Embed Size (px)
Citation preview

JOURNAL OF BACTEIOLOGY, Mar. 1969, p. 1283-1290Copyright @ 1969 American Society for Microbiology
Vol. 97, No. 3Printed in U.S.A.
Repression of the Histidine Operon: Effect ofthe First Enzyme on the Kinetics of Repression
JOHN S. KOVACH, M. A. BERBERICH,1 PAL VENETIANER,9 AND ROBERT F. GOLDBERGERLaboratory of Chemical Biology, National Institute ofArthritis and Metabolic Diseases, Bethesda, Maryland 20014
Received for publication 21 December 1968
Kinetic studies on repression of the enzymes for histidine biosynthesis in Sal-monella typhimurium showed that, upon addition of histidine to a derepressed cul-ture, the enzymes became repressed in a temporal sequence which corresponds withthe positional sequence of the genes in the histidine operon. This serial pattern ofrepression occurred under conditions in which the feedback site of the first en-zyme for histidine biosynthesis is intact. When this site was rendered nonfunctionalthe pattern of repression was changed so that all of the enzymes became repressedconcomitantly. These results suggest that the first enzyme for histidine biosynthesisplays a hitherto unrecognized role in control of the histidine system.
The pathway for histidine biosynthesis inSalmonella typhimurium, elucidated largelythrough the work of B. N. Ames and co-workers(2, 5, 22, 27, 28), consists of a series of 10 reactionseach catalyzed by a specific enzyme. The geneswhich specify the structures of these enzymes arelocated in a small segment of the Salmonellachromosome known as the histidine operon (3,14). The histidine operon functions as a unit inresponse to the level of histidine available to theorganism. As originally shown by Ames andGarry (1), the differential rate of synthesis of thehistidine enzymes increases when histidine be-comes the limiting growth factor. That is, theoperon becomes derepressed. When excess histi-dine is added to a derepressed culture, the differ-ential rate of synthesis of these enzymes declines.That is, the operon becomes repressed.
Kinetic studies on the repression process arepresented here. These studies reveal that theenzyme encoded in the first gene of the histidineoperon may be involved in control of the histidinesystem. This involvement is demonstrated byexamining the manner in which translation of thepolycistronic histidine messenger ribonucleic acid(mRNA) ceases (19, 30).
MATERIALS AND METHODSBacterial strains. The organisms employed in these
studies were the LT-2 (wild type) strain of S. typhi-murium; the nonpolar, "complete" histidine auxo-
'Research Fellow, Helen Hay Whitney Foundation.'Visiting Scientist, NIAMD, 1965-66, on leave from Institute
of Medical Chemistry, University of Medicine, Budapest, Hun-gary.
trophs hisG52, hisG1102 hisIF135, hisG109 hisIF135,hisC2, hisA30, and hisIF135; the nonpolar, "leaky"histidine auxotrophs hisG1306, hisB59, and hisEll;the constitutive mutant hisS1520; and the polar non-sense mutant hisG428. These organisms were obtainedfrom the collection of P. E. Hartman.
Growth conditions. Cells were routinely grown inminimal medium (31) with glucose at 0.5% in 2 litersof culture medium in a 4-liter flask. The cultures wereaerated vigorously in a New Brunswick rotary shakerat 37 C. The histidine auxotrophs and the constitutivemutant were grown in the presence of a sufficientamount of L-histidine to support growth to an opticaldensity (OD) at 700 nm of approximately 0.35 (3.4 X10' cells per ml). After depletion of histidine from themedium, growth was supported by L-histidinol (2.5X 10- M; Cyclo Chemical Co.) in the case of the"complete" auxotrophs and by the endogenous syn-thesis of histidine in the case of the "leaky" auxotrophsand the constitutive mutant. Derepression in the wild-type organism was obtained when the culture hadreached a cell density of 3.4 X 108 cells per ml by theaddition of either DL-2-thiazolalanine (Cyclo ChemicalCo.; final concentration, 0.1 mM) or 3-amino-1 ,2,4-triazole (aminotriazole, final concentration, 20 mM;Aldrich Chemical Co.). In all experiments, the growthrate during derepression was the same (doubling time4 hr), and derepression was allowed to proceed for 30min. Repression was then effected by the addition ofL-histidine at a final concentration of 0.3 mm. Thegrowth rate always returned immediately to thatcharacteristic of repressed cells (doubling time 50min).
Experimental design. In a typical experiment,samples (50 to 100 ml) were withdrawn from the cul-ture periodically, so that 10 samples were collectedduring derepression and 10 samples were collectedafter the addition of histidine. Each sample was im-mediately mnixed with ice and excess histidine. After
1283
on March 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
KOVACH ET AL.
all samples had been collected, they were centrifuged.The pelleted cells were washed with 0.05 M tris (hy-droxymethyl)aminomethane (Tris) buffer (pH 8.0),containing histidine at a concentration of 0.1 mM,recentrifuged, and finally resuspended in 4 ml of 0.05M Tris buffer (pH 8.0). Extracts were prepared fromthe suspensions with a French pressure cell (AmericanInstrument Co.) at 6,000 psi, clarified by centrifuga-tion, and assayed immediately.
Assays and substrates. Protein was determined bythe method of Lowry et al. (15). Assays for the en-zymes corresponding to the first gene (G), third gene(C), fourth gene (B), sixth gene (A), seventh gene (F),and eighth gene (I) of the histidine operon were per-formed as previously described (2, 5, 17, 27, 28).Enzyme levels are expressed as the amount of activityper milligram of protein. Substrates were obtained asfollows: L-histidinol phosphate was purchased fromCyclo Chemical Co.; N-(5'-phospho-D-ribosylformi-mino) - 5 -amino- 1 - (5'-phosphoribosyl)-4-imidazole-carboxamide and N-1-(5"-phosphoribosyl)adenosinemonophosphate were synthesized enzymatically (27)from 5-phosphoribosyl-1-pyrophosphate (P-L Bio-chemicals).
RESULTSRepression in histidine auxotrophs. We studied
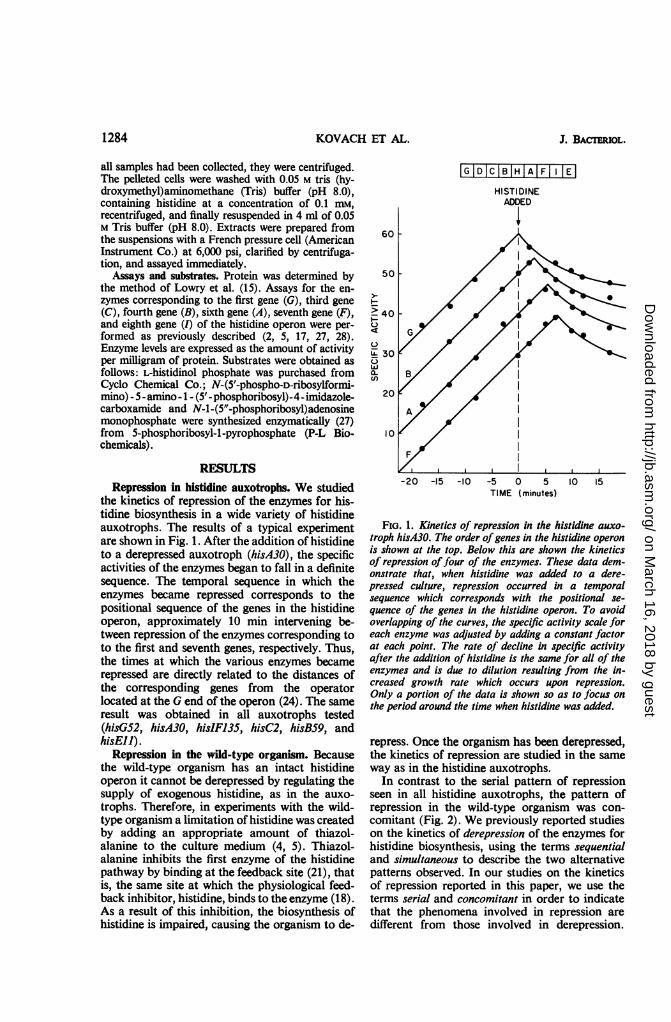
the kinetics of repression of the enzymes for his-tidine biosynthesis in a wide variety of histidineauxotrophs. The results of a typical experimentare shown in Fig. 1. After the addition of histidineto a derepressed auxotroph (hisA30), the specificactivities of the enzymes began to fall in a definitesequence. The temporal sequence in which theenzymes became repressed corresponds to thepositional sequence of the genes in the histidineoperon, approximately 10 min intervening be-tween repression of the enzymes corresponding toto the first and seventh genes, respectively. Thus,the times at which the various enzymes becamerepressed are directly related to the distances ofthe corresponding genes from the operatorlocated at the G end of the operon (24). The sameresult was obtained in all auxotrophs tested(hisG52, hisA30, hisIF135, hisC2, hisB59, andhisEll).
Repression in the wild-type organism. Becausethe wild-type organism has an intact histidineoperon it cannot be derepressed by regulating thesupply of exogenous histidine, as in the auxo-trophs. Therefore, in experiments with the wild-type organism a limitation of histidine was createdby adding an appropriate amount of thiazol-alanine to the culture medium (4, 5). Thiazol-alanine inhibits the first enzyme of the histidinepathway by binding at the feedback site (21), thatis, the same site at which the physiological feed-back inhibitor, histidine, binds to the enzyme (18).As a result of this inhibition, the biosynthesis ofhistidine is impaired, causing the organism to de-
IGIDICIBIHTAIFT I 1ElHISTI DINE
60
50
> 400
0iZ 30wa.
20
I01
-5 0 5 10TIME (minutes)
FIG. 1. Kinetics of repression in the histidine auxo-troph hisA30. The order ofgenes in the histidine operonis shown at the top. Below this are shown the kineticsof repression offour of the enzymes. These data dem-onstrate that, when histidine was added to a dere-pressed culture, repression occurred in a temporalsequence which corresponds with the positional se-quence of the genes in the histidine operon. To avoidoverlapping of the curves, the specific activity scale foreach enzyme was adjusted by adding a constant factorat each point. The rate of decline in specific activityafter the addition of histidine is the same for all of theenzymes and is due to dilution resulting from the in-creased growth rate which occurs upon repression.Only a portion of the data is shown so as to focus onthe period around the time when histidine was added.
repress. Once the organism has been derepressed,the kinetics of repression are studied in the sameway as in the histidine auxotrophs.
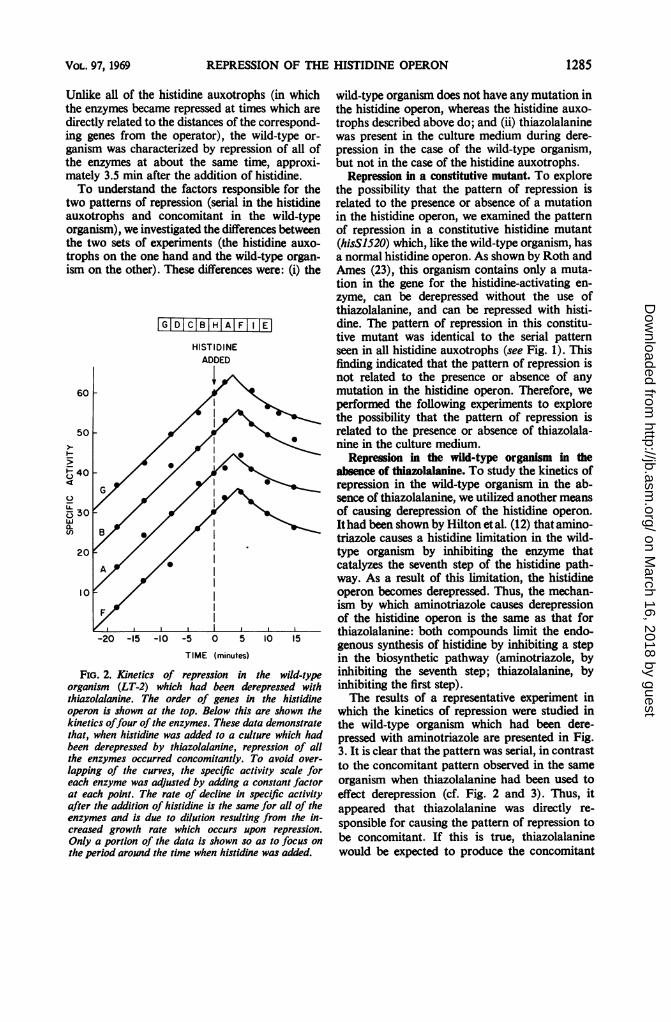
In contrast to the serial pattern of repressionseen in all histidine auxotrophs, the pattern ofrepression in the wild-type organism was con-comitant (Fig. 2). We previously reported studieson the kinetics of derepression of the enzymes forhistidine biosynthesis, using the terms sequentialand simultaneous to describe the two alternativepatterns observed. In our studies on the kineticsof repression reported in this paper, we use theterms serial and concomitant in order to indicatethat the phenomena involved in repression aredifferent from those involved in derepression.
1284 J. BAC-rERIOL.
on March 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
REPRESSION OF THE HISTIDINE OPERON
Unlike all of the histidine auxotrophs (in whichthe enzymes became repressed at times which aredirectly related to the distances of the correspond-ing genes from the operator), the wild-type or-ganism was characterized by repression of all ofthe enzymes at about the same time, approxi-mately 3.5 min after the addition of histidine.To understand the factors responsible for the
two patterns of repression (serial in the histidineauxotrophs and concomitant in the wild-typeorganism), we investigated the differences betweenthe two sets of experiments (the histidine auxo-trophs on the one hand and the wild-type organ-ism on the other). These differences were: (i) the
IGIDICIBIHIAIFI |E
60
50
F 40
0
1530w
20
IOl
HISTIDINEADDED
TIME (minutes)
Fio. 2. Kinetics of repression in the wild-typeorganism (LT-2) which had been derepressed withthiazolalanine. The order of genes in the histidineoperon is shown at the top. Below this are shown thekinetics offour of the enzymes. These data demonstratethat, when histidine was added to a culture which hadbeen derepressed by thiazolalanine, repression of allthe enzymes occurred concomitantly. To avoid over-
lapping of the curves, the specific activity scale foreach enzyme was adjusted by adding a constant factorat each point. The rate of decline in specific activityafter the addition of histidine is the same for all of theenzymes and is due to dilution resulting from the in-creased growth rate which occurs upon repression.Only a portion of the data is shown so as to focus onthe period around the time when histidine was added.
wild-type organism does not have any mutation inthe histidine operon, whereas the histidine auxo-trophs described above do; and (ii) thiazolalaninewas present in the culture medium during dere-pression in the case of the wild-type organism,but not in the case of the histidine auxotrophs.
Repression in a constitutive mutant. To explorethe possibility that the pattern of repression isrelated to the presence or absence of a mutationin the histidine operon, we examined the patternof repression in a constitutive histidine mutant(hisS1520) which, like the wild-type organism, hasa normal histidine operon. As shown by Roth andAmes (23), this organism contains only a muta-tion in the gene for the histidine-activating en-zyme, can be derepressed without the use ofthiazolalanine, and can be repressed with histi-dine. The pattern of repression in this constitu-tive mutant was identical to the serial patternseen in all histidine auxotrophs (see Fig. 1). Thisfinding indicated that the pattern of repression isnot related to the presence or absence of anymutation in the histidine operon. Therefore, weperformed the following experiments to explorethe possibility that the pattern of repression isrelated to the presence or absence of thiazolala-nine in the culture medium.
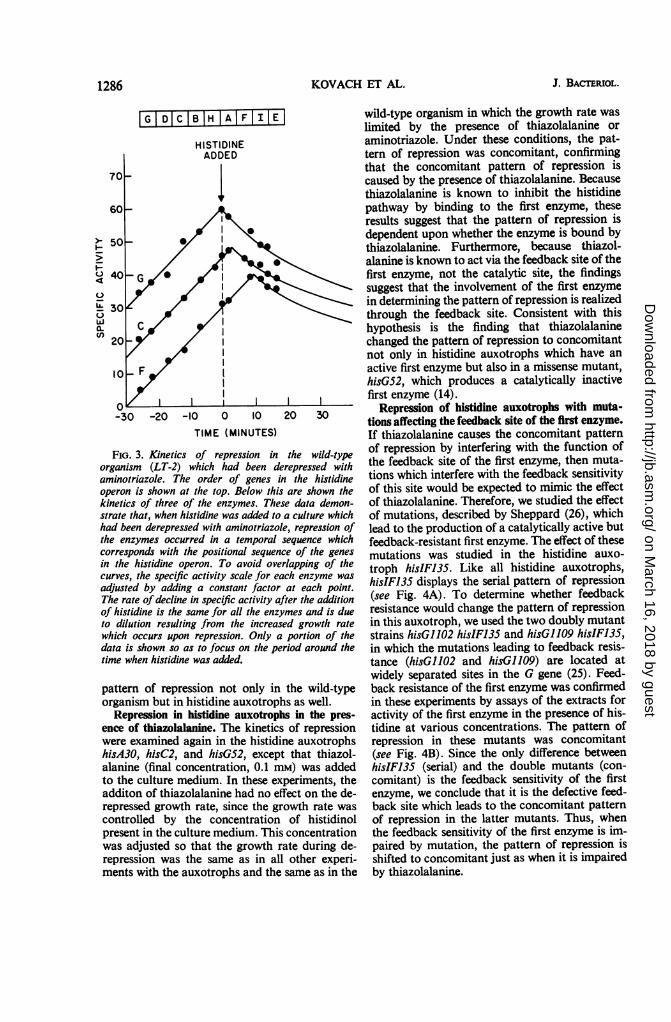
Repression in the wild-type organism in theabsence of thiazolalanine. To study the kinetics ofrepression in the wild-type organism in the ab-sence of thiazolalanine, we utilized another meansof causing derepression of the histidine operon.Ithad been shown by Hilton etal. (12) that amino-triazole causes a histidine limitation in the wild-type organism by inhibiting the enzyme thatcatalyzes the seventh step of the histidine path-way. As a result of this limitation, the histidineoperon becomes derepressed. Thus, the mechan-ism by which aminotriazole causes derepressionof the histidine operon is the same as that forthiazolalanine: both compounds limit the endo-genous synthesis of histidine by inhibiting a stepin the biosynthetic pathway (aminotriazole, byinhibiting the seventh step; thiazolalanine, byinhibiting the first step).The results of a representative experiment in
which the kinetics of repression were studied inthe wild-type organism which had been dere-pressed with aminotriazole are presented in Fig.3. It is clear that the pattern was serial, in contrastto the concomitant pattern observed in the sameorganism when thiazolalanine had been used toeffect derepression (cf. Fig. 2 and 3). Thus, itappeared that thiazolalanine was directly re-sponsible for causing the pattern of repression tobe concomitant. If this is true, thiazolalaninewould be expected to produce the concomitant
1285VOL. 97, 1969
on March 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
KOVACH ET AL.
[-GI DC B1H AI F II EJIHISTIDINEADDED
-30 -20 -10 0 10 20 30
TIME (MINUTES)
FIG. 3. Kinetics of repression in the wild-typeorganism (LT-2) which had been derepressed withaminotriazole. The order of genes in the histidineoperon is shown at the top. Below this are shown thekinetics of three of the enzymes. These data demon-strate that, when histidine was added to a culture whichhad been derepressed with aminotriazole, repression ofthe enzymes occurred in a temporal sequence whichcorresponds with the positional sequence of the genes
in the histidine operon. To avoid overlapping of thecurves, the specific activity scale for each enzyme was
adjusted by adding a constant factor at each point.The rate of decline in specific activity after the additionof histidine is the same for all the enzymes and is dueto dilution resulting from the increased growth rate
which occurs upon repression. Only a portion of thedata is shown so as to focus on the period around thetime when histidine was added.
pattern of repression not only in the wild-typeorganism but in histidine auxotrophs as well.
Repression in histidine auxotrophs in the pres-
ence of thiazolalanine. The kinetics of repressionwere examined again in the histidine auxotrophshisA30, hisC2, and hisGS2, except that thiazol-alanine (final concentration, 0.1 mM) was addedto the culture medium. In these experiments, theadditon of thiazolalanine had no effect on the de-repressed growth rate, since the growth rate was
controlled by the concentration of histidinolpresent in the culture medium. This concentrationwas adjusted so that the growth rate during de-repression was the same as in all other experi-ments with the auxotrophs and the same as in the
wild-type organism in which the growth rate waslimited by the presence of thiazolalanine oraminotriazole. Under these conditions, the pat-tern of repression was concomitant, confirmingthat the concomitant pattern of repression iscaused by the presence of thiazolalanine. Becausethiazolalanine is known to inhibit the histidinepathway by binding to the first enzyme, theseresults suggest that the pattern of repression isdependent upon whether the enzyme is bound bythiazolalanine. Furthermore, because thiazol-alanine is known to act via the feedback site of thefirst enzyme, not the catalytic site, the findingssuggest that the involvement of the first enzymein determining the pattern of repression is realizedthrough the feedback site. Consistent with thishypothesis is the finding that thiazolalaninechanged the pattern of repression to concomitantnot only in histidine auxotrophs which have anactive first enzyme but also in a missense mutant,hisG52, which produces a catalytically inactivefirst enzyme (14).
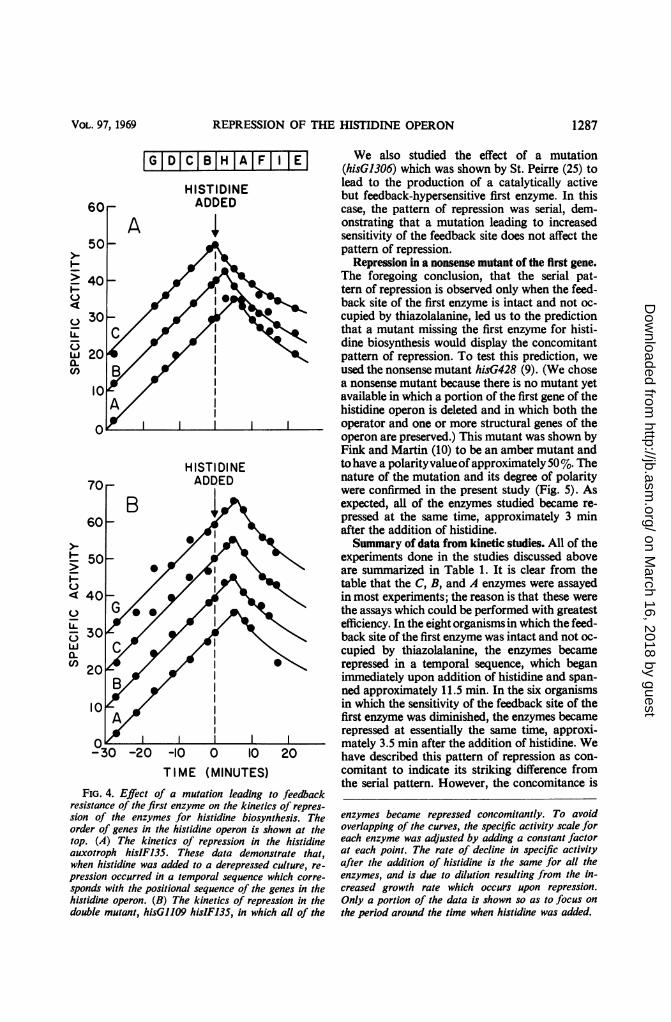
Repression of histidine auxotrophs with muta-tions affecting the feedback site of the first enzyme.If thiazolalanine causes the concomitant patternof repression by interfering with the function ofthe feedback site of the first enzyme, then muta-tions which interfere with the feedback sensitivityof this site would be expected to mimic the effectof thiazolalanine. Therefore, we studied the effectof mutations, described by Sheppard (26), whichlead to the production of a catalytically active butfeedback-resistant first enzyme. The effect of thesemutations was studied in the histidine auxo-troph hisIF135. Like all histidine auxotrophs,hisIF135 displays the serial pattern of repression(see Fig. 4A). To determine whether feedbackresistance would change the pattern of repressionin this auxotroph, we used the two doubly mutantstrains hisG1102 hisIF135 and hisG1109 hisIF135,in which the mutations leading to feedback resis-tance (hisG1102 and hisG109) are located atwidely separated sites in the G gene (25). Feed-back resistance of the first enzyme was confirmedin these experiments by assays of the extracts foractivity of the first enzyme in the presence of his-tidine at various concentrations. The pattern ofrepression in these mutants was concomitant(see Fig. 4B). Since the only difference betweenhisIF135 (serial) and the double mutants (con-comitant) is the feedback sensitivity of the firstenzyme, we conclude that it is the defective feed-back site which leads to the concomitant patternof repression in the latter mutants. Thus, whenthe feedback sensitivity of the first enzyme is im-paired by mutation, the pattern of repression isshifted to concomitant just as when it is impairedby thiazolalanine.
-
4
UIL
wa.U)
1286 J. BACTERIOL.
on March 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
REPRESSION OF THE HISTIDINE OPERON
IGIDICIBIHIAIFII IEIHISTIDINEADDED
-30 -20 -10 0 10 20
TIME (MINUTES)
FIG. 4. Effect of a mutation leading to feedbackresistance of thefirst enzyme on the kinetics of repres-
sion of the enzymes for histidine biosynthesis. Theorder of genes in the histidine operon is shown at thetop. (A) The kinetics of repression in the histidineauxotroph hisIF135. These data demonstrate that,when histidine was added to a derepressed culture, re-
pression occurred in a temporal sequence which corre-
sponds with the positional sequence of the genes in thehistidine operon. (B) The kinetics of repression in thedouble mutant, hisG1109 hisIF135, in which all of the
We also studied the effect of a mutation(hisG1306) which was shown by St. Peirre (25) tolead to the production of a catalytically activebut feedback-hypersensitive first enzyme. In thiscase, the pattern of repression was serial, dem-onstrating that a mutation leading to increasedsensitivity of the feedback site does not affect thepattern of repression.
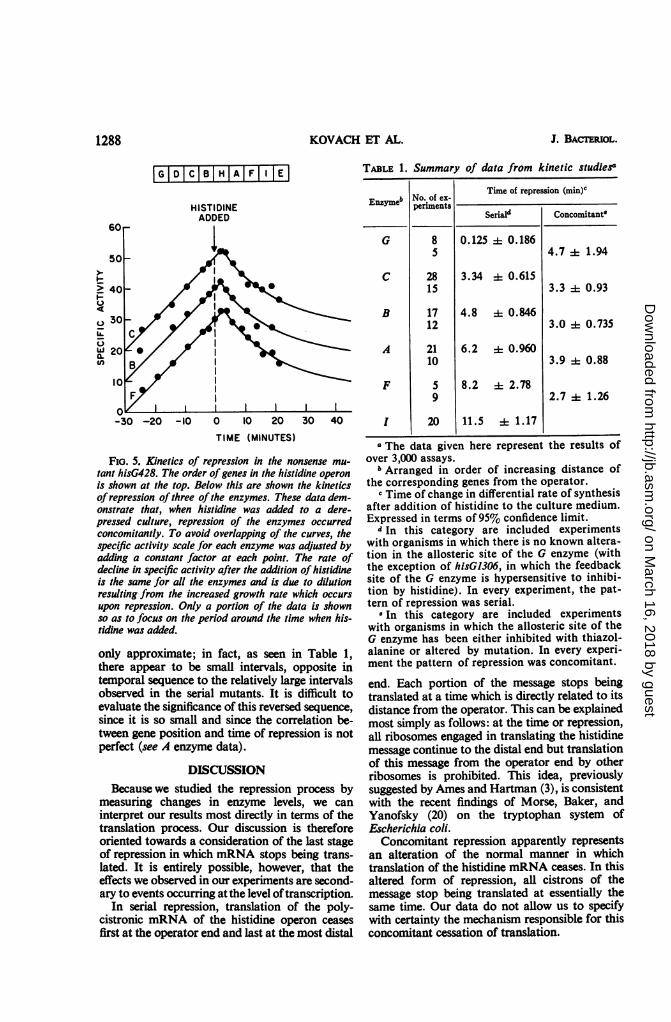
Repression in a nonsense mutant of the first gene.The foregoing conclusion, that the serial pat-tern of repression is observed only when the feed-back site of the first enzyme is intact and not oc-cupied by thiazolalanine, led us to the predictionthat a mutant missing the first enzyme for histi-dine biosynthesis would display the concomitantpattern of repression. To test this prediction, weused the nonsense mutant hisG428 (9). (We chosea nonsense mutant because there is no mutant yetavailable in which a portion of the first gene of thehistidine operon is deleted and in which both theoperator and one or more structural genes of theoperon are preserved.) This mutant was shown byFink and Martin (10) to be an amber mutant andto have a polarityvalueofapproximately 50%. Thenature of the mutation and its degree of polaritywere confirmed in the present study (Fig. 5). Asexpected, all of the enzymes studied became re-pressed at the same time, approximately 3 minafter the addition of histidine.Summary of data from kinetic studies. All of the
experiments done in the studies discussed aboveare summarized in Table 1. It is clear from thetable that the C, B, and A enzymes were assayedin most experiments; the reason is that these werethe assays which could be performed with greatestefficiency. In the eight organisms in which the feed-back site of the first enzyme was intact and not oc-cupied by thiazolalanine, the enzymes becamerepressed in a temporal sequence, which beganimmediately upon addition of histidine and span-ned approximately 11.5 min. In the six organismsin which the sensitivity of the feedback site of thefirst enzyme was diminished, the enzymes becamerepressed at essentially the same time, approxi-mately 3.5 min after the addition of histidine. Wehave described this pattern of repression as con-comitant to indicate its striking difference fromthe serial pattern. However, the concomitance is
enzymes became repressed concomitantly. To avoidoverlapping of the curves, the specific activity scale foreach enzyme was adjusted by adding a constant factorat each point. The rate of decline in specific activityafter the addition of histidine is the same for all theenzymes, and is due to dilution resulting from the in-creased growth rate which occurs upon repression.Only a portion of the data is shown so as to focus on
the period around the time when histidine was added.
I-
b.
LA5
wa.(I)
LLZ5wa.Cf)
VOL. 97, 1969 1287
on March 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
KOVACH ET AL.
IGID|ICIBIHIA|F I El
4
C-)
ILw
a.(I)
HISTIDINEADDED
TIME (MINUTES)
FIG. 5. Kinetics of repression in the nonsense mu-
tant hisG428. The order of genes in the histidine operon
is shown at the top. Below this are shown the kineticsofrepression ofthree ofthe enzymes. These data dem-onstrate that, when histidine was added to a dere-pressed culture, repression of the enzymes occurredconcomitantly. To avoid overlapping of the curves, thespecific activity scale for each enzyme was adjusted byadding a constant factor at each point. The rate ofdecline in specific activity after the addition of histidineis the same for all the enzymes and is due to dilutionresulting from the increased growth rate which occurs
upon repression. Only a portion of the data is shownso as to focus on the period around the time when his-tidine was added.
only approximate; in fact, as seen in Table 1,there appear to be small intervals, opposite intemporal sequence to the relatively large intervalsobserved in the serial mutants. It is difficult toevaluate the significance of this reversed sequence,
since it is so small and since the correlation be-tween gene position and time of repression is notperfect (see A enzyme data).
DISCUSSIONBecause we studied the repression process by
measuring changes in enzyme levels, we can
interpret our results most directly in terms of thetranslation process. Our discussion is thereforeoriented towards a consideration of the last stageof repression in which mRNA stops being trans-lated. It is entirely possible, however, that theeffects we observed in our experiments are second-ary to events occurring at the level oftranscription.
In serial repression, translation of the poly-cistronic mRNA of the histidine operon ceases
first at the operator end and last at the most distal
TABLE 1. Summary of data from kinetic studies"
Time of repression (min)'Enzymeb No. ofpe-
Seriald Concomitant
G 8 0.125 i 0.1865 4.7 A 1.94
C 28 3.34 i 0.61515 3.3 4 0.93
B 17 4.8 i0.84612 3.0 + 0.735
A 21 6.2 + 0.96010 3.9 0.88
F 5 8.2 2.789 2.7 + 1.26
1 20 11.5 : 1.17
"The data given here represent the results ofover 3,000 assays.
b Arranged in order of increasing distance ofthe corresponding genes from the operator.
c Time of change in differential rate of synthesisafter addition of histidine to the culture medium.Expressed in terms of 95% confidence limit.
d In this category are included experimentswith organisms in which there is no known altera-tion in the allosteric site of the G enzyme (withthe exception of hisG1306, in which the feedbacksite of the G enzyme is hypersensitive to inhibi-tion by histidine). In every experiment, the pat-tern of repression was serial.
e In this category are included experimentswith organisms in which the allosteric site of theG enzyme has been either inhibited with thiazol-alanine or altered by mutation. In every experi-ment the pattern of repression was concomitant.
end. Each portion of the message stops beingtranslated at a time which is directly related to itsdistance from the operator. This can be explainedmost simply as follows: at the time or repression,all ribosomes engaged in translating the histidinemessage continue to the distal end but translationof this message from the operator end by otherribosomes is prohibited. This idea, previouslysuggested by Ames and Hartman (3), is consistentwith the recent findings of Morse, Baker, andYanofsky (20) on the tryptophan system ofEscherichia coli.Concomitant repression apparently represents
an alteration of the normal manner in whichtranslation of the histidine mRNA ceases. In thisaltered form of repression, all cistrons of themessage stop being translated at essentially thesame time. Our data do not allow us to specifywith certainty the mechanism responsible for thisconcomitant cessation of translation.
1288 J. BACrl3RIOL.
on March 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

REPRESSION OF THE HISTIDINE OPERON
Whatever may be the difference between thetwo mechanisms responsible for the normal, serialpattern and the altered, concomitant pattern ofrepression, the molecular basis for the differencemust involve the first enzyme for histidine bio-synthesis. Theoretically, this involvement couldbe realized either through the catalytic activity ofthe enzyme or through some other activity of theenzyme. The possibility that the catalytic activityof first enzymes of metabolic pathways may beinvolved in control has recently been stressed byTomkins and Ames (29). They discussed the idea,termed "metabolite induction" (29), that in cer-tain bacterial operons derepression can occur onlywhen there is a shortage of the end product of thepathway, when substrate for the pathway is pres-ent, and when the first enzyme of the pathway iscatalytically active (not inhibited by the end prod-uct). However, it appears unlikely to us that thecatalytic activity of the first enzyme is involved indetermining the pattern of repression, becausewe do not see a consistent correlation between thelevel of intermediates of the histidine pathway andthe pattern of repression. For example, hisG52produces none of the intermediates of the path-way, yet displays either mode of repression, de-pending upon the presence or absence of thiazol-alanine. We favor the view that some other ac-tivity of the first enzyme is involved in determin-ing the pattern of repression, an activity which isrelated to, but not identical with, feedback in-hibition. In keeping with this view is the consis-tent correlation between the pattern of repressionand the functional state of the feedback site of theenzyme. Whenever this site is functionally im-paired or is occupied by thiazolalanine, the patternof repression is concomitant; whenever this site isintact and not occupied by thiazolalanine, thepattern of repression is serial.
It is notable that in three of the systems foramino acid biosynthesis (histidine, tryptophan,and leucine), the first structural gene of the operon(that is, the gene closest to the operator) specifiesthe structure of the first and feedback-sensitive,enzyme of the biosynthetic pathway. This correla-tion between first gene and first enzyme is un-likely to be fortuitous. It is more reasonable toassume that this correlation is a consequence ofevolutionary pressures. Perhaps some survivalvalue was achieved by a regulatory mechanismwhich requires that the newly synthesized firstenzyme be located close to the operator end of themRNA. Regulatory mechanisms involving thefeedback sites of enzymes encoded in the firstgenes of operons have been discussed on theoreti-cal grounds by Gruber and Campagne (11), Maasand McFall (16), Cline and Bock (8), and Kosh-land and Kirtley (13). The findings reported here
suggest that the functional state of the feedbacksite of the first enzyme for histidine biosynthesisinfluences, directly or indirectly, the mechanismby which translation of the histidine mRNAceases. Although the physiological significance ofthis phenomenon is not yet understood, it sug-gests that the first enzyme may play a previouslyunrecognized role in regulation of the histidinesystem.
ACKNOWLEDGMENTSWe acknowledge the excellent technical assistance of Marilyn
Meyers. We also thank C. B. Anfinsen for his support and en-couragement, and B. N. Ames, G. F-L. Ames, P. E. Hartman,M. Levinthal, R. G. Martin, G. M. Tomkins, and H. J. Whitfield,Jr., for their participation in many helpful discussions and fortheir critical reading of the manuscript.
LIERATURE CITED1. Ames, B. N., and B. J. Garry. 1959. Coordinate repression of
the synthesis of four histidine biosynthetic enzymes byhistidine. Proc. Natl. Acad. Sci. U.S. 45:1453-1461.
2. Ames, B. N., B. Garry, and L. A. Herzenberg. 1960. Thegenetic control of the enzymes of hisddine biosynthesis inSalmonella typhimurium. J. Gen. Microbiol. 22:369-378.
3. Ames, B. N., and P. E. Hartman. 1963. The histidine operon.Cold Spring Harbor Symp. Quant. Biol. 28:349-356.
4. Ames, B. N., P. E. Hartman, and F. Jacob. 1963. Chromo-somal alterations affecting the regulation of histidinebiosynthetic enzymes in Salmonella. J. Mol. Biol. 7:23-42.
5. Ames, B. N., R. G. Martin, and B. Garry. 1961. The firststep of histidine biosynthesis. J. Biol. Chem. 236:2019-2026.
6. Berberich, M. A., J. S. Kovach, and R. F. Goldberger. 1967.Chain initiation in a polycistronic message: sequentialversus simultaneous derepression of the enzymes forhistidine biosynthesis in Salmonella typhimurium. Proc.Natl. Acad. Sci. U.S. 57:1857-1864.
7. Berberich, M. A., P. Venetianer, and R. F. Goldberger. 1966.Alternative modes of derepression of the histidine operonobserved in Salmonella typhimurium. J. Biol. Chem.241:4425-4433.
8. Cline, A. L., and R. M. Bock. 1966. Translational control ofgene expression. Cold Spring Harbor Symp. Quant. Biol.31:321-333.
9. Fink, G. R., T. Klopotowski, and B. N. Ames. 1967. Histi-dine regulatory mutants in Salmonella typhimurium. IV. Apositive selection for polar histidine-requiring mutantsfrom histidine operator constitutive mutants. J. Mol. Biol.30:81-95.
10. Fink, G. R., and R. G. Martin. 1967. Translation and polar-ity in the histidine operon. II. Polarity in the histidineoperon. J. Mol. Biol. 30:97-107.
11. Gruber, M,, and R. N. Campagne. 1965. Regulation of pro-tein synthesis: an alternative to the repressor-operator hy-pothesis. Koninkl. Ned. Akad. Wetenschap. Proc. Ser. C68:270-276.
12. Hilton, J. L., P. S. Keamery, and B. N. Ames. 1965. Mode ofaction of the herbicide, 3-amino-1-2-4-triazole (amitrole):inhibition of an enzyme of histidine biosynthesis. Arch.Biochem. Biophys. 112:544-547.
13. Koshland, D. E., Jr., and M. E. Kirtley. 1966. Protein struc-ture in reiation to cell dynamics and differentiation, p. 217-249. In M. Locke (ed.), Major problems in developmentalbiology, Academic Press Inc., New York.
14. Loper, J. C., M. Grabnar, R. C. Stahl, Z. Hartman, andP. E. Hartman. 1964. Genes and proteins involved in histi-dine biosynthesis in Salmonella. Brookhaven Symp. Biol.17:15-50.
1289VOL. 97, 1969
on March 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

KOVACH ET AL.
15. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Ran-dall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
16. Maas, W., and E. McFall. 1964. Genetic aspects of metaboliccontrol. Ann. Rev. Microbiol. 18.95-110.
17. Margolies, M. N., and R. F. Goldberger. 1966. Isolation ofthe fourth enzyme (isomerase) of histidine biosynthesisfrom Salmonella typhimurlum. J. Biol. Chem. 241:3262-3269.
18. Martin, R. G. 1963. The first enzyme in histidine biosynthe-sis: the nature of feedback inhibition by histidine. J. Biol.Chem. 238:257-268.
19. Martin, R. G. 1963. The one operon-one messenger theoryof transcription. Cold Spring Harbor Symp. Quant. Biol.28:357-361.
20. Morse, D. E., R. F. Baker, and C. Yanofsky. 1968. Transla-tion of the tryptophan messenger RNA of Escherichla coll.Proc. Natl. Acad. Sci. U.S. 60:1428-1435.
21. Moyed, H. S. 1961. Interference with the feed-back controlof histidine biosynthesis. J. Biol. Chem. 236:2261-2267.
22. Moyed, H. S., and B. Magasanik. 1960. The biosynthesis ofthe imidazole ring of histidine. J. Biol. Chem. 235:149-153.
23. Roth, J. R., and B. N. Ames. 1966. Histidine regulatory mu-tants in Salmonella typhlmurlum. II. Histidine regulatorymutants having altered histidyl-tRNA synthetase. J. Mol.Biol. 22:325-334.
24. Roth, J. R., D. N. Anton, and P. E. Hartman. 1966. Histi-dine regulatory mutants in Salmonella typhimurium. I. Iso-lation and general properties. J. Mol. Biol. 22:305-323.
25. St. Pierre, M. L. 1968. Mutations creating a new initiationpoint for expression of the histidine operon in Salmonellatyphimurium. J. Mol. Biol. 35:1-12.
26. Sheppard, D. W. 1964. Mutants of Salmonella typhlmuriumresistant to feedback inhibition by L-histidine. Genetics20:611-623.
27. Smith, D. W. E., and B. N. Ames. 1964. Intermediates inthe early steps of histidine biosynthesis. J. Biol. Chem.239:1848-1855.
28. Smith, D. W. E., and B. N. Ames. 1965. Phosphoribosylade-nosine monophosphate, an intermediate in histidine bio-synthesis. J. Biol. Chem. 240:3056-3063.
29. Tomkins, G. M., and B. N. Ames. 1966. The operon conceptin bacteria and in higher organisms. Natl. Cancer Inst.Monograph 27, p. 221-234.
30. Venetianer, P., M. A. Berberich, and R. F. Goldberger.1968. Studies on the size of the messenger RNA transcribedfrom the histidine operon during simultaneous and sequen-
tial derepression. Biochim. Biophys. Acta 166:124-133.31. Vogel, H. J., and B. M. Bonner. 1956. Acetylornithinase of
Escherlchla colf: partial purification and some properties.J. Biol. Chem. 218:97-106.
1290 J. BACTERIOL.
on March 16, 2018 by guest
http://jb.asm.org/
Dow
nloaded from





![Research Article CrystalStructureofL-Histidinium2 ...chloride monohydrate [2], L-histidine tetrafluoroborate [3], L-histidine hydrochloride monohydrate [4], L-histidine hydrofluoride](https://img.pdfslide.us/doc/110x75/60b51c180636315681384205/research-article-crystalstructureofl-histidinium2-chloride-monohydrate-2.jpg)























