Bull. 879 (119 pp.).
Golia, R.T., Stewart, J.H., 1984. Depositional environments and
paleogeography of the upper Miocene Wassuk Group, west
central Nevada. Sediment. Geol. 38, 159–180.
Graham, A., 1999. Late Cretaceous and Cenozoic History of
North American Vegetation. Oxford University Press, New
York. 350 pp.
Greene, R.C., 1973. Petrology of the welded tuff of Devine
Canyon, southeastern Oregon. U. S. Geol. Surv. Prof. Pap. 797
(26 pp.).
Gregory, J.T., 1942. Pliocene vertebrates from Big Spring Canyon,
South Dakota. Univ. Calif. Publ. Geol. Sci. 26, 307–442.
Gustafson, E.P., 1978. The vertebrate fauna of the Pliocene Ringold
Formation, south-central Washington. Bull. Mus. Nat. Hist.
Univ. Oregon 23 (62 pp.).
Gustafson, E.P., 1985. Soricids (Mammalian: Insectivora) from
the Blufftop local fauna, Blancan Ringold Formation of
central Washington and correlation of the Ringold Formation.
J. Vertebr. Paleontol. 5, 88–92.
2Hall, E.R., 1930. Rodents and lagomorphs from the Late Tertiary
of Fish Lake Valley, Nevada. Univ. Calif. Publ. Geol. Sci. 19,
295–312.
Harden, J., 1982. A quantitative index of soil development from
field descriptions: examples from a chronosequence in central
California. Geoderma 28, 1–28.
Harden, J., 1990. Soil development on stable landforms and
implications for landscape studies. Geomorphology 3, 391–398.
Hannibal, H., 1912. A synopsis of the recent and Tertiary
freshwater Mollusca of the Californian province, based upon
an ontogenetic classification. Proc. Malacol. Soc. Lond. 10,
112–211.
Hayek, L.-A.C., Bernor, R.L., Solounias, N., Steigerwald, P., 1992.
Preliminary studies of hipparionine horse diet as measured by
tooth wear. Ann. Zool. Fenn. 28, 187–200.
Hermanson, J.W., MacFadden, B.J., 1992. Evolutionary and func-
tional morphology of the shoulder region and stay apparatus in
fossil and extant horses (Equidae). J. Vertebr. Paleontol. 12,
377–386.
Hermanson, J.W., MacFadden, B.J., 1996. Evolutionary and func-
tional morphology of the knee in fossil and extant horses
(Equidae). J. Vertebr. Paleontol. 16, 349–357.
Herrington, H.B., Taylor, D.W., 1958. Pliocene and Pleistocene
Sphaeriidae (Pelecypoda) from the central United States. Occas.
Pap. Mich. Univ. Dept. Zool. 596 (23 pp.).
Hibbard, C.W., Phillis, L.F., 1945. The occurrence of Eucastor and
Epigaulus in the lower Pliocene of Trego County, Kansas. Bull.
Univ. Kans. 30, 549–555.
Hoffman, J., Prothero, D.R., 2002. Magnetic stratigraphy of the
upper Miocene (early Hemphillian) Rattlesnake Formation,
central Oregon. Abstr. Geol. Soc. Amer. 36 (6), 135.
Honey, J.G., Harrison, J.A., Prothero, D.R., Stevens, M., 1998.
Camelidae. In: Janis, C.M., Scott, K.M., Jacobs, L.L. (Eds.),
Evolution of Tertiary Mammals of North America, Terrestrial
Carnivores, Ungulates and Ungulatelike Mammals, vol. 1.
Cambridge University Press, Cambridge, pp. 439–462.
Humphreys, E.D., Dueker, K.G., Schutt, D.L., Smith, R.B., 2000.
Beneath Yellowstone: evaluating plume and non-plume models
using teleseismic images of the upper mantle. GSA Today 10
(12), 1–7.
Hutchison, J.H., 1968. Fossil Talpidae (Insectivora, Mammalia)
from the later Tertiary of Oregon. Bull. Mus. Nat. Hist. Univ.
Oregon 11 (117 pp.).
Itoigawa, J., Yamanoi, T., 1990. Climatic optimum in the mid-
Neogene of the Japanese Islands. In: Tsuchi, P. (Ed.), Pacific
Neogene Events. Tokyo University Press, Tokyo, pp. 3–15.
Jackson, R.B., Banner, J.L., Jobbagy, E.G., Pockman, W.T., Wall,
T.H., 2002. Ecosystem carbon loss with woody plant invasion of
grasslands. Nature 418, 623–626.
Jacobs, B., Kingston, J.D., Jacobs, L., 1999. The origin of grass-
dominated ecosystems. Mo. Bot. Gard. Ann. 399, 673–676.
Jahren, A.H., Arens, N.C., Sarmiento, G., Guerro, J., Armstrong, R.,
2001. Terrestrial record of global methane hydrate dissociation
in the Early Cretaceous. Geology 29, 159–162.
Janis, C.M., 2002. The origin and evolution of the North American
grassland biome: the story from the hoofed mammals. Palae-
ogeogr. Palaeoclimatol. Palaeoecol. 177, 183–198.
Janis, C.M., Wilheim, P.B., 1993. Were there mammalian pursuit
predators in the Tertiary? Dances with wolf avatars. J. Mamm.
Evol. 1, 103–125.
Janis, C.M., Gordon, J.J., Ilius, A.W., 1994. Modelling equid/
ruminant competition in the fossil record. Hist. Biol. 8, 15–29.
Janis, C.M., Damuth, J., Theodor, J.M., 2000. Miocene ungulates
and terrestrial primary productivity: where have all the browsers
gone? Proc. U.S. Natl. Acad. Sci. 97, 7899–7904.
Johnson, J.A., Binger, G.B., Hooper, P.R., Hawkesworth, C.J.,
1996. Implications of structure and stratigraphy in the Juntura
area, east-central Oregon. Abstr. Geol. Soc. Amer. 28 (5), 78.
Kellogg, E.A., 1999. Phylogenetic aspects of the evolution of C4
photosynthesis. In: Sage, R.F., Monson, R.K. (Eds.), Plant
Biology. Academic Press, New York, pp. 411–444.
Kennett, J.P., Cannariato, K.G., Hendy, I.L., Behl, R.J., 2002.
Methane Hydrates in Quaternary Climate Change. American
Geophysical Union, Washington. 224 pp.

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 121
Kingston, J.D., Marino, B.D., Hill, A., 1994. Isotopic evidence for
Neogene hominid environments in the Kenya Rift Valley.
Science 264, 955–959.
Knowlton, F.H., 1898. The fossil plants of the Payette Formation.
U.S. Geol. Surv. Ann. Rep. 18 (3), 721–744.
Koch, P.L., Clyde, W.C., Hepple, R.P., Fogel, M.L., Wing, S.L.,
Zachos, J.C., 2003. Carbon and oxygen isotope records
from paleosols spanning the Paleocene–Eocene boundary,
Bighorn Basin. In: Wing, S.L., Gingerich, P.D., Schmitz, N.,
Thomas, E. (Eds.), Causes and consequences of globally warm
climates in the early Paleogene, Spec. Pap. Geol. Soc. Amer.
369, pp. 49–64.
Kohn, M.J., Miselis, J.L., Fremd, T.J., 2002. Oxygen isotope
evidence for progressive uplift of the Cascade Range, Oregon.
Earth Planet. Sci. Lett. 6426, 1–15.
Kohno, N., 1997. The first record of an amphicyonid (Mammalia,
Carnivora) from Japan, and its implications for amphicyonid
paleobiogeography. Paleontol. Res. 1, 311–315.
Korth, W.W., 1994. The Tertiary Record of Rodents in North
America. Plenum Press, New York. 319 pp.
Kqrschner, W.M., Van Der Burgh, J., Visscher, H., Dilcher, D.L.,
1996. Oak leaves as biosensors of late Neogene and early
Pleistocene paleoatmospheric CO2 concentrations. Mar. Micro-
paleontol. 27, 299–312.
Lambert, W.D., 1992. The feeding habits of the shovel-tusked
gomphotheres: evidence from tusk wear patterns. Paleobiology
18, 132–147.
Lambert, W.D., Shoshani, J., 1998. Proboscidea. In: Janis, C.M.,
Scott, K.M., Jacobs, L.L. (Eds.), Evolution of Tertiary Mammals
of North America, Terrestrial Carnivores, Ungulates and
Ungulatelike Mammals, vol. 1. Cambridge University Press,
Cambridge, pp. 606–621.
Lander, B., 1998. Oreodontoidea. In: Janis, C.M., Scott, K.M.,
Jacobs, L.L. (Eds.), Evolution of Tertiary Mammals of North
America, Terrestrial Carnivores, Ungulates and Ungulatelike
Mammals vol. 1. Cambridge University Press, Cambridge,
pp. 402–425.
Lear, C.H., Elderfield, H., Wilson, P.A., 2000. Cenozoic deep-sea
temperatures and global ice volumes from Mg/Ca in benthic
foraminiferal calcite. Science 287, 269–272.
Leidy, J., 1858. Notice of remains of extinct Vertebrata, from the
valley of the Niobrara River, collected during the expedition of
1857, in Nebraska, under the command of Lieut. G.K. Warren.
Proc. Acad. Nat. Sci. Philadelphia 10, 10–29.
Leopold, E.B., Denton, M.F., 1987. Comparative age of grassland
and steppe east and west of the northern Rocky Mountains. Mo.
Bot. Gard. Ann. 74, 841–867.
Leopold, E.B., Wright, V.C., 1985. Pollen profiles of the
Plio–Pleistocene transition in the Snake River Plain, Idaho.
In: Smiley, C.J. (Ed.), Late Cenozoic History of the Paci-
fic Northwest. American Association for the Advancement
of Science, San Francisco, pp. 323–334.
Lesquereux, L., 1878. Report on the auriferous gravel deposits of
the Sierra Nevada. Harv. Mus Comp. Zool. Mem. 6 (62 pp.).
Li, Q.-Y., McGowran, B., 2000. Miocene foraminifera from Lakes
Entrance oil shaft, Gippsland, southeastern Australia. Mem.
Assoc. Australas. Palaeontol. 22 (142 pp.).
Lindsay, E.H., 1972. Small mammal fossils from the Barstow
Formation, California. Univ. Calif. Publ. Geol. Sci. 93, 1–104.
Lundberg, J.G., 1975. The fossil catfish of North America. Mus.
Paleontol. Univ. Mich. Pap. Paleontol. 2 (11), 1–51.
MacFadden, B.F., 1984. Systematics and phylogeny of Hipparion,
Neohipparion, Nannippus, and Cormohipparion (Mammalia,
Equidae) from the Miocene and Pliocene of the New World.
Bull. Am. Mus. Nat. Hist. 179, 1–175.
MacFadden, B.F., 1992. Fossil Horses. Cambridge University Press,
Cambridgee. 369 pp.
MacFadden, B.F., Skinner, M.F., 1977. Earliest known Hipparion
from Holarctica. Nature 265, 532–533.
MacGinitie, H.D., 1934. The Trout Creek flora of southeastern
Oregon. Carnegie Inst. Wash. Publ. 416, 21–68.
MacLeod, K.G., Smith, R.M.H., Koch, P.L., Ward, P.D., 2000.
Timing of mammal-like reptile extinction across the Permian–
Triassic boundary in South Africa. Geology 28, 227–230.
Madden, C.T., Storer, J.E., 1985. The Proboscidea from the middle
Miocene Wood Mountain Formation, Saskatchewan. Can. J.
Earth Sci. 22, 1345–1350.
Martin, L.D., 1994. Cenozoic climatic history from a biological
perspective. TER-QUA Symp. Ser. 2, 39–56.
Martin, L.D., 1998. Felidae. In: Janis, C.M., Scott, K.M., Jacobs,
L.L. (Eds.), Evolution of Tertiary Mammals of North America,
Terrestrial Carnivores, Ungulates and Ungulatelike mammals,
vol. 1. Cambridge University Press, Cambridge, pp. 236–242.
Matthew, W.D., Cook, H.J., 1909. A Pliocene fauna from western
Nebraska. Bull. Am. Mus. Nat. Hist. 26, 361–414.
McBirney, A.R., Sutter, J.F., Naslund, H.R., Sutton, K.G., White,
C.M., 1974. Episodic volcanism in the central Oregon Cascade
Range. Geology 2, 585–589.
Meehan, T.J., 1996. Climate control of evolutionary trends in
Cenozoic mammals. Suppl. J. Vertebr. Paleontol. 16, 52.
Meehan, T.J., 1999. Ecomorph analysis of North American
Cenozoic mammals. Suppl. J. Vertebr. Paleontol. 19, 63.
Meehan, T.J., Martin, L.D., 1994. Iterative evolution in the North
American Cenozoic. Suppl. J. Vertebr. Paleontol. 14, 38.
Merriam, J.C., 1916. Mammalian remains from a Tertiary
formation at Ironside, Oregon. Univ. Calif. Publ. Geol. Sci.
10, 129–135.
Miller, R.R., Smith, G.R., 1967. New fossil fishes from Plio–
Pleistocene Lake Idaho. Occas. Pap. Mus. Zool. Univ. Mich.
654, 1–24.
Moore, E.J., 1963. Miocene marine mollusks from the Astoria
Formation in Oregon. U. S. Geol. Surv. Prof. Pap. 419, 109.
Morgan, J.K., Morgan, N.H., 1995. A new species of Capromeryx
(Mammalia: Artiodactyla) from the Taunton local fauna of
Washington, and the correlation with other Blancan faunas of
Washington and Idaho. J. Vertebr. Paleontol. 15, 160–170.
Morgan, M.E., Kingston, J.D., Marino, B.D., 1994. Carbon isotopic
evidence for the emergence of C4 plants from Pakistan and
Kenya. Nature 367, 162–165.
Nambudiri, W.M.V., Tidwell, W.D., Smith, B.N., Hebbert, N.P.,
1978. A C4 plant from the Pliocene. Nature 276, 816–817.
Nepstad, D.C., de Carvelho, C.R., Davodson, E.A., Jipp, P.H.,
Lefebvre, P.R., Negreiros, G.H., de Silva, E.D., Stone, T.A.,
Trumbore, S.A., Vieira, S., 1994. The role of deep roots in the

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123122
hydrological and carbon cycle of Amazonian pastures and
forests. Nature 372, 666–669.
Neumayr, M., Paul, C.M., 1875. Die Congerien- und Paludien-
schichten Slavoniens und deren Fauna. Abh. K.-K. Geol.
Reichs. Wien 7/3, 1–106.
Oleinik, A., Marincovich, L., 2001. Biogeographic and stable
isotopic evidence for middle Miocene warming in the
high latitude North Pacific. Abstr. Geol. Soc. Amer. 33 (6),
A386.
Oliver, E., 1934. A Miocene flora from the Blue Mountains. Publ.
Carnegie Inst. Wash. 455, 1–27.
Orr, E.L., Orr, W.N., 1999. Oregon Fossils. Kendall/Hunt,
Dubuque, Iowa. 381 pp.
Orr, E.L., Orr, W.N., Baldwin, E., 1992. Geology of Oregon.
Kendall/Hunt, Dubuque, Iowa. 254 pp.
Owen-Smith, R.N., 1988. Megaherbivores. Cambridge University
Press, Cambridge. 369 pp.
Pagani, M., Freeman, K.H., Arthure, M.A., 1999. Miocene
atmospheric CO2 concentration and expansion of C4 grasses.
Science 285, 878–976.
Palmer, A.R., 1957. Miocene arthropods from the Mojave Desert,
California. U. S. Geol. Surv. Prof. Pap. 294G, 237–280.
Passy, B.H., Cerling, T.E., Perkins, M.E., Voorhies, M.R., Harris,
J.M., Tucker, S.T., 2002. Environmental change in the Great
Plains: an isotopic record from fossil horses. J. Geol. 110,
123–140.
Pearson, P.N., Palmer, M.R., 2000. Atmospheric carbon dioxide
concentrations over the past 60 million years. Nature 406,
695–699.
Perkins, M.E., Brown, F.H., Nash, W.F., McIntosh, W., Williams,
S.K., 1998. Sequence, age and source of silicic fallout tuffs in
middle to late Miocene basins of northern Basin and Range
Province. Bull. Geol. Soc. Am. 110, 344–360.
Priest, G.R., 1990. Volcanic and tectonic evolution of the
Cascade volcanic arc, central Oregon. J. Geophys. Res. 95,
19383–19599.
Prothero, D.R., 1998. The chronological, climatic, and paleogeo-
graphic background to North American mammalian evolution.
In: Janis, C.M., Scott, K.M., Jacobs, L.L. (Eds.), Evolution of
Tertiary Mammals of North America, Terrestrial Carnivores,
Ungulates and Ungulatelike Mammals, vol. 1. Cambridge
University Press, Cambridge, pp. 1–35.
Raymo, M.E., Ruddiman, W.F., 1992. Tectonic forcing of late
Cenozoic climate. Nature 359, 112–122.
Reef, J.W., 1983 The Unity Reservoir tuff–breccia and associated
volcanic rocks, Baker County, Oregon. Unpub. MSc thesis.
Washington State University, Pullman. 128 p.
Retallack, G.J., 1994. The environmental factor approach
to the interpretation of paleosols. In: Amundson, R.,
Harden, J., Singer, M. (Eds.), Factors in soil formation: a
fiftieth anniversary retrospective, Soil Sci. Soc. Amer. Spec.
Publ., vol. 33, pp. 31–64.
Retallack, G.J., 1997. A Colour Guide to Paleosols. Wiley,
Chichester. 175 pp.
Retallack, G.J., 1997. Neogene expansion of the North American
prairie. Palaios 12, 380–390.
Retallack, G.J., 2000. Depth to pedogenic carbonate horizon as a
paleoprecipitation indicator? Comment and reply. Geology 28,
572–573.
Retallack, G.J., 2001. Cenozoic expansion of grasslands and
climatic cooling. J. Geol. 109, 407–426.
Retallack, G.J., 2001. Soils of the Past. Blackwell, Oxford. 404 pp.
Retallack, G.J., 2001. A 300-million-year record of atmos-
pheric carbon dioxide from fossil plant cuticles. Nature 411,
287–290.
Retallack, G.J., 2002. Carbon dioxide and climate over the past 300
Myr. Philos. Trans. R. Soc. Lond. B360, 659–673.
Retallack, G.J., 2004. Late Oligocene bunch grassland and early
Miocene sod grassland paleosols from central Oregon, U.S.A.
Palaeogeogr. Palaeoclimatol. Palaeoecol. 207, 203–237.
Retallack, G.J., Bestland, E.A., Fremd, T.J., 2000. Eocene and
Oligocene paleosols of central Oregon. Spec. Pap. Geol. Soc.
Amer. 344, 192.
Retallack, G.J., Tanaka, S., Tate, T., 2002a. Late Miocene advent of
tall grassland paleosols in Oregon. Palaeogeogr. Palaeoclimatol.
Palaeoecol. 183, 329–354.
Retallack, G.J., Wynn, J.G., Benefit, B.R., McCrossin, M.L., 2002b.
Paleosols and paleoenvironments of the middle Miocene
Maboko Formation, Kenya. J. Hum. Evol. 42, 659–708.
Retallack, G.J., Wynn, J.G., Fremd, T.J., 2004. Glacial–interglacial-
scale paleoclimatic change without large ice sheets in the
Oligocene of central Oregon. Geology 32, 297–300.
Rigsby, B.J., 1965 Linguistic relations in the southern Plateau.
Unpub. PhD thesis. Univ. Oregon, Eugene, 269 pp.
Robyn, T.L., 1977. Geology and petrology of the Strawberry
Volcanics, NE Oregon. Unpub PhD thesis. Univ. Oregon,
Eugene. 224 pp.
Ruffner, J.A., 1985. Climates of the States, vol. 2. Gale, Detroit.
vols 1572 pp.
Ruxton, B.B., 1968. Rates of weathering of Quaternary volcanic ash
in northeast Papua. Trans. 9th Int. Soil Sci. Congr. Adelaide,
vol. 4, pp. 367–376.
Sage, R.F., Wedin, O.A., Li, M., 1999. Biogeography of C4
photosynthesis: pattern and controlling factors. In: Sage, R.F.,
Monson, R.K. (Eds.), Plant Biology. Academic Press, San Diego,
pp. 313–373.
Sawyer, J.O., Keeler-Wolf, T., 1995. A Manual of California
Vegetation. California Native Plant Society, Sacramento. 471 pp.
Schultz, C.B., Stout, T.M., 1980. Ancient soils and climate changes
in the central Great Plains. Trans. Nebr. Acad. Sci. 8, 187–205.
Schwarz, T., 1997. Lateritic paleosols in central Germany and
implications for Miocene palaeoclimates. Palaeogeogr. Palae-
oclimatol. Palaeoecol. 129, 37–50.
Sheldon, N.D., 2003. Pedogenesis and geochemical alteration of the
Picture Gorge Subgroup, Columbia River Basalt, Oregon. Geol.
Soc. Amer. Bull. 115, 1377–1387.
Sheldon, N.D., Retallack, G.J., 2001. Equation for compaction of
paleosols due to burial. Geology 29, 247–250.
Sheldon, N.D., Retallack, G.J., Tanaka, S., 2002. Geochemical
climofunctions from North American soils and application to
paleosols across the Eocene–Oligocene boundary in Oregon.
J. Geol. 110, 687–696.

G.J. Retallack / Palaeogeography, Palaeoclimatology, Palaeoecology 214 (2004) 97–123 123
Shotwell, J.A., 1958. Evolution and biogeography of the Aplodon-
tid and Mylagaulid rodents. Evolution 12, 451–484.
Shotwell, J.A., 1963. Pliocene mammalian communities of the
Juntura Basin. Trans. Am. Philos. Soc. 53, 7–21.
Shotwell, J.A., 1967. Peromyscus of the late Tertiary in Oregon.
Bull. Mus. Nat. Hist. Univ. Oregon 5, 35.
Shotwell, J.A., 1970. Pliocene mammals of southeast Oregon and
adjacent Idaho. Bull. Mus. Nat. Hist. Univ. Oregon 17, 103.
Shotwell, J.A., Russell, D.E., 1963. Mammalian fauna of the upper
Juntura Formation, the Black Butte local fauna. Trans. Am.
Philos. Soc. 53, 42–69.
Smith, G.R., Morgan, N., Gustafson, E., 2000. Fishes of the Mio–
Pliocene Ringold Formation, Washington: Pliocene capture of
the Snake River by the Columbia River. Publ. Paleontol. Univ.
Mich. 32, 47.
Soil Survey Staff, 1999. Keys to Soil Taxonomy. Pocahontas Press,
Blacksburg. 600 pp.
Solounias, N., 1981. The Turolian Fauna from the Island of Samos,
Greece: with Special Emphasis on the Hyaenids and the Bovids.
Karger, Basel. 232 pp.
Stace, H.C.T., Hubble, G.D., Brewer, R., Northcote, K.H., Sleeman,
J.R., Mulcahy, M.J., Hallsworth, E.G., 1968. A Handbook of
Australian Soils. Rellim, Adelaide. 435 pp.
Strfmberg, C.A.E., 2002. The origin and spread of grass-dominated
ecosystems in the late Tertiary of North America: preliminary
results concerning the evolution of hypsodonty. Palaeogeogr.
Palaeoclimatol. Palaeoecol. 177, 59–75.
Taggart, R.E., Cross, A.T., 1990. Plant successions and interruptions
in Miocene volcanic deposits, Pacific Northwest. In: Lockley,
M.G., Rice, A. (Eds.), Volcanism and fossil biotas, Spec. Pap.
Geol. Soc. Amer., vol. 244, pp. 57–68.
Taggart, R.E., Cross, A.T., Satchell, L., 1982. Effects of periodic
volcanism on Miocene vegetation distribution in eastern Oregon
and western Idaho. Proc. 3rd Nth Amer. Paleont. Conv., vol. 2,
pp. 535–540.
Tate, T.A., 1998 Micromorphology of loessial soils and paleosols
on aggrading landscapes of the Columbia Plateau. Unpub.
MSc Thesis Dept. Geology Western Washington Univ. Pull-
man. 192 pp.
Taylor, D.W., 1963. Mollusks of the Black Butte local fauna. Trans.
Am. Philos. Soc. 53, 35–41.
Taylor, D.W., 1985. Evolution of freshwater drainages and molluscs
in western North America. In: Smiley, C.J. (Ed.), Late Cenozoic
History of the Pacific Northwest. American Association for the
Advancement of Science, San Francisco, pp. 265–320.
Tedford, R.H., 1961. Clarendonian insectivores from the Ricardo
Formation, Kern County, California. Bull. S. C. Acad. Sci. 60,
57–76.
Tedford, R.H., Barghoorn, S.F., 1993. Neogene stratigraphy and
mammalian biochronology of the Espanola Basin, New Mexico.
Bull. N. M. Mus. Nat. Hist. Sci. 2, 159–168.
Thayer, T.P., 1957. Some relations of later Tertiary volcanology and
structure in eastern Oregon. Proc. 20th Int. Geol. Congr.,
Mexico, vol. 1956, pp. 231–245. sect. 1 Volcanologia del
Cenozoico 1.
Thayer, T.P., Brown, C.E., 1973. Ironside Mountain, Oregon: a late
Tertiary volcanic and structural enigma. Bull. Geol. Soc. Am.
84, 489–498.
Thomasson, J.R., 1979. Late Cenozoic grasses and angiosperms from
Kansas, Nebraska and Colorado: biostratigraphy and relation-
ships with living taxa. Bull. Geol. Surv. Kans. 218, 68.
Tobien, H., 1972. Status of the genus Serridentinus Osborn 1923
(Proboscidea, Mammalia) and related forms. Mainzer Geowiss.
Mitt. 1, 143–191.
Van Landingham, S.L., 1967. Paleoecology and microfloristics of
Miocene diatomites from the Otis Basin–Juntura region of
Harney and Malheur Counties, Oregon, Beih. Nova Hedwigia
16, 1–77.
Van TassellL, J., Ferns, M., McConnell, V., Smith, G.R., 2001. The
mid-Pliocene Imbler fish fossils, Grande Ronde Valley, Union
County, Oregon, and the connection between Lake Idaho and
the Columbia River. Or. Geol. 63, 77–96.
Veizer, J., Godderis, Y., Francois, L.M., 2000. Evidence for
decoupling of atmospheric CO2 and global climate during the
Phanerozoic eon. Nature 408, 698–701.
Walker, G.W. (Ed.), Geology of the Blue Mountains region of
Oregon, Idaho and Washington, U. S. Geol. Surv. Prof. Pap.,
1457, 135.
Walker, G.W., MacLeod, N.S., 1991. Geologic Map of Oregon:
1:500,000. U.S. Geological Survey, Denver. 2 sheets.
Webb, S.D., 1983. The rise and fall of the late Miocene ungulate
fauna in North America. In: Nitecki, M.N. (Ed.), Coevolution.
University of Chicago Press, Chicago, pp. 276–306.
Webb, S.D., 1998. Hornless ruminants. In: Janis, C.M., Scott,
K.M., Jacobs, L.L. (Eds.), Evolution of Tertiary Mammals of
North America, Terrestrial Carnivores, Ungulates and Ungu-
latelike mammals, vol. 1. Cambridge University Press, Cam-
bridge, pp. 463–476.
Whistler, D.P., Burbank, D.W., 1992. Miocene biostratigraphy and
biochronology of the Dove Spring Formation, Mojave Desert,
California and characterization of the Clarendonian mammal
age in California. Bull. Geol. Soc. Am. 104, 644–658.
Wignall, P.B., 2001. Large igneous provinces and mass extinctions.
Earth Sci. Rev. 272, 1155–1158.
Wolfe, J.A., 1994. Tertiary climate changes at the middle latitudes
of western North America. Palaeogeogr. Palaeoclimatol. Palae-
oecol. 108, 195–205.
Wolfe, J.A., Tanai, T., 1980. The Miocene Seldovia Point flora of the
Kenai Group, Alaska. U. S. Geol. Surv. Prof. Pap. 1105, 52.
Wolfe, J.A., Schorn, H.E., Forest, C.E., Molnar, P., 1997.
Paleobotanical evidence for high altitudes in Nevada during
the Miocene. Science 276, 1672–1675.
Wright, D.B., 1998. Tayassuidae. In: Janis, C.M., Scott, K.M.,
Jacobs, L.L. (Eds.), Evolution of Tertiary Mammals of North
America, Terrestrial Carnivores, Ungulates and Ungulatelike
mammals, vol. 1. Cambridge University Press, Cambridge,
pp. 389–401.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., Billups, K., 2001.
Trends, rhythms, and aberrations in global climate, 65 Ma to
present. Science 292, 686–693.