Embed Size (px)
Citation preview

Research ArticleAntioxidant Supplement Inhibits Skeletal Muscle ConstitutiveAutophagy rather than Fasting-Induced Autophagy in Mice
Zhengtang Qi12 Qiang He12 Liu Ji12 and Shuzhe Ding12
1 Key Laboratory of Adolescent Health Assessment and Exercise Intervention Ministry of EducationEast China Normal University Shanghai 200241 China
2 College of Physical Education and Health East China Normal University Shanghai 200241 China
Correspondence should be addressed to Shuzhe Ding szdingtyxxecnueducn
Received 8 January 2014 Revised 28 April 2014 Accepted 16 May 2014 Published 15 June 2014
Academic Editor Marcelo Paes de Barros
Copyright copy 2014 Zhengtang Qi et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
In this study we tested the hypothesis that NAC administration leads to reduced oxidative stress and thus to decreased expressionof autophagy markers in youngmice Our results reveal that NAC administration results in reduced muscle mRNA levels of severalautophagy markers including Beclin-1 Atg7 LC3 Atg9 and LAMP2 However NAC supplement fails to block the activationof skeletal muscle autophagy in response to fasting because fasting significantly increases the mRNA level of several autophagymarkers and LC3 lipidation We further examined the effects of NAC administration on mitochondrial antioxidant capacity infed and 24-hour fasted mice Our results clearly show that NAC administration depresses the expression of manganese superoxidedismutase (MnSOD) and TP53-induced glycolysis and apoptosis regulator (TIGAR) both of which play a predominant antioxidantrole in mitochondria by reducing ROS level In addition we found no beneficial effect of NAC supplement on muscle mass but itcan protect from muscle loss in response to fasting Collectively our findings indicate that ROS is required for skeletal muscleconstitutive autophagy rather than starvation-induced autophagy and that antioxidant NAC inhibits constitutive autophagy by theregulation of mitochondrial ROS production and antioxidant capacity
1 Introduction
Autophagy a conserved intracellular process carrying outdegradation of organelles and proteins has been linked toapoptosis and cell death serving both muscle growth andmitochondrial quality control [1 2] Autophagy-related genes(Atgs) are required for autophagic signaling pathway and theformation of autophagosome Beclin-1 (Atg6) is an importantcomponent of the Class III phosphoinositide 3-kinase (PI3K)complex which plays a positive role in the initial step ofautophagosome formation Beclin-1 recruits the other Atgproteins to the Class III PI3K complex and thus starts the nextstep of autophagy [3] Atg7 Atg9 and LC3 all play importantroles in elongation and formation of the autophagosome [4]LC3-II the lipidated form of LC3 was identified as an intrin-sic autophagosomal membrane marker in autophagy [5]Finally lysosomal-associated membrane protein-2 (LAMP-2) is responsible for the fusion of autophagosomes with
lysosomes LAMP-2 deletion causes the accumulation ofautophagosomes and a decrease in autolysosomes [6]
Autophagy can be carried out in stressed or unstressedstate Under conditions of stress such as nutrient depri-vation starvation and hypoxia autophagy allows cells tosurvive by releasing energy substrates via degradation ofcellular constituents In the stressed state of fasting themarker proteins for autophagy were increased in skele-tal muscle Further the autophagic response preferentiallyoccurred in fast-switch muscle during fasting this differencein autophagy regulation resulted from differential signal-ing transduction in slow- and fast-twitch skeletal muscle[7 8] In unstressed state autophagy contributes to theelimination of damaged organelles and proteins thereforeimpaired autophagy is implicated in the pathogenesis ofheart disease diabetes obesity neurodegenerative diseaseand cancer [9] Autophagy plays a dual role in cancer actingboth as a tumor suppressor by preventing the accumulation of
Hindawi Publishing CorporationOxidative Medicine and Cellular LongevityVolume 2014 Article ID 315896 10 pageshttpdxdoiorg1011552014315896
2 Oxidative Medicine and Cellular Longevity
damaged proteins and organelles and as a tumor promoterby promoting cell survival [10] In skeletal muscle autophagyis required to maintain muscle mass and mitochondrialfunction [11] impaired autophagy contributes to muscleatrophy in glycogen storage disease [12] Although autophagyis increased during muscle wasting and the induction ofautophagy precedes muscle loss [13] inhibiting autophagyis unable to protect skeletal muscle from atrophy duringdenervation and fasting instead it promotes greater mus-cle loss [14] These findings indicate that skeletal muscleautophagy at the basal level is both detrimental to causemus-cle protein degradation and beneficial to promote musclecell survival and mitochondrial quality control A completeunderstanding of the mechanisms responsible for skeletalmuscle autophagy in stressed and unstressed state remainsunclear
Regardless of being in the stressed state and at the basallevel autophagy induction is involved in reactive oxygenspecies (ROS) and cellular redox regulation Mitochondriaplay an important role in the regulation of autophagy bygenerating endogenous ROS on one hand and scavengingROS on the other hand [15] Thus maintaining mito-chondrial homeostasis by eliminating defective mitochon-dria is required for the maintenance of redox homeostasis[16] Under oxidative stress defective autophagy may alsoinduce abnormal mitochondrial accumulation In contrastto the increase in mitochondrial mass mitochondrial DNAwas observed to decrease with mitochondrial dysfunctionunder the oxidative conditions in cardiac myoblast cells[17] Compared with fragmented mitochondria elongatedmitochondria are more resistant to ROS-induced damageand mitochondrial autophagy (mitophagy) ROS-inducedmitochondrial damage may be an important upstream acti-vator of mitophagy [18] In many cell types cucurbitacin-induced autophagy was mediated by increased production ofmitochondrial-derived ROS and subsequently activation ofextracellular signal-regulated kinase (ERK) and c-jun NH2-terminal kinase (JNK) [19] These results suggest that theredox state is both the cause and the result of the interplaybetween autophagy and mitochondrial turnover Usuallyantioxidant supplement is used to treat some diseases byimproving redox state N-Acetylcysteine (NAC) is a precursorto cysteine which participates in the general antioxidantactivities of the body NAC can improve redox state andprotect mitochondrial function so as to prevent and reversethe progression of some diseases including neurodegen-erative diseases and diabetes [20ndash22] However previouswork has shown that a proper mitochondrial turnover isROS-dependent Antioxidant supplement inhibited mito-chondrial biogenesis in skeletal muscle by reducing ROS level[23 24]
In regard to the redox regulation in skeletal muscle pre-vious work has shown that NAC is an effective antioxidant fortreating muscular dystrophy (DMD) a severe degenerativemuscle disease [25] NAC prevents apoptosis and promotescell survival by activating ERK pathway so as to treat certaindegenerative diseases [26] In addition NAC has been usedas a tool for investigating the role of ROS in numerous bio-logical and pathological processes [22] In this study we used
antioxidant NAC to change the redox state in mice andobserved their autophagic responses to short-term fastingand the basal level of autophagy in skeletal muscle To inves-tigate the role of antioxidant administration in autophagywe tested the hypothesis that (1) the importance of ROS inconstitutive autophagy in skeletal muscle is different fromthat in the stress state of fasting and that (2) mitochondrialantioxidant capacity is attenuated by NAC and involved inautophagy regulation Our results support our hypothesis asNAC did repress numerous markers of autophagy and fail toprevent fasting-induced autophagy in skeletal muscle Thesenovel findings reveal that ROS is not required for skeletalmuscle autophagy resulting from fasting but important forskeletal muscle autophagy in unstressed state
2 Results
21 NAC Decreases Mitochondrial ROS Production but Failsto Protect against Fasting-Induced Mitochondrial Lipid Per-oxidation In our study mitochondrial ROS production wasdecreased in the NAC group (119875 lt 001) and increased inthe Fast group (119875 lt 005) compared with the Control group(Figure 1(c)) Mitochondrial GSH GSSG was elevated in theNAC group (119875 lt 005) and decreased in the Fast group(119875 lt 005) compared with the Control group (Figure 1(b))Importantly our results show thatNAC failed to protectmito-chondria against fasting-induced lipid peroxidation in theNAC + Fast group (119875 lt 005) However in the mice withoutNAC treatment fasting did not increasemitochondrialMDAlevel (Figure 1(a))
22 NAC Increases Glycogen Content in Skeletal Musclebut Fails to Protect against Fasting-Induced Glycogen Con-sumption Our results show that fasting reduced glycogencontent in skeletal muscle independent of NAC (119875 lt 001Figure 2(a)) In addition there was a significant increase(119875 lt 005) in skeletal muscle glycogen in the NAC groupcompared with Control (Figure 2(a)) Also the glycogencontent in the liver was significantly decreased (119875 lt 005)in the Fast group compared with Control but NAC treatmentbefore fasting prevented this decrease (Figure 2(b)) Similarlyquadriceps muscle weight was significantly decreased (119875 lt005) after fasting butNAC treatment prevented this decrease(Figure 2(c))
23 NAC Downregulates Autophagy Markers in Skeletal Mus-cle but Fails to Protect against Fasting-Induced UpregulationOur results show that mRNA expression of these Atgs inskeletal muscle was significantly decreased in the NAC groupcomparedwith Control HowevermRNA expression of thosegenes except Atg9 was increased significantly in the NAC+ Fast group compared with NAC group (Figure 3(a)) Weobserved that the activity of caspases 3 and 9 was significantlydecreased in the NAC group compared with Control andfasting significantly increased the activity of caspases 3 and9 in vehicle-treated mice (Figure 3(b)) Approximately redgastrocnemius muscle had a higher mRNA expression ofAtgs than white gastrocnemius muscle (Figure 3(c))
Oxidative Medicine and Cellular Longevity 3
002040608
112141618
Control Fast NAC NAC + Fast
Con
trol (
)
MDA
(a)
0
02
04
06
08
1
12
14
16
Control Fast NAC NAC + Fast
GSH GSSG
Con
trol (
)
lowast
lowast
(b)
0
05
1
15
2
25
Control Fast NAC
ROS level
NAC + Fast
Con
trol (
)
lowast
lowastlowast
(c)
Figure 1Oxidative stress biomarkers in the isolatedmitochondria of gastrocnemiusmuscleweremeasured after short-term fasting (a)MDAmethane dicarboxylic aldehyde (b) GSH glutathione (c) ROS production Values are means plusmn SE (fold differences) lowast119875 lt 005 lowastlowast119875 lt 001significantly different versus Control
119875 lt 005 significantly different versus NAC
24 NAC Represses TIGAR Expression but Fails to Protectagainst Fasting-Induced LC3 Lipidation In our study TIGARexpression was repressed significantly in the NAC groupcompared with Control group (119875 lt 001 Figures 4(a)and 4(c)) TIGARb expression was significantly increasedin the Fast group compared with Control group (119875 lt001 Figure 4(c)) and TIGARa expression was significantlyincreased in the NAC + Fast group compared with NACgroup (119875 lt 005 Figure 4(c)) In addition the ratio of LC3II to LC3 I (LC3 IILC3 I) was also quantified as a marker ofLC3 cleavage Fasting significantly increased LC3 lipidationindependent of NAC treatment (119875 lt 001 Figures 4(a) and4(b))
25 NAC Decreases MnSOD Expression and MitochondrialAntioxidant Capacity Our results show that mRNA expres-sion of CuZn-SOD MnSOD in skeletal muscle was signifi-cantly decreased in the NAC group compared with Control(119875 lt 001 Figures 5(a) and 5(b)) Similarly MnSOD activitywas significantly decreased in the NAC group compared withControl (119875 lt 001 Figure 5(c)) Also fasting had no effectson MnSOD expression and activity (Figure 5(c))
3 Discussion
These experiments provide novel and important informationregarding the effects of NAC administration on skeletalmuscle autophagy First our findings support the hypothesis
that NAC treatment dramatically decreases the expression ofnumerous markers of skeletal muscle autophagy at the basallevel rather than under the stress state of fasting Furtherour results reveal that NAC treatment is potential to lowermitochondrial antioxidant capacity through the inhibition ofthe antioxidant enzymes in mitochondria
31 NAC Administration Depresses Skeletal Muscle Consti-tutive Autophagy Significantly constitutive autophagy atthe basal level differs from stress-induced autophagy underconditions of nutrition deprivation In stress state enhancedautophagy favors cell survival by the degradation of cel-lular constituents and the recycling of macromolecules tosupport cellular metabolism Glycogen in skeletal muscleand liver can be utilized to produce energy or glucoseduring fasting so glycogen catabolism is also regarded asa peculiar autophagy outside lysosome [27] In response tostarvation the activation of autophagy is essential for themost efficient degradation of glycogen [28] In unstressedstate the basal level of autophagy in most tissues ensuresthe turnover of long-lived proteins and the elimination ofdysfunctional organelles Mammalian target of rapamycincomplex I (mTORC1) is regarded as the dominant regulatorof autophagy induction in skeletal muscle In young TSC1-(tuberous sclerosis complex 1-) deficient mice constitu-tive and starvation-induced autophagy was blocked at theinduction steps via mTORC1-mediated inhibition of ULK1(Unc-51-like kinase 1) [29] Recent studies suggested that
4 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
Control Fast NAC
Gly
coge
n co
nten
t (m
mol
g)
Skeletal muscle
NAC + Fast
lowast
lowastlowast
(a)
0
05
1
15
2
Control Fast NAC
Gly
coge
n co
nten
t (m
mol
g)
Liver
NAC + Fast
lowast
(b)
200
220
240
260
280
300
Left Right
Qua
dric
eps w
eigh
t (m
g)
ControlFast
NACNAC + Fast
lowastlowast
(c)
Figure 2Glycogen content and quadricepsmuscleweightweremeasured after short-term fasting (a)Glycogen content in quadricepsmuscle(b) Glycogen content in liver (c) Quadriceps muscle weight Values are means plusmn SE lowast119875 lt 005 lowastlowast119875 lt 001 significantly different versusControl
119875 lt 001 significantly different versus NAC
endogenous ROS promoted skeletal muscle autophagy at thebasal level and in response to acute nutrient starvation Thiseffect was mediated through regulation of autophagosomeinitiation and PI3KAKT inhibition [30] Using a potentialautophagy-targeting nutrient excessive muscle autophagy intype 2 diabetic rats was ameliorated through the downregu-lation of ROS signaling leading to improvement of glucosemetabolism reduction of oxidative stress and inhibition ofmitochondrial loss and dysfunction [31] ROS scavengerNACwas reported to diminish both endoplasmic reticulum (ER)stress and autophagy markers in skeletal muscle cells [32]Further NAC attenuated AMPK activation under starvationcondition and enhanced mTOR signaling in cervical cancercell The authors concluded that mitochondria-generatedROS induces autophagy mediated by the AMPK pathwayunder starvation conditions [33] Consistent with the previ-ous studies and our hypothesis our results show that NACsignificantly decreases mRNA expression of numerousmark-ers of skeletalmuscle autophagy and increases skeletalmuscleglycogenThese results indicate that the basal level of skeletalmuscle autophagy is ROS-dependent Constitutive autophagyin skeletal muscle is determined by redox state However ourresults also show that NAC fails to prevent fasting-inducedautophagy and glycogen reduction Specifically short-termfasting after NAC administration still increases the mRNAexpression of numerousmarkers of autophagy and the level of
LC3 lipidation These results indicate that ROS-independentautophagy occurs in skeletal muscle under starvation condi-tions In addition the activation of apoptosis also contributedto muscle loss relevant to oxidative stress Our data showthat NAC blocks fasting-induced activation of caspases 3and 9 and quadriceps muscle loss Collectively our resultsindicate that ROS is not required for fasting-induced auto-phagy but important for constitutive autophagy in skeletalmuscle
Additionally our results show that the expression ofmost Atgs is higher in red muscle than in white muscleThis difference in constitutive autophagy may result fromdifferential PGC-1120572 signaling and mitochondrial content inslow- and fast-twitch skeletal muscle Two similar works atleast partly support this postulation fiber type conversionfrom fast to slow fibers by PGC-1120572 activates lysosomaland autophagosomal biogenesis [34] as well as mitochon-drial biogenesis in skeletal muscle PGC-1120572 overexpressionincreases the expression of genes related to muscle repairand autophagy [35] Thus we suggest that the level ofconstitutive autophagy in slowmuscle fibers is higher than infast muscle fibers However PGC-1120572 overexpression duringaging protects from sarcopenia and metabolic disease byreducing apoptosis and autophagy [36] Further studiesneed to determine the relationship between mitochondrialbiogenesis and constitutive autophagy
Oxidative Medicine and Cellular Longevity 5
0
02
04
06
08
1
12
14
Beclin-1Atg6 Atg7 LC3Atg8 Atg9 LAMP-2
mRN
A le
vel (
Con
trol (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowastlowastlowast
lowastlowastlowast
(a)
0
05
1
15
2
25
Caspase-3 Caspase-8 Caspase-9
Enzy
me a
ctiv
ity (C
ontro
l (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowast
lowastlowast
(b)
0
05
1
15
2
25
3
mRN
A in
skel
etal
mus
cle(r
elat
ive e
xpre
ssio
n)
Young mice
Atg7 Atg9 Beclin-1 LAMP-2 LC3 p53
WGRG
lowastlowast
lowast
lowast lowast
(c)
Figure 3 Autophagy-related gene expressions were analyzed by real-time PCR in gastrocnemius muscle (a) mRNA level of Beclin-1 Atg7LC3 Atg9 and LAMP2 (b) Caspases 3 8 and 9 enzyme activity in gastrocnemius muscle (c) Autophagy-related gene expressions in redgastrocnemius (RG) versus white gastrocnemius (WG) Values are means plusmn SE (fold differences) lowast119875 lt 005 lowastlowast119875 lt 001 significantly dif-ferent versus Control
119875 lt 005 significantly different versus NAC
32 NAC Administration Lowers Mitochondrial AntioxidantCapacity In regard to the effects of antioxidant supple-ment on skeletal muscle autophagy we cannot ignore therole of mitochondria On one hand endogenous ROS ismainly generated by mitochondria on the other handmitochondrial antioxidant enzyme is able to scavenge ROSand reduce oxidative stress Recent works have suggestedthat mitochondrial-derived ROS induces autophagy [30 33]thus mitochondrial antioxidant capacity may be responsiblefor the activation of autophagy In the previous studiesNAC has been considered as a mitochondrial enhancerfor improving redox status and mitochondrial dysfunctionand reversing the progression of some diseases [37 38]Consistent with the previous results our data show that NACreduces mitochondrial ROS level and increases the ratio ofGSH GSSG suggesting that reduced ROS production andelevated GSH GSSG ratio in mitochondria are involved inthe inhibition of skeletal muscle constitutive autophagy Fur-ther NAC in part prevents fasting-induced oxidative stress inmitochondria but not autophagic responses indicating thatskeletal muscle autophagy under starvation is independentof mitochondrial-derived ROS In contrast to our results
in fasted mice lower nutrient supply or calorie restrictionleads to a lower production of mitochondrial ROS in skeletalmuscle [39] because the lower mitochondrial membranepotential favors reducing mitochondrial ROS production[40] In skeletal muscle of both rodents and humans high-fatdiet increases H
2O2emission of mitochondria [41] However
in response to acute nutrient deprivation mitochondrialROS levels significantly increase in skeletal muscle cells [30]These findings strongly indicate that ROS production inmitochondria is nutrient-sensitizing [42] It is noteworthythat chronic mild nutrient restriction differs from acutenutrient deprivation in the regulation of mitochondrial ROSproduction Thus the differences in constitutive and acutestress-induced autophagy may result from differential ROSlevels in skeletal muscle
Manganese superoxide dismutase (MnSOD) is a vitalantioxidant enzyme in the matrix of mitochondria by scav-enging ROS MnSOD knockdown led to chronic oxidativestress increased autophagy and mitochondrial biogenesiswithin the distal nephrons [43] In brain MnSOD knock-down increased oxidative stress and instead suppressedthe levels of autophagy stimulators [44] To our surprise
6 Oxidative Medicine and Cellular Longevity
LC3-I
TIGAR
LC3-II
Con
trol
Fast
NAC
NAC
+ F
ast
120573-Actin
(a)
0
05
1
15
2
Control Fast NAC
LC3
lipid
atio
n (C
ontro
l (
))
LC3-IILC3-I
NAC + Fast
lowastlowast
(b)
0
5
10
15
20
25
30
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
TIGARaTIGARb
lowastlowastlowastlowast
lowastlowast
(c)
Figure 4 LC3 lipidation and expressions of TIGAR were measured in skeletal muscle after short-term fasting (a) Representative westernblots of LC3 TIGAR (b) LC3-IILC3-I ratio (c) mRNA expression of TIGAR Values are means plusmn SE lowastlowast119875 lt 001 significantly differentversus Control
119875 lt 005 119875 lt 001 significantly different versus NAC
our results show that NAC decreases MnSODmRNA expres-sion and its enzyme activity CuZn-SOD expression is alsorepressed Fasting fails to rescue the decline in MnSODexpression Our data suggest that although NAC adminis-tration reduces ROS level and increases GSH GSSG exoge-nous antioxidant supplementmay lower inherent antioxidantcapacity in mitochondria ROS can lead to MnSOD upreg-ulation through ERK2 translocation into the nucleus anddissociation of p53 from its inhibitory protein mouse doubleminute 2 (MDM2) [45] Similarly heat stress producedROS in fibroblasts NAC blocked the increase in MnSODlevels by heat stress through reducing ROS production [46]Therefore a proper level of ROS is required for MnSODexpression Likewise we observe that TIGAR is repressedin mRNA and protein level by NAC Although TIGAR isnot an antioxidant enzyme that eliminates ROS directlyTIGAR plays an important antioxidant role in the regu-lation of autophagy [47] TIGAR inhibits glycolysis andpromotes respiration In TIGAR-expressing cells ROS levelwas redu-ced whereas glutathione was elevated [48] FurtherTIGAR inhibited autophagy by suppressing ROS level [47]TIGAR knockout increased mitophagy and ROS productionin myocardium followed by Bnip3 activation which is an
initiator of mitophagy Further NAC reversed the activationof Bnip3 and mitophagy [49] Under hypoxia mitochondriallocalization of TIGAR can control mitochondrial ROS levelsand protect from cell death [50] Therefore TIGAR is apotential antioxidant gene formitochondria Collectively ourresults indicate that NAC treatment with young mice atten-uates the inherent antioxidant capacity in skeletal musclesuggesting that ROS is also required for the maintenance ofmitochondrial antioxidant capacity
In summary our study provides the first evidencethat NAC decreases skeletal muscle constitutive autophagybut does not protect skeletal muscle from fasting-inducedautophagy in young mice In addition we also showed thatNAC attenuates the inherent antioxidant capacity in skeletalmuscle by depressing the expression of MnSOD and TIGARWe interpret these findings as an indication that ROS isrequired for the maintenance of both constitutive autophagyand antioxidant capacity in skeletal muscle Although NACsupplement is regarded as an effective intervention forimprovingmitochondrial function it should be reconsideredcarefully for young and healthy individuals when NACor the other antioxidants are used to promote musclegrowth
Oxidative Medicine and Cellular Longevity 7
0
20
40
60
80
100
120
140
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
SOD1 (CuZn-SOD)
lowastlowast
(a)
mRN
A le
vel (
vers
us 120573
-act
in)
0
50
100
150
200
250
Control Fast NAC NAC + Fast
SOD2 (MnSOD)
lowastlowast
(b)
0
02
04
06
08
1
12
14
Control Fast NAC NAC + Fast
Con
trol (
)
MnSOD activity
lowastlowast
(c)
Figure 5 Antioxidant capacity wasmeasured in gastrocnemiusmuscle after short-term fasting (a) mRNA expression of SOD1 (CuZn-SOD)(b) mRNA expression of SOD2 (MnSOD) (c) MnSOD activity in the isolated mitochondria of gastrocnemius muscle Values are means plusmnSE lowastlowast119875 lt 001 significantly different versus Control
119875 lt 005 significantly different versus NAC
4 Materials and Methods
41 Animals Male ICRCD-1 mice were purchased fromSino-British SipprBK Lab Animal Ltd CO (ShanghaiChina) at four to five weeks of age (Young 228 plusmn 12 g) Ani-mals were housed in a temperature-controlled environment(23 plusmn 2∘C) with 12 12 h light-dark cycles where they wereprovided diets and water ad libitum Mice were randomlyassigned to one of four experimental groups (1) Controlno drug treatment (Control 119899 = 8) (2) Fast no drugtreatment (Fast 119899 = 8) (3) Control treated with NAC (NAC119899 = 10) and (4) Fast treated with NAC (NAC + Fast119899 = 10) All experiments were performed in accordance withthe guidelines for the use of laboratory animals publishedby China Ministry of Health (no 55 order ordained on 25Jan 1998) All experimental procedureswere approved by theExperimental Animal Care andUse Committee at East ChinaNormal University (ECNU 2006-05)
42 Drugs and Short-TermFasting TheantioxidantN-acetyl-cysteine (NAC Sigma) was dissolved in DMSO The miceassigned to NAC groups received intraperitoneal injection ofNAC for 3weeks (100mgkg bodyweight2 day) As a controlthe vehicle (10 DMSO in saline) was injected to the controlmice For studies on the effects of food deprivation the mice
assigned to Fast groups were deprived of food for 24 hr beforesacrifice These mice had free access to drinking water
43 Mitochondrial Isolation At the end of the experimentalperiod animals in each group were acutely anesthetized withpentobarbital sodium Quadriceps gastrocnemius muscleand liver were removed completely and weighed For mito-chondrial isolation fresh gastrocnemius muscle was rinsedwith PBS and put into ice-cold isolation buffer (0075Msucrose 0225M sorbitol 1mM EGTA 01 fatty acid-freebovine serum albumin (BSA) and 10mM Tris-HCl pH 74)Tissues were sheared carefully to mince rinsed to get ridof residual blood and then homogenized in 1mL isolationbuffer per 100mg tissue The homogenate was centrifugedat 1000 g for 5min at 4∘C using a Beckman centrifuge(Avanti J-26XP) the resulting supernatant was decanted andsaved The pellet was washed once with isolation buffer Thesupernatant was combined and centrifuged at 9000 g for10min at 4∘C The mitochondrial pellet was washed andcentrifuged twice at 15000 g for 2min at 4∘C with isolationbuffer [51] Mitochondrial protein content was assayed usingBSA as a standard according to Bradford
44Mitochondrial ROSGeneration and Antioxidant CapacityThe dichlorofluorescin diacetate (DCF-DA) was used as
8 Oxidative Medicine and Cellular Longevity
a probe to detect mitochondrial ROS generation The assaywas performed on freshly isolated mitochondria Samples(30 120583g proteins) were incubated with 10 120583L of DCF-DA(10 120583molI) for 3 h at 37∘C The fluorescent product formedwas quantified by spectrofluorometer at the 485525 nmChanges in fluorescence were expressed as arbitrary unitTo assay mitochondrial lipid peroxidation MDA levels weremeasured using an assay kit according to the manufac-turerrsquos protocol (Nanjing Jiancheng Biotech Nanjing China)Briefly isolated mitochondria were incubated at 37∘C for 3 htreated with buffer and centrifuged at 10000timesg for 15minThe supernatants were incubated with thiobarbituric acid(TBA) and the absorbance of the supernatants wasmeasuredusing a spectrophotometer at a wavelength of 535 nm Alsothe GSHGSSG ratio and MnSOD activity were measuredusing an assay kit (Nanjing Jiancheng Biotech NanjingChina) according to themanufacturerrsquos protocol respectively
45 Caspases Activity Caspases 3 8 and 9 activity wasdetermined with an assay kit (Beyotime China) accordingto the manufacturerrsquos instructions Under the caspase-3acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) canbe changed into the yellow product p-nitroaniline (pNA)Under the caspase-8 acetyl-Ile-Glu-Thr-Asp p-nitroanilide(Ac-IETD-pNA) can be changed into the yellow prod-uct pNA Under the caspase-9 acetyl-Leu-Glu-His-Asp p-nitroanilide (Ac-LEHD-pNA) can be changed into the yellowproduct pNA Finally pNA content was measured usingthe spectrophotometer at an absorbance of 405 nm Proteincontent was assayed using BSA as a standard according toBradford
46 Glycogen Content Glycogen content was determinedwith an assay kit (Nanjing Jianchen Biotech China) accord-ing to the manufacturerrsquos instructions
47 Real-Time PCR Total RNA was prepared from sim100mgof frozen tissues using TRIzol (Invitrogen Chromos Singa-pore) and purified according to the instructions includedDouble-stranded cDNA was synthesized from sim1 120583g of totalRNA using ReverTra Ace qPCR RT kit (TOYOBO OsakaJapan) Real-time PCR reactions were set up using the SYBR-Green PCR kit (TOYOBO Osaka Japan) and were cycled inStepOne Real-Time PCR System (Applied Biosystems CAUSA) as previously described [52] The abundance of targetmRNA was normalized to that of 120573-actin Primer pairs weredesigned based on GenBank reference sequences and listedin Table S1 available online in Supplementary Material athttpdxdoiorg1011552014315896
48 Western Blotting The isolated muscle tissue (sim200mg)was weighed and cut into pieces at 4∘C and 2mL (10times dryweight of isolated muscle fibers) of ice-cold buffer (50mMTris HCl 150mMNaCl 1mM EDTA 02mM PMSF 1 NP-40 pH 74 cocktail) was added The tissue pieces were thenhomogenized after which homogenates were centrifuged for10min at 8000 g and 4∘C The total protein content of thesupernatant was quantified using bicinchoninic acid reagents
and BSA standards (Shanghai Sangon Shanghai China)Theprotein samples were immunoblotted as previously described[52] Visualization of reaction bands was performed by 331015840-diaminobenzidine staining (Shanghai Sangon China) andscanned densitometrically 120573-Actin was used to standardizefor the amount of protein loaded
49 Statistical Analysis Data are reported as mean plusmn SEMStatistical differences between treatments were determinedusing ANOVA For all tests the significance level was set at119875 lt 005
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from the National NaturalScience Foundation of China (Grant nos 31300977 and31171142) This work was also supported by the Key Labora-tory Construction Project of Adolescent Health Assessmentand Exercise Intervention of Ministry of Education EastChinaNormalUniversity (no 40500-541235-14203004)Theauthors also thank postgraduate students at Professor Dingrsquoslab for the technical support on animal care biochemicalassays PCR and western blot
References
[1] M Sandri ldquoAutophagy in skeletal musclerdquo FEBS Letters vol584 no 7 pp 1411ndash1416 2010
[2] G Twig B Hyde and O S Shirihai ldquoMitochondrial fusionfission and autophagy as a quality control axis the bioenergeticviewrdquo Biochimica et Biophysica Acta Bioenergetics vol 1777 no9 pp 1092ndash1097 2008
[3] Q Sun W Fan K Chen X Ding S Chen and Q ZhongldquoIdentification of Barkor as a mammalian autophagy-specificfactor for Beclin 1 and class III phosphatidylinositol 3-kinaserdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 49 pp 19211ndash19216 2008
[4] M C Maiuri E Zalckvar A Kimchi and G Kroemer ldquoSelf-eating and self-killing crosstalk between autophagy and apop-tosisrdquo Nature Reviews Molecular Cell Biology vol 8 no 9 pp741ndash752 2007
[5] T Ueno W Sato Y Horie et al ldquoLoss of Pten a tumor suppres-sor causes the strong inhibition of autophagy without affectingLC3 lipidationrdquo Autophagy vol 4 no 5 pp 692ndash700 2008
[6] F FortunatoH Burgers F Bergmann et al ldquoImpaired autolyso-some formation correlates with lamp-2 depletion role of apop-tosis autophagy and necrosis in pancreatitisrdquoGastroenterologyvol 137 no 1 pp 350ndash360 2009
[7] F A Graca D A Goncalves W A Silveira et al ldquoEpinephrinedepletion exacerbates the fasting-induced protein breakdown infast-twitch skeletal musclesrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 305 pp E1483ndashE14942013
[8] T Ogata Y Oishi M Higuchi and I Muraoka ldquoFasting-relatedautophagic response in slow- and fast-twitch skeletal musclerdquo
Oxidative Medicine and Cellular Longevity 9
Biochemical andBiophysical ResearchCommunications vol 394no 1 pp 136ndash140 2010
[9] A Doria M Gatto and L Punzi ldquoAutophagy in human healthand diseaserdquo New England Journal of Medicine vol 368 no 19p 1845 2013
[10] S Fulda ldquoAutophagy and cell deathrdquo Autophagy vol 8 no 8pp 1250ndash1251 2012
[11] E Masiero L Agatea C Mammucari et al ldquoAutophagy isrequired to maintain muscle massrdquo Cell Metabolism vol 10 no6 pp 507ndash515 2009
[12] A C Nascimbeni M Fanin E Masiero C Angelini and MSandri ldquoImpaired autophagy contributes to muscle atrophy inglycogen storage disease type II patientsrdquo Autophagy vol 8 no11 pp 1697ndash1700 2012
[13] M Sandri ldquoProtein breakdown in muscle wasting role ofautophagy-lysosome and ubiquitin-proteasomerdquo InternationalJournal of Biochemistry and Cell Biology vol 45 pp 2121ndash21292013
[14] E Masiero and M Sandri ldquoAutophagy inhibition inducesatrophy and myopathy in adult skeletal musclesrdquo Autophagyvol 6 no 2 pp 307ndash309 2010
[15] R Scherz-Shouval and Z Elazar ldquoRegulation of autophagy byROS physiology and pathologyrdquoTrends in Biochemical Sciencesvol 36 no 1 pp 30ndash38 2011
[16] C R Morales Z Pedrozo S Lavandero and J A Hill ldquoOxi-dative stress and autophagy in cardiovascular homeostasisrdquoAntioxidants amp Redox Signaling vol 20 pp 507ndash518 2014
[17] C Luo Y Li H Wang et al ldquoMitochondrial accumulationunder oxidative stress is due to defects in autophagyrdquo Journalof Cellular Biochemistry vol 114 no 1 pp 212ndash219 2013
[18] Y Wang Y Nartiss B Steipe G A McQuibban and PK Kim ldquoROS-induced mitochondrial depolarization initi-ates PARK2PARKIN-dependent mitochondrial degradationby autophagyrdquo Autophagy vol 8 no 10 pp 1462ndash1476 2012
[19] T Zhang Y Li K A Park et al ldquoCucurbitacin induces auto-phagy through mitochondrial ROS production which counter-acts to limit caspase-dependent apoptrdquo Autophagy vol 8 no 4pp 559ndash576 2012
[20] R Sandhir A Sood A Mehrotra and S S Kamboj ldquoN-acetyl-cysteine reverses mitochondrial dysfunctions and behavioralabnormalities in 3-nitropropionic acid-induced Huntingtonsdiseaserdquo Neurodegenerative Diseases vol 9 no 3 pp 145ndash1572012
[21] S S Kamboj and R Sandhir ldquoProtective effect of N-acetyl-cysteine supplementation onmitochondrial oxidative stress andmitochondrial enzymes in cerebral cortex of streptozotocin-treated diabetic ratsrdquoMitochondrion vol 11 no 1 pp 214ndash2222011
[22] M ZafarullahW Q Li J Sylvester andM Ahmad ldquoMolecularmechanisms of N-acetylcysteine actionsrdquo Cellular and Molecu-lar Life Sciences vol 60 no 1 pp 6ndash20 2003
[23] N A Strobel J M Peake A Matsumoto S A Marsh J SCoombes and G D Wadley ldquoAntioxidant supplementationreduces skeletal muscle mitochondrial biogenesisrdquo Medicineand Science in Sports and Exercise vol 43 no 6 pp 1017ndash10242011
[24] M-C Gomez-Cabrera E Domenech M Romagnoli et alldquoOral administration of vitamin C decreases muscle mitochon-drial biogenesis and hampers training-induced adaptationsin endurance performancerdquo The American Journal of ClinicalNutrition vol 87 no 1 pp 142ndash149 2008
[25] N P Whitehead C Pham O L Gervasio and D G Allen ldquoN-Acetylcysteine ameliorates skeletal muscle pathophysiology inmdx micerdquo Journal of Physiology vol 586 no 7 pp 2003ndash20142008
[26] C-H Lin S-C Kuo L-J Huang and P-W Gean ldquoNeuropro-tective effect ofN-acetylcysteine on neuronal apoptosis inducedby a synthetic gingerdione compound involvement of ERK andp38 phosphorylationrdquo Journal of Neuroscience Research vol 84no 7 pp 1485ndash1494 2006
[27] O BKotoulas S AKalamidas andD J Kondomerkos ldquoGlyco-gen autophagy in glucose homeostasisrdquo Pathology Research andPractice vol 202 no 9 pp 631ndash638 2006
[28] J Zirin J Nieuwenhuis and N Perrimon ldquoRole of autophagyin glycogen breakdown and its relevance to chloroquinemyopa-thyrdquo PLoS Biology vol 11 no 11 Article ID e1001708 2013
[29] P Castets S Lin N Rion et al ldquoSustained activation ofmTORC1 in skeletalmuscle inhibits constitutive and starvation-induced autophagy and causes a severe late-onset myopathyrdquoCell Metabolism vol 17 no 5 pp 731ndash744 2013
[30] M Rahman M Mofarrahi A S Kristof B Nkengfac S Hareland S N Hussain ldquoReactive oxygen species regulation ofautophagy in skeletal musclesrdquoAntioxidants amp Redox Signalingvol 20 no 3 pp 443ndash459 2014
[31] J Yan Z Feng J Liu et al ldquoEnhanced autophagy plays a cardi-nal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats ameliorating effects of (-)-epigallocatechin-3-gallaterdquo Journal of Nutritional Biochemistry vol 23 no 7 pp716ndash724 2012
[32] LV Yuzefovych S IMusiyenkoG LWilson and L I RachekldquoMitochondrial DNA damage and dysfunction and oxidativestress are associated with endoplasmic reticulum stress proteindegradation and apoptosis in high fat diet-induced InsulinresistancemicerdquoPLoSONE vol 8 no 1 Article ID e54059 2013
[33] L Li Y Chen and S B Gibson ldquoStarvation-induced autophagyis regulated bymitochondrial reactive oxygen species leading toAMPK activationrdquo Cellular Signalling vol 25 no 1 pp 50ndash652013
[34] S Takikita C Schreiner R Baum et al ldquoFiber type conversionby PGC-1120572 activates lysosomal and autophagosomal biogenesisin both unaffected and pompe skeletal musclerdquo PLoS ONE vol5 no 12 Article ID e15239 2010
[35] K Hollinger D Gardan-Salmon C Santana D Rice E Snellaand J T Selsby ldquoRescue of dystrophic skeletal muscle byPGC-1120572 involves restored expression of dystrophin-associatedprotein complex components and satellite cell signalingrdquo TheAmerican Journal of Physiology Regulatory Integrative andComparative Physiology vol 305 no 1 pp R13ndashR23 2013
[36] T Wenz S G Rossi R L Rotundo B M Spiegelman and CT Moraes ldquoIncreased muscle PGC-1120572 expression protects fromsarcopenia and metabolic disease during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 48 pp 20405ndash20410 2009
[37] I Grattagliano P Portincasa T Cocco et al ldquoEffect of dietaryrestriction and N-acetylcysteine supplementation on intestinalmucosa and liver mitochondrial redox status and function inaged ratsrdquo Experimental Gerontology vol 39 no 9 pp 1323ndash1332 2004
[38] S Kumar and S L Sitasawad ldquoN-acetylcysteine prevents glu-coseglucose oxidase-induced oxidative stress mitochondrialdamage and apoptosis in H9c2 cellsrdquo Life Sciences vol 84 no11-12 pp 328ndash336 2009
10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Oxidative Medicine and Cellular Longevity
damaged proteins and organelles and as a tumor promoterby promoting cell survival [10] In skeletal muscle autophagyis required to maintain muscle mass and mitochondrialfunction [11] impaired autophagy contributes to muscleatrophy in glycogen storage disease [12] Although autophagyis increased during muscle wasting and the induction ofautophagy precedes muscle loss [13] inhibiting autophagyis unable to protect skeletal muscle from atrophy duringdenervation and fasting instead it promotes greater mus-cle loss [14] These findings indicate that skeletal muscleautophagy at the basal level is both detrimental to causemus-cle protein degradation and beneficial to promote musclecell survival and mitochondrial quality control A completeunderstanding of the mechanisms responsible for skeletalmuscle autophagy in stressed and unstressed state remainsunclear
Regardless of being in the stressed state and at the basallevel autophagy induction is involved in reactive oxygenspecies (ROS) and cellular redox regulation Mitochondriaplay an important role in the regulation of autophagy bygenerating endogenous ROS on one hand and scavengingROS on the other hand [15] Thus maintaining mito-chondrial homeostasis by eliminating defective mitochon-dria is required for the maintenance of redox homeostasis[16] Under oxidative stress defective autophagy may alsoinduce abnormal mitochondrial accumulation In contrastto the increase in mitochondrial mass mitochondrial DNAwas observed to decrease with mitochondrial dysfunctionunder the oxidative conditions in cardiac myoblast cells[17] Compared with fragmented mitochondria elongatedmitochondria are more resistant to ROS-induced damageand mitochondrial autophagy (mitophagy) ROS-inducedmitochondrial damage may be an important upstream acti-vator of mitophagy [18] In many cell types cucurbitacin-induced autophagy was mediated by increased production ofmitochondrial-derived ROS and subsequently activation ofextracellular signal-regulated kinase (ERK) and c-jun NH2-terminal kinase (JNK) [19] These results suggest that theredox state is both the cause and the result of the interplaybetween autophagy and mitochondrial turnover Usuallyantioxidant supplement is used to treat some diseases byimproving redox state N-Acetylcysteine (NAC) is a precursorto cysteine which participates in the general antioxidantactivities of the body NAC can improve redox state andprotect mitochondrial function so as to prevent and reversethe progression of some diseases including neurodegen-erative diseases and diabetes [20ndash22] However previouswork has shown that a proper mitochondrial turnover isROS-dependent Antioxidant supplement inhibited mito-chondrial biogenesis in skeletal muscle by reducing ROS level[23 24]
In regard to the redox regulation in skeletal muscle pre-vious work has shown that NAC is an effective antioxidant fortreating muscular dystrophy (DMD) a severe degenerativemuscle disease [25] NAC prevents apoptosis and promotescell survival by activating ERK pathway so as to treat certaindegenerative diseases [26] In addition NAC has been usedas a tool for investigating the role of ROS in numerous bio-logical and pathological processes [22] In this study we used
antioxidant NAC to change the redox state in mice andobserved their autophagic responses to short-term fastingand the basal level of autophagy in skeletal muscle To inves-tigate the role of antioxidant administration in autophagywe tested the hypothesis that (1) the importance of ROS inconstitutive autophagy in skeletal muscle is different fromthat in the stress state of fasting and that (2) mitochondrialantioxidant capacity is attenuated by NAC and involved inautophagy regulation Our results support our hypothesis asNAC did repress numerous markers of autophagy and fail toprevent fasting-induced autophagy in skeletal muscle Thesenovel findings reveal that ROS is not required for skeletalmuscle autophagy resulting from fasting but important forskeletal muscle autophagy in unstressed state
2 Results
21 NAC Decreases Mitochondrial ROS Production but Failsto Protect against Fasting-Induced Mitochondrial Lipid Per-oxidation In our study mitochondrial ROS production wasdecreased in the NAC group (119875 lt 001) and increased inthe Fast group (119875 lt 005) compared with the Control group(Figure 1(c)) Mitochondrial GSH GSSG was elevated in theNAC group (119875 lt 005) and decreased in the Fast group(119875 lt 005) compared with the Control group (Figure 1(b))Importantly our results show thatNAC failed to protectmito-chondria against fasting-induced lipid peroxidation in theNAC + Fast group (119875 lt 005) However in the mice withoutNAC treatment fasting did not increasemitochondrialMDAlevel (Figure 1(a))
22 NAC Increases Glycogen Content in Skeletal Musclebut Fails to Protect against Fasting-Induced Glycogen Con-sumption Our results show that fasting reduced glycogencontent in skeletal muscle independent of NAC (119875 lt 001Figure 2(a)) In addition there was a significant increase(119875 lt 005) in skeletal muscle glycogen in the NAC groupcompared with Control (Figure 2(a)) Also the glycogencontent in the liver was significantly decreased (119875 lt 005)in the Fast group compared with Control but NAC treatmentbefore fasting prevented this decrease (Figure 2(b)) Similarlyquadriceps muscle weight was significantly decreased (119875 lt005) after fasting butNAC treatment prevented this decrease(Figure 2(c))
23 NAC Downregulates Autophagy Markers in Skeletal Mus-cle but Fails to Protect against Fasting-Induced UpregulationOur results show that mRNA expression of these Atgs inskeletal muscle was significantly decreased in the NAC groupcomparedwith Control HowevermRNA expression of thosegenes except Atg9 was increased significantly in the NAC+ Fast group compared with NAC group (Figure 3(a)) Weobserved that the activity of caspases 3 and 9 was significantlydecreased in the NAC group compared with Control andfasting significantly increased the activity of caspases 3 and9 in vehicle-treated mice (Figure 3(b)) Approximately redgastrocnemius muscle had a higher mRNA expression ofAtgs than white gastrocnemius muscle (Figure 3(c))
Oxidative Medicine and Cellular Longevity 3
002040608
112141618
Control Fast NAC NAC + Fast
Con
trol (
)
MDA
(a)
0
02
04
06
08
1
12
14
16
Control Fast NAC NAC + Fast
GSH GSSG
Con
trol (
)
lowast
lowast
(b)
0
05
1
15
2
25
Control Fast NAC
ROS level
NAC + Fast
Con
trol (
)
lowast
lowastlowast
(c)
Figure 1Oxidative stress biomarkers in the isolatedmitochondria of gastrocnemiusmuscleweremeasured after short-term fasting (a)MDAmethane dicarboxylic aldehyde (b) GSH glutathione (c) ROS production Values are means plusmn SE (fold differences) lowast119875 lt 005 lowastlowast119875 lt 001significantly different versus Control
119875 lt 005 significantly different versus NAC
24 NAC Represses TIGAR Expression but Fails to Protectagainst Fasting-Induced LC3 Lipidation In our study TIGARexpression was repressed significantly in the NAC groupcompared with Control group (119875 lt 001 Figures 4(a)and 4(c)) TIGARb expression was significantly increasedin the Fast group compared with Control group (119875 lt001 Figure 4(c)) and TIGARa expression was significantlyincreased in the NAC + Fast group compared with NACgroup (119875 lt 005 Figure 4(c)) In addition the ratio of LC3II to LC3 I (LC3 IILC3 I) was also quantified as a marker ofLC3 cleavage Fasting significantly increased LC3 lipidationindependent of NAC treatment (119875 lt 001 Figures 4(a) and4(b))
25 NAC Decreases MnSOD Expression and MitochondrialAntioxidant Capacity Our results show that mRNA expres-sion of CuZn-SOD MnSOD in skeletal muscle was signifi-cantly decreased in the NAC group compared with Control(119875 lt 001 Figures 5(a) and 5(b)) Similarly MnSOD activitywas significantly decreased in the NAC group compared withControl (119875 lt 001 Figure 5(c)) Also fasting had no effectson MnSOD expression and activity (Figure 5(c))
3 Discussion
These experiments provide novel and important informationregarding the effects of NAC administration on skeletalmuscle autophagy First our findings support the hypothesis
that NAC treatment dramatically decreases the expression ofnumerous markers of skeletal muscle autophagy at the basallevel rather than under the stress state of fasting Furtherour results reveal that NAC treatment is potential to lowermitochondrial antioxidant capacity through the inhibition ofthe antioxidant enzymes in mitochondria
31 NAC Administration Depresses Skeletal Muscle Consti-tutive Autophagy Significantly constitutive autophagy atthe basal level differs from stress-induced autophagy underconditions of nutrition deprivation In stress state enhancedautophagy favors cell survival by the degradation of cel-lular constituents and the recycling of macromolecules tosupport cellular metabolism Glycogen in skeletal muscleand liver can be utilized to produce energy or glucoseduring fasting so glycogen catabolism is also regarded asa peculiar autophagy outside lysosome [27] In response tostarvation the activation of autophagy is essential for themost efficient degradation of glycogen [28] In unstressedstate the basal level of autophagy in most tissues ensuresthe turnover of long-lived proteins and the elimination ofdysfunctional organelles Mammalian target of rapamycincomplex I (mTORC1) is regarded as the dominant regulatorof autophagy induction in skeletal muscle In young TSC1-(tuberous sclerosis complex 1-) deficient mice constitu-tive and starvation-induced autophagy was blocked at theinduction steps via mTORC1-mediated inhibition of ULK1(Unc-51-like kinase 1) [29] Recent studies suggested that
4 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
Control Fast NAC
Gly
coge
n co
nten
t (m
mol
g)
Skeletal muscle
NAC + Fast
lowast
lowastlowast
(a)
0
05
1
15
2
Control Fast NAC
Gly
coge
n co
nten
t (m
mol
g)
Liver
NAC + Fast
lowast
(b)
200
220
240
260
280
300
Left Right
Qua
dric
eps w
eigh
t (m
g)
ControlFast
NACNAC + Fast
lowastlowast
(c)
Figure 2Glycogen content and quadricepsmuscleweightweremeasured after short-term fasting (a)Glycogen content in quadricepsmuscle(b) Glycogen content in liver (c) Quadriceps muscle weight Values are means plusmn SE lowast119875 lt 005 lowastlowast119875 lt 001 significantly different versusControl
119875 lt 001 significantly different versus NAC
endogenous ROS promoted skeletal muscle autophagy at thebasal level and in response to acute nutrient starvation Thiseffect was mediated through regulation of autophagosomeinitiation and PI3KAKT inhibition [30] Using a potentialautophagy-targeting nutrient excessive muscle autophagy intype 2 diabetic rats was ameliorated through the downregu-lation of ROS signaling leading to improvement of glucosemetabolism reduction of oxidative stress and inhibition ofmitochondrial loss and dysfunction [31] ROS scavengerNACwas reported to diminish both endoplasmic reticulum (ER)stress and autophagy markers in skeletal muscle cells [32]Further NAC attenuated AMPK activation under starvationcondition and enhanced mTOR signaling in cervical cancercell The authors concluded that mitochondria-generatedROS induces autophagy mediated by the AMPK pathwayunder starvation conditions [33] Consistent with the previ-ous studies and our hypothesis our results show that NACsignificantly decreases mRNA expression of numerousmark-ers of skeletalmuscle autophagy and increases skeletalmuscleglycogenThese results indicate that the basal level of skeletalmuscle autophagy is ROS-dependent Constitutive autophagyin skeletal muscle is determined by redox state However ourresults also show that NAC fails to prevent fasting-inducedautophagy and glycogen reduction Specifically short-termfasting after NAC administration still increases the mRNAexpression of numerousmarkers of autophagy and the level of
LC3 lipidation These results indicate that ROS-independentautophagy occurs in skeletal muscle under starvation condi-tions In addition the activation of apoptosis also contributedto muscle loss relevant to oxidative stress Our data showthat NAC blocks fasting-induced activation of caspases 3and 9 and quadriceps muscle loss Collectively our resultsindicate that ROS is not required for fasting-induced auto-phagy but important for constitutive autophagy in skeletalmuscle
Additionally our results show that the expression ofmost Atgs is higher in red muscle than in white muscleThis difference in constitutive autophagy may result fromdifferential PGC-1120572 signaling and mitochondrial content inslow- and fast-twitch skeletal muscle Two similar works atleast partly support this postulation fiber type conversionfrom fast to slow fibers by PGC-1120572 activates lysosomaland autophagosomal biogenesis [34] as well as mitochon-drial biogenesis in skeletal muscle PGC-1120572 overexpressionincreases the expression of genes related to muscle repairand autophagy [35] Thus we suggest that the level ofconstitutive autophagy in slowmuscle fibers is higher than infast muscle fibers However PGC-1120572 overexpression duringaging protects from sarcopenia and metabolic disease byreducing apoptosis and autophagy [36] Further studiesneed to determine the relationship between mitochondrialbiogenesis and constitutive autophagy
Oxidative Medicine and Cellular Longevity 5
0
02
04
06
08
1
12
14
Beclin-1Atg6 Atg7 LC3Atg8 Atg9 LAMP-2
mRN
A le
vel (
Con
trol (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowastlowastlowast
lowastlowastlowast
(a)
0
05
1
15
2
25
Caspase-3 Caspase-8 Caspase-9
Enzy
me a
ctiv
ity (C
ontro
l (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowast
lowastlowast
(b)
0
05
1
15
2
25
3
mRN
A in
skel
etal
mus
cle(r
elat
ive e
xpre
ssio
n)
Young mice
Atg7 Atg9 Beclin-1 LAMP-2 LC3 p53
WGRG
lowastlowast
lowast
lowast lowast
(c)
Figure 3 Autophagy-related gene expressions were analyzed by real-time PCR in gastrocnemius muscle (a) mRNA level of Beclin-1 Atg7LC3 Atg9 and LAMP2 (b) Caspases 3 8 and 9 enzyme activity in gastrocnemius muscle (c) Autophagy-related gene expressions in redgastrocnemius (RG) versus white gastrocnemius (WG) Values are means plusmn SE (fold differences) lowast119875 lt 005 lowastlowast119875 lt 001 significantly dif-ferent versus Control
119875 lt 005 significantly different versus NAC
32 NAC Administration Lowers Mitochondrial AntioxidantCapacity In regard to the effects of antioxidant supple-ment on skeletal muscle autophagy we cannot ignore therole of mitochondria On one hand endogenous ROS ismainly generated by mitochondria on the other handmitochondrial antioxidant enzyme is able to scavenge ROSand reduce oxidative stress Recent works have suggestedthat mitochondrial-derived ROS induces autophagy [30 33]thus mitochondrial antioxidant capacity may be responsiblefor the activation of autophagy In the previous studiesNAC has been considered as a mitochondrial enhancerfor improving redox status and mitochondrial dysfunctionand reversing the progression of some diseases [37 38]Consistent with the previous results our data show that NACreduces mitochondrial ROS level and increases the ratio ofGSH GSSG suggesting that reduced ROS production andelevated GSH GSSG ratio in mitochondria are involved inthe inhibition of skeletal muscle constitutive autophagy Fur-ther NAC in part prevents fasting-induced oxidative stress inmitochondria but not autophagic responses indicating thatskeletal muscle autophagy under starvation is independentof mitochondrial-derived ROS In contrast to our results
in fasted mice lower nutrient supply or calorie restrictionleads to a lower production of mitochondrial ROS in skeletalmuscle [39] because the lower mitochondrial membranepotential favors reducing mitochondrial ROS production[40] In skeletal muscle of both rodents and humans high-fatdiet increases H
2O2emission of mitochondria [41] However
in response to acute nutrient deprivation mitochondrialROS levels significantly increase in skeletal muscle cells [30]These findings strongly indicate that ROS production inmitochondria is nutrient-sensitizing [42] It is noteworthythat chronic mild nutrient restriction differs from acutenutrient deprivation in the regulation of mitochondrial ROSproduction Thus the differences in constitutive and acutestress-induced autophagy may result from differential ROSlevels in skeletal muscle
Manganese superoxide dismutase (MnSOD) is a vitalantioxidant enzyme in the matrix of mitochondria by scav-enging ROS MnSOD knockdown led to chronic oxidativestress increased autophagy and mitochondrial biogenesiswithin the distal nephrons [43] In brain MnSOD knock-down increased oxidative stress and instead suppressedthe levels of autophagy stimulators [44] To our surprise
6 Oxidative Medicine and Cellular Longevity
LC3-I
TIGAR
LC3-II
Con
trol
Fast
NAC
NAC
+ F
ast
120573-Actin
(a)
0
05
1
15
2
Control Fast NAC
LC3
lipid
atio
n (C
ontro
l (
))
LC3-IILC3-I
NAC + Fast
lowastlowast
(b)
0
5
10
15
20
25
30
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
TIGARaTIGARb
lowastlowastlowastlowast
lowastlowast
(c)
Figure 4 LC3 lipidation and expressions of TIGAR were measured in skeletal muscle after short-term fasting (a) Representative westernblots of LC3 TIGAR (b) LC3-IILC3-I ratio (c) mRNA expression of TIGAR Values are means plusmn SE lowastlowast119875 lt 001 significantly differentversus Control
119875 lt 005 119875 lt 001 significantly different versus NAC
our results show that NAC decreases MnSODmRNA expres-sion and its enzyme activity CuZn-SOD expression is alsorepressed Fasting fails to rescue the decline in MnSODexpression Our data suggest that although NAC adminis-tration reduces ROS level and increases GSH GSSG exoge-nous antioxidant supplementmay lower inherent antioxidantcapacity in mitochondria ROS can lead to MnSOD upreg-ulation through ERK2 translocation into the nucleus anddissociation of p53 from its inhibitory protein mouse doubleminute 2 (MDM2) [45] Similarly heat stress producedROS in fibroblasts NAC blocked the increase in MnSODlevels by heat stress through reducing ROS production [46]Therefore a proper level of ROS is required for MnSODexpression Likewise we observe that TIGAR is repressedin mRNA and protein level by NAC Although TIGAR isnot an antioxidant enzyme that eliminates ROS directlyTIGAR plays an important antioxidant role in the regu-lation of autophagy [47] TIGAR inhibits glycolysis andpromotes respiration In TIGAR-expressing cells ROS levelwas redu-ced whereas glutathione was elevated [48] FurtherTIGAR inhibited autophagy by suppressing ROS level [47]TIGAR knockout increased mitophagy and ROS productionin myocardium followed by Bnip3 activation which is an
initiator of mitophagy Further NAC reversed the activationof Bnip3 and mitophagy [49] Under hypoxia mitochondriallocalization of TIGAR can control mitochondrial ROS levelsand protect from cell death [50] Therefore TIGAR is apotential antioxidant gene formitochondria Collectively ourresults indicate that NAC treatment with young mice atten-uates the inherent antioxidant capacity in skeletal musclesuggesting that ROS is also required for the maintenance ofmitochondrial antioxidant capacity
In summary our study provides the first evidencethat NAC decreases skeletal muscle constitutive autophagybut does not protect skeletal muscle from fasting-inducedautophagy in young mice In addition we also showed thatNAC attenuates the inherent antioxidant capacity in skeletalmuscle by depressing the expression of MnSOD and TIGARWe interpret these findings as an indication that ROS isrequired for the maintenance of both constitutive autophagyand antioxidant capacity in skeletal muscle Although NACsupplement is regarded as an effective intervention forimprovingmitochondrial function it should be reconsideredcarefully for young and healthy individuals when NACor the other antioxidants are used to promote musclegrowth
Oxidative Medicine and Cellular Longevity 7
0
20
40
60
80
100
120
140
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
SOD1 (CuZn-SOD)
lowastlowast
(a)
mRN
A le
vel (
vers
us 120573
-act
in)
0
50
100
150
200
250
Control Fast NAC NAC + Fast
SOD2 (MnSOD)
lowastlowast
(b)
0
02
04
06
08
1
12
14
Control Fast NAC NAC + Fast
Con
trol (
)
MnSOD activity
lowastlowast
(c)
Figure 5 Antioxidant capacity wasmeasured in gastrocnemiusmuscle after short-term fasting (a) mRNA expression of SOD1 (CuZn-SOD)(b) mRNA expression of SOD2 (MnSOD) (c) MnSOD activity in the isolated mitochondria of gastrocnemius muscle Values are means plusmnSE lowastlowast119875 lt 001 significantly different versus Control
119875 lt 005 significantly different versus NAC
4 Materials and Methods
41 Animals Male ICRCD-1 mice were purchased fromSino-British SipprBK Lab Animal Ltd CO (ShanghaiChina) at four to five weeks of age (Young 228 plusmn 12 g) Ani-mals were housed in a temperature-controlled environment(23 plusmn 2∘C) with 12 12 h light-dark cycles where they wereprovided diets and water ad libitum Mice were randomlyassigned to one of four experimental groups (1) Controlno drug treatment (Control 119899 = 8) (2) Fast no drugtreatment (Fast 119899 = 8) (3) Control treated with NAC (NAC119899 = 10) and (4) Fast treated with NAC (NAC + Fast119899 = 10) All experiments were performed in accordance withthe guidelines for the use of laboratory animals publishedby China Ministry of Health (no 55 order ordained on 25Jan 1998) All experimental procedureswere approved by theExperimental Animal Care andUse Committee at East ChinaNormal University (ECNU 2006-05)
42 Drugs and Short-TermFasting TheantioxidantN-acetyl-cysteine (NAC Sigma) was dissolved in DMSO The miceassigned to NAC groups received intraperitoneal injection ofNAC for 3weeks (100mgkg bodyweight2 day) As a controlthe vehicle (10 DMSO in saline) was injected to the controlmice For studies on the effects of food deprivation the mice
assigned to Fast groups were deprived of food for 24 hr beforesacrifice These mice had free access to drinking water
43 Mitochondrial Isolation At the end of the experimentalperiod animals in each group were acutely anesthetized withpentobarbital sodium Quadriceps gastrocnemius muscleand liver were removed completely and weighed For mito-chondrial isolation fresh gastrocnemius muscle was rinsedwith PBS and put into ice-cold isolation buffer (0075Msucrose 0225M sorbitol 1mM EGTA 01 fatty acid-freebovine serum albumin (BSA) and 10mM Tris-HCl pH 74)Tissues were sheared carefully to mince rinsed to get ridof residual blood and then homogenized in 1mL isolationbuffer per 100mg tissue The homogenate was centrifugedat 1000 g for 5min at 4∘C using a Beckman centrifuge(Avanti J-26XP) the resulting supernatant was decanted andsaved The pellet was washed once with isolation buffer Thesupernatant was combined and centrifuged at 9000 g for10min at 4∘C The mitochondrial pellet was washed andcentrifuged twice at 15000 g for 2min at 4∘C with isolationbuffer [51] Mitochondrial protein content was assayed usingBSA as a standard according to Bradford
44Mitochondrial ROSGeneration and Antioxidant CapacityThe dichlorofluorescin diacetate (DCF-DA) was used as
8 Oxidative Medicine and Cellular Longevity
a probe to detect mitochondrial ROS generation The assaywas performed on freshly isolated mitochondria Samples(30 120583g proteins) were incubated with 10 120583L of DCF-DA(10 120583molI) for 3 h at 37∘C The fluorescent product formedwas quantified by spectrofluorometer at the 485525 nmChanges in fluorescence were expressed as arbitrary unitTo assay mitochondrial lipid peroxidation MDA levels weremeasured using an assay kit according to the manufac-turerrsquos protocol (Nanjing Jiancheng Biotech Nanjing China)Briefly isolated mitochondria were incubated at 37∘C for 3 htreated with buffer and centrifuged at 10000timesg for 15minThe supernatants were incubated with thiobarbituric acid(TBA) and the absorbance of the supernatants wasmeasuredusing a spectrophotometer at a wavelength of 535 nm Alsothe GSHGSSG ratio and MnSOD activity were measuredusing an assay kit (Nanjing Jiancheng Biotech NanjingChina) according to themanufacturerrsquos protocol respectively
45 Caspases Activity Caspases 3 8 and 9 activity wasdetermined with an assay kit (Beyotime China) accordingto the manufacturerrsquos instructions Under the caspase-3acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) canbe changed into the yellow product p-nitroaniline (pNA)Under the caspase-8 acetyl-Ile-Glu-Thr-Asp p-nitroanilide(Ac-IETD-pNA) can be changed into the yellow prod-uct pNA Under the caspase-9 acetyl-Leu-Glu-His-Asp p-nitroanilide (Ac-LEHD-pNA) can be changed into the yellowproduct pNA Finally pNA content was measured usingthe spectrophotometer at an absorbance of 405 nm Proteincontent was assayed using BSA as a standard according toBradford
46 Glycogen Content Glycogen content was determinedwith an assay kit (Nanjing Jianchen Biotech China) accord-ing to the manufacturerrsquos instructions
47 Real-Time PCR Total RNA was prepared from sim100mgof frozen tissues using TRIzol (Invitrogen Chromos Singa-pore) and purified according to the instructions includedDouble-stranded cDNA was synthesized from sim1 120583g of totalRNA using ReverTra Ace qPCR RT kit (TOYOBO OsakaJapan) Real-time PCR reactions were set up using the SYBR-Green PCR kit (TOYOBO Osaka Japan) and were cycled inStepOne Real-Time PCR System (Applied Biosystems CAUSA) as previously described [52] The abundance of targetmRNA was normalized to that of 120573-actin Primer pairs weredesigned based on GenBank reference sequences and listedin Table S1 available online in Supplementary Material athttpdxdoiorg1011552014315896
48 Western Blotting The isolated muscle tissue (sim200mg)was weighed and cut into pieces at 4∘C and 2mL (10times dryweight of isolated muscle fibers) of ice-cold buffer (50mMTris HCl 150mMNaCl 1mM EDTA 02mM PMSF 1 NP-40 pH 74 cocktail) was added The tissue pieces were thenhomogenized after which homogenates were centrifuged for10min at 8000 g and 4∘C The total protein content of thesupernatant was quantified using bicinchoninic acid reagents
and BSA standards (Shanghai Sangon Shanghai China)Theprotein samples were immunoblotted as previously described[52] Visualization of reaction bands was performed by 331015840-diaminobenzidine staining (Shanghai Sangon China) andscanned densitometrically 120573-Actin was used to standardizefor the amount of protein loaded
49 Statistical Analysis Data are reported as mean plusmn SEMStatistical differences between treatments were determinedusing ANOVA For all tests the significance level was set at119875 lt 005
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from the National NaturalScience Foundation of China (Grant nos 31300977 and31171142) This work was also supported by the Key Labora-tory Construction Project of Adolescent Health Assessmentand Exercise Intervention of Ministry of Education EastChinaNormalUniversity (no 40500-541235-14203004)Theauthors also thank postgraduate students at Professor Dingrsquoslab for the technical support on animal care biochemicalassays PCR and western blot
References
[1] M Sandri ldquoAutophagy in skeletal musclerdquo FEBS Letters vol584 no 7 pp 1411ndash1416 2010
[2] G Twig B Hyde and O S Shirihai ldquoMitochondrial fusionfission and autophagy as a quality control axis the bioenergeticviewrdquo Biochimica et Biophysica Acta Bioenergetics vol 1777 no9 pp 1092ndash1097 2008
[3] Q Sun W Fan K Chen X Ding S Chen and Q ZhongldquoIdentification of Barkor as a mammalian autophagy-specificfactor for Beclin 1 and class III phosphatidylinositol 3-kinaserdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 49 pp 19211ndash19216 2008
[4] M C Maiuri E Zalckvar A Kimchi and G Kroemer ldquoSelf-eating and self-killing crosstalk between autophagy and apop-tosisrdquo Nature Reviews Molecular Cell Biology vol 8 no 9 pp741ndash752 2007
[5] T Ueno W Sato Y Horie et al ldquoLoss of Pten a tumor suppres-sor causes the strong inhibition of autophagy without affectingLC3 lipidationrdquo Autophagy vol 4 no 5 pp 692ndash700 2008
[6] F FortunatoH Burgers F Bergmann et al ldquoImpaired autolyso-some formation correlates with lamp-2 depletion role of apop-tosis autophagy and necrosis in pancreatitisrdquoGastroenterologyvol 137 no 1 pp 350ndash360 2009
[7] F A Graca D A Goncalves W A Silveira et al ldquoEpinephrinedepletion exacerbates the fasting-induced protein breakdown infast-twitch skeletal musclesrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 305 pp E1483ndashE14942013
[8] T Ogata Y Oishi M Higuchi and I Muraoka ldquoFasting-relatedautophagic response in slow- and fast-twitch skeletal musclerdquo
Oxidative Medicine and Cellular Longevity 9
Biochemical andBiophysical ResearchCommunications vol 394no 1 pp 136ndash140 2010
[9] A Doria M Gatto and L Punzi ldquoAutophagy in human healthand diseaserdquo New England Journal of Medicine vol 368 no 19p 1845 2013
[10] S Fulda ldquoAutophagy and cell deathrdquo Autophagy vol 8 no 8pp 1250ndash1251 2012
[11] E Masiero L Agatea C Mammucari et al ldquoAutophagy isrequired to maintain muscle massrdquo Cell Metabolism vol 10 no6 pp 507ndash515 2009
[12] A C Nascimbeni M Fanin E Masiero C Angelini and MSandri ldquoImpaired autophagy contributes to muscle atrophy inglycogen storage disease type II patientsrdquo Autophagy vol 8 no11 pp 1697ndash1700 2012
[13] M Sandri ldquoProtein breakdown in muscle wasting role ofautophagy-lysosome and ubiquitin-proteasomerdquo InternationalJournal of Biochemistry and Cell Biology vol 45 pp 2121ndash21292013
[14] E Masiero and M Sandri ldquoAutophagy inhibition inducesatrophy and myopathy in adult skeletal musclesrdquo Autophagyvol 6 no 2 pp 307ndash309 2010
[15] R Scherz-Shouval and Z Elazar ldquoRegulation of autophagy byROS physiology and pathologyrdquoTrends in Biochemical Sciencesvol 36 no 1 pp 30ndash38 2011
[16] C R Morales Z Pedrozo S Lavandero and J A Hill ldquoOxi-dative stress and autophagy in cardiovascular homeostasisrdquoAntioxidants amp Redox Signaling vol 20 pp 507ndash518 2014
[17] C Luo Y Li H Wang et al ldquoMitochondrial accumulationunder oxidative stress is due to defects in autophagyrdquo Journalof Cellular Biochemistry vol 114 no 1 pp 212ndash219 2013
[18] Y Wang Y Nartiss B Steipe G A McQuibban and PK Kim ldquoROS-induced mitochondrial depolarization initi-ates PARK2PARKIN-dependent mitochondrial degradationby autophagyrdquo Autophagy vol 8 no 10 pp 1462ndash1476 2012
[19] T Zhang Y Li K A Park et al ldquoCucurbitacin induces auto-phagy through mitochondrial ROS production which counter-acts to limit caspase-dependent apoptrdquo Autophagy vol 8 no 4pp 559ndash576 2012
[20] R Sandhir A Sood A Mehrotra and S S Kamboj ldquoN-acetyl-cysteine reverses mitochondrial dysfunctions and behavioralabnormalities in 3-nitropropionic acid-induced Huntingtonsdiseaserdquo Neurodegenerative Diseases vol 9 no 3 pp 145ndash1572012
[21] S S Kamboj and R Sandhir ldquoProtective effect of N-acetyl-cysteine supplementation onmitochondrial oxidative stress andmitochondrial enzymes in cerebral cortex of streptozotocin-treated diabetic ratsrdquoMitochondrion vol 11 no 1 pp 214ndash2222011
[22] M ZafarullahW Q Li J Sylvester andM Ahmad ldquoMolecularmechanisms of N-acetylcysteine actionsrdquo Cellular and Molecu-lar Life Sciences vol 60 no 1 pp 6ndash20 2003
[23] N A Strobel J M Peake A Matsumoto S A Marsh J SCoombes and G D Wadley ldquoAntioxidant supplementationreduces skeletal muscle mitochondrial biogenesisrdquo Medicineand Science in Sports and Exercise vol 43 no 6 pp 1017ndash10242011
[24] M-C Gomez-Cabrera E Domenech M Romagnoli et alldquoOral administration of vitamin C decreases muscle mitochon-drial biogenesis and hampers training-induced adaptationsin endurance performancerdquo The American Journal of ClinicalNutrition vol 87 no 1 pp 142ndash149 2008
[25] N P Whitehead C Pham O L Gervasio and D G Allen ldquoN-Acetylcysteine ameliorates skeletal muscle pathophysiology inmdx micerdquo Journal of Physiology vol 586 no 7 pp 2003ndash20142008
[26] C-H Lin S-C Kuo L-J Huang and P-W Gean ldquoNeuropro-tective effect ofN-acetylcysteine on neuronal apoptosis inducedby a synthetic gingerdione compound involvement of ERK andp38 phosphorylationrdquo Journal of Neuroscience Research vol 84no 7 pp 1485ndash1494 2006
[27] O BKotoulas S AKalamidas andD J Kondomerkos ldquoGlyco-gen autophagy in glucose homeostasisrdquo Pathology Research andPractice vol 202 no 9 pp 631ndash638 2006
[28] J Zirin J Nieuwenhuis and N Perrimon ldquoRole of autophagyin glycogen breakdown and its relevance to chloroquinemyopa-thyrdquo PLoS Biology vol 11 no 11 Article ID e1001708 2013
[29] P Castets S Lin N Rion et al ldquoSustained activation ofmTORC1 in skeletalmuscle inhibits constitutive and starvation-induced autophagy and causes a severe late-onset myopathyrdquoCell Metabolism vol 17 no 5 pp 731ndash744 2013
[30] M Rahman M Mofarrahi A S Kristof B Nkengfac S Hareland S N Hussain ldquoReactive oxygen species regulation ofautophagy in skeletal musclesrdquoAntioxidants amp Redox Signalingvol 20 no 3 pp 443ndash459 2014
[31] J Yan Z Feng J Liu et al ldquoEnhanced autophagy plays a cardi-nal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats ameliorating effects of (-)-epigallocatechin-3-gallaterdquo Journal of Nutritional Biochemistry vol 23 no 7 pp716ndash724 2012
[32] LV Yuzefovych S IMusiyenkoG LWilson and L I RachekldquoMitochondrial DNA damage and dysfunction and oxidativestress are associated with endoplasmic reticulum stress proteindegradation and apoptosis in high fat diet-induced InsulinresistancemicerdquoPLoSONE vol 8 no 1 Article ID e54059 2013
[33] L Li Y Chen and S B Gibson ldquoStarvation-induced autophagyis regulated bymitochondrial reactive oxygen species leading toAMPK activationrdquo Cellular Signalling vol 25 no 1 pp 50ndash652013
[34] S Takikita C Schreiner R Baum et al ldquoFiber type conversionby PGC-1120572 activates lysosomal and autophagosomal biogenesisin both unaffected and pompe skeletal musclerdquo PLoS ONE vol5 no 12 Article ID e15239 2010
[35] K Hollinger D Gardan-Salmon C Santana D Rice E Snellaand J T Selsby ldquoRescue of dystrophic skeletal muscle byPGC-1120572 involves restored expression of dystrophin-associatedprotein complex components and satellite cell signalingrdquo TheAmerican Journal of Physiology Regulatory Integrative andComparative Physiology vol 305 no 1 pp R13ndashR23 2013
[36] T Wenz S G Rossi R L Rotundo B M Spiegelman and CT Moraes ldquoIncreased muscle PGC-1120572 expression protects fromsarcopenia and metabolic disease during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 48 pp 20405ndash20410 2009
[37] I Grattagliano P Portincasa T Cocco et al ldquoEffect of dietaryrestriction and N-acetylcysteine supplementation on intestinalmucosa and liver mitochondrial redox status and function inaged ratsrdquo Experimental Gerontology vol 39 no 9 pp 1323ndash1332 2004
[38] S Kumar and S L Sitasawad ldquoN-acetylcysteine prevents glu-coseglucose oxidase-induced oxidative stress mitochondrialdamage and apoptosis in H9c2 cellsrdquo Life Sciences vol 84 no11-12 pp 328ndash336 2009
10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 3
002040608
112141618
Control Fast NAC NAC + Fast
Con
trol (
)
MDA
(a)
0
02
04
06
08
1
12
14
16
Control Fast NAC NAC + Fast
GSH GSSG
Con
trol (
)
lowast
lowast
(b)
0
05
1
15
2
25
Control Fast NAC
ROS level
NAC + Fast
Con
trol (
)
lowast
lowastlowast
(c)
Figure 1Oxidative stress biomarkers in the isolatedmitochondria of gastrocnemiusmuscleweremeasured after short-term fasting (a)MDAmethane dicarboxylic aldehyde (b) GSH glutathione (c) ROS production Values are means plusmn SE (fold differences) lowast119875 lt 005 lowastlowast119875 lt 001significantly different versus Control
119875 lt 005 significantly different versus NAC
24 NAC Represses TIGAR Expression but Fails to Protectagainst Fasting-Induced LC3 Lipidation In our study TIGARexpression was repressed significantly in the NAC groupcompared with Control group (119875 lt 001 Figures 4(a)and 4(c)) TIGARb expression was significantly increasedin the Fast group compared with Control group (119875 lt001 Figure 4(c)) and TIGARa expression was significantlyincreased in the NAC + Fast group compared with NACgroup (119875 lt 005 Figure 4(c)) In addition the ratio of LC3II to LC3 I (LC3 IILC3 I) was also quantified as a marker ofLC3 cleavage Fasting significantly increased LC3 lipidationindependent of NAC treatment (119875 lt 001 Figures 4(a) and4(b))
25 NAC Decreases MnSOD Expression and MitochondrialAntioxidant Capacity Our results show that mRNA expres-sion of CuZn-SOD MnSOD in skeletal muscle was signifi-cantly decreased in the NAC group compared with Control(119875 lt 001 Figures 5(a) and 5(b)) Similarly MnSOD activitywas significantly decreased in the NAC group compared withControl (119875 lt 001 Figure 5(c)) Also fasting had no effectson MnSOD expression and activity (Figure 5(c))
3 Discussion
These experiments provide novel and important informationregarding the effects of NAC administration on skeletalmuscle autophagy First our findings support the hypothesis
that NAC treatment dramatically decreases the expression ofnumerous markers of skeletal muscle autophagy at the basallevel rather than under the stress state of fasting Furtherour results reveal that NAC treatment is potential to lowermitochondrial antioxidant capacity through the inhibition ofthe antioxidant enzymes in mitochondria
31 NAC Administration Depresses Skeletal Muscle Consti-tutive Autophagy Significantly constitutive autophagy atthe basal level differs from stress-induced autophagy underconditions of nutrition deprivation In stress state enhancedautophagy favors cell survival by the degradation of cel-lular constituents and the recycling of macromolecules tosupport cellular metabolism Glycogen in skeletal muscleand liver can be utilized to produce energy or glucoseduring fasting so glycogen catabolism is also regarded asa peculiar autophagy outside lysosome [27] In response tostarvation the activation of autophagy is essential for themost efficient degradation of glycogen [28] In unstressedstate the basal level of autophagy in most tissues ensuresthe turnover of long-lived proteins and the elimination ofdysfunctional organelles Mammalian target of rapamycincomplex I (mTORC1) is regarded as the dominant regulatorof autophagy induction in skeletal muscle In young TSC1-(tuberous sclerosis complex 1-) deficient mice constitu-tive and starvation-induced autophagy was blocked at theinduction steps via mTORC1-mediated inhibition of ULK1(Unc-51-like kinase 1) [29] Recent studies suggested that
4 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
Control Fast NAC
Gly
coge
n co
nten
t (m
mol
g)
Skeletal muscle
NAC + Fast
lowast
lowastlowast
(a)
0
05
1
15
2
Control Fast NAC
Gly
coge
n co
nten
t (m
mol
g)
Liver
NAC + Fast
lowast
(b)
200
220
240
260
280
300
Left Right
Qua
dric
eps w
eigh
t (m
g)
ControlFast
NACNAC + Fast
lowastlowast
(c)
Figure 2Glycogen content and quadricepsmuscleweightweremeasured after short-term fasting (a)Glycogen content in quadricepsmuscle(b) Glycogen content in liver (c) Quadriceps muscle weight Values are means plusmn SE lowast119875 lt 005 lowastlowast119875 lt 001 significantly different versusControl
119875 lt 001 significantly different versus NAC
endogenous ROS promoted skeletal muscle autophagy at thebasal level and in response to acute nutrient starvation Thiseffect was mediated through regulation of autophagosomeinitiation and PI3KAKT inhibition [30] Using a potentialautophagy-targeting nutrient excessive muscle autophagy intype 2 diabetic rats was ameliorated through the downregu-lation of ROS signaling leading to improvement of glucosemetabolism reduction of oxidative stress and inhibition ofmitochondrial loss and dysfunction [31] ROS scavengerNACwas reported to diminish both endoplasmic reticulum (ER)stress and autophagy markers in skeletal muscle cells [32]Further NAC attenuated AMPK activation under starvationcondition and enhanced mTOR signaling in cervical cancercell The authors concluded that mitochondria-generatedROS induces autophagy mediated by the AMPK pathwayunder starvation conditions [33] Consistent with the previ-ous studies and our hypothesis our results show that NACsignificantly decreases mRNA expression of numerousmark-ers of skeletalmuscle autophagy and increases skeletalmuscleglycogenThese results indicate that the basal level of skeletalmuscle autophagy is ROS-dependent Constitutive autophagyin skeletal muscle is determined by redox state However ourresults also show that NAC fails to prevent fasting-inducedautophagy and glycogen reduction Specifically short-termfasting after NAC administration still increases the mRNAexpression of numerousmarkers of autophagy and the level of
LC3 lipidation These results indicate that ROS-independentautophagy occurs in skeletal muscle under starvation condi-tions In addition the activation of apoptosis also contributedto muscle loss relevant to oxidative stress Our data showthat NAC blocks fasting-induced activation of caspases 3and 9 and quadriceps muscle loss Collectively our resultsindicate that ROS is not required for fasting-induced auto-phagy but important for constitutive autophagy in skeletalmuscle
Additionally our results show that the expression ofmost Atgs is higher in red muscle than in white muscleThis difference in constitutive autophagy may result fromdifferential PGC-1120572 signaling and mitochondrial content inslow- and fast-twitch skeletal muscle Two similar works atleast partly support this postulation fiber type conversionfrom fast to slow fibers by PGC-1120572 activates lysosomaland autophagosomal biogenesis [34] as well as mitochon-drial biogenesis in skeletal muscle PGC-1120572 overexpressionincreases the expression of genes related to muscle repairand autophagy [35] Thus we suggest that the level ofconstitutive autophagy in slowmuscle fibers is higher than infast muscle fibers However PGC-1120572 overexpression duringaging protects from sarcopenia and metabolic disease byreducing apoptosis and autophagy [36] Further studiesneed to determine the relationship between mitochondrialbiogenesis and constitutive autophagy
Oxidative Medicine and Cellular Longevity 5
0
02
04
06
08
1
12
14
Beclin-1Atg6 Atg7 LC3Atg8 Atg9 LAMP-2
mRN
A le
vel (
Con
trol (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowastlowastlowast
lowastlowastlowast
(a)
0
05
1
15
2
25
Caspase-3 Caspase-8 Caspase-9
Enzy
me a
ctiv
ity (C
ontro
l (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowast
lowastlowast
(b)
0
05
1
15
2
25
3
mRN
A in
skel
etal
mus
cle(r
elat
ive e
xpre
ssio
n)
Young mice
Atg7 Atg9 Beclin-1 LAMP-2 LC3 p53
WGRG
lowastlowast
lowast
lowast lowast
(c)
Figure 3 Autophagy-related gene expressions were analyzed by real-time PCR in gastrocnemius muscle (a) mRNA level of Beclin-1 Atg7LC3 Atg9 and LAMP2 (b) Caspases 3 8 and 9 enzyme activity in gastrocnemius muscle (c) Autophagy-related gene expressions in redgastrocnemius (RG) versus white gastrocnemius (WG) Values are means plusmn SE (fold differences) lowast119875 lt 005 lowastlowast119875 lt 001 significantly dif-ferent versus Control
119875 lt 005 significantly different versus NAC
32 NAC Administration Lowers Mitochondrial AntioxidantCapacity In regard to the effects of antioxidant supple-ment on skeletal muscle autophagy we cannot ignore therole of mitochondria On one hand endogenous ROS ismainly generated by mitochondria on the other handmitochondrial antioxidant enzyme is able to scavenge ROSand reduce oxidative stress Recent works have suggestedthat mitochondrial-derived ROS induces autophagy [30 33]thus mitochondrial antioxidant capacity may be responsiblefor the activation of autophagy In the previous studiesNAC has been considered as a mitochondrial enhancerfor improving redox status and mitochondrial dysfunctionand reversing the progression of some diseases [37 38]Consistent with the previous results our data show that NACreduces mitochondrial ROS level and increases the ratio ofGSH GSSG suggesting that reduced ROS production andelevated GSH GSSG ratio in mitochondria are involved inthe inhibition of skeletal muscle constitutive autophagy Fur-ther NAC in part prevents fasting-induced oxidative stress inmitochondria but not autophagic responses indicating thatskeletal muscle autophagy under starvation is independentof mitochondrial-derived ROS In contrast to our results
in fasted mice lower nutrient supply or calorie restrictionleads to a lower production of mitochondrial ROS in skeletalmuscle [39] because the lower mitochondrial membranepotential favors reducing mitochondrial ROS production[40] In skeletal muscle of both rodents and humans high-fatdiet increases H
2O2emission of mitochondria [41] However
in response to acute nutrient deprivation mitochondrialROS levels significantly increase in skeletal muscle cells [30]These findings strongly indicate that ROS production inmitochondria is nutrient-sensitizing [42] It is noteworthythat chronic mild nutrient restriction differs from acutenutrient deprivation in the regulation of mitochondrial ROSproduction Thus the differences in constitutive and acutestress-induced autophagy may result from differential ROSlevels in skeletal muscle
Manganese superoxide dismutase (MnSOD) is a vitalantioxidant enzyme in the matrix of mitochondria by scav-enging ROS MnSOD knockdown led to chronic oxidativestress increased autophagy and mitochondrial biogenesiswithin the distal nephrons [43] In brain MnSOD knock-down increased oxidative stress and instead suppressedthe levels of autophagy stimulators [44] To our surprise
6 Oxidative Medicine and Cellular Longevity
LC3-I
TIGAR
LC3-II
Con
trol
Fast
NAC
NAC
+ F
ast
120573-Actin
(a)
0
05
1
15
2
Control Fast NAC
LC3
lipid
atio
n (C
ontro
l (
))
LC3-IILC3-I
NAC + Fast
lowastlowast
(b)
0
5
10
15
20
25
30
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
TIGARaTIGARb
lowastlowastlowastlowast
lowastlowast
(c)
Figure 4 LC3 lipidation and expressions of TIGAR were measured in skeletal muscle after short-term fasting (a) Representative westernblots of LC3 TIGAR (b) LC3-IILC3-I ratio (c) mRNA expression of TIGAR Values are means plusmn SE lowastlowast119875 lt 001 significantly differentversus Control
119875 lt 005 119875 lt 001 significantly different versus NAC
our results show that NAC decreases MnSODmRNA expres-sion and its enzyme activity CuZn-SOD expression is alsorepressed Fasting fails to rescue the decline in MnSODexpression Our data suggest that although NAC adminis-tration reduces ROS level and increases GSH GSSG exoge-nous antioxidant supplementmay lower inherent antioxidantcapacity in mitochondria ROS can lead to MnSOD upreg-ulation through ERK2 translocation into the nucleus anddissociation of p53 from its inhibitory protein mouse doubleminute 2 (MDM2) [45] Similarly heat stress producedROS in fibroblasts NAC blocked the increase in MnSODlevels by heat stress through reducing ROS production [46]Therefore a proper level of ROS is required for MnSODexpression Likewise we observe that TIGAR is repressedin mRNA and protein level by NAC Although TIGAR isnot an antioxidant enzyme that eliminates ROS directlyTIGAR plays an important antioxidant role in the regu-lation of autophagy [47] TIGAR inhibits glycolysis andpromotes respiration In TIGAR-expressing cells ROS levelwas redu-ced whereas glutathione was elevated [48] FurtherTIGAR inhibited autophagy by suppressing ROS level [47]TIGAR knockout increased mitophagy and ROS productionin myocardium followed by Bnip3 activation which is an
initiator of mitophagy Further NAC reversed the activationof Bnip3 and mitophagy [49] Under hypoxia mitochondriallocalization of TIGAR can control mitochondrial ROS levelsand protect from cell death [50] Therefore TIGAR is apotential antioxidant gene formitochondria Collectively ourresults indicate that NAC treatment with young mice atten-uates the inherent antioxidant capacity in skeletal musclesuggesting that ROS is also required for the maintenance ofmitochondrial antioxidant capacity
In summary our study provides the first evidencethat NAC decreases skeletal muscle constitutive autophagybut does not protect skeletal muscle from fasting-inducedautophagy in young mice In addition we also showed thatNAC attenuates the inherent antioxidant capacity in skeletalmuscle by depressing the expression of MnSOD and TIGARWe interpret these findings as an indication that ROS isrequired for the maintenance of both constitutive autophagyand antioxidant capacity in skeletal muscle Although NACsupplement is regarded as an effective intervention forimprovingmitochondrial function it should be reconsideredcarefully for young and healthy individuals when NACor the other antioxidants are used to promote musclegrowth
Oxidative Medicine and Cellular Longevity 7
0
20
40
60
80
100
120
140
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
SOD1 (CuZn-SOD)
lowastlowast
(a)
mRN
A le
vel (
vers
us 120573
-act
in)
0
50
100
150
200
250
Control Fast NAC NAC + Fast
SOD2 (MnSOD)
lowastlowast
(b)
0
02
04
06
08
1
12
14
Control Fast NAC NAC + Fast
Con
trol (
)
MnSOD activity
lowastlowast
(c)
Figure 5 Antioxidant capacity wasmeasured in gastrocnemiusmuscle after short-term fasting (a) mRNA expression of SOD1 (CuZn-SOD)(b) mRNA expression of SOD2 (MnSOD) (c) MnSOD activity in the isolated mitochondria of gastrocnemius muscle Values are means plusmnSE lowastlowast119875 lt 001 significantly different versus Control
119875 lt 005 significantly different versus NAC
4 Materials and Methods
41 Animals Male ICRCD-1 mice were purchased fromSino-British SipprBK Lab Animal Ltd CO (ShanghaiChina) at four to five weeks of age (Young 228 plusmn 12 g) Ani-mals were housed in a temperature-controlled environment(23 plusmn 2∘C) with 12 12 h light-dark cycles where they wereprovided diets and water ad libitum Mice were randomlyassigned to one of four experimental groups (1) Controlno drug treatment (Control 119899 = 8) (2) Fast no drugtreatment (Fast 119899 = 8) (3) Control treated with NAC (NAC119899 = 10) and (4) Fast treated with NAC (NAC + Fast119899 = 10) All experiments were performed in accordance withthe guidelines for the use of laboratory animals publishedby China Ministry of Health (no 55 order ordained on 25Jan 1998) All experimental procedureswere approved by theExperimental Animal Care andUse Committee at East ChinaNormal University (ECNU 2006-05)
42 Drugs and Short-TermFasting TheantioxidantN-acetyl-cysteine (NAC Sigma) was dissolved in DMSO The miceassigned to NAC groups received intraperitoneal injection ofNAC for 3weeks (100mgkg bodyweight2 day) As a controlthe vehicle (10 DMSO in saline) was injected to the controlmice For studies on the effects of food deprivation the mice
assigned to Fast groups were deprived of food for 24 hr beforesacrifice These mice had free access to drinking water
43 Mitochondrial Isolation At the end of the experimentalperiod animals in each group were acutely anesthetized withpentobarbital sodium Quadriceps gastrocnemius muscleand liver were removed completely and weighed For mito-chondrial isolation fresh gastrocnemius muscle was rinsedwith PBS and put into ice-cold isolation buffer (0075Msucrose 0225M sorbitol 1mM EGTA 01 fatty acid-freebovine serum albumin (BSA) and 10mM Tris-HCl pH 74)Tissues were sheared carefully to mince rinsed to get ridof residual blood and then homogenized in 1mL isolationbuffer per 100mg tissue The homogenate was centrifugedat 1000 g for 5min at 4∘C using a Beckman centrifuge(Avanti J-26XP) the resulting supernatant was decanted andsaved The pellet was washed once with isolation buffer Thesupernatant was combined and centrifuged at 9000 g for10min at 4∘C The mitochondrial pellet was washed andcentrifuged twice at 15000 g for 2min at 4∘C with isolationbuffer [51] Mitochondrial protein content was assayed usingBSA as a standard according to Bradford
44Mitochondrial ROSGeneration and Antioxidant CapacityThe dichlorofluorescin diacetate (DCF-DA) was used as
8 Oxidative Medicine and Cellular Longevity
a probe to detect mitochondrial ROS generation The assaywas performed on freshly isolated mitochondria Samples(30 120583g proteins) were incubated with 10 120583L of DCF-DA(10 120583molI) for 3 h at 37∘C The fluorescent product formedwas quantified by spectrofluorometer at the 485525 nmChanges in fluorescence were expressed as arbitrary unitTo assay mitochondrial lipid peroxidation MDA levels weremeasured using an assay kit according to the manufac-turerrsquos protocol (Nanjing Jiancheng Biotech Nanjing China)Briefly isolated mitochondria were incubated at 37∘C for 3 htreated with buffer and centrifuged at 10000timesg for 15minThe supernatants were incubated with thiobarbituric acid(TBA) and the absorbance of the supernatants wasmeasuredusing a spectrophotometer at a wavelength of 535 nm Alsothe GSHGSSG ratio and MnSOD activity were measuredusing an assay kit (Nanjing Jiancheng Biotech NanjingChina) according to themanufacturerrsquos protocol respectively
45 Caspases Activity Caspases 3 8 and 9 activity wasdetermined with an assay kit (Beyotime China) accordingto the manufacturerrsquos instructions Under the caspase-3acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) canbe changed into the yellow product p-nitroaniline (pNA)Under the caspase-8 acetyl-Ile-Glu-Thr-Asp p-nitroanilide(Ac-IETD-pNA) can be changed into the yellow prod-uct pNA Under the caspase-9 acetyl-Leu-Glu-His-Asp p-nitroanilide (Ac-LEHD-pNA) can be changed into the yellowproduct pNA Finally pNA content was measured usingthe spectrophotometer at an absorbance of 405 nm Proteincontent was assayed using BSA as a standard according toBradford
46 Glycogen Content Glycogen content was determinedwith an assay kit (Nanjing Jianchen Biotech China) accord-ing to the manufacturerrsquos instructions
47 Real-Time PCR Total RNA was prepared from sim100mgof frozen tissues using TRIzol (Invitrogen Chromos Singa-pore) and purified according to the instructions includedDouble-stranded cDNA was synthesized from sim1 120583g of totalRNA using ReverTra Ace qPCR RT kit (TOYOBO OsakaJapan) Real-time PCR reactions were set up using the SYBR-Green PCR kit (TOYOBO Osaka Japan) and were cycled inStepOne Real-Time PCR System (Applied Biosystems CAUSA) as previously described [52] The abundance of targetmRNA was normalized to that of 120573-actin Primer pairs weredesigned based on GenBank reference sequences and listedin Table S1 available online in Supplementary Material athttpdxdoiorg1011552014315896
48 Western Blotting The isolated muscle tissue (sim200mg)was weighed and cut into pieces at 4∘C and 2mL (10times dryweight of isolated muscle fibers) of ice-cold buffer (50mMTris HCl 150mMNaCl 1mM EDTA 02mM PMSF 1 NP-40 pH 74 cocktail) was added The tissue pieces were thenhomogenized after which homogenates were centrifuged for10min at 8000 g and 4∘C The total protein content of thesupernatant was quantified using bicinchoninic acid reagents
and BSA standards (Shanghai Sangon Shanghai China)Theprotein samples were immunoblotted as previously described[52] Visualization of reaction bands was performed by 331015840-diaminobenzidine staining (Shanghai Sangon China) andscanned densitometrically 120573-Actin was used to standardizefor the amount of protein loaded
49 Statistical Analysis Data are reported as mean plusmn SEMStatistical differences between treatments were determinedusing ANOVA For all tests the significance level was set at119875 lt 005
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from the National NaturalScience Foundation of China (Grant nos 31300977 and31171142) This work was also supported by the Key Labora-tory Construction Project of Adolescent Health Assessmentand Exercise Intervention of Ministry of Education EastChinaNormalUniversity (no 40500-541235-14203004)Theauthors also thank postgraduate students at Professor Dingrsquoslab for the technical support on animal care biochemicalassays PCR and western blot
References
[1] M Sandri ldquoAutophagy in skeletal musclerdquo FEBS Letters vol584 no 7 pp 1411ndash1416 2010
[2] G Twig B Hyde and O S Shirihai ldquoMitochondrial fusionfission and autophagy as a quality control axis the bioenergeticviewrdquo Biochimica et Biophysica Acta Bioenergetics vol 1777 no9 pp 1092ndash1097 2008
[3] Q Sun W Fan K Chen X Ding S Chen and Q ZhongldquoIdentification of Barkor as a mammalian autophagy-specificfactor for Beclin 1 and class III phosphatidylinositol 3-kinaserdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 49 pp 19211ndash19216 2008
[4] M C Maiuri E Zalckvar A Kimchi and G Kroemer ldquoSelf-eating and self-killing crosstalk between autophagy and apop-tosisrdquo Nature Reviews Molecular Cell Biology vol 8 no 9 pp741ndash752 2007
[5] T Ueno W Sato Y Horie et al ldquoLoss of Pten a tumor suppres-sor causes the strong inhibition of autophagy without affectingLC3 lipidationrdquo Autophagy vol 4 no 5 pp 692ndash700 2008
[6] F FortunatoH Burgers F Bergmann et al ldquoImpaired autolyso-some formation correlates with lamp-2 depletion role of apop-tosis autophagy and necrosis in pancreatitisrdquoGastroenterologyvol 137 no 1 pp 350ndash360 2009
[7] F A Graca D A Goncalves W A Silveira et al ldquoEpinephrinedepletion exacerbates the fasting-induced protein breakdown infast-twitch skeletal musclesrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 305 pp E1483ndashE14942013
[8] T Ogata Y Oishi M Higuchi and I Muraoka ldquoFasting-relatedautophagic response in slow- and fast-twitch skeletal musclerdquo
Oxidative Medicine and Cellular Longevity 9
Biochemical andBiophysical ResearchCommunications vol 394no 1 pp 136ndash140 2010
[9] A Doria M Gatto and L Punzi ldquoAutophagy in human healthand diseaserdquo New England Journal of Medicine vol 368 no 19p 1845 2013
[10] S Fulda ldquoAutophagy and cell deathrdquo Autophagy vol 8 no 8pp 1250ndash1251 2012
[11] E Masiero L Agatea C Mammucari et al ldquoAutophagy isrequired to maintain muscle massrdquo Cell Metabolism vol 10 no6 pp 507ndash515 2009
[12] A C Nascimbeni M Fanin E Masiero C Angelini and MSandri ldquoImpaired autophagy contributes to muscle atrophy inglycogen storage disease type II patientsrdquo Autophagy vol 8 no11 pp 1697ndash1700 2012
[13] M Sandri ldquoProtein breakdown in muscle wasting role ofautophagy-lysosome and ubiquitin-proteasomerdquo InternationalJournal of Biochemistry and Cell Biology vol 45 pp 2121ndash21292013
[14] E Masiero and M Sandri ldquoAutophagy inhibition inducesatrophy and myopathy in adult skeletal musclesrdquo Autophagyvol 6 no 2 pp 307ndash309 2010
[15] R Scherz-Shouval and Z Elazar ldquoRegulation of autophagy byROS physiology and pathologyrdquoTrends in Biochemical Sciencesvol 36 no 1 pp 30ndash38 2011
[16] C R Morales Z Pedrozo S Lavandero and J A Hill ldquoOxi-dative stress and autophagy in cardiovascular homeostasisrdquoAntioxidants amp Redox Signaling vol 20 pp 507ndash518 2014
[17] C Luo Y Li H Wang et al ldquoMitochondrial accumulationunder oxidative stress is due to defects in autophagyrdquo Journalof Cellular Biochemistry vol 114 no 1 pp 212ndash219 2013
[18] Y Wang Y Nartiss B Steipe G A McQuibban and PK Kim ldquoROS-induced mitochondrial depolarization initi-ates PARK2PARKIN-dependent mitochondrial degradationby autophagyrdquo Autophagy vol 8 no 10 pp 1462ndash1476 2012
[19] T Zhang Y Li K A Park et al ldquoCucurbitacin induces auto-phagy through mitochondrial ROS production which counter-acts to limit caspase-dependent apoptrdquo Autophagy vol 8 no 4pp 559ndash576 2012
[20] R Sandhir A Sood A Mehrotra and S S Kamboj ldquoN-acetyl-cysteine reverses mitochondrial dysfunctions and behavioralabnormalities in 3-nitropropionic acid-induced Huntingtonsdiseaserdquo Neurodegenerative Diseases vol 9 no 3 pp 145ndash1572012
[21] S S Kamboj and R Sandhir ldquoProtective effect of N-acetyl-cysteine supplementation onmitochondrial oxidative stress andmitochondrial enzymes in cerebral cortex of streptozotocin-treated diabetic ratsrdquoMitochondrion vol 11 no 1 pp 214ndash2222011
[22] M ZafarullahW Q Li J Sylvester andM Ahmad ldquoMolecularmechanisms of N-acetylcysteine actionsrdquo Cellular and Molecu-lar Life Sciences vol 60 no 1 pp 6ndash20 2003
[23] N A Strobel J M Peake A Matsumoto S A Marsh J SCoombes and G D Wadley ldquoAntioxidant supplementationreduces skeletal muscle mitochondrial biogenesisrdquo Medicineand Science in Sports and Exercise vol 43 no 6 pp 1017ndash10242011
[24] M-C Gomez-Cabrera E Domenech M Romagnoli et alldquoOral administration of vitamin C decreases muscle mitochon-drial biogenesis and hampers training-induced adaptationsin endurance performancerdquo The American Journal of ClinicalNutrition vol 87 no 1 pp 142ndash149 2008
[25] N P Whitehead C Pham O L Gervasio and D G Allen ldquoN-Acetylcysteine ameliorates skeletal muscle pathophysiology inmdx micerdquo Journal of Physiology vol 586 no 7 pp 2003ndash20142008
[26] C-H Lin S-C Kuo L-J Huang and P-W Gean ldquoNeuropro-tective effect ofN-acetylcysteine on neuronal apoptosis inducedby a synthetic gingerdione compound involvement of ERK andp38 phosphorylationrdquo Journal of Neuroscience Research vol 84no 7 pp 1485ndash1494 2006
[27] O BKotoulas S AKalamidas andD J Kondomerkos ldquoGlyco-gen autophagy in glucose homeostasisrdquo Pathology Research andPractice vol 202 no 9 pp 631ndash638 2006
[28] J Zirin J Nieuwenhuis and N Perrimon ldquoRole of autophagyin glycogen breakdown and its relevance to chloroquinemyopa-thyrdquo PLoS Biology vol 11 no 11 Article ID e1001708 2013
[29] P Castets S Lin N Rion et al ldquoSustained activation ofmTORC1 in skeletalmuscle inhibits constitutive and starvation-induced autophagy and causes a severe late-onset myopathyrdquoCell Metabolism vol 17 no 5 pp 731ndash744 2013
[30] M Rahman M Mofarrahi A S Kristof B Nkengfac S Hareland S N Hussain ldquoReactive oxygen species regulation ofautophagy in skeletal musclesrdquoAntioxidants amp Redox Signalingvol 20 no 3 pp 443ndash459 2014
[31] J Yan Z Feng J Liu et al ldquoEnhanced autophagy plays a cardi-nal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats ameliorating effects of (-)-epigallocatechin-3-gallaterdquo Journal of Nutritional Biochemistry vol 23 no 7 pp716ndash724 2012
[32] LV Yuzefovych S IMusiyenkoG LWilson and L I RachekldquoMitochondrial DNA damage and dysfunction and oxidativestress are associated with endoplasmic reticulum stress proteindegradation and apoptosis in high fat diet-induced InsulinresistancemicerdquoPLoSONE vol 8 no 1 Article ID e54059 2013
[33] L Li Y Chen and S B Gibson ldquoStarvation-induced autophagyis regulated bymitochondrial reactive oxygen species leading toAMPK activationrdquo Cellular Signalling vol 25 no 1 pp 50ndash652013
[34] S Takikita C Schreiner R Baum et al ldquoFiber type conversionby PGC-1120572 activates lysosomal and autophagosomal biogenesisin both unaffected and pompe skeletal musclerdquo PLoS ONE vol5 no 12 Article ID e15239 2010
[35] K Hollinger D Gardan-Salmon C Santana D Rice E Snellaand J T Selsby ldquoRescue of dystrophic skeletal muscle byPGC-1120572 involves restored expression of dystrophin-associatedprotein complex components and satellite cell signalingrdquo TheAmerican Journal of Physiology Regulatory Integrative andComparative Physiology vol 305 no 1 pp R13ndashR23 2013
[36] T Wenz S G Rossi R L Rotundo B M Spiegelman and CT Moraes ldquoIncreased muscle PGC-1120572 expression protects fromsarcopenia and metabolic disease during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 48 pp 20405ndash20410 2009
[37] I Grattagliano P Portincasa T Cocco et al ldquoEffect of dietaryrestriction and N-acetylcysteine supplementation on intestinalmucosa and liver mitochondrial redox status and function inaged ratsrdquo Experimental Gerontology vol 39 no 9 pp 1323ndash1332 2004
[38] S Kumar and S L Sitasawad ldquoN-acetylcysteine prevents glu-coseglucose oxidase-induced oxidative stress mitochondrialdamage and apoptosis in H9c2 cellsrdquo Life Sciences vol 84 no11-12 pp 328ndash336 2009
10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Oxidative Medicine and Cellular Longevity
0
05
1
15
2
Control Fast NAC
Gly
coge
n co
nten
t (m
mol
g)
Skeletal muscle
NAC + Fast
lowast
lowastlowast
(a)
0
05
1
15
2
Control Fast NAC
Gly
coge
n co
nten
t (m
mol
g)
Liver
NAC + Fast
lowast
(b)
200
220
240
260
280
300
Left Right
Qua
dric
eps w
eigh
t (m
g)
ControlFast
NACNAC + Fast
lowastlowast
(c)
Figure 2Glycogen content and quadricepsmuscleweightweremeasured after short-term fasting (a)Glycogen content in quadricepsmuscle(b) Glycogen content in liver (c) Quadriceps muscle weight Values are means plusmn SE lowast119875 lt 005 lowastlowast119875 lt 001 significantly different versusControl
119875 lt 001 significantly different versus NAC
endogenous ROS promoted skeletal muscle autophagy at thebasal level and in response to acute nutrient starvation Thiseffect was mediated through regulation of autophagosomeinitiation and PI3KAKT inhibition [30] Using a potentialautophagy-targeting nutrient excessive muscle autophagy intype 2 diabetic rats was ameliorated through the downregu-lation of ROS signaling leading to improvement of glucosemetabolism reduction of oxidative stress and inhibition ofmitochondrial loss and dysfunction [31] ROS scavengerNACwas reported to diminish both endoplasmic reticulum (ER)stress and autophagy markers in skeletal muscle cells [32]Further NAC attenuated AMPK activation under starvationcondition and enhanced mTOR signaling in cervical cancercell The authors concluded that mitochondria-generatedROS induces autophagy mediated by the AMPK pathwayunder starvation conditions [33] Consistent with the previ-ous studies and our hypothesis our results show that NACsignificantly decreases mRNA expression of numerousmark-ers of skeletalmuscle autophagy and increases skeletalmuscleglycogenThese results indicate that the basal level of skeletalmuscle autophagy is ROS-dependent Constitutive autophagyin skeletal muscle is determined by redox state However ourresults also show that NAC fails to prevent fasting-inducedautophagy and glycogen reduction Specifically short-termfasting after NAC administration still increases the mRNAexpression of numerousmarkers of autophagy and the level of
LC3 lipidation These results indicate that ROS-independentautophagy occurs in skeletal muscle under starvation condi-tions In addition the activation of apoptosis also contributedto muscle loss relevant to oxidative stress Our data showthat NAC blocks fasting-induced activation of caspases 3and 9 and quadriceps muscle loss Collectively our resultsindicate that ROS is not required for fasting-induced auto-phagy but important for constitutive autophagy in skeletalmuscle
Additionally our results show that the expression ofmost Atgs is higher in red muscle than in white muscleThis difference in constitutive autophagy may result fromdifferential PGC-1120572 signaling and mitochondrial content inslow- and fast-twitch skeletal muscle Two similar works atleast partly support this postulation fiber type conversionfrom fast to slow fibers by PGC-1120572 activates lysosomaland autophagosomal biogenesis [34] as well as mitochon-drial biogenesis in skeletal muscle PGC-1120572 overexpressionincreases the expression of genes related to muscle repairand autophagy [35] Thus we suggest that the level ofconstitutive autophagy in slowmuscle fibers is higher than infast muscle fibers However PGC-1120572 overexpression duringaging protects from sarcopenia and metabolic disease byreducing apoptosis and autophagy [36] Further studiesneed to determine the relationship between mitochondrialbiogenesis and constitutive autophagy
Oxidative Medicine and Cellular Longevity 5
0
02
04
06
08
1
12
14
Beclin-1Atg6 Atg7 LC3Atg8 Atg9 LAMP-2
mRN
A le
vel (
Con
trol (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowastlowastlowast
lowastlowastlowast
(a)
0
05
1
15
2
25
Caspase-3 Caspase-8 Caspase-9
Enzy
me a
ctiv
ity (C
ontro
l (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowast
lowastlowast
(b)
0
05
1
15
2
25
3
mRN
A in
skel
etal
mus
cle(r
elat
ive e
xpre
ssio
n)
Young mice
Atg7 Atg9 Beclin-1 LAMP-2 LC3 p53
WGRG
lowastlowast
lowast
lowast lowast
(c)
Figure 3 Autophagy-related gene expressions were analyzed by real-time PCR in gastrocnemius muscle (a) mRNA level of Beclin-1 Atg7LC3 Atg9 and LAMP2 (b) Caspases 3 8 and 9 enzyme activity in gastrocnemius muscle (c) Autophagy-related gene expressions in redgastrocnemius (RG) versus white gastrocnemius (WG) Values are means plusmn SE (fold differences) lowast119875 lt 005 lowastlowast119875 lt 001 significantly dif-ferent versus Control
119875 lt 005 significantly different versus NAC
32 NAC Administration Lowers Mitochondrial AntioxidantCapacity In regard to the effects of antioxidant supple-ment on skeletal muscle autophagy we cannot ignore therole of mitochondria On one hand endogenous ROS ismainly generated by mitochondria on the other handmitochondrial antioxidant enzyme is able to scavenge ROSand reduce oxidative stress Recent works have suggestedthat mitochondrial-derived ROS induces autophagy [30 33]thus mitochondrial antioxidant capacity may be responsiblefor the activation of autophagy In the previous studiesNAC has been considered as a mitochondrial enhancerfor improving redox status and mitochondrial dysfunctionand reversing the progression of some diseases [37 38]Consistent with the previous results our data show that NACreduces mitochondrial ROS level and increases the ratio ofGSH GSSG suggesting that reduced ROS production andelevated GSH GSSG ratio in mitochondria are involved inthe inhibition of skeletal muscle constitutive autophagy Fur-ther NAC in part prevents fasting-induced oxidative stress inmitochondria but not autophagic responses indicating thatskeletal muscle autophagy under starvation is independentof mitochondrial-derived ROS In contrast to our results
in fasted mice lower nutrient supply or calorie restrictionleads to a lower production of mitochondrial ROS in skeletalmuscle [39] because the lower mitochondrial membranepotential favors reducing mitochondrial ROS production[40] In skeletal muscle of both rodents and humans high-fatdiet increases H
2O2emission of mitochondria [41] However
in response to acute nutrient deprivation mitochondrialROS levels significantly increase in skeletal muscle cells [30]These findings strongly indicate that ROS production inmitochondria is nutrient-sensitizing [42] It is noteworthythat chronic mild nutrient restriction differs from acutenutrient deprivation in the regulation of mitochondrial ROSproduction Thus the differences in constitutive and acutestress-induced autophagy may result from differential ROSlevels in skeletal muscle
Manganese superoxide dismutase (MnSOD) is a vitalantioxidant enzyme in the matrix of mitochondria by scav-enging ROS MnSOD knockdown led to chronic oxidativestress increased autophagy and mitochondrial biogenesiswithin the distal nephrons [43] In brain MnSOD knock-down increased oxidative stress and instead suppressedthe levels of autophagy stimulators [44] To our surprise
6 Oxidative Medicine and Cellular Longevity
LC3-I
TIGAR
LC3-II
Con
trol
Fast
NAC
NAC
+ F
ast
120573-Actin
(a)
0
05
1
15
2
Control Fast NAC
LC3
lipid
atio
n (C
ontro
l (
))
LC3-IILC3-I
NAC + Fast
lowastlowast
(b)
0
5
10
15
20
25
30
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
TIGARaTIGARb
lowastlowastlowastlowast
lowastlowast
(c)
Figure 4 LC3 lipidation and expressions of TIGAR were measured in skeletal muscle after short-term fasting (a) Representative westernblots of LC3 TIGAR (b) LC3-IILC3-I ratio (c) mRNA expression of TIGAR Values are means plusmn SE lowastlowast119875 lt 001 significantly differentversus Control
119875 lt 005 119875 lt 001 significantly different versus NAC
our results show that NAC decreases MnSODmRNA expres-sion and its enzyme activity CuZn-SOD expression is alsorepressed Fasting fails to rescue the decline in MnSODexpression Our data suggest that although NAC adminis-tration reduces ROS level and increases GSH GSSG exoge-nous antioxidant supplementmay lower inherent antioxidantcapacity in mitochondria ROS can lead to MnSOD upreg-ulation through ERK2 translocation into the nucleus anddissociation of p53 from its inhibitory protein mouse doubleminute 2 (MDM2) [45] Similarly heat stress producedROS in fibroblasts NAC blocked the increase in MnSODlevels by heat stress through reducing ROS production [46]Therefore a proper level of ROS is required for MnSODexpression Likewise we observe that TIGAR is repressedin mRNA and protein level by NAC Although TIGAR isnot an antioxidant enzyme that eliminates ROS directlyTIGAR plays an important antioxidant role in the regu-lation of autophagy [47] TIGAR inhibits glycolysis andpromotes respiration In TIGAR-expressing cells ROS levelwas redu-ced whereas glutathione was elevated [48] FurtherTIGAR inhibited autophagy by suppressing ROS level [47]TIGAR knockout increased mitophagy and ROS productionin myocardium followed by Bnip3 activation which is an
initiator of mitophagy Further NAC reversed the activationof Bnip3 and mitophagy [49] Under hypoxia mitochondriallocalization of TIGAR can control mitochondrial ROS levelsand protect from cell death [50] Therefore TIGAR is apotential antioxidant gene formitochondria Collectively ourresults indicate that NAC treatment with young mice atten-uates the inherent antioxidant capacity in skeletal musclesuggesting that ROS is also required for the maintenance ofmitochondrial antioxidant capacity
In summary our study provides the first evidencethat NAC decreases skeletal muscle constitutive autophagybut does not protect skeletal muscle from fasting-inducedautophagy in young mice In addition we also showed thatNAC attenuates the inherent antioxidant capacity in skeletalmuscle by depressing the expression of MnSOD and TIGARWe interpret these findings as an indication that ROS isrequired for the maintenance of both constitutive autophagyand antioxidant capacity in skeletal muscle Although NACsupplement is regarded as an effective intervention forimprovingmitochondrial function it should be reconsideredcarefully for young and healthy individuals when NACor the other antioxidants are used to promote musclegrowth
Oxidative Medicine and Cellular Longevity 7
0
20
40
60
80
100
120
140
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
SOD1 (CuZn-SOD)
lowastlowast
(a)
mRN
A le
vel (
vers
us 120573
-act
in)
0
50
100
150
200
250
Control Fast NAC NAC + Fast
SOD2 (MnSOD)
lowastlowast
(b)
0
02
04
06
08
1
12
14
Control Fast NAC NAC + Fast
Con
trol (
)
MnSOD activity
lowastlowast
(c)
Figure 5 Antioxidant capacity wasmeasured in gastrocnemiusmuscle after short-term fasting (a) mRNA expression of SOD1 (CuZn-SOD)(b) mRNA expression of SOD2 (MnSOD) (c) MnSOD activity in the isolated mitochondria of gastrocnemius muscle Values are means plusmnSE lowastlowast119875 lt 001 significantly different versus Control
119875 lt 005 significantly different versus NAC
4 Materials and Methods
41 Animals Male ICRCD-1 mice were purchased fromSino-British SipprBK Lab Animal Ltd CO (ShanghaiChina) at four to five weeks of age (Young 228 plusmn 12 g) Ani-mals were housed in a temperature-controlled environment(23 plusmn 2∘C) with 12 12 h light-dark cycles where they wereprovided diets and water ad libitum Mice were randomlyassigned to one of four experimental groups (1) Controlno drug treatment (Control 119899 = 8) (2) Fast no drugtreatment (Fast 119899 = 8) (3) Control treated with NAC (NAC119899 = 10) and (4) Fast treated with NAC (NAC + Fast119899 = 10) All experiments were performed in accordance withthe guidelines for the use of laboratory animals publishedby China Ministry of Health (no 55 order ordained on 25Jan 1998) All experimental procedureswere approved by theExperimental Animal Care andUse Committee at East ChinaNormal University (ECNU 2006-05)
42 Drugs and Short-TermFasting TheantioxidantN-acetyl-cysteine (NAC Sigma) was dissolved in DMSO The miceassigned to NAC groups received intraperitoneal injection ofNAC for 3weeks (100mgkg bodyweight2 day) As a controlthe vehicle (10 DMSO in saline) was injected to the controlmice For studies on the effects of food deprivation the mice
assigned to Fast groups were deprived of food for 24 hr beforesacrifice These mice had free access to drinking water
43 Mitochondrial Isolation At the end of the experimentalperiod animals in each group were acutely anesthetized withpentobarbital sodium Quadriceps gastrocnemius muscleand liver were removed completely and weighed For mito-chondrial isolation fresh gastrocnemius muscle was rinsedwith PBS and put into ice-cold isolation buffer (0075Msucrose 0225M sorbitol 1mM EGTA 01 fatty acid-freebovine serum albumin (BSA) and 10mM Tris-HCl pH 74)Tissues were sheared carefully to mince rinsed to get ridof residual blood and then homogenized in 1mL isolationbuffer per 100mg tissue The homogenate was centrifugedat 1000 g for 5min at 4∘C using a Beckman centrifuge(Avanti J-26XP) the resulting supernatant was decanted andsaved The pellet was washed once with isolation buffer Thesupernatant was combined and centrifuged at 9000 g for10min at 4∘C The mitochondrial pellet was washed andcentrifuged twice at 15000 g for 2min at 4∘C with isolationbuffer [51] Mitochondrial protein content was assayed usingBSA as a standard according to Bradford
44Mitochondrial ROSGeneration and Antioxidant CapacityThe dichlorofluorescin diacetate (DCF-DA) was used as
8 Oxidative Medicine and Cellular Longevity
a probe to detect mitochondrial ROS generation The assaywas performed on freshly isolated mitochondria Samples(30 120583g proteins) were incubated with 10 120583L of DCF-DA(10 120583molI) for 3 h at 37∘C The fluorescent product formedwas quantified by spectrofluorometer at the 485525 nmChanges in fluorescence were expressed as arbitrary unitTo assay mitochondrial lipid peroxidation MDA levels weremeasured using an assay kit according to the manufac-turerrsquos protocol (Nanjing Jiancheng Biotech Nanjing China)Briefly isolated mitochondria were incubated at 37∘C for 3 htreated with buffer and centrifuged at 10000timesg for 15minThe supernatants were incubated with thiobarbituric acid(TBA) and the absorbance of the supernatants wasmeasuredusing a spectrophotometer at a wavelength of 535 nm Alsothe GSHGSSG ratio and MnSOD activity were measuredusing an assay kit (Nanjing Jiancheng Biotech NanjingChina) according to themanufacturerrsquos protocol respectively
45 Caspases Activity Caspases 3 8 and 9 activity wasdetermined with an assay kit (Beyotime China) accordingto the manufacturerrsquos instructions Under the caspase-3acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) canbe changed into the yellow product p-nitroaniline (pNA)Under the caspase-8 acetyl-Ile-Glu-Thr-Asp p-nitroanilide(Ac-IETD-pNA) can be changed into the yellow prod-uct pNA Under the caspase-9 acetyl-Leu-Glu-His-Asp p-nitroanilide (Ac-LEHD-pNA) can be changed into the yellowproduct pNA Finally pNA content was measured usingthe spectrophotometer at an absorbance of 405 nm Proteincontent was assayed using BSA as a standard according toBradford
46 Glycogen Content Glycogen content was determinedwith an assay kit (Nanjing Jianchen Biotech China) accord-ing to the manufacturerrsquos instructions
47 Real-Time PCR Total RNA was prepared from sim100mgof frozen tissues using TRIzol (Invitrogen Chromos Singa-pore) and purified according to the instructions includedDouble-stranded cDNA was synthesized from sim1 120583g of totalRNA using ReverTra Ace qPCR RT kit (TOYOBO OsakaJapan) Real-time PCR reactions were set up using the SYBR-Green PCR kit (TOYOBO Osaka Japan) and were cycled inStepOne Real-Time PCR System (Applied Biosystems CAUSA) as previously described [52] The abundance of targetmRNA was normalized to that of 120573-actin Primer pairs weredesigned based on GenBank reference sequences and listedin Table S1 available online in Supplementary Material athttpdxdoiorg1011552014315896
48 Western Blotting The isolated muscle tissue (sim200mg)was weighed and cut into pieces at 4∘C and 2mL (10times dryweight of isolated muscle fibers) of ice-cold buffer (50mMTris HCl 150mMNaCl 1mM EDTA 02mM PMSF 1 NP-40 pH 74 cocktail) was added The tissue pieces were thenhomogenized after which homogenates were centrifuged for10min at 8000 g and 4∘C The total protein content of thesupernatant was quantified using bicinchoninic acid reagents
and BSA standards (Shanghai Sangon Shanghai China)Theprotein samples were immunoblotted as previously described[52] Visualization of reaction bands was performed by 331015840-diaminobenzidine staining (Shanghai Sangon China) andscanned densitometrically 120573-Actin was used to standardizefor the amount of protein loaded
49 Statistical Analysis Data are reported as mean plusmn SEMStatistical differences between treatments were determinedusing ANOVA For all tests the significance level was set at119875 lt 005
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from the National NaturalScience Foundation of China (Grant nos 31300977 and31171142) This work was also supported by the Key Labora-tory Construction Project of Adolescent Health Assessmentand Exercise Intervention of Ministry of Education EastChinaNormalUniversity (no 40500-541235-14203004)Theauthors also thank postgraduate students at Professor Dingrsquoslab for the technical support on animal care biochemicalassays PCR and western blot
References
[1] M Sandri ldquoAutophagy in skeletal musclerdquo FEBS Letters vol584 no 7 pp 1411ndash1416 2010
[2] G Twig B Hyde and O S Shirihai ldquoMitochondrial fusionfission and autophagy as a quality control axis the bioenergeticviewrdquo Biochimica et Biophysica Acta Bioenergetics vol 1777 no9 pp 1092ndash1097 2008
[3] Q Sun W Fan K Chen X Ding S Chen and Q ZhongldquoIdentification of Barkor as a mammalian autophagy-specificfactor for Beclin 1 and class III phosphatidylinositol 3-kinaserdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 49 pp 19211ndash19216 2008
[4] M C Maiuri E Zalckvar A Kimchi and G Kroemer ldquoSelf-eating and self-killing crosstalk between autophagy and apop-tosisrdquo Nature Reviews Molecular Cell Biology vol 8 no 9 pp741ndash752 2007
[5] T Ueno W Sato Y Horie et al ldquoLoss of Pten a tumor suppres-sor causes the strong inhibition of autophagy without affectingLC3 lipidationrdquo Autophagy vol 4 no 5 pp 692ndash700 2008
[6] F FortunatoH Burgers F Bergmann et al ldquoImpaired autolyso-some formation correlates with lamp-2 depletion role of apop-tosis autophagy and necrosis in pancreatitisrdquoGastroenterologyvol 137 no 1 pp 350ndash360 2009
[7] F A Graca D A Goncalves W A Silveira et al ldquoEpinephrinedepletion exacerbates the fasting-induced protein breakdown infast-twitch skeletal musclesrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 305 pp E1483ndashE14942013
[8] T Ogata Y Oishi M Higuchi and I Muraoka ldquoFasting-relatedautophagic response in slow- and fast-twitch skeletal musclerdquo
Oxidative Medicine and Cellular Longevity 9
Biochemical andBiophysical ResearchCommunications vol 394no 1 pp 136ndash140 2010
[9] A Doria M Gatto and L Punzi ldquoAutophagy in human healthand diseaserdquo New England Journal of Medicine vol 368 no 19p 1845 2013
[10] S Fulda ldquoAutophagy and cell deathrdquo Autophagy vol 8 no 8pp 1250ndash1251 2012
[11] E Masiero L Agatea C Mammucari et al ldquoAutophagy isrequired to maintain muscle massrdquo Cell Metabolism vol 10 no6 pp 507ndash515 2009
[12] A C Nascimbeni M Fanin E Masiero C Angelini and MSandri ldquoImpaired autophagy contributes to muscle atrophy inglycogen storage disease type II patientsrdquo Autophagy vol 8 no11 pp 1697ndash1700 2012
[13] M Sandri ldquoProtein breakdown in muscle wasting role ofautophagy-lysosome and ubiquitin-proteasomerdquo InternationalJournal of Biochemistry and Cell Biology vol 45 pp 2121ndash21292013
[14] E Masiero and M Sandri ldquoAutophagy inhibition inducesatrophy and myopathy in adult skeletal musclesrdquo Autophagyvol 6 no 2 pp 307ndash309 2010
[15] R Scherz-Shouval and Z Elazar ldquoRegulation of autophagy byROS physiology and pathologyrdquoTrends in Biochemical Sciencesvol 36 no 1 pp 30ndash38 2011
[16] C R Morales Z Pedrozo S Lavandero and J A Hill ldquoOxi-dative stress and autophagy in cardiovascular homeostasisrdquoAntioxidants amp Redox Signaling vol 20 pp 507ndash518 2014
[17] C Luo Y Li H Wang et al ldquoMitochondrial accumulationunder oxidative stress is due to defects in autophagyrdquo Journalof Cellular Biochemistry vol 114 no 1 pp 212ndash219 2013
[18] Y Wang Y Nartiss B Steipe G A McQuibban and PK Kim ldquoROS-induced mitochondrial depolarization initi-ates PARK2PARKIN-dependent mitochondrial degradationby autophagyrdquo Autophagy vol 8 no 10 pp 1462ndash1476 2012
[19] T Zhang Y Li K A Park et al ldquoCucurbitacin induces auto-phagy through mitochondrial ROS production which counter-acts to limit caspase-dependent apoptrdquo Autophagy vol 8 no 4pp 559ndash576 2012
[20] R Sandhir A Sood A Mehrotra and S S Kamboj ldquoN-acetyl-cysteine reverses mitochondrial dysfunctions and behavioralabnormalities in 3-nitropropionic acid-induced Huntingtonsdiseaserdquo Neurodegenerative Diseases vol 9 no 3 pp 145ndash1572012
[21] S S Kamboj and R Sandhir ldquoProtective effect of N-acetyl-cysteine supplementation onmitochondrial oxidative stress andmitochondrial enzymes in cerebral cortex of streptozotocin-treated diabetic ratsrdquoMitochondrion vol 11 no 1 pp 214ndash2222011
[22] M ZafarullahW Q Li J Sylvester andM Ahmad ldquoMolecularmechanisms of N-acetylcysteine actionsrdquo Cellular and Molecu-lar Life Sciences vol 60 no 1 pp 6ndash20 2003
[23] N A Strobel J M Peake A Matsumoto S A Marsh J SCoombes and G D Wadley ldquoAntioxidant supplementationreduces skeletal muscle mitochondrial biogenesisrdquo Medicineand Science in Sports and Exercise vol 43 no 6 pp 1017ndash10242011
[24] M-C Gomez-Cabrera E Domenech M Romagnoli et alldquoOral administration of vitamin C decreases muscle mitochon-drial biogenesis and hampers training-induced adaptationsin endurance performancerdquo The American Journal of ClinicalNutrition vol 87 no 1 pp 142ndash149 2008
[25] N P Whitehead C Pham O L Gervasio and D G Allen ldquoN-Acetylcysteine ameliorates skeletal muscle pathophysiology inmdx micerdquo Journal of Physiology vol 586 no 7 pp 2003ndash20142008
[26] C-H Lin S-C Kuo L-J Huang and P-W Gean ldquoNeuropro-tective effect ofN-acetylcysteine on neuronal apoptosis inducedby a synthetic gingerdione compound involvement of ERK andp38 phosphorylationrdquo Journal of Neuroscience Research vol 84no 7 pp 1485ndash1494 2006
[27] O BKotoulas S AKalamidas andD J Kondomerkos ldquoGlyco-gen autophagy in glucose homeostasisrdquo Pathology Research andPractice vol 202 no 9 pp 631ndash638 2006
[28] J Zirin J Nieuwenhuis and N Perrimon ldquoRole of autophagyin glycogen breakdown and its relevance to chloroquinemyopa-thyrdquo PLoS Biology vol 11 no 11 Article ID e1001708 2013
[29] P Castets S Lin N Rion et al ldquoSustained activation ofmTORC1 in skeletalmuscle inhibits constitutive and starvation-induced autophagy and causes a severe late-onset myopathyrdquoCell Metabolism vol 17 no 5 pp 731ndash744 2013
[30] M Rahman M Mofarrahi A S Kristof B Nkengfac S Hareland S N Hussain ldquoReactive oxygen species regulation ofautophagy in skeletal musclesrdquoAntioxidants amp Redox Signalingvol 20 no 3 pp 443ndash459 2014
[31] J Yan Z Feng J Liu et al ldquoEnhanced autophagy plays a cardi-nal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats ameliorating effects of (-)-epigallocatechin-3-gallaterdquo Journal of Nutritional Biochemistry vol 23 no 7 pp716ndash724 2012
[32] LV Yuzefovych S IMusiyenkoG LWilson and L I RachekldquoMitochondrial DNA damage and dysfunction and oxidativestress are associated with endoplasmic reticulum stress proteindegradation and apoptosis in high fat diet-induced InsulinresistancemicerdquoPLoSONE vol 8 no 1 Article ID e54059 2013
[33] L Li Y Chen and S B Gibson ldquoStarvation-induced autophagyis regulated bymitochondrial reactive oxygen species leading toAMPK activationrdquo Cellular Signalling vol 25 no 1 pp 50ndash652013
[34] S Takikita C Schreiner R Baum et al ldquoFiber type conversionby PGC-1120572 activates lysosomal and autophagosomal biogenesisin both unaffected and pompe skeletal musclerdquo PLoS ONE vol5 no 12 Article ID e15239 2010
[35] K Hollinger D Gardan-Salmon C Santana D Rice E Snellaand J T Selsby ldquoRescue of dystrophic skeletal muscle byPGC-1120572 involves restored expression of dystrophin-associatedprotein complex components and satellite cell signalingrdquo TheAmerican Journal of Physiology Regulatory Integrative andComparative Physiology vol 305 no 1 pp R13ndashR23 2013
[36] T Wenz S G Rossi R L Rotundo B M Spiegelman and CT Moraes ldquoIncreased muscle PGC-1120572 expression protects fromsarcopenia and metabolic disease during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 48 pp 20405ndash20410 2009
[37] I Grattagliano P Portincasa T Cocco et al ldquoEffect of dietaryrestriction and N-acetylcysteine supplementation on intestinalmucosa and liver mitochondrial redox status and function inaged ratsrdquo Experimental Gerontology vol 39 no 9 pp 1323ndash1332 2004
[38] S Kumar and S L Sitasawad ldquoN-acetylcysteine prevents glu-coseglucose oxidase-induced oxidative stress mitochondrialdamage and apoptosis in H9c2 cellsrdquo Life Sciences vol 84 no11-12 pp 328ndash336 2009
10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
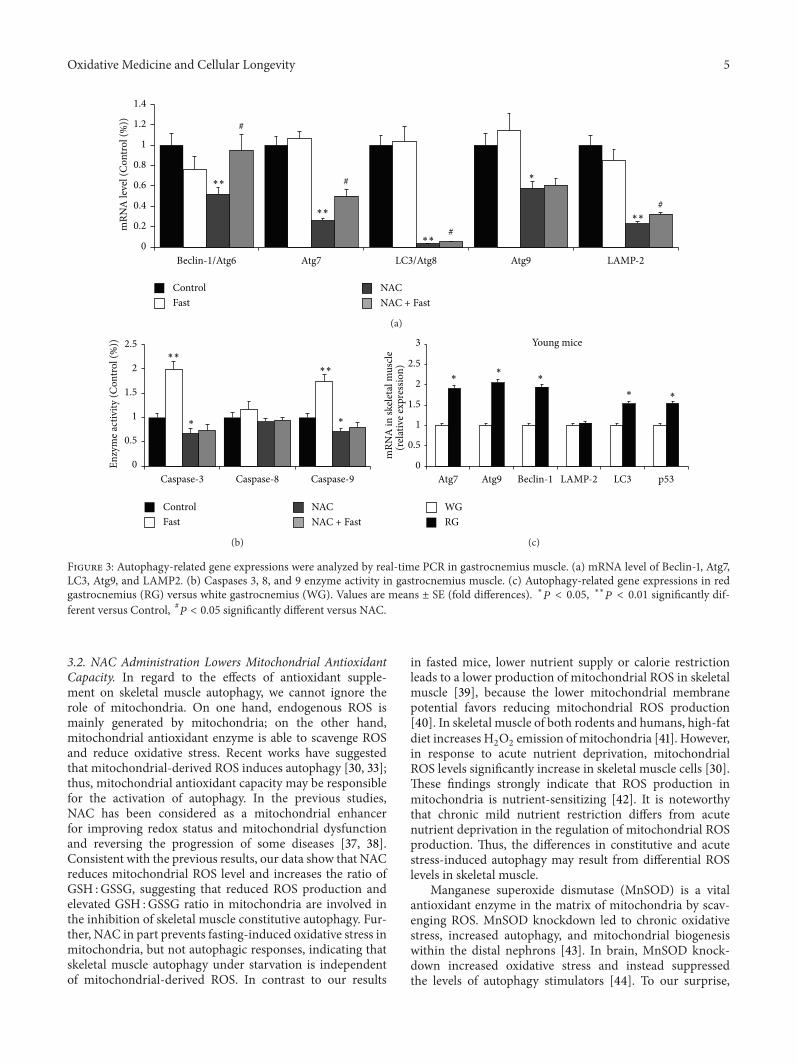
Oxidative Medicine and Cellular Longevity 5
0
02
04
06
08
1
12
14
Beclin-1Atg6 Atg7 LC3Atg8 Atg9 LAMP-2
mRN
A le
vel (
Con
trol (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowastlowastlowast
lowastlowastlowast
(a)
0
05
1
15
2
25
Caspase-3 Caspase-8 Caspase-9
Enzy
me a
ctiv
ity (C
ontro
l (
))
ControlFast
NACNAC + Fast
lowastlowast
lowastlowast
lowastlowast
(b)
0
05
1
15
2
25
3
mRN
A in
skel
etal
mus
cle(r
elat
ive e
xpre
ssio
n)
Young mice
Atg7 Atg9 Beclin-1 LAMP-2 LC3 p53
WGRG
lowastlowast
lowast
lowast lowast
(c)
Figure 3 Autophagy-related gene expressions were analyzed by real-time PCR in gastrocnemius muscle (a) mRNA level of Beclin-1 Atg7LC3 Atg9 and LAMP2 (b) Caspases 3 8 and 9 enzyme activity in gastrocnemius muscle (c) Autophagy-related gene expressions in redgastrocnemius (RG) versus white gastrocnemius (WG) Values are means plusmn SE (fold differences) lowast119875 lt 005 lowastlowast119875 lt 001 significantly dif-ferent versus Control
119875 lt 005 significantly different versus NAC
32 NAC Administration Lowers Mitochondrial AntioxidantCapacity In regard to the effects of antioxidant supple-ment on skeletal muscle autophagy we cannot ignore therole of mitochondria On one hand endogenous ROS ismainly generated by mitochondria on the other handmitochondrial antioxidant enzyme is able to scavenge ROSand reduce oxidative stress Recent works have suggestedthat mitochondrial-derived ROS induces autophagy [30 33]thus mitochondrial antioxidant capacity may be responsiblefor the activation of autophagy In the previous studiesNAC has been considered as a mitochondrial enhancerfor improving redox status and mitochondrial dysfunctionand reversing the progression of some diseases [37 38]Consistent with the previous results our data show that NACreduces mitochondrial ROS level and increases the ratio ofGSH GSSG suggesting that reduced ROS production andelevated GSH GSSG ratio in mitochondria are involved inthe inhibition of skeletal muscle constitutive autophagy Fur-ther NAC in part prevents fasting-induced oxidative stress inmitochondria but not autophagic responses indicating thatskeletal muscle autophagy under starvation is independentof mitochondrial-derived ROS In contrast to our results
in fasted mice lower nutrient supply or calorie restrictionleads to a lower production of mitochondrial ROS in skeletalmuscle [39] because the lower mitochondrial membranepotential favors reducing mitochondrial ROS production[40] In skeletal muscle of both rodents and humans high-fatdiet increases H
2O2emission of mitochondria [41] However
in response to acute nutrient deprivation mitochondrialROS levels significantly increase in skeletal muscle cells [30]These findings strongly indicate that ROS production inmitochondria is nutrient-sensitizing [42] It is noteworthythat chronic mild nutrient restriction differs from acutenutrient deprivation in the regulation of mitochondrial ROSproduction Thus the differences in constitutive and acutestress-induced autophagy may result from differential ROSlevels in skeletal muscle
Manganese superoxide dismutase (MnSOD) is a vitalantioxidant enzyme in the matrix of mitochondria by scav-enging ROS MnSOD knockdown led to chronic oxidativestress increased autophagy and mitochondrial biogenesiswithin the distal nephrons [43] In brain MnSOD knock-down increased oxidative stress and instead suppressedthe levels of autophagy stimulators [44] To our surprise
6 Oxidative Medicine and Cellular Longevity
LC3-I
TIGAR
LC3-II
Con
trol
Fast
NAC
NAC
+ F
ast
120573-Actin
(a)
0
05
1
15
2
Control Fast NAC
LC3
lipid
atio
n (C
ontro
l (
))
LC3-IILC3-I
NAC + Fast
lowastlowast
(b)
0
5
10
15
20
25
30
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
TIGARaTIGARb
lowastlowastlowastlowast
lowastlowast
(c)
Figure 4 LC3 lipidation and expressions of TIGAR were measured in skeletal muscle after short-term fasting (a) Representative westernblots of LC3 TIGAR (b) LC3-IILC3-I ratio (c) mRNA expression of TIGAR Values are means plusmn SE lowastlowast119875 lt 001 significantly differentversus Control
119875 lt 005 119875 lt 001 significantly different versus NAC
our results show that NAC decreases MnSODmRNA expres-sion and its enzyme activity CuZn-SOD expression is alsorepressed Fasting fails to rescue the decline in MnSODexpression Our data suggest that although NAC adminis-tration reduces ROS level and increases GSH GSSG exoge-nous antioxidant supplementmay lower inherent antioxidantcapacity in mitochondria ROS can lead to MnSOD upreg-ulation through ERK2 translocation into the nucleus anddissociation of p53 from its inhibitory protein mouse doubleminute 2 (MDM2) [45] Similarly heat stress producedROS in fibroblasts NAC blocked the increase in MnSODlevels by heat stress through reducing ROS production [46]Therefore a proper level of ROS is required for MnSODexpression Likewise we observe that TIGAR is repressedin mRNA and protein level by NAC Although TIGAR isnot an antioxidant enzyme that eliminates ROS directlyTIGAR plays an important antioxidant role in the regu-lation of autophagy [47] TIGAR inhibits glycolysis andpromotes respiration In TIGAR-expressing cells ROS levelwas redu-ced whereas glutathione was elevated [48] FurtherTIGAR inhibited autophagy by suppressing ROS level [47]TIGAR knockout increased mitophagy and ROS productionin myocardium followed by Bnip3 activation which is an
initiator of mitophagy Further NAC reversed the activationof Bnip3 and mitophagy [49] Under hypoxia mitochondriallocalization of TIGAR can control mitochondrial ROS levelsand protect from cell death [50] Therefore TIGAR is apotential antioxidant gene formitochondria Collectively ourresults indicate that NAC treatment with young mice atten-uates the inherent antioxidant capacity in skeletal musclesuggesting that ROS is also required for the maintenance ofmitochondrial antioxidant capacity
In summary our study provides the first evidencethat NAC decreases skeletal muscle constitutive autophagybut does not protect skeletal muscle from fasting-inducedautophagy in young mice In addition we also showed thatNAC attenuates the inherent antioxidant capacity in skeletalmuscle by depressing the expression of MnSOD and TIGARWe interpret these findings as an indication that ROS isrequired for the maintenance of both constitutive autophagyand antioxidant capacity in skeletal muscle Although NACsupplement is regarded as an effective intervention forimprovingmitochondrial function it should be reconsideredcarefully for young and healthy individuals when NACor the other antioxidants are used to promote musclegrowth
Oxidative Medicine and Cellular Longevity 7
0
20
40
60
80
100
120
140
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
SOD1 (CuZn-SOD)
lowastlowast
(a)
mRN
A le
vel (
vers
us 120573
-act
in)
0
50
100
150
200
250
Control Fast NAC NAC + Fast
SOD2 (MnSOD)
lowastlowast
(b)
0
02
04
06
08
1
12
14
Control Fast NAC NAC + Fast
Con
trol (
)
MnSOD activity
lowastlowast
(c)
Figure 5 Antioxidant capacity wasmeasured in gastrocnemiusmuscle after short-term fasting (a) mRNA expression of SOD1 (CuZn-SOD)(b) mRNA expression of SOD2 (MnSOD) (c) MnSOD activity in the isolated mitochondria of gastrocnemius muscle Values are means plusmnSE lowastlowast119875 lt 001 significantly different versus Control
119875 lt 005 significantly different versus NAC
4 Materials and Methods
41 Animals Male ICRCD-1 mice were purchased fromSino-British SipprBK Lab Animal Ltd CO (ShanghaiChina) at four to five weeks of age (Young 228 plusmn 12 g) Ani-mals were housed in a temperature-controlled environment(23 plusmn 2∘C) with 12 12 h light-dark cycles where they wereprovided diets and water ad libitum Mice were randomlyassigned to one of four experimental groups (1) Controlno drug treatment (Control 119899 = 8) (2) Fast no drugtreatment (Fast 119899 = 8) (3) Control treated with NAC (NAC119899 = 10) and (4) Fast treated with NAC (NAC + Fast119899 = 10) All experiments were performed in accordance withthe guidelines for the use of laboratory animals publishedby China Ministry of Health (no 55 order ordained on 25Jan 1998) All experimental procedureswere approved by theExperimental Animal Care andUse Committee at East ChinaNormal University (ECNU 2006-05)
42 Drugs and Short-TermFasting TheantioxidantN-acetyl-cysteine (NAC Sigma) was dissolved in DMSO The miceassigned to NAC groups received intraperitoneal injection ofNAC for 3weeks (100mgkg bodyweight2 day) As a controlthe vehicle (10 DMSO in saline) was injected to the controlmice For studies on the effects of food deprivation the mice
assigned to Fast groups were deprived of food for 24 hr beforesacrifice These mice had free access to drinking water
43 Mitochondrial Isolation At the end of the experimentalperiod animals in each group were acutely anesthetized withpentobarbital sodium Quadriceps gastrocnemius muscleand liver were removed completely and weighed For mito-chondrial isolation fresh gastrocnemius muscle was rinsedwith PBS and put into ice-cold isolation buffer (0075Msucrose 0225M sorbitol 1mM EGTA 01 fatty acid-freebovine serum albumin (BSA) and 10mM Tris-HCl pH 74)Tissues were sheared carefully to mince rinsed to get ridof residual blood and then homogenized in 1mL isolationbuffer per 100mg tissue The homogenate was centrifugedat 1000 g for 5min at 4∘C using a Beckman centrifuge(Avanti J-26XP) the resulting supernatant was decanted andsaved The pellet was washed once with isolation buffer Thesupernatant was combined and centrifuged at 9000 g for10min at 4∘C The mitochondrial pellet was washed andcentrifuged twice at 15000 g for 2min at 4∘C with isolationbuffer [51] Mitochondrial protein content was assayed usingBSA as a standard according to Bradford
44Mitochondrial ROSGeneration and Antioxidant CapacityThe dichlorofluorescin diacetate (DCF-DA) was used as
8 Oxidative Medicine and Cellular Longevity
a probe to detect mitochondrial ROS generation The assaywas performed on freshly isolated mitochondria Samples(30 120583g proteins) were incubated with 10 120583L of DCF-DA(10 120583molI) for 3 h at 37∘C The fluorescent product formedwas quantified by spectrofluorometer at the 485525 nmChanges in fluorescence were expressed as arbitrary unitTo assay mitochondrial lipid peroxidation MDA levels weremeasured using an assay kit according to the manufac-turerrsquos protocol (Nanjing Jiancheng Biotech Nanjing China)Briefly isolated mitochondria were incubated at 37∘C for 3 htreated with buffer and centrifuged at 10000timesg for 15minThe supernatants were incubated with thiobarbituric acid(TBA) and the absorbance of the supernatants wasmeasuredusing a spectrophotometer at a wavelength of 535 nm Alsothe GSHGSSG ratio and MnSOD activity were measuredusing an assay kit (Nanjing Jiancheng Biotech NanjingChina) according to themanufacturerrsquos protocol respectively
45 Caspases Activity Caspases 3 8 and 9 activity wasdetermined with an assay kit (Beyotime China) accordingto the manufacturerrsquos instructions Under the caspase-3acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) canbe changed into the yellow product p-nitroaniline (pNA)Under the caspase-8 acetyl-Ile-Glu-Thr-Asp p-nitroanilide(Ac-IETD-pNA) can be changed into the yellow prod-uct pNA Under the caspase-9 acetyl-Leu-Glu-His-Asp p-nitroanilide (Ac-LEHD-pNA) can be changed into the yellowproduct pNA Finally pNA content was measured usingthe spectrophotometer at an absorbance of 405 nm Proteincontent was assayed using BSA as a standard according toBradford
46 Glycogen Content Glycogen content was determinedwith an assay kit (Nanjing Jianchen Biotech China) accord-ing to the manufacturerrsquos instructions
47 Real-Time PCR Total RNA was prepared from sim100mgof frozen tissues using TRIzol (Invitrogen Chromos Singa-pore) and purified according to the instructions includedDouble-stranded cDNA was synthesized from sim1 120583g of totalRNA using ReverTra Ace qPCR RT kit (TOYOBO OsakaJapan) Real-time PCR reactions were set up using the SYBR-Green PCR kit (TOYOBO Osaka Japan) and were cycled inStepOne Real-Time PCR System (Applied Biosystems CAUSA) as previously described [52] The abundance of targetmRNA was normalized to that of 120573-actin Primer pairs weredesigned based on GenBank reference sequences and listedin Table S1 available online in Supplementary Material athttpdxdoiorg1011552014315896
48 Western Blotting The isolated muscle tissue (sim200mg)was weighed and cut into pieces at 4∘C and 2mL (10times dryweight of isolated muscle fibers) of ice-cold buffer (50mMTris HCl 150mMNaCl 1mM EDTA 02mM PMSF 1 NP-40 pH 74 cocktail) was added The tissue pieces were thenhomogenized after which homogenates were centrifuged for10min at 8000 g and 4∘C The total protein content of thesupernatant was quantified using bicinchoninic acid reagents
and BSA standards (Shanghai Sangon Shanghai China)Theprotein samples were immunoblotted as previously described[52] Visualization of reaction bands was performed by 331015840-diaminobenzidine staining (Shanghai Sangon China) andscanned densitometrically 120573-Actin was used to standardizefor the amount of protein loaded
49 Statistical Analysis Data are reported as mean plusmn SEMStatistical differences between treatments were determinedusing ANOVA For all tests the significance level was set at119875 lt 005
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from the National NaturalScience Foundation of China (Grant nos 31300977 and31171142) This work was also supported by the Key Labora-tory Construction Project of Adolescent Health Assessmentand Exercise Intervention of Ministry of Education EastChinaNormalUniversity (no 40500-541235-14203004)Theauthors also thank postgraduate students at Professor Dingrsquoslab for the technical support on animal care biochemicalassays PCR and western blot
References
[1] M Sandri ldquoAutophagy in skeletal musclerdquo FEBS Letters vol584 no 7 pp 1411ndash1416 2010
[2] G Twig B Hyde and O S Shirihai ldquoMitochondrial fusionfission and autophagy as a quality control axis the bioenergeticviewrdquo Biochimica et Biophysica Acta Bioenergetics vol 1777 no9 pp 1092ndash1097 2008
[3] Q Sun W Fan K Chen X Ding S Chen and Q ZhongldquoIdentification of Barkor as a mammalian autophagy-specificfactor for Beclin 1 and class III phosphatidylinositol 3-kinaserdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 49 pp 19211ndash19216 2008
[4] M C Maiuri E Zalckvar A Kimchi and G Kroemer ldquoSelf-eating and self-killing crosstalk between autophagy and apop-tosisrdquo Nature Reviews Molecular Cell Biology vol 8 no 9 pp741ndash752 2007
[5] T Ueno W Sato Y Horie et al ldquoLoss of Pten a tumor suppres-sor causes the strong inhibition of autophagy without affectingLC3 lipidationrdquo Autophagy vol 4 no 5 pp 692ndash700 2008
[6] F FortunatoH Burgers F Bergmann et al ldquoImpaired autolyso-some formation correlates with lamp-2 depletion role of apop-tosis autophagy and necrosis in pancreatitisrdquoGastroenterologyvol 137 no 1 pp 350ndash360 2009
[7] F A Graca D A Goncalves W A Silveira et al ldquoEpinephrinedepletion exacerbates the fasting-induced protein breakdown infast-twitch skeletal musclesrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 305 pp E1483ndashE14942013
[8] T Ogata Y Oishi M Higuchi and I Muraoka ldquoFasting-relatedautophagic response in slow- and fast-twitch skeletal musclerdquo
Oxidative Medicine and Cellular Longevity 9
Biochemical andBiophysical ResearchCommunications vol 394no 1 pp 136ndash140 2010
[9] A Doria M Gatto and L Punzi ldquoAutophagy in human healthand diseaserdquo New England Journal of Medicine vol 368 no 19p 1845 2013
[10] S Fulda ldquoAutophagy and cell deathrdquo Autophagy vol 8 no 8pp 1250ndash1251 2012
[11] E Masiero L Agatea C Mammucari et al ldquoAutophagy isrequired to maintain muscle massrdquo Cell Metabolism vol 10 no6 pp 507ndash515 2009
[12] A C Nascimbeni M Fanin E Masiero C Angelini and MSandri ldquoImpaired autophagy contributes to muscle atrophy inglycogen storage disease type II patientsrdquo Autophagy vol 8 no11 pp 1697ndash1700 2012
[13] M Sandri ldquoProtein breakdown in muscle wasting role ofautophagy-lysosome and ubiquitin-proteasomerdquo InternationalJournal of Biochemistry and Cell Biology vol 45 pp 2121ndash21292013
[14] E Masiero and M Sandri ldquoAutophagy inhibition inducesatrophy and myopathy in adult skeletal musclesrdquo Autophagyvol 6 no 2 pp 307ndash309 2010
[15] R Scherz-Shouval and Z Elazar ldquoRegulation of autophagy byROS physiology and pathologyrdquoTrends in Biochemical Sciencesvol 36 no 1 pp 30ndash38 2011
[16] C R Morales Z Pedrozo S Lavandero and J A Hill ldquoOxi-dative stress and autophagy in cardiovascular homeostasisrdquoAntioxidants amp Redox Signaling vol 20 pp 507ndash518 2014
[17] C Luo Y Li H Wang et al ldquoMitochondrial accumulationunder oxidative stress is due to defects in autophagyrdquo Journalof Cellular Biochemistry vol 114 no 1 pp 212ndash219 2013
[18] Y Wang Y Nartiss B Steipe G A McQuibban and PK Kim ldquoROS-induced mitochondrial depolarization initi-ates PARK2PARKIN-dependent mitochondrial degradationby autophagyrdquo Autophagy vol 8 no 10 pp 1462ndash1476 2012
[19] T Zhang Y Li K A Park et al ldquoCucurbitacin induces auto-phagy through mitochondrial ROS production which counter-acts to limit caspase-dependent apoptrdquo Autophagy vol 8 no 4pp 559ndash576 2012
[20] R Sandhir A Sood A Mehrotra and S S Kamboj ldquoN-acetyl-cysteine reverses mitochondrial dysfunctions and behavioralabnormalities in 3-nitropropionic acid-induced Huntingtonsdiseaserdquo Neurodegenerative Diseases vol 9 no 3 pp 145ndash1572012
[21] S S Kamboj and R Sandhir ldquoProtective effect of N-acetyl-cysteine supplementation onmitochondrial oxidative stress andmitochondrial enzymes in cerebral cortex of streptozotocin-treated diabetic ratsrdquoMitochondrion vol 11 no 1 pp 214ndash2222011
[22] M ZafarullahW Q Li J Sylvester andM Ahmad ldquoMolecularmechanisms of N-acetylcysteine actionsrdquo Cellular and Molecu-lar Life Sciences vol 60 no 1 pp 6ndash20 2003
[23] N A Strobel J M Peake A Matsumoto S A Marsh J SCoombes and G D Wadley ldquoAntioxidant supplementationreduces skeletal muscle mitochondrial biogenesisrdquo Medicineand Science in Sports and Exercise vol 43 no 6 pp 1017ndash10242011
[24] M-C Gomez-Cabrera E Domenech M Romagnoli et alldquoOral administration of vitamin C decreases muscle mitochon-drial biogenesis and hampers training-induced adaptationsin endurance performancerdquo The American Journal of ClinicalNutrition vol 87 no 1 pp 142ndash149 2008
[25] N P Whitehead C Pham O L Gervasio and D G Allen ldquoN-Acetylcysteine ameliorates skeletal muscle pathophysiology inmdx micerdquo Journal of Physiology vol 586 no 7 pp 2003ndash20142008
[26] C-H Lin S-C Kuo L-J Huang and P-W Gean ldquoNeuropro-tective effect ofN-acetylcysteine on neuronal apoptosis inducedby a synthetic gingerdione compound involvement of ERK andp38 phosphorylationrdquo Journal of Neuroscience Research vol 84no 7 pp 1485ndash1494 2006
[27] O BKotoulas S AKalamidas andD J Kondomerkos ldquoGlyco-gen autophagy in glucose homeostasisrdquo Pathology Research andPractice vol 202 no 9 pp 631ndash638 2006
[28] J Zirin J Nieuwenhuis and N Perrimon ldquoRole of autophagyin glycogen breakdown and its relevance to chloroquinemyopa-thyrdquo PLoS Biology vol 11 no 11 Article ID e1001708 2013
[29] P Castets S Lin N Rion et al ldquoSustained activation ofmTORC1 in skeletalmuscle inhibits constitutive and starvation-induced autophagy and causes a severe late-onset myopathyrdquoCell Metabolism vol 17 no 5 pp 731ndash744 2013
[30] M Rahman M Mofarrahi A S Kristof B Nkengfac S Hareland S N Hussain ldquoReactive oxygen species regulation ofautophagy in skeletal musclesrdquoAntioxidants amp Redox Signalingvol 20 no 3 pp 443ndash459 2014
[31] J Yan Z Feng J Liu et al ldquoEnhanced autophagy plays a cardi-nal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats ameliorating effects of (-)-epigallocatechin-3-gallaterdquo Journal of Nutritional Biochemistry vol 23 no 7 pp716ndash724 2012
[32] LV Yuzefovych S IMusiyenkoG LWilson and L I RachekldquoMitochondrial DNA damage and dysfunction and oxidativestress are associated with endoplasmic reticulum stress proteindegradation and apoptosis in high fat diet-induced InsulinresistancemicerdquoPLoSONE vol 8 no 1 Article ID e54059 2013
[33] L Li Y Chen and S B Gibson ldquoStarvation-induced autophagyis regulated bymitochondrial reactive oxygen species leading toAMPK activationrdquo Cellular Signalling vol 25 no 1 pp 50ndash652013
[34] S Takikita C Schreiner R Baum et al ldquoFiber type conversionby PGC-1120572 activates lysosomal and autophagosomal biogenesisin both unaffected and pompe skeletal musclerdquo PLoS ONE vol5 no 12 Article ID e15239 2010
[35] K Hollinger D Gardan-Salmon C Santana D Rice E Snellaand J T Selsby ldquoRescue of dystrophic skeletal muscle byPGC-1120572 involves restored expression of dystrophin-associatedprotein complex components and satellite cell signalingrdquo TheAmerican Journal of Physiology Regulatory Integrative andComparative Physiology vol 305 no 1 pp R13ndashR23 2013
[36] T Wenz S G Rossi R L Rotundo B M Spiegelman and CT Moraes ldquoIncreased muscle PGC-1120572 expression protects fromsarcopenia and metabolic disease during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 48 pp 20405ndash20410 2009
[37] I Grattagliano P Portincasa T Cocco et al ldquoEffect of dietaryrestriction and N-acetylcysteine supplementation on intestinalmucosa and liver mitochondrial redox status and function inaged ratsrdquo Experimental Gerontology vol 39 no 9 pp 1323ndash1332 2004
[38] S Kumar and S L Sitasawad ldquoN-acetylcysteine prevents glu-coseglucose oxidase-induced oxidative stress mitochondrialdamage and apoptosis in H9c2 cellsrdquo Life Sciences vol 84 no11-12 pp 328ndash336 2009
10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Oxidative Medicine and Cellular Longevity
LC3-I
TIGAR
LC3-II
Con
trol
Fast
NAC
NAC
+ F
ast
120573-Actin
(a)
0
05
1
15
2
Control Fast NAC
LC3
lipid
atio
n (C
ontro
l (
))
LC3-IILC3-I
NAC + Fast
lowastlowast
(b)
0
5
10
15
20
25
30
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
TIGARaTIGARb
lowastlowastlowastlowast
lowastlowast
(c)
Figure 4 LC3 lipidation and expressions of TIGAR were measured in skeletal muscle after short-term fasting (a) Representative westernblots of LC3 TIGAR (b) LC3-IILC3-I ratio (c) mRNA expression of TIGAR Values are means plusmn SE lowastlowast119875 lt 001 significantly differentversus Control
119875 lt 005 119875 lt 001 significantly different versus NAC
our results show that NAC decreases MnSODmRNA expres-sion and its enzyme activity CuZn-SOD expression is alsorepressed Fasting fails to rescue the decline in MnSODexpression Our data suggest that although NAC adminis-tration reduces ROS level and increases GSH GSSG exoge-nous antioxidant supplementmay lower inherent antioxidantcapacity in mitochondria ROS can lead to MnSOD upreg-ulation through ERK2 translocation into the nucleus anddissociation of p53 from its inhibitory protein mouse doubleminute 2 (MDM2) [45] Similarly heat stress producedROS in fibroblasts NAC blocked the increase in MnSODlevels by heat stress through reducing ROS production [46]Therefore a proper level of ROS is required for MnSODexpression Likewise we observe that TIGAR is repressedin mRNA and protein level by NAC Although TIGAR isnot an antioxidant enzyme that eliminates ROS directlyTIGAR plays an important antioxidant role in the regu-lation of autophagy [47] TIGAR inhibits glycolysis andpromotes respiration In TIGAR-expressing cells ROS levelwas redu-ced whereas glutathione was elevated [48] FurtherTIGAR inhibited autophagy by suppressing ROS level [47]TIGAR knockout increased mitophagy and ROS productionin myocardium followed by Bnip3 activation which is an
initiator of mitophagy Further NAC reversed the activationof Bnip3 and mitophagy [49] Under hypoxia mitochondriallocalization of TIGAR can control mitochondrial ROS levelsand protect from cell death [50] Therefore TIGAR is apotential antioxidant gene formitochondria Collectively ourresults indicate that NAC treatment with young mice atten-uates the inherent antioxidant capacity in skeletal musclesuggesting that ROS is also required for the maintenance ofmitochondrial antioxidant capacity
In summary our study provides the first evidencethat NAC decreases skeletal muscle constitutive autophagybut does not protect skeletal muscle from fasting-inducedautophagy in young mice In addition we also showed thatNAC attenuates the inherent antioxidant capacity in skeletalmuscle by depressing the expression of MnSOD and TIGARWe interpret these findings as an indication that ROS isrequired for the maintenance of both constitutive autophagyand antioxidant capacity in skeletal muscle Although NACsupplement is regarded as an effective intervention forimprovingmitochondrial function it should be reconsideredcarefully for young and healthy individuals when NACor the other antioxidants are used to promote musclegrowth
Oxidative Medicine and Cellular Longevity 7
0
20
40
60
80
100
120
140
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
SOD1 (CuZn-SOD)
lowastlowast
(a)
mRN
A le
vel (
vers
us 120573
-act
in)
0
50
100
150
200
250
Control Fast NAC NAC + Fast
SOD2 (MnSOD)
lowastlowast
(b)
0
02
04
06
08
1
12
14
Control Fast NAC NAC + Fast
Con
trol (
)
MnSOD activity
lowastlowast
(c)
Figure 5 Antioxidant capacity wasmeasured in gastrocnemiusmuscle after short-term fasting (a) mRNA expression of SOD1 (CuZn-SOD)(b) mRNA expression of SOD2 (MnSOD) (c) MnSOD activity in the isolated mitochondria of gastrocnemius muscle Values are means plusmnSE lowastlowast119875 lt 001 significantly different versus Control
119875 lt 005 significantly different versus NAC
4 Materials and Methods
41 Animals Male ICRCD-1 mice were purchased fromSino-British SipprBK Lab Animal Ltd CO (ShanghaiChina) at four to five weeks of age (Young 228 plusmn 12 g) Ani-mals were housed in a temperature-controlled environment(23 plusmn 2∘C) with 12 12 h light-dark cycles where they wereprovided diets and water ad libitum Mice were randomlyassigned to one of four experimental groups (1) Controlno drug treatment (Control 119899 = 8) (2) Fast no drugtreatment (Fast 119899 = 8) (3) Control treated with NAC (NAC119899 = 10) and (4) Fast treated with NAC (NAC + Fast119899 = 10) All experiments were performed in accordance withthe guidelines for the use of laboratory animals publishedby China Ministry of Health (no 55 order ordained on 25Jan 1998) All experimental procedureswere approved by theExperimental Animal Care andUse Committee at East ChinaNormal University (ECNU 2006-05)
42 Drugs and Short-TermFasting TheantioxidantN-acetyl-cysteine (NAC Sigma) was dissolved in DMSO The miceassigned to NAC groups received intraperitoneal injection ofNAC for 3weeks (100mgkg bodyweight2 day) As a controlthe vehicle (10 DMSO in saline) was injected to the controlmice For studies on the effects of food deprivation the mice
assigned to Fast groups were deprived of food for 24 hr beforesacrifice These mice had free access to drinking water
43 Mitochondrial Isolation At the end of the experimentalperiod animals in each group were acutely anesthetized withpentobarbital sodium Quadriceps gastrocnemius muscleand liver were removed completely and weighed For mito-chondrial isolation fresh gastrocnemius muscle was rinsedwith PBS and put into ice-cold isolation buffer (0075Msucrose 0225M sorbitol 1mM EGTA 01 fatty acid-freebovine serum albumin (BSA) and 10mM Tris-HCl pH 74)Tissues were sheared carefully to mince rinsed to get ridof residual blood and then homogenized in 1mL isolationbuffer per 100mg tissue The homogenate was centrifugedat 1000 g for 5min at 4∘C using a Beckman centrifuge(Avanti J-26XP) the resulting supernatant was decanted andsaved The pellet was washed once with isolation buffer Thesupernatant was combined and centrifuged at 9000 g for10min at 4∘C The mitochondrial pellet was washed andcentrifuged twice at 15000 g for 2min at 4∘C with isolationbuffer [51] Mitochondrial protein content was assayed usingBSA as a standard according to Bradford
44Mitochondrial ROSGeneration and Antioxidant CapacityThe dichlorofluorescin diacetate (DCF-DA) was used as
8 Oxidative Medicine and Cellular Longevity
a probe to detect mitochondrial ROS generation The assaywas performed on freshly isolated mitochondria Samples(30 120583g proteins) were incubated with 10 120583L of DCF-DA(10 120583molI) for 3 h at 37∘C The fluorescent product formedwas quantified by spectrofluorometer at the 485525 nmChanges in fluorescence were expressed as arbitrary unitTo assay mitochondrial lipid peroxidation MDA levels weremeasured using an assay kit according to the manufac-turerrsquos protocol (Nanjing Jiancheng Biotech Nanjing China)Briefly isolated mitochondria were incubated at 37∘C for 3 htreated with buffer and centrifuged at 10000timesg for 15minThe supernatants were incubated with thiobarbituric acid(TBA) and the absorbance of the supernatants wasmeasuredusing a spectrophotometer at a wavelength of 535 nm Alsothe GSHGSSG ratio and MnSOD activity were measuredusing an assay kit (Nanjing Jiancheng Biotech NanjingChina) according to themanufacturerrsquos protocol respectively
45 Caspases Activity Caspases 3 8 and 9 activity wasdetermined with an assay kit (Beyotime China) accordingto the manufacturerrsquos instructions Under the caspase-3acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) canbe changed into the yellow product p-nitroaniline (pNA)Under the caspase-8 acetyl-Ile-Glu-Thr-Asp p-nitroanilide(Ac-IETD-pNA) can be changed into the yellow prod-uct pNA Under the caspase-9 acetyl-Leu-Glu-His-Asp p-nitroanilide (Ac-LEHD-pNA) can be changed into the yellowproduct pNA Finally pNA content was measured usingthe spectrophotometer at an absorbance of 405 nm Proteincontent was assayed using BSA as a standard according toBradford
46 Glycogen Content Glycogen content was determinedwith an assay kit (Nanjing Jianchen Biotech China) accord-ing to the manufacturerrsquos instructions
47 Real-Time PCR Total RNA was prepared from sim100mgof frozen tissues using TRIzol (Invitrogen Chromos Singa-pore) and purified according to the instructions includedDouble-stranded cDNA was synthesized from sim1 120583g of totalRNA using ReverTra Ace qPCR RT kit (TOYOBO OsakaJapan) Real-time PCR reactions were set up using the SYBR-Green PCR kit (TOYOBO Osaka Japan) and were cycled inStepOne Real-Time PCR System (Applied Biosystems CAUSA) as previously described [52] The abundance of targetmRNA was normalized to that of 120573-actin Primer pairs weredesigned based on GenBank reference sequences and listedin Table S1 available online in Supplementary Material athttpdxdoiorg1011552014315896
48 Western Blotting The isolated muscle tissue (sim200mg)was weighed and cut into pieces at 4∘C and 2mL (10times dryweight of isolated muscle fibers) of ice-cold buffer (50mMTris HCl 150mMNaCl 1mM EDTA 02mM PMSF 1 NP-40 pH 74 cocktail) was added The tissue pieces were thenhomogenized after which homogenates were centrifuged for10min at 8000 g and 4∘C The total protein content of thesupernatant was quantified using bicinchoninic acid reagents
and BSA standards (Shanghai Sangon Shanghai China)Theprotein samples were immunoblotted as previously described[52] Visualization of reaction bands was performed by 331015840-diaminobenzidine staining (Shanghai Sangon China) andscanned densitometrically 120573-Actin was used to standardizefor the amount of protein loaded
49 Statistical Analysis Data are reported as mean plusmn SEMStatistical differences between treatments were determinedusing ANOVA For all tests the significance level was set at119875 lt 005
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from the National NaturalScience Foundation of China (Grant nos 31300977 and31171142) This work was also supported by the Key Labora-tory Construction Project of Adolescent Health Assessmentand Exercise Intervention of Ministry of Education EastChinaNormalUniversity (no 40500-541235-14203004)Theauthors also thank postgraduate students at Professor Dingrsquoslab for the technical support on animal care biochemicalassays PCR and western blot
References
[1] M Sandri ldquoAutophagy in skeletal musclerdquo FEBS Letters vol584 no 7 pp 1411ndash1416 2010
[2] G Twig B Hyde and O S Shirihai ldquoMitochondrial fusionfission and autophagy as a quality control axis the bioenergeticviewrdquo Biochimica et Biophysica Acta Bioenergetics vol 1777 no9 pp 1092ndash1097 2008
[3] Q Sun W Fan K Chen X Ding S Chen and Q ZhongldquoIdentification of Barkor as a mammalian autophagy-specificfactor for Beclin 1 and class III phosphatidylinositol 3-kinaserdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 49 pp 19211ndash19216 2008
[4] M C Maiuri E Zalckvar A Kimchi and G Kroemer ldquoSelf-eating and self-killing crosstalk between autophagy and apop-tosisrdquo Nature Reviews Molecular Cell Biology vol 8 no 9 pp741ndash752 2007
[5] T Ueno W Sato Y Horie et al ldquoLoss of Pten a tumor suppres-sor causes the strong inhibition of autophagy without affectingLC3 lipidationrdquo Autophagy vol 4 no 5 pp 692ndash700 2008
[6] F FortunatoH Burgers F Bergmann et al ldquoImpaired autolyso-some formation correlates with lamp-2 depletion role of apop-tosis autophagy and necrosis in pancreatitisrdquoGastroenterologyvol 137 no 1 pp 350ndash360 2009
[7] F A Graca D A Goncalves W A Silveira et al ldquoEpinephrinedepletion exacerbates the fasting-induced protein breakdown infast-twitch skeletal musclesrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 305 pp E1483ndashE14942013
[8] T Ogata Y Oishi M Higuchi and I Muraoka ldquoFasting-relatedautophagic response in slow- and fast-twitch skeletal musclerdquo
Oxidative Medicine and Cellular Longevity 9
Biochemical andBiophysical ResearchCommunications vol 394no 1 pp 136ndash140 2010
[9] A Doria M Gatto and L Punzi ldquoAutophagy in human healthand diseaserdquo New England Journal of Medicine vol 368 no 19p 1845 2013
[10] S Fulda ldquoAutophagy and cell deathrdquo Autophagy vol 8 no 8pp 1250ndash1251 2012
[11] E Masiero L Agatea C Mammucari et al ldquoAutophagy isrequired to maintain muscle massrdquo Cell Metabolism vol 10 no6 pp 507ndash515 2009
[12] A C Nascimbeni M Fanin E Masiero C Angelini and MSandri ldquoImpaired autophagy contributes to muscle atrophy inglycogen storage disease type II patientsrdquo Autophagy vol 8 no11 pp 1697ndash1700 2012
[13] M Sandri ldquoProtein breakdown in muscle wasting role ofautophagy-lysosome and ubiquitin-proteasomerdquo InternationalJournal of Biochemistry and Cell Biology vol 45 pp 2121ndash21292013
[14] E Masiero and M Sandri ldquoAutophagy inhibition inducesatrophy and myopathy in adult skeletal musclesrdquo Autophagyvol 6 no 2 pp 307ndash309 2010
[15] R Scherz-Shouval and Z Elazar ldquoRegulation of autophagy byROS physiology and pathologyrdquoTrends in Biochemical Sciencesvol 36 no 1 pp 30ndash38 2011
[16] C R Morales Z Pedrozo S Lavandero and J A Hill ldquoOxi-dative stress and autophagy in cardiovascular homeostasisrdquoAntioxidants amp Redox Signaling vol 20 pp 507ndash518 2014
[17] C Luo Y Li H Wang et al ldquoMitochondrial accumulationunder oxidative stress is due to defects in autophagyrdquo Journalof Cellular Biochemistry vol 114 no 1 pp 212ndash219 2013
[18] Y Wang Y Nartiss B Steipe G A McQuibban and PK Kim ldquoROS-induced mitochondrial depolarization initi-ates PARK2PARKIN-dependent mitochondrial degradationby autophagyrdquo Autophagy vol 8 no 10 pp 1462ndash1476 2012
[19] T Zhang Y Li K A Park et al ldquoCucurbitacin induces auto-phagy through mitochondrial ROS production which counter-acts to limit caspase-dependent apoptrdquo Autophagy vol 8 no 4pp 559ndash576 2012
[20] R Sandhir A Sood A Mehrotra and S S Kamboj ldquoN-acetyl-cysteine reverses mitochondrial dysfunctions and behavioralabnormalities in 3-nitropropionic acid-induced Huntingtonsdiseaserdquo Neurodegenerative Diseases vol 9 no 3 pp 145ndash1572012
[21] S S Kamboj and R Sandhir ldquoProtective effect of N-acetyl-cysteine supplementation onmitochondrial oxidative stress andmitochondrial enzymes in cerebral cortex of streptozotocin-treated diabetic ratsrdquoMitochondrion vol 11 no 1 pp 214ndash2222011
[22] M ZafarullahW Q Li J Sylvester andM Ahmad ldquoMolecularmechanisms of N-acetylcysteine actionsrdquo Cellular and Molecu-lar Life Sciences vol 60 no 1 pp 6ndash20 2003
[23] N A Strobel J M Peake A Matsumoto S A Marsh J SCoombes and G D Wadley ldquoAntioxidant supplementationreduces skeletal muscle mitochondrial biogenesisrdquo Medicineand Science in Sports and Exercise vol 43 no 6 pp 1017ndash10242011
[24] M-C Gomez-Cabrera E Domenech M Romagnoli et alldquoOral administration of vitamin C decreases muscle mitochon-drial biogenesis and hampers training-induced adaptationsin endurance performancerdquo The American Journal of ClinicalNutrition vol 87 no 1 pp 142ndash149 2008
[25] N P Whitehead C Pham O L Gervasio and D G Allen ldquoN-Acetylcysteine ameliorates skeletal muscle pathophysiology inmdx micerdquo Journal of Physiology vol 586 no 7 pp 2003ndash20142008
[26] C-H Lin S-C Kuo L-J Huang and P-W Gean ldquoNeuropro-tective effect ofN-acetylcysteine on neuronal apoptosis inducedby a synthetic gingerdione compound involvement of ERK andp38 phosphorylationrdquo Journal of Neuroscience Research vol 84no 7 pp 1485ndash1494 2006
[27] O BKotoulas S AKalamidas andD J Kondomerkos ldquoGlyco-gen autophagy in glucose homeostasisrdquo Pathology Research andPractice vol 202 no 9 pp 631ndash638 2006
[28] J Zirin J Nieuwenhuis and N Perrimon ldquoRole of autophagyin glycogen breakdown and its relevance to chloroquinemyopa-thyrdquo PLoS Biology vol 11 no 11 Article ID e1001708 2013
[29] P Castets S Lin N Rion et al ldquoSustained activation ofmTORC1 in skeletalmuscle inhibits constitutive and starvation-induced autophagy and causes a severe late-onset myopathyrdquoCell Metabolism vol 17 no 5 pp 731ndash744 2013
[30] M Rahman M Mofarrahi A S Kristof B Nkengfac S Hareland S N Hussain ldquoReactive oxygen species regulation ofautophagy in skeletal musclesrdquoAntioxidants amp Redox Signalingvol 20 no 3 pp 443ndash459 2014
[31] J Yan Z Feng J Liu et al ldquoEnhanced autophagy plays a cardi-nal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats ameliorating effects of (-)-epigallocatechin-3-gallaterdquo Journal of Nutritional Biochemistry vol 23 no 7 pp716ndash724 2012
[32] LV Yuzefovych S IMusiyenkoG LWilson and L I RachekldquoMitochondrial DNA damage and dysfunction and oxidativestress are associated with endoplasmic reticulum stress proteindegradation and apoptosis in high fat diet-induced InsulinresistancemicerdquoPLoSONE vol 8 no 1 Article ID e54059 2013
[33] L Li Y Chen and S B Gibson ldquoStarvation-induced autophagyis regulated bymitochondrial reactive oxygen species leading toAMPK activationrdquo Cellular Signalling vol 25 no 1 pp 50ndash652013
[34] S Takikita C Schreiner R Baum et al ldquoFiber type conversionby PGC-1120572 activates lysosomal and autophagosomal biogenesisin both unaffected and pompe skeletal musclerdquo PLoS ONE vol5 no 12 Article ID e15239 2010
[35] K Hollinger D Gardan-Salmon C Santana D Rice E Snellaand J T Selsby ldquoRescue of dystrophic skeletal muscle byPGC-1120572 involves restored expression of dystrophin-associatedprotein complex components and satellite cell signalingrdquo TheAmerican Journal of Physiology Regulatory Integrative andComparative Physiology vol 305 no 1 pp R13ndashR23 2013
[36] T Wenz S G Rossi R L Rotundo B M Spiegelman and CT Moraes ldquoIncreased muscle PGC-1120572 expression protects fromsarcopenia and metabolic disease during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 48 pp 20405ndash20410 2009
[37] I Grattagliano P Portincasa T Cocco et al ldquoEffect of dietaryrestriction and N-acetylcysteine supplementation on intestinalmucosa and liver mitochondrial redox status and function inaged ratsrdquo Experimental Gerontology vol 39 no 9 pp 1323ndash1332 2004
[38] S Kumar and S L Sitasawad ldquoN-acetylcysteine prevents glu-coseglucose oxidase-induced oxidative stress mitochondrialdamage and apoptosis in H9c2 cellsrdquo Life Sciences vol 84 no11-12 pp 328ndash336 2009
10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 7
0
20
40
60
80
100
120
140
Control Fast NAC NAC + Fast
mRN
A le
vel (
vers
us 120573
-act
in)
SOD1 (CuZn-SOD)
lowastlowast
(a)
mRN
A le
vel (
vers
us 120573
-act
in)
0
50
100
150
200
250
Control Fast NAC NAC + Fast
SOD2 (MnSOD)
lowastlowast
(b)
0
02
04
06
08
1
12
14
Control Fast NAC NAC + Fast
Con
trol (
)
MnSOD activity
lowastlowast
(c)
Figure 5 Antioxidant capacity wasmeasured in gastrocnemiusmuscle after short-term fasting (a) mRNA expression of SOD1 (CuZn-SOD)(b) mRNA expression of SOD2 (MnSOD) (c) MnSOD activity in the isolated mitochondria of gastrocnemius muscle Values are means plusmnSE lowastlowast119875 lt 001 significantly different versus Control
119875 lt 005 significantly different versus NAC
4 Materials and Methods
41 Animals Male ICRCD-1 mice were purchased fromSino-British SipprBK Lab Animal Ltd CO (ShanghaiChina) at four to five weeks of age (Young 228 plusmn 12 g) Ani-mals were housed in a temperature-controlled environment(23 plusmn 2∘C) with 12 12 h light-dark cycles where they wereprovided diets and water ad libitum Mice were randomlyassigned to one of four experimental groups (1) Controlno drug treatment (Control 119899 = 8) (2) Fast no drugtreatment (Fast 119899 = 8) (3) Control treated with NAC (NAC119899 = 10) and (4) Fast treated with NAC (NAC + Fast119899 = 10) All experiments were performed in accordance withthe guidelines for the use of laboratory animals publishedby China Ministry of Health (no 55 order ordained on 25Jan 1998) All experimental procedureswere approved by theExperimental Animal Care andUse Committee at East ChinaNormal University (ECNU 2006-05)
42 Drugs and Short-TermFasting TheantioxidantN-acetyl-cysteine (NAC Sigma) was dissolved in DMSO The miceassigned to NAC groups received intraperitoneal injection ofNAC for 3weeks (100mgkg bodyweight2 day) As a controlthe vehicle (10 DMSO in saline) was injected to the controlmice For studies on the effects of food deprivation the mice
assigned to Fast groups were deprived of food for 24 hr beforesacrifice These mice had free access to drinking water
43 Mitochondrial Isolation At the end of the experimentalperiod animals in each group were acutely anesthetized withpentobarbital sodium Quadriceps gastrocnemius muscleand liver were removed completely and weighed For mito-chondrial isolation fresh gastrocnemius muscle was rinsedwith PBS and put into ice-cold isolation buffer (0075Msucrose 0225M sorbitol 1mM EGTA 01 fatty acid-freebovine serum albumin (BSA) and 10mM Tris-HCl pH 74)Tissues were sheared carefully to mince rinsed to get ridof residual blood and then homogenized in 1mL isolationbuffer per 100mg tissue The homogenate was centrifugedat 1000 g for 5min at 4∘C using a Beckman centrifuge(Avanti J-26XP) the resulting supernatant was decanted andsaved The pellet was washed once with isolation buffer Thesupernatant was combined and centrifuged at 9000 g for10min at 4∘C The mitochondrial pellet was washed andcentrifuged twice at 15000 g for 2min at 4∘C with isolationbuffer [51] Mitochondrial protein content was assayed usingBSA as a standard according to Bradford
44Mitochondrial ROSGeneration and Antioxidant CapacityThe dichlorofluorescin diacetate (DCF-DA) was used as
8 Oxidative Medicine and Cellular Longevity
a probe to detect mitochondrial ROS generation The assaywas performed on freshly isolated mitochondria Samples(30 120583g proteins) were incubated with 10 120583L of DCF-DA(10 120583molI) for 3 h at 37∘C The fluorescent product formedwas quantified by spectrofluorometer at the 485525 nmChanges in fluorescence were expressed as arbitrary unitTo assay mitochondrial lipid peroxidation MDA levels weremeasured using an assay kit according to the manufac-turerrsquos protocol (Nanjing Jiancheng Biotech Nanjing China)Briefly isolated mitochondria were incubated at 37∘C for 3 htreated with buffer and centrifuged at 10000timesg for 15minThe supernatants were incubated with thiobarbituric acid(TBA) and the absorbance of the supernatants wasmeasuredusing a spectrophotometer at a wavelength of 535 nm Alsothe GSHGSSG ratio and MnSOD activity were measuredusing an assay kit (Nanjing Jiancheng Biotech NanjingChina) according to themanufacturerrsquos protocol respectively
45 Caspases Activity Caspases 3 8 and 9 activity wasdetermined with an assay kit (Beyotime China) accordingto the manufacturerrsquos instructions Under the caspase-3acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) canbe changed into the yellow product p-nitroaniline (pNA)Under the caspase-8 acetyl-Ile-Glu-Thr-Asp p-nitroanilide(Ac-IETD-pNA) can be changed into the yellow prod-uct pNA Under the caspase-9 acetyl-Leu-Glu-His-Asp p-nitroanilide (Ac-LEHD-pNA) can be changed into the yellowproduct pNA Finally pNA content was measured usingthe spectrophotometer at an absorbance of 405 nm Proteincontent was assayed using BSA as a standard according toBradford
46 Glycogen Content Glycogen content was determinedwith an assay kit (Nanjing Jianchen Biotech China) accord-ing to the manufacturerrsquos instructions
47 Real-Time PCR Total RNA was prepared from sim100mgof frozen tissues using TRIzol (Invitrogen Chromos Singa-pore) and purified according to the instructions includedDouble-stranded cDNA was synthesized from sim1 120583g of totalRNA using ReverTra Ace qPCR RT kit (TOYOBO OsakaJapan) Real-time PCR reactions were set up using the SYBR-Green PCR kit (TOYOBO Osaka Japan) and were cycled inStepOne Real-Time PCR System (Applied Biosystems CAUSA) as previously described [52] The abundance of targetmRNA was normalized to that of 120573-actin Primer pairs weredesigned based on GenBank reference sequences and listedin Table S1 available online in Supplementary Material athttpdxdoiorg1011552014315896
48 Western Blotting The isolated muscle tissue (sim200mg)was weighed and cut into pieces at 4∘C and 2mL (10times dryweight of isolated muscle fibers) of ice-cold buffer (50mMTris HCl 150mMNaCl 1mM EDTA 02mM PMSF 1 NP-40 pH 74 cocktail) was added The tissue pieces were thenhomogenized after which homogenates were centrifuged for10min at 8000 g and 4∘C The total protein content of thesupernatant was quantified using bicinchoninic acid reagents
and BSA standards (Shanghai Sangon Shanghai China)Theprotein samples were immunoblotted as previously described[52] Visualization of reaction bands was performed by 331015840-diaminobenzidine staining (Shanghai Sangon China) andscanned densitometrically 120573-Actin was used to standardizefor the amount of protein loaded
49 Statistical Analysis Data are reported as mean plusmn SEMStatistical differences between treatments were determinedusing ANOVA For all tests the significance level was set at119875 lt 005
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from the National NaturalScience Foundation of China (Grant nos 31300977 and31171142) This work was also supported by the Key Labora-tory Construction Project of Adolescent Health Assessmentand Exercise Intervention of Ministry of Education EastChinaNormalUniversity (no 40500-541235-14203004)Theauthors also thank postgraduate students at Professor Dingrsquoslab for the technical support on animal care biochemicalassays PCR and western blot
References
[1] M Sandri ldquoAutophagy in skeletal musclerdquo FEBS Letters vol584 no 7 pp 1411ndash1416 2010
[2] G Twig B Hyde and O S Shirihai ldquoMitochondrial fusionfission and autophagy as a quality control axis the bioenergeticviewrdquo Biochimica et Biophysica Acta Bioenergetics vol 1777 no9 pp 1092ndash1097 2008
[3] Q Sun W Fan K Chen X Ding S Chen and Q ZhongldquoIdentification of Barkor as a mammalian autophagy-specificfactor for Beclin 1 and class III phosphatidylinositol 3-kinaserdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 49 pp 19211ndash19216 2008
[4] M C Maiuri E Zalckvar A Kimchi and G Kroemer ldquoSelf-eating and self-killing crosstalk between autophagy and apop-tosisrdquo Nature Reviews Molecular Cell Biology vol 8 no 9 pp741ndash752 2007
[5] T Ueno W Sato Y Horie et al ldquoLoss of Pten a tumor suppres-sor causes the strong inhibition of autophagy without affectingLC3 lipidationrdquo Autophagy vol 4 no 5 pp 692ndash700 2008
[6] F FortunatoH Burgers F Bergmann et al ldquoImpaired autolyso-some formation correlates with lamp-2 depletion role of apop-tosis autophagy and necrosis in pancreatitisrdquoGastroenterologyvol 137 no 1 pp 350ndash360 2009
[7] F A Graca D A Goncalves W A Silveira et al ldquoEpinephrinedepletion exacerbates the fasting-induced protein breakdown infast-twitch skeletal musclesrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 305 pp E1483ndashE14942013
[8] T Ogata Y Oishi M Higuchi and I Muraoka ldquoFasting-relatedautophagic response in slow- and fast-twitch skeletal musclerdquo
Oxidative Medicine and Cellular Longevity 9
Biochemical andBiophysical ResearchCommunications vol 394no 1 pp 136ndash140 2010
[9] A Doria M Gatto and L Punzi ldquoAutophagy in human healthand diseaserdquo New England Journal of Medicine vol 368 no 19p 1845 2013
[10] S Fulda ldquoAutophagy and cell deathrdquo Autophagy vol 8 no 8pp 1250ndash1251 2012
[11] E Masiero L Agatea C Mammucari et al ldquoAutophagy isrequired to maintain muscle massrdquo Cell Metabolism vol 10 no6 pp 507ndash515 2009
[12] A C Nascimbeni M Fanin E Masiero C Angelini and MSandri ldquoImpaired autophagy contributes to muscle atrophy inglycogen storage disease type II patientsrdquo Autophagy vol 8 no11 pp 1697ndash1700 2012
[13] M Sandri ldquoProtein breakdown in muscle wasting role ofautophagy-lysosome and ubiquitin-proteasomerdquo InternationalJournal of Biochemistry and Cell Biology vol 45 pp 2121ndash21292013
[14] E Masiero and M Sandri ldquoAutophagy inhibition inducesatrophy and myopathy in adult skeletal musclesrdquo Autophagyvol 6 no 2 pp 307ndash309 2010
[15] R Scherz-Shouval and Z Elazar ldquoRegulation of autophagy byROS physiology and pathologyrdquoTrends in Biochemical Sciencesvol 36 no 1 pp 30ndash38 2011
[16] C R Morales Z Pedrozo S Lavandero and J A Hill ldquoOxi-dative stress and autophagy in cardiovascular homeostasisrdquoAntioxidants amp Redox Signaling vol 20 pp 507ndash518 2014
[17] C Luo Y Li H Wang et al ldquoMitochondrial accumulationunder oxidative stress is due to defects in autophagyrdquo Journalof Cellular Biochemistry vol 114 no 1 pp 212ndash219 2013
[18] Y Wang Y Nartiss B Steipe G A McQuibban and PK Kim ldquoROS-induced mitochondrial depolarization initi-ates PARK2PARKIN-dependent mitochondrial degradationby autophagyrdquo Autophagy vol 8 no 10 pp 1462ndash1476 2012
[19] T Zhang Y Li K A Park et al ldquoCucurbitacin induces auto-phagy through mitochondrial ROS production which counter-acts to limit caspase-dependent apoptrdquo Autophagy vol 8 no 4pp 559ndash576 2012
[20] R Sandhir A Sood A Mehrotra and S S Kamboj ldquoN-acetyl-cysteine reverses mitochondrial dysfunctions and behavioralabnormalities in 3-nitropropionic acid-induced Huntingtonsdiseaserdquo Neurodegenerative Diseases vol 9 no 3 pp 145ndash1572012
[21] S S Kamboj and R Sandhir ldquoProtective effect of N-acetyl-cysteine supplementation onmitochondrial oxidative stress andmitochondrial enzymes in cerebral cortex of streptozotocin-treated diabetic ratsrdquoMitochondrion vol 11 no 1 pp 214ndash2222011
[22] M ZafarullahW Q Li J Sylvester andM Ahmad ldquoMolecularmechanisms of N-acetylcysteine actionsrdquo Cellular and Molecu-lar Life Sciences vol 60 no 1 pp 6ndash20 2003
[23] N A Strobel J M Peake A Matsumoto S A Marsh J SCoombes and G D Wadley ldquoAntioxidant supplementationreduces skeletal muscle mitochondrial biogenesisrdquo Medicineand Science in Sports and Exercise vol 43 no 6 pp 1017ndash10242011
[24] M-C Gomez-Cabrera E Domenech M Romagnoli et alldquoOral administration of vitamin C decreases muscle mitochon-drial biogenesis and hampers training-induced adaptationsin endurance performancerdquo The American Journal of ClinicalNutrition vol 87 no 1 pp 142ndash149 2008
[25] N P Whitehead C Pham O L Gervasio and D G Allen ldquoN-Acetylcysteine ameliorates skeletal muscle pathophysiology inmdx micerdquo Journal of Physiology vol 586 no 7 pp 2003ndash20142008
[26] C-H Lin S-C Kuo L-J Huang and P-W Gean ldquoNeuropro-tective effect ofN-acetylcysteine on neuronal apoptosis inducedby a synthetic gingerdione compound involvement of ERK andp38 phosphorylationrdquo Journal of Neuroscience Research vol 84no 7 pp 1485ndash1494 2006
[27] O BKotoulas S AKalamidas andD J Kondomerkos ldquoGlyco-gen autophagy in glucose homeostasisrdquo Pathology Research andPractice vol 202 no 9 pp 631ndash638 2006
[28] J Zirin J Nieuwenhuis and N Perrimon ldquoRole of autophagyin glycogen breakdown and its relevance to chloroquinemyopa-thyrdquo PLoS Biology vol 11 no 11 Article ID e1001708 2013
[29] P Castets S Lin N Rion et al ldquoSustained activation ofmTORC1 in skeletalmuscle inhibits constitutive and starvation-induced autophagy and causes a severe late-onset myopathyrdquoCell Metabolism vol 17 no 5 pp 731ndash744 2013
[30] M Rahman M Mofarrahi A S Kristof B Nkengfac S Hareland S N Hussain ldquoReactive oxygen species regulation ofautophagy in skeletal musclesrdquoAntioxidants amp Redox Signalingvol 20 no 3 pp 443ndash459 2014
[31] J Yan Z Feng J Liu et al ldquoEnhanced autophagy plays a cardi-nal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats ameliorating effects of (-)-epigallocatechin-3-gallaterdquo Journal of Nutritional Biochemistry vol 23 no 7 pp716ndash724 2012
[32] LV Yuzefovych S IMusiyenkoG LWilson and L I RachekldquoMitochondrial DNA damage and dysfunction and oxidativestress are associated with endoplasmic reticulum stress proteindegradation and apoptosis in high fat diet-induced InsulinresistancemicerdquoPLoSONE vol 8 no 1 Article ID e54059 2013
[33] L Li Y Chen and S B Gibson ldquoStarvation-induced autophagyis regulated bymitochondrial reactive oxygen species leading toAMPK activationrdquo Cellular Signalling vol 25 no 1 pp 50ndash652013
[34] S Takikita C Schreiner R Baum et al ldquoFiber type conversionby PGC-1120572 activates lysosomal and autophagosomal biogenesisin both unaffected and pompe skeletal musclerdquo PLoS ONE vol5 no 12 Article ID e15239 2010
[35] K Hollinger D Gardan-Salmon C Santana D Rice E Snellaand J T Selsby ldquoRescue of dystrophic skeletal muscle byPGC-1120572 involves restored expression of dystrophin-associatedprotein complex components and satellite cell signalingrdquo TheAmerican Journal of Physiology Regulatory Integrative andComparative Physiology vol 305 no 1 pp R13ndashR23 2013
[36] T Wenz S G Rossi R L Rotundo B M Spiegelman and CT Moraes ldquoIncreased muscle PGC-1120572 expression protects fromsarcopenia and metabolic disease during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 48 pp 20405ndash20410 2009
[37] I Grattagliano P Portincasa T Cocco et al ldquoEffect of dietaryrestriction and N-acetylcysteine supplementation on intestinalmucosa and liver mitochondrial redox status and function inaged ratsrdquo Experimental Gerontology vol 39 no 9 pp 1323ndash1332 2004
[38] S Kumar and S L Sitasawad ldquoN-acetylcysteine prevents glu-coseglucose oxidase-induced oxidative stress mitochondrialdamage and apoptosis in H9c2 cellsrdquo Life Sciences vol 84 no11-12 pp 328ndash336 2009
10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Oxidative Medicine and Cellular Longevity
a probe to detect mitochondrial ROS generation The assaywas performed on freshly isolated mitochondria Samples(30 120583g proteins) were incubated with 10 120583L of DCF-DA(10 120583molI) for 3 h at 37∘C The fluorescent product formedwas quantified by spectrofluorometer at the 485525 nmChanges in fluorescence were expressed as arbitrary unitTo assay mitochondrial lipid peroxidation MDA levels weremeasured using an assay kit according to the manufac-turerrsquos protocol (Nanjing Jiancheng Biotech Nanjing China)Briefly isolated mitochondria were incubated at 37∘C for 3 htreated with buffer and centrifuged at 10000timesg for 15minThe supernatants were incubated with thiobarbituric acid(TBA) and the absorbance of the supernatants wasmeasuredusing a spectrophotometer at a wavelength of 535 nm Alsothe GSHGSSG ratio and MnSOD activity were measuredusing an assay kit (Nanjing Jiancheng Biotech NanjingChina) according to themanufacturerrsquos protocol respectively
45 Caspases Activity Caspases 3 8 and 9 activity wasdetermined with an assay kit (Beyotime China) accordingto the manufacturerrsquos instructions Under the caspase-3acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) canbe changed into the yellow product p-nitroaniline (pNA)Under the caspase-8 acetyl-Ile-Glu-Thr-Asp p-nitroanilide(Ac-IETD-pNA) can be changed into the yellow prod-uct pNA Under the caspase-9 acetyl-Leu-Glu-His-Asp p-nitroanilide (Ac-LEHD-pNA) can be changed into the yellowproduct pNA Finally pNA content was measured usingthe spectrophotometer at an absorbance of 405 nm Proteincontent was assayed using BSA as a standard according toBradford
46 Glycogen Content Glycogen content was determinedwith an assay kit (Nanjing Jianchen Biotech China) accord-ing to the manufacturerrsquos instructions
47 Real-Time PCR Total RNA was prepared from sim100mgof frozen tissues using TRIzol (Invitrogen Chromos Singa-pore) and purified according to the instructions includedDouble-stranded cDNA was synthesized from sim1 120583g of totalRNA using ReverTra Ace qPCR RT kit (TOYOBO OsakaJapan) Real-time PCR reactions were set up using the SYBR-Green PCR kit (TOYOBO Osaka Japan) and were cycled inStepOne Real-Time PCR System (Applied Biosystems CAUSA) as previously described [52] The abundance of targetmRNA was normalized to that of 120573-actin Primer pairs weredesigned based on GenBank reference sequences and listedin Table S1 available online in Supplementary Material athttpdxdoiorg1011552014315896
48 Western Blotting The isolated muscle tissue (sim200mg)was weighed and cut into pieces at 4∘C and 2mL (10times dryweight of isolated muscle fibers) of ice-cold buffer (50mMTris HCl 150mMNaCl 1mM EDTA 02mM PMSF 1 NP-40 pH 74 cocktail) was added The tissue pieces were thenhomogenized after which homogenates were centrifuged for10min at 8000 g and 4∘C The total protein content of thesupernatant was quantified using bicinchoninic acid reagents
and BSA standards (Shanghai Sangon Shanghai China)Theprotein samples were immunoblotted as previously described[52] Visualization of reaction bands was performed by 331015840-diaminobenzidine staining (Shanghai Sangon China) andscanned densitometrically 120573-Actin was used to standardizefor the amount of protein loaded
49 Statistical Analysis Data are reported as mean plusmn SEMStatistical differences between treatments were determinedusing ANOVA For all tests the significance level was set at119875 lt 005
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from the National NaturalScience Foundation of China (Grant nos 31300977 and31171142) This work was also supported by the Key Labora-tory Construction Project of Adolescent Health Assessmentand Exercise Intervention of Ministry of Education EastChinaNormalUniversity (no 40500-541235-14203004)Theauthors also thank postgraduate students at Professor Dingrsquoslab for the technical support on animal care biochemicalassays PCR and western blot
References
[1] M Sandri ldquoAutophagy in skeletal musclerdquo FEBS Letters vol584 no 7 pp 1411ndash1416 2010
[2] G Twig B Hyde and O S Shirihai ldquoMitochondrial fusionfission and autophagy as a quality control axis the bioenergeticviewrdquo Biochimica et Biophysica Acta Bioenergetics vol 1777 no9 pp 1092ndash1097 2008
[3] Q Sun W Fan K Chen X Ding S Chen and Q ZhongldquoIdentification of Barkor as a mammalian autophagy-specificfactor for Beclin 1 and class III phosphatidylinositol 3-kinaserdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 105 no 49 pp 19211ndash19216 2008
[4] M C Maiuri E Zalckvar A Kimchi and G Kroemer ldquoSelf-eating and self-killing crosstalk between autophagy and apop-tosisrdquo Nature Reviews Molecular Cell Biology vol 8 no 9 pp741ndash752 2007
[5] T Ueno W Sato Y Horie et al ldquoLoss of Pten a tumor suppres-sor causes the strong inhibition of autophagy without affectingLC3 lipidationrdquo Autophagy vol 4 no 5 pp 692ndash700 2008
[6] F FortunatoH Burgers F Bergmann et al ldquoImpaired autolyso-some formation correlates with lamp-2 depletion role of apop-tosis autophagy and necrosis in pancreatitisrdquoGastroenterologyvol 137 no 1 pp 350ndash360 2009
[7] F A Graca D A Goncalves W A Silveira et al ldquoEpinephrinedepletion exacerbates the fasting-induced protein breakdown infast-twitch skeletal musclesrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 305 pp E1483ndashE14942013
[8] T Ogata Y Oishi M Higuchi and I Muraoka ldquoFasting-relatedautophagic response in slow- and fast-twitch skeletal musclerdquo
Oxidative Medicine and Cellular Longevity 9
Biochemical andBiophysical ResearchCommunications vol 394no 1 pp 136ndash140 2010
[9] A Doria M Gatto and L Punzi ldquoAutophagy in human healthand diseaserdquo New England Journal of Medicine vol 368 no 19p 1845 2013
[10] S Fulda ldquoAutophagy and cell deathrdquo Autophagy vol 8 no 8pp 1250ndash1251 2012
[11] E Masiero L Agatea C Mammucari et al ldquoAutophagy isrequired to maintain muscle massrdquo Cell Metabolism vol 10 no6 pp 507ndash515 2009
[12] A C Nascimbeni M Fanin E Masiero C Angelini and MSandri ldquoImpaired autophagy contributes to muscle atrophy inglycogen storage disease type II patientsrdquo Autophagy vol 8 no11 pp 1697ndash1700 2012
[13] M Sandri ldquoProtein breakdown in muscle wasting role ofautophagy-lysosome and ubiquitin-proteasomerdquo InternationalJournal of Biochemistry and Cell Biology vol 45 pp 2121ndash21292013
[14] E Masiero and M Sandri ldquoAutophagy inhibition inducesatrophy and myopathy in adult skeletal musclesrdquo Autophagyvol 6 no 2 pp 307ndash309 2010
[15] R Scherz-Shouval and Z Elazar ldquoRegulation of autophagy byROS physiology and pathologyrdquoTrends in Biochemical Sciencesvol 36 no 1 pp 30ndash38 2011
[16] C R Morales Z Pedrozo S Lavandero and J A Hill ldquoOxi-dative stress and autophagy in cardiovascular homeostasisrdquoAntioxidants amp Redox Signaling vol 20 pp 507ndash518 2014
[17] C Luo Y Li H Wang et al ldquoMitochondrial accumulationunder oxidative stress is due to defects in autophagyrdquo Journalof Cellular Biochemistry vol 114 no 1 pp 212ndash219 2013
[18] Y Wang Y Nartiss B Steipe G A McQuibban and PK Kim ldquoROS-induced mitochondrial depolarization initi-ates PARK2PARKIN-dependent mitochondrial degradationby autophagyrdquo Autophagy vol 8 no 10 pp 1462ndash1476 2012
[19] T Zhang Y Li K A Park et al ldquoCucurbitacin induces auto-phagy through mitochondrial ROS production which counter-acts to limit caspase-dependent apoptrdquo Autophagy vol 8 no 4pp 559ndash576 2012
[20] R Sandhir A Sood A Mehrotra and S S Kamboj ldquoN-acetyl-cysteine reverses mitochondrial dysfunctions and behavioralabnormalities in 3-nitropropionic acid-induced Huntingtonsdiseaserdquo Neurodegenerative Diseases vol 9 no 3 pp 145ndash1572012
[21] S S Kamboj and R Sandhir ldquoProtective effect of N-acetyl-cysteine supplementation onmitochondrial oxidative stress andmitochondrial enzymes in cerebral cortex of streptozotocin-treated diabetic ratsrdquoMitochondrion vol 11 no 1 pp 214ndash2222011
[22] M ZafarullahW Q Li J Sylvester andM Ahmad ldquoMolecularmechanisms of N-acetylcysteine actionsrdquo Cellular and Molecu-lar Life Sciences vol 60 no 1 pp 6ndash20 2003
[23] N A Strobel J M Peake A Matsumoto S A Marsh J SCoombes and G D Wadley ldquoAntioxidant supplementationreduces skeletal muscle mitochondrial biogenesisrdquo Medicineand Science in Sports and Exercise vol 43 no 6 pp 1017ndash10242011
[24] M-C Gomez-Cabrera E Domenech M Romagnoli et alldquoOral administration of vitamin C decreases muscle mitochon-drial biogenesis and hampers training-induced adaptationsin endurance performancerdquo The American Journal of ClinicalNutrition vol 87 no 1 pp 142ndash149 2008
[25] N P Whitehead C Pham O L Gervasio and D G Allen ldquoN-Acetylcysteine ameliorates skeletal muscle pathophysiology inmdx micerdquo Journal of Physiology vol 586 no 7 pp 2003ndash20142008
[26] C-H Lin S-C Kuo L-J Huang and P-W Gean ldquoNeuropro-tective effect ofN-acetylcysteine on neuronal apoptosis inducedby a synthetic gingerdione compound involvement of ERK andp38 phosphorylationrdquo Journal of Neuroscience Research vol 84no 7 pp 1485ndash1494 2006
[27] O BKotoulas S AKalamidas andD J Kondomerkos ldquoGlyco-gen autophagy in glucose homeostasisrdquo Pathology Research andPractice vol 202 no 9 pp 631ndash638 2006
[28] J Zirin J Nieuwenhuis and N Perrimon ldquoRole of autophagyin glycogen breakdown and its relevance to chloroquinemyopa-thyrdquo PLoS Biology vol 11 no 11 Article ID e1001708 2013
[29] P Castets S Lin N Rion et al ldquoSustained activation ofmTORC1 in skeletalmuscle inhibits constitutive and starvation-induced autophagy and causes a severe late-onset myopathyrdquoCell Metabolism vol 17 no 5 pp 731ndash744 2013
[30] M Rahman M Mofarrahi A S Kristof B Nkengfac S Hareland S N Hussain ldquoReactive oxygen species regulation ofautophagy in skeletal musclesrdquoAntioxidants amp Redox Signalingvol 20 no 3 pp 443ndash459 2014
[31] J Yan Z Feng J Liu et al ldquoEnhanced autophagy plays a cardi-nal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats ameliorating effects of (-)-epigallocatechin-3-gallaterdquo Journal of Nutritional Biochemistry vol 23 no 7 pp716ndash724 2012
[32] LV Yuzefovych S IMusiyenkoG LWilson and L I RachekldquoMitochondrial DNA damage and dysfunction and oxidativestress are associated with endoplasmic reticulum stress proteindegradation and apoptosis in high fat diet-induced InsulinresistancemicerdquoPLoSONE vol 8 no 1 Article ID e54059 2013
[33] L Li Y Chen and S B Gibson ldquoStarvation-induced autophagyis regulated bymitochondrial reactive oxygen species leading toAMPK activationrdquo Cellular Signalling vol 25 no 1 pp 50ndash652013
[34] S Takikita C Schreiner R Baum et al ldquoFiber type conversionby PGC-1120572 activates lysosomal and autophagosomal biogenesisin both unaffected and pompe skeletal musclerdquo PLoS ONE vol5 no 12 Article ID e15239 2010
[35] K Hollinger D Gardan-Salmon C Santana D Rice E Snellaand J T Selsby ldquoRescue of dystrophic skeletal muscle byPGC-1120572 involves restored expression of dystrophin-associatedprotein complex components and satellite cell signalingrdquo TheAmerican Journal of Physiology Regulatory Integrative andComparative Physiology vol 305 no 1 pp R13ndashR23 2013
[36] T Wenz S G Rossi R L Rotundo B M Spiegelman and CT Moraes ldquoIncreased muscle PGC-1120572 expression protects fromsarcopenia and metabolic disease during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 48 pp 20405ndash20410 2009
[37] I Grattagliano P Portincasa T Cocco et al ldquoEffect of dietaryrestriction and N-acetylcysteine supplementation on intestinalmucosa and liver mitochondrial redox status and function inaged ratsrdquo Experimental Gerontology vol 39 no 9 pp 1323ndash1332 2004
[38] S Kumar and S L Sitasawad ldquoN-acetylcysteine prevents glu-coseglucose oxidase-induced oxidative stress mitochondrialdamage and apoptosis in H9c2 cellsrdquo Life Sciences vol 84 no11-12 pp 328ndash336 2009
10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Oxidative Medicine and Cellular Longevity 9
Biochemical andBiophysical ResearchCommunications vol 394no 1 pp 136ndash140 2010
[9] A Doria M Gatto and L Punzi ldquoAutophagy in human healthand diseaserdquo New England Journal of Medicine vol 368 no 19p 1845 2013
[10] S Fulda ldquoAutophagy and cell deathrdquo Autophagy vol 8 no 8pp 1250ndash1251 2012
[11] E Masiero L Agatea C Mammucari et al ldquoAutophagy isrequired to maintain muscle massrdquo Cell Metabolism vol 10 no6 pp 507ndash515 2009
[12] A C Nascimbeni M Fanin E Masiero C Angelini and MSandri ldquoImpaired autophagy contributes to muscle atrophy inglycogen storage disease type II patientsrdquo Autophagy vol 8 no11 pp 1697ndash1700 2012
[13] M Sandri ldquoProtein breakdown in muscle wasting role ofautophagy-lysosome and ubiquitin-proteasomerdquo InternationalJournal of Biochemistry and Cell Biology vol 45 pp 2121ndash21292013
[14] E Masiero and M Sandri ldquoAutophagy inhibition inducesatrophy and myopathy in adult skeletal musclesrdquo Autophagyvol 6 no 2 pp 307ndash309 2010
[15] R Scherz-Shouval and Z Elazar ldquoRegulation of autophagy byROS physiology and pathologyrdquoTrends in Biochemical Sciencesvol 36 no 1 pp 30ndash38 2011
[16] C R Morales Z Pedrozo S Lavandero and J A Hill ldquoOxi-dative stress and autophagy in cardiovascular homeostasisrdquoAntioxidants amp Redox Signaling vol 20 pp 507ndash518 2014
[17] C Luo Y Li H Wang et al ldquoMitochondrial accumulationunder oxidative stress is due to defects in autophagyrdquo Journalof Cellular Biochemistry vol 114 no 1 pp 212ndash219 2013
[18] Y Wang Y Nartiss B Steipe G A McQuibban and PK Kim ldquoROS-induced mitochondrial depolarization initi-ates PARK2PARKIN-dependent mitochondrial degradationby autophagyrdquo Autophagy vol 8 no 10 pp 1462ndash1476 2012
[19] T Zhang Y Li K A Park et al ldquoCucurbitacin induces auto-phagy through mitochondrial ROS production which counter-acts to limit caspase-dependent apoptrdquo Autophagy vol 8 no 4pp 559ndash576 2012
[20] R Sandhir A Sood A Mehrotra and S S Kamboj ldquoN-acetyl-cysteine reverses mitochondrial dysfunctions and behavioralabnormalities in 3-nitropropionic acid-induced Huntingtonsdiseaserdquo Neurodegenerative Diseases vol 9 no 3 pp 145ndash1572012
[21] S S Kamboj and R Sandhir ldquoProtective effect of N-acetyl-cysteine supplementation onmitochondrial oxidative stress andmitochondrial enzymes in cerebral cortex of streptozotocin-treated diabetic ratsrdquoMitochondrion vol 11 no 1 pp 214ndash2222011
[22] M ZafarullahW Q Li J Sylvester andM Ahmad ldquoMolecularmechanisms of N-acetylcysteine actionsrdquo Cellular and Molecu-lar Life Sciences vol 60 no 1 pp 6ndash20 2003
[23] N A Strobel J M Peake A Matsumoto S A Marsh J SCoombes and G D Wadley ldquoAntioxidant supplementationreduces skeletal muscle mitochondrial biogenesisrdquo Medicineand Science in Sports and Exercise vol 43 no 6 pp 1017ndash10242011
[24] M-C Gomez-Cabrera E Domenech M Romagnoli et alldquoOral administration of vitamin C decreases muscle mitochon-drial biogenesis and hampers training-induced adaptationsin endurance performancerdquo The American Journal of ClinicalNutrition vol 87 no 1 pp 142ndash149 2008
[25] N P Whitehead C Pham O L Gervasio and D G Allen ldquoN-Acetylcysteine ameliorates skeletal muscle pathophysiology inmdx micerdquo Journal of Physiology vol 586 no 7 pp 2003ndash20142008
[26] C-H Lin S-C Kuo L-J Huang and P-W Gean ldquoNeuropro-tective effect ofN-acetylcysteine on neuronal apoptosis inducedby a synthetic gingerdione compound involvement of ERK andp38 phosphorylationrdquo Journal of Neuroscience Research vol 84no 7 pp 1485ndash1494 2006
[27] O BKotoulas S AKalamidas andD J Kondomerkos ldquoGlyco-gen autophagy in glucose homeostasisrdquo Pathology Research andPractice vol 202 no 9 pp 631ndash638 2006
[28] J Zirin J Nieuwenhuis and N Perrimon ldquoRole of autophagyin glycogen breakdown and its relevance to chloroquinemyopa-thyrdquo PLoS Biology vol 11 no 11 Article ID e1001708 2013
[29] P Castets S Lin N Rion et al ldquoSustained activation ofmTORC1 in skeletalmuscle inhibits constitutive and starvation-induced autophagy and causes a severe late-onset myopathyrdquoCell Metabolism vol 17 no 5 pp 731ndash744 2013
[30] M Rahman M Mofarrahi A S Kristof B Nkengfac S Hareland S N Hussain ldquoReactive oxygen species regulation ofautophagy in skeletal musclesrdquoAntioxidants amp Redox Signalingvol 20 no 3 pp 443ndash459 2014
[31] J Yan Z Feng J Liu et al ldquoEnhanced autophagy plays a cardi-nal role in mitochondrial dysfunction in type 2 diabetic Goto-Kakizaki (GK) rats ameliorating effects of (-)-epigallocatechin-3-gallaterdquo Journal of Nutritional Biochemistry vol 23 no 7 pp716ndash724 2012
[32] LV Yuzefovych S IMusiyenkoG LWilson and L I RachekldquoMitochondrial DNA damage and dysfunction and oxidativestress are associated with endoplasmic reticulum stress proteindegradation and apoptosis in high fat diet-induced InsulinresistancemicerdquoPLoSONE vol 8 no 1 Article ID e54059 2013
[33] L Li Y Chen and S B Gibson ldquoStarvation-induced autophagyis regulated bymitochondrial reactive oxygen species leading toAMPK activationrdquo Cellular Signalling vol 25 no 1 pp 50ndash652013
[34] S Takikita C Schreiner R Baum et al ldquoFiber type conversionby PGC-1120572 activates lysosomal and autophagosomal biogenesisin both unaffected and pompe skeletal musclerdquo PLoS ONE vol5 no 12 Article ID e15239 2010
[35] K Hollinger D Gardan-Salmon C Santana D Rice E Snellaand J T Selsby ldquoRescue of dystrophic skeletal muscle byPGC-1120572 involves restored expression of dystrophin-associatedprotein complex components and satellite cell signalingrdquo TheAmerican Journal of Physiology Regulatory Integrative andComparative Physiology vol 305 no 1 pp R13ndashR23 2013
[36] T Wenz S G Rossi R L Rotundo B M Spiegelman and CT Moraes ldquoIncreased muscle PGC-1120572 expression protects fromsarcopenia and metabolic disease during agingrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 106 no 48 pp 20405ndash20410 2009
[37] I Grattagliano P Portincasa T Cocco et al ldquoEffect of dietaryrestriction and N-acetylcysteine supplementation on intestinalmucosa and liver mitochondrial redox status and function inaged ratsrdquo Experimental Gerontology vol 39 no 9 pp 1323ndash1332 2004
[38] S Kumar and S L Sitasawad ldquoN-acetylcysteine prevents glu-coseglucose oxidase-induced oxidative stress mitochondrialdamage and apoptosis in H9c2 cellsrdquo Life Sciences vol 84 no11-12 pp 328ndash336 2009
10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Oxidative Medicine and Cellular Longevity
[39] L Bevilacqua J J Ramsey K Hagopian R Weindruch andM-E Harper ldquoEffects of short- and medium-term calorierestriction on muscle mitochondrial proton leak and reactiveoxygen species productionrdquo The American Journal of Physiol-ogy Endocrinology and Metabolism vol 286 no 5 pp E852ndashE861 2004
[40] S S Korshunov V P Skulachev and A A Starkov ldquoHigh pro-tonic potential actuates a mechanism of production of reactiveoxygen species in mitochondriardquo FEBS Letters vol 416 no 1pp 15ndash18 1997
[41] E J Anderson M E Lustig K E Boyle et al ldquoMitochondrialH2O2emission and cellular redox state link excess fat intake
to insulin resistance in both rodents and humansrdquo Journal ofClinical Investigation vol 119 no 3 pp 573ndash581 2009
[42] V M Gohil S A Sheth R Nilsson et al ldquoNutrient-sensitizedscreening for drugs that shift energy metabolism from mito-chondrial respiration to glycolysisrdquo Nature Biotechnology vol28 no 3 pp 249ndash255 2010
[43] N Parajuli and L A MacMillan-Crow ldquoRole of reduced man-ganese superoxide dismutase in ischemia-reperfusion injury apossible trigger for autophagy and mitochondrial biogenesisrdquoThe American Journal of Physiology Renal Physiology vol 304no 3 pp F257ndashF267 2013
[44] S L Mehta Y Lin W Chen et al ldquoManganese superoxide dis-mutase deficiency exacerbates ischemic brain damage underhyperglycemic conditions by altering autophagyrdquo TranslationalStroke Research vol 2 no 1 pp 42ndash50 2011
[45] Z Li K Shi L Guan et al ldquoROS leads toMnSOD upregulationthrough ERK2 translocation and p53 activation in selenite-induced apoptosis of NB4 cellsrdquo FEBS Letters vol 584 no 11pp 2291ndash2297 2010
[46] S B Mustafi P K Chakraborty R S Dey and S RahaldquoHeat stress upregulates chaperone heat shock protein 70 andantioxidant manganese superoxide dismutase through reactiveoxygen species (ROS) p38MAPK and Aktrdquo Cell Stress andChaperones vol 14 no 6 pp 579ndash589 2009
[47] K Bensaad E C Cheung and K H Vousden ldquoModulation ofintracellular ROS levels by TIGAR controls autophagyrdquo EMBOJournal vol 28 no 19 pp 3015ndash3026 2009
[48] C Wanka J P Steinbach and J Rieger ldquoTp53-induced gly-colysis and apoptosis regulator (TIGAR) protects glioma cellsfrom starvation-induced cell death by up-regulating respirationand improving cellular redox homeostasisrdquo Journal of BiologicalChemistry vol 287 no 40 pp 33436ndash33446 2012
[49] A Hoshino S Matoba E Iwai-Kanai et al ldquoP53-TIGARaxis attenuates mitophagy to exacerbate cardiac damage afterischemiardquo Journal of Molecular and Cellular Cardiology vol 52no 1 pp 175ndash184 2012
[50] E C Cheung R L Ludwig andKH Vousden ldquoMitochondriallocalization of TIGAR under hypoxia stimulates HK2 andlowers ROS and cell deathrdquoProceedings of theNational Academyof Sciences of the United States of America vol 109 no 50 pp20491ndash20496 2012
[51] E Fernandez-VizarraG Ferrın A Perez-Martos P Fernandez-SilvaM Zeviani and J A Enrıquez ldquoIsolation ofmitochondriafor biogenetical studies an updaterdquoMitochondrion vol 10 no3 pp 253ndash262 2010
[52] Z Qi J He Y Su et al ldquoPhysical exercise regulates p53 activitytargeting SCO
2and increases Mitochondrial COX biogenesis
in cardiac muscle with agerdquo PLoS ONE vol 6 no 7 Article IDe21140 2011
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom





























