Embed Size (px)
Citation preview

Department of Applied Chemistry and Microbiology, Division of Nutritionand
Department of Biological and Environmental Sciences, Division of GeneticsUniversity of Helsinki
Minna Pekkinen
REGULATION OF OSTEOBLAST DIFFERENTIATION AND GENE EXPRESSION BYPHOSPHODIESTERASES AND BIOACTIVE PEPTIDES
ACADEMIC DISSERTATION
To be presented, with the permission of the Faculty of Biosciences of the University ofHelsinki, for public criticism in the large lecture hall, Bulevardi 18, Helsinki, on 28 March2008, at 12 noon.
HELSINKI 2008

2
Supervisors
Acting Professor Christel LambergAllardtDepartment of Applied Chemistry and MicrobiologyUniversity of HelsinkiFinland
Dr. Mikael AhlströmDepartment of Applied Chemistry and MicrobiologyUniversity of HelsinkiFinland
Reviewers
Acting Professor Petri LehenkariDepartment of AnatomyUniversity of OuluFinland
Adjunct Professor Anitta MahonenInstitute of BiomedicineDeparment of Medical BiochemistryUniversity of KuopioFinland
Opponent
Professor Kalervo VäänänenInstitute of BiomedicineDepartment of AnatomyUniversity of TurkuFinland
© Minna Pekkinen 2008 Yliopistopaino
ISNB 9789529235285 (paperback)ISNB 9789521045929 (PDF, http//:ethesis.helsinki.fi)

3
Contents
Abstract… … … … … … … … … … … … … … … … … … … … … … … … … … … … … … 6List of original publications… … … … … … … … … … … … … … … … … … … … … … .8
Abbreviations… … … … … … … … … … … … … … … … … … … … … … … … … … … ...9
1. Introduction… … … … … … … … … … … … … … … … … … … … … … … … … .112. Review of the literature...................................................................................13
2.1 Bone biology… … … … … … … … … … … … … … … … … … … … … … … ..132.1.1 Bone tissue and bone cells.............................................................132.1.2 Osteoblastic differentiation............................................................132.1.3 Osteoblastic regulation of osteoclast differentiation......................172.1.4 Bone remodelling and its regulation...............................................182.1.5 Unbalanced bone remodelling: osteoporosis … … … … … … .........192.1.6 Glucocorticoids and bone.............................................................. .20
2.2 Cyclic nucleotide signalling and phosphodiesterases (PDEs).....................212.2.1 Cyclic adenosine monophosphate (cAMP) signalling.....................212.2.2 Cyclic guanosine monophosphate (cGMP) signalling....................222.2.3 cAMP and cGMP PDEs...................................................................23
2.2.3.1 PDE1 family............................................................................272.2.3.2 PDE2 family...........................................................................292.2.3.3 PDE3 family...........................................................................292.2.3.4 PDE4 family...........................................................................302.2.3.5 PDE5 family...........................................................................312.2.3.6 PDE6 family...........................................................................312.2.3.7 PDE7 family...........................................................................312.2.3.8 PDE8 family...........................................................................322.2.3.9 PDE9 family...........................................................................322.2.1.10 PDE10 family.........................................................................332.2.1.11 PDE11 family.........................................................................33
2.2.4 Regulation of activity and compartmentalization of PDEs.............332.2.5 Inhibitors of PDEs...........................................................................342.2.6 Bone metabolism and PDE inhibitors… … … ...… … … … ..............34
2.3 Other signal transduction pathways associated with PDEs...................... ..352.3.1 Exchange protein directly activated by cAMP (EPAC)
and PDEs........................................................................352.3.2 Mitogenactivated protein kinase (MAPK) and PDEs....................362.3.3 PKC and PDEs................................................................................36

4
2.4 Bioactive peptides........................................................................................362.4.1 Bioactive peptides in bone...............................................................38
3. Aims of the study… … … … … … … … … … … … … … … … … … … … … … ..… .394. Materials and methods… … … … … … … … … … … … … … … … … … … … .......40
4.1 Cell culture...................................................................................................404.1.1 Osteosarcoma cell lines.................................................................404.1.2 Human osteoblasts........................................................................404.1.3 Human mesenchymal stem cell (hMSC)derived osteoblasts.......40
4.2 PDE inhibitors..............................................................................................414.3 Primers..........................................................................................................414.4 cAMP analyses.............................................................................................434.5 Qsepharose chromatography.......................................................................434.6 Enzyme activity analyses.............................................................................44
4.6.1 Assay of PDE activity...................................................................444.6.2 Assay of ALP activity...................................................................44
4.7 PDE silencing...............................................................................................444.8 Isoleucineprolineproline (IPP), valineprolineproline (VPP) and leucine lysineproline (LKP) tripeptides...................................................................454.9 Cell proliferation analyses............................................................................454.10 Mineralization analyses..............................................................................454.11 mRNA expression analyses........................................................................46
4.11.1 RNA isolation..............................................................................464.11.2 cDNA microarrays.......................................................................464.11.3 cDNA synthesis...........................................................................464.11.4 Semiquantitative reverse transcriptase polymerase chain
reaction (RTPCR).......................................................................474.11.5 Quantitative Realtime PCR (qRTPCR).....................................47
5. Results and discussion… ..… … … … … … … … … … … … … … … … … … … … ..485.1 Identification of PDE families in human osteoblastic cells.............485.2 Identification of PDE subtypes in human osteoblastic cells............515.3 PDE7 and PDE8A silencing by siRNAs..........................................525.4 PDE7 silencing has potential for enhancing osteogenic gene expression and differentiation in hMSCderived osteoblasts...........535.5 PDE7 inhibition enhances cAMP signalling, differentiation and mineralization in hMSCderived osteoblasts....................................565.6 Downregulation of cAMPPDE by dexamethasone in humanosteosarcoma cells...................................................................................585.7 Influence of dexamethasone on PDE subtype mRNA expression in MG63 and SaOS2 cells............................................585.8 Effects of PDEs in osteoblasts..........................................................59

5
5.9 Influences of bioactive peptides IPP, VPP and LKP on gene expression and proliferation in hMSCderived osteoblasts.............60
6. Conclusions… … … … … … … … … … … … … … … … … … … … … … .................637. Acknowledgements… … … … … … … … … … … … … … … … … … … … … … … .658. References… … … … … … … … … … … … … … … … … … … … … … … … … … 66

6
Abstract
Bone is a mineralized tissue that enables multiple mechanical and metabolic functions to becarried out in the skeleton. Bone contains distinct cell types: osteoblasts (boneforming cells),osteocytes (mature osteoblast that embedded in mineralized bone matrix) and the osteoclasts(boneresorbing cells). Remodelling of bone begins early in foetal life, and once the skeletonis fully formed in young adults, almost all of the metabolic activity is in this form. Bone isconstantly destroyed or resorbed by osteoclasts and then replaced by osteoblasts. Many bonediseases, i.e. osteoporosis, also known as bone loss, typically reflect an imbalance in skeletalturnover.
The cyclic adenosine monophosphate (cAMP) and the cyclic guanosine monophosphate(cGMP) are second messengers involved in a variety of cellular responses to suchextracellular agents as hormones and neurotransmitters. In the hormonal regulation of bonemetabolism, i.e. via parathyroid hormone (PTH), parathyroid hormonerelated peptide(PTHrp) and prostaglandin E2 signal via cAMP. cAMP and cGMP are formed by adenylateand guanylate cyclases and are degraded by phosphodiesterases (PDEs). PDEs determine theamplitudes of cyclic nucleotidemediated hormonal responses and modulate the duration ofthe signal. The activities of the PDEs are regulated by multiple inputs from other signallingsystems and are crucial points of crosstalk between the pathways.
Foodderived bioactive peptides are reported to express a variety of functions in vivo. Theangiotensinconverting enzymes (ACEs) are involved in the regulation of the specificmaturation or degradation of a number of mammalian bioactive peptides. The bioactivepeptides offer also a nutriceutical and a nutrigenomic aspect to bone cell biology.
The aim of this study was to investigate the influence of PDEs and bioactive peptides on theactivation and the differentiation of human osteoblast cells. The profile of PDEs in humanosteoblastlike cells and the effect of glucocorticoids on the function of cAMP PDEs, wereinvestigated at the mRNA and enzyme levels. The effects of PDEs on bone formation andosteoblast gene expression were determined with chemical inhibitors and siRNAs (shortinterfering RNAs). The influence of bioactive peptides on osteoblast gene expression andproliferation was studied at the mRNA and cellular levels.
This work provides information on how PDEs are involved in the function and thedifferentiation of osteoblasts. The findings illustrate that genespecific silencing with anRNA interference (RNAi) method is useful in inhibiting, the gene expression of specificPDEs and further, PDE7 inhibition upregulates several osteogenic genes and increases bALPactivity and mineralization in human mesenchymal stem cellsderived osteoblasts. PDEsappear to be involved in a mechanism by which glucocorticoids affect cAMP signaling. Thismay provide a potential route in the formation of glucocorticoidinduced bone loss, involvingthe downregulation of cAMPPDE. PDEs may play an important role in the regulation of

7
osteoblastic differentiation. Isoleucineprolineproline (IPP), a bioactive peptide, possessesthe potential to increase osteoblast proliferation, differentiation and signalling.

8
LIST OF ORIGINAL PUBLICATIONS
This thesis is based on the following original publications, which are referred to in the text bytheir Roman numerals (IIV):
I Ahlström, M., Pekkinen M., Huttunen, M., LambergAllardt C. (2005): Cyclicnucleotide phosphodiesterases (PDEs) in human osteoblastic cells; effect ofPDE inhibition on cAMP accumulation. Cellular & Molecular Biology Letters10(2): 305319.
II Pekkinen, M., Ahlström, M.E.B, Riehle, U., Huttunen, M.M, LambergAllardtC.J.E. (2007): Effects of phosphodiesterase 7 Inhibition by RNA interference onthe gene expression and differentiation of human mesenchymal stem cellderived osteoblasts. Bone (accepted).
III Ahlström, M.*, Pekkinen, M.*, Huttunen, M., LambergAllardt, C. (2005):Dexamethasone downregulates cAMPphosphodiesterase in humanosteosarcoma cells. Biochemical Pharmacology 69: 267275.
IV Huttunen, M.M.*, Pekkinen, M.*, Ahlström, M.E.B, LambergAllardt C. J.E.(2007): Effects of bioactive peptides isoleucineprolineproline (IPP), valineprolineproline (VPP) and leucinelysineproline (LKP) on gene expression ofosteoblasts, differentiated from human mesenchymal stem cells. British Journalof Nutrition 98: 780788.
*These authors contributed equally.
These publications have been reproduced with the kind permission of theircopyright holders. In addition, some unpublished material is presented.

9
ABBREVIATIONSaa amino acidAC adenylyl cyclaseACEs angiotensincoverting enzymesAKAP Akinase anchoring proteinATF1 activating transcription factor 1ATP adenosine triphosphatebALP bonespecific alkaline phosphataseBMPs bone morphogenetic proteinsBRL 50481 3(N,Ndimethylsulfonamido)4methylnitrobenzeneBSA bovine serum albuminBSP bone sialoproteinCaM calmodulincAMP cyclic adenosine monophosphateCdc42 cell division cycle 42cGMP cyclic guanosine monophoshateCRE cAMP response elementCREB cAMP response elementbinding proteinCREM cAMP responseelement modulator proteinCol1 collagen type 1Dex dexamethasonedsRNA doublestranded RNAECM extracellular matrixEHNA erythro9[2hydroxy3nonyl] adenineEPAC exchange protein directly activated by cyclic AMPERK extracellular signalregulated kinaseFCS foetal calf serumFGF fibroblast growth factorFSP1 fibroblastspecific protein 1GAPDH glyceraldehyde3phosphate dehydrogenaseGCs glucocorticoidshMSC human mesenchymal stem cellIBMX 3isobutyl1methylxanthineIGFI insulinlike growth factor 1Ihh Indian hedgehogIPP isoleucineprolineprolineKm Michaelis constantLKP leucinelysineprolineMAPK mitogenactivated protein kinaseMC mesangial cellmRNA messenger RNAMSCF cytokine macrophage colonystimulating factor

10
8MMX 8methoxymethyl3isobutyl1methylxanthineMSC bone marrow stroma mesenchymal stem cellMSCF macrophage colonystimulating factorMyoD transcription factor for myogenesisNO nitric oxideOCN osteocalcinOPG osteoprotegerinOPN osteopontinOsx osterixPAS perarntsimPCA principal component analysisPDE phosphodiesterasePGE2 prostaglandin E2 prostaglandinsPGs prostaglandinsPKA protein kinase APKB protein kinase BPKC protein kinase CPKG protein kinase GPKI protein kinase inhibitorPLC phospholipase CPPAR 2 peroxisome proliferatoractivated receptorgamma2PTH parathyroid hormonePTHR PTH/PTHrp receptorPTHrp parathyroid hormonerelated peptideRANK receptor activator for nuclear factor BRANKL receptor activator for nuclear factor B ligandRap1 repressor activator protein 1REC response regulator receiverRGD arginineglycineaspartic acidRho ras homolog geneRNAi RNA interferenceRunx2 runt related transcription factor 2siRNAs short interfering RNAsSox9 SRYbox containing gene 9TGF transforming growth factor TRANCE receptor activator of NF{kappa} B ligandVmax maximal velocity constantVDR 1,25dihydroxyvitamin D3 receptorVEGF vascular endothelial growth factorVPP valineprolineprolineWnt a combination of the genes (Wg) wingless and Int

11
1. Introduction
Bone has two main functions: it forms a rigid skeleton and plays a central role in calcium andphosphate homeostasis. Bone modelling is a process associated with growth and reshaping ofbones in childhood and adolescence. Throughout human life, bone tissue is subject to acontinuous turnover process whereby old bone is removed and replaced by new bone by thecoupled processes of bone resorption and bone formation. Bone is a very active organ and itsfixity after death is in sharp contrast to its ceaseless activity during life. In the adult skeleton,the continuous activity largely comprises to the process of bone remodelling. Bone isconstantly destroyed or resorbed by the osteoclasts (boneresorbing cells) and then replacedby the osteoblasts (boneforming cells). This process is regulated by other bone cells, namelyosteocytes, hormones, cytokines, prostaglandins and growth factors (Raisz 1999, Ducy et al.2000, Teitelbaum 2000). A misbalance between bone resorption and formation may lead tomany bone diseases, e.g., osteoporosis and osteopetrosis. Osteoporosis, also known as boneloss, typically reflects an imbalance in skeletal turnover such that bone resorption exceedsbone formation. In osteopetrosis, the misbalance is caused by a defect in osteoclastogenesis orby dysfunction of osteoclasts.
The cyclic nucleotides cAMP and cGMP are second messengers involved in a variety ofcellular responses to extracellular agents such as hormones and neurotransmitters. Forexample, parathyroid hormone (PTH), an important hormone acting together with 1,25dihydroxyvitamin D and calcitonin regulates bone metabolism, signalling via cAMP. cAMPand cGMP are formed by adenylate and guanylate cyclases and are degraded byphosphodiesterases (PDEs). PDEs determine the amplitudes of cyclic nucleotidemediatedhormonal responses and modulate the duration of the signal. The activities of PDEs areregulated by multiple inputs from other signalling systems and are crucial points of crosstalkbetween the pathways (Soderling and Beavo 2000, Lugnier 2006). PDEs are an important partof signal transduction in the cell, but very little is known about their role in bone cells.
During the last decade, fundamental studies have opened a new field of research dealing withbioactive or biogenic substances derived from foods (Meisel 2001). The angiotensinconverting enzymes (ACEs) are involved in the regulation of the specific maturation ordegradation of a number of mammalian bioactive peptides. The antihypertensity effects ofbioactive peptides on blood pressure have been shown in several studies (e.g. Mitsui et al.2004), and the bioactive peptides offered also provide a nutrigenomic aspect to bone cellbiology.
This work was conducted to clarify the involvement of PDEs and bioactive peptides in thefunction and differentiation of human osteoblast cells and to provide more insight into the roleof PDEs in the regulation of bone metabolism. Elucidation of the role of PDEs in bone cells is

12
important because bone metabolism, especially bone formation, could be affected via PDEs.The potential of bioactive peptides to increase osteogenic effects was also investigated.

13
2. Review of the literature
2.1 Bone biology2.1.1 Bone tissue and bone cells
Bone tissue is highly organized and consists of a mineral phase of hydroxyapatite andamorphous calcium phosphate crystals deposited in an organic matrix. It contains the distinctcell types of osteoblasts, osteocytes, osteoclasts and lining cells. The osteoblast is the cellresponsible for collagenand other bone proteins, forming the bone organic matrix, whichconsists of approximately 90% type I collagen and 10% different noncollagenous proteins,such as osteocalcin, matrix Gla protein, bone sialoprotein, osteopontin and fibronectin (Aubinand Liu 1996). Osteoblasts also have an important role in the subsequent mineralization of thematrix. Once bone is mineralized, some of the osteoblasts are retained on the bone surface ininactive form and named as the bonelining cells (Parfit 1994). Osteocytes are osteoblasts thathave been trapped within the bone matrix during the process of bone formation. Osteocytescommunicate with one another and with cells at the bone surface via a meshwork of cellprocesses that run through canaliculi in the bone matrix (Kogiannis and Noble, 2007). Thus,bone cells form a functional network within which cells at all stages of bone formation, fromthe preosteoblast to the mature osteocyte, remain connected. Osteoclasts are multinucleatedcells formed by the fusion of the monocyte/macrophage family. They are responsible for boneresorption, playing a central role in formation of the skeleton and regulation of its mass. Theyattach on the bone surface, secreting acids and lysosomal enzymes into the space providedbetween their apical surfaces, and the mineralized bone surface (Ducy et al. 2000, Teitelbaum2000).
2.1.2 Osteoblastic differentiation
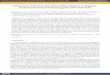
Bone marrow stromal mesenchymal stem cells (MSCs) have strong proliferative potential andthe capacity to form bone, cartilage, adipocytes and fibrous tissue. MSCs have been found ina variety of other adult tissues and can differentiate into myoblasts, haematocytes andpossibly even neural tissues (Reyes et al. 2001, Jiang et al. 2002a, 2002b, 2003). And, inexample, osteogenic differentiation potential of human bone marrow MSCs does not decreaseby age at late adulthood (Leskelä et al. 2003). Commitment of MSCs to tissuespecific celltypes is orchestrated by morphogens, developmental signalling pathways and transcriptionalregulators that serve as key factors (Figure 1A, Table 1).
The process of osteoblast differentiation can be subdivided in to three stages: 1) proliferation,2) extracellular matrix synthesis and maturation and 3) mineralization. Each stage ischaracterized by expression of distinguishing osteoblast markers. The most frequently usedmarkers of osteoblast differentiation are alkaline phosphatase (ALP), collagen type 1 (Col1),osteopontin (OPN), bone sialoprotein (BSP), osteocalcin (OCN) and PTH/PTHrp receptor
A B
Figure 1. (A) Commitment of mesenchymal stem cells to tissuespecific cell types. Some of the known signalling proteins, growth factorsand transcription factors playing key regulator roles in the mesenchymal lineages are indicated. Also shown is noticeable plasticity betweenosteoblasts and adipocytes. (B) Osteoblastogenesis. Selected markers of the stages of maturation are used as examples. Modified fromLian et al. (2003).
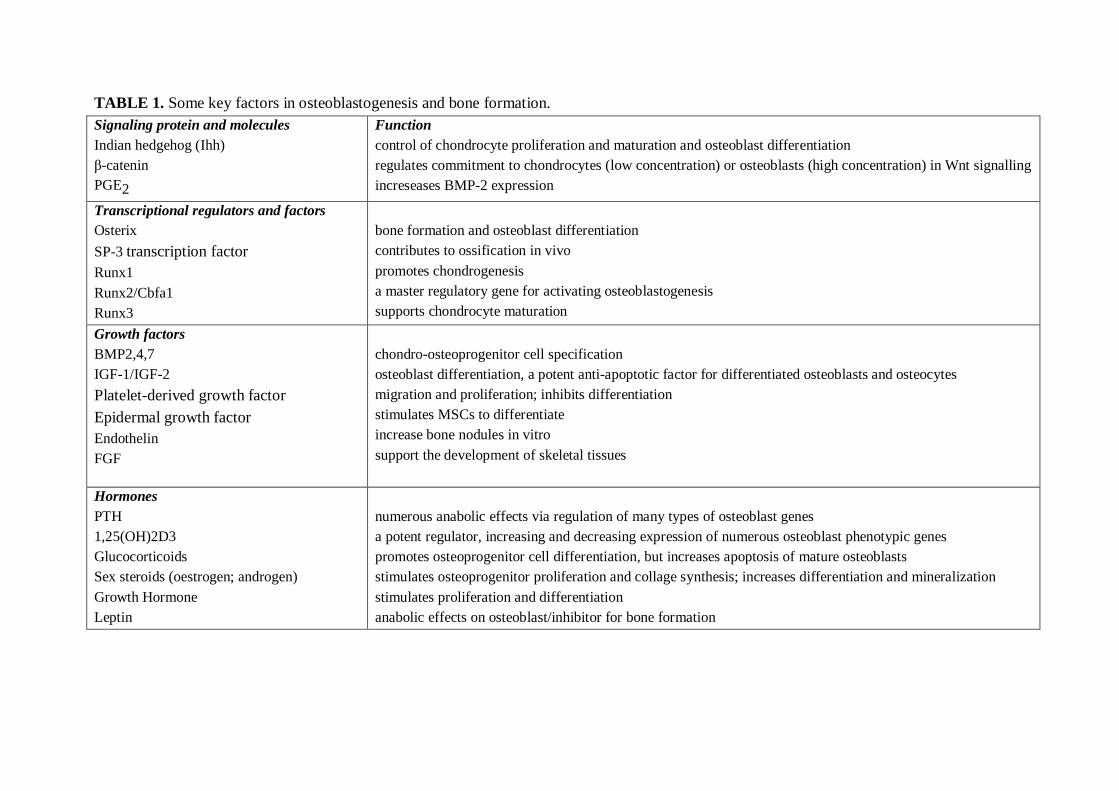
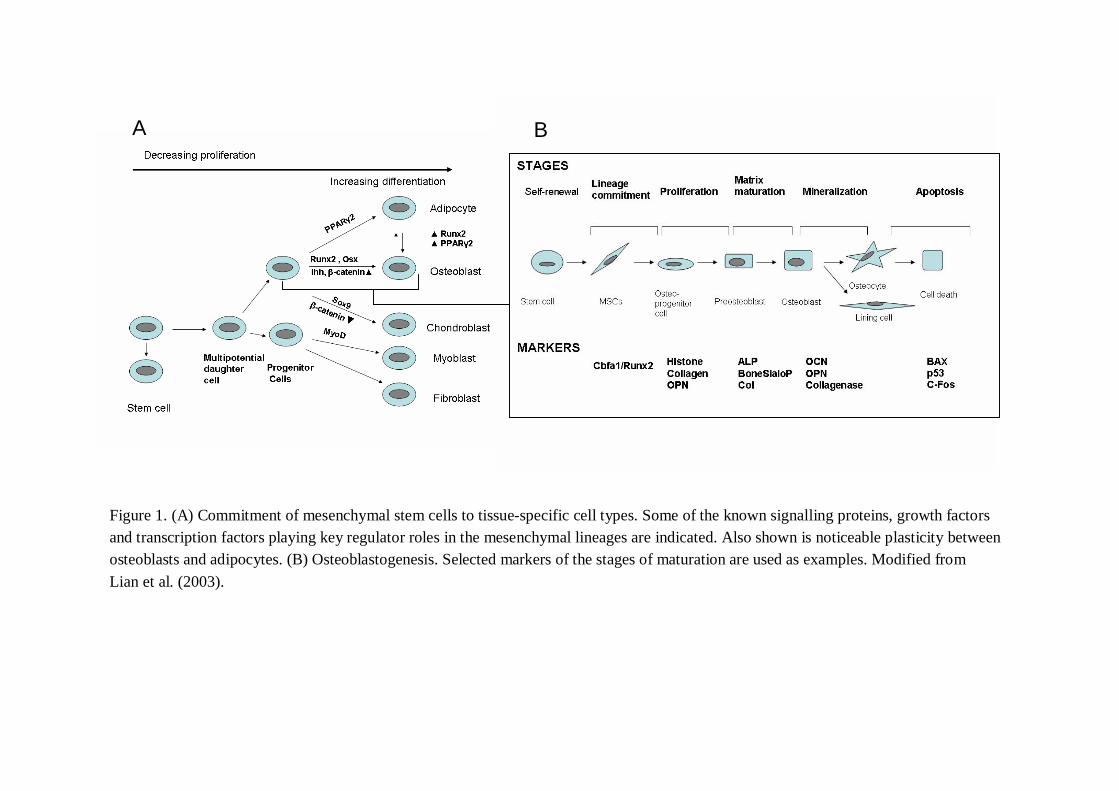
TABLE 1. Some key factors in osteoblastogenesis and bone formation.Signaling protein and moleculesIndian hedgehog (Ihh)catenin
PGE2
Functioncontrol of chondrocyte proliferation and maturation and osteoblast differentiationregulates commitment to chondrocytes (low concentration) or osteoblasts (high concentration) in Wnt signallingincreseases BMP2 expression
Transcriptional regulators and factorsOsterixSP3 transcription factorRunx1Runx2/Cbfa1Runx3
bone formation and osteoblast differentiationcontributes to ossification in vivopromotes chondrogenesisa master regulatory gene for activating osteoblastogenesissupports chondrocyte maturation
Growth factorsBMP2,4,7IGF1/IGF2Plateletderived growth factorEpidermal growth factorEndothelinFGF
chondroosteoprogenitor cell specificationosteoblast differentiation, a potent antiapoptotic factor for differentiated osteoblasts and osteocytesmigration and proliferation; inhibits differentiationstimulates MSCs to differentiateincrease bone nodules in vitrosupport the development of skeletal tissues
HormonesPTH1,25(OH)2D3GlucocorticoidsSex steroids (oestrogen; androgen)Growth HormoneLeptin
numerous anabolic effects via regulation of many types of osteoblast genesa potent regulator, increasing and decreasing expression of numerous osteoblast phenotypic genespromotes osteoprogenitor cell differentiation, but increases apoptosis of mature osteoblastsstimulates osteoprogenitor proliferation and collage synthesis; increases differentiation and mineralizationstimulates proliferation and differentiationanabolic effects on osteoblast/inhibitor for bone formation

16
(PTHR). In general, ALP, BSP and Col1 are early markers for osteoblast differentiation,while PTHR and OCN appears late, parallel with mineralization. Osteoprogenitor cells willarise under the appropriate stimulus from MSCs, differentiated preosteoblasts, osteoblasts andosteocytes (Figure 1B).
The canonical Wnt/ catenin pathway provides early development cues that indirectlymediate the initial cascade of gene expression for skeletal development, bone formation andosteoblast differentiation (Church et al. 2002, Logan et al. 2004). Bone morphogeneticproteins [BMPs, members of the transforming growth factor (TGF ) family] playimportant roles in directing fate decisions for MCSs. TGF can provide competence for earlystages of chondroblastic and osteoblastic differentiation, but it inhibits myogenesis,adipogenesis and latestage osteoblast differentiation.
BMPs also inhibit adipogenesis and myogenesis, but they strongly promote osteoblastdifferentiation (Boden et al. 1997, Hughes et al. 1995, Abe et al. 2000). The necessity of therunt homology domain factor Runx2 (Cbfa1/AML3/PEBP 1) and the zing finger proteinosterix in bone formation has been ascertained by genetic studies (Banerjee et al. 1997, Ducyet al. 1997, Choi et al. 2000, Nakashima et al. 2002). Because Runx2 expression arises earlierthan osterix, these two factors may represent a temporal sequence of regulation crucial for theearly and final stages of osteoblast differentiation (Nakashima et al. 2002, Komori 2005,2006). The influence of transcription factors on lineage determination and differentiation isrevealed by their capability to cause transdifferentiation of cells to other phenotypes whenthese factors are overexpressed or suppressed. For example, forced overexpression ofperoxisome proliferationactivated receptor 2 (PPAR 2) in osteoblasts will modify theirphenotype to adipocytes, whereas forced expression of Runx2 in adipocytes or other nonosseous cells results in acquisition of a differentiated osteoblast phenotype (Nuttall et al.1998, LeckaCzernik et al. 1999, Jeon et al. 2003)(Figure 1).
The developmental expression patterns of transcription factors during osteoblast maturationreflect their roles as key determinants of osteoblast differentiation (Figure 1B). Many steroidand polypeptide hormones, growth factors and cytokines regulate not only the growth ofosteoprogenitors and/or their progression to mature osteoblasts but also osteoblast activity andapoptosis (for review, see Xing and Boyce 2005). For example, PTH stimulates growth ofosteoprogenitor populations, while also inhibiting osteoblast and osteocyte apoptosis (Boyceet al. 2002). Leptin is secreted largely by adipose tissue, but also by human mesenchymalstem cells (hMSCs) and osteoblasts at the mineralization stage. It enhances proliferation,collagen synthesis, matrix deposition and mineralization (Gordeladze et al. 2002) and servesas an antiapoptotic agent in several cell systems, including bone marrow stem cells andosteoblasts (Shimabukuro et al. 1998, Takeda et al. 1998, Konopleva et al. 1999).
Among important concepts in regulation of osteoblast differentiation and activity is the ideathat local or endogenous production of particular growth factors and cytokines undergoes

17
changes required for osteoprogenitor differentiation to proceed and for osteoblasts to modifytheir activity. For example, insulinlike growth factor 1 (IGFI) expression is markedlyincreased during early osteoblast recruitment, but declines dramatically as these cells undergodifferentiation. Data from both IGFI or IGFI receptordeficient mice and mice with targetedoverexpression of IGFI in osteoblasts show that IGFI is involved not only in embryonicbone development but also postnatal bone formation and remodelling (Zhao et al. 2000,Zhang et al. 2002). Fibroblast growth factor 8 (FGF8) has been demonstrated to regulatesdifferent stages of MSC differentiation toward osteogenic lineage in vitro and to increase theosteogenic capacity of bone marrow cells at the early stage of their differentiation. However,continued exposure of osteoblastic cultures to FGF8 leads to the inhibition of bone formation(Valta et al. 2006).
2.1.3 Osteoblastic regulation of osteoclast differentiation
During skeletal development and throughout life, the cells from the osteoblast lineagesynthesize and secrete molecules that in turn initiate and control osteoclast differentiation(Raisz 1999). Osteoclast differentiation calls for the binding of macrophage colonystimulating factor (MSCF) to its receptor as well as the binding of the soluble differentiationfactor receptor activator of NFB ligand (RANKL) to its receptor (RANK) on osteoclastprecursor cells (Teitelbaum and Ross 2003). One of the first factors cloned that regulatesosteoclast differentiation was osteoprotegerin. Osteoprotegerin, originally characterized as anovel secreted member of the tumor necrosis factor receptor superfamily, was later found toinhibit spontaneous or induced bone resorption and cause osteopetrosis, the opposite ofosteoporosis. Osteoprotegerin acts as a decoy receptor that binds to RANKL and prevents itfrom interacting with its receptor (Figure 2). RANKL and osteoprotegerin, both of which areproduced by osteoblasts at different stages of maturity (Gori et al. 2000), account for some ofthe signals in osteoblast–osteoclast communication. In conjunction with MSCF, the RANK–RANKL–osteoprotegerin system regulates osteoclast differentiation. Thus, mice and humansdeficient in osteoprotegerin have a high rate of bone loss (bone resorption that exceeds boneformation; Whyte et al. 2002). As in other high bone turnover states, antiresorptive agentscan still reduce both bone formation and resorption and compensate for osteoprotegerindeficiency. The signal(s) that couples resorption and formation remains elusive, althoughseveral of the anabolic ligands, such as BMPs and TGFß, are stored in bone matrix as bone isformed and are released during bone resorption and can thus act on osteoblasts and precursorsin the vicinity. BMPs could also enhance osteoclast differentiation from progenitors(Hentunen et al. 1998, Itoh et al. 2001). Signals from osteoblasts to osteoclasts can beprovided by RANKL and osteoprotegerin. In this case, preosteoblasts express a high level ofRANKL relative to osteoprotegerin, which stimulates osteoclast differentiation and function.More mature osteoblasts, by contrast, express high levels of osteoprotegerin relative toRANKL, which inhibits osteoclast differentiation and function (Gori et al. 2000).

18
Figure 2. Interactions among osteoblasts and osteoclasts in bone resorption and formation andsome factors that modulate their differentiation and function.
2.1.4 Bone remodelling and its regulation
Remodelling of bone begins early in foetal life, and once the skeleton is fully formed inyoung adults, almost all of the metabolic activity is in this form. The process involving boneresorption and formation is initiated by mechanical signals and is controlled by local andsystemic factors that regulate osteoblast and osteoclast differentiation and function. Boneremodelling is not a random process and takes place in bone remodelling units comprisingosteoblasts, osteoclasts and their precursors, in which resorption and formation are coupled.Osteocytes are also involved in the process by soluble factors or directly through cell to cellcontacts (Zhao et al. 2002, Heino et al. 2002, 2004). Bone resorption is likely the initial eventthat occurs in response to local mechanical stress signals. The bone remodelling cycleinvolves a complex series of sequential steps that are highly regulated (Raisz 1999). The mainphases are 1) osteoclastic resorption, 2) reversal phase, 3) preosteoblastic migration anddifferentiation into osteoblasts 4) osteoblastic matrix formation and 5) mineralization. Thecomplete bone remodelling cycle at each microscopic site takes about 46 months, with the

19
resorption process lasting about two weeks and bone formation several months (Hadjidakisand Androulakis 2006).
The metabolic functions of the skeleton are served in large part by two calciumregulatinghormones, PTH and 1,25dihydroxy vitamin D. A third hormone, calcitonin, which caninhibit bone resorption, is a potent inhibitor of bone resorption and is used clinically in thetreatment of osteoporosis. The US Food and Drug Administration has approved teriparatide,the Nterminal (134) fragment of recombinant human PTH, for the treatment of osteoporosis(Mishaela et al. 2002, Hodsman et al. 2005). In many countries in Europe, teriparatide is usedafter an unsuccessful course of bisphosphonate therapy or after a patient has had a previousosteoporotic fracture (Canalis 2007). PTH regulates serum ionized calcium concentration andits primary target cells in bone tissue are the osteoblasts (Raisz 1999). The increase in boneformation after PTH administration in vivo has been attributed to increased production ofosteoprogenitors and differentiation of osteoblasts and decreased apoptosis of existingosteoblasts (Jilka et al. 1999). A number of studies in animals and humans have demonstratedthat shortterm, intermittent administration of low doses of PTH has a significant anaboliceffect on bone (for review, see Jilka 2007). PTH affects several typical bone formationassociated features in the osteoblast such as collagen synthesis (Dempster et al. 1993), ALPactivity (Majeska et al. 1982) and synthesis of OPN (Noda and Rodan 1989). The receptor forPTH belongs to the superfamily of the seven membranes spanning G proteincoupledreceptors (Birnbaumer 1990). The effects of PTH are known to be mediated by cAMP andcalcium and by the activation of protein kinase C (PKC) (Sprague et al. 1996, Ahlström andLambergAllardt 1997). Plasma PTH concentration tends to increase with age, and this mayproduce an increase in bone turnover and a loss of bone mass, particularly of cortical bone.
Other systemic hormones are important in regulating skeletal growth. Growth hormone canstimulate bone formation and resorption (Rosen et al. 1998), glucocorticoids are necessary forbone cell differentiation during development (Advani et al. 1997) and thyroid hormones canalso stimulate bone resorption and formation and are critical for maintenance of normal boneremodelling (Kawaguchi et al. 1994). Probably the most important systemic hormone inmaintaining normal bone turnover is oestrogen. Oestrogen defiency leads to an increase inbone remodelling in which resorption outstrips formation and bone mass decreases.Testosterone increases osteoblast formation and affects bone formation rather than boneresorption (Khosla et al. 2002). Many local factors, cytokines, prostaglandins (PGs) andgrowth factors also regulate bone remodelling.
2.1.5 Unbalanced bone remodelling: osteoporosis
The reduced bone mass, for instance, in osteoporosis results from an imbalance between boneresorption and formation, with the rate of resorption exceeding that of formation. Excessivebone resorption causes changes in the microstructure of the bone matrix, which make bones

20
prone to fracture. Osteoporosis has been proposed to result from a failure of resident bonecells to respond appropriately to mechanical load–bearing signals and to control bone massand architecture (Lanyon and Skerry 2001). In the majority of conditions that lead toosteoporosis, such as ageassociated gonadal hormone deficiency, remodelling rates are high.Although bone formation and resorption are both increased, the rate of bone formation isinsufficient to keep up with resorption. RANKL has been identified as a crucial cytokine forthe formation and activation of osteoclasts. The effects of RANKL are physiologicallycompensated by the decoy receptor osteoprotegerin (OPG). The RANKL/OPG system isactivated in favour of RANKL in oestrogen deficiency, inflammation, bone malignancies andduring treatment with glucocorticoids, which enhances the ratio of RANKL to OPG, thuspromoting osteoclastogenesis, accelerating bone resorption and inducing bone loss (Simonetat al. 1997, Hsu et al. 1999). The balance between proliferation, differentiation and apoptosisof bone cells decides the size of osteoclast or osteoblast populations. Bone cells continuouslyreceive signals that control their proliferation, activity and survival from neighbouring cells,the bone matrix and hormones. Life history before and after menopause and therapeuticinterventions with drugs, such as sex hormones, glucocorticoids, PTH and bisphosphonates,are therefore partly determined by the survival of osteoclasts, osteoblasts and osteocytes,which contribute to the bone mass and its microarchitecture (Weinstein et al. 1998, Jilka et al.1999, Borton et al. 2001, Calvi et al. 2001, Gordeladze et al. 2002, Iwata et al. 2006).
2.1.6 Glucocorticoids and bone
Glucocorticoids (GCs) play an important role in modifying the metabolic activity andproliferation of bone cells. GCs are required for in vitro bone nodule formation andmineralization, as well as for bone marrow MSC proliferation and osteogenic differentiation(Bellows et al. 1987, Shalhoub et al. 1992, Jaiswal et al. 1997, Bellows et al. 1998). Used inlongterm therapy as immunosuppressive and antiinflammatory drugs, glucocorticoids leadto a reduction in bone formation, enabling the development of osteoporosis (Eastell et al.1998, Patschan et al. 2001). GSs have two major inhibitory effect on bone formation: inhibitcell proliferation and decrease the population of cells capable of synthesizing type I collagen,as well as downregulate collagen gene expression in osteoblasts (Mahonen et al. 1998). GCsdecrease osteoblast and osteocyte activities and survival and have the opposite effects onosteoclasts, i.e. increased maturation and activity as well as decreased apoptosis (Weinstein etal. 1998, O`Brien 2004, Canalis 2005). GCs exposure enhances RANKL expression andinhibits OPG production by osteoblasts (Hofbauer et al. 1999), and also suppresses OPGserum levels in vivo, thus elevating the RANKL/OPG ratio (Sasaki et al. 2001). In addition,GCs enhance the expression and activity of PPAR 2, a transcription factor that suppressesosteoblastogenesis and supports adipogenesis (Shi et al. 2000). Most studies have not foundan association between GCs and increased PTH levels (Canalis et al. 2004), but GCs mightenhance the sensitivity to PTH by changing the number of PTH receptors and their affinity toPTH (Urena et al. 1994).

21
2.2 Cyclic nucleotide signalling and phosphodiesterases (PDEs)
2.2.1 Cyclic adenosine monophosphate (cAMP) signalling
The cAMP is a second messengers involved in a variety of cellular responses to suchextracellular agents such as hormones, growth factors and neurotransmitters. cAMP is a partof the intracellular signalling pathway of several hormones involved in the regulation of bonemetabolism such as PTH, parathyroid hormonerelated peptide (PTHrp) and prostaglandin E2(Beavo 1995, Soderling and Beavo 2000). The level of intracellular cAMP is regulated by thebalance between the activity of two types of enzymes: adenylyl cyclase (AC) and the cyclicnucleotide PDE. Both enzymes are encoded by a large number of genes that differ in theirexpression patterns and mechanisms of regulation. The cAMPdependent signal transductioncascades from cellsurface hormone receptors to AC activation through the “couplingproteins” that we know as heterotrimeric G proteins (Ross and Gilman 1977, Benovic et al.1986, Lohse et al. 1990). Nine different ACs exist in mammals. Their activities are stimulatedby interaction with the subunit of Gs proteins. Gs subunits are coupled with differenttypes of membrane receptors in heterotrimeric complexes together with the ß and subunits,from which they dissociate after the binding of specific ligands (Taussig et al. 1995).Degradation of cAMP is regulated by a large family of PDEs (Beavo 1995). The activities ofACs and PDEs are regulated positively and negatively by other signalling systems, such ascalcium signalling [through calmodulin (CaM), CaMdependent protein kinase II and IV, andcalcineurin], subunits of other G proteins (e.g. Gi, Go and Gq proteins), inositol lipids (byPKC) and receptor tyrosine kinases [through ERK and protein kinase B (PKB)] (Gilman1990, Daub et al. 1996, Dessauer and Gilman 1997, Luttrell et al. 1999, Fisher et al. 2003).
Three major targets of cAMP have been identified: protein kinase A (PKA), the GTPexchange protein for the small GTPase or the exchange protein directly activated by cAMP(EPAC) and the cyclic nucleotidegated ion channels (Kopperud et al. 2003). cAMP couldalso directly regulate the mitogenactivated protein kinase (MAPK) pathway by binding toand activating EPAC and repressor activator protein 1 (Rap1) (Ehses et al. 2003). Althoughthe role of Rap1 in the MAPK pathway is incompletely understood, it seems to act by bindingto and activating mitogenactivated protein kinase kinase kinase BRaf and/or inhibiting theRasRaf (Ras as a G protein and Raf as a mitogenactivated protein kinase kinase) pathway(Zhong 1995, Stork and Schmitt 2002).
PKA is composed of a complex of two regulatory (R) subunits and two catalytic (C) subunits.Four genes (RI , RIß, RII and RIIß) encode the R subunits, and three (C , Cß, C andPrKX) encode the C subunits (Skalhegg and Tasken 2000). PKA is activated by the binding ofcAMP to the R subunits, which induces their dissociation from the C subunit (for review, seeTaylor et al. 2005) Apart from the R subunits, PKA activity is downregulated by protein

22
kinase inhibitors (PKIs). Three known isoforms of PKI , ß and ) are encoded by differentgenes, each of which has a specific expression pattern. Interestingly, PKI could act as achaperone for nuclear export of the C subunit, thereby interfering with the extent of PKAactivity in the nucleus (Taylor et al. 2005, Dalton and Dewey 2006). An important stepforward in our understanding of the action of PKA has been the identification of PKAanchoring proteins (AKAPs). These proteins allow specificity in cAMP signal transduction byplacing PKA close to specific effectors and substrates (Huang et al. 1997).
A large number of proteins have been identified as substrates for PKA. PKA modulates thecAMP response by phosphorylating other components of the cAMP pathway such asreceptors, ACs and PDEs. Depending on the cellular system involved, phosphorylation ofthese proteins results in an enhancement of the cAMP signal (e.g. repressing the activity ofPDE1) (Giorgi et al. 2002, Goraya et al. 2004) or in a rapid downregulation of the pathway toobtain a transient effect (e.g. by repressing the activity of AC V and AC VI, or the 2adrenergic receptor) (for review, see Beazly and Watts 2006). Regulation of transcription byPKA is mainly achieved by direct phosphorylation of transcription factors. cAMP responseelementbinding protein (CREB), cAMP responseelement modulator protein (CREM) andactivating transcription factor 1 (ATF1), originally identified as the activators that respond tocAMP, are phosphorylated by PKA in their activation domains. Several lines of evidencesupport the notion that CREB, CREM and ATF1 can be phosphorylated by many differentkinases (Ziff 1990, Foulkes et al. 1991, Rehfuss et al. 1991, Lalli and SassoneCorsi 1994,SassoneCorsi 1995).
2.2.2 Cyclic guanosine monophosphate (cGMP) signalling
Natriuretic peptides and nitric oxide (NO) activate the cGMP/cGMPdependent protein kinaseG (PKG) signalling pathway. The role of cGMP as a second messenger has long beenrealized, but the mechanism of action has only been addressed in the last couple of decades(Francis and Corbin 1994, Forte and Currie 1995). Most of cGMP's downstream actions(Eigenthaler et al. 1999), and possibly some of cAMP's actions (White et al. 2000), involveactivation of PKGs. Unlike cAMP, however, cGMP has been found to regulate other proteinclasses such as ion channels (Mery et al. 1991, Delay et al. 1997, Hagen et al. 1998, Anthonyet al. 2000) and PDEs (Francis et al. 1994). The two forms of mammalian PKG have distinctand overlapping tissue distributions. Type I isoforms exist as alpha and beta splice variants,are cytosolic and have been recognized in the brain (Tsou et al. 1993), smooth muscle(Komalavilas and Lincoln 1996), platelets, cardiac myocytes (Kwak et al. 1995) and kidney(Gambaryan et al. 1996). The type II isoform is myristylated and membranebound and ispresent in intestinal mucosa (French et al. 1995), kidney (Gambaryan et al. 1996), brain (elHusseini et al. 1995) and bone (Pfeifer et al. 1996). Both PKGs contain aminoterminalleucine zipper motifs, and, in contrast to PKA, exist as homodimers in native tissues (Francisand Corbin 1994).

23
Little is known about PKG regulation of gene expression, and gene expression profiling isjust beginning to contribute to the growing list of cGMPregulated genes (Pilz and Broderik2005). Expression of the immediate early genes cfos and junB, but not of cjun (a componentof the transcription factor activator protein 1) or protooncogene junD, is increased uponactivation of the cGMP pathway. Furthermore, this induction can be inhibited with theselective PKG inhibitor KT5823 (Haby et al. 1994). cAMP response element (CRE)dependent gene transcription is induced by transfected PKG, but weakly, as compared withPKA (Collins and Uhler 1999). Both nitric oxide and atrial natriuretic peptide activate the cfos promoter through a guanylyl cyclase and PKGdependent mechanism (Gudi et al. 1996,Idriss et al. 1999, Lincoln et al. 2006). Many other genes probably are targeted bycGMP/PKG, including vascular endothelium growth factor (VEGF) and the VEGF receptor(Tuder et al. 1995, Dulak et al. 2000), smooth muscle actin and calponin (Boerth et al. 1997,Lincoln et al. 1998), but whether these genes are activated directly by PKG or indirectlythrough crosstalk with other signalling systems is obscure. Reexpression of PKG in culturesof mammalian cells that produce low levels of endogenous protein has been helpful in testingthe requirement for PKG in various signal transduction systems. Gudi et al. (1997) observedthat a specific cGMPdependent nuclear localization signal controls PKG translocation to thenucleus and transactivation of the fos promoter. These data suggest that the cGMP/PKGsystem plays a potentially important and direct role in transcriptional regulation. However,Collins and Uhler (1999) observed exclusively cytoplasmic localization of both PKGisoforms and could only demonstrate nuclear translocation and transcriptional activation whenthe protein was mutated to a chimeric form containing the C subunit of PKG and the PKAcatalytic domain. These findings indicate that PKGs, in contrast to PKA, may, in fact, utilizeadditional cytoplasmic factors to regulate gene expression.
2.2.3. cAMP and cGMP PDEs
The second messengers, cAMP and cGMP, are inactivated by the cyclic nucleotide PDEs(Figure 3). Mammalian PDEs have an HD domain (a superfamily of metaldependentphosphohydrolases; Aravind and Koonin 1998), the Cterminal half and show high affinityfor cAMP and/or cGMP. Protein domains involved in regulation of PDE enzymatic activityand subcellular localization are mainly present in the Nterminal half. Some PDEs havephosphorylation sites targeted by protein kinases and lipid modification sites. Approximately270 aa in the Cterminal catalytic domain are conserved, with a sequence identity of 35%50% among different PDE families (Figure 4). Some PDE families are composed of 2 4subfamily genes showing sequence identity of more than 70% and having identical proteindomain organization. Twentyone PDE genes have been identified in humans, rats, and mice.They are categorized into 11 structurally, biochemically and pharmacologically distinct PDEfamilies, PDE1PDE11 (Manganiello et al. 1995, Soderling et al. 1998a, 1998b, Fawcett et al.
24
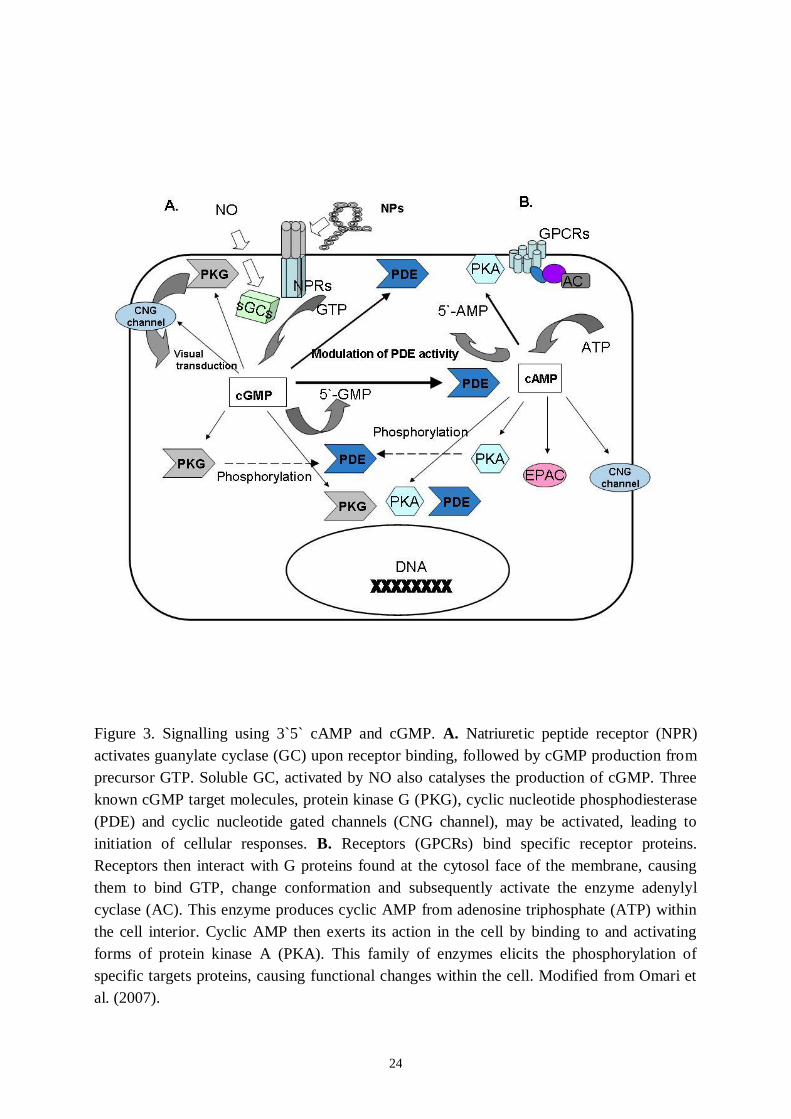
Figure 3. Signalling using 3`5` cAMP and cGMP. A. Natriuretic peptide receptor (NPR)activates guanylate cyclase (GC) upon receptor binding, followed by cGMP production fromprecursor GTP. Soluble GC, activated by NO also catalyses the production of cGMP. Threeknown cGMP target molecules, protein kinase G (PKG), cyclic nucleotide phosphodiesterase(PDE) and cyclic nucleotide gated channels (CNG channel), may be activated, leading toinitiation of cellular responses. B. Receptors (GPCRs) bind specific receptor proteins.Receptors then interact with G proteins found at the cytosol face of the membrane, causingthem to bind GTP, change conformation and subsequently activate the enzyme adenylylcyclase (AC). This enzyme produces cyclic AMP from adenosine triphosphate (ATP) withinthe cell interior. Cyclic AMP then exerts its action in the cell by binding to and activatingforms of protein kinase A (PKA). This family of enzymes elicits the phosphorylation ofspecific targets proteins, causing functional changes within the cell. Modified from Omari etal. (2007).

25
Figure 4. General structure of phosphodiesterases. The different domains of the PDEs aredrawn as boxes connected by wires. PDEs share a common structure, with a conservedcatalytic domain (light blue) flanked by regulator domains (grey and white) the Nterminusand Cterminus. The white box corresponds to regulator domain 1, where severalphosphorylation sites or allosteric binding sites have been mapped. Modified from Mehats etal. (2002).
2000, Conti and Beavo 2007), some of which specifically hydrolyse cAMP (PDE3, PDE4,PDE7 and PDE8) and others cGMP (PDE5, PDE6 and PDE9). The PDEs belonging to thePDE1, PDE2, PDE10 and PDE11 families are dualsubstrate enzymes, hydrolysing bothcAMP and cGMP (Soderling et al. 1998a, 1998b, Fawcett et al. 2000). Most PDE familiescontain multiple genes (subtypes), and most of these genes can produce multiple transcripts orvariants (Conti and Jin 2000). The unique characteristics of each PDE gene family are definedby protein domains located in the Nterminal to the catalytic unit. As shown in Figure 5,approximately half of the PDE gene families (PDE2, PDE5, PDE6, PDE10 and PDE11) havea protein domain termed GAF in tandem and are therefore designated a GAFPDE
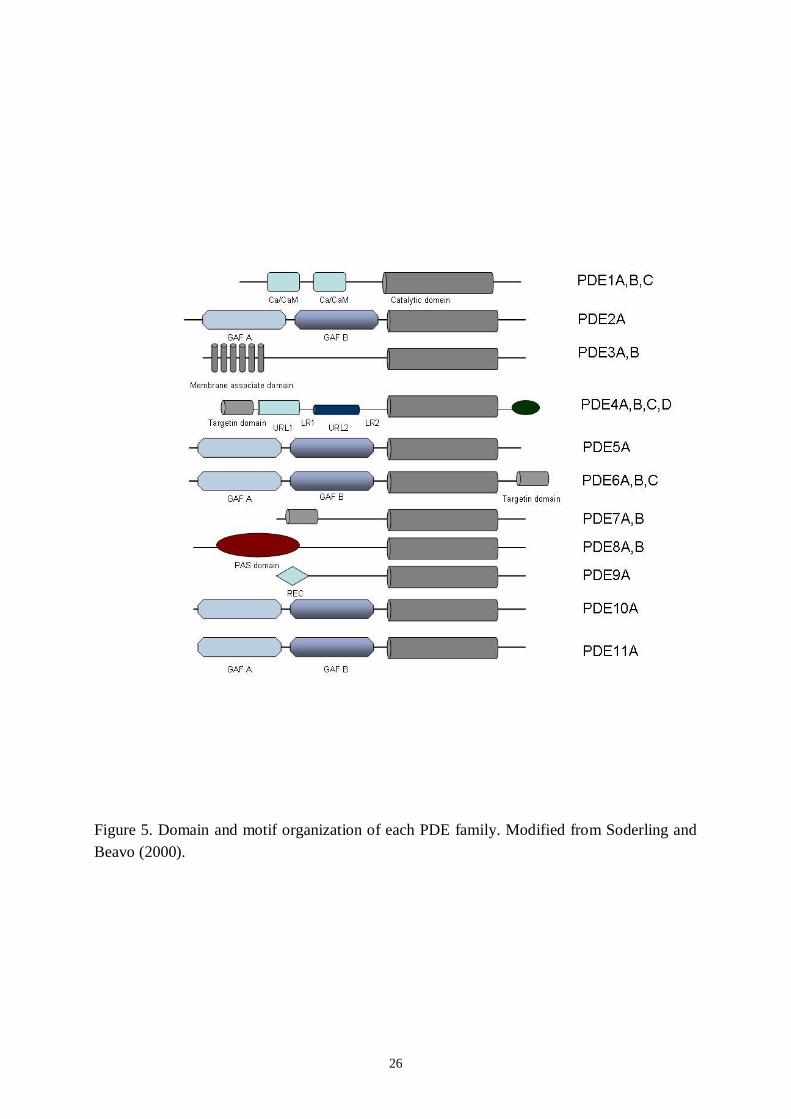
26
Figure 5. Domain and motif organization of each PDE family. Modified from Soderling andBeavo (2000).

27
subfamily.The known functions of GAF domains are cGMP bindingmediated allostericregulation and dimerization of GAFPDEs. Some GAF domains have also been reported tobind cAMP. Other PDEs (PDE1, PDE3, PDE4 and PDE7–9) have no GAF domain and belongto the non–GAFPDE subfamily. PDE1 contains a Ca2+/CaMbinding site, PDE3 has atransmembrane domain, PDE4 has upstream conserved regions (UCRs) and PDE8 has aresponse regulator receiver (REC) domain and a per–arnt–sim (PAS) domain. PDE7 andPDE9 have no specific protein domain in addition to the PDE catalytic domain.
Each PDE family is distinguished functionally by its unique combination of enzymaticcharacteristics and pharmacological inhibitory profiles (Table 2). Different families, subtypesand variants are varied in different tissues and cell types. One of the main roles of the PDEs isto determine the amplitude of cyclic nucleotidemediated hormonal responses and tomodulate the duration of the signal. Therefore, the PDEs can function as modulators of thecyclic nucleotidemediated signal transduction pathways (Manganiello et al. 1995, Mehats etal. 2002, Housley et al. 2003, Conti et al. 2003). The activities of PDE are regulated bymultiple inputs from other signalling systems and PDEs are crucial in crosstalk betweenpathways.
In the nomenclature used for each PDE isozyme (e.g. HsPDE1A1), the first two lettersindicate the animal species and the first Arabic number after PDE designates the PDE genefamily. This number is followed by a single capital letter indicating a distinct subfamily gene.The last Arabic number indicates a specific splice variant or a specific transcript generatedfrom a unique transcription initiation site (http://depts.washington.edu/pde/pde.html).
2.2.3.1 PDE1 family
Three subfamily genes, PDE1A–C with various splice variants encode Ca2+/CaMdependentcAMP and cGMPhydrolysing PDEs. PDE1 is mainly present in the cytosolic fraction;however PDE1 has also been found in the fibres of several neurons from the dorsal rootganglion (Giorgio et al. 2002). In humans, PDE1A shows high affinity for cGMP (Loughneyet al. 1996). PDE1B hydrolyses cGMP with a Km value lower and a Vmax value higher thanthose for cAMP (Bender et al. 2005). High affinity for both cAMP and cGMP is observedwith PDE1C (Loughney et al. 1996). In cells expressing PDE1, hormones that raise cytosolicCa2+ would active PDE1, in this way diminishing the cyclic AMP response to hormonesstimulating cAMP or cGMP synthesis. Sustained Ca2+ entry into the cells is required toactivate PDE1A in the astroma cell line, and PDE1A reportedly cannot discriminate betweenthe different sources of Ca2+ entry (Goraya et al. 2004). By contrast, in vivo phosphorylationof PDE1 would likely result in the potention of cAMP or cGMP accumulation, involving theelevation of cytosolic Ca2+ and activation of guanylyl or adenylyl cyclases, and would play amajor role in the amplification and prolongation of cyclic nucleotide effects. Differentalterations in tissue and cell PDE1 isozymes may occur by shortterm (phosphorylation,proteolysis) or longterm regulation (at the mRNA level) in response to hormonal stimulation
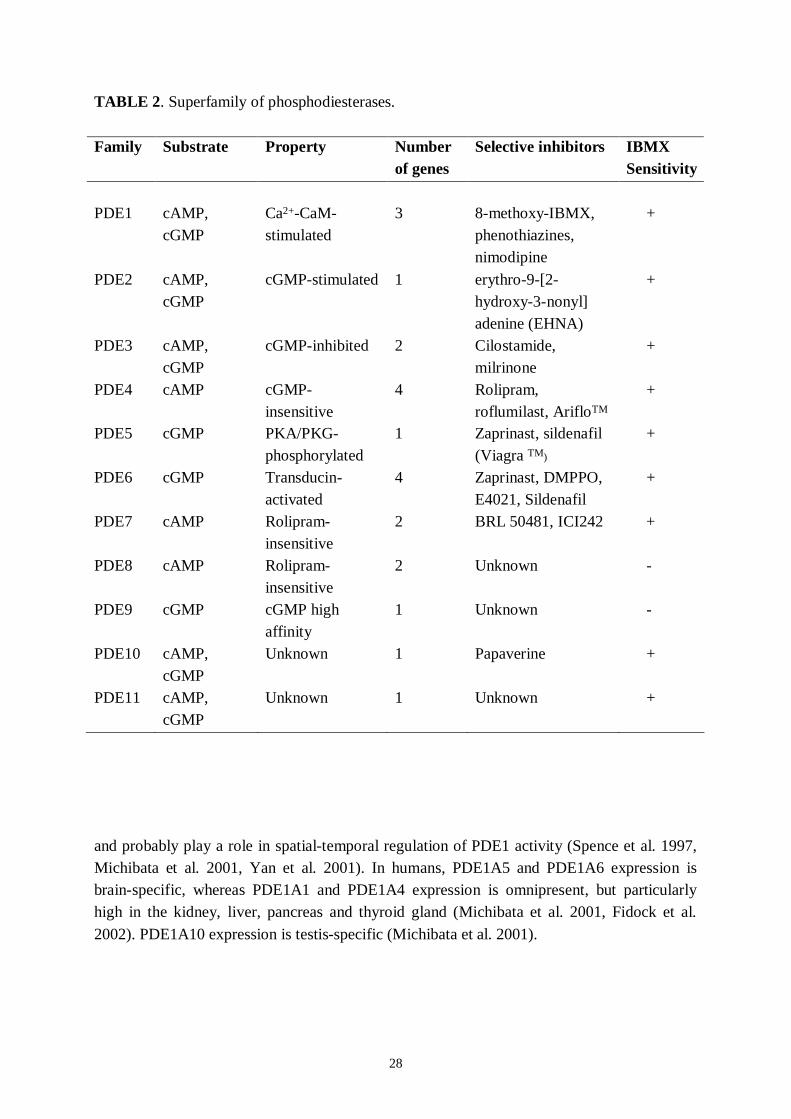
28
TABLE 2. Superfamily of phosphodiesterases.
Family Substrate Property Numberof genes
Selective inhibitors IBMXSensitivity
PDE1 cAMP,cGMP
Ca2+CaMstimulated
3 8methoxyIBMX,phenothiazines,nimodipine
+
PDE2 cAMP,cGMP
cGMPstimulated 1 erythro9[2hydroxy3nonyl]adenine (EHNA)
+
PDE3 cAMP,cGMP
cGMPinhibited 2 Cilostamide,milrinone
+
PDE4 cAMP cGMPinsensitive
4 Rolipram,roflumilast, ArifloTM
+
PDE5 cGMP PKA/PKGphosphorylated
1 Zaprinast, sildenafil(Viagra TM)
+
PDE6 cGMP Transducinactivated
4 Zaprinast, DMPPO,E4021, Sildenafil
+
PDE7 cAMP Rolipraminsensitive
2 BRL 50481, ICI242 +
PDE8 cAMP Rolipraminsensitive
2 Unknown
PDE9 cGMP cGMP highaffinity
1 Unknown
PDE10 cAMP,cGMP
Unknown 1 Papaverine +
PDE11 cAMP,cGMP
Unknown 1 Unknown +
and probably play a role in spatialtemporal regulation of PDE1 activity (Spence et al. 1997,Michibata et al. 2001, Yan et al. 2001). In humans, PDE1A5 and PDE1A6 expression isbrainspecific, whereas PDE1A1 and PDE1A4 expression is omnipresent, but particularlyhigh in the kidney, liver, pancreas and thyroid gland (Michibata et al. 2001, Fidock et al.2002). PDE1A10 expression is testisspecific (Michibata et al. 2001).

29
2.2.3.2 PDE2 family
PDE2A hydrolyses both cGMP and cAMP with similar maximal rates and relatively high Km
values. PDE2A is allosterically stimulated by cGMP binding to its GAF domain (Martinez etal. 2002), enabling the simultaneous regulation of both cAMP and cGMP signalling. PDE2was shown to play a major feedback role by restoring the basal level in cyclic nucleotides inresponse to hormonal stimulation of the adrenal gland (MacFarland et al. 1991). Threevariants of a single gene were cloned for PDE2: PDE2A1, PDE2A2 and PDE2A3(Sonnenburg et al. 1991, Yang et al. 1994, Rosman et al. 1997). PDE2 activity is upregulatedin vivo at the posttranscriptional level by PKC under 4 betaphorhol 12myristate 13 acetate(Geoffroy et al. 1999). In endothelial cells, PDE2 is upregulated during phenotype changes(Keravis et al. 2000) as well as under stimulation by VEGF (Favot et al. 2004), indicatingPDE2 participation in endothelial cell proliferation. PDE2 protein is mainly present in theadrenal medulla, heart, rat ventricle (Yanaka et al. 2003), brown adipose tissue (Coudray et al.1999), liver and brain.
2.2.3.3 PDE3 family
PDE3A and PDE3B, subfamily genes of PDE3, show a high affinity for both cAMP andcGMP. A low Vmax value for cGMP compared with that for cAMP allows cGMP to functionas a competitive inhibitor for cAMP hydrolysis (Degerman et al. 1997). Therefore, PDE3s aretermed cGMPinhibited cAMP PDEs. The presence of a 44aa insert in the catalytic domain isa unique characteristic of the PDE3 family. Another special feature is the presence of Nterminal hydrophobic membrane association domains (Wechsler et al. 2002). Shorttermactivation of PDE3 was firstly demonstrated in rat fat cells in response to insulin andisoprenaline stimulations, which induce PDE3 phosphorylation (Degerman et al. 1990). Asingle phosphorylated serine site (Ser 302) was identified (Rahn et al. 1996). This site couldbe phosphorylated either by PKA or by PKB, but the major site of PKB phosphorylation isSer 273 (Kitamura et al. 1999). Possible activation of PDE3B by phosphoinositide 3kinasephosphorylation (Rondinone et al. 2000) was recently implicated in the hypothalamic actionof leptin on feeding (Zhao et al. 2002) as well as in cell insulin secretion (Zhao et al. 1998).Furthermore, hormonal stimulation (insulin, glucagon) was shown to activate in vivo PDE3associated with the Golgiendosomal fraction (Geoffroy et al. 2001). PDE 3A is mainlypresent in the heart muscle cells, platelets, vascular smooth muscle cells and oocytes(Reinhardt et al. 1995, Degerman et al.1997, Wechsler et al. 2002), whereas PDE3B is mainlyassociated with adipocytes, hepatocytes and spermatocytes (Miki et al. 1996).

30
2.2.3.4 PDE4 family
The PDE4 family exclusively hydrolyses cAMP (Km=24 µM). Currently, the PDE4 familyrepresents the largest and most studied PDE family, comprising four highly similar subfamilygenes, PDE4AD, that encode rolipramsensitive PDEs. The PDE4 family includes variousalternative splice variant groups encoding long PDE4 and short PDE4 isozymes, with aminimum of 35 different PDE4 proteins (Houslay 2001) based on the presence or absence ofNterminal upstream conserved region (UCR) domains (Bolger et al. 1993). The longformvariants have UCR1, linker region (LR) 1, UCR2, LR2, and a catalytic domain. The shortform variants contain LR1UCR2LR2 and UCR2 (truncated)LR2 in the Nterminal region.UCR1, which includes one PKA phosphorylation site, is connected to UCR2 by LR1. UCR2has a hydrophilic Nterminal region, which intramolecularly interacts with the hydrophobic Cterminal portion of UCR1 (Beard et al. 2000). UCR1 and UCR2 are involved in PDE4enzymatic regulation (Sette and Conti 1996, Hoffman et al. 1999, Grange et al. 2000) throughUCR2 interaction with the catalytic domain (Lim et al. 1999) and have also been reported toparticipate in PDE4 dimerization (Richter and Conti 2002). Although the PDE4 catalytic sitebinds to the competitive inhibitor rolipram, the affinity of this site for rolipram can changemarkedly depending on the conformation of the enzyme. PDE4 activity is controlled byphosphorylation, a relationship with a protein or endogenous mediator and proteolysis. Thepresence of an acceptor site in UCR1 for PKAmediated phosphorylation allows a fast changein PDE4 activity. In vivo, this augmentation of PDE4 activity was shown as a result of aextended increase of cAMP resulting from hormonal stimulation; this was demonstrated to bea shortterm feedback mechanism allowing cAMP levels to return to the basal cellular state(Sette et al. 1994, Oki et al. 2000). An extended accretion of cAMP induces a longtermregulation of PDE4 induction. Folliclestimulating hormone and dibutyryl cAMP treatmentwas shown to start the specific induction of shortform PDE4D1 and PDE4D2 in rat Sertollicells, whereas the expression of the longform PDE4D3 was unchanged (Swinnen et al.1991). Conti et al. (1995) suggested that the PDE4D gene had two distinct transcriptionalunits, the unit controlling the shortform expression being upregulated by cAMP.Correspondingly, cAMPdependent upregulation was shown for PDE4A and PDE4B inU937 (Torphy et al. 1995), for PDE4B2 in human monocytes (Manning et al. 1996) and,more recently, in human myometrial cells (Oger et al. 2002). Furthermore, the downregulation of PDE4A and PDE4B could be connected to PDE4D upregulation, as shown inendothelial cells during phenotypic variance (Keravis et al. 2000), whereas PDE4A was upregulated and PDE4B and PDE4D were downregulated in the brain during fluoxetinetreatment (Miro et al. 2002). In humans, PDE4APDE4D expression is fundamentallyubiquitous (Engels et al. 1994, 1995, Bolger et al. 1997, Wang et al. 1999, Wang et al. 2003),with a variantspecific tissue distribution pattern, but PDE4A has been reported to berelatively high in the brain (Bolger et al. 1993) and PDE4B low in the lung and absent inblood (Engels et al. 1995).

31
2.2.3.5 PDE5 family
To date, only one gene product has been reported for this family, PDE5A, which has twoGAF domains (GAF A and GAF B) in the Nterminal half and specifically hydrolyses cGMP.The GAF A domain of PDE5A has been reported to be responsible for this enzyme`sallosteric binding to cGMP (Liu et al. 2002), and therefore PDE5A is termed a cGMPbindingcGMPspecific PDE. One PKG and PKAdependent phosphorylation site in the Nterminalregion is related to activation of the PDE5A enzyme (Corbin et al. 2000). cGMP binding tothe PDE5A GAF A domain promotes phosphorylation, which not only activates the catalyticfunction but also increases cGMPbinding affinity (Corbin et al. 2000, Francis et al. 2002,Zoraghi et al. 2005). In humans, PDE5A transcripts are abundantly found in various tissues,especially in smooth muscle tissues, and are also detected in platelets (Yanaga et al. 1998,Kotera et al. 1999). PDE5A1 and PDE5A2 transcripts are widely distributed (Kotera et al.1999, Lin et al. 2000). In contrast, specific expression of PDE5A3 in smooth and/or cardiacmuscle has been suggested (Lin et al. 2000).
2.2.3.6 PDE6 family
In retinal rod and cone cells, the level of cGMP, a second messenger in visual signaltransduction, is tightly controlled through regulated cGMP hydrolysis by PDE6AC subfamilygenes (photoreceptor PDEs) (Cote 2004). Lightactivated transducin stimulates PDE6 activityby removing the inhibitory subunit . Membrane hyperpolarization is caused by cGMPelimination, leading to an electrical cellular response. PDE6 and PDE6ß subunits encoded bythe PDE6A and PDE6B genes, respectively, form a holoenzyme, rod PDE, with two copies ofthe smaller inhibitory subunit (PDE6 ) encoded by PDE6G. The GAF A domain is relatedto PDE6 ß heterodimerization (Muradov et al. 2003). In cone cells, a homodimer of 2 'subunits (PDE6 ') encoded by PDE6C comprises cone PDE with conespecific inhibitorysubunits encoded by PDE6H. Thus, regulation of PDE6 activity by small inhibitory subunits isa unique aspect of this PDE family. Several different genetic diseases caused by defects inPDE6 function or expression have been characterized; in humans, for instance, autosomaldominant congenital stationary night blindness and autosomal recessive retinitis pigmentosahave been attributed to PDE6B mutations (Gal et al. 1994).
2.2.3.7 PDE7 family
This family includes two genes, PDE7A and PDE7B, that encode rolipraminsensitive highaffinity cAMPspecific PDEs (Km value approximately 0.2 µmol/L) (Michaeli et al. 1993).The PDE7 family contains neither GAF domains nor regulatory domains (Beavo 1995).PDE7A and PDE7B subtypes show approximately 70% homology (Gardner et al. 2000,Sasaki et al. 2000). A PKA pseudosubstrate site is present in the Nterminus of the PDE7Asubfamily. Alternative splicing for PDE7A and tissuespecific expression of PDE7 splice

32
variants have been identified (Bloom and Beavo 1996, Han et al. 1997). PDE7A1 mRNA andprotein are distributed ubiquitously across human proinflammatory and immune cells (Smithet al. 2003). On the other hand, PDE7A2, which is generated by 5'splicing and differs fromPDE7A1 at its Nterminus (Bloom and Beavo 1996, Han et al. 1997), has never been detectedat the protein level, despite unequivocal identification of its mRNA (Smith et al. 2003). Thedistribution of PDE7A3 is largely unknown, but in human it has been found in Tlymphocytes(Glavas et al. 2001) and may also be present in many PDE7A1expressing cells, as bothtranscripts are probably regulated by the same promoter (TorrasLlort and Azorin, 2003). Inrats, three Nterminal splice variants, PDE7B13, exist. Only PDE7B1 has been identified inhumans and mice and it is abundant in the brain, liver, heart, thyroid glands and skeletalmuscles (Gardner et al. 2000).
2.2.3.8 PDE8 family
PDE8A and PDE8B are the subfamily genes of PDE8. PDE8s are highaffinity cAMPspecific PDEs insensitive to rolipram and 3isobutyl1methylxanthine (IBMX) (Gamanumaet al. 2003), and contain response regulator receiver (REC) and PAS domains in the Nterminal portion. The REC domain functions as a receiver of signals from the sensorcomponent in a 2component signal transduction system in lower organisms. The PAS domainis involved in the binding of small ligands and protein–protein interactions (Huang et al.1993). However, regulation of PDE8s via REC or PAS domain is unknown, and obviousPDE8 activity has not yet been demonstrated in either tissue or cell extracts. PDE8Atranscripts are expressed in various tissues and are abundant in the testis, ovary, smallintestine and colon (Fisher et al. 1998). The expression of PDE8B seems to be confined to thethyroid gland and the brain (Kobayashi et al. 2003).
2.2.3.9 PDE9 family
PDE9A is the only reported subfamily gene of PDE9, although more than 20 splice variantshave been observed in humans, indicating that the PDE9A gene appears to have a complexregulation of expression (Rentero et al. 2003). PDE9A specifically hydrolyses cGMP withhigh affinity (Fisher et al. 1998, Guipponi et al. 1998). However, no reports are available onthe regulation of PDE9A activity or on the presence of endogenous PDE9A activity in eithertissue or cell extracts. IBMXinsensitive cGMP PDE activity, which has been shown to date,is likely attributable to PDE9. PDE9 mRNA is highly conserved between species and iswidely distributed throughout the rodent brain (Van Staveren et al. 2002). PDE9A mRNA isexpressed in the spleen, small intestine, brain, colon, prostate, kidney and placenta (Fisher etal. 1998, Guipponi et al. 1998). In humans, the PDE9A5 splice variant is highly expressed inimmune tissues (Wang et al. 2003).

33
2.2.3.10 PDE10 family
PDE10A contains two Nterminal GAF domains and hydrolyses both cAMP and cGMP. Twomajor variants, PDE10A1 and PDE10A2, and several minor variants have been identified inhumans (Fujishige et al. 1999a, 2000). High affinity for cAMP inhibits cGMP hydrolysis,making this enzyme a cAMPinhibited dualsubstrate PDE. Among newly discovered PDEs,the enzymatic activity of PDE10A is clearly demonstrated in tissue extracts (Fujishige et al.1999b). The enzymatic activity of a chimeric construct of the PDE10A GAF domain andcyanobacterial adenyl cyclase is stimulated by cAMP, suggesting a possible allostericmodulation of PDE10A activity by cAMP (GrossLangenhoff et al. 2006). PDE10 transcriptsare particularly abundant in the brain, thyroid and testis. The PDE10 family was recentlyshown to be associated with a progressive neurodegenerative disease, Huntington`s disease(Hebb et al. 2004, Hu et al. 2004).
2.2.3.11 PDE11 family
The most recently identified family, PDE11, contains four Nterminal variants, PDE11A1PDE11A4. A fulllength form, PDE11A4, contains two GAF domains and a catalytic domain.PDE11A hydrolyses both cAMP and cGMP with similar Km values (Fawcett et al. 2000,Hetman et al. 2000, Yuasa et al. 2000). Although a cyanobacterial adenyl cyclase fused withPDE11A4 GAF domains is activated by cGMP (GrossLangenhoff et al. 2006), no reportsexist on the allosteric regulation of the PDE11A enzyme. PDE11A activity has not yet beenclearly demonstrated in tissue or cell extracts. In humans, PDE11A expression is strong in theprostate and moderate in the testis and several other tissues (Fawcett et al. 2000, Yuasa et al.2000).
2.2.4 Regulation of activity and compartmentalization of PDEs
PDEs exist universally, from prokaryotes to eukaryotes. In all organisms, multiple PDEsappear to be involved in tight feedback regulation of cAMP and cGMP. Observations alsoimply that different combinations of PDEs can regulate the same general function in differentspecies. Divergent PDE expression patterns have been detected in the hearts of the rodentsand humans. Two general types of PDE regulation occur. Shortterm regulation involves theactivation of second messenger pathways and consequent allosteric modulation of enzymeactivity, and longterm regulation occurs through increased enzyme synthesis. Several PDEfamilies are subject to one or both forms of regulation. Functional evidence indicates thatcyclic nucleotide signals are confined in compartments and that different PDEs, as well as thepresence of PDE/PKA or PDE/PKG complexes, may contribute to establishment orstabilization of these compartments. However, the exact mechanism restricting cyclic

34
nucleotide diffusion remains unclear, and convincing evidence for PDEgenerated cyclicnucleotide gradients is still lacking (for review, see Conti and Beavo 2007).
2.2.5 Inhibitors of PDEs
The PDEs are targets for a number of synthetic PDE isozyme inhibitors. Theophylline was thefirst chemical inhibitor described for PDEs (Butcher and Sutherland 1962). One decade later,a new xanthine analogue, IBMX, was shown to be 100fold more potent than theophylline(for review, see Chasin and Harris 1976). These two compounds are both nonspecific PDEinhibitors. The last decade, numerous more or less selective inhibitors have been designed andsynthesized for scientific and pharmacological purposes. Some of them inhibit PDE activityvery potently in the nanomolar and subnanomolar ranges and are specific to certain PDEfamilies. Thus, in the span of several decades, the potency of PDE inhibitors has increased by108, and the specificity of some of them by 104. Recently, several selective inhibitors forindividual genes within the PDE family have been discovered and synthesized, e.g. OPC33540 inhibits PDE3A (IC50= 0.32 nM) (Sudo et al. 2000) and IC242 inhibits PDE7Aselectively, with an IC50 of 0.37 µM (Lee et al. 2002a, 2002b). Efforts are now focused ondeveloping thirdgeneration inhibitors that are subtypespecific. PDE inhibitors areexperimental tools singularly useful in studies of the role of PDEs in cyclic nucleotidesignalling and also represent a novel class of pharmacological agents suited for “signaltransduction pharmacotherapy” (Levitzki 1997). Currently, PDE1, PDE2, PDE3, PDE4,PDE5, PDE6, PDE7 and PDE10 have more or less selective inhibitors, but no selectivechemical inhibitors have yet been described for PDE8, PDE9 or PDE11 (Table 2). Because oftheir great market potential and therapeutic relevance, PDE inhibitors became recognized asimportant therapeutical agents for a variety of diseases, such as heart failure, depression,asthma, inflammation and erectile dysfunction (Conti et al. 1995, Torphy 1998, Mehats et al.2002, Rotella 2002, Halena and Siegel 2007). Several PDE inhibitors are currently on the
market, e.g. sildenfil (Viagra®), tadalafil (Cialis®) and vardenafil (Levitra®); all of these areinhibitors of PDE5.
2.2.6 Bone metabolism and PDE inhibitors
Several studies have shown that selective chemical PDE inhibitors promote osteoblasticdifferentiation and increase bone formation both in vivo and in vitro (Miyamoto et al. 1997,2003, Kinoshita et al. 2000, Wakabayashi et al. 2002, Scutt et al. 2004). Most of these effectsare likely to be mediated by elevated cAMP levels, although cAMPindependent effects havealso been reported (Rawadi et al. 2001). In osteoblasts, for example, cAMP produced inresponse to PTH or PGs regulates osteoblastic differentiation (Kumegawa et al. 1984,Farndale et al. 1988, Partridge et al. 1994, Ishizuya et al. 1997). There are corresponding datathat show that administration of PTH or PGs also leads to increases in cancellous bone

35
volume in animal models (Jee et al. 1985, 1987, High 1987, Finkelstein et al.1994, Whitfieldand Morley 1995, Reeve 1996). In rat bone marrow culture, XT44 (a PDE4 inhibitor) androlipram stimulated mineralnodule formation, whereas it inhibited osteoclastlike cellformation in mouse bone marrow culture (Waki et al. 1999). Therefore, PDE inhibitors areconsidered to be promising candidates for antiosteoporosis drug therapies. A PDE4 inhibitor,XT611, has been shown to inhibit osteoclast multinucleation or maturation by actingsynergistically on osteoclast progenitors with prostaglandin E2 (PGE2) secreted from stromalcells, but not by influencing the celltocell interaction between stromal cells and osteoclastprogenitors (Yamagami et al. 2003). Rolipram has augmented the inhibitory effects of PGE2on osteoclast progenitor cell viability; thus combined treatment with PGE2 and rolipramsuppresses osteoclast formation by directly reducing osteoclast progenitor cell viability (Parkand Yim 2007). By contrast, PDE inhibitors have been shown to stimulate osteoclastformation by inducing receptor activator of NF{kappa} B ligand (TRANCE) expression inmice calvarial osteoblasts and UAMS32 cells (Takami et al. 2005). Pentoxifylline androlipram have reportedly been to increase bone mass in animals given these drugs in dailyinjections (Kinoshita et al. 2000). PTH as well as PDE inhibitors induced osteoclast formationin a mouse coculture system. Injections of PTH to animals once per day produced a netanabolic effect in vivo (Tam et al. 1982, Fu et al. 2002). Continuous exposure to PTH leads toan increase in both bone formation and bone resorption, with a net loss of bone mass (Tam etal. 1982). PDE inhibitors may induce bone loss if administered continuously (Takami et al.2005).
2.3 Other signal transduction pathways associated with PDEs
PDE isoenzymes are targets of intricate regulatory mechanisms that integrate differentsignalling pathways, function in feedback to modulate signalling pathways and serve aseffectors of signalling pathways. An additional property that only recently is being explored isthe subcellular distribution of PDEs and their involvement in establishing microdomains andchannelling of signals. Future pathway studies are anticipated to shed light on the role ofPDEs in cyclic nucleotide signal transduction and connections between other signal pathways.
2.3.1 Exchange protein directly activated by cAMP (EPAC) and PDEs
In the heart, signalling events such as the onset of cardiac hypertrophy are influenced bymusclespecific mAKAP signalling complexes that target PKA, the EPAC and PDE4 (DodgeKafka et al. 2005). Mediation of signalling events by AKAPs might also have a role inlipolysis in adipocytes. cAMP either stimulates or inhibits PKB phosphorylation, actingthrough different effectors, in rat adipocytes. Hence, a cAMP pool that activates EPAC and isregulated by PDE3B and PDE4 appears to mediate a decrease in PKB phosphorylation. PKB

36
phosphorylation, and therefore activation, in adipocytes is regulated by an orchestrated crosstalk between insulin and cAMP pathways, where PDE3B seems to play an important role(ZmudaTrzebiatowska et al. 2007).
2.3.2 Mitogenactivated protein kinase (MAPK) and PDEs
PDEs have been shown to be involved in MAPK signalling; e.g. pentoxifylline, a nonselective PDE inhibitor, induces osteoblastic differentiation in pluripotent mesenchymal celllines C3H10T1/2 and C2C12. Pentoxifylline treatment enhances osteoblast differentiation,which occurs via signalling by MAPK cascades and is independent of PKA activation(Rawadi et al. 2001). Rat mesangial cells (MCs) possess functionally compartmentalizedintracellular pools of cAMP that are differentially regulated by cAMP PDEs. Inhibition ofPDE3, but not PDE4, has been demonstrated to suppress MC mitogenesis (Tsuboi et al.1996). MC mitogenesis is regulated through "negative crosstalk" between cAMP PKA andextracellular signalregulated kinase (ERK) signalling. PDE3 inhibitors act by suppressingERK activation. Other targets of cAMP signalling directed by PDE3 include cyclins D, E andA, and the cell cycle inhibitor p21 (Cheng et al. 2004).
2.3.3 PKC and PDEs
PDEs have been reported to be involved in the noncanonical Wnt signalling pathway. Wntbinds to a Frizzled receptor and possibly also the coreceptor, Knypek or Ror2. This can leadto activation of a pathway involving calcium/CaMdependent kinase II and protein kinase C.Frizzled may activate heterotrimeric GTPbinding proteins, leading to activation ofphospholipase C (PLC) and PDE. Frizzled can also recruit Dishevelled, leading to activationof small GTPbinding proteins, such as Rho and Cdc42 (Veeman et al. 2003). PKC, cAMP,PLC, Ca2+ and PDEs have been shown to be involved oocyte maturation in amphibians(Toranzo et al. 2007). The roles of PKC and PDE have been demonstrated in opioidagonistinduced reductions in cAMP levels in the isolated irisciliary bodies of the rabbit(DortchCarnes and Potter 2002). In addition, opioid receptor stimulation in rat heart hasshown PDE4 activation by PKC (Bian et al. 2000).
2.4 Bioactive peptides
During the last decade, fundamental studies have opened a new field of research concerningbioactive or biogenic substances derived from foods (Meisel 2001). Proteins and peptidesfrom food have been found to be physiologically active or bioactive either in a direct mannerthrough their presence in the undisturbed food itself or after their release from the respectivehost proteins by hydrolysis in vivo or in vitro, e.g. in cheese ripening (Rizzello et al. 2005a),

37
food fermentation (Fitzgerald and Murray 2006, Korhonen and Pihlanto 2006) or enzymaticreactions in the gut after the ingestion of foods containing precursor proteins (Meisel et al.2003, HernandezLedesma et al. 2004). Bioactive peptides have been found, among others, inmeat, fish, egg, soy and grain products, but thus far most have been isolated from milkbasedproducts. Functionally bioactive peptides are usually unknown sequences 25 aa in length andhave a proline residue in the Cterminus (Curtis et al. 2002). Two human di/tripeptidetransporters have been identified; hPepT1 is expressed both in the small intestine and in theproximal tubule, whereas hPepT2 is expressed only in the proximal tubule (see Nielsen andBrodin 2003, Steffansen et al. 2004). The cellular mechanisms and signalling of bioactivepeptides remain obscure.
Different health effects have been attributed to foodderived peptides. Antimicrobial peptideshave been recognized in many protein hydrolysates, particularly in milk. The most studied arethe lactoferricins, which are derived from bovine and human lactoferrin (Kitts and Weiler2003). In addition, antibacterial peptides have also been recognized in s1casein and s2casein (Rizzello et al. 2005b, McCann et al. 2006). Antimicrobial peptides operate againstdifferent Grampositive and Gramnegative bacteria, yeast and filamentous fungi. The ACEsare involved in the regulation of the specific maturation or degradation of a number ofmammalian bioactive peptides. Since the first detection of exogenous ACE inhibitors in snakevenom (Ondetti et al. 1977), a large number of ACE peptides have been discovered in thedigestion of different food proteins, being established particularly in milk but also in fish andmeat. ACE inhibitory peptides are generally shortchain peptides, often carrying polar aminoacid residues like proline (Fuglsang et al. 2003) The hypertensive and immunomodulatorpeptides ValProPro and IleProPro, for example, can be released from precursor proteins
casein and casein) by enzymes from Lactobacillus helveticus (Hata et al. 1996,Korhonen and Pihlanto 2003). The antihypertensity activities of bioactive peptides on bloodpressure have been shown in several studies (e.g. Mitsui et al. 2004, Miguel and Aleixandre2006). Hypocholesterolemic effects have been reported for casein and wheyderived peptides,in addition to peptides from soy protein (Hori et al. 2001). Antithrombotic and antioxidantactivities of bioactive peptides (Kitts and Weiler 2003) have been shown, as has enhancementof mineral absorption and/or bioavailability in rats (Erba et al. 2002) and in the humanintestine (Chabance et al. 1998, Meisel et al. 2003). Immunomodulatory peptides can improveimmune cell functions, measured as lymphocyte proliferation, natural killer cell activity,antibody synthesis and cytokine regulation (Horiguchi et al. 2005, Fitzgerald and Murray2006). Furthermore, immunomodulatory peptides might reduce allergic reactions in atopichumans and augment mucosal immunity in the gastrointestinal tract (Korhonen and Pihlanto2003). Cytochemical studies have given increasing evidence that bioactive peptides modulatethe proliferation, differentiation and apoptosis of various cell types. Some milkderivedpeptides, for instance, have been shown to trigger apoptosis, mainly in malignant cells,whereas normal cells seem to be less susceptible (Meisel and Fitzgerald 2003). Peptides havealso been observed to have opioid activities; for example, opioid receptor ligands withagonistic activity originate from different milk proteins and exert naloxoneinhibitable opioid

38
activities in both receptor studies and bioassays (Kayser and Meisel 1996, Mullally et al.1996).
2.4.1 Bioactive peptides in bone
Effects of bioactive peptides have been studied in dental/orthopaedic biomaterials as well asin basic bone biology. Proactive or newgeneration biomaterials achieve specific, timely anddesirable responses from surrounding cells and tissues. Osteoblast adhesion is a crucialprocess at bone implant interfaces before multiple osteoblast functions (such as proliferationand mineralization), which occur after cell adhesion and are necessary for the success of animplant (Dee 1996a). In the past two decades, many cell adhesion molecules have beendiscovered, and their functions in cell morphology, locomotion, mitosis, cytokinesis,phagocytosis and maintenance of cell polarity have been investigated (Pavalko and Otey1994, Haass et al. 2005, Turowski et al. 2005). A key finding remains that the amino acidsequence arginineglycineaspartic acid (RGD) in fibronectin serves as a primary cellattachment cue. The RGD sequence is expressed in many extracellular matrix (ECM)molecules, and the responsiveness of the RGD sequence for all attachments has been shownnumerous times. The small synthetic peptides (often hundreds of daltons) that contain theamino acid sequence RGD can apparently mediate cell attachment similarly to theirconsiderably larger parental molecules (100 000 Daltons). According to Craig et al. (1995),the potential exists to develop RGDbased therapeutics that function either as agonist, topromote the interaction of cells and tissues with artificial matrices or as antagonists to controlthe nature of cellcell and cellECM interactions. The coating of biomaterials with RGDpeptides offers an attractive strategy for controlling the cellmaterial interface and achieving abioactive implant (Kantlehner et al. 1999, Verrier et al. 2002). Peptides have been shown tosupport and enhance osteoblast adhesion (Dee et al. 1998, Schuler et al. 2006), proliferation(Huag et al. 2003, Benoit and Anseth, 2005) and mineralization (Dee et al. 1996b). Based onthe RGD structure, synthetic RGDmimetic compounds have been designed. These mimeticcompounds are effective inhibitors of bone resorption in vitro and in vivo (James et al. 1996,Engleman et al. 1997).
In addition, the effect of other small peptides (without RGD sequence), such as isoleucineprolineproline (IPP) and valineprolineproline (VPP), have been studied in bone cells. IPPand VPP, fermented from milk by Lactobacillus helveticus, increase activation of boneformation in osteoblast culture, but do not have an impact on osteoclast activation and boneresorption in vitro (Narva et al. 2004). L. helveticusfermented milk has been demonstrated toprevent bone loss in ovariectomized rats, but this phenomenon could not be seen followingsynthetic VPP treatment (Narva et al. 2007).

39
3. Aims of the study
This study focused on the areas of bone cell biology and signalling transduction. PDEs are animportant part of signal transduction in the cell, but very little is known about them in bonecells. The inhibition of some PDEs could have therapeutic potential in the treatment ofosteoporosis. Short bioactive peptides have been shown to have modulatory effects on bonebiology.
The aim of this study was to identify PDE families in human osteoblasts and osteosarcomacells as well as to investigate the role of PDEs and bioactive peptides on the activation and thedifferentiation of human osteoblast cells.
Specific aims were as follows:
1) To identify the expression of PDE families in human osteoblasts.2) To investigate the effects of PDE inhibition on osteoblast differentiation and function
with RNA interference (RNAi) and pharmacological inhibitors.3) To analyse the expression and regulation of different PDEs during osteoblast
differentiation at the mRNA and enzyme levels.4) To examine the effects of bioactive peptides on gene expression in the osteoblast.

40
4. Materials and methods
4.1 Cell culture
4.1.1 Osteosarcoma cell lines (I, III, IV)
Human and rat osteosarcoma cell lines, widely used as models for osteoblasts, were used instudies I, III and IV. The human osteosarcoma MG63 (III), which is considered to represent amature osteoblastic phenotype was the primarily used cell line, whereas SaOS2 cells (I, III)were mainly used to confirm results obtained with MG63 cells (III) or NHOst (I). Both celllines were purchased from the American Type Culture Collection. MG63 cells were culturedin αMEM supplemented with 10% foetal calf serum and SaOS2 cells in McCoy`s 5A,supplemented with 12.5% foetal calf serum, with antibiotics (50 IU/ml penicillin and 50µg/ml streptomycin) in a humidified atmosphere with 5% CO2 at 37°C. UMR106 (IV) ratosteosarcoma cells (American Type Culture Collection) were grown in Dulbecco’s modifiedEagle’s medium (DMEM), supplemented with 10% foetal calf serum (FCS), 50 IU penicillinand 50 g/ml streptomycin at 37°C in a humified atmosphere with 5% CO2 on 96–well plates.The cells were seeded at 1 x 104 cells/cm².
4.1.2 Human osteoblasts (I)
Normal human osteoblasts (NHOst) and all the cell culture reagents for the NHOst cells werepurchased from Cambrex, BioWhittaker. Two strains of NHOst cells isolated from bone chipsfrom female donors were used. The cells were seeded at a density of 5000 cells/cm2 andcultured in osteoblast growth medium supplemented with 10% FCS, 50 µg/ml ascorbic acid, 5mM ßglycerophosphate, 200 nM hydrocortisone21hemisuccinate and 0.1%gentamin/amphotericinB solution, at 37ºC in a 5% CO2/95% air atmosphere on plastic Petridishes.
4.1.3 Human mesenchymal stem cells (hMSC)derived osteoblasts (II, IV)
Human bone marrowderived mesenchymal stem cells (hMSC, Cambrex) were cultured inmesenchymal stem cell growth medium with mesenchymal cell growth supplements(Cambrex) at 37°C in a humified atmosphere with 5% CO2 on 100mm plastic dishes at aseeding density of 3100 cells/cm2. The differentiation into osteoblasts was performedaccording to the instructions of the supplier after the cultures became 50% confluent. Thedifferentiation medium consisted of MEM supplemented with 10% foetal bovine serum,108 M dexamethasone (Dex), 50 g/ml Lascorbic acid, 10 mM –glycerophosphate, 100U/ml penicillin and 100 U/ml streptomycin and was changed every 34 days.

41
4.2 PDE inhibitors
The PDE chemical inhibitors used in the studies are listed in Table 3.
TABLE 3. PDE inhibitors used in Studies IIII.
Family Inhibitor Study
PDE1 8MMX (8methoxymethyl3isobutyl1methylxanthine) I, III
PDE2 EHNA I, III
PDE3 Cilostamide (NcyclohexylNmethyl4[1,2dihydro2oxo6quinolyloxy] butyramide)
III
PDE3 Milrinone (6methyl2oxo5pyridin4yl1Hpyridine3carbonitrile)
I
PDE4 Rolipram (4[3{cyclopentyloxy}4methoxyphenyl]2pyrrolidinone)
I, II, III
PDE7 BRL 50481 (3(N,Ndimethylsulfonamido)4methylnitrobenzene)
II
PDE3 cGMP I, III
nonspecific PDEinhibition (PDE17,1011)
IBMX I, III
4.3 Primers (I, II, III, IV)
The primers were used as presented in Table 4.
TABLE 4. Primers used in Studies IIV. Forward primer (F), Reverse Primer (R).cDNA Sequence StudyPDE1A F: AAAATGGGGATGACAAAAAAGAA
R: GATATTCATTTCTTCTTCTTGCATI, III
PDE1B F: ATGCAGGATGATGAGATGAAR: CGGAAGAATTCCTCCAT
I, III
PDE1C F: AGACAGGGTGACAGAGAAGCR: TGGCCTTCTCCTCTTTGGGT
I, III
PDE3A F: TCACCTCTCCAAGGGACTCCTR: CAGCATGTAAAACATCAGTGGC
I, III
42
PDE4A F: CTCGCACAAGTTCAAAAGGATGR: GCCTCCAGCGTAATCCGACA
I, III
PDE4B F: GACATTGCAACAGAAGACAAG TCCCCR: ACTCAAGTAACTGAAGGCCAGGTGG
II
PDE4B F: AGCTCATGACCCAGATAAGTGR: ATAACCATCTTCCTGAGTGTC
I, III
PDE4C F: ACCTCAGCGCCAAGCAGCR: TGGGTGGGAAAGTGAAGCAGG
I
PDE4C F: ACACTGAACTCCTGTCCCCTGAAGR: GATGTGACTCAAGAGTGACCACTGG
III
PDE4D F: CCCTGGACTGTTATCATGCAR: TGATTGGACACACCAGGATG
I, III
PDE7A F: GGACGTGGGAATTAAGCAAGCR: TCCTCACTGCTCGACTGTTCT
I, II, III
PDE7B F: TGCACAGGACAGGCACTTTAR: CTTCTGTGCTGCCTGGGCAA
I, II, III
PDE8A F: GCC TGT TTC CTG GACAAA CR: GCA TTA CGG ACA ACT CTT CTC
II
PDE8A F: CAGCCAGAGACGACACTCTTCCATR: ATGATCTTAGCGTTGACTCGGAGC
III
PDE10A F: GTGTGGTGCAGATGGTCAACR: GGGTGAATTGCATGAGACCT
III
PDE11A F: ATCTGCCTCAGTATCCCCCTR: CCACCAGGAAAAGAGAGCAG
III
GAPDH F: AAGGCTGGGGCTCATTTGR: CCACCAGGAAAAGAGAGCAG
III
Betaactin F: CATCGTCACCAACTGGGACGACR: CGTGGCCATCTCTTGCTCGAAG
I
Betaactin F: AGGCCAACCGCGAGAAGATGACCR: GAAGTCCAGGGCGACGTAGCAC
II, IV
Osteocalcin F: ATGAGAGCCCTCACACTCCTCGR: GTCAGCCAACTGGTCACAGTCC
II, IV
Caspase8 F: AGGAGGAGATGGAAAGGGAACTTR: ACCTCAATTCTGATCTGCTCACTTCT
II, IV
CREB5 F: GCTTTGGTGCTTTTCTCCAGR: GGTGACACCACAGCACAAAC
II, IV
Betacatenin F: TTCTGGTGCCACTACCACAGCR: TGCATGCCCTCATCTAATGTC
II
Vitamin Dreceptor
F: CCAGTTCGTGTGAATGATGGR: CCTTTTGGATGCTGTAACTG
II
BMP1 F: AGGCCCACTTCTTCTCAGAAAAGR: GAAGAGAGCACCAGTTCTGTCCG
II
BMP2 F: GGAGAAGGAGGAGGCAAGR: GACACGTCCATTGAAAGAGC
IV
BMP5 F: AAGAAGACAAGAAGGACTAAAAATATR: GTAGAGATCCAGCATAAAGAGAGGT
II, IV
PTHrp F. GTCTCAGCCGCCGCCTCAAR: GGAAGAATCGTCGCCGTAAA
IV
T7promotor+ PDE7
F: TAATACGACTCACTATAGGTTCAAGAAGATTACR: TAATACGACTCACTATAGGTTTCACTCCACT
II

43
T7promotor + PDE8A
F: TAATACGACTCACTATAGGAAGAAGGTAGCAGR: TAATACGACTCACTATAGGTTCATTGTACGAG
II
4.4 cAMP analyses (I, II, III)
Measurement of cAMP accumulation was determined with a luminometric cAMP assay(cAMPGlo TM Assay, Promega) two days after transfections with PDE7 siRNAs (II) or usingthe method of Frandsen and Krishna (1976), as described earlier (Ahlström et al. 2000).Briefly, the cells were grown in 24well plates without treatment (I) or treated with eithervehicle (ethanol) or Dex (1100 nM) for 48 h (III). The cells were then induced with forskolinfor 7 min at 37°C and rinsed twice with icecold phosphatebuffered saline (PBS) (III) orwashed with Hanks Balanced Salt Solution (HBSS) (I). For determination of basal cAMPlevels, cells were incubated at 37°C for 30 min before washing with icecold PBS (III) orHBSS (I). cAMP was extracted from the cells with 96% ethanol at 18°C for 3 h. The extractwas evaporated with N2 gas and dissolved in assay buffer (0.05 M sodium acetate, pH 6.2). Ifnecessary, the sample was further diluted with assay buffer, and the cAMP concentration wasdetermined by radioimmunoassay with iodinated cAMP as tracer.
4.5 Qsepharose chromatography (I, III)
Five confluent cultures of either NHOst, SaOS2 or MG63 cells, grown on 100mm plasticdishes were washed twice with PBS buffer, harvested with a cell scraper into 6 ml of icecoldhomogenization buffer (buffer A), containing 20 mM Bistris, pH 6.5, 5 mMmercaptoethanol, 0.1% of a protease inhibitor cocktail (PIC), consisting of 0.5 mg/ml each ofbestain, pepstatin A, aprotinin, leupeptin, E64 and 12.5 mg/ml AEBSF. The harvested cellswere then homogenized on ice by ten passages with a Teflon/glass homogenizer. Thehomogenization was repeated three times. The homogenate was then centrifuged for 20 minat 20 000 g. Five millilit of the supernatant was diluted with 20 ml of buffer B, containing 20mM Bistris, pH 6.5, 0.1 M sodium acetate, 0.02% PIC (v/v), 0.1 mM EDTA, 1 mMbenzamidine and 1 mM mercaptoethanol, filtered through a 0.22µm syringe filter andapplied to a column (5ml bed volume) of Qsepharose High Performance (AmershamPharmacia Biotech) previously equilibrated with buffer B. After washing the column with 10bed volumes, the PDE activities were eluted with a 0.11.0 M linear sodium acetate gradientin buffer B, at a flow rate of 2.5 ml/min. Fractions of 2 ml were collected into tubescontaining 50 µl of 5% bovine serum albumin (BSA) and assayed for either cAMPPDE orcGMPPDE activity, as described below.

44
4.6 Enzyme activity analyses
4.6.1 Assay of PDE activity (I, II, III)
PDE activity was assayed according to the method described by Thompson and Appleman(1971). Samples from either Qsepharose elutions or wholecell homogenates were added tothe incubation buffer (40 mM TrisHCl, pH 8.0, 0.1% PIC, 0.05% BSA, 5 mM MgCl2, 1 mMEGTA, 0.5 µM cAMP and 3H –cAMP (100000 cpm/tube) to give a final reaction volume of
200 µl. For the cGMPPDE assay, cAMP was replaced with 0.25 µM cGMP and 3H –cGMP.The samples were incubated at 34°C for 3060 min, followed by boiling for 1 min. Aftercooling on ice, 50100 µl of Crotalus atrox snake venom nucleotidase (1 mg/ml, Sigma) wasadded and incubated at 34°C for 10 min. Then 500 µl of a 1:3 slurry of AGIX8 resin(BioRad) was added, and the tubes were mixed and centrifuged for 5 min at 8000 rpm. Theradioactivity of the supernatant was determined with a βcounter. Selective inhibitors used fordifferent PDE types are summarized in Table 3.
4.6.2 Assay of ALP activity (II)
Bonespecific alkaline phosphatase (bALP) activity was measured with a immunoenzymetricassay (OCTEIATM Ostase® BAP, Immunodiagnostic Systems). Prior to measurements, thecells were washed with PBS and after addition of 300 l of Tris –Triton (0.1 M Tris–base,0.2% Triton X1000), the cells were frozen at 70 C. After thawing, the cells were centrifuged(12000 g, 5 min). The supernatant was used for analysis. The ALP activity was corrected withtotal protein content.
4.7 PDE silencing (II)
PDE silencing was performed with siRNAs, and the PDE7 selective inhibitor BRL 50481(Table 3) was mainly used to confirm results obtain with PDE7 siRNAs. The short doublestranded RNA (dsRNA) mixtures for PDE7 and PDE8 inhibition were generated from PDE7and PDE8 PCR products. Primers with appended T7 promoters sequenced for the PDE7 PCRproducts were designed to amplify both PDE7A and PDE7B. PDE8 PCR products weredesigned to amplify only PDE8A. The BLAST database program (NBCI) was used to confirmthe specificity of the target sequences. The large dsRNAs were first synthesized by in vitrotranscription from the DNA templates. The large (1501500 bp) dsRNAs were processed byShortCut RNase III digestion in the presence of manganese, which produced a heterogeneouspopulation of short (1825 bp) siRNAs. For generation of siRNAs, we used a ShortCut RNAikit (New England BioLabs). After 1214 days of differentiation, the osteoblast phenotype was

45
confirmed by staining cultures and control cultures with a histological alkaline phosphatasekit (Sigma). At this stage, the medium was changed to a low serum (2.5%), antibioticfreemedium, and 24 h later, the siRNA mixtures were transfected into the cells by Lipofectamin2000 (Invitrogen). Control cells were simultaneously treated with lipofectamin withoutsiRNAs. The transfection time varied between 6 and 24 h, and the cell culture was thenchanged to a new differentiation medium. The transfection efficiency was determined withBLOCKiT™ Fluorescent Oligos (Invitrogen) according to the manufacturer`s instructions.
4.8 Isoleucineprolineproline (IPP), valineprolineproline (VPP) and leucinelysineproline(LKP) tripeptides (IV)
The peptides were synthesized by 9fluorenylmethyloxycarbonyl (Fmoc) chemistry andpurified using HPLC reverse phase columns. Peptides were synthesized in the Core Facilityfor Synthetic Peptides, Division of Biochemistry, Department of Biological andEnvironmental Sciences, University of Helsinki, Finland. Peptides were dissolved in sterilewater prior to administration.
4.9 Cell proliferation analyses (II, IV)
The proliferation rate was assayed with a luminometric ATP detection assay (ATPlite, PerkinElmer) 48 h after the siRNA transfections (II), with multilabel counter (Victor 3, PerkinElmer) or measured the activity for DNA synthesis (IV) with a Cell Proliferation ELISA kit(Roche Diagnostics). The 5bromo2’deoxyuridine (BrdU) binds to the newly synthesizedcellular DNA. BrdU incorporation was measured with a spectroscopic plate reader at 450 nm(Multiskan Ex, Thermo Labsystems).
4.10 Mineralization analyses (II)
Mineralization was assessed by Alizarin Red S staining in cells grown on 96well platesusing the method described by Stanford et al. (1995). After staining, the dye was eluated with10% cetylpyridinium chloride in 10 mM sodium phosphate (pH 7.0, 15 min, RT). Absorbancewas measured with a spectroscopic plate reader at 590 nm.

46
4.11 mRNA expression analyses
4.11.1 RNA isolation (IIV)
RNA of the osteoblastic cells was extracted using the QuickPrep Micro mRNA purificationkit (Amersham Pharmacia Biotech) or the RNeasy Protect mini kit (Qiagen GmbH). TheRNA concentration was measured at 260 nm with a spectrophotometer. The integrity of theRNA was confirmed by 1.2% formaldehyde agarose gel electrophoresis.
4.11.2 cDNA microarrays (II, IV)
The cDNA microarray hybridizations were performed at the Finnish DNA Microarray Centreof the Turku Centre for Biotechnology. The expression profiles of osteoblasts differentiatedfrom hMSC were compared after a 48h treatment with 35 nM PDE7 or PDE8A siRNAmixtures (III), or after a 24 h treatment with 50 M IPP, VPP or LKP (IV), with Hum16Kprotocol. The array contains 16 000 human gene probes (accession number: AMEXP557,http://www.ebi.ac.uk/ arrayexpress/). Samples were labelled with FluoroLink Cy3dUTP andCy5dUTP (Amersham Biosciences) using 20 g of total RNA for direct labelling andhybridized using an augmented reference design (Kerr and Churchill 2001). Discrete imagesfor Cy3 and Cy5 dyes were obtained using a Scan Express laserscanning microscope(Packard BioSciences) and gene transcriptlevels were determined from the fluorescenceintensities of scanned data image files with the QuantArray Microarray Analysis software(Packard BioSciences). Ttest and geneexpression profiles were analysed by KensingtonDiscovery Edition 2.0 software. At least a 1.6 fold difference in expression was used incontrol/treatment comparison. The cDNA microarray experiment is available at accessionnumber EMEXP1021 (II) and EMEXP885 (IV) (http://www.ebi.ac.uk/arrayexpress/). Thegene expression data for IPP treated osteoblasts were analysed by principal componentanalysis (PCA) to determine the main components affected. Results were consideredsignificant at a 95% significance level (p < 0.05) (IV).
4.11.3 cDNA synthesis (I, II, III, IV)
cDNA was prepared from 12 µg of total RNA or mRNA by reverse transcription with thefollowing buffer and conditions: RT buffer (Promega) containing 50 mM TrisHCl (pH 8.3)75 mM KCl, 3 mM MgCl2 and 10 mM DTT, 13 U RNAguard, Rnase inhibitor (AmershamPharmacia Biotech), 0.9 mM each of dATP, dCTP, dGTP and dTTP, 15 pmol Oligo(dT) 15primer (Promega) and 100 U MMLVreverse transcriptase primer (Promega) in a finalvolume of 25 µl, with incubation for 1 h at 37°C.

47
4.11.4 Semiquantitative reverse transcriptase polymerase chain reaction (RTPCR) (I, III)
PCR amplifications were conducted in 30µl aliquots containing 24 µl of cDNA, 1x buffer(Finnzymes), comprising 10 mM TrisHCl, pH 8.8 at 25°C, 1.5 mM MgCl2, 50 mM KCl and
0.1% Triton X100, 0.2 mM each of dATP, dCTP, dGTP and dTTP, 0.35 µM of each primerand 1 U of DyNAzyme DNA polymerase (Finnzymes). Primers are listed in Table 4. Theamplification protocol was run on a PTC100 thermocycler (MJ Research). After denaturationat 94°C for 4 min, the reactions underwent cycles of denaturing for 1 min at 95°C, annealingfor 90 s at 5863°C and extension for 2 min at 72°C. This was followed by a single step at72°C for 10 min. To ensure that the PCR was in the exponential phase, the conditions of thesemiquantitative RTPCR were optimised for the number of PCR cycles and the amount oftotal RNA applied to the reactions for the PDE subtypes/isoforms and for glyceraldehyde3phosphate dehydrogenase (GAPDH) or actin, which was used as an external standard.Human brain RNA and a cocktail of RNA from rat spleen, brain, kidney and heart was usedto prepare cDNAs used as positive controls. Negative controls included samples in whichcDNA synthesis was performed in the absence of reverse transcriptase. Reactions in which nocDNA was added were also performed to check for possible nonspecific products. Volumesof 10 µl of the PCRamplified products were analysed on 1.5% agarose gels containingethidium bromide. Gel results were quantified by AlphaDigiDoc™ System 1000 (AlphaInnotech Corporation). The results were analysed as a ratio of integrated optical density of thePDE subtypes/isoforms to the external standard.
4.11.5 Quantitative Realtime PCR (qRTPCR) (II, IV)
PCR amplification was performed in a realtime quantitative PCR engine Mx3000P(Stratagene). The PCR reactions contained cDNA templates and Brilliant SYBR Green QPCRMaster mix kit (Stratagene) in a 25µl reaction volume containing 200 nM of each genespecific primer (Table 4). Fluorescence data were collected during the annealing step andanalysed with Mx3000P software. Amplification was obtained by denaturing at 95°C for 10min, followed by 40 cycles of denaturing at 95°C for 30 s, annealing at 58°C for 1 min, andextension at 72°C for 30 s. The amplification cycles were followed by for 1 min at 95°C andfor 30 s at 55°C, and data for dissociation plots were collected as the temperature wasreturned to 95°C. cDNA from human brain total RNA (BD Biosciences) was used as acalibration standard, and the data were normalized with amplification data of ßactin. Allreactions were run in triplicate with three dilution concentrations, and the mean value wasused to calculate the ratio of target gene/ßactin expression in each sample. Using the ratio inthe untreated sample as a standard (100), the relative ratio of treated sample was presented asthe relative expression level of the target gene.
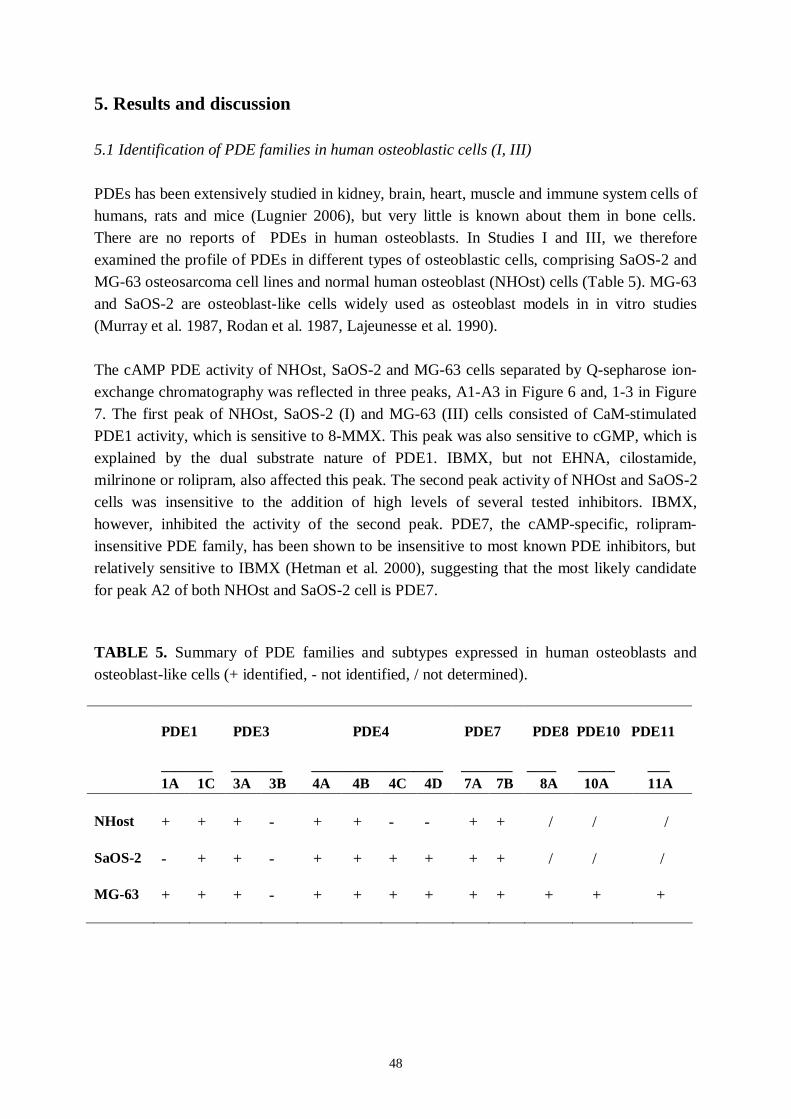
48
5. Results and discussion
5.1 Identification of PDE families in human osteoblastic cells (I, III)
PDEs has been extensively studied in kidney, brain, heart, muscle and immune system cells ofhumans, rats and mice (Lugnier 2006), but very little is known about them in bone cells.There are no reports of PDEs in human osteoblasts. In Studies I and III, we thereforeexamined the profile of PDEs in different types of osteoblastic cells, comprising SaOS2 andMG63 osteosarcoma cell lines and normal human osteoblast (NHOst) cells (Table 5). MG63and SaOS2 are osteoblastlike cells widely used as osteoblast models in in vitro studies(Murray et al. 1987, Rodan et al. 1987, Lajeunesse et al. 1990).
The cAMP PDE activity of NHOst, SaOS2 and MG63 cells separated by Qsepharose ionexchange chromatography was reflected in three peaks, A1A3 in Figure 6 and, 13 in Figure7. The first peak of NHOst, SaOS2 (I) and MG63 (III) cells consisted of CaMstimulatedPDE1 activity, which is sensitive to 8MMX. This peak was also sensitive to cGMP, which isexplained by the dual substrate nature of PDE1. IBMX, but not EHNA, cilostamide,milrinone or rolipram, also affected this peak. The second peak activity of NHOst and SaOS2cells was insensitive to the addition of high levels of several tested inhibitors. IBMX,however, inhibited the activity of the second peak. PDE7, the cAMPspecific, rolipraminsensitive PDE family, has been shown to be insensitive to most known PDE inhibitors, butrelatively sensitive to IBMX (Hetman et al. 2000), suggesting that the most likely candidatefor peak A2 of both NHOst and SaOS2 cell is PDE7.
TABLE 5. Summary of PDE families and subtypes expressed in human osteoblasts andosteoblastlike cells (+ identified, not identified, / not determined).
PDE1 PDE3 PDE4 PDE7 PDE8 PDE10 PDE11
_______ _______ __________________ _______ ____ _____ ___1A 1C 3A 3B 4A 4B 4C 4D 7A 7B 8A 10A 11A
NHost + + + + + + + / / /
SaOS2 + + + + + + + + / / /
MG63 + + + + + + + + + + + +
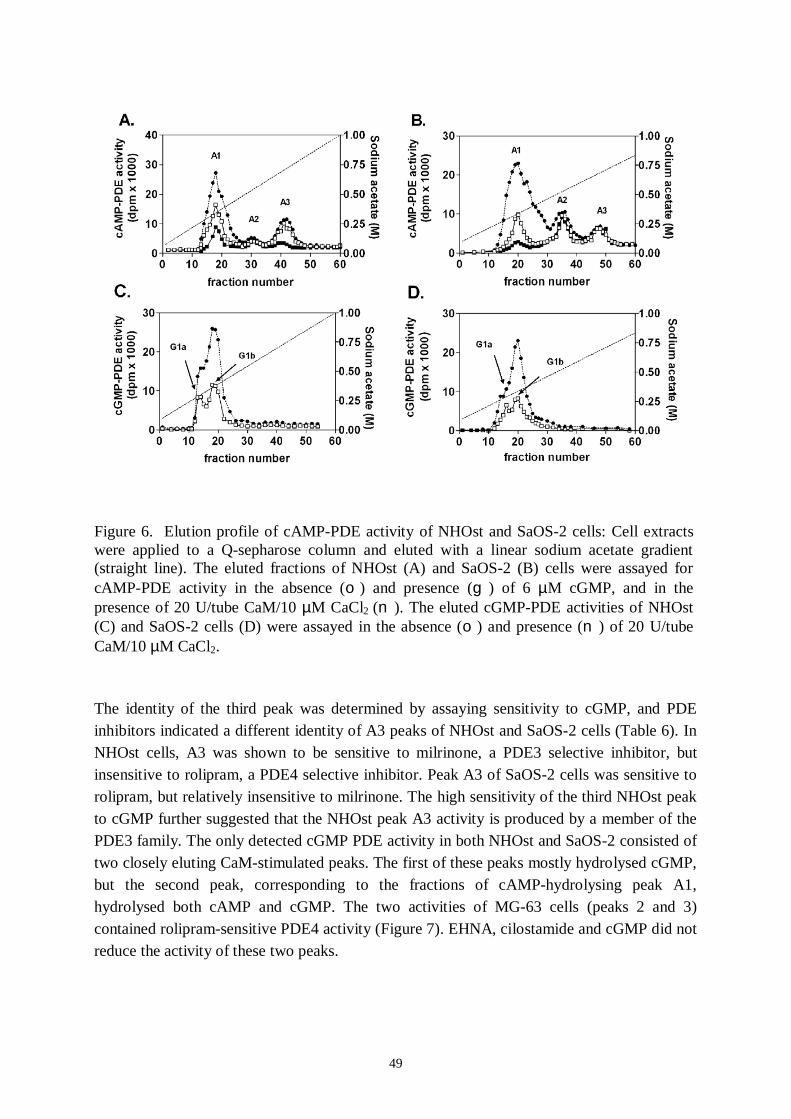
49
Figure 6. Elution profile of cAMPPDE activity of NHOst and SaOS2 cells: Cell extractswere applied to a Qsepharose column and eluted with a linear sodium acetate gradient(straight line). The eluted fractions of NHOst (A) and SaOS2 (B) cells were assayed forcAMPPDE activity in the absence (o) and presence (g) of 6 µM cGMP, and in thepresence of 20 U/tube CaM/10 µM CaCl2 (n). The eluted cGMPPDE activities of NHOst(C) and SaOS2 cells (D) were assayed in the absence (o) and presence (n) of 20 U/tubeCaM/10 µM CaCl2.
The identity of the third peak was determined by assaying sensitivity to cGMP, and PDEinhibitors indicated a different identity of A3 peaks of NHOst and SaOS2 cells (Table 6). InNHOst cells, A3 was shown to be sensitive to milrinone, a PDE3 selective inhibitor, butinsensitive to rolipram, a PDE4 selective inhibitor. Peak A3 of SaOS2 cells was sensitive torolipram, but relatively insensitive to milrinone. The high sensitivity of the third NHOst peakto cGMP further suggested that the NHOst peak A3 activity is produced by a member of thePDE3 family. The only detected cGMP PDE activity in both NHOst and SaOS2 consisted oftwo closely eluting CaMstimulated peaks. The first of these peaks mostly hydrolysed cGMP,but the second peak, corresponding to the fractions of cAMPhydrolysing peak A1,hydrolysed both cAMP and cGMP. The two activities of MG63 cells (peaks 2 and 3)contained rolipramsensitive PDE4 activity (Figure 7). EHNA, cilostamide and cGMP did notreduce the activity of these two peaks.
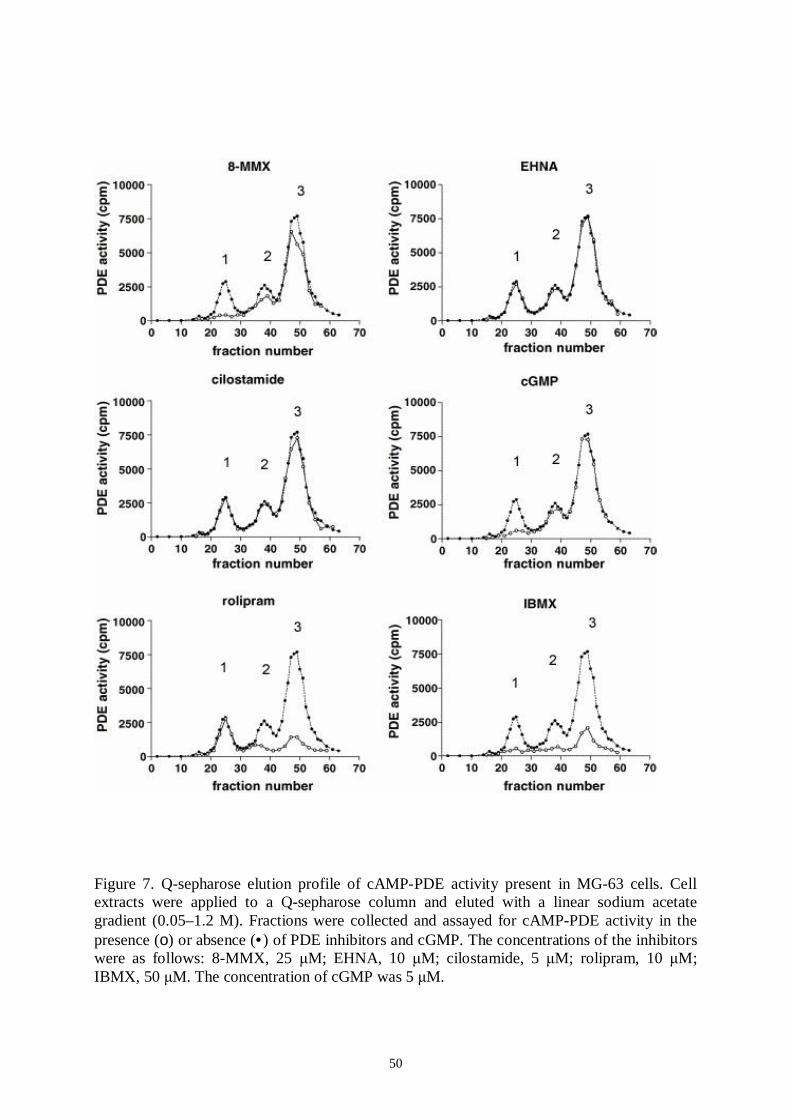
50
Figure 7. Qsepharose elution profile of cAMPPDE activity present in MG63 cells. Cellextracts were applied to a Qsepharose column and eluted with a linear sodium acetategradient (0.05–1.2 M). Fractions were collected and assayed for cAMPPDE activity in thepresence (ο) or absence (•) of PDE inhibitors and cGMP. The concentrations of the inhibitorswere as follows: 8MMX, 25 M; EHNA, 10 M; cilostamide, 5 M; rolipram, 10 M;IBMX, 50 M. The concentration of cGMP was 5 M.
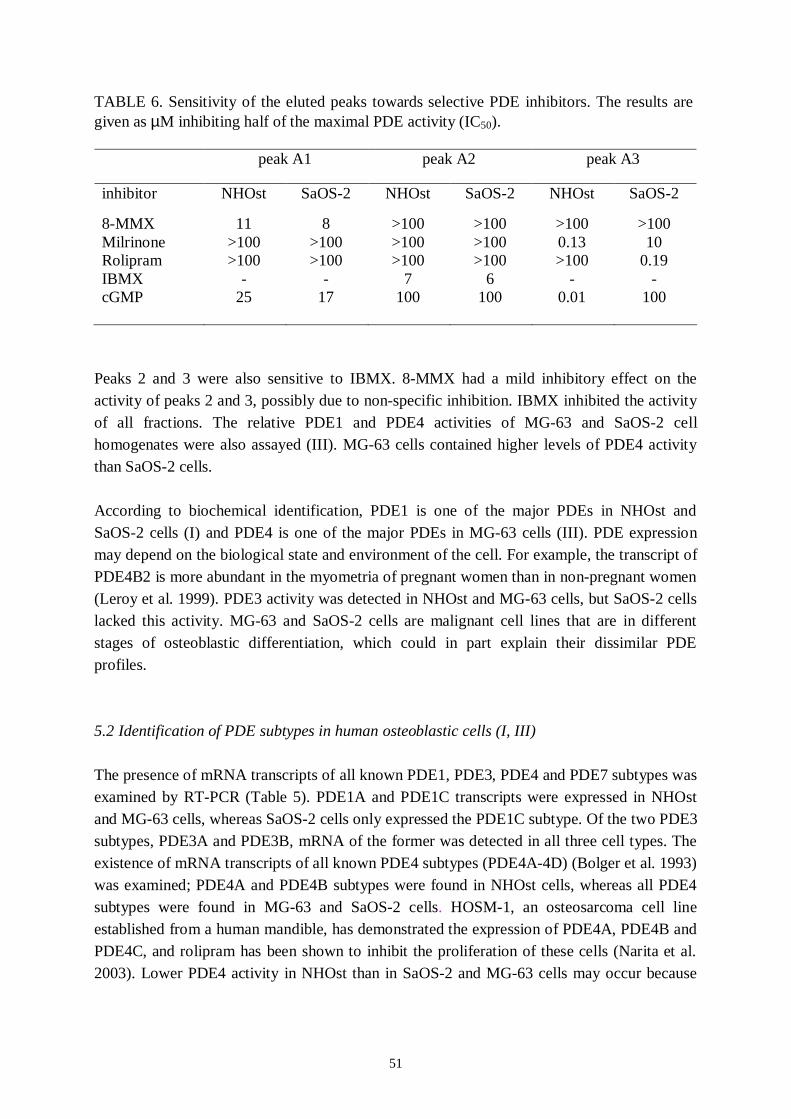
51
TABLE 6. Sensitivity of the eluted peaks towards selective PDE inhibitors. The results aregiven as µM inhibiting half of the maximal PDE activity (IC50).
peak A1 peak A2 peak A3
inhibitor NHOst SaOS2 NHOst SaOS2 NHOst SaOS2
8MMX 11 8 >100 >100 >100 >100Milrinone >100 >100 >100 >100 0.13 10Rolipram >100 >100 >100 >100 >100 0.19IBMX 7 6 cGMP 25 17 100 100 0.01 100
Peaks 2 and 3 were also sensitive to IBMX. 8MMX had a mild inhibitory effect on theactivity of peaks 2 and 3, possibly due to nonspecific inhibition. IBMX inhibited the activityof all fractions. The relative PDE1 and PDE4 activities of MG63 and SaOS2 cellhomogenates were also assayed (III). MG63 cells contained higher levels of PDE4 activitythan SaOS2 cells.
According to biochemical identification, PDE1 is one of the major PDEs in NHOst andSaOS2 cells (I) and PDE4 is one of the major PDEs in MG63 cells (III). PDE expressionmay depend on the biological state and environment of the cell. For example, the transcript ofPDE4B2 is more abundant in the myometria of pregnant women than in nonpregnant women(Leroy et al. 1999). PDE3 activity was detected in NHOst and MG63 cells, but SaOS2 cellslacked this activity. MG63 and SaOS2 cells are malignant cell lines that are in differentstages of osteoblastic differentiation, which could in part explain their dissimilar PDEprofiles.
5.2 Identification of PDE subtypes in human osteoblastic cells (I, III)
The presence of mRNA transcripts of all known PDE1, PDE3, PDE4 and PDE7 subtypes wasexamined by RTPCR (Table 5). PDE1A and PDE1C transcripts were expressed in NHOstand MG63 cells, whereas SaOS2 cells only expressed the PDE1C subtype. Of the two PDE3subtypes, PDE3A and PDE3B, mRNA of the former was detected in all three cell types. Theexistence of mRNA transcripts of all known PDE4 subtypes (PDE4A4D) (Bolger et al. 1993)was examined; PDE4A and PDE4B subtypes were found in NHOst cells, whereas all PDE4subtypes were found in MG63 and SaOS2 cells. HOSM1, an osteosarcoma cell lineestablished from a human mandible, has demonstrated the expression of PDE4A, PDE4B andPDE4C, and rolipram has been shown to inhibit the proliferation of these cells (Narita et al.2003). Lower PDE4 activity in NHOst than in SaOS2 and MG63 cells may occur because

52
cells expressed only PDE4A and PDE4B subtypes. The expression of PDE4C and PDE4Dcould be involved in the proliferation and the malignant phenotype of osteosarcoma cells.
The rolipraminsensitive/IBMXsensitive cAMPspecific PDE activity detected in NHOst,SaOS2 and MG63 cells indicated the presence of PDE7. PDE7 has two subtypes inmammals; both mRNA transcripts (PDE7A and PDE7B) were detected in all studied cells. Inaddition, the presence of mRNA transcripts of the remaining cAMP PDEs (PDE8A, PDE10Aand PDE11A) was identified in MG63 cells.
Profiles of PDE subtypes in different osteoblastic cells have previously been studied in ratUMR106 osteoblastlike cells (Ahlström and LambergAllardt 1999, 2000), mouseosteoblastic cell line MC3T3E1 and bone marrow stromal cell line ST2 (Wakabayashi et al.2002). In mouse cell lines, PDE1, PDE2, PDE3, PDE4, PDE7, PDE8 and PDE9 mRNAtranscripts have been identified. In rat UMR106 osteoblastlike cells, the main PDE fractionsare PDE1 and PDE4, but PDE2, PDE3, PDE7 and PDE8 expression has also been detected.These findings are line with our results from human osteoblasts and osteosarcoma cells MG63 and SaOS2. PDE1, PDE2, PDE3, PDE4, PDE7 and PDE8 may be the major PDEs inosteoblasts, but profiles and subtypes could vary during the differentiation and withenvironmental changes.
5.3 PDE7 and PDE8A silencing by siRNAs (II)
In in vivo and in vitro studies, the inhibition of PDE4 has been demonstrated to increaseosteoblastic differentiation and bone formation in rats and mice (Miyamoto et al. 1997, 2003,Kinoshita et al. 2000, Wakabayashi et al. 2002, Scutt et al. 2004), but the effect of PDE7inhibition on bone biology has not previously been investigated. At the beginning of ourstudy, no chemical inhibitors for PDE7 or PDE8 were available. siRNAs have provided thefield of molecular biology with a new and efficient tool to efficiently knockdown specificgenes (Mousses et al. 2003, Bantounas et al. 2004, Semizarov et al. 2004). We investigatedthe effect of specific silencing of phosphodiesterase subtypes PDE7A, PDE7B and PDE8Aon gene expression in hMSCderived osteoblasts. For the silencing of specific PDEs, siRNAsgenerated from PDE7 and PDE8 were utilized. Quantitative realtime PCR was used toconfirm the specific PDE silencing. PDE7 and PDE8 inhibition by RNAi significantlydecreased the gene expression of PDE7A, PDE7B and PDE8A. At the level of mRNA, PDE7and PDE8 inhibition by RNAi decreased the gene expression of PDE7A by 6070%, PDE7Bby 4050% and PDE8A by 30%.
Since the discovery of the RNAi method various genes involved in the cell cycle, apoptosis,differentiation and signalling have successfully been inhibited in different mammalian celltypes, in vitro and in vivo, to elucidate gene function, identify drug targets and treat a varietyof diseases. βcatenin expression has been silencing, leading to the inhibition of growth of

53
colon cancer cells (Verma et al. 2003). The reduction of cyclin B1 levels in HeLa cells bysiRNAs caused inhibition of proliferation by arresting cells in the G2 phase and inducingapoptosis (Yuan et al. 2006). RNAi targeting TGFβ1 significantly ameliorated theprogression of matrix expansion in experimental glomerulonephritis (Takabatake et al. 2005).In addition, the gene expression of PDE4B and PDE4D has been reduced with syntheticsiRNAs in human embryonic kidney (HEK293B2) cells (Lynch et al. 2005). Silencingefficiency of siRNAs is dependent on the sequence of siRNAs, the number of siRNAmolecules and the cell doubling rate. In slowly proliferating or nondividing cells, siRNAsseem to have a longer gene silencing effect than in rapidly dividing cells (Pai et al. 2006). Forexample, siRNA activity lasts for 37 days in proliferating cells, yet persists for three weeksor more in terminally differentiated cells such as neurons (Omi et al. 2004). We used hMSCderived osteoblasts in our study, when cells differentiated the rate of proliferation has becomelimited (Amedee et al. 1994). Our results show that specific gene silencing with the RNAimethod is useful for inhibiting the gene expression of PDE7 and PDE8 subtypes.
5.4 PDE7 silencing has potential for enhancing osteogenic gene expression anddifferentiation in hMSCcells derived osteoblasts (II)
Gene expression profiling was performed using a Hum16K cDNA microarray. The arraycontains 16 000 human genes probes and is designed for genomewide screening. The arraytherefore contains a limited number of known key factors for osteoblastogenesis and boneformation; Runx 2, Rankl and Lrp5 genes are missing. Treatment for 48 h with PDE7 siRNAsupregulated 435 and downregulated 60 loci ( 1.6fold difference). The respective numbersfor siRNA treatment of PDE8A were 38 and 96. Selected microarray results are displayed inTable 7. Quantitative realtime PCR results confirmed up/downregulation of a set of genesfrom the microarray results (Table 8). The qRTPCR results were in line with the microarrayanalysis. PDE7 silencing upregulated the gene encoding OCN, a noncollagenous bonespecific matrix protein secreted by normal maturing osteoblasts (Price et al. 1976). OCNexpression is highly upregulated by 1,25(OH)2D3 in MG63 osteosarcoma cells (Mahonen etal. 1990, Lajeunesse et al. 1991). In addition, the activation of the cAMP/PKA signallingpathway increases OCN mRNA and the secretion of OCN in rat osteoblastlike cells (Noda etal. 1988, Theofan et al. 1989). In human osteosarcoma cells, the results of cAMP/PKAactivation on OCN expression are conflicting; OCN expression has been reported to be eitherup or downregulated by compounds that elevate cAMP (Lajeunesse et al. 1991, Drissi et al.1997, Boguslawski et al. 2000).

54
TABLE 7. Effects of PDE7 and PDE8A RNAi silencing on gene expression in hMSCderived humanosteoblasts. A set of interesting genes were selected from cDNA microarray results._________________________________________________________________________________________
Accession number Gene/protein Foldchange___________________________________________________________________________Inhibition of PDE7
UpregulatedEnzymes and metabolismH87471 Kynureninase (Lkynurenine hydrolase) 2.1DifferentiationAA779457 Bone morphogenetic protein 5 3.5
Catenin betalike 1 2.0Calciumsensing receptor 2.0Glutamate receptor metabotropic 3 3.1Tumour necrosis factor (ligand)superfamily member 10 3.2
ApoptosisN94588 CASP8 and FADDlike apoptosis regulator 2.1
DownregulatedHormone and growth factor receptors
Vitamin D (1,25 dihydroxyvitamin D3 receptor) 1.7Fibroblast growth factor receptor 5.4
ApoptosisT95823 cyclin L2 1.8
Inhibition of PDE8A
UpregulatedN55459 Metallothionein 1F (functional) 1.6R01682 Metalloproteaserelated protein 1 1.6DownregulatedApoptosisN94588 CASP8 and FADDlike apoptosis regulator 1.9DifferentiationAA045327 Osteoglycin (osteoinductive factor, mimecan) 1.5R56774 Bone morphogenetic protein 1 1.5___________________________________________________________________________Foldchange describes the relative expression of the given factor in osteoblast cells treated for48 h with 35 nM PDE7 or PDE8A siRNAs.

55
TABLE 8. Summary of qPCR results. Effects of PDE7 and PDE8A RNAi silencing on geneexpression in hMSCderived human osteoblasts.Gene PDE7 silencing PDE8A silencing
OCN ↑ ↔βcatenin ↑ ↔Caspase 8 ↑ ↓CREB5 ↑ ↔VDR ↓ ↔BMP1 ↔ ↓BMP5 ↑ ↔PDE7A ↓ ↔PDE7B ↓ ↔PDE8A ↔ ↓PDE4A ↔ ↔PDE4B ↔ ↔Ostoglycin ↔ ↓
The effects of gene expression are indicated as follows: upregulation ↑, downregulation ↓,no effect ↔.
The BMPs are known to be important in the process of osteoblast differentiation,proliferation, apoptosis and morphogenesis (Canalis et al. 2003). The BMP family can bedivided into several subgroups based on BMP gene sequence identity. BMP1 is unrelated tothe other BMPs and does not regulate the growth and differentiation of skeletal cells (Uzel etal. 2001). BMP5, BMP6, BMP7 and BMP8 belong to the same subgroup and all haveosteoinductive properties (Wozney 2002). In our study, PDE7 inhibition caused a threefoldupregulation of BMP5 gene expression. The BMPs have been shown to interact closely withWnt peptides. The Wnt proteins are secreted by osteoblasts and are required for BMPmediated stimulation of osteoblast proliferation and function (Nakashima et al. 2004, Nelsonand Nusse 2004). Inhibition of PDE7 upregulated catenin, a protein that plays a central rolein the Wnt signalling pathway (Rawadi and RomanRoman 2005), suggesting that PDE7could participate in Wntmediated effects on osteoblast differentiation. In parallel withdifferentiation, osteoblasts show increased apoptosis, which is established to be associatedwith the activation of caspases (Canalis et al. 2005). Evidence indicates that caspasemediatedactivation of apoptosis is necessary for osteoblastic differentiation (Mogi and Togari 2003).The effect of PDE7 silencing was to significantly upregulate caspase8. In contrast tocaspase8, vitamin D receptor (VDR) mRNA was reduced in PDE7silenced cells. VDRknockout mice show increased bone volume and density compared with wildtype mice(Tanaka and Seino 2004), and osteoblasts lacking VDR have enhanced differentiation,manifested as an increase in both ALP activity and mineralized matrix formation (Sooy et al.2005). The increased expression of caspase8 and the decreased expression of VDR seen inPDE7silenced cells therefore support PDE7 inhibition promoting an osteoblastic geneexpression pattern. The transcription factors of the CREB family stimulate basal transcription

56
of CREcontaining genes and mediate induction of transcription upon phosphorylation byprotein kinase A. CREB has been demonstrated to have a major role in the expression ofskeletonspecific genes (Sakamoto et al. 1998). In our study, silencing of PDE7 upregulatedCREB5, which further suggests that PDE7 is involved in gene expression during osteoblasticdifferentiation.
In human T cells, PDE7 has the potential to regulate cell function, including cytokineproduction, proliferation and expression of activation markers (Nakata et al. 2002). PDE8Asilencing was also performed, resulting in decreased BMP1, osteoglycin and collagen 1 geneexpression. As the expression of these genes is known to be associated with the osteoblasticphenotype, PDE8A inhibition seems unlikely to enhance the differentiation of osteoblasts. Infact, PDE8 silencing appears to have a negative effect on osteoblastic differentiation. Themicroarray findings of PDE7 silencing illustrate upregulation of several osteogenic geneexpressions, which could reflect augmented osteoblastic differentiation.
5.5 PDE7 inhibition enhances cAMP signalling, differentiation and mineralization in hMSCderived osteoblasts (II)
The effects of PDE7 silencing on cAMP accumulation were studied to evaluate the role ofPDE7 on cAMP signalling. The cells were cultured in differentiation media for two weeks.PDE7 was silenced with siRNAs for 48 h, and cAMP synthesis was then stimulated with 10µM forskolin for 20 min. PDE7 silencing increased the forskolinstimulated cAMP responsemore than threefold. As the PDE7 silencing increased both cAMP accumulation andosteocalcin expression, we investigated the possible involvement of PKA in PDE7 siRNAstimulated osteocalcin expression. Treatment with 5 µM PKA inhibitor H89 after siRNAtransfections completely abolished the effects of PDE7 siRNAs on OCN expression.
To evaluate bALP activity in hMSCderived osteoblasts, the transfections were performedafter 13 days of differentiation and bALP activity was measured two weeks aftertransfections. The treatment with PDE7 siRNAs significantly increased the bALP levelscompared with control cells. Silencing PDE8 did not significantly change bALP. Inundifferentiated hMSC cells, bALP levels were low (Figure 8A). Mineralization was assessedby Alizarin red S staining. Cells were treated with siRNAs or with 20 µM of the PDE7selective inhibitor BRL 50481 (3(N,Ndimethylsulfonamido)4methylnitrobenzene). After2122 days of differentiation, the mineralization of PDE7 siRNAtreated cells was increasedthreefold compared with controls (Figure 8B). Treatment with PDE8A siRNAs had no effecton mineralization (Figure 8C). Treatment with 20 µM BRL50481 also increasedmineralization (Figure 8D).
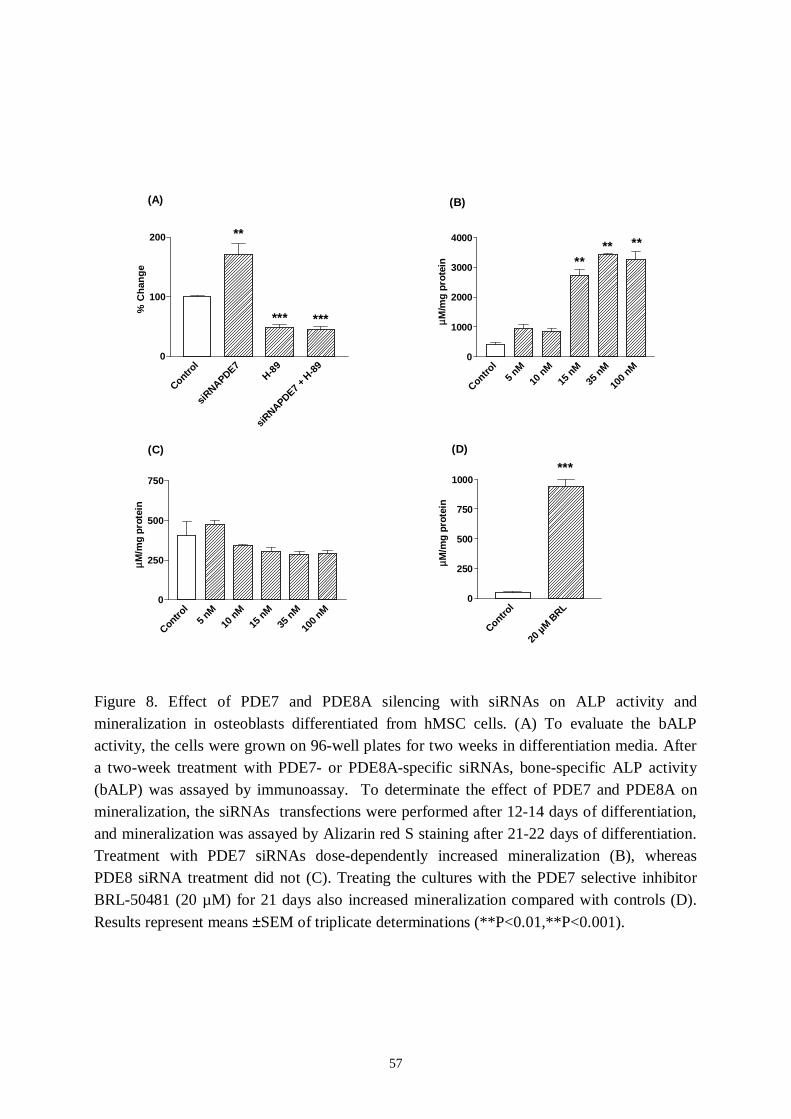
57
Control
siRNAPDE7
H89
siRNAPDE7 +
H89
0
100
200 **
*** *** % C
hang
e
Control
5 nM
10 nM
15 nM
35 nM
100 n
M0
1000
2000
3000
4000
**** **
µM
/mg
prot
ein
Control
5 nM
10 nM
15 nM
35 nM
100 n
M0
250
500
750
µM
/mg
prot
ein
Control
20 µM B
RL0
250
500
750
1000***
µM
/mg
prot
ein
(A) (B)
(C) (D)
Figure 8. Effect of PDE7 and PDE8A silencing with siRNAs on ALP activity andmineralization in osteoblasts differentiated from hMSC cells. (A) To evaluate the bALPactivity, the cells were grown on 96well plates for two weeks in differentiation media. Aftera twoweek treatment with PDE7 or PDE8Aspecific siRNAs, bonespecific ALP activity(bALP) was assayed by immunoassay. To determinate the effect of PDE7 and PDE8A onmineralization, the siRNAs transfections were performed after 1214 days of differentiation,and mineralization was assayed by Alizarin red S staining after 2122 days of differentiation.Treatment with PDE7 siRNAs dosedependently increased mineralization (B), whereasPDE8 siRNA treatment did not (C). Treating the cultures with the PDE7 selective inhibitorBRL50481 (20 µM) for 21 days also increased mineralization compared with controls (D).Results represent means ±SEM of triplicate determinations (**P<0.01,**P<0.001).

58
Silencing of PDE7 increased both bALP activity and mineralization in hMSCderivedosteoblasts, implying that PDE7 also has an impact on osteoblast cell functions. Our studyshows that inhibition by PDE7specific siRNAs and by the chemical PDE7 inhibitor BRL50481 can control osteoblastic differentiation and mineralization.
5.6 Downregulation of cAMPPDE by dexamethasone in human osteosarcoma cells (III)
The effect of glucocorticoids on the function of PDEs was studied at the mRNA and enzymelevels in vitro. Dex, a synthetic glucocorticoid, has been shown to increase cell osteogenicdifferentiation and proliferation in bone marrow mesenchymal stem cell (Bellows et al. 1987,Shalhoub et al. 1992, Jaiswal et al. 1997, Bellows et al. 1998), but impair the proliferative andmetabolic activity of osteoblasts (Patschan et al. 2001). Treatment of MG63 cells with Dexsignificantly decreased the cAMPPDE activity in a dose and timedependent manner. Theeffect of Dex on cAMPPDE activity was reduced by up to 50%, with the halfmaximalinhibition of the cAMPPDE activity being 5 nM. Treatment with 100 nm Dex for 48 h didnot affect the cGMPPDE activity in MG63 cells, indicating that Dex specifically affectscAMPPDE activity. The effect of Dex on the cAMPPDE activity of SaOS2 cells was lesspronounced, probably due to the lower PDE4 activity in these cells. The sensitivity ofhomogenates towards PDE inhibitors EHNA and cilostamide was not affected by Dextreatment. However, homogenates of the Dextreated cells were, as expected, less sensitive torolipram. The Dex treatment also resulted in homogenates that were slightly less sensitive to8MMX. Stimulation with forskolin or rolipram separately only increased the cAMP levels inthe cells moderately. Simultaneous treatment with rolipram and forskolin resulted in 20foldand 60fold increases in cAMP levels in MG63 and SaOS2 cells, respectively. Treatmentwith Dex did not increase cAMP levels in either cell type. However, the Dex treatmentsignificantly reduced the potency of rolipram to increase forskolinstimulated cAMPresponse. We detected a substantial decrease in the cAMPspecific PDE activity aftertreatment with Dex. The Dex treatment alone did not affect the intracellular cAMP levels, buttaking into account that PDE4 is abundant PDE type in both MG63 and SaOS2 cells, thereduction of the cAMP response in the Dextreated cell might better describe the effect of analtered PDE profile of the cells. A previous study with SaOS2 cells has presented similarresults, showing that Dex alone does not affect cAMP levels (Mori et al. 1999).
5.7 Influence of dexamethasone on PDE subtype mRNA expression in MG63 and SaOS2cells (III)
Expression of the following cAMPPDE subtypes was detected by reverse transcriptase PCR:PDE1A, PDE1C, PDE2A, PDE3A, PDE4A, PDE4B, PDE4C, PDE4D, PDE7A, PDE7B,PDE8A, PDE10A and PDE11A (Table 5). Of these subtypes, Dex reduced the mRNA ofPDE4A and PDE4B subtypes in MG63 and SaOS2 cells. The decrease in PDE4 mRNA was

59
dose and timedependent, with the halfmaximal inhibition being 5–10 nM Dex, withsignificant effects seen within 4 h of treatment. Further analysis revealed a reduction in themRNA of the PDE4A4 and PDE4B1 isoforms. Transcription of PDE4A1, 4A7, 4A10, 4B2and 4B3 was also evident, but we could not detect any reduction with Dex treatment in theseisoforms. The observed effects on cAMPPDE activity and cAMP accumulation wereaccompanied by a decrease in the mRNA levels of PDE4A4 and PDE4B1 isoforms. Thesepresented results thereby support the hypothesis that PDEs play a role in Dexmediatedeffects on cAMP signalling in osteoblasts. In rat cultured osteoblasts, two glucocorticoidactions have been shown: the stimulation of adenylate cyclase and the inhibition of PDE(Chen and Feldman 1979). In addition, treatment with Dex increases PTHR mRNAexpression in ROS 17/2.8 cells, but not in opossum kidney cells. Glucocorticoids mightenhance sensitivity to PTH by changing the number of PTH receptors and their affinity toPTH; this suggests the involvement of a posttranscriptional mechanism (Urena et al. 1994).
5.8 Effects of PDEs in osteoblasts
In summary, Studies IIII revealed that the profiles of PDEs in human osteoblasts are similarto these in rat and mouse osteoblasts. The main PDEs are PDE1, PDE2, PDE3, PDE4, PDE7and PDE8, but families and subtypes can vary depending on differentiation stage andcircumstances around the cells. In Study II, we investigated the effects of silencing of PDE7and PDE8 on gene expression and differentiation in hMSCderived osteoblasts. Inhibition ofPDE7 enhances differentiation and mineralization. The inhibitors of PDE4 have earlier beenshown to increase osteoblastic differentiation and bone formation. At the sequence level, thePDE7 family is closely related to the PDE4 family (Omori et al. 2007). This could be onereason why effects of PDE inhibition are osteogenic in hMSCderived osteoblasts. Asynthetic glucocorticoid, Dex, could alter the profile of PDE4 subtypes in osteoblastic cells.Dex did not have a direct effect on intracellular cAMP levels, but did cause a reduction incAMP response. Our results suggest an additional mechanism by which glucocorticoids couldbe involved in bone loss.
Only a small number of studies exist on the effects of PDEs in bone or bone cells, and most ofthese have investigated the effect of PDE4 inhibitors. In rats inoculated with Walker 256/Scarcinoma to induce an artificial osteoporotic state, bone loss was reduced by over 50% aftera twoweek treatment with the PDE4 selective inhibitor denbufylline (Miyamoto et al. 1997).PDE4 inhibitors rolipram, XT44 and XT611 have been shown to increase mineralizednodule formation (Waki et al. 1999) and PGE2mediated anabolic effects (Miyamoto et al.2003) in rat bone marrow culture. XT44 also increases the bone mineral density ofovariectomized rats. Rolipram and pentoxifylline, a nonselective inhibitor, have beendemonstrated to increase both cortical and cancellous bone mass in mice (Kinoshita et al.2000). Daily injections of pentoxifylline have been reported to enhance bone formation inBMPimpregnated collagen disks implanted into the back muscles of mice (Horiuchi et al.

60
2001). Pentoxifylline also promotes the osteoblastic differentiation of ST2 mouse bonemarrowderived stromal cells, although a recent report suggests that the effects ofpentoxifylline are protein kinase Aindependent (Rawadi et al. 2001). More information aboutthe role of other PDEs in bone formation and how these enzymes are involved in theregulation of bone metabolism is needed.
Future studies should clarify the profile of PDEs in various differentiation stages, from stemcells to osteoblasts, osteocytes and osteoclast. The role of PDE7 in bone metabolism could beinvestigated in vivo with, for instance, knockout mice. Tcell functional studies haverevealed that PDE7A knockout mice have approximately 10% lower body fat than PDE7A /+ and PDE7A / mice (Yang et al. 2003). The lower body fat in PDE7A knockout micemight also indicate changes in bone tissue.
5.9 Influences of bioactive peptides IPP, VPP and LKP on gene expression and proliferationin hMSCderived osteoblasts (IV)
The Hum16K cDNA microarray data analysis revealed that the bioactive peptide IPPregulated more genes in hMSCderived osteoblasts than the bioactive peptides VPP and LKP.Treatments with 20 µM tripeptides showed that IPP upregulated 270 genes and downregulated 100 genes. The respective numbers for VPP were 25 and 10, and for LKP 16 and14. IPP not only regulated a larger number of genes than VPP or LKP, but also was the onlytripeptide to upregulate osteogenic differentiation factors.
The PCA included 245/270 upregulated genes and 47/100 downregulated genes in thedifferent components. The PCA revealed 45% of the variation in upregulated genes from theIPPtreated osteoblasts to be attributed to the first principal component (upregulated celldifferentiation). The second principal component explained 25.3% (upregulated cell growth)and the third 22.6% (upregulated cell transcription) of the variation (Table 9). The firstcomponent (115 genes) had heavy positive accumulations of genes related to differentiation,receptors, mitochondria, apoptosis, enzymes, signal transduction, membrane architecture,membrane function and membrane transport. The second component (47 genes) had heavypositive accumulations of genes related to cell adhesion, osteogenic differentiation and cellgrowth and proliferation, while and third component (83 genes) had heavy positiveassociations with genes related to transcription, splicing, translation, EST and ‘unknownfunctions’. The upregulation of these genes indicates that IPP enhances osteoblastproliferation and differentiation.
Our results also show that IPP increases UMR106 and hMSC proliferation, but notproliferation of mature osteoblasts. When the state of differentiation increases, the
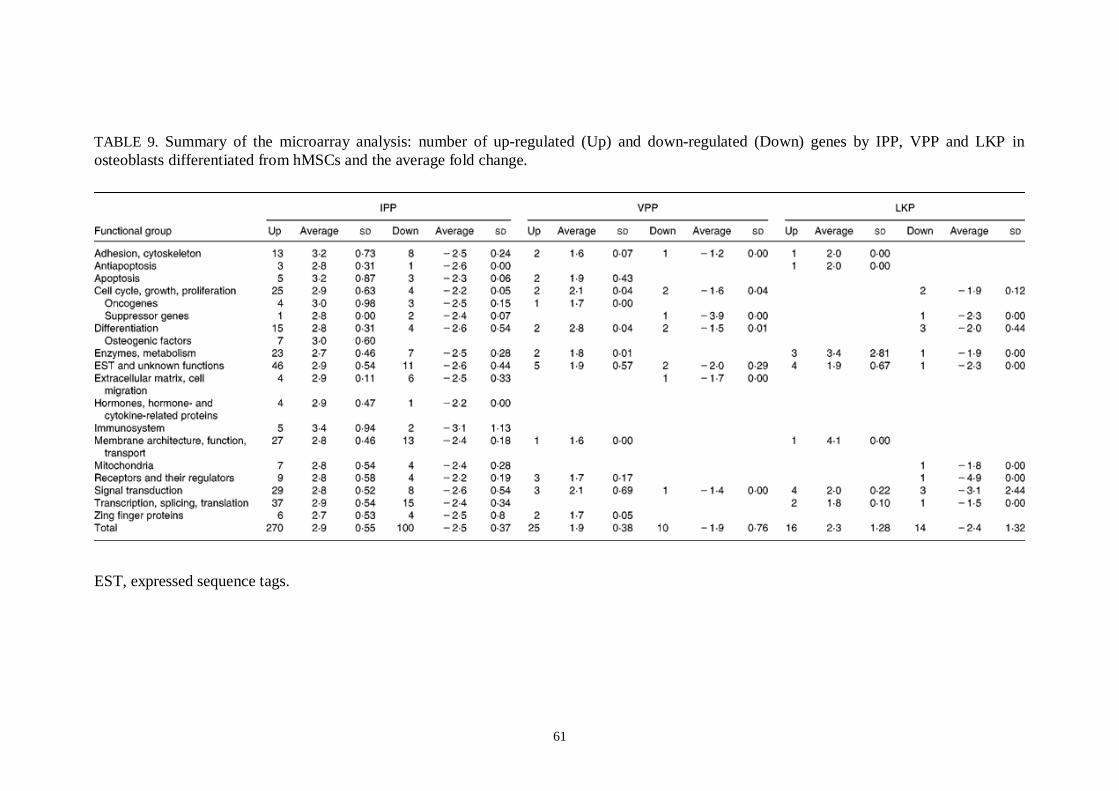
61
TABLE 9. Summary of the microarray analysis: number of upregulated (Up) and downregulated (Down) genes by IPP, VPP and LKP inosteoblasts differentiated from hMSCs and the average fold change.
EST, expressed sequence tags.
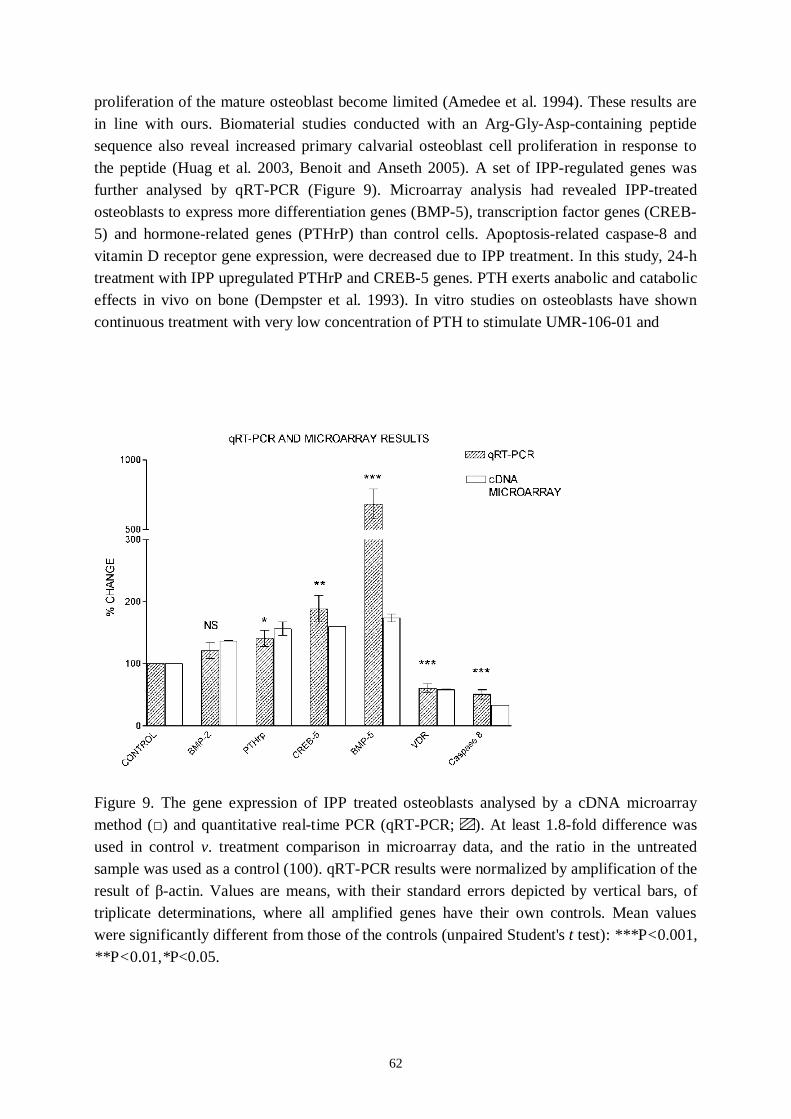
62
proliferation of the mature osteoblast become limited (Amedee et al. 1994). These results arein line with ours. Biomaterial studies conducted with an ArgGlyAspcontaining peptidesequence also reveal increased primary calvarial osteoblast cell proliferation in response tothe peptide (Huag et al. 2003, Benoit and Anseth 2005). A set of IPPregulated genes wasfurther analysed by qRTPCR (Figure 9). Microarray analysis had revealed IPPtreatedosteoblasts to express more differentiation genes (BMP5), transcription factor genes (CREB5) and hormonerelated genes (PTHrP) than control cells. Apoptosisrelated caspase8 andvitamin D receptor gene expression, were decreased due to IPP treatment. In this study, 24htreatment with IPP upregulated PTHrP and CREB5 genes. PTH exerts anabolic and cataboliceffects in vivo on bone (Dempster et al. 1993). In vitro studies on osteoblasts have showncontinuous treatment with very low concentration of PTH to stimulate UMR10601 and
Figure 9. The gene expression of IPP treated osteoblasts analysed by a cDNA microarraymethod ( ) and quantitative realtime PCR (qRTPCR; ). At least 1.8fold difference wasused in control v. treatment comparison in microarray data, and the ratio in the untreatedsample was used as a control (100). qRTPCR results were normalized by amplification of theresult of actin. Values are means, with their standard errors depicted by vertical bars, oftriplicate determinations, where all amplified genes have their own controls. Mean valueswere significantly different from those of the controls (unpaired Student's t test): ***P<0.001,**P<0.01,*P<0.05.

63
primary osteoblast proliferation (Dempster et al. 1993, Swarthout et al. 2001). PTHrp is apolypeptide hormone sharing a common sequence with PTH (Walsh et al. 1997), and both canfunction with the PTHR (Walsh et al. 1997). PTHrp produced by osteoblasts is thought tofunction locally. Within the skeletal microenvironment, PTHrp propels pluripotential bonemarrow stromal cells towards the osteogenic lineage and also exerts antiapoptotic effects(Karaplis and Goltzman 2000). The ability of PTH to drive changes in gene expression isdependent upon activation of such transcription factors as the CREB (Swarthout et al. 2002).In response to PTH, CREB is phosphorylated and in turn activates transcription. Theincreased proliferation in IPPtreated hMSC could be due to increased PTHrp and CREB.Because cell proliferation and maturation are sequential processes, increased numbers ofpluripotent cells and their augmented maturation into osteoblasts could increase the totalnumber of osteoblasts. The ability of IPP to induce BMP5 production can increase osteoblastdifferentiation and augment fracture repair, a feature associated with a proteinrich diet(Bastow et al. 1983, Delmi et al. 1990, Rizzoli and Bonjour 2004).
In summary, IPP, one of the three investigated tripeptides, increase the gene expression ofosteogenic factors in hMSCderived osteoblasts. IPP has also shown to increase matrixproduction, mineralization and decrease RANKL/OPG ration in the same cells (Huttunen etal. 2007). Tripeptides could transport via hPepT1 transporter both in the small intestine and inthe proximal tubule or hPepT2 in the proximal tubule (see Nielsen and Brodin 2003,Steffansen et al. 2004), but the transporter and signaling mechanisms in bone cells is stillunclear. Forthcoming studies could focus on the the receptor interactions and cellularmechanisms of bioactive tripeptides in bone cells.
6. Conclusions
We identified PDE families and their subtypes present in normal human osteoblasts and inhuman osteosarcoma (SaOS2, MG63) cell lines. The main difference in PDE profiles wasthe obvious lack of PDE3 activity in osteosarcoma cells, while PDE3 contributed largely tothe total PDE activity of normal human osteoblasts. The major PDE subtype of MG63 cellswas PDE4, and MG63 cells also contained higher levels of PDE4 activity than SaOS2 cells.The reason for this difference in PDE profiles is unknown. The osteosarcoma cell lines mightrepresent a different differentiation stage that is associated with high levels of PDE4expression, or PDE profiles might have altered during cell transformation from the normal tothe malignant phenotype.
Our results showed that inhibition by PDE7specific siRNAs and by the chemical PDE7inhibitor BRL50481 can control osteoblastic differentiation and mineralization. The findingsalso indicated that genespecific silencing with the RNAi method is useful for inhibiting thegene expression of specific PDEs. The silencing of PDE7 upregulates several osteogenic

64
genes at the mRNA level, and PDE7 may therefore play an important role in the regulation ofosteoblastic differentiation.
At the mRNA and enzyme levels, Dex, a synthetic glucocorticoid, decreased gene expressionof PDE4 subtypes 4A and 4B, but did not affect other cAMPPDEs in MG63 and SaOS2cells. The observed effects on cAMPPDE activity and cAMP accumulation wereaccompanied by a decrease in the mRNA levels of PDE4A4 and PDE4B1 isoforms. Theresults support the hypothesis that PDEs play a role in Dexmediated effects in cAMPsignalling in osteoblasts. PDEs appears to be involved in a mechanism by whichglucocorticoids affect cAMP signalling, demonstrating a new possible route in the formationof glucocorticoidinduced bone loss involving the downregulation of cAMPPDE.
Based on our results, tripeptide IPP increases cell proliferation at the cellular and mRNAlevels. Genes associated with differentiation were upregulated and apoptosis inducers weredownregulated. These findings indicate that IPP enhances gene expression in such a way thatincreases the differentiation of hMSC into osteoblasts and prolongs their viability.
This work has shown that PDEs possess mechanisms that enhance osteoblast differentiationand affect cAMP signalling by glucocorticoids. Bioactive peptide IPP augmented the potentialto upregulate osteogenic and cell viability genes in osteoblasts. Bioactive peptides, such asIPP, might well contribute to the positive effects that dietary protein has on bone mineralmass. Understanding the role of PDEs in bone cells is important since bone metabolism,especially bone formation, could be affected by PDEs. These enzymes are potentiallysignificant targets for the development of new signal transduction therapies for the treatmentof metabolic bone diseases, for example osteoporosis. However, additional research is neededto elucidate the mechanisms of PDEs and bioactive peptides on osteoblast differentiation andbone formation.

65
7. Acknowledgements
This work was carried out at the Calcium Research Unit, Division of Nutrition, Department ofApplied Chemistry and Microbiology, University of Helsinki. I am grateful to ProfessorsAntti Ahlström and Marja Mutanen for providing excellent working facilities.
I also wish to thank Professor Tapio Palva, Head of the Division of Genetics, Department ofEnvironmental and Biosciences, for interest in my work and for guiding me through thebureaucracy.
My warmest gratitude is due to my supervisors, Dr. Christel LambergAllardt for theopportunity to work with her group and Dr. Mikael Ahlström for introducing me to the worldof bone cells and phosphodiesterases. Both (of you) gave me a great deal of scientificfreedom, guidance when needed and were always ready to listen to my ideas.
I am grateful to the reviewers, Dr. Petri Lehenkari and Dr. Anitta Mahonen, their advice,constractive criticism and insightful comments that substantially improved this thesis.
I am grateful to my authoreditor, Carol Ann Pelli, for revising the English language of themanuscript.
I thank present and past colleagues at the Calcium Research Unit; Heini Karp, MerjaKärkkäinen, Virpi Kemi, Marika Laaksonen, Zahirul Islam, Heli Viljakainen and particularlyMinna Huttunen for stimulating teamwork, help and enthusiasm, and Ulrike Riehle for hercontribution and for being such a nice person. It has been a pleasure to share the ups anddowns of academic life with all of you.
My heartfelt thanks are due to my friends; providing for relaxing company and all kinds offun over the years. I greatly appreciate the unwavering support of my mother Kerttu, mybrother Harri, my motherinlaw Elise and her husband Juhani. Finally, my deepest love andgratitude are owed to my husband Pasi and dauhgter Iisa. I am so lucky to have you in mylife.
This work has been financially supported by the Finnish Graduate School in MusculoSkeletalDiseases, the National Technology Agency of Finland (TEKES), the Oskar ÖflundFoundation, the Finnish Konkordia Fund, the Finnish Cultural Foundation, the Ella ja GeorgEhrnrooth Foundation and the Betty Väänänen Foundation.
Sipoo, March 2008
Minna Pekkinen

66
8. References
Abe, E., Yamamoto, M., Taguchi, Y., LeckaCzernik, B., O’Brien, C., Economides, A.N.,Stahl, N., Jilka, R.L., Manolagas, S.C. (2000) Essential requirement of BMPs2/4 for bothosteoblast and osteoclast formation in murine bone marrow cultures from adult mice:antagonism by noggin. J Bone Miner Res 15:663673.Advani, S., LaFrancis, D., Bogdanovic, E., Taxel, P., RaisZ, L.G.; Kream, B.E. (1997)Dexamethasone suppresses in vivo levels of bone collagen synthesis in neonatal mice. Bone20, 4146.Ahlström, M., LambergAllardt, C. (1997) Rapid protein kinase Amediated activation ofcyclic AMPphosphodiesterase by parathyroid hormone in UMR106 osteoblastlike cells. JBone Miner Res 12, 172178.Ahlström, M., LambergAllardt, C. (1999) Regulation of Adenosine 3`,5´cyclicmonophosphate (cAMP) accumulation in UMR106 osteoblastlike cells: Role of cAMPphoshodiesterase and cAMP efflux. Bio Pharmacol 58, 13351340.Ahlström, M., LambergAllardt, C. (2000) Inactivation of atrial natriuretic factorstimulated cyclic guanosine 3`,5´monophosphate (cGMP) in UMR106 osteoblastlike cells.Bio Pharmacol 59, 11331139.Amedee, J., Bareille, R., Rouais, F., Cunningham, N., Reddi, H., Harmand, M.F. (1994)Osteogenin (bone morphogenic protein 3) inhibits proliferation and stimulates differentiationof osteoprogenitors in human bone marrow. Differentiation 58, 157164.Anthony, T.L., Brooks, H.L., Boassa, D., Leonov, S., Yanochko, G.M., Regan, J.W.,Yool, A.J. (2000) Cloned human aquaporin1 is a cyclic GMPgated ion channel. MolPharmacol 57, 57688.Aravind, L., Koonin, E.V. (1998) The HD domain defines a new superfamily of metaldependent phosphohydrolases. Trends Biochem Sci 23, 469472.Aubin, J.E. and Liu, F. (1996) The osteoblast lineage. In: Bilezikian, J.P., Raisz, L.G. andRodan, G.A., Editors, 1996. Principles of Bone Biology, Academic Press, New York, pp. 51–67.Banerjee, C., McCabe, L.R., Choi, JY., Hiebert, S.W., Stein, J.L., Stein, G.S., Lian, J.B.(1997) Runt homology domain proteins in osteoblast differentiation: AML3/CBFA1 is amajor component of a bone specific complex. J Cell Biochem 66, 18.Bantounas, I., Phylactou, L.A., Uney, J.B. (2004) RNA interference and the use of smallinterfering RNA to study gene function in mammalian systems. J Mol Endocrinol 33, 545457.Bastow, M.D., Rawlings, J., Allison, S.P. (1983) Benefits of supplementary tube feedingafter fractured neck of femur: a randomised controlled trial. Br Med J Clin Res 287, 15891592.Beavo, J.A. (1995) Cyclic nucleotide phosphodiesterases: Functional implications of multipleisoforms. Physil Rev 75, 725748.Beazely, M.A., Watts, V.J. (2006) Regulatory properties of adenylate cyclases type 5 and 6:A progress report. European Journal of Pharmacology 535, 112.

67
Beard, M.B., Olsen, A.E., Jones, R.E., Erdogan, S., Houslay, M.D., Bolger, G.B. (2000)UCR1 and UCR2 domains unique to the cAMPspecific phosphodiesterase family form adiscrete module via electrostatic interactions. J Biol Chem 275, 1034958.Bellows, C.G., Aubin, J.E., Heersche, J.N. (1987) Physiological concentrations ofglucocorticoids stimulate formation of bone nodules from isolated rat calvaria cells in vitro.Endocrinology, 121, 19851992.Bellows, C.G., Ciaccia, A., Heersche, J.N. (1998) Osteoprogenitor cells in cell populationsderived from mouse and rat calvaria differ in their response to corticosterone, cortisol, andcortisone. Bone 23, 119125.Bender, A.T., Ostenson, C.L., Wang, E.H., Beavo, J.A. (2005) Selective upregulation ofPDE1B2 upon monocytetomacrophage differentiation. Proc Natl Acad Sci USA 102, 497502.Benoit, D.S., Anseth, K.S. (2005) The effect on osteoblast function of colocalized RGD andPHSRN epitopes on PEG surfaces. Biomaterials 26, 52095220.Benovic, J.L., Strasser, R.H., Caron, M.G, Lefkowitz, R.J. (1986) Betaadrenergicreceptor kinase: identification of a novel protein kinase that phosphorylates the agonistoccupied form of the receptor. Proc Natl Acad Sci U S A 83, 2797801.Bian, J.S., Zhang, W.M., Pei, J.M., Wong, T.M. (2000) The role of phosphodiesterase inmediating the effect of protein kinase C on cyclic AMP accumulation upon kappaopioidreceptor stimulation in the rat heart. J Pharmacol Exp Ther 292, 10651070.Birnbaumer, L. (1990) G proteins in signal transduction. Ann Rev Pharmacol Toxicol 30,675705.Bloom, T.J., Beavo, J.A. (1996) Identification and tissuespecific expression of PDE7phosphodiesterase splice variants. Proc Natl Acad Sci U S A 93, 1418892.Boden, S.D., Hair, G., Titus, L., Racine, M., McCuaig, K., Wozney, J.M., Nanes, M.S.(1997) Glucocorticoidinduced differentiation of fetal rat calvarial osteoblasts is mediated bybone morphogenetic protein6. Endocrinology 138:28202828.Boerth, N.J., Dey, N.B., Cornwell, T.L., Lincoln, T.M. (1997) Cyclic GMPdependentprotein kinase regulates vascular smooth muscle cell phenotype. J Vasc Res 34, 24559.Bolger, G., Michaeli, T., Martins, T., St John, T., Steiner, B., Rodgers, L., Riggs, M.,Wigler, M., Ferguson, K. (1993) A family of human phosphodiesterases homologous to thedunce learning and memory gene product of Drosophila melanogaster are potential targets forantidepressant drugs. Mol Cell Biol 13, 655871.Bolger, G.B., Erdogan, S., Jones, R.E., Loughney, K., Scotland, G., Hoffmann, R.,Wilkinson, I., Farrell, C., Houslay, M.D. (1997) Characterization of five different proteinsproduced by alternatively spliced mRNAs from the human cAMPspecific phosphodiesterasePDE4D gene. Biochem J 328, 53948.Borton, A.J., Frederick, J.P., Datto, M.B., Wang, X.F., Weinstein, R.S. (2001) The loss ofSmad3 results in a lower rate of bone formation and osteopenia through dysregulation ofosteoblast differentiation and apoptosis. J Bone Mineral Res 16, 175464.

68
Boyce, B.F., Xing, L., Jilka, R.J., Bellido, T., Weinstein, R.S., Parfitt, A.M., Manolagas,S.C. (2002) Apoptosis in bone cells. In: Bilezikian, J.P., Raisz, L.G., Rodan, G.A. (eds.)Principles of Bone Biology, 2nd ed. Academic Press, San Diego, CA, USA, pp 151168.Boguslawski, G., Hale, L.V., Yu, XP., Miles, R.R., Onyia, J.E., Santerre, R.F.,Chandrasekhar, S. (2000) Activation of Osteocalcin Transcription Involves Interaction ofProtein Kinase A and Protein Kinase Cdependent Pathways. J Biol Chem 275, 9991006.Butcher, R.W., Sutherland, E.W. (1962) Adenosine 3',5'phosphate in biological materials.I. Purification and properties of cyclic 3',5'nucleotide phosphodiesterase and use of thisenzyme to characterize adenosine 3',5'phosphate in human urine. J Biol Chem 237, 124450.Calvi, L.M., Sims, N.A., Hunzelman, J.L., Knight, M.C., Giovannetti, A., Saxton, J.M.,Kronenberg, H.M., Baron, R., Schipani, E. (2001) Activated parathyroidhormone/parathyroid hormonerelated protein receptor in osteoblastic cells differentiallyaffects cortical and trabecular bone. J Clin Invest 107, 27786.Canalis, E. (2005) Mechanisms of glucocorticoid action in bone. Curr Osteopor Rep 3, 98102.Canalis, E., Bilezikian, J.P., Angeli, A., Giustina, A. (2004) Perspectives on glucocorticoidinduced osteoporosis. Bone 34, 593598.Canalis, E., Deregowski, V., Pereira, R.C., Gazzerro, E. (2005) Signals that determine thefate of osteoblastic cells. J Endocrinol Invest 28, 37.Canalis, E., Economides, A.N., Gazzerro, E. (2003) Bone morphogenetic proteins, theirantagonists, and the skeleton. Endocr Rev 24, 218235.Canalis, E., Giustina, A., Bilezikian, J.P. (2007) Mechanisms of anabolic therapies forosteoporosis. New Engl J Med 357, 905916.Chabance, B., Marteau, P., Rambaud, J.C., MiglioreSamour, D., Boynard, M.,Perrotin, P., Guillet, R., Jolles, P., Fiat, A.M. (1998) Casein peptide release and passage tothe blood in humans during digestion of milk or yogurt. Biochimie 80, 155165.Chasin, M., Harris, D.N. (1976) Inhibitory and activators of cyclic nucleotidephosphodiesterase. Adv Cycl Nucleotide Res 7, 22564.Chen, T.L., Feldman, D. (1979) Glucocorticoid receptors and actions in subpopulations ofcultured rat bone cells. Mechanism of dexamethasone potentiation of parathyroid hormonestimulated cyclic AMP production. J Clin Invest 63, 750758.Cheng, J., Thompson, M.A., Walker, H.J., Gray, C.E., Diaz Encarnacion, M.M.,Warner, G.M., Grande, J.P. (2004) Differential regulation of mesangial cell mitogenesis bycAMP phosphodiesterase isozymes 3 and 4. Am J Physiol Renal Physiol 287, F940953.Choi, JY., Pratap, J., Javed, A., Zaidi, S.K., van Wijnen, A.J., Lian, J.B., Stein, J.L.,Jones, S.C., Stein, G.S. (2000) In vivo replacement of the Runx/Cbfa1 gene with a mutantgene lacking a subnuclear targeting signal and transcriptional regulatory functions results insevere skeletal abnormalities. J Bone Miner Res 15, S157.Church, VL., FrancisWest, P. (2002) Wnt signalling during limb development. Int J DevBiol 46 (7),92736.

69
Collins, S.P., Uhler, M.D. (1999) Cyclic AMP and cyclic GMPdependent protein kinasesdiffer in their regulation of cyclic AMP response elementdependent gene transcription. J BiolChem 274, 8391404.Conti, M., Iona, S., Cuomo, M., Swinnen, J.V., Odeh, J., Svoboda, M.E. (1995)Characterization of a hormoneinducible, high affinity adenosine 3'5'cyclic monophosphatephosphodiesterase from the rat Sertoli cell. Biochemistry 34, 797987.Conti, M., Jin, S.L.C. (2000) The molecular biology of cyclic nucleotide phoshodiesterases.Prog Nuleic Acid Res Mol Biol 63, 138.Conti, M., Richter, W., Mehats, C., Livera, G., Park, JY., Jin, C. (2003) Cyclic AMPspecificPDE4 phosphodiesterases as critical components of cyclic AMP signaling. J Biol Chem.278:54936.Corbin, J.D., Turko, I.V., Beasley, A., Francis, S.H. (2000) Phosphorylation ofphosphodiesterase5 by cyclic nucleotidedependent protein kinase alters its catalytic andallosteric cGMPbinding activities. Eur J Biochem 267, 27607.Cote, R.H. (2004) Characteristics of photoreceptor PDE (PDE6): similarities and differencesto PDE5. Int J Impot Res 16 Suppl 1, S2833.Coudray, C., Charon, C., Komas, N., Mory, G., DiotDupuy, F., Manganiello, V., Ferre,P., Bazin, R. (1999) Evidence for the presence of several phosphodiesterase isoforms inbrown adipose tissue of Zucker rats: modulation of PDE2 by the fa gene expression. FEBSLett 456, 20710.Craig, W.S. Cheng, S., Mullen, D.G., Blevitt, J., Pierschbacher, M.D. (1995) Concept andprogress in the development of RGDcontaining peptide pharmaceuticals. Biopolymers 37,157175.Dalton, G.D., Dewey, W.L. (2006) Protein kinase inhibitor peptide (PKI): a family ofendogenous neuropeptides that modulate neuronal cAMPdependent protein kinase function.Neuropeptides 40, 2334.Daub, H., Weiss, F.U., Wallasch, C., Ullrich, A. (1996) Role of transactivation of the EGFreceptor in signaling by Gprotein coupled receptors. Nature 379, 557560.Dee, K.C. (1996a) Considerations for design of proactive dental/orthopaedic implantbiomaterials. Ph.D. Thesis, Rensselaer Polytechnic Institute, Troy, New York, 1996.Dee, K.C., Rueger, D.C., Andersen, T.T., Bizios, R. (1996b) Conditions which promotemineralization at the boneimplant interface: a model in vitro study. Biomaterials 17, 209215.Dee, K.C., Andersen, T.T., Bizios, R. (1998) Design and function of novel osteoblastadhesive peptides for chemical modification of biomaterials. J Biomed Mater Res 40, 371377.Degerman, E., Smith, C.J., Tornqvist, H., Vasta, V., Belfrage, P., Manganiello, V.C.(1990) Evidence that insulin and isoprenaline activate the cGMPinhibited lowKm cAMPphosphodiesterase in rat fat cells by phosphorylation. Proc Natl Acad Sci U S A 87, 5337.Degerman, E., Belfrage, P., Manganiello, V.C. (1997) Structure, localization, andregulation of cGMPinhibited phosphodiesterase (PDE3). J Biol Chem 272, 68236.

70
Delay, R.J., Dubin, A.E., Dionne, V.E. (1997) A cyclic nucleotidedependent chlorideconductance in olfactory receptor neurons. J Membr Biol 159, 5360.Delmi, M., Rapin, C.H., Bengoa, J.M., Delmas, P.D., Vasey, H., Bonjour, J.P. (1990)Dietary supplementation in elderly patients with fractured neck of the femur. Lancet 335,10131016.Dempster, D.W., Cosman, F., Parisien, M., Shen, V. (1993) Anabolic actions ofparathyroid hormone on bone. Endocr Rev 14, 690709.Dessauer, C.W., Gilman, A.G. (1997) The catalytic mechanism of mammalian adenylylcyclase. Equilibrium binding and kinetic analysis of Psite inhibition. J Biol Chem 272,27787–27795.DodgeKafka, K.L., Soughayer, J., Pare, G.C., Carlisle Michel, J.J., Langeberg, L.K.,Kapiloff, M.S., Scott, J.D. (2005) The protein kinase A anchoring protein mAKAPcoordinates two integrated cAMP effectors pathways. Nature 22, 574578.DortchCarnes, J., Potter, D.E. (2002) Inhibition of cAMP accumulation by kappareceptoractivation in isolated irisciliary bodies: role of phosphodiesterase and protein kinase C. JPharmacol Exp Ther 301, 599604.Drissi, H., Hott, M., Marie, P.J., Lasmoles, F. (1997) Expression of the CT/CGRP geneand its regulation by dibutyryl cyclic adenosine monophosphate in human osteoblastic cells. JBone Mineral Res 12, 18051814.Ducy, P., Zhang, R., Geoffrey, V., Ridall, A.L., Karsenty G. (1997) Osf2/Cbfa1 : Atranscriptional activator of osteoblast differentiation. Cell 89, 747754.Ducy, P., Schinke, T., Karsenty G. (2000) The osteoblast: A sophisticated fibroblast undercentral surveillance.Science 289, 15011504.Dulak, J., Jozkowicz, A., DembinskaKiec, A., Guevara, I., Zdzienicka, A., ZmudzinskaGrochot, D., Florek, I., Wojtowicz, A., Szuba, A., Cooke, J.P. (2000) Nitric oxide inducesthe synthesis of vascular endothelial growth factor by rat vascular smooth muscle cells.Arterioscler Thromb Vasc Biol 20, 659666.Eastell, R.,Reid,DM., Compston, J., Cooper, C., Fogelman, I., Francis, RM., Hosking, DJ.,Purdie, DW., Ralston, SH., Reeve, J., Russell, RG., Stevenson, JC., Torgerson, DJ. (1998) AUK Consensus Group on management of glucocorticoidinduced osteoporosis: an update.Journal of Internal Medicine 244(4):27192.Ehses, J.A., Casilla, V.R., Doty, T., Pospisilik, J.A., Winter, K.D., Demuth, H.U.,Pederson, R.A., McIntosh, C.H. (2003) Glucosedependent insulinotropic polypeptidepromotes beta(INS1) cell survival via cyclic adenosine monophosphatemediated caspase3inhibition and regulation of p38 mitogenactivated protein kinase. Endocrinology 144, 443345.Eigenthaler, M., Lohmann, S.M., Walter, U., Pilz, RB. (1999) Signal transduction bycGMPdependent protein kinases and their emerging roles in the regulation of cell adhesionand gene expression. Rev Physiol Biochem Pharmacol 135, 173209.Elbashir, S.M., Harborth, J., Weber, K., Tuschl, T. (2002) Analysis of gene function insomatic mammalian cells using small interfering RNAs. Methods (Duluth) 26, 199213.

71
elHusseini, A.E., Bladen, C., Vincent, S.R. (1995) Molecular characterization of a type IIcyclic GMPdependent protein kinase expressed in the rat brain. J Neurochem 64, 28147.Engels, P., Fichtel, K., Lubbert, H. (1994) Expression and regulation of human and ratphosphodiesterase type IV isogenes. FEBS Lett 350, 2915.Engels, P., Sullivan, M., Muller, T., Lubbert, H. (1995) Molecular cloning and functionalexpression in yeast of a human cAMPspecific phosphodiesterase subtype (PDE IVC). FEBSLett 358, 30510.Engleman, V.W., Nickols, G.A., Ross, F.P., Horton, M.A., Griggs, D.W., Settle, S.L.,Ruminski, P.G., Teitelbaum, S.L. (1997) A peptidomimetic antagonist of the alpha(v)beta3integrin inhibits bone resorption in vitro and prevents osteoporosis in vivo. J Clin Invest 99,22842292.Erba, D., Ciappellano, S., Testolin, G. (2002) Effect of the ratio of casein phosphopeptidesto calcium (w/w) on passive calcium transport in the distal small intestine of rats. Nutrition18, 743746.Farndale, R.W., Sandy, J.R., Atkinson, S.J., Pennington, S.R., Meghji, S., Meikle, M.C.(1988) Parathyroid hormone and prostaglandin E2 stimulate both inositol phosphates andcyclic AMP accumulation in mouse osteoblast cultures. Biochem J 252, 263268.Favot, L., Keravis, T., Lugnier, C. (2004) Modulation of VEGFinduced endothelial cellcycle protein expression through cyclic AMP hydrolysis by PDE2 and PDE4. ThrombHaemost 92, 63445.Fawcett, L., Baxendale, R., Stacey, P., McGrouther, C., Harrow, I., Soderling, S.,Hetman, J., Beavo, J.A., Pillips, S.C. (2000) Molecular cloning and characterization of adistinct human phoshodiesterase gene family: PDE 11A. Proc Natl Acad Sci USA 97, 37023707.Fidock, M., Miller, M., Lanfear, J. (2002) Isolation and differential tissue distribution oftwo human cDNAs encoding PDE1 splice variants. Cell Signal 14, 5360.Finkelstein, J.S., Klibanski, A., Schaefer, E.H., Hornstein, M.D., Schiff, I., Neer, R.M.(1994) Parathyroid hormone for the prevention of bone loss induced by estrogen deficiency.N Engl J Med 331, 16181623.Fischer, O.M., Hart, S., Gschwind, A., Ullrich, A. (2003) EGFR signal transactivation incancer cells. Biochem Soc Trans 31, 12031208.Fisher, D.A., Smith, J.F., Pillar, J.S., St Denis, S.H., Cheng, J.B. (1998) Isolation andcharacterization of PDE8A, a novel human cAMPspecific phosphodiesterase. BiochemBiophys Res Commun 246, 5707.Fisher, D.A., Smith, J.F., Pillar, J.S., St Denis, S.H., Cheng, J.B. (1998) Isolation andcharacterization of PDE9A, a novel human cGMPspecific phosphodiesterase. J Biol Chem273, 1555964.Forte, L.R., Currie, M.G. (1995) Guanylin: a peptide regulator of epithelial transport.FASEB J 9, 64350.Foulkes, N.S., Borrelli, E., SassoneCorsi, P. (1991) CREM gene: use of alternative DNAbinding domains generates multiple antagonists of cAMPinduced transcription. Cell 64, 739749.

72
Francis, S.H., Colbran, J.L., McAllisterLucas, L.M. Corbin, J.D. (1994) Zinc interactionsand conserved motifs of the cGMPbinding cGMPspecific phosphodiesterase suggest that itis a zinc hydrolase. J Biol Chem 269, 2247780.Francis, S.H., Corbin, J.D. (1994) Progress in understanding the mechanism and function ofcyclic GMPdependent protein kinase. Adv Pharmacol 26, 11570.Francis, S.H., Bessay, E.P., Kotera, J., Grimes, K.A., Liu, L., Thompson, W.J., Corbin,J.D. (2002) Phosphorylation of isolated human phosphodiesterase5 regulatory domaininduces an apparent conformational change and increases cGMP binding affinity. J BiolChem 277, 475817.Frandsen, E.K., Krishna, G. A. (1976) A simple ultrasensitive method for the assay ofcyclic AMP and cyclic GMP in tissues. Life Sci 76,529541.French, P.J., Bijman, J., Edixhoven, M., Vaandrager, A.B., Scholte, B.J., Lohmann,S.M., Nairn, A.C., de Jonge, H.R. (1995) Isotypespecific activation of cystic fibrosistransmembrane conductance regulatorchloride channels by cGMPdependent protein kinaseII. J Biol Chem 270, 2662631.Fitzgerald, R.J., Murray, B.A. (2006) Bioactive peptides and lactic fermantations. Int JDairy Technol 59, 118125.Fu, Q., Jilka, R.L., Manolagas, S.C., O'Brien, C.A. (2002) Parathyroid hormone stimulatesreceptor activator of NFkappa B ligand and inhibits osteoprotegerin expression via proteinkinase A activation of cAMPresponse elementbinding protein. J Biol Chem 277, 4886848875.Fuglsang, A., Rattray, F.P., Nilsson, D., Nyborg, N.C.B. (2003) Lactic acid bacteria:inhibition of angiotensin converting enzyme in vitro and in vivo. Anton Leeuw Int J G 83, 2734.Fujishige, K., Kotera, J., Michibata, H., Yuasa, K., Takebayashi, S., Okumura, K.,Omori, K. (1999) Cloning and characterization of a novel human phosphodiesterase thathydrolyzes both cAMP and cGMP (PDE10A). J Biol Chem 274, 1843845.Fujishige, K., Kotera, J., Omori, K. (1999) Striatum and testisspecific phosphodiesterasePDE10A isolation and characterization of a rat PDE10A. Eur J Biochem 266, 111827.Fujishige, K., Kotera, J., Yuasa, K., Omori, K. (2000) The human phosphodiesterasePDE10A gene genomic organization and evolutionary relatedness with other PDEs containingGAF domains. Eur J Biochem 267, 594351.Gamanuma, M., Yuasa, K., Sasaki, T., Sakurai, N., Kotera, J., Omori, K. (2003)Comparison of enzymatic characterization and gene organization of cyclic nucleotidephosphodiesterase 8 family in humans. Cell Signal 15, 56574.Gambaryan, S., Hausler, C., Markert, T., Pohler, D., Jarchau, T., Walter, U., Haase, W.,Kurtz, A., Lohmann, S.M. (1996) Expression of type II cGMPdependent protein kinase inrat kidney is regulated by dehydration and correlated with renin gene expression. J Clin Inv98, 66270.Gardner, C., Robas, N., Cawkill, D., Fidock, M. (2000) Cloning and characterization of thehuman and mouse PDE7B, a novel cAMPspecific cyclic nucleotide phosphodiesterase.Biochem Bioph Res 272, 18692.

73
Geoffroy, V., Fouque, F., Nivet, V., Clot, J.P., Lugnier, C., Desbuquois, B., Benelli C.(1999) Activation of a cGMPstimulated cAMP phosphodiesterase by protein kinase C in aliver Golgiendosomal fraction. Eur J Biochem 259, 892900.Geoffroy, V., Fouque, F., Lugnier, C., Desbuquois, B., Benelli, C. (2001) Characterizationof an in vivo hormonally regulated phosphodiesterase 3 (PDE3) associated with a liver Golgiendosomal fraction. Arch Biochem Biophys 387, 15462.Gilman, A.G. (1990) Regulation of adenylyl cyclase by G proteins. Adv Second MessengerPhosphoprotein Res 24,51–57.Giorgi, M., Giordano, D., Rosati, J., Tata, A.M., AugustiTocco, G. (2002) Differentialexpression and localization of CaMdependent phosphodiesterase genes during ontogenesis ofchick dorsal root ganglion. J Neurochem 80, 9709.Glavas, N.A., Ostenson, C., Schaefer, J.B., Vasta, V., Beavo, J.A. (2001) T cell activationupregulates cyclic nucleotide phosphodiesterases 8A1 and 7A3. Proc Natl Acad Sci U S A98, 631924.Conti, M., Beavo,J. (2007) Biochemistry and physiology of cyclic nucleotidephosphodiesterases: essential components in cyclic nucleotide signaling. Annu Rev Biochem76, 481511.Goraya, T.A., Masada, N., Ciruela, A., Cooper, D.M. (2004) Sustained entry of Ca2+ isrequired to activate Ca2+calmodulindependent phosphodiesterase 1A. J Biol Chem 279,40494504.Gori, F., Hofbauer, L.C., Dunstan, C.R., Spelsberg, T.C., Khosla, S., Riggs, B.L. (2000)The expression of osteoprotegerin and RANK ligand and the support of osteoclast formationby stromalosteoblast lineage cells is developmentally regulated. Endocrinology 141, 476876.Gordeladze, J.O., Drevon, C.A., Syversen, U., Reseland, J.E. (2002) Leptin stimulateshuman osteoblastic cell proliferation, de novo collagen synthesis, and mineralization: Impacton differentiation markers, apoptosis, and osteoclastic signaling. J Cell Biochem 85, 825836.Grange, M., Sette, C., Cuomo, M., Conti, M., Lagarde, M., Prigent, A.F., Nemoz, G.(2000) The cAMPspecific phosphodiesterase PDE4D3 is regulated by phosphatidic acidbinding. Consequences for cAMP signaling pathway and characterization of a phosphatidicacid binding site. J Biol Chem 275, 3337987.GrossLangenhoff, M., Hofbauer, K., Weber, J., Schultz, A., Schultz, J.E. (2006) cAMPis a ligand for the tandem GAF domain of human phosphodiesterase 10 and cGMP for thetandem GAF domain of phosphodiesterase 11. J Biol Chem 281, 28416.Gudi, T., Huvar, I., Meinecke, M., Lohmann, S.M., Boss, G.R., Pilz, R.B. (1996)Regulation of gene expression by cGMPdependent protein kinase. Transactivation of the cfos promoter. J Biol Chem 271, 4597600.Gudi, T., Lohmann, S.M., Pilz, R.B. (1997) Regulation of gene expression by cyclic GMPdependent protein kinase requires nuclear translocation of the kinase: identification of anuclear localization signal. Mol Cell Biol 17, 524454.Guipponi, M., Scott, H.S., Kudoh, J., Kawasaki, K., Shibuya, K., Shintani, A., Asakawa,S., Chen, H., Lalioti, M.D., Rossier, C., Minoshima, S., Shimizu, N., Antonarakis, S.E.

74
(1998) Identification and characterization of a novel cyclic nucleotide phosphodiesterase gene(PDE9A) that maps to 21q22.3: alternative splicing of mRNA transcripts, genomic structureand sequence. Hum Genet 103, 38692.Haass, N.K., Smalley, K.S., Li, L., Herlyn, M. (2005) Adhesion, migration andcommunication in melanocytes and melanoma. Pigment Cell Res 18, 150159.Haby, C., Lisovoski, F., Aunis, D., Zwiller, J. (1994) Stimulation of the cyclic GMPpathway by NO induces expression of the immediate early genes cfos and junB in PC12cells. J Neurochem 62, 496501.Hadjidakis, D.J., Androulakis, I.I. (2006) Bone remodeling. Ann NY Acad Sci 1092, 385396.Hagen, V., Dzeja, C., Bendig, J., Baeger, I., Kaupp, U.B. (1998) Novel caged compoundsof hydrolysisresistant 8BrcAMP and 8BrcGMP: photolabile NPE esters. J PhotochemPhotobiol B 42, 718.Halene, T.B., Siegel, S.J. (2007) PDE inhibitors in psychiatryfuture options for dementia,depression and schizophrenia? Drug Discov Today 12, 870878.Han, P., Zhu, X., Michaeli, T. (1997) Alternative splicing of the high affinity cAMPspecificphosphodiesterase (PDE7A) mRNA in human skeletal muscle and heart. J Biol Chem 272,161527.Hannon, G.J. (2002) RNA interference. Nature 418, 244251.Hata, Y., Yamamoto, M., Ohni, M., Nakajima, K., Nakamura, Y., Takano, T. (1996) Aplacebocontrolled study of the effect of sour milk on blood pressure in hypertensive subjects.Am J Clin Nutr 64, 767771.Hebb, A.L., Robertson, H.A., DenovanWright, E.M. (2004) Striatal phosphodiesterasemRNA and protein levels are reduced in Huntington's disease transgenic mice prior to theonset of motor symptoms. Neuroscience 123, 96781.Heino, T.J., Hentunen, T.A., Vaananen, H.K. (2002) Osteocytes inhibit osteoclastic boneresorption through transforming growth factorbeta: enhancement by estrogen. J CellBiochem 85, 185197.Heino, T.J., Hentunen, T.A., Vaananen, H.K. (2004) Conditioned medium from osteocytesstimulates the proliferation of bone marrow mesenchymal stem cells and their differentiationinto osteoblasts. Exp Cell Res 294, 458468.Hentunen, T.A., Lakkakorpi, P.T., Tuukkanen, J., Lehenkari, P.P., Sampath, T.K.,Vaananen, H.K. (1995) Effects of recombinant human osteogenic protein1 on thedifferentiation of osteoclastlike cells and bone resorption. Biochem Biophys Res Commun209, 433443.HernandezLedesma, B., Amigo, L., Ramos, M., Recio, I. (2004) Angiotensin convertingenzyme inhibitory activity in commercial fermented products. Formation of peptides undersimulated gastrointestinal digestion. J Agric Food Chem 52, 15041510.Hetman, J.M., Soderling, S.H., Glavas, N.A., Beavo, J.A. (2000) Cloning andcharacterization of PDE7B, a cAMPspecific phosphodiesterase. Proc Natl Acad Sci USA97,4726.

75
High, W.B. (1987) Effects of orally administered prostaglandin E2 on cortical bone turnoverin adult dogs: a histomorphometric study. Bone 8, 363373.Hodsman, AB., Bauer, DC., Dempster, DW., Dian, L., Hanley, DA., Harris, ST.,Kendler, DL., McClung, MR., Miller, PD., Olszynski, WP., Orwoll, E., Yuen, CK. (2005)Parathyroid hormone and teriparatide for the treatment of osteoporosis: a review of theevidence and suggested guidelines for its use. Endocrine Reviews 2005;26(5):688703.Hofbauer, L.C., Gori, F., Riggs, B.L., Lacey, D.L., Dunstan, C.R., Spelsberg, T.C.,Khosla, S. (1999) Stimulation of osteoprotegerin ligand and inhibition of osteoprotegerinproduction by glucocorticoids in human osteoblastic lineage cells: potential paracrinemechanisms of glucocorticoidinduced osteoporosis. Endocrinology 140, 43824389.Hoffmann, R., Baillie, G.S., MacKenzie, S.J., Yarwood, S.J., Houslay, M.D. (1999) TheMAP kinase ERK2 inhibits the cyclic AMPspecific phosphodiesterase HSPDE4D3 byphosphorylating it at Ser579. EMBO J 18, 893903.Hori, G., Wang, M.F., Chan, Y.C., Komatsu, T., Wong, Y., Chen, T.H., Yamamoto, K.,Nagaoka, S., Yamamoto, S. (2001) Soy protein hydrolyzate with bound phospholipidsreduces serum cholesterol levels in hypercholesterolemic adult male volunteers. BiosciBiotechnol Biochem 65, 7278.Horiguchi, N., Horiguchi, H., Suzuki, Y. (2005) Effect of wheat gluten hydrolysate on theimmune system in healthy human subjects. Biosci Biotechnol Biochem 69, 24452449.Houslay, M.D. (2001) PDE4 cAMPspecific phosphodiesterases. Prog Nucleic Acid Res MolBiol 69, 249315.Houslay, MD., Adams, DR. (2003) PDE4 cAMP phosphodiesterases: modular enzymes thatorchestrate signalling crosstalk, desensitization and compartmentalization. BiochemicalJournal. 370:118.Hsu, H., Lacey, D.L., Dunstan, C.R., Solovyev, I., Colombero, A., Timms, E., Tan, H.L.,Elliott, G., Kelley, M.J., Sarosi, I., Wang, L., Xia, X.Z., Elliott, R., Chiu, L., Black, T.,Scully, S., Capparelli, C., Morony, S., Shimamoto, G., Bass, M.B., Boyle, W.J. (1999)Tumor necrosis factor receptor family member RANK mediates osteoclast differentiation andactivation induced by osteoprotegerin ligand. Proc Natl Acad Sci U S A 96, 35403545.Hu, H., McCaw, E.A., Hebb, A.L., Gomez, G.T., DenovanWright, E.M. (2004) Mutanthuntingtin affects the rate of transcription of striatumspecific isoforms of phosphodiesterase10A. Eur J Neurosci 20, 335163.Huag, H., Zhao, Y., Liu, Z., Zhang, Y., Zhang, H., Fu, T., Ma, X. (2003) Enhancedosteoblast functions on RGD immobilized surface. J Oral Implantol 29, 7379.Huang, L.J., Durick, K., Weiner, J.A., Chun, J., Taylor, S.S. (1997) Identification of anovel protein kinase A anchoring protein that binds both type I and type II regulatorysubunits. J Biol Chem 272, 805764.Huang, Z.J., Edery, I., Rosbash, M. (1993) PAS is a dimerization domain common toDrosophila period and several transcription factors. Nature 364, 25962.Hughes, F.J., Collyer, J., Stanfield, M., Goodman, S.A. (1995) The effects of bonemorphogenetic protein2, 4, and 6 on differentiation of rat osteoblast cells in vitro.Endocrinology 136:26712677.

76
Huttunen, M.M., Pekkinen, M., Ahlström, M.E.B, LambergAllardt, C.J.E. (2007) Longterm effects of tripeptide IleProPro on osteoblast differentiation in vitro. J Nutr Biochemdoi:10.1016/j.jnutbio.2007.09.006.Idriss, S.D., Gudi, T., Casteel, D.E., Kharitonov, V.G., Pilz, R.B., Boss, G.R. (1999) Nitricoxide regulation of gene transcription via soluble guanylate cyclase and type I cGMPdependent protein kinase. J Biol Chem 274, 948993.Ishizuya, T., Yokose, S., Hori, M., Noda, T., Suda, T., Yoshiki, S., Yamaguchi, A. (1997)Parathyroid hormone exerts disparate effects on osteoblast differentiation depending onexposure time in rat osteoblastic cells. J Clin Invest 99, 29612970.Itoh, K., Udagawa, N., Katagiri, T., Iemura, S., Ueno, N., Yasuda, H., Higashio, K.,Quinn, J.M., Gillespie, M.T., Martin, T.J., Suda, T., Takahashi, N. (2001) Bonemorphogenetic protein 2 stimulates osteoclast differentiation and survival supported byreceptor activator of nuclear factorkappaB ligand. Endocrinology 142, 36563662.Iwata, K., Li, J., Follet, H., Phipps, R.J., Burr, D.B. (2006) Bisphosphonates suppressperiosteal osteoblast activity independently of resorption in rat femur and tibia. Bone 39,10538.Jaiswal, N., Haynesworth, S.E., Caplan, A.I., Bruder, S.P. (1997) Osteogenicdifferentiation of purified, cultureexpanded human mesenchymal stem cells in vitro. J CellBiochem 64, 295312.James, I.E., Dodds, R.A., Olivera, D.L., Nuttall, M.E., Gowen, M. (1996) Humanosteoclastomaderived stromal cells: correlation of the ability to form mineralized nodules invitro with formation of bone in vivo. J Bone Miner Res 11, 14531460.Jee, W.S., Ueno, K., Deng, Y.P., Woodbury, D.M. (1985) The effects of prostaglandin E2in growing rats: increased metaphyseal hard tissue and corticoendosteal bone formation.Calcif Tissue Int 37, 148157.Jee, W.S., Ueno, K., Kimmel, D.B., Woodbury, D.M., Price, P., Woodbury, L.A. (1987)The role of bone cells in increasing metaphyseal hard tissue in rapidly growing rats treatedwith prostaglandin E2. Bone 8, 171178.Jeon, M.J., Kim, J.A., Kwon, S.H., Kim, S.W., Park, K.S., Park, S.W., Kim, S.Y., Shin, C.S.(2003) Activation of peroxisome proliferatoractivated receptorgamma inhibits the Runx2mediated transcription of osteocalcin in osteoblasts. J Biol Chem 278, 232707.Jiang, Y., Vaessen, B., Lenvik, T., Blackstad, M., Reyes, M., Verfaillie, C.M. (2002a)Multipotent progenitor cells can be isolated from postnatal murine bone marrow, muscle, andbrain, Exp Hematol 30, 896–904.Jiang, Y., Jahagirdar, B.N., Reinhardt, R.L., Schwartz, R.E., Keene, C.D., OrtizGonzalez, X.R., Reyes, M., Lenvik, T., Lund, T., Blackstad, M., Du, J., Aldrich, S.,Lisberg, A., Low, W., Largaespada, D.A., Verfaillie, C.M. (2002b) Pluripotency ofmesenchymal stem cells derived from adult marrow, Nature 418, 41–49.Jiang, Y., Henderson, D., Blackstad, M., Chen, A., Miller, R.F., Verfaillie, C.M. (2003)Neuroectodermal differentiation from mouse multipotent adult progenitor cells. Proc NatlAcad Sci USA 100 (Suppl. 1), 118554–118560.

77
Jilka, R.L. (2007) Molecular and cellular mechanisms of the anabolic effect of intermittentPTH. Bone 40, 14341446.Jilka, R.L., Weinstein, R.S., Bellido, T., Parfitt, A.M., Manolagas, S.C. (1998) Osteoblastprogrammed cell death (apoptosis): modulation by growth factors and cytokines. J BoneMiner Res 13, 793802.Jilka, R.L., Weinstein, R.S., Bellido, T., Roberson, P., Parfitt, A.M., Manolagas, S.C.(1999) Increased bone formation by prevention of osteoblast apoptosis with parathyroidhormone. J Clin Invest 104, 43946.Karaplis, A.C., Goltzman, D. (2000) PTH and PTHrP effects on the skeleton. Rev EndocrMetab Disord 1, 331341.Kawaguchi, H., Pilbeam, C.C., Raisz, L.G. (1994) Anabolic effects of 3,3 ,5trooodothyronine and triiodothyroacetic acid in cultured neonatal mouse parietal bones.Endocrinology 135, 971976.Kayser, H., Meisel, H. (1996) Stimulation of human peripheral blood lymphocytes bybioactive peptides derived from bovine milk proteins. FEBS Lett 383, 1820.Keravis, T., Komas, N., Lugnier, C. (2000) Cyclic nucleotide hydrolysis in bovine aorticendothelial cells in culture: differential regulation in cobblestone and spindle phenotypes. JVasc Res 37, 23549.Kerr, M.K., Churchill, G.A. (2001) Experimental design for gene expression microarrays.Biostatistics 2:183201.Khosla, S., Atkinson, E.J., Dunstan, C.R, O'Fallon, W.M. (2002) Effect of estrogen versustestosterone on circulating osteoprotegerin and other cytokine levels in normal elderly men.J Clin Endocrinol Metab 87, 15501554.Kinoshita, T. Kobayashi, S., Ebara, S., Yoshimura, Y., Horiuchi, H., Tsutsumimoto T.,Wakabayashi, S., Takaoka, K. (2000) Phosphodiesterase inhibitors, pentoxifylline androlipram, increase bone mass mainly by promoting bone formation in normal mice. Bone 27,811817.Kitamura, T., Kitamura, Y., Kuroda, S., Hino, Y., Ando, M., Kotani, K., Konishi, H.,Matsuzaki, H., Kikkawa, U., Ogawa, W., Kasuga, M. (1999) Insulininducedphosphorylation and activation of cyclic nucleotide phosphodiesterase 3B by the serinethreonine kinase Akt. Mol Cell Biol 19, 628696.Kitts, D.D., Weiler, K. (2003) Bioactive proteins and peptides from food sources.Applications of bioprocesses used in isolation and recovery. Curr Pharm Des 9, 13091323.Kobayashi, T., Gamanuma, M., Sasaki, T., Yamashita, Y., Yuasa, K., Kotera, J., Omori,K. (2003) Molecular comparison of rat cyclic nucleotide phosphodiesterase 8 family: uniqueexpression of PDE8B in rat brain. Gene 319, 2131.Kogianni, G., Noble, B.S. (2007) The biology of osteocytes. Curr Osteoporos Rep 5, 816.Koh, AJ., Beecher, CA., Rosol, TJ., McCauley, LK. (1999) 3',5'Cyclic adenosinemonophosphate activation in osteoblastic cells: effects on parathyroid hormone1 receptorsand osteoblastic differentiation in vitro. Endocrinology. 140, 315462.

78
Komalavilas, P., Lincoln, T.M. (1996) Phosphorylation of the inositol 1,4,5trisphosphatereceptor. Cyclic GMPdependent protein kinase mediates cAMP and cGMP dependentphosphorylation in the intact rat aorta. J Biol Chem 271, 219338.Komori, T. (2005) Regulation of skeletal development by the Runx family of transcriptionfactors. J Cell Biochem 95, 445453.Komori, T. (2006) Regulation of osteoblast differentiation by transcription factors. . J CellBiochem 99, 12339.Konopleva, M., Mikhail, A., Estrov, Z., Zhao, S., Harris, D., SanchezWilliams, G.,Kornblau, S.M., Dong, J., Kliche, K.O., Jiang, S., Snodgrass, H.R., Estey, E.H.,Andreeff, M. (1999) Expression and function of leptin receptor isoforms in myeloid leukemiaand myeloplastic syndromes: proliferative and antiapoptotic activities. Blood 93, 16681676.Kopperud, R., Krakstad, C., Selheim, F., Doskeland, S.O. (2003) cAMP effectormechanisms. Novel twists for an ‘old’ signaling system, FEBS Letters 546, 121–126.Korhonen, H., Pihlanto, A. (2003) Foodderived bioactive peptidesopportunities fordesigning future foods. Curr Pharm Des 9, 12971308.Korhonen, H., Pihlanto A. (2006) Bioactive peptides: production and functionality. IntDairy J 16, 945960.Kulkarni, NH., Halladay, DL., Miles, RR., Gilbert, LM., Frolik, CA., Galvin, RJ.,Martin, TJ., Gillespie, MT., Onyia, JE. (2005) Effects of parathyroid hormone on Wntsignaling pathway in bone. Journal of Cellular Biochemistry. 95(6):117890.Kumegawa, M., Ikeda, E., Tanaka, S., Haneji, T., Yora, T., Sakagishi, Y., Minami, N.,Hiramatsu, M. (1984) The effects of prostaglandin E2, parathyroid hormone, 1,25dihydroxycholecalciferol, and cyclic nucleotide analogs on alkaline phosphatase activity inosteoblastic cells. Calcif Tissue Int 36, 7276.Kwak, B.R., Saez, J.C., Wilders, R., Chanson, M., Fishman, G.I., Hertzberg, E.L.,Spray, D.C., Jongsma, H.J. (1995) Effects of cGMPdependent phosphorylation on rat andhuman connexin43 gap junction channels. Pflugers Archiv European Journal of Physiology430, 7708.Lalli, E., SassoneCorsi, P. (1994) Signal transduction and gene regulation: the nuclearresponse to cAMP. J Biol Chem 269, 1735962.Lanyon, L., and Skerry, T. (2001) Postmenopausal osteoporosis as a failure of bone'sadaptation to functional loading: a hypothesis. J Bone Miner Res 16, 1937–1947.Lajeunesse, D., Frondoza, C., Schoffield, B., Sacktor, B. (1990) Osteocalcin secretion bythe human osteosarcoma cell line MG63. J Bone Miner Res 5, 91522.Lajeunesse, D., Kiebzak, G.M., Frondoza, C., Sacktor, B. (1991) Regulation of osteocalcinsecretion by human primary bone cells and by the human osteosarcoma cell line MG63.Bone Miner 14, 237–250.LeckaCzernik, B., Gubrik, I., Moerman, E.J., Kajkenova, O., Lipschitz, D.A.,Manolagas, S.C., Jilka, R.L. (1999) Inhibition of Osf2/Cbfa1 expression and terminalosteoblast differentiation by PPARgamma2. J Cell Biochem 74, 357371.Leroy, M.J., Mehats, C., DucGoiran, P., Tanguy, G., Robert, B., Dallot, E., Mignot,T.M., Grange, G., Ferre, F. (1999) Effect of pregnancy on PDE4 cAMPspecific

79
phosphodiesterase messenger ribonucleic acid expression in human myometrium. Cell Signal11, 3137.Leskela, H.V., Risteli, J., Niskanen, S., Koivunen, J., Ivaska, K.K., Lehenkari, P. (2003)Osteoblast recruitment from stem cells does not decrease by age at late adulthood. BiochemBiophys Res Commun 311, 10081013.Levitzki, A. (1997) Targeting signal transduction for disease therapy. Medical Oncology.14(2), 8389.Lian, J.B, Stein, G.S., Aubin, J.E. Bone formation: Maturation and Functional Activities ofOsteoblast Lineage Cells. In Primers on the Metabolic Bone Diseases and Disorders ofMineral Metabolism; Favus, M.J., Ed.; American Society of Bone and Mineral Research:Washington D.C., 2003; Chapter 3.Lim, J., Pahlke, G., Conti, M. (1999) Activation of the cAMPspecific phosphodiesterasePDE4D3 by phosphorylation. Identification and function of an inhibitory domain. J BiolChem 274, 1967785.Lincoln, T.M., Dey, N.B., Boerth, N.J., Cornwell, T.L., Soff, G.A. (1998) Nitric oxidecyclic GMP pathway regulates vascular smooth muscle cell phenotypic modulation:implications in vascular diseases. Acta Physiol Scand 164, 50715.Lincoln, T.M., Wu, X., Sellak, H., Dey, N., Choi, C.S. (2006) Regulation of vascularsmooth muscle cell phenotype by cyclic GMP and cyclic GMPdependent protein kinase.Front Biosci 11, 356367.Liu, L., Underwood, T., Li, H., Pamukcu, R., Thompson, W.J. (2002) Specific cGMPbinding by the cGMP binding domains of cGMPbinding cGMP specific phosphodiesterase.Cell Signal 14, 4551.Logan, CY., Nusse, R. (2004) The Wnt signaling pathway in development and disease. AnnuRev Cell Dev Biol 20,781810.Lohse, M.J., Benovic, J.L., Codina, J., Caron, M.G., Lefkowitz, R.J. (1990) betaArrestin:a protein that regulates betaadrenergic receptor function. Science 248(4962):154750.Loughney, K., Martins, T.J., Harris, E.A., Sadhu, K., Hicks, J.B., Sonnenburg, W.K.,Beavo, J.A., Ferguson, K. (1996) Isolation and characterization of cDNAs corresponding totwo human calcium, calmodulinregulated, 3',5'cyclic nucleotide phosphodiesterases. J BiolChem 271, 796806.Lugnier, C. (2006) Cyclic nucleotide phosphodiesterase (PDE) superfamily: a new target forthe development of specific therapeutic agents. Pharmacol Ther 109, 366398.Luttrell, L., Daaka, Y., Lefkowitz, R. (1999) Regulation of tyrosine kinase cascades by Gproteincoupled receptors. Curr Opin Cell Biol 11, 177183.LykkeAndersen, K. (2006) Regulation of gene expression in mouse embryos and itsembryonic cells through RNAi. Molecular Biotechnology. 34, 2718.Lynch, M.J., Baillie, G.S., Mohamed, A., Li, X., Maisonneuve, C., Klussmann, E., vanHeeke, G., Houslay, M.D. (2005) RNA silencing identifies PDE4D5 as the functionallyrelevant cAMP phosphodiesterase interacting with beta arrestin to control the protein kinaseA/AKAP79mediated switching of the beta2adrenergic receptor to activation of ERK inHEK293B2 cells. J Biol Chem 280, 3317833189.

80
MacFarland, R.T., Zelus, B.D., Beavo, J.A. (1991) High concentrations of a cGMPstimulated phosphodiesterase mediate ANPinduced decreases in cAMP and steroidogenesisin adrenal glomerulosa cells. J Biol Chem 266, 13642.Mahonen, A., Pirskanen, A., Keinanen, R., Maenpaa, P.H. (1990) Effect of 1,25(OH)2D3on its receptor mRNA levels and osteocalcin synthesis in human osteosarcoma cells. BiochimBiophys Acta 1048, 3037.Mahonen, A., Jukkola, A., Risteli, L., Risteli, J., Maenpaa, P.H. (1998) Type I procollagensynthesis is regulated by steroids and related hormones in human osteosarcoma cells. J CellBiochem 68, 151163.Majeska, R.J., Rodan, G.A. (1982) Alkaline phoshatase inhibition by parathyroid hormoneand isoproterenol in a clonal rat osteosarcoma cell line: Possible mediation by cyclic AMP.Calcif Tissue Int 34, 5966.Manganiello, V.C., Murata, T., Taira, M., Belfrage, P., Degerman, E. (1995) Diversity incyclic nucleotide phoshodiesterase isozyme families. Arch Biochem Biophys 322, 113.Manning, C.D., McLaughlin, M.M., Livi, G.P., Cieslinski, L.B., Torphy, T.J., Barnette,M.S. (1996) Prolonged beta adrenoceptor stimulation upregulates cAMP phosphodiesteraseactivity in human monocytes by increasing mRNA and protein for phosphodiesterases 4A and4B. J Pharmacol Exp Ther 276, 8108.Martinez, S.E., Wu, A.Y., Glavas, N.A., Tang, X.B., Turley, S., Hol, W.G., Beavo, J.A.(2002) The two GAF domains in phosphodiesterase 2A have distinct roles in dimerization andin cGMP binding. Proc Natl Acad Sci USA 99, 132605.McCann, K.B., Shiell, B.J., Michalski, W.P., Lee, A., Wan, J., Roginski, H., Coverty,M.J. (2006) Isolation and characterisation of a novel antibacterial peptide from bovine s1casein. Int Dairy J 16, 316323.Mehats, C., Andersen, CB., Filopanti, M., Jin, SL., Conti, M. (2002) Cyclic nucleotidephosphodiesterases and their role in endocrine cell signaling. Trends in Endocrinology &Metabolism 13, 2935.Meisel, H. (2001) Bioactive peptides from milk proteins: a perspective for consumers andproducers. Austr. J. Dairy Technol. 56:8391.Meisel, H., FitzGerald, R.J. (2003) Biofunctional peptides from milk proteins: mineralbinding and cytomodulatory effects. Curr Pharm Des 9, 12891295.Meisel, H., Bernard, H., FairweatherTait, S., FitzGerald, R.J., Hartmann, R., Lane,C.N., McDonagh, D., Teucher, B., Wal, J.M. (2003) Detection of caseinophosphopeptidesin the distal ileostomy fluid of human subjects. Br J Nutr 89, 351359.Mery, P.F., Lohmann, S.M., Walter, U., Fischmeister, R. (1991) Ca2+ current is regulatedby cyclic GMPdependent protein kinase in mammalian cardiac myocytes. Proc Natl AcadUSA 88, 1197201.Michibata, H., Yanaka, N., Kanoh, Y., Okumura, K., Omori, K. (2001) HumanCa2+/calmodulindependent phosphodiesterase PDE1A: novel splice variants, their specificexpression, genomic organization, and chromosomal localization. Biochim Biophys Acta1517, 27887.

81
Michaeli, T., Bloom, T.J., Martins, T., Loughney, K., Ferguson, K., Riggs, M., Rodgers,L., Beavo, J.A., Wigler, M. (1993) Isolation and characterization of a previously undetectedhuman cAMP phosphodiesterase by complementation of cAMP phosphodiesterasedeficientSaccharomyces cerevisiae. J Biol Chem 268, 1292532.Miguel, M., Aleixandre, A. (2006) Antihypertensive peptides derived from egg proteins. JNutr 136, 14571460.Miki, T., Taira, M., Hockman, S., Shimada, F., Lieman, J., Napolitano, M., Ward, D.,Taira, M., Makino, H., Manganiello, V.C. (1996) Characterization of the cDNA and geneencoding human PDE3B, the cGIP1 isoform of the human cyclic GMPinhibited cyclicnucleotide phosphodiesterase family. Genomics 36, 47685.Miro, X., PerezTorres, S., Artigas, F., Puigdomenech, P., Palacios, J.M., Mengod, G.(2002) Regulation of cAMP phosphodiesterase mRNAs expression in rat brain by acute andchronic fluoxetine treatment. An in situ hybridization study. Neuropharmacology 43, 114857.Mishaela, R., Rubin, MD., Bilezikian, JP. (2002) New anabolic therapies in osteoporosis.Current Opinion in Rheumatology 2002(14):433440.Mitsui T., Nomura S., Itakura A., Mizutani S. (2004) Role of aminopeptidases in the bloodpressure regulation. Biol Pharmacoll Bull 27(6):76871Miyamoto, K., Suzuki, H., Yamamoto, S., Saitoh, Y., Ochiai, E., Moritani, S., Yokogawa,K., Waki, Y., Kasugai, S., Sawanishi, H., Yamagami, H. (2003) Prostaglandin E2mediatedanabolic effect of a novel inhibitor of phosphodiesterase 4, XT611, in the in vitro bonemarrow culture. J Bone Miner Res 18:14717.Miyamoto, K., Waki, Y., Horita, T., Kasugai, S., Ohya, K. (1997) Reduction of bone lossby denbufylline, an inhibitor of phosphodiesterase 4. Biochem Pharmacol 54, 6136177.Miyamoto, K., Suzuki, H., Yamamoto, S., Saitoh, Y., Ochiai, E., Moritani, S., Yokogawa,Ng KW. Romas E. Donnan L. Findlay DM. (1997) Bone biology. Baillieres ClinicalEndocrinology & Metabolism. 11(1):122.Mogi, M., Togari, A. (2003) Activation of caspases is required for osteoblasticdifferentiation. J Biol Chem 278, 4747747482.Mori, K., Shioi, A., Jono, S., Nishizawa, Y., Morii, H. (1999) Dexamethasone enhances Invitro vascular calcification by promoting osteoblastic differentiation of vascular smoothmuscle cells. Arterioscler Thromb Vascul Biol 19, 21122118.Mousses, S., Caplen, N.J., Cornelison, R., Weaver, D., Basik, M., Hautaniemi, S.,Elkahloun, A.G., Lotufo, R.A., Choudary, A., Dougherty, E.R., Suh, E., Kallioniemi, O.(2003) RNAi microarray analysis in cultured mammalian cells. Genome Res 13, 23412347.Mullally, M.M., Meisel, H., FitzGerald, R.J. (1996) Synthetic peptides corresponding toalphalactalbumin and betalactoglobulin sequences with angiotensinIconverting enzymeinhibitory activity. Biological Chemistry HoppeSeyler 377, 25926Muradov, K.G., Boyd, K.K., Martinez, S.E., Beavo, J.A., Artemyev, N.O. (2003) TheGAFa domains of rod cGMPphosphodiesterase 6 determine the selectivity of the enzymedimerization. J Biol Chem 278, 10594601.

82
Murray, E., Provvedini, D., Curran, D., Catherwood, B., Sussman, H., Manolagas, S.(1987) Characterization of a human osteoblastic osteosarcoma cell line (SAOS2) with highbone alkaline phosphatase activity. J Bone Miner Res 2, 2318.Nakashima, K., Zhou, X., Kunkel, G., Zhang, Z., Deng, J.M, Behringer, R.R., deCrombrugghe, B. (2002) The novel zing fingercontaining transcription factor osterix isrequired for osteoblast differentiation and bone formation. Cell 108, 1729.Nakashima, A., Katagiri, T., Tamura, M. (2004) Crosstalk between Wnt and bonemorphogenetic protein 2 (BMP2) signaling in differentiation pathway of C2C12 myoblasts. JBiol Chem 280, 3766037668.Nakata, A., Ogawa, K., Sasaki, T., Koyama, N., Wada, K., Kotera, J., Kikkawa, H.,Omori, K., Kaminuma, O. (2002) Potential role of phosphodiesterase 7 in human T cellfunction: comparative effects of two phosphodiesterase inhibitors. Clin Exp Immunol 128,460466.Narita, M., Murata, T., Shimizu, K., Sugiyama, T., Nakagawa, T., Manganiello, VC.,Tagawa, T. (2003) Phosphodiesterase 4 in osteoblastic osteosarcoma cells as a potentialtarget for growth inhibition. AntiCancer Drug. 14, 377381.Narva, M., Halleen, J., Vaananen, K., Korpela, R. (2004) Effects of Lactobacillushelveticus fermented milk on bone cells in vitro. Life Sci 75, 17271734.Narva, M., Rissanen, J., Halleen, J., Vapaatalo, H., Vaananen, K., Korpela, R. (2007)Effects of bioactive peptide, valylprolylproline (VPP), and lactobacillus helveticusfermented milk containing VPP on bone loss in ovariectomized rats. Ann Nutr Metab 51, 6574.Nelson, W.J., Nusse, R. (2004) Convergence of Wnt, betacatenin, and cadherin pathways.Science 303, 14831487.Nielsen, C.U., Brodin, B. (2003) Di/tripeptide transporters as drug delivery targets:regulation of transport under physiological and pathophysiological conditions Curr DrugTargets 4, 373388.Noda, M., Yoon, K., Rodan, G.A. (1988) Cyclic AMPmediated stabilization of osteocalcinmRNA in rat osteoblastlike cells treated with parathyroid hormone. J Biol Chem 263,18574–18577.Noda, M., Rodan, G.A. (1989) Transcriptional regulation of osteopontin production in ratosteoblastlike cells by parathyroid hormone. J Cell Biol 108, 713718.Nuttall, M.E., Patton, A.J., Olivera, D.L., Nadeau, D.P., Gowen, M. (1998) Humantrabecular bone cells are able to express both osteoblastic and adipocytic phenotype:implications for osteopenic disorders. J Bone Miner Res 13, 37182.O'Brien, C.A., Jia, D., Plotkin, L.I., Bellido, T., Powers, C.C., Stewart, S.A., Manolagas,S.C., Weinstein, R.S. (2004) Glucocorticoids act directly on osteoblasts and osteocytes toinduce their apoptosis and reduce bone formation and strength. Endocrinology 145, 18351841.Oger, S., Mehats, C., Dallot, E., Ferre, F., Leroy, M.J. (2002) Interleukin1beta inducesphosphodiesterase 4B2 expression in human myometrial cells through a prostaglandin E2

83
and cyclic adenosine 3',5'monophosphatedependent pathway. J Clin Endocrinol Metab 87,552431.Ondetti, M.A., Rubin, B., Cushman, D.W. (1977) Design of specific inhibitors ofangiotensinconverting enzyme: new class of orally active antihypertensive agents. Science196, 441444.Oki, N., Takahashi, S.I., Hidaka, H., Conti, M. (2000) Short term feedback regulation ofcAMP in FRTL5 thyroid cells. Role of PDE4D3 phosphodiesterase activation. J Biol Chem275, 108317.Omi, K., Tokunaga, K., Hohjoh, H. (2004) Longlasting RNAi activity in mammalianneurons. FEBS Lett 558, 8995.Omori, K., Kotera, J. (2007) Overview of PDEs and their regulation. Circ Res 100, 309327.Pai, S.I., Lin, Y.Y., Macaes, B., Meneshian, A., Hung, C.F., Wu, T.C. (2006) Prospects ofRNA interference therapy for cancer. Gene Ther 13, 464477.Parfitt, A.M. (1994) Osteonal and hemiosteonal remodeling: the spatial and temporalframework for signal traffic in adult human bone. J Cell Biochem 55(3):27386.Park, H., Yim, M. (2007) Rolipram, a phosphodiesterase 4 inhibitor, suppresses PGE2induced osteoclast formation by lowering osteoclast progenitor cell viability. Arch PharmaRes 30, 486492.Partridge, N.C., Bloch, S.R., Pearman, A.T. (1994) Signal transduction pathways mediatingparathyroid hormone regulation of osteoblastic gene expression. J Cell Biochem 55, 321327.Patschan, D., Loddenkemper, K., Buttgereit, F. (2001) Molecular mechanisms ofglucocorticoidinduced osteoporosis. Bone 29(6):498505.Pavalko, F.M., Otey, C.A. (1994) Role of adhesion molecule cytoplasmic domains inmediating interactions with the cytoskeleton. Proc Soc Exp Biol Med 205, 282293.Pfeifer, A., Aszodi, A., Seidler, U., Ruth, P., Hofmann, F., Fassler, R. (1996) Intestinalsecretory defects and dwarfism in mice lacking cGMPdependent protein kinase II. Science274, 20822086.Pilz, R.B, Broderick K.E. (2005) Role of cyclic GMP in gene regulation. Front Biosci 10,12391268.Price, P.A., Otsuka, A.A., Poser, J.W., Kristaponis, J., Raman, N. (1976) Characterizationof a gammacarboxyglutamic acidcontaining protein from bone. P Natl Acad Sci USA 73,14471451.Rahn, T., Ronnstrand, L., Leroy, M.J., Wernstedt, C., Tornqvist, H., Manganiello, V.C.,Belfrage, P., Degerman, E. (1996) Identification of the site in the cGMPinhibitedphosphodiesterase phosphorylated in adipocytes in response to insulin and isoproterenol. JBiol Chem 271, 1157580.Raisz, L.G. (1999) Physiology and pathophysiology of bone remodeling. Clin Chem 45,13531358.Rawadi, G., Ferrer, C., SpinellaJaegle, S., RomanRoman, S., Bouali, Y., Baron, R.(2001) 1(5oxohexyl)3,7Dimethylxanthine, a phosphodiesterase inhibitor, activates MAPKcascades and promotes osteoblast differentiation by a mechanism independent of PKA

84
activation (pentoxifylline promotes osteoblast differentiation). Endocrinology 142(11):467382.Rawadi, G., RomanRoman, S. (2005) Wnt signalling pathway: a new target for thetreatment of osteoporosis. Expert Opin Ther Tar 9, 10631077.Reeve, J. (1996) PTH: a future role in the management of osteoporosis?. J Bone Miner Res11, 440445.Rehfuss, R.P., Walton, K.M., Loriaux, M.M., Goodman, R.H. (1991) The cAMPregulated enhancerbinding protein ATF1 activates transcription in response to cAMPdependent protein kinase A. J Biol Chem 266, 184314.Reinhardt, R.R., Chin, E., Zhou, J., Taira, M., Murata, T., Manganiello, V.C., Bondy,C.A. (1995) Distinctive anatomical patterns of gene expression for cGMPinhibited cyclicnucleotide phosphodiesterases. J Clin Invest 95, 152838.Rentero, C., Monfort, A., Puigdomenech, P. (2003) Identification and distribution ofdifferent mRNA variants produced by differential splicing in the human phosphodiesterase9A gene. Biochem Biophys Res Commun 301, 68692.Reyes, M., Verfaillie, C.M. (2001) Characterization of multipotent adult progenitor cells, asubpopulation of mesenchymal stem cells, Ann. N. Y. Acad. Sci. 938, 231–233.Richter, W., Conti, M. (2002) Dimerization of the type 4 cAMPspecific phosphodiesterasesis mediated by the upstream conserved regions (UCRs). J Biol Chem 277, 4021221.Rizzoli, R., Bonjour, J.P. (2004) Dietary protein and bone health.[comment]. J Bone MinerRes 19, 527531.Rizzello, C.G., Losito, I., Gobbetti, M., Carbonara, T., De Bari, M.D., Zambonin, P.G.(2005a) Antibacterial activities of peptides from the watersoluble extracts of Italian cheesevarieties. J Dairy Sci 88, 23482360.Rizzello, C.G., Losito, I., Gobbetti, M., Carbonara, T., De Bari, M.D., Zambonin, P.G.(2005b) Antibacterial activities of peptides from the watersoluble extracts of Italian cheesevarieties. J Dairy Sci 88, 23482360.Rodan, S.B., Imai, Y., Thiede, M.A., Wesolowski, G., Thompson, D., BarShavit, Z.,Shull, S., Mann, K., Rodan, G.A. (1987) Characterization of a human osteosarcoma cell line(Saos2) with osteoblastic properties. Cancer Research 47, 49616.Rondinone, C.M., Carvalho, E., Rahn, T., Manganiello, V.C., Degerman, E., Smith, U.P.(2000) Phosphorylation of PDE3B by phosphatidylinositol 3kinase associated with theinsulin receptor. J Biol Chem 275, 100938.Rosen, C.J., Donahue, L.R. (1998) Insulin like growth factors and bonethe osteoporosisconnection revisited. Proc Soc Exp Biol Med 219, 17.Rosman, G.J., Martins, T.J., Sonnenburg, W.K., Beavo, J.A., Ferguson, K., Loughney,K. (1997) Isolation and characterization of human cDNAs encoding a cGMPstimulated 3',5'cyclic nucleotide phosphodiesterase. Gene 191, 8995.Ross, E.M., Gilman, A.G. (1977) Resolution of some components of adenylate cyclasenecessary for catalytic activity. J Biol Chem 252, 69669.Rotella, DP. (2002) Phosphodiesterase 5 inhibitors: current status and potential applications.Nature Reviews. Drug Discovery 1(9):67482.

85
Sakamoto, M.K., Suzuki, K., Takiya, S., Yoshimura, Y., Imai, T., Matsumoto, A.,Nakamura, S. (1998) Developmental profiles of phosphorylated and unphosphorylatedCREBs in murine calvarial MC3T3E1 cells. J Biochem 3, 399407.Sasaki, N., Kusano, E., Ando, Y., Yano, K., Tsuda, E., Asano, Y. (2001) Glucocorticoiddecreases circulating osteoprotegerin (OPG): possible mechanism for glucocorticoid inducedosteoporosis. Nephrol Dial Transplant 16, 479482.Sasaki, T., Kotera, J., Yuasa, K., Omori, K. (2000) Identification of human PDE7B, acAMPspecific phosphodiesterase. Biochem Bioph Res Co 271, 57583.SassoneCorsi, P. (1995) Transcription factors responsive to cAMP. Annu Rev Cell Dev Biol11, 35577.Schaffner, P., Meyer, J., Dard, M., Wenz, R., Nies, B., Verrier, S., Kessler, H.,Kantlehner, M. (1999) Induced tissue integration of bone implants by coating with boneselective RGDpeptides in vitro and in vivo studies. J Mater Scie Mater Med 10, 837839.Scherr, M., Eder, M. (2007) Gene silencing by small regulatory RNAs in mammalian cells.Cell Cycle 6, 4449.Schuler, M., Owen, G.R., Hamilton, D.W., de Wild, M., Textor, M., Brunette, D.M.,Tosatti, S.G. (2006) Biomimetic modification of titanium dental implant model surfacesusing the RGDSPpeptide sequence: a cell morphology study. Biomaterials 27, 40034015.Scutt, A., Beier, N., Fittschen, C. (2004) EMD273316 & EMD95833, type 4phosphodiesterase inhibitors, stimulate fibroblasticcolony formation by bone marrow cellsvia direct inhibition of PDE4 and the induction of endogenous prostaglandin synthesis. BMCPharmacology 4(1);10.Semizarov, D., Kroeger, P., Fesik, S. (2004) siRNAmediated gene silencing: a globalgenome view. Nucleic Acids Res 32, 38363845.Sette, C., Iona, S., Conti, M. (1994) The shortterm activation of a rolipramsensitive,cAMPspecific phosphodiesterase by thyroidstimulating hormone in thyroid FRTL5 cells ismediated by a cAMPdependent phosphorylation. J Biol Chem 269, 924552.Sette, C., Conti, M. (1996) Phosphorylation and activation of a cAMPspecificphosphodiesterase by the cAMPdependent protein kinase. Involvement of serine 54 in theenzyme activation. J Biol Chem 271, 1652634.Shalhoub, V., Conlon, D., Tassinari, M., Quinn, C., Partridge, N., Stein, G.S., Lian, J.B.(1992) Glucocorticoids promote development of the osteoblast phenotype by selectivelymodulating expression of cell growth and differentiation associated genes. J Cell Biochem 50,425440.Shi, X.M., Blair, H.C., Yang, X., McDonald, J.M., Cao, X. (2000) Tandem repeat ofC/EBP binding sites mediates PPARgamma2 gene transcription in glucocorticoidinducedadipocyte differentiation. J Cell Biochem 76, 518527.Shimabukuro, M., Zhou, YT., Levi, M., Unger, RH. (1998) Fattyacid induced beta cellapoptosis: a link between obesity and diabetes. Proc Natl Acad Sci USA 95, 24982501.Simonet, W.S., Lacey, D.L., Dunstan, C.R., Kelley, M., Chang, M.S., Luthy, R., Nguyen,H.Q., Wooden, S., Bennett, L., Boone, T., Shimamoto, G., DeRose, M., Elliott, R.,Colombero, A., Tan, H.L., Trail, G., Sullivan, J., Davy, E., Bucay, N., RenshawGegg, L.,

86
Hughes, T.M., Hill, D., Pattison, W., Campbell, P., Sander, S., Van, G., Tarpley, J.,Derby, P., Lee, R., Boyle, W.J. (1997) Osteoprotegerin: a novel secreted protein involved inthe regulation of bone density. Cell 89, 309319.Skalhegg, B.S., Tasken, K. (2000) Specificity in the cAMP/PKA signaling pathway.Differential expression,regulation, and subcellular localization of subunits of PKA. FrontBiosci 5, D678693.Smith, S.J., BrookesFazakerley, S., Donnelly, L.E., Barnes, P.J., Barnette, M.S.,Giembycz, M.A. (2003) Ubiquitous expression of phosphodiesterase 7A in humanproinflammatory and immune cells. Am J Physiol Lung Cell Mol Physiol 284, L27989.Soderling, S.H., Bayuga, S.J., Beavo, J.A. (1998a) Cloning and characterization of a cAMPspecific cyclic nucleotide phoshodiesterase. Proc Natl Acad Sci USA 95, 89918996.Soderling, S.H., Bayuga, S.J., Beavo, J.A. (1998b) Identification and characterization of anovel family of cyclic nucleotide phoshodiesterases. J Biol Chem 273, 1555315558.Soderling, S.H., Beavo, J.A. (2000) Regulation of cAMP and cGMP signaling: newphosphodiesterases and new functions. Curr Opin Cell Biol 12,1749.Sonnenburg, W.K., Mullaney, P.J., Beavo, J.A. (1991) Molecular cloning of a cyclic GMPstimulated cyclic nucleotide phosphodiesterase cDNA. Identification and distribution ofisozyme variants. J Biol Chem 266, 1765561.Sooy, K., Sabbagh, Y., Demay, M.B. (2005) Osteoblasts lacking the vitamin D receptordisplay enhanced osteogenic potential in vitro. J Cell Biochem 94, 8187.Sprague, S.M., Popovtzer, M.M., DranitzkiElhalel, M., Wald, H. (1996) Parathyroidhormoneinduced calcium efflux from cultured bone is mediated by protein kinase Ctranslocation. Am J Physiol 271, F113946.Spence, S., Rena, G., Sullivan, M., Erdogan, S., Houslay, M.D. (1997) Receptormediatedstimulation of lipid signalling pathways in CHO cells elicits the rapid transient induction ofthe PDE1B isoform of Ca2+/calmodulinstimulated cAMP phosphodiesterase. Biochem J321, 15763.Stanford, C.M., Jacobson, P.A., Eanes, E.D., Lembke, L.A., Midura, R.J. (1995) RapidlyForming Apatitic Mineral in an Osteoblastic Cell Line (UMR 10601 BSP). J Biol Chem 270,9420–9428.Steffansen, B., Nielsen, C.U., Brodin, B., Eriksson, A.H., Andersen, R., Frokjaer, S.(2004) Intestinal solute carriers: an overview of trends and strategies for improving oral drugabsorption. Eur J Pharm Sci 21, 316.Stork, P.J., Schmitt, J.M. (2002) Crosstalk between cAMP and MAP kinase signaling in theregulation of cell proliferation. Trends Cell Biol 12, 258266.Swarthout, J.T., Doggett, T.A., Lemker, J.L., Partridge, N.C. (2001) Stimulation ofextracellular signalregulated kinases and proliferation in rat osteoblastic cells by parathyroidhormone is protein kinase Cdependent. J Biol Chem 276, 75867592.Swarthout, J.T., D'Alonzo, R.C., Selvamurugan, N., Partridge, N.C. (2002) Parathyroidhormonedependent signaling pathways regulating genes in bone cells. Gene 282, 117.

87
Swinnen, J.V., Tsikalas, K.E., Conti, M. (1991) Properties and hormonal regulation of twostructurally related cAMP phosphodiesterases from the rat Sertoli cell. J Biol Chem 266,183707.Takabatake, Y., Isaka, Y., Mizui, M., Kawachi, H., Shimizu, F., Ito T., Hori, M., Imai, E.(2005) Exploring RNA interference as a therapeutic strategy for renal disease. Gene Ther 12,965973.Takami, M., Cho, E.S., Lee, S.Y., Kamijo, R., Yim, M. (2005) Phosphodiesterase inhibitorsstimulate osteoclast formation via TRANCE/RANKL expression in osteoblasts: possibleinvolvement of ERK and p38 MAPK pathways. FEBS Lett 579, 832838.Takeda, K., Kaisho, Y., Yoshida, N., Takeda, J., Kishimoto, T., Akira, S. (1998) STAT3activation is responsible for IL6dependent T cell proliferation through preventing apoptosis:generation and characterizaion of T cellspecific STAT3deficient mice. J Immunol 161,46524660.Tam, C.S., Heersche, J.N., Murray, T.M., Parsons, J.A. (1982) Parathyroid hormonestimulates the bone apposition rate independently of its resorptive action: differential effectsof intermittent and continuous administration. Endocrinology 110, 506512.Tanaka, H., Seino, Y. (2004) Direct action of 1,25dihydroxyvitamin D on bone: VDRKObone shows excessive bone formation in normal mineral condition. J Steroid Biochem 8990,343345.Taussig, R., Gilman, A.G. (1995) Mammalian membranebound adenylyl cyclases, J BiolChem 270, 1–4.Taylor, S.S., Kim, C., Vigil, D., Haste, N.M., Yang, J., Wu, J., Anand, G.S. (2005)Dynamics of signaling by PKA. Biochimi Biophys Acta 1754, 2537.Teitelbaum, S.L. (2000) Bone resorption by osteoclast. Science 289, 15041508.Teitelbaum SL. Ross FP. (2003) Genetic regulation of osteoclast development and function.Nature Reviews Genetics 4(8), 63849.Theofan, G., Price, P.A. (1989) Bone Gla protein messenger ribonucleic acid is regulated byboth 1,25dihydroxyvitamin D3 and 3',5'cyclic adenosine monophosphate in ratosteosarcoma cells. Mol Endocrinol 3, 3643.Thompson, W.J., Appleman, M.M. (1971) Multiple cyclic nucleotide phosphodiesteraseactivities from rat brain. Biochemistry 10, 311316.Toranzo, G.S., Oterino, J., Zelarayan, L., Bonilla, F., Buhler, M.I. (2007) Spontaneous andLHinduced maturation in Bufo arenarum oocytes: importance of gap junctions. Zygote 15,6580.Torphy, T.J., Zhou, H.L., Foley, J.J., Sarau, H.M., Manning, C.D., Barnette, M.S. (1995)Salbutamol upregulates PDE4 activity and induces a heterologous desensitization of U937cells to prostaglandin E2. Implications for the therapeutic use of betaadrenoceptor agonists. JBiol Chem 270, 23598604.Torphy, TJ. (1998) Phosphodiesterase isozymes: molecular targets for novel antiasthmaagents. American Journal of Respiratory & Critical Care Medicine 157(2):35170.TorrasLlort, M., Azorin, F. (2003) Functional characterization of the humanphosphodiesterase 7A1 promoter. Biochem J 373, 83543.

88
Tsuboi, Y., Shankland, S.J., Grande, J.P., Walker, H.J., Johnson, R.J., Dousa, T.P. (1996)Suppression of mesangial proliferative glomerulonephritis development in rats by inhibitorsof cAMP phosphodiesterase isozymes types III and IV. J Clin Invest 98, 262270.Tsou, K., Snyder, G.L., Greengard, P. (1993) Nitric oxide/cGMP pathway stimulatesphosphorylation of DARPP32, a dopamine and cAMPregulated phosphoprotein, in thesubstantia nigra. Proc Natl Acad Sci USA 90, 34625.Tuder, R.M., Flook, B.E., Voelkel, N.F. (1995) Increased gene expression for VEGF and theVEGF receptors KDR/Flk and Flt in lungs exposed to acute or to chronic hypoxia.Modulation of gene expression by nitric oxide. J Clin Invest. 95, 1798807.Turowski, P., Adamson, P., Greenwood, J. (2005) Pharmacological targeting of ICAM1signaling in brain endothelial cells: potential for treating neuroinflammation. Cell MolNeurobiol 25, 153170.Urena, P., IidaKlein, A., Kong, X.F., Juppner, H., Kronenberg, H.M., AbouSamra,A.B., Segre, G.V. (1994) Regulation of parathyroid hormone (PTH)/PTHrelated peptidereceptor messenger ribonucleic acid by glucocorticoids and PTH in ROS 17/2.8 and OK cells.Endocrinology 134, 451456.Uzel, M.I., Scott, I.C., BabakhanlouChase, H., Palamakumbura, A.H., Pappano, W.N.,Hong, HH., Greenspan, D.S., Trackman, P.C. (2001) Multiple bone morphogeneticprotein 1related mammalian metalloproteinases process prolysyl oxidase at the correctphysiological site and control lysyl oxidase activation in mouse embryo fibroblast cultures. JBiol Chem 276, 2253722543.Van Sande, J., Lefort, A., Beebe, S., Roger, P., Perret, J., Corbin, J., Dumont, J.E. (1989)Pairs of cyclic AMP analogs, that are specifically synergistic for type I and type II cAMPdependent protein kinases, mimic thyrotropin effects on the function, differentiationexpression and mitogenesis of dog thyroid cells. Eur J Biochem 183,699708.van Staveren, W.C., Glick, J., Markerinkvan Ittersum, M., Shimizu, M., Beavo, J.A.,Steinbusch, H.W., de Vente, J. (2002) Cloning and localization of the cGMPspecificphosphodiesterase type 9 in the rat brain. J Neurocytol 31, 72941.Veeman, M.T., Axelrod, J.D., Moon, R.T. (2003) A second canon. Functions andmechanisms of betacateninindependent Wnt signaling. Dev Cell 5, 367377.Verma, U.N., Surabhi RM. Schmaltieg, A., Becerra, C., Gaynor, R.B. (2003) Smallinterfering RNAs directed against betacatenin inhibit the in vitro and in vivo growth of coloncancer cells. Clin Cancer Res 9, 12911300.Verrier, S., Pallu, S., Bareille, R., Jonczyk, A., Meyer, J., Dard, M., Amedee, J. (2002)Function of linear and cyclic RGDcontaining peptides in osteoprogenitor cells adhesionprocess. Biomaterials 23, 585596.Zhang, M., Xuan, S., Bouxsein, M.L., von Stechow, D., Akeno, N., Faugere, M.C.,Malluche, H., Zhao, G., Rosen, C.J., Efstratiadis, A., Clemens, T.L. (2002) Osteoblastspecific knockout of the insulinlike growth factor (IGF) receptor gene reveals an essentialrole of IGF signaling in bone matrix mineralization. J Biol Chem 277, 4400512.Zhao, A.Z., Bornfeldt, K.E., Beavo, J.A. (1998) Leptin inhibits insulin secretion byactivation of phosphodiesterase 3B. J Clin Invest 102, 86973.

89
Zhao, A.Z., Huan, J.N., Gupta, S., Pal, R., Sahu, A. (2002) A phosphatidylinositol 3kinasephosphodiesterase 3Bcyclic AMP pathway in hypothalamic action of leptin on feeding. NatNeurosci 5, 7278.Zhao,G., MonierFaugere, M.C., Langub, M.C., Geng, Z., Nakayama, T., Pike, J.W.,Chernausek, S.D., Rosen, C.J., Donahue, L.R., Malluche, H.H., Fagin, J.A., Clemens,T.L. (2000) Targeted overexpression of insulinlike growth factor I to osteoblasts oftransgenic mice: increased trabecular bone volume without increased osteoblast proliferation.Endocrinology 141, 267482.Zhao, S., Zhang, Y.K., Harris, S., Ahuja, S.S., Bonewald, L.F. (2002) MLOY4 osteocytelike cells support osteoclast formation and activation. J Bone Miner Res 17, 20682079.Zhong, Y. (1995) Mediation of PACAPlike neuropeptide transmission by coactivation ofRas/Raf and cAMP signal transduction pathways in Drosophila. Nature 375, 58892.Ziff, EB. (1990) Transcription factors: a new family gathers at the cAMP response site.Trends Genet 6, 6972.ZmudaTrzebiatowska, E., Manganiello, V., Degerman, E. (2007) Novel maechanism ofthe regulation of Protein kinase B in adipocytes; implications for Protein kinase A, Epac,Phoshpodiesterases 3 and 4. Cell Signal 19, 8186.Zoraghi, R., Bessay, E.P., Corbin, J.D., Francis, S.H. (2005) Structural and functionalfeatures in human PDE5A1 regulatory domain that provide for allosteric cGMP binding,dimerization, and regulation. J Biol Chem 280, 1205163.Yamagami, H., Nishioka, T., Ochiai, E., Fukushima, K., Nomura, M., Kasugai, S.,Moritani, S., Yokogawa, K., Miyamoto, K. (2003) Inhibition of osteoclastogenesis by aphosphodiesterase 4 inhibitor XT611 through synergistic action with endogenousprostaglandin E2. Biochem Pharmacol 66, 801807.Yan, C., Zhao, A.Z., Sonnenburg, W.K., Beavo, J.A. (2001) Stage and cellspecificexpression of calmodulindependent phosphodiesterases in mouse testis. Biol Reprod 64,174654.Yanaka, N., Kurosawa, Y., Minami, K., Kawai, E., Omori, K. (2003) cGMPphosphodiesterase activity is upregulated in response to pressure overload of rat ventricles.Biosci Biochem 67, 9739.Yang, G., McIntyre, K.W., Townsend, R.M., Shen, H.H., Pitts, W.J., Dodd, J.H., Nadler,S.G., McKinnon, M., Watson, A.J. (2003) Phosphodiesterase 7Adeficient mice havefunctional T cells. J Immunol 171, 64146420.Yang, Q., Paskind, M., Bolger, G., Thompson, W.J., Repaske, D.R., Cutler, L.S.,Epstein, P.M. (1994) A novel cyclic GMP stimulated phosphodiesterase from rat brain.Biochem Biophys Res Commun 205, 18508.Yuan, J., Kramer, A., Matthess, Y., Yan, R., Spankuch, B., Gatje, R., Knecht, R.,Kaufmann, M., Strebhardt, K. (2006) Stable gene silencing of cyclin B1 in tumor cellsincreases susceptibility to taxol and leads to growth arrest in vivo. Oncogene 25, 17531762.Xing, L., Boyce, BF. (2005) Regulation of apoptosis in osteoclasts and osteoblastic cells.Biochem Biophys Res Commun 328, 709720.

90
Wakabayashi, S., Tsutsumimoto, T., Kawasaki, S., Kinoshita, T., Horiuchi, H., Taoka,K. (2002) Involvement of Phoshodiesterase Isozymes in Osteoblastic Differentiation. J BoneMiner Res 12, 172178.Waki, Y., Horita, T., Miyamoto, K., Ohya, K., Kasugai, S. (1999) Effects of XT44, aphosphodiesterase 4 inhibitor, in osteoblastgenesis and osteoclastgenesis in culture and itstherapeutic effects Japanese Journal of Pharmacology. 79(4):47783.Walsh, C.A., Bowler, W.B., Bilbe, G., Fraser, W.D., Gallagher, J.A. (1997) Effects ofPTH on PTHrP gene expression in human osteoblasts: upregulation with the kinetics of animmediate early gene. Biochem Biophys Res Commun 239, 155159.Valta, M.P., Hentunen, T., Qu, Q., Valve, E.M., Harjula, A., Seppanen, J.A., Vaananen,H.K., Harkonen, P.L. (2006) Regulation of osteoblast differentiation: a novel function forfibroblast growth factor 8. Endocrinology. 147, 21712182.Wang, D., Deng, C., BugajGaweda, B., Kwan, M., Gunwaldsen, C., Leonard, C., Xin,X., Hu, Y., Unterbeck, A., De Vivo, M. (2003) Cloning and characterization of novelPDE4D isoforms PDE4D6 and PDE4D7. Cell Signal 15, 88391.Wang, J.H., Sharma, R.K., Mooibroek, M.J. (1990) Calmodulinstimulated cyclicnucleotide phosphodiesterases. In J. Beavo, & M.D. Houslay (Eds.), Cyclic NucleotidePhosphodiesterases: Structure, Regulation and Drug Action (pp.1959). John Wiley and SonsLtd.Wang, P., Wu, P., Ohleth, K.M., Egan, R.W., Billah, M.M. (1999) Phosphodiesterase 4B2is the predominant phosphodiesterase species and undergoes differential regulation of geneexpression in human monocytes and neutrophils. Mol Pharmacol 56, 1704.Wang, P., Wu, P., Egan, R.W., Billah, M.M. (2003) Identification and characterization of anew human type 9 cGMPspecific phosphodiesterase splice variant (PDE9A5). Differentialtissue distribution and subcellular localization of PDE9A variants. Gene 314, 1527.Wechsler, J., Choi, Y.H., Krall, J., Ahmad, F., Manganiello, V.C., Movsesian, M.A.(2002) Isoforms of cyclic nucleotide phosphodiesterase PDE3A in cardiac myocytes. J BiolChem 277, 380728.Weinstein, R.S., Jilka, R.L., Parfitt, A.M., Manolagas, S.C. (1998) Inhibition ofosteoblastogenesis and promotion of apoptosis of osteoblasts and osteocytes byglucocorticoids. Potential mechanisms of their deleterious effects on bone. J Clin Invest 102,27482.White, R.E., Kryman, J.P., ElMowafy, A.M., Han, G., Carrier, G.O. (2000) cAMPdependent vasodilators crossactivate the cGMPdependent protein kinase to stimulateBK(Ca) channel activity in coronary artery smooth muscle cells. Circ Res 86, 897905.Whitfield, J.F., Morley, P. (1995) Small bonebuilding fragments of parathyroid hormone:new therapeutic agents for osteoporosis. Trends Pharmacol Sci 16, 382386.Whyte, M.P., Obrecht, S.E., Finnegan, P.M., Jones, J.L., Podgornik, M.N., McAlister,W.H., Mumm, S. (2002) Osteoprotegerin deficiency and juvenile Paget's disease. NewEngland Journal of Medicine 347, 17584.Wozney, J.M. (2002) Overview of Bone Morphogenetic Proteins. Spine 27, 5258.
Erratum sheet
Page 17- line 34, Hentunen et al. 1995 not 1998Page 20- line 28, GCs not GSsPage 57- in Figure 8 (A) and publication II (manuscript), page 25- in Figure 6 (A)
(A)
Control
siRNAPDE7
H-89
siRNAPDE7 +
H-89
0
100
200
% C
hang
e
**
*** ***
Should be
(A)
hMSC
Contro
l
siRNAPDE7
siRNAPDE8A
0
250
500
750
1000
**
bALP
ng/
mg
prot
ein
Page 73 – Reference Conti and Beavo (2007) should be on page 69Page 90 – Reference Valta et al. (2006) should be on page 88