Embed Size (px)
Citation preview

Repin-Induced Neurotoxicity in Rodents1
Mario Robles, Ben H. Choi,2 Bora Han, Karen Santa Cruz, and Ronald C. KimDivision of Neuropathology, Department of Pathology, University of California, Irvine, California 92697-4800
Received December 10, 1997; accepted March 21, 1998
Russian knapweed is a perennial weed found inmany parts of the world, including southern Califor-nia. Chronic ingestion of this plant by horses has beenreported to cause equine nigropallidal encephalomala-cia (ENE), which is associated with a movement disor-der simulating Parkinson’s disease (PD). Repin, a prin-cipal ingredient purified from Russian knapweed, is asesquiterpene lactone containing an a-methylenebu-tyrolactone moiety and epoxides and is a highly reac-tive electrophile that can readily undergo conjugationwith various biological nucleophiles, such as proteins,DNA, and glutathione (GSH). We show in this studythat repin is highly toxic to C57BL/6J mice and Spra-gue–Dawley rats and acutely induces uncoordinatedlocomotion associated with postural tremors, hypother-mia, and inability to respond to sonic and tactilestimuli. We also show that repin intoxication reducesstriatal and hippocampal GSH and increases totalstriatal dopamine (DA) levels in mice. Striatal microdi-alysis in rats, however, has demonstrated a significantreduction of extracellular DA levels. These findings,coupled with the absence of any demonstrable changein striatal DOPAC levels, suggest that repin acts byinhibiting DA release, a hypothesis that is furthersupported by our demonstration that, in cultured PC12cells, repin inhibits the release of DA without affectingits uptake. We believe, therefore, that inhibition of DArelease represents one of the earliest pathogeneticevents in ENE, leading eventually to striatal extra-cellular DA denervation, oxidative stress, and degen-eration of nigrostriatal pathways. Since the neuro-toxic effects of repin appear to be mediated viaoxidative stress, and since repin is a natural productisolated from a plant in our environment that cancause a movement disorder associated with degenera-tion of nigrostriatal pathways, clarification of the mech-anism of repin neurotoxicity may provide new insightsinto our understanding of the pathogenesis of PD. r 1998
Academic Press
Key Words: Parkinson’s disease; Russian knapweed;repin; sesquiterpene lactone; neurotoxicity; rodents;glutathione; oxidative stress.
INTRODUCTION
Repin is a novel natural compound extracted andpurified from Russian knapweed (Centaurea repens), aperennial weed that is rapidly taking over pastures inmany parts of the world due to its extensive rootletsystem and its ability to withstand harsh environmen-tal conditions (30). Ingestion of this weed by horses hasbeen reported to cause equine nigropallidal encephalo-malacia (ENE), a condition that is associated withsigns and symptoms similar to those of PD (3, 4, 9, 34).The disorder is characterized by a fixed ‘‘wooden’’ facialexpression with hypertonia of the muscles of the muzzleand face, idle chewing and tongue flicking, and im-paired eating and drinking, followed by hypokinesiaand death. Neuropathological examination revealedbilateral necrosis of the anterior globus pallidus andthe substantia nigra (SN). An almost identical disorderdevelops in horses after ingestion of a similar kind ofweed called yellow star thistle (17). The biologicallyactive ingredient in these plants is a sesquiterpenelactone (SQL), and repin is the principal SQL, represent-ing approximately 1% of the dry weight of Russianknapweed. The chemical structure of repin is character-ized by the presence of epoxides and an a-methylenebu-tyrolactone moiety (Fig. 1). Thus, repin is a highlyreactive electrophile that can readily undergo conjuga-tion with various biological nucleophiles, such as DNA,proteins, and glutathione (GSH).
Previous studies in our laboratory have shown thatPC12 cells and cultured neonatal mouse astrocytesundergo dose-dependent reduction in both cell viabilityand GSH levels following repin exposure (24, 26). Inaddition, repin was shown to enhance significantly therate of reactive oxygen species (ROS) production inPC12 cells (26, 29) as determined by the dichlorofluores-cein diacetate method (1). These findings suggestedthat oxidative stress may play a prominent role in themechanism of repin toxicity.
1 Presented in part at the 71st annual meeting of the AmericanAssociation of Neuropathologists, San Antonio, TX.
2 To whom correspondence and reprint requests should be ad-dressed. Fax: (714) 824-2160. E-mail: [email protected].
EXPERIMENTAL NEUROLOGY 152, 129–136 (1998)ARTICLE NO. EN986826
129 0014-4886/98 $25.00Copyright r 1998 by Academic Press
All rights of reproduction in any form reserved.

There has been increasing interest in searching forenvironmental neurotoxins that may cause or contrib-ute to the development of PD (32). The discovery that1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)is the toxic agent responsible for the sudden onset of aparkinsonian syndrome and that MPTP selectivelydamages nigral neurons (14, 15) provided a new andchallenging impetus to the field. Although the precisemechanism by which ingestion of Russian knapweedinduces a disorder simulating PD remains unclear, thefact that chronic exposure to a natural product in theenvironment can lead to a movement disorder associ-ated with degeneration of neurons in the SN and basalganglia is worthy of further investigation. Accordingly,we have extracted and purified repin from Russianknapweed and examined its neurotoxic effects in orderto clarify the pathogenesis of ENE.
MATERIALS AND METHODS
Isolation and Purification of Repin
Russian knapweed was collected from Riverside andIrvine, CA. Repin was extracted and purified accordingto the method of Robles (1995). Briefly, the aerial partsof Centaurea repens (250 g) were extracted twice with 2L of chloroform, filtered, and concentrated in a rotatoryevaporator. The crude extract was then dissolved inmethanol, cooled to 4.0°C for 20 min and filtered toremove waxes and other nonpolar substances. Thisextract was placed onto a Sephadex LH-20 gel andeluted with methanol. Fractions were collected andcombined according to their respective thin layer chro-matography (TLC) Rf values in a 7:3 chloroform-methanol solvent system. Further purification wasachieved using silica gel column chromatography withan eluting gradient of chloroform to 100% methanol.Preparatory TLC was used for the final purification ofrepin using a 6:4 chloroform–acetone solvent system.Repin was dissolved in 0.1% DMSO for experimentaluse. Best yield of repin is obtained from Centaurea
repens harvested at spring time when the plant beginsto bloom. In general, approximately 200 mg of repin canbe purified from 250 g of well-dried Centaurea repens.
Animals
C57BL/6J mice weighing 25–30 g and Sprague–Dawley rats weighing 270–300 g, obtained from Simon-sen (Gilroy, CA), were used. Animals were housed in avivarium and cared for in accordance with the GuidingPrinciples in the Use of Animals in Toxicology. Animalexperiments were conducted with approval from theInstitutional Animal Care & Use Committee of theUniversity of California (Irvine).
Experiments
Mice were injected intraperitoneally (IP) with 10 and20 mg/kg body weight of repin and total brain GSHlevels were determined 3, 6, 9, and 12 h later. Toexamine the effects of repin on GSH levels in specificregions of the brain, IP injections of repin (60 mg/kg)were made and the GSH levels in the striatum, hippo-campus, cerebral cortex, cerebellum, and brain stemdetermined at 6 h. The levels of thiobarbiturate-reactive substances (TBARS) were also determined invarious regions of the rat brain, after IP injection ofrepin (20 mg/kg), at 6 and 12 h.
Striatal DA and DOPAC levels were determined aftervarying doses of repin in mice following either IPinjection (20 or 10 mg/kg per day for 3 days) oradministration via gastric tube (20 or 10 mg/kg every 2days for 6 days and 10 mg/kg twice a day for 14 days),using high performance liquid chromatography (HPLC)coupled with electrochemical (EC) detection at 6 h.
We used microdialysis to examine the effects of repinon extracellular DA levels in the striatum. Sprague–Dawley rats were anesthetized with phenobarbital andmounted on a stereotaxic apparatus and prepared forintracranial microdialysis brain perfusion.Amicrodialy-sis probe (Carnegie Medicine CMA/10, Solma, Sweden)was washed for 30 min with Ringer’s solution prior toits implantation in the striatum. A hole was drilled atthe top of the cranium using the following coordinates:AP 5 4.0 mm, LM 5 3.5 mm, VD 5 2.5 mm from thebregma (19). The microdialysis probe was inserted andthe striatum perfused with Ringer’s solution (1.0 µl/min) for at least 60 min before repin injection (20mg/kg, IP). The dialysate was collected every 20 mininto 5 µl of 0.1 M perchloric acid and analyzed byHPLC-EC.
At least five animals were used for each category ofdeterminations in all experimental and control groups.Control animals received physiological saline in placeof repin. Motor activities and other behavioral abnor-malities were recorded on video. Following anesthesia,mice were perfused with physiological saline and the
FIG. 1. Chemical structure of repin. Repin is a highly reactiveelectrophile that contains an a-methylenebutyrolactone moiety (en-closed in a square) with an exocyclic double bond (arrow). Thepresence of two epoxides (empty arrows) is also shown.
130 ROBLES ET AL.

brains rapidly excised. Selected regions of the brainwere dissected out under a dissecting microscope andquickly frozen in liquid nitrogen until used.
DA and DOPAC Assays
The striatum dissected was weighed, immediatelyfrozen in liquid nitrogen, and stored at 270°C untilanalyzed. The tissue was homogenized in 200 µl ofice-cooled 0.1 M perchloric acid containing 0.1% ofdisodium EDTA and 10% sodium metabisulfite. Thehomogenates were centrifuged and filtered through0.2-µm microfilter tubes. The supernatants were ana-lyzed for DA and DOPAC using reverse-phase HPLCcoupled with EC detection. The HPLC system consistedof a Rainin solvent delivery pump (Model HPLC, Rainin,Woburn, MA), a 6-port injector (Model 7125, Rheodyne,Berkeley, CA), and an analytical column UltrasphereODS-C18, 15 cm 3 4.6 mm i.d., particle size 3 mm,ESA, Bedford, MA). An electrochemical detector (Coulo-chem ESA detector Model 5100A), an analytical cell(Model 5011, ESA) set at 10.22 V, and a guard cell(Model 5020, ESA) set at 10.25 V were used. Themobile phase for HPLC consisted of 100 mM sodiumphosphate, 0.1 mM EDTA, 1.0 mM heptanesulfonicacid, 10% v/v acetonitrile, and 0.01% triethylamine (pH3.0, adjusted with phosphoric acid). The solution wasfiltered and degassed before being passed through thecolumn at an isocratic flow rate of 0.6 ml/min. Quantita-tive determination of neurochemical levels was per-formed using Dynamax HPLC Method Manager soft-ware (Rainin, Woburn, MA), comparing sample peakareas with those of known standards under identicalconditions.
GSH Assay
Total GSH was determined using the Tietze method(33). Briefly, samples of tissue homogenates were mixedwith GSH reductase (100 I.U./mg) in the presence of200 µM Ellman’s reagent [5,58-dithiobis-(2-nitroben-zoic acid)]. Measurements of absorbance changes at 412nm were made for 1 min using a Gilford spectrophotom-eter. All reactions were carried out at 25°C. Total GSHconcentrations were expressed as mM/g of protein.Protein content was measured by the method of Lowryet al. (16).
Lipid Peroxidation Products Assay
TBARS were measured according to the method ofOhkawa et al. (18). Briefly, tissue samples were homog-enized in 0.1 g/ml Tris-phosphate buffer, incubated at37°C for 1 h, and mixed with 40% TCA and freshlymade 0.2% TBA. Samples were then boiled for 15 minand mixed with 70% TCA. MDA standards were pre-
pared at 10, 20, 50, and 100 nM concentrations. Sampleswere read at 532 nm absorbance.
Histological and Immunocytochemical Studies
Brain tissues fixed in 10% formalin were embeddedin paraffin. Representative sections of brains obtainedfrom at least five animals each of both control andrepin-intoxicated mice 6 h after IP injection of repin(10, 20, and 10 mg/kg/day for 3 days) were examinedafter staining with hematoxylin and eosin, Biel-schowsky and Luxol Fast Blue-cresyl-violet stains.Additional sections were processed for immunoperoxi-dase staining for glial fibrillary acidic protein (GFAP),tyrosine hydroxylase (TH), and TdT-mediated dUTPnick end labeling (TUNEL).
DA Release and Uptake Studies Using PC12 Cells
PC12 cells were obtained from ATCC (Rockville,MD). Growth medium consisted of Dulbecco’s modifiedEagle’s medium (DMEM) with high glucose (IrvineScientific), supplemented with 5% horse serum, 10%fetal calf serum, and 1% penicillin/streptomycin. Thecells were grown in an incubator in an atmosphere of95% air and 5% CO2 maintained at 37°C.
DA release by PC12 cells was determined by incubat-ing cells (1–2 3 106/ml) with 5, 10, and 20 µM repin for60 min, either (1) in normal culture medium or (2) inKrebs-Ringer (KR) buffer containing Ca21 (1.2 mM).Following incubation, the supernatant was separatedby centrifugation and used for measurement of DArelease. A 50-µl aliquot of ice-cold antioxidant solutioncontaining 0.1% sodium metabisulfite and 0.5% EDTAin 0.1 M perchloric acid was added to 450 µl of sample.The cell pellet was lysed with antioxidant solution tomeasure the intracellular DA. The cell lysates weremeasured using an HPLC coupled to an electrochemi-cal detector. DA release was expressed as a percentageof the transmitter secreted into the media relative tothe total cellular content. DA uptake was examined inPC 12 cells following incubation with 10–100 nM[3H]DA (22.1 Ci/mmol, Dupont, New England NuclearProducts, Boston, MA) in 1.0 ml of KR buffer atapproximately 2 3 106 cells/ml, and aliquotting into1.7-ml microfuge tubes for 10 min at 37°C. Subse-quently, samples were centrifuged and washed with 2.0ml of ice-cold KR buffer. The radioactivity remaininginside the cells was extracted and measured in a liquidscintillation counter.
Statistical Analyses
Differences between experimental groups were as-sessed by Student’s t test. The acceptance level of
131REPIN NEUROTOXICITY
significance was P , 0.05, using a two-tailed distribu-tion.
RESULTS
Preliminary tests revealed that a single IP bolusinjection of repin at a dosage of 50 mg/kg body weight oradministration of repin at 100 mg/kg body weightthrough a gastric tube was lethal after 12 to 24 h inC57BL/6J mice. Heavily intoxicated mice showedmarked reduction in motor activity, ataxia, hypother-mia, loss of appetite, and lethargy followed by death.Twitching of the limbs was frequently observed duringthe first 3 h. Repin administration through a gastrictube or by IP injection produced similar behavioralabnormalities initially, but the animals soon recovered.Sprague–Dawley rats showed more severe toxic signsas compared to those of mice and demonstrated pecu-liar postures and locomotor disturbances characterizedby dragging of the hind limbs and/or turning overbackward within 15 min of repin injection; markedhypothermia was also observed.
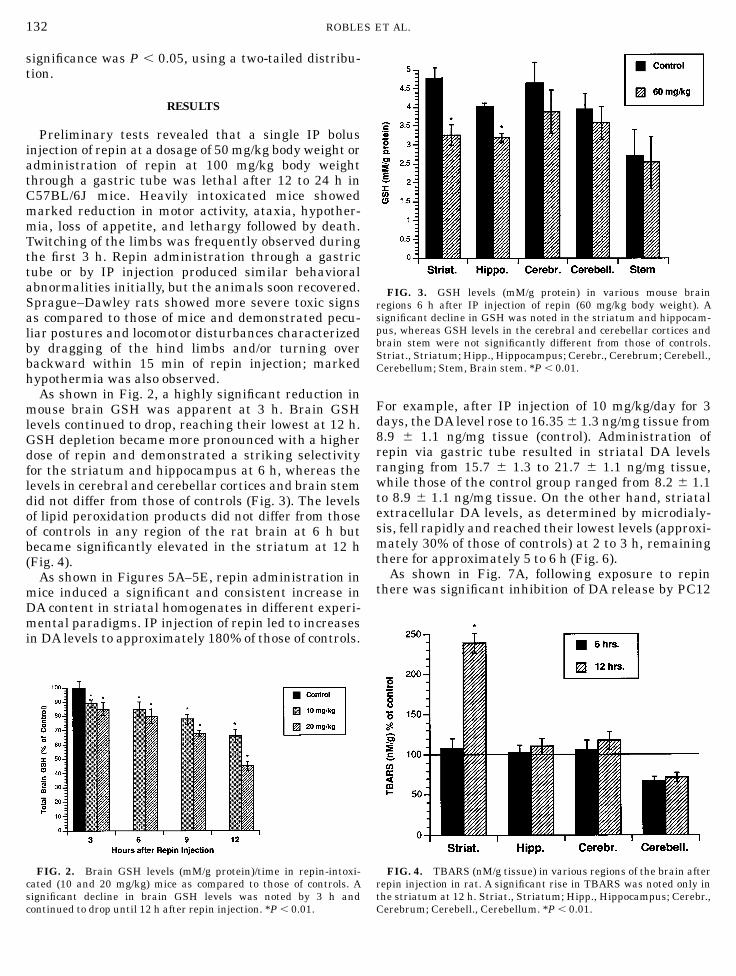
As shown in Fig. 2, a highly significant reduction inmouse brain GSH was apparent at 3 h. Brain GSHlevels continued to drop, reaching their lowest at 12 h.GSH depletion became more pronounced with a higherdose of repin and demonstrated a striking selectivityfor the striatum and hippocampus at 6 h, whereas thelevels in cerebral and cerebellar cortices and brain stemdid not differ from those of controls (Fig. 3). The levelsof lipid peroxidation products did not differ from thoseof controls in any region of the rat brain at 6 h butbecame significantly elevated in the striatum at 12 h(Fig. 4).
As shown in Figures 5A–5E, repin administration inmice induced a significant and consistent increase inDA content in striatal homogenates in different experi-mental paradigms. IP injection of repin led to increasesin DA levels to approximately 180% of those of controls.
For example, after IP injection of 10 mg/kg/day for 3days, the DA level rose to 16.35 6 1.3 ng/mg tissue from8.9 6 1.1 ng/mg tissue (control). Administration ofrepin via gastric tube resulted in striatal DA levelsranging from 15.7 6 1.3 to 21.7 6 1.1 ng/mg tissue,while those of the control group ranged from 8.2 6 1.1to 8.9 6 1.1 ng/mg tissue. On the other hand, striatalextracellular DA levels, as determined by microdialy-sis, fell rapidly and reached their lowest levels (approxi-mately 30% of those of controls) at 2 to 3 h, remainingthere for approximately 5 to 6 h (Fig. 6).
As shown in Fig. 7A, following exposure to repinthere was significant inhibition of DA release by PC12
FIG. 2. Brain GSH levels (mM/g protein)/time in repin-intoxi-cated (10 and 20 mg/kg) mice as compared to those of controls. Asignificant decline in brain GSH levels was noted by 3 h andcontinued to drop until 12 h after repin injection. *P , 0.01.
FIG. 3. GSH levels (mM/g protein) in various mouse brainregions 6 h after IP injection of repin (60 mg/kg body weight). Asignificant decline in GSH was noted in the striatum and hippocam-pus, whereas GSH levels in the cerebral and cerebellar cortices andbrain stem were not significantly different from those of controls.Striat., Striatum; Hipp., Hippocampus; Cerebr., Cerebrum; Cerebell.,Cerebellum; Stem, Brain stem. *P , 0.01.
FIG. 4. TBARS (nM/g tissue) in various regions of the brain afterrepin injection in rat. A significant rise in TBARS was noted only inthe striatum at 12 h. Striat., Striatum; Hipp., Hippocampus; Cerebr.,Cerebrum; Cerebell., Cerebellum. *P , 0.01.
132 ROBLES ET AL.
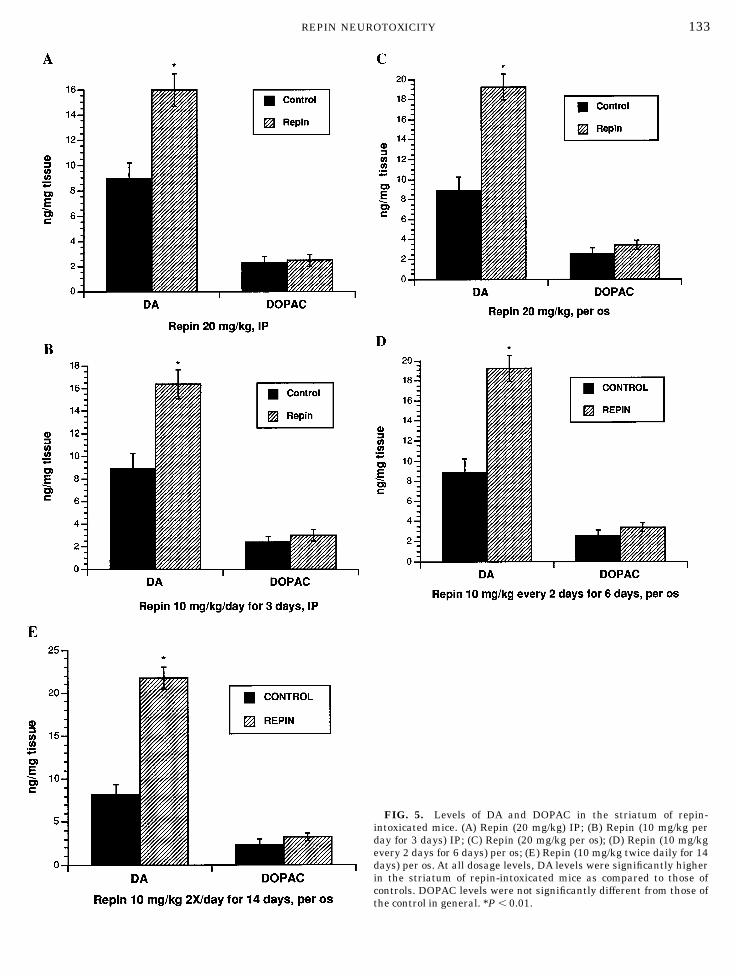
FIG. 5. Levels of DA and DOPAC in the striatum of repin-intoxicated mice. (A) Repin (20 mg/kg) IP; (B) Repin (10 mg/kg perday for 3 days) IP; (C) Repin (20 mg/kg per os); (D) Repin (10 mg/kgevery 2 days for 6 days) per os; (E) Repin (10 mg/kg twice daily for 14days) per os. At all dosage levels, DA levels were significantly higherin the striatum of repin-intoxicated mice as compared to those ofcontrols. DOPAC levels were not significantly different from those ofthe control in general. *P , 0.01.
133REPIN NEUROTOXICITY
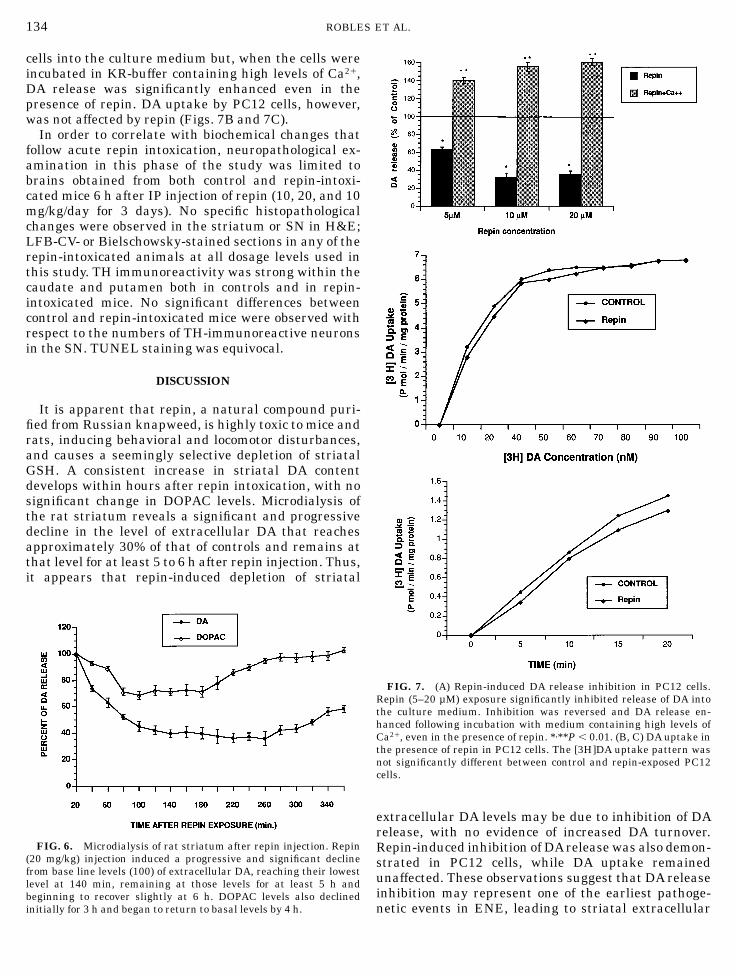
cells into the culture medium but, when the cells wereincubated in KR-buffer containing high levels of Ca21,DA release was significantly enhanced even in thepresence of repin. DA uptake by PC12 cells, however,was not affected by repin (Figs. 7B and 7C).
In order to correlate with biochemical changes thatfollow acute repin intoxication, neuropathological ex-amination in this phase of the study was limited tobrains obtained from both control and repin-intoxi-cated mice 6 h after IP injection of repin (10, 20, and 10mg/kg/day for 3 days). No specific histopathologicalchanges were observed in the striatum or SN in H&E;LFB-CV- or Bielschowsky-stained sections in any of therepin-intoxicated animals at all dosage levels used inthis study. TH immunoreactivity was strong within thecaudate and putamen both in controls and in repin-intoxicated mice. No significant differences betweencontrol and repin-intoxicated mice were observed withrespect to the numbers of TH-immunoreactive neuronsin the SN. TUNEL staining was equivocal.
DISCUSSION
It is apparent that repin, a natural compound puri-fied from Russian knapweed, is highly toxic to mice andrats, inducing behavioral and locomotor disturbances,and causes a seemingly selective depletion of striatalGSH. A consistent increase in striatal DA contentdevelops within hours after repin intoxication, with nosignificant change in DOPAC levels. Microdialysis ofthe rat striatum reveals a significant and progressivedecline in the level of extracellular DA that reachesapproximately 30% of that of controls and remains atthat level for at least 5 to 6 h after repin injection. Thus,it appears that repin-induced depletion of striatal
extracellular DA levels may be due to inhibition of DArelease, with no evidence of increased DA turnover.Repin-induced inhibition of DA release was also demon-strated in PC12 cells, while DA uptake remainedunaffected. These observations suggest that DA releaseinhibition may represent one of the earliest pathoge-netic events in ENE, leading to striatal extracellular
FIG. 6. Microdialysis of rat striatum after repin injection. Repin(20 mg/kg) injection induced a progressive and significant declinefrom base line levels (100) of extracellular DA, reaching their lowestlevel at 140 min, remaining at those levels for at least 5 h andbeginning to recover slightly at 6 h. DOPAC levels also declinedinitially for 3 h and began to return to basal levels by 4 h.
FIG. 7. (A) Repin-induced DA release inhibition in PC12 cells.Repin (5–20 µM) exposure significantly inhibited release of DA intothe culture medium. Inhibition was reversed and DA release en-hanced following incubation with medium containing high levels ofCa21, even in the presence of repin. *,**P , 0.01. (B, C) DA uptake inthe presence of repin in PC12 cells. The [3H]DA uptake pattern wasnot significantly different between control and repin-exposed PC12cells.
134 ROBLES ET AL.

DA depletion, oxidative stress, and eventual degenera-tion of nigrostriatal pathways.
Although it remains to be determined through whatmechanism repin inhibits DA release, it is possible thata similar sequence of events may take place in PD.Although clues to the etiology of PD have generallybeen sought within the SN because of the presence, atthat site, of intense neuronal loss and Lewy inclusionbody formation, it is quite possible that the diseaseprocess originates in the terminal portion of the nigro-striatal projection (8).
Postmortem biochemical measurements in PD brainshave shown a reduction in mitochondrial GSH in theSN but not in other brain regions (20, 22), associatedwith an increase in lipid peroxides (5, 11). Further-more, it has been reported that the reduction of GSH inthe SN correlated with the severity of the PD (22). Anincrease in the concentration of nonheme iron in the SNof PD patients has also been reported (5, 6, 22). All ofthese findings support the concept that oxidative stressmay play a role in the degeneration of nigral neurons inPD.
The biochemical changes observed following repinintoxication, namely, depletion of GSH and an increasein lipid peroxidation products in defined regions of thebrain, also suggest a pathogenetic role for oxidativestress. Our previous finding of dose-dependent cytotox-icity in PC12 cells associated with a marked reductionin GSH levels and a significant enhancement in therate of ROS production following repin exposure (26,29) supports this contention. Furthermore, enzymaticand nonenzymatic oxidative deamination of accumu-lated striatal DA due to inhibition of its release is likelyto contribute to oxidant stress and may possibly lead toretrograde degeneration of the SN neurons.
A number of neurotoxic compounds, such as6-hydroxydopamine and MPTP, are known to induceneuronal degeneration via oxidative stress. It has beenamply demonstrated that MPTP causes selective de-struction of dopaminergic neurons due to the actions ofits toxic byproduct, 1-methyl-4-phenylpyridinium(MPP1). MPP1 is a free radical generated by the actionof monoamine oxidase within glial cells that inhibitsthe activity of complex I of the mitochondrial electrontransport chain. Mitochondrial complex I deficiencyhas also been described in a familial form of PD (28).The strong affinity of repin for sulfhydryl radicals andits ready conjugation with GSH appear to play a role inrepin toxicity, but depletion of GSH may not be the solecause of its toxic action (26). It has been suggested thatSQLs induce toxic effects through depletion of nonpro-tein thiols within tissues (13). Once depleted of protec-tive GSH, tissues would then become more susceptibleto secondary insult by SQLs.
Several SQLs have been identified as highly toxic,causing allergic contact dermatitis (25, 27) or lethality
(7, 12). The neurotoxic effects of sesquiterpenoids werealso demonstrated using cultures of central nervoussystem cells. For example, in the presence of 80 nMrepin, neurite extension of chick embryo sensory neu-rons was markedly inhibited (30). Sesquiterpenoidsfrom yellow star thistle also were reported to causesevere neurotoxic effects in primary cultures of fetal ratbrain cells (2, 35). Since many vegetables, includinglettuce and artichokes, contain SQLs, the population isexposed to many different types and amounts of SQLs.Lettuce is known to be affected by at least 11 viraldiseases (23), and two multiply virus-resistant wildspecies commonly used in lettuce breeding programshave been found to contain rather high levels of threemajor SQLs. However, lettuce lines that derived fromL. saligna appear to contain much smaller amounts ofSQLs, and yet show equally effective resistance toviruses (31). In view of the potential toxicity of SQLs,the content and type of SQLs in plants and vegetables,including lettuce, should be of concern to breeders andconsumers alike.
In our study, morphological evaluation of repin-intoxicated mouse brains revealed no readily apparentdegeneration of TH-positive neurons as compared tothose of controls. This is presumably due to the rela-tively short interval of time between repin intoxicationand sacrifice. In a separate study, we have describedapoptosis in repin-exposed PC12 cells and its modifica-tion by thiol-containing antioxidants, such as GSH-glycoside and lipoic acid (10). It remains to be deter-mined whether or not chronic repin intoxication willinduce specific morphological changes in the SN or inother brain regions associated with depletion of DAand/or movement disorders.
Although it is premature to state that the neurotoxiceffects of repin can be used as an animal model to studythe pathogenesis of PD, since the acute toxic effects ofrepin are exerted through oxidative stress in a mannerhighly selective to nigrostriatal dopaminergic path-ways, and since repin is a purified compound extractedfrom a plant in our environment that is capable ofinducing movement disorders in horses simulating PD,further studies of the morphologic, biochemical, andfunctional effects of repin may prove useful in elucidat-ing mechanisms of neuronal death and in assessing therole of possible environmental factors in PD.
ACKNOWLEDGMENTS:
Authors thank Drs. A. G. Kanthasamy and D. Truong for technicalassistance and helpful discussions related to the project.
REFERENCES
1. Bondy, S. C., S. F. Ali, and C. P. LeBel. 1992. Reactive oxygenspecies formation as a biomarker of methylmercury and tri-methyltin neurotoxicity. Neurotoxicology 13: 637–648.
135REPIN NEUROTOXICITY

2. Cheung, C. H. K., B. Costall, M. Hamburger, K. Hostettmann,R. J. Naylor, Y. Wang, and P. Jenner. 1992. Toxic effects ofsolstitialin a 13-acetate and cynaropicrin from centaurea solsti-tialis L. (Asteraceae) in cell cultures of foetal rat brain. Neuro-pharmacology 31: 271–277.
3. Cordy, D. R. 1954. Nigropallidal encephalomalacia in horsesassociated with ingestion of yellow star thistle. J. Neuropathol.Exp. Neurol. 13: 330–342.
4. Cordy, D. R. 1978. Centaurea species and equine nigropallidalencephalomacia. In Effects of Poisonous Plants on Livestock(R. F. Keeler, K. R. Van Kampen, and L. F. James, Eds.), pp.327–336. Academic Press, New York.
5. Dexter, D. T., C. J. Carter, R. R. Wells, A. J. Lees, F. Agid, Y. Agid,P. Jenner, and C. D. Marsden. 1989. Increased nigral ironcontent and alterations in other metal ions occurring in brain inParkinson’s disease. J. Neurochem. 52: 1830–1836.
6. Dexter, D. T., A. Carayon, F. Javoy-Agid, Y. Agid, F. R. Wells,S. E. Daniel, A. J. Lees, P. Jenner, and C. D. Marsden. 1991.Alterations in the levels of iron, ferritin and other trace inParkinson’s disease and other neurodegenerative diseases affect-ing the basal ganglia. Brain 114: 1953–1975.
7. Dollahite, J. W., W. T. Hardy, and J. B. Menson. 1964. Toxicity ofHelenium microcephalum (Smallhead sneezeweed). J. Am. Vet.Assoc. 145: 694–696.
8. Forno, L. S. 1995. Pathological considerations in the etiology ofParkinson’s disease. In Etiology of Parkinson’s Disease (J. H.Ellenberg, W. C. Koller, and J. W. Langston, Eds.), pp. 65–95.Marcel Dekker, New York.
9. Fowler, M. E. 1965. Nigropallidal encephalomalacia in thehorse. J. Am. Vet. Med. Assoc. 147: 607–616.
10. Han, B., M. Robles, and B. H. Choi. 1998. Repin, a sesquiterpenelactone isolated from Russian knapweed, induces apoptosis inPC12 cells. Society of Toxicology Annual Meeting, Seattle, WA.[Abstract]
11. Jenner, P. 1991. Oxidative stress as a cause of Parkinson’sdisease. Acta Neurol. Scand. 84: 6–15.
12. Jollow, D. J., S. S. Thorgesson, W. Z. Potter, M. Hashimoto, andJ. R. Mitchell 1974. Acetominophen-induced hepatic necrosis.Pharmacology 12: 251–271.
13. Kim, H. K. 1980. Toxicity of sesquiterpene lactones. Res. Com-mun. Chem. Pathol. Pharmacol. 28: 189–192.
14. Langston, J. W., P. Ballard, J. W. Tetrud, and I. Irwin. 1983.Chronic parkinsonism in humans due to a product of meperi-dine analog synthesis. Science 219: 979–980.
15. Langston, J. W., I. Irwin, and G. A. Ricaurte. 1987. Neurotoxins,parkinsonism and Parkinson’s disease. Pharmacol. Ther. 32:19–49.
16. Lowry, O. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the folin phenol reagent. J.Biol. Chem. 193: 265–275.
17. Mettler, F. A., and G. M. Stern. 1963. Observations on the toxiceffects of yellow star thistle. J. Neuropathol. Exp. Neurol. 22:164–169.
18. Ohkawa, H., N. Ohishi, and K. Yagi. 1979. Assays in lipidperoxides in animal tissues by thiobarbituric acid reaction.Analyt. Biochem. 95: 351–358.
19. Paxinos, G., and C. Watson. 1982. The Rat Brain in StereotaxicCoordinates. Academic Press, New York.
20. Perry, T. L., D. V. Godin, and S. Hansen. 1982. Parkinson’sdisease ‘‘a disorder due to nigral glutathione deficiency?’’ Neuro-sci. Lett. 33: 305–310.
21. Picman, A. K., and E. F. Schneider. 1993. Inhibition of fungalgrowth by selected sesquiterpene lactones. Biochem. Syst. Ecol.21: 307–314.
22. Riederer, P., E. Sofic, W. D. Rausch, B. Schmidt, G. P. Reynolds,K. Jellinger, and M. B. Youdim. 1989. Transitional metals,ferritin, glutathione and ascorbic acid in parkinsonian brains. J.Neurochem. 52: 515–520.
23. Robinson, R. W., and R. Providenti. 1993. Breeding lettuce forviral resistance. In Resistance to Viral Disease of Vegetables:Genetics & Breeding. (M. Kyle, and D. Wallace, Eds.), pp. 61–79.Timber Press, Portland, OR.
24. Robles, M. 1995. Toxic Effects of Repin, a Neurotoxic Sesquiter-pene Lactone. Ph.D. Dissertation, University of California,Irvine.
25. Robles, M., M. Aregullin, J. West, and E. Rodriguez. 1995.Recent studies on the zoopharmacognosy, pharmacology andneurotoxicology of sesquiterpene lactones. Planta Med. 61:199–203.
26. Robles, M., N. Wang, R. Kim, and B. H. Choi. 1997. Cytotoxiceffects of repin, a principal sesquiterpene lactone of Russianknapweed. J. Neurosci. Res. 47: 90–97.
27. Rodriguez, E., G. Towers, and J. C. Mitchell. 1976. Biologicalactivities of sesquiterpene lactones (Review) Phytochemistry 15:1573–1580.
28. Schapira, A. H. V., J. M. Cooper, D. Dexter, J. B. Clark, P. Jenner,and C. D. Marsden. 1990. Mitochondrial complex I deficiency inParkinson’s disease. J. Neurochem. 54: 823–827.
29. Shin, J., M. Robles, and B. H. Choi. 1996. Oxidative stress intoxic effects of repin, a sesquiterpene lactone isolated fromRussian knapweed. Society of Toxicology Annual Meeting, Ana-heim, CA. [Abstract]
30. Stevens, K. L., R. J. Riopelle, and R. Y. Young. 1990. Repin, asesquiterpene lactone from acroptilon repens posessing excep-tional biological activity. J. Nat. Prod. 53: 218–221.
31. Tamaki, H., R. W. Robinson, J. L. Anderson, and G. S. Stoews-and. 1995. Sesquiterpene lactones in virus-resistant lettuce. J.Agric. Food Chem. 43: 6–8.
32. Tanner, C. M., and J. W. Langston. 1990. Do environmentaltoxins cause Parkinson’s disease? A critical review. Neurology40: 17–30.
33. Tietze, F. 1969. Enzymatic method for quantitative determina-tion of nanogram amounts of total and oxidized glutathione:applications to mammalian blood and other tissues. Analyt.Biochem. 27: 502–522.
34. Young, S., W. W. Brown, and B. Klinger. 1970. Nigropallidalencephalomalacia in horses fed Russian knapweed (Centaurearepens L.). Am. J. Vet. Res. 31: 1393–1404.
35. Wang, Y., M. Hamburger, C. H. K. Cheng, B. Costall, R. J.Naylor, P. Jenner, and K. Hostettmann. 1991. Neurotoxic sesqui-terpenoids from the yellow star thistle Centaurea solstitialis L.(Asteraceae). Helv. Chim. Acta 74: 117–123.
136 ROBLES ET AL.





























