Embed Size (px)
Citation preview

1
Dysregulation of Nova-mediated alternative pre-mRNA splicing in
the orbital frontal cortex of schizophrenia and bipolar disorders
Taylor Brown

2
ABSTRACT
Schizophrenia (SCZ) and bipolar disorder (BPD) are two devastating mental disorders,
garnering substantial costs to the affected individuals and society. The underlying etiology of
SCZ and BPD remains largely unknown, but SCZ and BPD are commonly viewed as a complex
disorder of brain development and plasticity involving alterations of multiple genes and various
neurobiological processes contributed by both genetic and environmental factors. Neuro-
oncological ventral antigens (Nova1 and Nova2) are neuron-specific RNA binding proteins or
splicing factors that regulate alternative pre-mRNA splicing, transcription, mRNA stability, and
trafficking of many neuronal genes, most of which are expressed in synapse and associated with
SCZ and BPD. This study is one of the first to describe the dysregulation of Nova proteins and
Nova-mediated splicing regulation in the postmortem orbital frontal cortex (OFC) of SCZ and
BPD patients. Particularly, Nova-regulated splicing in a NMDA receptor subunit (GRIN1), a
GABAA receptor subunits (GABRG2) and a calcium channel (CACNA1B) was found to be
significantly altered in the OFC of SCZ and BPD patients, most likely due to an increased
expression of Nova proteins. In addition, steady-state mRNA levels in several Nova targets
including ARHGAP21, CACNA1B, CAMK2G, EPHA5 and PLCB4, were upregulated in the
OFC of BPD and/or SCZ patients, suggesting potential Nova-mediated regulation at steady-state
mRNA level. These findings strongly endorse a novel hypothesis that dysregulation of Nova-
mediated splicing regulation in various brain regions plays an important role in development and
pathogenesis of SCZ and BPD. Most importantly, it may provide a new perspective on the
underlying etiology of SCZ and BPD, and may potentially lead to the development of novel
therapeutics for the treatment and/or prevention of SCZ and BPD. The current study may also
have an impact on studies of other psychiatric disorders such as depression and autism, some of
which have clinical and pathophysiological features similar to those seen in SCZ and BPD.

3
INTRODUCTION
Schizophrenia (SCZ) and bipolar disorders (BPD) are chronic, severe and prevalent
mental illnesses, together affecting approximately 1.5% of the population1, 2, and characterized
by affective and cognitive symptoms. SCZ patients often suffer from positive symptoms such as
hallucinations and delusions, and negative symptoms such as alogia, avolition and flat affect, and
depression. BPD patients usually experience abnormally intense emotional states or mood
episodes that can be manic, or depressive, or mixed. When psychosis is exhibited, schizophrenia
and bipolar disorders are at times mistaken for each other due to their many similarities. Both
SCZ and BPD patients have a higher suicidal rate than those nit affected with mental illness.
There are no medications that can cure SCZ and BPD, largely because the causes of SCZ and
BPD are still unknown, and most treatments using antipsychotic drugs simply focus on relieving
the symptoms by blocking receptors or transporters, and are often prescribed for both SCZ and
BPD patients. Although some of the anti-psychotic medications, such as Abilfy and Risperdol,
are prescribed to SCZ patients, they are also used as off-label medications for psychosis and with
mania. Therefore, these devastating neuropsychiatric disorders create substantial financial and
emotional burdens for affected individuals, their families and our society.
The underlying etiology of SCZ and BPD remains largely unknown despite decades of
intense research efforts. Nevertheless, SCZ and BPD should be considered as complex disorders
of brain development and plasticity involving alterations of multiple genes and various
neurobiological processes contributed by both genetic and environmental factors3-‐11. The
heritability estimated from twin/family studies is quite high (0.81 for SCZ and 0.75 for BPD)10,
which is consistent with estimates of single nucleotide polymorphism (SNP) heritabilities for
SCZ and BPD from recent genome-wide association (GWA) studies12. Linkage and genome-

4
wide association studies have suggested associations of several chromosomal regions and
multiple genes with SCZ and BPD11, 13. Several hypotheses involving neurotransmitter
dysregulation, such as the dopamine, serotonin and glutamine, calcium channel dysfunction,
immune system and neurodevelopmental dysfunction have been proposed to define the
underlying mechanisms of developing SCZ and BPD14-‐21. For example, the dopamine hypothesis
explains dysfunction of dopaminergic system in SCZ and BPD14, whereas the glutamate
hypothesis emphasizes disturbed glutamate pathways mainly involved the N-methyl-D-aspartate
(NMDA) receptor15. Neurodevelopmental hypothesis associates neruodevelopmental defects
during the prenatal and early postnatal period with SCZ and BPD22. Using neuroimaging such as
magnetic resonance imaging (MRI), neuromorphological abnormalities have been reported in a
number of brain regions of SCZ and BPD, such as the frontal lobe including the orbital frontal
cortex, temporal lobe, somatosensory and motor cortices, thalamus and hippocampus23-‐43, some
areas that were investigated in this study.
Neuro-oncological ventral antigens (Nova) including Nova1 and Nova2 were initially
identified in paraneoplastic opsoclonus myoclonus ataxia (POMA), an autoimmune neurologic
disease, as neuron-specific RNA-binding proteins44-47. Nova family proteins include Nova1 with
a molecular weight about 55 kDa on SDS PAGE, and Nova2 expressed as two forms, one with ~
55 kDa and the other, ~72 kDa (Supernova). Both Nova1 and Nova2 proteins are primarily
expressed in the central nervous system, particular in neocortical regions47. Nova proteins bind to
YCAY-rich elements of their target transcripts to regulate their alternative splicing, stability,
trafficking and transport46-48. Nova-mediated splicing regulation acts through YCAY-rich
elements that are commonly located at exon and its adjacent introns, and either enhances or
represses exon splicing depending upon where they bind and how they interact with other

5
splicing factors (Fig. 1)49-51.
Nova proteins also regulate
steady-state mRNA levels of
their target transcripts through
YCAY-rich elements located
at 5’-untranslated region
(UTR), coding regions and 3’-
UTR, as well as adjacent
introns. Importantly, a large
number of Nova-regulated
targets are proteins critical for
synaptic function and plasticity, such as NMDA receptors, calcium channels, GABAA receptors,
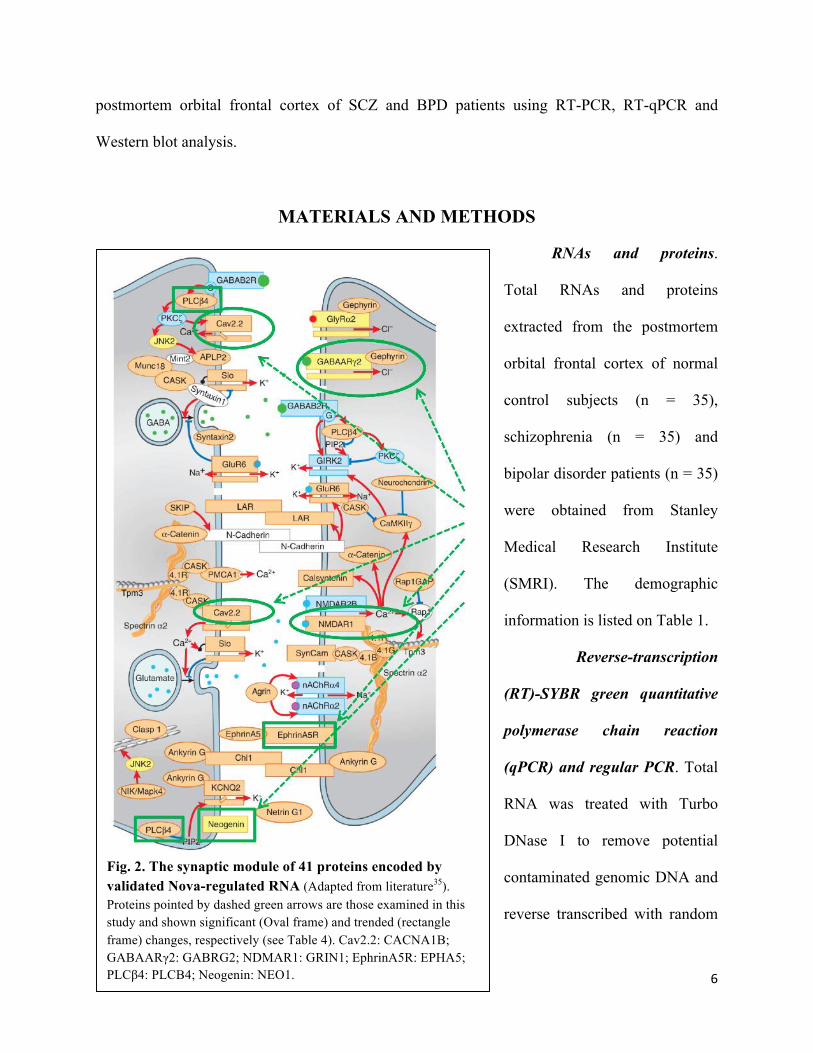
neuregulin-1, and neurexin-1 (Fig. 2)52, which have all been associated with SCZ and BPD. For
example, 6 of 32 proteins located at 22q11 whose deletion was found in 30% of SCZ patients are
the Nova targets: UFD1L, TXNRD7, COMT, ARVCF, RANBP1 and LZTR153. Moreover,
Changes in alternative splicing of several individual genes have been reported in SCZ and
BPD54-57. Many of these splicing events are regulated by Nova. For example, 5 of 8 genes
(ADAR, BIRC6, MAPK14, STXBP2 and ZDHHC3) showing significant alternative splicing
changes in SCZ patients belong to Nova-regulated targets54. All these observations provide the
rationales for a novel hypothesis that dysregulation of Nova proteins and Nova-mediated
alternative splicing plays an important role in development and pathogenesis of SCZ and BPD.
The present study examines this hypothesis by analyzing RNA and protein samples from the
Fig.1. Nova-mediated splicing regulation (Adapted from literature49-51). Shaded light orange and blue areas are the distribution of Nova binding sites (YCAY clusters45,49) positively and negatively regulated by Nova proteins for exon splicing, respectively. Fox, SR proteins, hnRNP and PTBP are also shown to influence Nova-mediated splicing74-81.
6
postmortem orbital frontal cortex of SCZ and BPD patients using RT-PCR, RT-qPCR and
Western blot analysis.
MATERIALS AND METHODS
RNAs and proteins.
Total RNAs and proteins
extracted from the postmortem
orbital frontal cortex of normal
control subjects (n = 35),
schizophrenia (n = 35) and
bipolar disorder patients (n = 35)
were obtained from Stanley
Medical Research Institute
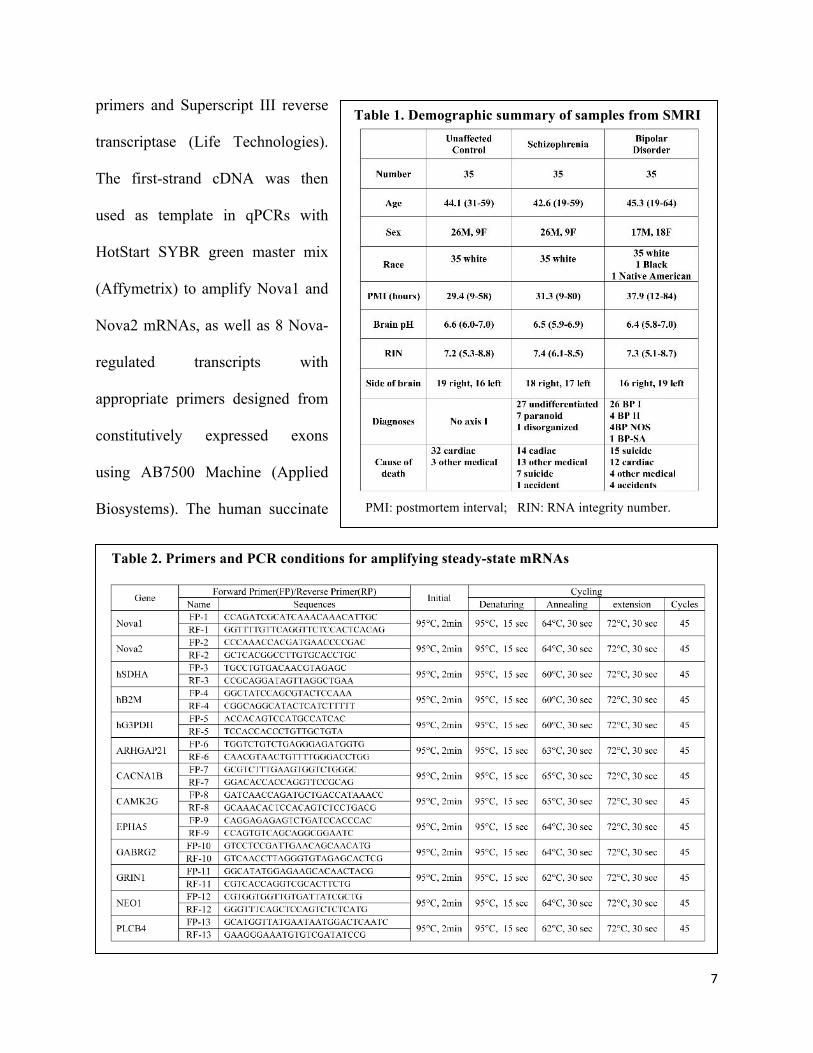
(SMRI). The demographic
information is listed on Table 1.
Reverse-transcription
(RT)-SYBR green quantitative
polymerase chain reaction
(qPCR) and regular PCR. Total
RNA was treated with Turbo
DNase I to remove potential
contaminated genomic DNA and
reverse transcribed with random
Fig. 2. The synaptic module of 41 proteins encoded by validated Nova-regulated RNA (Adapted from literature35). Proteins pointed by dashed green arrows are those examined in this study and shown significant (Oval frame) and trended (rectangle frame) changes, respectively (see Table 4). Cav2.2: CACNA1B; GABAARγ2: GABRG2; NDMAR1: GRIN1; EphrinA5R: EPHA5; PLCβ4: PLCB4; Neogenin: NEO1.
7
primers and Superscript III reverse
transcriptase (Life Technologies).
The first-strand cDNA was then
used as template in qPCRs with
HotStart SYBR green master mix
(Affymetrix) to amplify Nova1 and
Nova2 mRNAs, as well as 8 Nova-
regulated transcripts with
appropriate primers designed from
constitutively expressed exons
using AB7500 Machine (Applied
Biosystems). The human succinate
Table 1. Demographic summary of samples from SMRI
PMI: postmortem interval; RIN: RNA integrity number.
Table 2. Primers and PCR conditions for amplifying steady-state mRNAs

8
dehydrogenase subunit A (hSDHA), humann beta-2 microglobulin (hB2M) and humen
glyceraldehyde 3-phosphate dehydrogenase (hG3PDH) were used to determine normalization
factor with (C(t)hSDHA x C(t)hB2M x C(t)hG3PDH)1/3 format. All variant C(t) values were normalized
with the normalization factor to obtain ∆C(t). (∆C(t) = C(t)variant – normalization factor). To
analyze Nova-mediated splicing exons, the same first-strand cDNA was used as template in
regular PCRs with appropriate primers designed from Nova-regulated splicing exons, which
generated two PCR fragments, the longer one as exon inclusion and the shorter one as exon
exclusion. Regular PCR conditions including annealing temperature and number of cycles were
optimized. PCR products were then analyzed on 3% of MetaPhor agarose gel, stained with
ethidium bromide, imaged and quantified on ImageLab 4.1, ChemiDoc MP System (Bio-Rad).
All PCR primers and conditions are listed in Tables 2 & 3. Exon inclusion (ψ) was calculated by
dividing the percentage of the bands representing exon inclusion by the total percentage of bands
representing exon inclusion and exclusion. Splicing changes (∆ψ) was determined by subtraction
between SCZ or BPD and Control. A positive ∆ψ value indicates enhanced exon inclusion by
Nova, and negative one suppressed by Nova.
Table 3. Primers and PCR conditions for amplifying Nova-regulated exons
*: The primer sequences were adapted from Tollervey et al82.

9
Western blot analysis. Protein samples were denatured in SDS sample buffer (25mM
Tris, 0.19M glycine, 2% SDS, 0.1M DTT, 0.25% β-mecaptoethanol) by heating at 100°C for 10
min., separated on pre-casted Any kD stain-free SDS polyacrylamide gel electrophoresis (PAGE)
(Bio-Rad), and transferred onto PVDF membranes using Trans-Blot Turbo (Bio-Rad). Gels were
UV-crosslinked before transfer to conjugate florescent substrate to any proteins on the gel for the
purpose of quantifying the amount of total proteins on transferred PVDF membranes that were
used in normalization for protein loading control. This stain-free technology is designed for
assessment of protein loading with clear advantages in accuracy and simplicity over traditional
method using an antibody against individual house-keeping protein such as Actin and GAPDH.
The PVDF membranes were then blocked in a block solution containing TTBS (10mM
Tris–HCl, pH 7.4, 150mM NaCl, and 0.05% Tween 20), 5% nonfat dried milk at room
temperature for 1 h and incubated with anti-Nova1 antibody (Santa Cruz, 1:1000 dilution) or
anti-Nova2 antibody (ProteinTech, 1:800 dilution) or anti-Nova antibody (a gift from Dr. Robert
Darnell, Rockefeller University (1:3000 dilution) in the block solution at 4°C overnight. After
washing with TTBS buffer, the membrane was incubated with appropriate peroxidase-
conjugated secondary antibodies in TTBS buffer at room temperature for 1 h. After washing with
TTBS buffer, the signals were determined by using Luminata Forte Western HRP substrate
(Millipore), imaged and analyzed on ImageLab 4.1, ChemiDoc MP System. Expression level of
Nova1 and Nova2 bands was determined by dividing band intensity of Nova proteins by
intensity of total protein bands shown on PVDF membranes by stain-free technology.
Statistical analysis. Student t-test was used to analyze statistical significance.
RESULTS
10
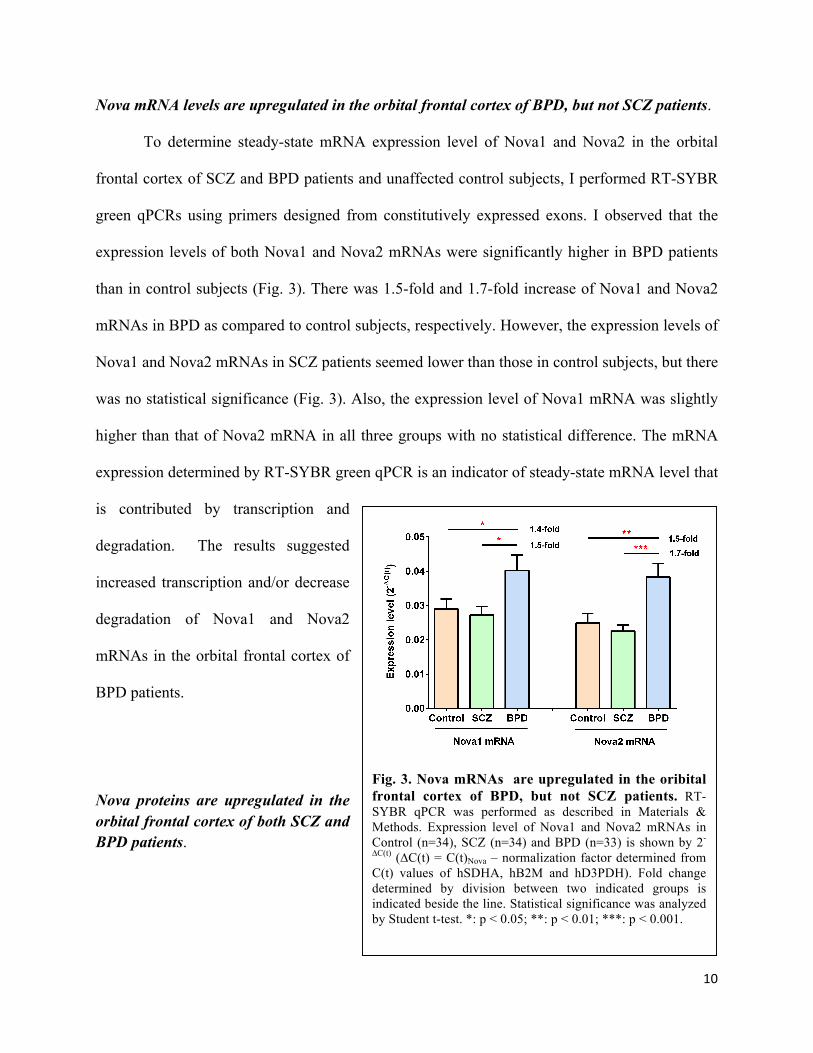
Nova mRNA levels are upregulated in the orbital frontal cortex of BPD, but not SCZ patients.
To determine steady-state mRNA expression level of Nova1 and Nova2 in the orbital
frontal cortex of SCZ and BPD patients and unaffected control subjects, I performed RT-SYBR
green qPCRs using primers designed from constitutively expressed exons. I observed that the
expression levels of both Nova1 and Nova2 mRNAs were significantly higher in BPD patients
than in control subjects (Fig. 3). There was 1.5-fold and 1.7-fold increase of Nova1 and Nova2
mRNAs in BPD as compared to control subjects, respectively. However, the expression levels of
Nova1 and Nova2 mRNAs in SCZ patients seemed lower than those in control subjects, but there
was no statistical significance (Fig. 3). Also, the expression level of Nova1 mRNA was slightly
higher than that of Nova2 mRNA in all three groups with no statistical difference. The mRNA
expression determined by RT-SYBR green qPCR is an indicator of steady-state mRNA level that
is contributed by transcription and
degradation. The results suggested
increased transcription and/or decrease
degradation of Nova1 and Nova2
mRNAs in the orbital frontal cortex of
BPD patients.
Nova proteins are upregulated in the orbital frontal cortex of both SCZ and BPD patients.
Fig. 3. Nova mRNAs are upregulated in the oribital frontal cortex of BPD, but not SCZ patients. RT-SYBR qPCR was performed as described in Materials & Methods. Expression level of Nova1 and Nova2 mRNAs in Control (n=34), SCZ (n=34) and BPD (n=33) is shown by 2-
∆C(t) (∆C(t) = C(t)Nova – normalization factor determined from C(t) values of hSDHA, hB2M and hD3PDH). Fold change determined by division between two indicated groups is indicated beside the line. Statistical significance was analyzed by Student t-test. *: p < 0.05; **: p < 0.01; ***: p < 0.001.
11
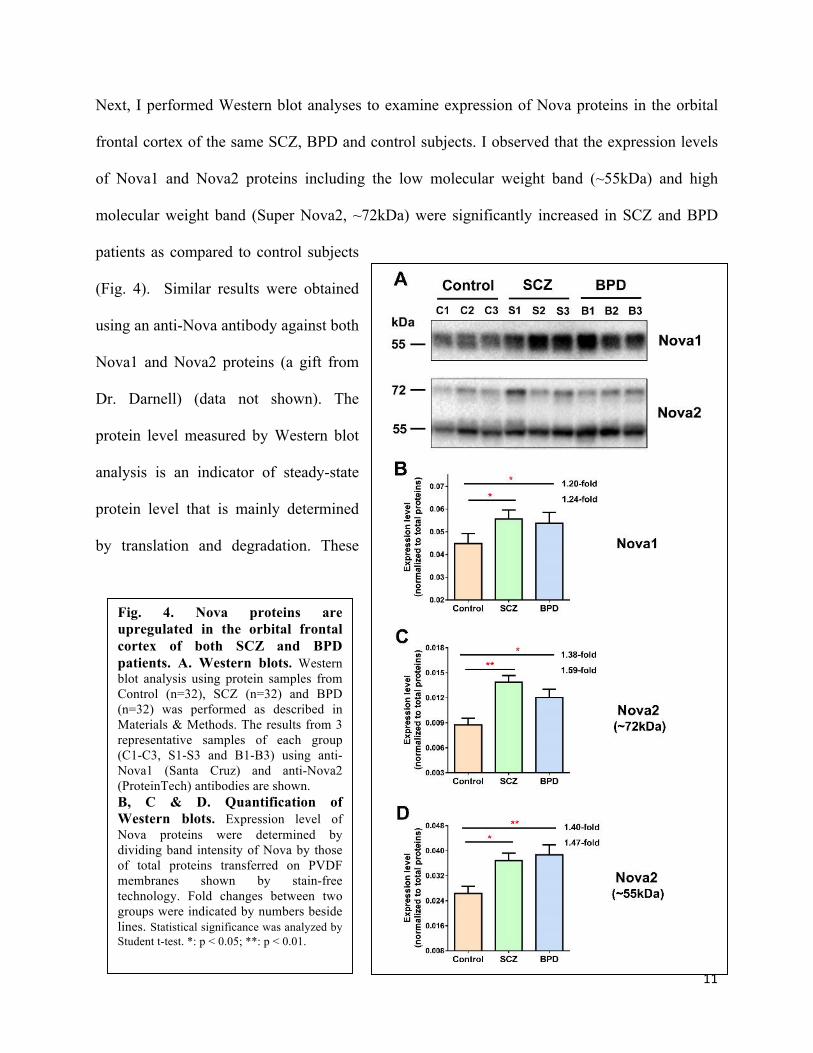
Next, I performed Western blot analyses to examine expression of Nova proteins in the orbital
frontal cortex of the same SCZ, BPD and control subjects. I observed that the expression levels
of Nova1 and Nova2 proteins including the low molecular weight band (~55kDa) and high
molecular weight band (Super Nova2, ~72kDa) were significantly increased in SCZ and BPD
patients as compared to control subjects
(Fig. 4). Similar results were obtained
using an anti-Nova antibody against both
Nova1 and Nova2 proteins (a gift from
Dr. Darnell) (data not shown). The
protein level measured by Western blot
analysis is an indicator of steady-state
protein level that is mainly determined
by translation and degradation. These
Fig. 4. Nova proteins are upregulated in the orbital frontal cortex of both SCZ and BPD patients. A. Western blots. Western blot analysis using protein samples from Control (n=32), SCZ (n=32) and BPD (n=32) was performed as described in Materials & Methods. The results from 3 representative samples of each group (C1-C3, S1-S3 and B1-B3) using anti-Nova1 (Santa Cruz) and anti-Nova2 (ProteinTech) antibodies are shown. B, C & D. Quantification of Western blots. Expression level of Nova proteins were determined by dividing band intensity of Nova by those of total proteins transferred on PVDF membranes shown by stain-free technology. Fold changes between two groups were indicated by numbers beside lines. Statistical significance was analyzed by Student t-test. *: p < 0.05; **: p < 0.01.

12
results suggested that in the orbital frontal cortex of both SCZ and BPD patients Nova proteins
are more efficiently translated and/or less degradated. The increased Nova proteins in BPD
patients were consistent with their upregulated levels of Nova mRNAs (Fig. 3), suggesting that
increased Nova mRNAs may contribute to the enhanced translation of Nova proteins in BPD
patients. On the other hand, the increased Nova proteins in SCZ patients were clearly not
contributed by the changes of their steady-state Nova mRNA levels (Fig. 3), suggesting
regulatory mechanisms at the post-transcriptional level.
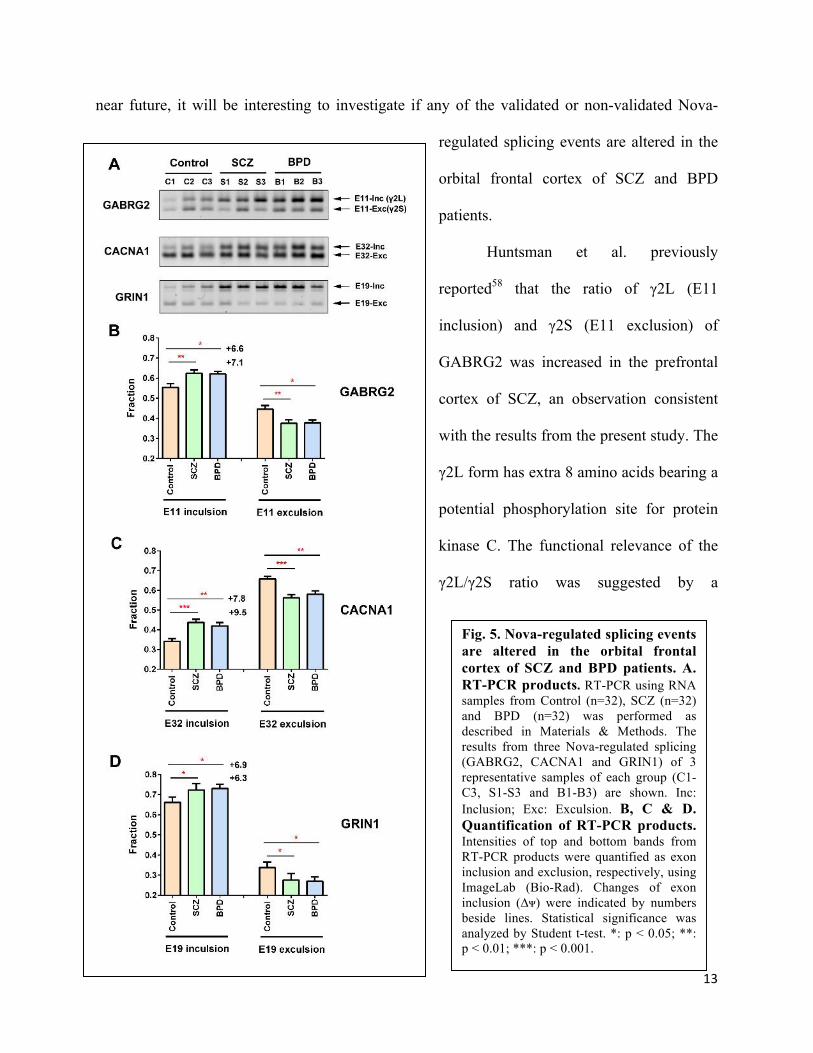
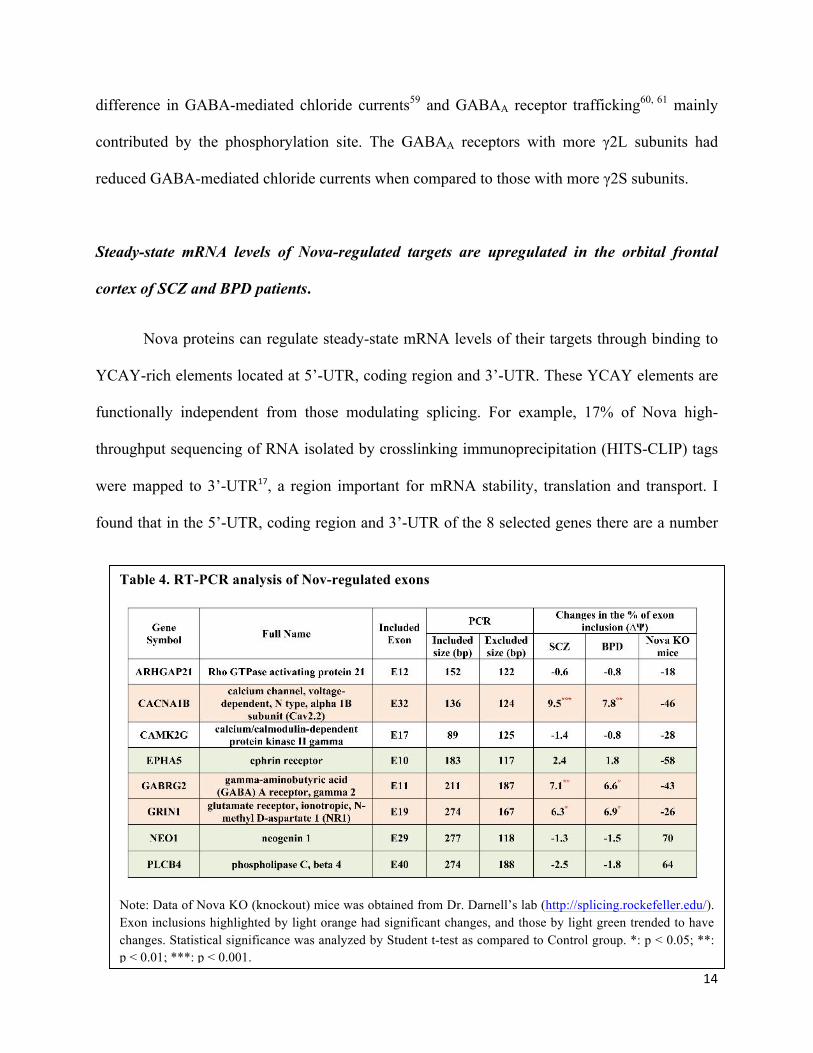
Nova-regulated splicing events are altered in the orbital frontal cortex of SCZ and BPD patients.
Nova proteins are neuron-specific splicing factors that regulate alternative splicing of
hundreds of genes, particularly neuronal genes richly expressed in synapse (Fig. 2)52. Increased
Nova proteins in SCZ and BPD patients raise questions as to whether or not these Nova-
regulated splicing events are consequently changed. I then performed RT-PCR to analyze eight
selected splicing events regulated by Nova proteins that were validated in mouse and human
exons52,82 (Table 4). I found that three Nova-regulated exons in CACNA1b, GABRG2 and
GRIN1 genes were significantly upregulated in the OFC of SCZ patients, and three Nova-
regulated exons in EPHA5, NEO1 and PLCB4 genes trended to be changed in SCZ and BPD
patients with no statistical difference, all of which were correlated with the increased Nova
proteins in these patients (Table 4 and Fig. 5). There were no changes in two Nova-regluated
exons in ARHGAP21 and CAMK2G genes. The results suggested that in SCZ and BPD patients
increased Nova proteins led to altered expression of some, but not all, Nova-regulated splicing
events or isoforms, and these alterations may contribute to development of SCZ and BPD. In the
13
near future, it will be interesting to investigate if any of the validated or non-validated Nova-
regulated splicing events are altered in the
orbital frontal cortex of SCZ and BPD
patients.
Huntsman et al. previously
reported58 that the ratio of γ2L (E11
inclusion) and γ2S (E11 exclusion) of
GABRG2 was increased in the prefrontal
cortex of SCZ, an observation consistent
with the results from the present study. The
γ2L form has extra 8 amino acids bearing a
potential phosphorylation site for protein
kinase C. The functional relevance of the
γ2L/γ2S ratio was suggested by a
Fig. 5. Nova-regulated splicing events are altered in the orbital frontal cortex of SCZ and BPD patients. A. RT-PCR products. RT-PCR using RNA samples from Control (n=32), SCZ (n=32) and BPD (n=32) was performed as described in Materials & Methods. The results from three Nova-regulated splicing (GABRG2, CACNA1 and GRIN1) of 3 representative samples of each group (C1-C3, S1-S3 and B1-B3) are shown. Inc: Inclusion; Exc: Exculsion. B, C & D. Quantification of RT-PCR products. Intensities of top and bottom bands from RT-PCR products were quantified as exon inclusion and exclusion, respectively, using ImageLab (Bio-Rad). Changes of exon inclusion (∆ᴪ) were indicated by numbers beside lines. Statistical significance was analyzed by Student t-test. *: p < 0.05; **: p < 0.01; ***: p < 0.001.
14
difference in GABA-mediated chloride currents59 and GABAA receptor trafficking60, 61 mainly
contributed by the phosphorylation site. The GABAA receptors with more γ2L subunits had
reduced GABA-mediated chloride currents when compared to those with more γ2S subunits.
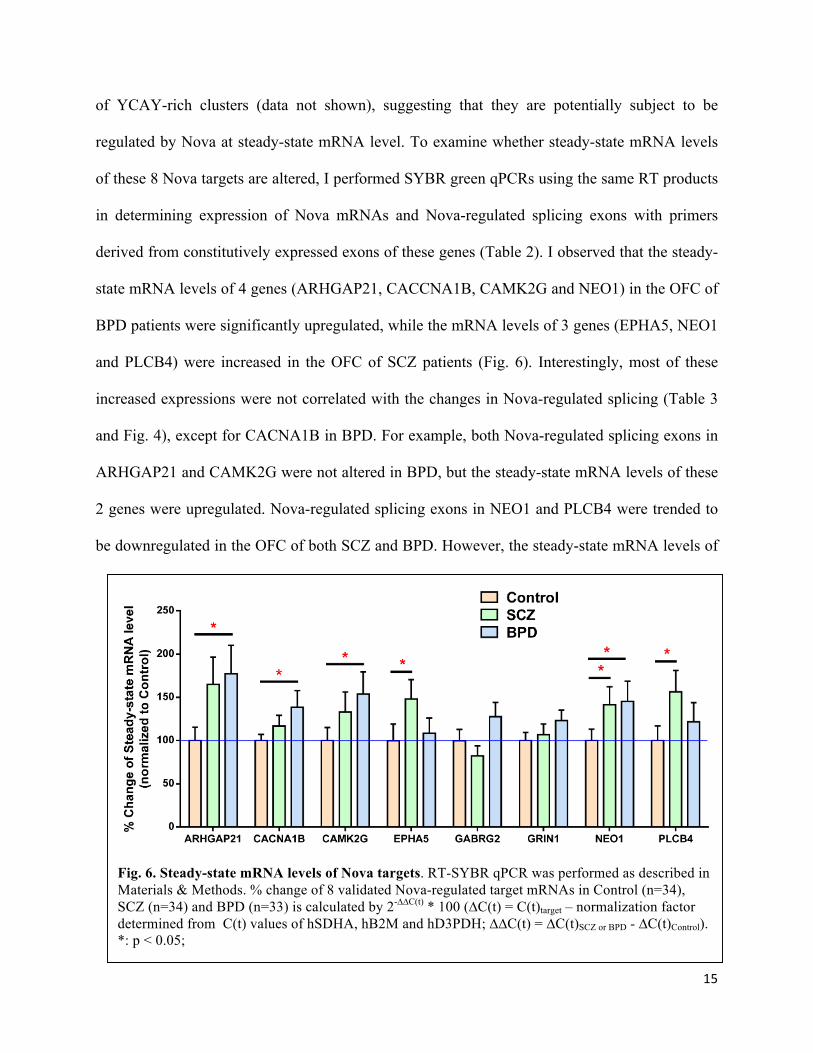
Steady-state mRNA levels of Nova-regulated targets are upregulated in the orbital frontal
cortex of SCZ and BPD patients.
Nova proteins can regulate steady-state mRNA levels of their targets through binding to
YCAY-rich elements located at 5’-UTR, coding region and 3’-UTR. These YCAY elements are
functionally independent from those modulating splicing. For example, 17% of Nova high-
throughput sequencing of RNA isolated by crosslinking immunoprecipitation (HITS-CLIP) tags
were mapped to 3’-UTR17, a region important for mRNA stability, translation and transport. I
found that in the 5’-UTR, coding region and 3’-UTR of the 8 selected genes there are a number
Table 4. RT-PCR analysis of Nov-regulated exons
Note: Data of Nova KO (knockout) mice was obtained from Dr. Darnell’s lab (http://splicing.rockefeller.edu/). Exon inclusions highlighted by light orange had significant changes, and those by light green trended to have changes. Statistical significance was analyzed by Student t-test as compared to Control group. *: p < 0.05; **: p < 0.01; ***: p < 0.001.
15
of YCAY-rich clusters (data not shown), suggesting that they are potentially subject to be
regulated by Nova at steady-state mRNA level. To examine whether steady-state mRNA levels
of these 8 Nova targets are altered, I performed SYBR green qPCRs using the same RT products
in determining expression of Nova mRNAs and Nova-regulated splicing exons with primers
derived from constitutively expressed exons of these genes (Table 2). I observed that the steady-
state mRNA levels of 4 genes (ARHGAP21, CACCNA1B, CAMK2G and NEO1) in the OFC of
BPD patients were significantly upregulated, while the mRNA levels of 3 genes (EPHA5, NEO1
and PLCB4) were increased in the OFC of SCZ patients (Fig. 6). Interestingly, most of these
increased expressions were not correlated with the changes in Nova-regulated splicing (Table 3
and Fig. 4), except for CACNA1B in BPD. For example, both Nova-regulated splicing exons in
ARHGAP21 and CAMK2G were not altered in BPD, but the steady-state mRNA levels of these
2 genes were upregulated. Nova-regulated splicing exons in NEO1 and PLCB4 were trended to
be downregulated in the OFC of both SCZ and BPD. However, the steady-state mRNA levels of
Fig. 6. Steady-state mRNA levels of Nova targets. RT-SYBR qPCR was performed as described in Materials & Methods. % change of 8 validated Nova-regulated target mRNAs in Control (n=34), SCZ (n=34) and BPD (n=33) is calculated by 2-∆∆C(t) * 100 (∆C(t) = C(t)target – normalization factor determined from C(t) values of hSDHA, hB2M and hD3PDH; ∆∆C(t) = ∆C(t)SCZ or BPD - ∆C(t)Control). *: p < 0.05;

16
these 2 genes were significantly increased in the OFC of SCZ and/or BPD. These results
suggested that Nova-mediated splicing regulation is independent from potential Nova-mediated
modulation at steady-state mRNA level, although it needs to be experimentally verified.
DISCUSSION
For the first time, the present study demonstrates alterations of Nova1 and Nova2
proteins, and their regulated splicing events in the orbital frontal cortex of SCZ and BPD patients.
This provides strong support for the hypothesis that dysregulation of Nova-mediated alternative
splicing in various brain regions plays an important role in development and pathogenesis of
SCZ and BPD. What makes the hypothesis more attractive is that Nova proteins regulate
expression of many protein isoforms that are expressed in synapses (Fig. 2), some of which have
already been linked to the pathogenesis of these disorders. For example, altered expression and
function of NMDA receptors and GABAA receptors have been extensively reported in various
brain regions of SCZ and BPD patients. Genetic variations in voltage-dependent calcium
channels, NMDA and GABAA receptors have been linked to SCZ and BPD19. As illustrated by
the present study, Nova-mediated alternative splicing of a NMDA receptor subunit (GRIN1), a
GABAA receptor subunit (GABRG2) and a voltage-dependent calcium channel subunit
(CACNA1) is dysregulated in the orbital frontal cortex of SCZ and BPD, as a result of the
increased Nova proteins. These findings raise a suggestive hypothesis that Nova-mediated
splicing regulation is a key player in development and pathogenesis of SCZ and BPD.
It should be noted that the magnitude of the dysregulation was not dramatic for all three
Nova-regulated exons, and the changes in the % of exon inclusion (∆ψ) ranged from 6.3 (GRIN1
in SCZ) to 9.5 (CACNA1B in SCZ) (Table 3 & Fig. 5). This may be correlated with similar
degree of changes in Nova proteins seen in SCZ and BPD patients, ranging from 1.20-fold

17
increase of Nova1 in BPD to 1.59-fold increase of Nova2(~72kDa) in SCZ (Fig. 4). However,
summation of these small changes in each of these three Nova-regulated events could be
dramatic. Nova proteins regulate alternative splicing of at least 48 validated splicing targets
including neurotransmitter receptors, cation channels, adhesion and scaffold proteins, all of
which are highly expressed in synapse52, in addition to others not yet validated. The present
study reveals that aproximately 37.5% of Nova-regulated splicing events (three of eight) were
dysregulated. Based on this rate, it will account for ~ 18 synaptic isoforms (37.5% of 48). One
can imagine that not only are there cumulative effects, but also synergistic effects among these
dysregulated proteins, which contribute greatly to alterations in synaptic functions and plasticity,
a pathological consequence commonly seen in SCZ and BPD patients. Although the functional
relevance of each Nova-regulated splicing isoform and the functional relationship among these
isoforms in SCZ and BPD needs to be further investigated, the current findings highlight the
potential influence of Nova-mediated splicing regulation in development and pathogenesis of
SCZ and BPD.
The present findings also raise questions regarding how Nova proteins are upregulated in
the orbital frontal cortex of SCZ and BPD patients. As demonstrated here, the underlying
mechanisms of Nova protein regulation are different between SCZ and BPD. In SCZ, the
increase of Nova proteins with no alteration of their mRNAs suggests regulatory mechanisms
mainly at the post-transcriptional level. However, upregulation of both Nova mRNAs and
proteins in BPD implicates the mechanisms at both transcription and/or post-transcription levels.
microRNAs (miRNAs) are a class of non-coding RNAs that regulate gene expression at both
transcription and translation levels, and play important roles in many biological processes such
as neuronal development and differentiation, synapse formation and neuronal plasticity.

18
Dysregulation of miRNAs has been linked to the cause of several brain disorders including SCZ,
BPS, major depression, Parkinson’s disease, as well as drug addiction62-69. It is most likely that
Nova mRNAs are regulated by miRNAs. Multiple miRNAs have been shown to target Nova
mRNAs based upon several computer models including miRBase, Targetscan, RNAhybrid,
RNA22 and miRWalk. Interestingly, Nova2 is a potential target of mir-185 located within 22q11
whose deletion was found in 30% of SCZ patients53, 70. In the PFC of Df(16)A+/- mice, a mouse
model mimicking human 22q11.2 microdeletion, 10 of 59 downregulated miRNAs potentially
targets Nova171, 72. Downregulation of these miRNAs may explain the upregulation of Nova
proteins in the OFC of SCZ and BPD patients, although speculated. Additionally, alterations of
several miRNAs previously reported in association with SCZ and BPD are also on the list of
miRNAs predicted to target Nova mRNAs, such as miR-13263 and miR-2273, suggesting that
these miRNAs also play an important role in regulating expression of Nova proteins and Nova-
mediated splicing isoforms. If confirmed, it will provide a functional link between miRNA and
Nova-mediated splicing regulation.
Several brain regions including the cingulate cortex, frontal cortex, temporal pole,
thalamus, and hippocampus have been indicated to be critically involved in pathogenesis of SCZ
and BPD. Although the present study only examined the orbital frontal cortex, I hope to extend
our study on expression of Nova proteins and Nova-mediated splicing isoforms in SCZ and BPD
to the other brain regions in the near future. It will be very interesting to see if the other brain
regions have the same or different changes as those observed in the orbital frontal cortex.
Alternative splicing involves coordination of multiple components including cis-acting
elements located within the exons and proximal or distal introns, trans-acting factors, including
the serine/arginine-rich (SR) family proteins, hnRNP proteins, the CELF protein family and

19
Nova proteins, that interact with the cis-acting elements, and their interactions with the basal
spliceosome. It has been shown that Nova-mediated splicing is greatly influenced by other
splicing factors such as Fox2, PTBP2, hnRNPA1, hnRNPH, hnRNPL and SR proteins (Fig. 1)50,
74-81. Now, it will be interesting to investigate whether these splicing factors are also dysregulated
in SCZ and BPD.
The present study used RNA and protein samples from dissected regions. One caveat of
using these samples is heterogeneity of various cell types in the samples that may differentially
express Nova mRNAs and proteins. It will be essential to examine the expression of Nova
mRNAs and proteins at an anatomic level by using in situ hybridization for Nova mRNAs and
immunohistochemistry for Nova proteins, if such brain tissues are available. The results from
these studies can further direct studies using laser capture microdissection approach that allows
collecting single cell or very small region for examining expression of Nova-regulated splicing
events.
Alterations of steady-state mRNAs in a number of genes bearing YCAY-rich elements at
5’-UTR, coding region and 3’-UTR in the OFC of SCZ and/or BPD patients raises questions
whether or not Nova proteins indeed regulate these targets, and if so, what the underlying
mechanisms are. Steady-state mRNA level is determined by many other factors such as
transcription factors that can also be altered in the OFC of SCZ and/or BPD patients. I believe
that alteration of Nova proteins can coordinate with changes of other factors, and such
coordination at least partly contributed to modulation of these steady-state mRNA levels.

20
CONCLUSION AND FUTURE WORK
The present study shows that Nova proteins and Nova-regulated splicing isoforms were
dysregulated in the postmortem orbital frontal cortex of SCZ and BPD, providing a strong
support for a new hypothesis that dysregulation of Nova-mediated splicing in brain plays an
important role in development and pathogenesis of SCZ and BPD. The significances of the
present findings are three-fold. First, majority of Nova-regulated genes are critical in
maintaining normal functions of synapse, particularly in the cortex regions. Summation of small
changes in splicing of these genes can potentially alter synaptic formation, development and
plasticity, leading to the development of SCZ and BPD. Alterations of just three Nova-regulated
splicing isoforms in GRIN1, GABA and CACNA1 genes discovered in the present study bridge
together of three systems, the NMDA receptor, the GABAA receptor and calcium channels,
whose alterations are commonly used for explaining the pathogenesis of SCZ and BPD.
Involvement of multiple Nova-regulated targets is also supporting the notion that SCZ and BPD
are complex brain disorders contributed by dysfunction of multiple genes. Second, further
exploring mechanisms underlying regulation of Nova mRNAs and proteins and their targets will
possibly link to regulation of miRNA and/or epigenetics, as well as other splicing factors,
providing even broad and deep understanding of the roles of Nova proteins in SCZ and BPD.
Third, if we can establish the functional relevance of Nova-regulated isoforms, it will lead to the
development of new drugs for the treatment and/or prevention of SCZ and BPD. For example,
we can use siRNA targeting Nova mRNAs to downregulate Nova proteins in the orbital frontal
cortex and therefore correct aberrant Nova-mediated splicing. Alternatively, we can deliver Nova
proteins into brain regions where Nova proteins are downregulated by using an appropriate viral

21
vector. Also by using morpholino antisense oligonucleotide approach, we can correct altered
splicing events.
The present study raises many intriguing questions that need to be addressed in the near
future regarding the role of Nova-mediated splicing regulation in development of SCZ and BPD.
1) How are the other 40 synaptic protein isoforms regulated by Nova proteins expressed in the
orbital frontal cortex of SCZ and BPD? 2) How are Nova proteins and Nova-regulated isoforms
expressed in other brain regions of SCZ and BPD? 3) What are the molecular mechanisms
underlying regulation of Nova mRNAs and proteins? 4) What is the functional relevance of
Nova-regulated isoforms, particularly for the genes already shown to be important in SCZ and
BPD, such as NMDA receptor (GRIN1), GABAA receptor (GABRG1) and calcium channel
(CACNA1) that are reported here? 5) Can we develop an animal model that mimics SCZ or BPD
in which Nova proteins and Nova-mediated splicing are dysregulated in a similar way to those in
the human postmortem brains? 6) Are other splicing factors, particularly for those related to
Nova-mediated splicing regulation, dysregulated in brain regions of SCZ and BPD? 7) What is
the impact of current antipsychotic drugs used for treating SCZ and BPD on expression of Nova
proteins and Nova-regulated isoforms? 8) What are the potentials for developing new
therapeutics to treat and/or prevent SCZ and BPD based upon the hypothesis raised by the
present study.

22
REFERENCES
1. Saha S, Chant D, and McGrath J (2008) Meta-analyses of the incidence and prevalence of schizophrenia: conceptual and methodological issues. Int J Methods Psychiatr Res 17 (1):55-61
2. Merikangas KR, Akiskal HS, Angst J, Greenberg PE, Hirschfeld RM, Petukhova M, and Kessler RC (2007) Lifetime and 12-month prevalence of bipolar spectrum disorder in the National Comorbidity Survey replication. Arch Gen Psychiatry 64 (5):543-552
3. Penzes P, Buonanno A, Passafaro M, Sala C, and Sweet RA (2013) Developmental vulnerability of synapses and circuits associated with neuropsychiatric disorders. J Neurochem 126 (2):165-182
4. Yin DM, Chen YJ, Sathyamurthy A, Xiong WC, and Mei L (2012) Synaptic dysfunction in schizophrenia. Adv Exp Med Biol 970:493-516
5. Balu DT and Coyle JT (2011) Neuroplasticity signaling pathways linked to the pathophysiology of schizophrenia. Neurosci Biobehav Rev 35 (3):848-870
6. Ross CA, Margolis RL, Reading SA, Pletnikov M, and Coyle JT (2006) Neurobiology of schizophrenia. Neuron 52 (1):139-153
7. Karam CS, Ballon JS, Bivens NM, Freyberg Z, Girgis RR, Lizardi-Ortiz JE, Markx S, Lieberman JA, and Javitch JA (2010) Signaling pathways in schizophrenia: emerging targets and therapeutic strategies. Trends Pharmacol Sci 31 (8):381-390
8. McCullumsmith RE, Clinton SM, and Meador-Woodruff JH (2004) Schizophrenia as a disorder of neuroplasticity. Int Rev Neurobiol 59:19-45
9. Arnold SE, Talbot K, and Hahn CG (2005) Neurodevelopment, neuroplasticity, and new genes for schizophrenia. Prog Brain Res 147:319-345
10. Sullivan PF, Daly MJ, and O'Donovan M (2012) Genetic architectures of psychiatric disorders: the emerging picture and its implications. Nat Rev Genet 13 (8):537-551
11. Segurado R, Detera-Wadleigh SD, Levinson DF, Lewis CM, Gill M, Nurnberger JI, Jr., Craddock N et al (2003) Genome scan meta-analysis of schizophrenia and bipolar disorder, part III: Bipolar disorder. Am J Hum Genet 73 (1):49-62
12. Lee SH, Ripke S, Neale BM, Faraone SV, Purcell SM, Perlis RH, Mowry BJ et al (2013) Genetic relationship between five psychiatric disorders estimated from genome-wide SNPs. Nat Genet 45 (9):984-994

23
13. Lewis CM, Levinson DF, Wise LH, DeLisi LE, Straub RE, Hovatta I, Williams NM et al (2003) Genome scan meta-analysis of schizophrenia and bipolar disorder, part II: Schizophrenia. Am J Hum Genet 73 (1):34-48
14. Toda M and Abi-Dargham A (2007) Dopamine hypothesis of schizophrenia: making sense of it all. Curr Psychiatry Rep 9 (4):329-336
15. Moghaddam B and Javitt D (2012) From revolution to evolution: the glutamate hypothesis of schizophrenia and its implication for treatment. Neuropsychopharmacology 37 (1):4-15
16. Kinros J, Reichenberg A, and Frangou S (2010) The neurodevelopmental theory of schizophrenia: evidence from studies of early onset cases. Isr J Psychiatry Relat Sci 47 (2):110-117
17. Altamura AC, Pozzoli S, Fiorentini A, and Dell'osso B (2013) Neurodevelopment and inflammatory patterns in schizophrenia in relation to pathophysiology. Prog Neuropsychopharmacol Biol Psychiatry 42:63-70
18. Geyer MA and Vollenweider FX (2008) Serotonin research: contributions to understanding psychoses. Trends Pharmacol Sci 29 (9):445-453
19. Giegling I, Genius J, Benninghoff J, and Rujescu D (2010) Genetic findings in schizophrenia patients related to alterations in the intracellular Ca-homeostasis. Prog Neuropsychopharmacol Biol Psychiatry 34 (8):1375-1380
20. Berridge MJ (2012) Calcium signalling remodelling and disease. Biochem Soc Trans 40 (2):297-309
21. Schizophrenia Research Forum (http://www.schizohreniaforum.org/). 2013. 22. Rapoport JL, Addington AM, Frangou S, and Psych MR (2005) The neurodevelopmental
model of schizophrenia: update 2005. Mol Psychiatry 10 (5):434-449
23. Williamson PC and Allman JM (2012) A framework for interpreting functional networks in schizophrenia. Front Hum Neurosci 6:184
24. Joshi D, Fung SJ, Rothwell A, and Weickert CS (2012) Higher gamma-aminobutyric acid neuron density in the white matter of orbital frontal cortex in schizophrenia. Biol Psychiatry 72 (9):725-733
25. Kumra S, Ashtari M, Wu J, Hongwanishkul D, White T, Cervellione K, Cottone J, and Szeszko PR (2011) Gray matter volume deficits are associated with motor and attentional impairments in adolescents with schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry 35 (4):939-943
26. Rosso IM, Makris N, Thermenos HW, Hodge SM, Brown A, Kennedy D, Caviness VS, Faraone SV, Tsuang MT, and Seidman LJ (2010) Regional prefrontal cortex gray matter volumes in youth at familial risk for schizophrenia from the Harvard Adolescent High Risk Study. Schizophr Res 123 (1):15-21

24
27. Takayanagi Y, Takahashi T, Orikabe L, Masuda N, Mozue Y, Nakamura K, Kawasaki Y, Itokawa M, Sato Y, Yamasue H, Kasai K, Okazaki Y, and Suzuki M (2010) Volume reduction and altered sulco-gyral pattern of the orbitofrontal cortex in first-episode schizophrenia. Schizophr Res 121 (1-3):55-65
28. Fornito A, Yucel M, Patti J, Wood SJ, and Pantelis C (2009) Mapping grey matter reductions in schizophrenia: an anatomical likelihood estimation analysis of voxel-based morphometry studies. Schizophr Res 108 (1-3):104-113
29. Nakamura M, Nestor PG, Levitt JJ, Cohen AS, Kawashima T, Shenton ME, and McCarley RW (2008) Orbitofrontal volume deficit in schizophrenia and thought disorder. Brain 131 (Pt 1):180-195
30. Szeszko PR, Robinson DG, Sevy S, Kumra S, Rupp CI, Betensky JD, Lencz T, Ashtari M, Kane JM, Malhotra AK, Gunduz-Bruce H, Napolitano B, and Bilder RM (2007) Anterior cingulate grey-matter deficits and cannabis use in first-episode schizophrenia. Br J Psychiatry 190:230-236
31. Gur RE, Cowell PE, Latshaw A, Turetsky BI, Grossman RI, Arnold SE, Bilker WB, and Gur RC (2000) Reduced dorsal and orbital prefrontal gray matter volumes in schizophrenia. Arch Gen Psychiatry 57 (8):761-768
32. Joseph R (1999) Frontal lobe psychopathology: mania, depression, confabulation, catatonia, perseveration, obsessive compulsions, and schizophrenia. Psychiatry 62 (2):138-172
33. Vita A, De PL, Deste G, and Sacchetti E (2013) Progressive loss of cortical gray matter in schizophrenia: a meta-analysis and meta-regression of longitudinal MRI studies. Transl Psychiatry 3:e275
34. Brunet-Gouet E and Decety J (2006) Social brain dysfunctions in schizophrenia: a review of neuroimaging studies. Psychiatry Res 148 (2-3):75-92
35. Shenton ME, Dickey CC, Frumin M, and McCarley RW (2001) A review of MRI findings in schizophrenia. Schizophr Res 49 (1-2):1-52
36. Taylor SF, Kang J, Brege IS, Tso IF, Hosanagar A, and Johnson TD (2012) Meta-analysis of functional neuroimaging studies of emotion perception and experience in schizophrenia. Biol Psychiatry 71 (2):136-145
37. Jardri R, Pouchet A, Pins D, and Thomas P (2011) Cortical activations during auditory verbal hallucinations in schizophrenia: a coordinate-based meta-analysis. Am J Psychiatry 168 (1):73-81
38. Andreasen NC, Arndt S, Swayze V, Cizadlo T, Flaum M, O'Leary D, Ehrhardt JC, and Yuh WT (1994) Thalamic abnormalities in schizophrenia visualized through magnetic resonance image averaging. Science 266 (5183):294-298

25
39. Corey-Bloom J, Jernigan T, Archibald S, Harris MJ, and Jeste DV (1995) Quantitative magnetic resonance imaging of the brain in late-life schizophrenia. Am J Psychiatry 152 (3):447-449
40. Honea R, Crow TJ, Passingham D, and Mackay CE (2005) Regional deficits in brain volume in schizophrenia: a meta-analysis of voxel-based morphometry studies. Am J Psychiatry 162 (12):2233-2245
41. Arnone D, Cavanagh J, Gerber D, Lawrie SM, Ebmeier KP, and McIntosh AM (2009) Magnetic resonance imaging studies in bipolar disorder and schizophrenia: meta-analysis. Br J Psychiatry 195 (3):194-201
42. Beyer JL, Young R, Kuchibhatla M, and Krishnan KR (2009) Hyperintense MRI lesions in bipolar disorder: A meta-analysis and review. Int Rev Psychiatry 21 (4):394-409
43. Freedman R (2007) Neuronal dysfunction and schizophrenia symptoms. Am J Psychiatry 164 (3):385-390
44. Buckanovich RJ, Posner JB, and Darnell RB (1993) Nova, the paraneoplastic Ri antigen, is homologous to an RNA-binding protein and is specifically expressed in the developing motor system. Neuron 11 (4):657-672
45. Buckanovich RJ, Yang YY, and Darnell RB (1996) The onconeural antigen Nova-1 is a neuron-specific RNA-binding protein, the activity of which is inhibited by paraneoplastic antibodies. J Neurosci 16 (3):1114-1122
46. Buckanovich RJ and Darnell RB (1997) The neuronal RNA binding protein Nova-1 recognizes specific RNA targets in vitro and in vivo. Mol Cell Biol 17 (6):3194-3201
47. Yang YY, Yin GL, and Darnell RB (1998) The neuronal RNA-binding protein Nova-2 is implicated as the autoantigen targeted in POMA patients with dementia. Proc Natl Acad Sci U S A 95 (22):13254-13259
48. Jensen KB, Dredge BK, Stefani G, Zhong R, Buckanovich RJ, Okano HJ, Yang YY, and Darnell RB (2000) Nova-1 regulates neuron-specific alternative splicing and is essential for neuronal viability. Neuron 25 (2):359-371
49. Ule J, Stefani G, Mele A, Ruggiu M, Wang X, Taneri B, Gaasterland T, Blencowe BJ, and Darnell RB (2006) An RNA map predicting Nova-dependent splicing regulation. Nature 444 (7119):580-586
50. Allen SE, Darnell RB, and Lipscombe D (2010) The neuronal splicing factor Nova controls alternative splicing in N-type and P-type CaV2 calcium channels. Channels (Austin ) 4 (6):483-489
51. Darnell RB (2013) RNA protein interaction in neurons. Annu Rev Neurosci 36:243-270

26
52. Ule J, Ule A, Spencer J, Williams A, Hu JS, Cline M, Wang H, Clark T, Fraser C, Ruggiu M, Zeeberg BR, Kane D, Weinstein JN, Blume J, and Darnell RB (2005) Nova regulates brain-specific splicing to shape the synapse. Nat Genet 37 (8):844-852
53. Earls LR and Zakharenko SS (2013) A Synaptic Function Approach to Investigating Complex Psychiatric Diseases. Neuroscientist
54. Glatt SJ, Cohen OS, Faraone SV, and Tsuang MT (2011) Dysfunctional gene splicing as a potential contributor to neuropsychiatric disorders. Am J Med Genet B Neuropsychiatr Genet 156B (4):382-392
55. Morikawa T and Manabe T (2010) Aberrant regulation of alternative pre-mRNA splicing in schizophrenia. Neurochem Int 57 (7):691-704
56. Hashimoto R, Straub RE, Weickert CS, Hyde TM, Kleinman JE, and Weinberger DR (2004) Expression analysis of neuregulin-1 in the dorsolateral prefrontal cortex in schizophrenia. Mol Psychiatry 9 (3):299-307
57. Liu X, Bates R, Yin DM, Shen C, Wang F, Su N, Kirov SA, Luo Y, Wang JZ, Xiong WC, and Mei L (2011) Specific regulation of NRG1 isoform expression by neuronal activity. J Neurosci 31 (23):8491-8501
58. Huntsman MM, Tran BV, Potkin SG, Bunney WE, Jr., and Jones EG (1998) Altered ratios of alternatively spliced long and short gamma2 subunit mRNAs of the gamma-amino butyrate type A receptor in prefrontal cortex of schizophrenics. Proc Natl Acad Sci U S A 95 (25):15066-15071
59. Krishek BJ, Xie X, Blackstone C, Huganir RL, Moss SJ, and Smart TG (1994) Regulation of GABAA receptor function by protein kinase C phosphorylation. Neuron 12 (5):1081-1095
60. Meier J and Grantyn R (2004) Preferential accumulation of GABAA receptor gamma 2L, not gamma 2S, cytoplasmic loops at rat spinal cord inhibitory synapses. J Physiol 559 (Pt 2):355-365
61. Alldred MJ, Mulder-Rosi J, Lingenfelter SE, Chen G, and Luscher B (2005) Distinct gamma2 subunit domains mediate clustering and synaptic function of postsynaptic GABAA receptors and gephyrin. J Neurosci 25 (3):594-603
62. Miller BH and Wahlestedt C (2010) MicroRNA dysregulation in psychiatric disease. Brain Res 1338:89-99
63. Miller BH, Zeier Z, Xi L, Lanz TA, Deng S, Strathmann J, Willoughby D, Kenny PJ, Elsworth JD, Lawrence MS, Roth RH, Edbauer D, Kleiman RJ, and Wahlestedt C (2012) MicroRNA-132 dysregulation in schizophrenia has implications for both neurodevelopment and adult brain function. Proc Natl Acad Sci U S A

27
64. Kim AH, Reimers M, Maher B, Williamson V, McMichael O, McClay JL, van den Oord EJ, Riley BP, Kendler KS, and Vladimirov VI (2010) MicroRNA expression profiling in the prefrontal cortex of individuals affected with schizophrenia and bipolar disorders. Schizophr Res 124 (1-3):183-191
65. Moreau MP, Bruse SE, David-Rus R, Buyske S, and Brzustowicz LM (2011) Altered microRNA expression profiles in postmortem brain samples from individuals with schizophrenia and bipolar disorder. Biol Psychiatry 69 (2):188-193
66. Wong J, Duncan CE, Beveridge NJ, Webster MJ, Cairns MJ, and Weickert CS (2013) Expression of NPAS3 in the human cortex and evidence of its posttranscriptional regulation by miR-17 during development, with implications for schizophrenia. Schizophr Bull 39 (2):396-406
67. Im HI and Kenny PJ (2012) MicroRNAs in neuronal function and dysfunction. Trends Neurosci 35 (5):325-334
68. Beveridge NJ and Cairns MJ (2012) MicroRNA dysregulation in schizophrenia. Neurobiol Dis 46 (2):263-271
69. Xu B, Karayiorgou M, and Gogos JA (2010) MicroRNAs in psychiatric and neurodevelopmental disorders. Brain Res 1338:78-88
70. Xu B, Hsu PK, Stark KL, Karayiorgou M, and Gogos JA (2013) Derepression of a neuronal inhibitor due to miRNA dysregulation in a schizophrenia-related microdeletion. Cell 152 (1-2):262-275
71. Stark KL, Xu B, Bagchi A, Lai WS, Liu H, Hsu R, Wan X, Pavlidis P, Mills AA, Karayiorgou M, and Gogos JA (2008) Altered brain microRNA biogenesis contributes to phenotypic deficits in a 22q11-deletion mouse model. Nat Genet 40 (6):751-760
72. Ingenuity (http://www/ingenuity.com/). 2013. 73. Moreau MP, Bruse SE, David-Rus R, Buyske S, and Brzustowicz LM (2011) Altered
microRNA expression profiles in postmortem brain samples from individuals with schizophrenia and bipolar disorder. Biol Psychiatry 69 (2):188-193
74. Zhang C, Frias MA, Mele A, Ruggiu M, Eom T, Marney CB, Wang H, Licatalosi DD, Fak JJ, and Darnell RB (2010) Integrative modeling defines the Nova splicing-regulatory network and its combinatorial controls. Science 329 (5990):439-443
75. Lipscombe D, Allen SE, and Toro CP (2013) Control of neuronal voltage-gated calcium ion channels from RNA to protein. Trends Neurosci 36 (10):598-609
76. Licatalosi DD, Yano M, Fak JJ, Mele A, Grabinski SE, Zhang C, and Darnell RB (2012) Ptbp2 represses adult-specific splicing to regulate the generation of neuronal precursors in the embryonic brain. Genes Dev 26 (14):1626-1642

28
77. Witten JT and Ule J (2011) Understanding splicing regulation through RNA splicing maps. Trends Genet 27 (3):89-97
78. Chabot B, Blanchette M, Lapierre I, and La Branch (1997) An intron element modulating 5' splice site selection in the hnRNP A1 pre-mRNA interacts with hnRNP A1. Mol Cell Biol 17 (4):1776-1786
79. Wang E and Cambi F (2009) Heterogeneous nuclear ribonucleoproteins H and F regulate the proteolipid protein/DM20 ratio by recruiting U1 small nuclear ribonucleoprotein through a complex array of G runs. J Biol Chem 284 (17):11194-11204
80. Hui J, Hung LH, Heiner M, Schreiner S, Neumuller N, Reither G, Haas SA, and Bindereif A (2005) Intronic CA-repeat and CA-rich elements: a new class of regulators of mammalian alternative splicing. EMBO J 24 (11):1988-1998
81. Polydorides AD, Okano HJ, Yang YY, Stefani G, and Darnell RB (2000) A brain-enriched polypyrimidine tract-binding protein antagonizes the ability of Nova to regulate neuron-specific alternative splicing. Proc Natl Acad Sci U S A 97 (12):6350-6355
82. Tollervey JR, Wang Z, Hortobagyi T, Witten JT, Zarnack K, Kayikci M, Clark TA,
Schweitzer AC, Rot G, Curk T, Zupan B, Rogelj B, Shaw CE, and Ule J (2011) Analysis of alternative splicing associated with aging and neurodegeneration in the human brain. Genome Res 21 (10):1572-1582
![C103-E099 FTIR Talk Letter Vol. 23 · First, to edit the report template, open the [Edit Printform] window. To display the [Edit Printform] window, click the [Edit Printform] tab](https://img.pdfslide.us/doc/110x75/5fa1f6920b4b01638f6ac9ce/c103-e099-ftir-talk-letter-vol-23-first-to-edit-the-report-template-open-the.jpg)




![FEC report [090819] final edit cover pic](https://img.pdfslide.us/doc/110x75/61fc83328d33c02b785e0a65/fec-report-090819-final-edit-cover-pic.jpg)























