Embed Size (px)
Citation preview

Receptive-Field Organization of
Monkey Superior Colliculus
MAX CYNADER AND NANCY BERMAN
Department of Psychology, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139
A NUMBER OF recent studies suggest that the mammalian superior colliculus plays a crucial role in visually guided behavior (33, 35). Ablation experiments indicate that lesions in the hamster superior colliculus can cause severe deficits in orienting to visual and auditory stimuli while leaving intact the ability to discriminate pat serns in situ .ations in whi ch no orienting com- ponent is necessary for the successful per- formance of the discrimination (32). Re- lated to this view is the idea that the col- liculus plays an important role in the con- trol of eye and head movements. After unilateral colliculectomy in the monkey, Denny-Brown (6) reports a fascinating cluster of eye-movement deficits. While the animal could look to either side or up and down, there was a slight divergent stra- bismus, and the optokinetic response was present only when the stripes entered the visual field from the operated side. Though the monkey could reach accurately for an object in the visual field contralateral to the lesion, he could not fixate the object, but only looked in its general direction. After bilateral colliculus lesions, monkeys showed fixed gaze, lack of reactivity to visual stimuli; lack of vocalization, and a variety of other deficits which led Denny-Brown to describe this structure as the “pattern setter of the nervous system.” Though other workers have failed to confirm the oculo- motor deficits reported in this study (2, 28), recent work usi *g more sophisticated methods for head immobilization and eye- movement recording has confirmed and ex- tended many of Denny-Brown’s original observations (M. Stryker, P. H. Schiller, and F. Koerner, unpublished observations). It
Received for publication July 6, 1971.
has long been known that stimulation of the colliculus results in head and eye move- ments (1, 3, 4, 12), and a recent study in alert monkeys indicates that saccadic eye movements of particular sizes and directions can be elicited by stimulation in different zones in the colliculus (29).
An understanding of the mechanisms subserving these postulated functions re- quires knowledge of the transformations of the visual input occurring at the superior colliculus. Several recent reports have focused on the receptive-field organization of the tectum opticum in carnivores, am- phibia, birds, and lower mammals (10, 20, 24, 37). Generally, units in the tectum re- spond well to moving stimuli and display little specificity for different stimulus shapes or orientations. ln the cat and ground squirrel many units show strong preferences for a particular direction of stimulus move- ment (26, 36), but these directional pref- erences are rare in colliculus units of other species (13, 18, 24). In the deeper layers of the colliculus, units responding well to novel stimuli and giving weaker responses on repeated stimulus presentation have been observed; multimodal responsivity has also been observed in cells in the deeper layers. However, few detailed reports have dealt with the response characteristics of single cells in the primate colliculus (18,
21) The object of this study, therefore, was
to describe the response characteristics of single units in the superior colliculus of the rhesus macaque (Macaca muEatta) and to study the topographic organization and functional subdivisions of this structure. Data from two cats are included to facili-

188 M. CYNADER AND N. BERMAN
tate comparison with a species which has been extensively studied (8, 25, 34, 36, 37).
MATERIALS AND METHODS
Eleven adult rhesus monkeys, weighing 3.0- 7.0 kg, and two adult cats were used, The animals were anesthetized with intravenous 5% thiopental sodium (Pentothal, Abbot) and light anesthesia was maintained throughout the sur- gical procedure. An endotracheal cannula was inserted and the head was placed in a stereo- taxic frame with raised eye and ear bars to allow a full field of vision, A trephine hole was cut in the skull. above the superior colliculus, and EEG electrodes were implanted over the contralateral visual cortex. The animal was paralyzed with intravenous gallamine tri- ethiodide (Flaxedil, Davis and Geck) at a high rate of injection initially and then maintained on a solution of gallamine (38.8 mg/hr) and 5% lactated dextrose in Ringer solution (7.4 ml/hr). The Pentothal was discontinued and the animal was artificially respired, using a Harvard ap- paratus 607 respiration pump, with a mixture of 60% nitrous oxide and 40y0 oxygen. Recta1 temperature was kept at 38.8 C with a heating pad. Expired CO, was monitored by infrared absorption using a Beckman physiologic gas analyzer and kept between 3.5 and 4.0%. Pupils were dilated with 0.25y0 scopolamine hydro- bromide (Isoptohyoscine, Alcon). A slit retino- scope was used to determine the correct lenses for focusing the eyes on a tangent screen at a distance of 57 inches from the eyes. Contact lenses of appropriate power were then placed on the cornea1 surfaces and the optic discs and foveas were mapped onto the screen with a reversing ophthalmoscope. The contact lenses were removed and the eyelids taped closed for 4 hr every night to allow oxygenation of the corneas. Some animals were maintained for 5 days, but almost aI1 of the data reported here are from animals maintained for 3 days.
The dura was incised and a 19-gauge needle containing a glass-coated platinum iridium microelectrode (41) was inserted at a loo angle from the Horsley-Clarke vertical in some pre- parations, or at a ZOO angle in others, to a point stereotactically determined to be 4 mm above the superior colliculus. Conventional amplifica- tion procedures were used, and the units and EEG were displayed on a Tektronix type 565 oscilloscope.
Receptive-field locations were determined us- ing a hand projector and were mapped onto the tangent screen for each eye separately. Back- ground illumination (3.5 cd/m2) was sufficient for low photopic conditions, and the stimuli
were 1-1.5 log units above threshold. Stimuli could be moved across the receptive field by driving a mirror galvanometer with a Wavetek 912 function generator. The function generator could also trigger a computer of average tran- sients (Mnemotron) which was used to produce poststimulus time histograms (9). The unit and Wavetek signal were recorded on magnetic tape for later analysis.
To correlate receptive-field positions in the visual field with recording positions in the col- liculus, some electrode tracks were marked with lesions made by passing 5 pa of current through the electrode tip for IO sec. At the end of the experiment, the animal was perfused with 0.9% saline followed by 10% formalin. The brain, fixed in 10% formalin, was aIIowed to sink in 30y0 sucrose in 10% formalin, embedded in albumin, and 30-p frozen sections were cut in the Horsley-Clarke plane. One brain was cut in the sagittal plane. The sections were stained with cresyl fast violet and recording positions were verified. A reconstruction of the superior colliculus, viewed from above, was constructed by measuring its perimeter on every 5th frontal section through the structure.
The procedure for cats differed from that already described for monkeys in the following ways: the cats were paralyzed by continuous infusion of a mixture of tubocurarine chloride (Squibb) at; a rate of 2.8 mg/hr, gallamine tri- ethiodide (X4 mg/hr), and 5% lactated dextrose in Ringer (3.4 ml/hr) (5). The nicitating mem- branes were retracted with lO% phenylephrine hydrochloride (Neosynephrine, Winthrop) and the pupils were dilated with O.Z5a/, scopolamine hydrobromide. Cats were maintained under N,O anesthesia for 36 hr.
RESULTS
Recefhx-field organization uf monkey superior colliculus
A total of 290 units was studied in 31 electrode penetrations in 11 monkeys. Units were identified by constancy of size and shape of their electrical waveform. On the first day of recording, the animal’s EEG showed periods of slow-wave activity inter- spersed with low-voltage fast activity. As the experiment proceeded, the periods of fast activity became less frequent and after about 30 hr of recording, the EEG showed only slow-wave activity. As the electrode penetrations were not normal to the surface of the colliculus, no conclusions about columnar organization are possible from these experiments. It was found, however,
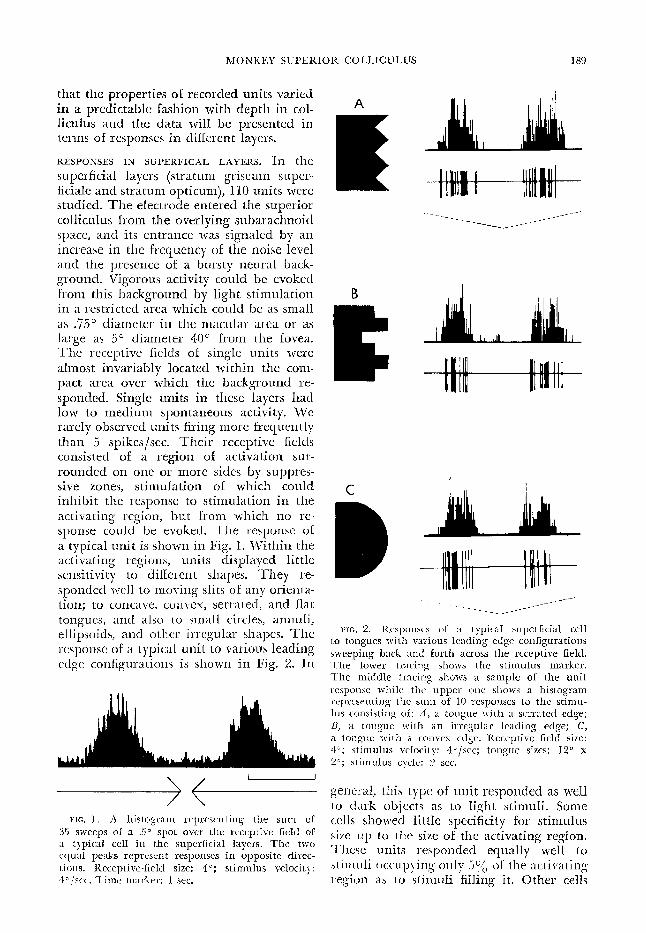
that tk properties of recorded units varied in a predictable fashion with depth in col- liculus and the data will be presented in terms of responses in different layers.
RESPONSES IN SUPERFICAL LAYERS. hl the
superficial layers (stratum griseum super- ficiale and stratum opticum), 1X0 units were studied. The electrode entered the superior colliculus from the overlying subarachnoid space, and its entrance was signaled by an increase in the frequency of the noise level and the presence of a bursty neural back- ground. Vigorous activity could be evoked from this background by light stimulation in a restricted area which could be as small as .%5G diameter in the macular area or as large as 5” diameter 40° from the fovea. The receptive Gelds of single units were almost invariably located within the com- pact area over which the background rc- sponded. Single units in these layers had low to medium spontaneous activity. We rarely observed units firing more frequently than 5 spikes/set. Their receptive fields consisted of a region of activation sur- rounded on one or more sides by suppres- sive zones, stimulation of which could inhibit the response to stimulation in the activating region, but from which no re- sponse could be evoked. Tile response of a typical unit is shown in Fig. X. Within the activating regions, units displayed little sensitivity to different shapes. They re- sponded ~~11 to moving slits of any orienta- tion; to concave, comcx, serrated, and Rat tongues, and also to small circles, annuli, ellipsoids, and other irregular shapes. The response of a typical unit to various leading edge configurations is shown in Fig. 2. 111
nn FiG, 1 . A hislogrnm rcprcscn t ing the sum of
35 sweeps of a ..!I* spot over the ruccptive field of ;t Iypicd cell in the superficial layers. The two cquaI peaks represent responses in opposite dircc- Lions. Receptive-field size: 40; stimulus velocity: 4 o ,krc~. Time marker: 1 sec.
A
E
6
II
C
B
FIG. 2. Kcsp”nS”S of a typical. superficial cell to tongues rG t 31 various leading edge configurations sweeping back and forth across the rcccptive field. The lower tracing shows the stimulus marker. The middle tracing shows a sample of the unit: wsponse while the upper one shows a histogram wprcscnting the sum of IO responses to the stimu- 10s consisting of: A, a tongue with a scrratcd edge; B, a tongue with an irregular leading edge; C, a tongue tz-i t 11 a convex edge. lieccptive field size: 4”; stimulus velocity: 4o/sec; tongue sizes: 12” x 2”; stimuIus cycle: 2 see.
gerwal, this type of- ulzit responded as well to cl;Irk objects as to light stimuli. Some cells showed little specificity for stimulus s.iLe up to the size of the activating region. These units responded equally well to st.irnrrli ocr:lrpyi ng only 5::: oC the activating x&n as to stimuli fil1in.g it. Other cells
190 M. CYNADER AND N. BERMAN
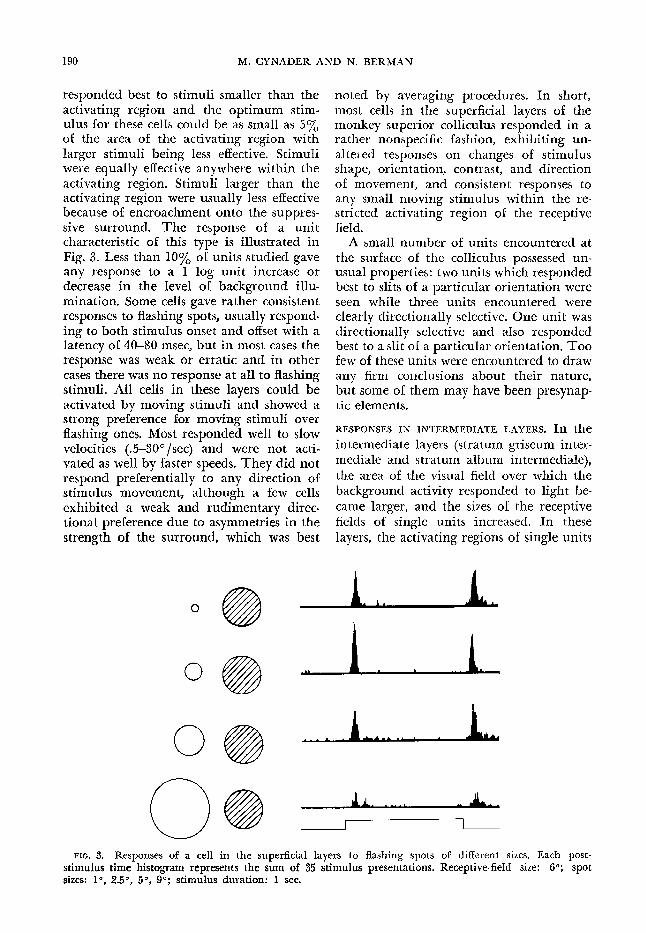
responded best to stimuli smaller than the activating region and the optimum stim- ulus for these cells could be as small as 5% of the area of the activating region with larger stimuli being less effective. Stimuli were equally effective anywhere within the activating region. Stimuli larger than the activating region were usually less effective because of encroachment onto the suppres- sive surround. The response of a unit characteristic of this type is illustrated in Fig. 3. Less than 10% of units studied gave any response to a 1 log unit increase or decrease in the level of background illu- mination. Some cells gave rather consistent responses to flashing spots, usually respond- ing to both stimulus onset and o&et with a latency of 40-W msec, but in most cases the response was weak or erratic and in other cases there was no response at all to flashing stimuli. All cells in these layers could be activated by moving stimuli and showed a strong preference for moving stimuli over flashing ones. Most responded well to slow velocities (.5-3P/set) and were not acti- vated as well by faster speeds. They did not respond preferentially to any direction of stimulus movement, although a few cells exhibited a weak and rudimentary direc- tional preference due to asymmetries in the strength of the surround, which was best
noted by averaging procedures. In short, most cells in the superficial layers of the monkey superior colliculus responded in a rather nonspecific fashion, exhibiting un- altered responses on changes of stimulus shape, orientation, contrast, and direction of movement, and consistent responses to any small moving stimulus within the re- stricted activating region of the receptive field.
A small number of units encountered at the surface of the colliculus possessed un- usual properties: two units which responded best to slits of a particular orientation were seen while three units encountered were clearly directionally selective. One unit was directionally selective and also responded best to a slit of a particular orientation. Too few of these units were encountered to draw any firm conclusions about their nature, but some of them may have been presynap- tic elements.
RESPONSES IN INTERMEDIATE LAYERS. In the intermediate layers (stratum griseum inter- mediale and stratum album intermediale), the area of the visual field over which the background activity responded to light be- came larger, and the sizes of the receptive fields of single units increased. In these layers, the activating regions of single units
0
FIG. 3. Responses of a cell in the superficial layers to flashing spots of different sizes. Each post- stimulus time histogram represents the sum of 35 stimulus presentations. Receptive-field size: 6O; spot sizes: 10, 2.50, 50, go; stimulus duration: X sec.
MONKEY SUPERIOR COLLICULUS 191
ranged from 1 to 70’ in diameter. Their shapes were again circular, ellipsoid, or in some instances approximately rectangular. The spontaneous activity of these cells was usually low. Most cells fired only a few times per minute. These cells gave weaker responses on repeated presentation of sub- optimal stimuli. We were able to compare the effectiveness of various stimuli by deter- mining the number of presentations re- quired for the response to disappear to each of them as well as by examining the strength of the intial response. These two measures showed a high positive correlation.
Like the cells in the superficial layers, cells in the intermediate 1 ayers were insen- sitive to details of stimulus shape or orien- tation. About 20% of the units studied in the intermediate and deep layers responded at least once to a I log unit change in dif- fuse illumination. About 30% of the units studied in the intermediate and deep layers appeared to be truly nonspecific and could best be described as “newness” or “novelty” detectors (23). They responded best to novel stimuli moving through the receptive field or appearing suddenly within- the field. Subsequent presentations of the same stimulus would evoke successively weaker responses. Repeated stimulation of one eye resulted in weaker responses from either
eye. The rate of response decrement on re- peated stimulus presentation varied with depth in the colliculus. At the transition point between the superficial and deeper layers, it manifested itself merely as a slight fat&ability which made it necessary to wait while mapping the receptive field. As the electrode was advanced, the response decre- ment on repeated stimulus presentation be- came so strong that a new stimulus rarely evoked more than one or two responses. Still deeper, cells were sometimes encoun- tered which, though still rather nonspecific, exhibited less rapid response decrement than those encountered previously. No clear-cut relationship between the cell’s re- sponses and the state of the EEG could be shown since after about 30 hr of recording, the EEG of the animal was always syn- chronized.
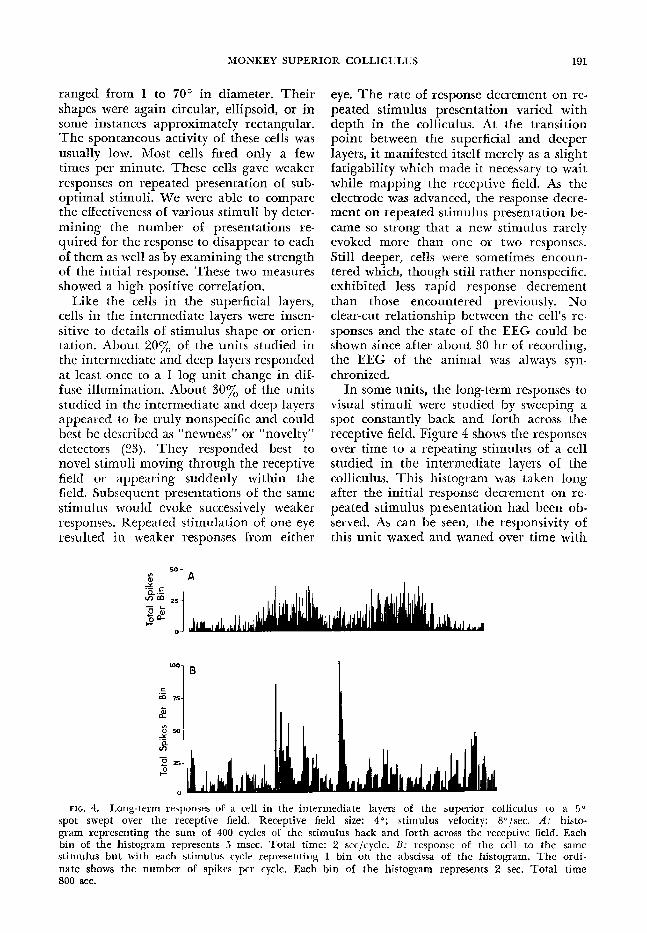
In some units, the long-term responses to visual stimuli were studied by sweeping a spot constantly back and forth across the receptive field. Figure 4 shows the responses over time to a repeating stimulus of a cell studied in the intermediate layers of the colliculus. This histogram was taken long after the initial response decrement on re- peated stimulus presentation had been ob- served. As can be seen, the responsivity of this unit waxed and waned over time with
*a t .- mm 25
0 !
L
= 75-
d
G 50- Y .-
3
2 25-
s
FIG. 4. Long-term responses of a cell in the intermediate layers of the superior colliculus to a 5” spot swept over the receptive field. Receptive field size: 4O; stimulus velocity: 8”/sec. A: histo- gram representing the sum of 400 cycles of the stimulus back and forth across the receptive field, Each bin of the histogram represents 5 msec. Total time: 2 secjcycle. B: response of the cell to the same stimulus but with each stimulus cycle representing 1 bin on the abscissa of the histogram. The ordi- nate shows the number of spikes per cycle. Each bin of the histogram represents 2 sec. Total time 800 sec.

192 M. CYNADER AND N. BERMAN
the unit occasionally responding to the stimulus as well as it had on the first pre- sentation. The spontaneous activitv of this unit was only a fkw spikes per min;te. His- togram B, therefore, represents variation in the unit’s responsiveness over time, and it cannot be accounted for bv variation in spontaneous activity.
Intermingled with these nonspecific cells in the intermediate layers, two other types of cells were found which resDonded to more specific aspects of the vi&I stimulus. One class of cells differed from all others studied by responding better to flashing stimuli than to smooth movement. The optimum stimulus for these cells, however, was a short jerky movement within the receptive field. Most of these cells re- sponded both to stimulus onset and offset, but a few gave stronger on- or off-responses. A cell which responded more strongly to on than to off when a flashing stimulus was used would respond better to the jerky stimulus entering the receptive field than to leaving it, and cells giving larger off-re- sponses responded better to the stimulus leaving the receptive field than entering it. These cells also possessed suppressive sur- rounds and responses were weaker to stimuli larger than the receptive field. Stimuli much smaller in size than the acti- vating region were usually the optimum stimulus. These cells were insensitive to de-
tails of stimulus shape or orientation as well as to direction of movement. Figure 5 shows the activity of a typical unit during smooth, sinusoidal, and jerky movement within the receptive field. While giving little or no response to the smooth move- ments, the unit followed each jerky move- ment of the stimulus. Some of these cells exhibited little or no response decrement to their optimum stimulus, but others ex- hibited erratic responsivity to the jerky stimulus. Some of these cells would begin responding to the jerky stimulus only after a few stimulus cycles. During a continuous presentation of the jerky stimulus, these cells would respond to every stimulus move- ment for several cycles, then would not respond for a few cycles, and later would again begin responding to the jerky move- ment.
Another group of cells in these laminae was optimally activated by a shadow tongue entering an illuminated area. There was no effect of differently shaped tongues, but light tongues, edges, slits, spots, annuli, and all other stimuli were far less effective in exciting these cells. These cells often had extremely strong suppressive surrounds and the size of the illuminated area into which the shadow tongue entered was often quite critical. Illuminating an area larger than the activating region in these cells resulted in much poorer responses to the shadow
FIG. 5. Responses of a unit in the intermediate layers to a Jo spot moved at a constant velocity (A), sinusoidally (B), and in a jerky manner (C) within the activating region. Receptive-field size 60; stim- lus excursion 40 in all cases; cycle length 500 msec.

tongue. The illuminated area could be smaller than the receptive field, but illu- minated areas less than one-tenth the area of the activating region usually gave poorer responses,
Other units fell into transition groups between these two types of cells and the novelty-detecting cells. We found cells which responded well to jerky stimulus movement and to entering shadows, but poorly to other stimuli; other cells were quite unspecific in their responses, habit- uating to all stimuli, but exhibiting some- what less rapid habituation to shadow tongues entering illuminated areas. 0 ther cells responded well to smooth slow move- ment initially, but this response rapidly weakened leaving a much more constant response to jerky movement. Approxi- mately one-sixth of all cells studied in the intermediate and deep layers could be con- sidered to fall into one of these transition groups, while about one-sixth of the cells in the intermediate and deep layers fell into each of the specific categories.
RESPONSES IN DEEP LAYERS. Within the deep layers (1.7-3.0 mm) (stratum griseum pro- fundum and stratum album profundum), the visual background became still more spread out and tactile, vibratory, or audi- tory stimuli could sometimes elicit re- sponses from the background. Most units which were encountered responded to a wide variety of large dark objects, giving, in general, poorer responses to light stimuli. These units were binocularly driven, gave weaker responses on repeated stimulus presentation, and had still larger receptive fields than cells in the superficial and intermediate layers. Some cells in the deep layers were subliminally facilitated by auditory stimulation occurring simulta- neously with the visual stimulus. Of all cells studied in the intermediate and deep layers, 8% were clearly multimodal and re-
sponded to visual and tactile stimuli, visual and auditory (or vibratory), or to stimula- tion of all three modalities. These cells gave weaker responses to repeated stimuli, and in two cases we could demonstrate that responses to repeated stimuli could be weakened independently in the three modali ties,
Ten cells studied in the deep layers re- sponded to stimuli in a manner which resembled that of the superficial cells. They exhibited no response decrement on re- peated stimulus presentation, responded well to flashing spots, to jerky and smooth movement, to all stimulus shapes and orientations, and equally to light stimuli and dark stimuli, Their receptive-field sizes were, however, much larger than those of the superficial cells, ranging in size from 3 to 40° in diameter.
About 10% of the cells found in the in- termediate and deep layers exhibited a strik- ing afterdischarge to various types of novel stimuli. This afterdischarge, which could last as long as 10 set, could rarely be evoked from a given cell more than a few times. Figure 6 shows the response of a cell of this type to a stimulus appearing suddenly within the receptive field and then remain- ing fixed in position.
DIRECTIONAL SELECTIVITY AND OCULAR DOM-
INANCE, Directional selectivity was observed in 5% of all cells studied. Cells exhibiting this property were either at the very surface of the colliculus or were scattered through- out the intermediate and deep layers. There was no clear arrangement of the preferred directions of these cells relative to the fovea such as that found in the cat (36, 37). These units were also binocularly driven and pos- sessed no clear orientation or shape speci- fici ty except for the one unit noted previously. It was noted that three of these units had their receptive fields located far from the area from which background ac-
MONKEY SUPERIOR COLLICULUS 193
I J
FIG. 6. Response of an afterdischarging unit in the intermediate layers to the sudden appearance of a novel stimulus within its receptive field. Note the long duration of the response although the stimuIus remained stationary after its initial appearance, Time marker: 1 sec. Triangle denotes stimu- lus onset.

194 M. CYNADER AND N, BERMAN
tivity could be evoked, an unusual property in nondirectional cells. This raises the possibility that some of these units are not of collicular origin. Figure 7 shows the
encountered cells which had nonoverlap- ping receptive fields, a very rare phe- nomenon in the monkey. As in- the monkey, the receptive fields of individual units con- sisted of activating regions flanked by sup- pressive surround areas which were asym- metric in strength in some cases. In the cat, the receptive fields were much larger than those of the monkey for the same retinal locus relative to the area centralis. As in the monkey, receptive fields increased in size with increasing depth in the superior colliculus. The activating regions were often elongated in cats, tending to be ellip- soid or approximately rectangular in shape, whereas those of the monkey were usually rounder in shape. These cells were also insensitive to small differences in stimulus shape or orientation and some were also optimally activated by stimuli much smaller than the activating region. The ocular dominance distribution of 25 cells studied in one cat is shown on the right-hand side of Fig. 7. This distribution differs from that of the monkey, where virtually all cells were activated equally by both eyes, in showing a greater spread of ocular domi- nance. Directional selectivity was observed in 68% of all cells studied in cats, and as noted by others (36, 37), the optimal direc- tion of stimulus movement for most cells
ocular dominance distribution of cells in the colliculus of the cat and monkey. As can be seen, all cells within the arka of binocular overlap were driven well bY either eye in monkeys. Receptive fields were located in approximately points in both retinas, but the
homologous special pre-
cautions necessary to prevent residual eYe movements (5) were not taken.
Receptive-field organization of cat cblliculus
Forty-eight cells were studied in seven penetrations in two cats. The area of the visual field over which background activity could be evoked was much less compact in the cat than in the monkey, and -it was of ten possible to record from successivelv
7%
50-
251
180
I
Monkey
24 3
I ’ I
2 3 4 I
5 ,
6 7 Equal lpsilateral y Contralateral c was awav from the area centralis.
In the deeper layers we also noted direc- tionally selective cells, some of which gave weaker responses to repeated stimuli. Un- like our experience in the monkey, we found no cells which exhibited strong pref- erences for jerky movement nor did we ob- serve cells specifically activated by shadow tongues entering illuminated areas.
Cat I6
25
I ( LontraLeral
I I
I I I 4 5 6 7
Equal lpsilateral y Topographic wganization
In one monkey, 15 penetrations were made through the colliculus, and the rela- tionship between receptive-field location in the visual field and unit position on the colliculus surface was established. If no unit was isolated within .3 mm from the surface, the center of the area of the visual field over which light stimulation evoked responses from the neural background was used to establish the location in the visual field. The fovea and optic disc were mapped on the tangent screen several times during
FIG. 7. Top of the figure indicates the ocular dominance distribution for 207 cells in the monkey colliculus and the lower part the distribution for 25 cells in the cat colliculus. The number on top of each bar gives the absolute number of cells in each category, The categories correspond to those defined by Hubel and Wiesel (15) Groups l-7 represent a contralateral to ipsilateral trend with cells in group I totally dominated by the contra- lateral eye, cells in group 4 being driven equally by both eyes, and cells in group 7 driven exclu- sively by the ipsilateral eye. Cells with receptive fields outside the area of binocular overlap are not included in this figure.
MONKEY SUPERIOR COLLICULUS
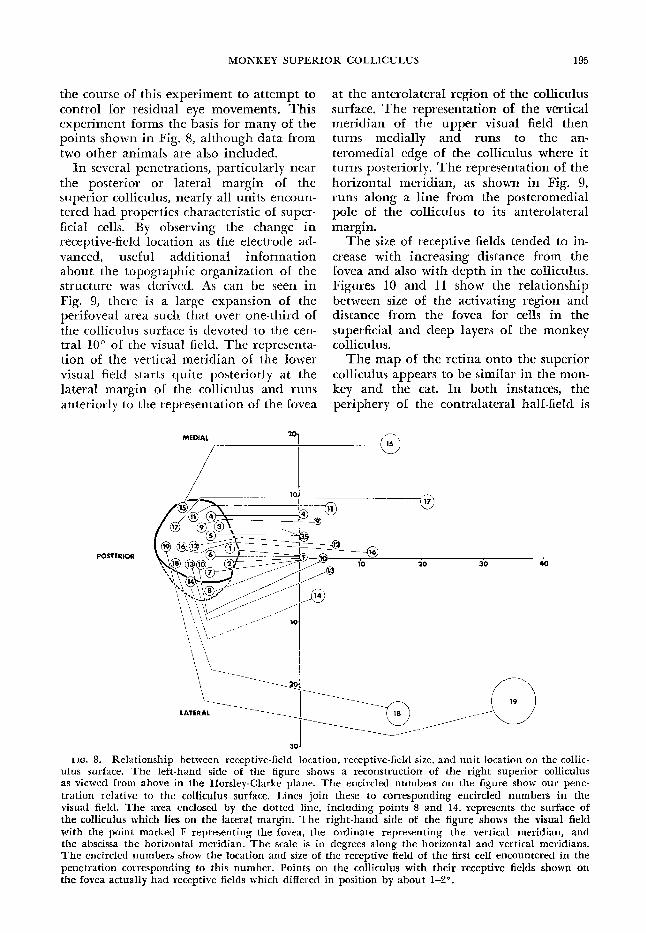
the course of this experiment to attempt to control for residual eye movements. This experiment forms the basis for many of the points shown in Fig. 8, although data from two other animals are also included.
In several penetrations, particularly near the posterior or lateral margin of the superior colliculus, nearly all units encoun- tered had properties characteristic of super- ficial cells. By observing the change in receptive-field location as the electrode ad- vanced, useful addi tionaf information about the topographic organization of the structure was derived. As can be seen in Fig. 9, there is a large expansion of the perifoveal area such that over one-third of the colliculus surface is devoted to the cen- tral 10” of the visual field. The representa- tion of the vertical meridian of the lower visual field starts quite posteriorly at the lateral margin of the colliculus and runs anteriorly to the representation of the fovea
MEDIAL 20
1
at the anterolateral region of the colliculus surface. The representation of the vertical meridian of the upper visual field then turns medially and runs to the an- teromedial edge of the colliculus where it turns posteriorly. The representation of the horizontal meridian, as shown in Fig. 9, runs along a line from the posteromedial pole of the colliculus to its anterolateral margin.
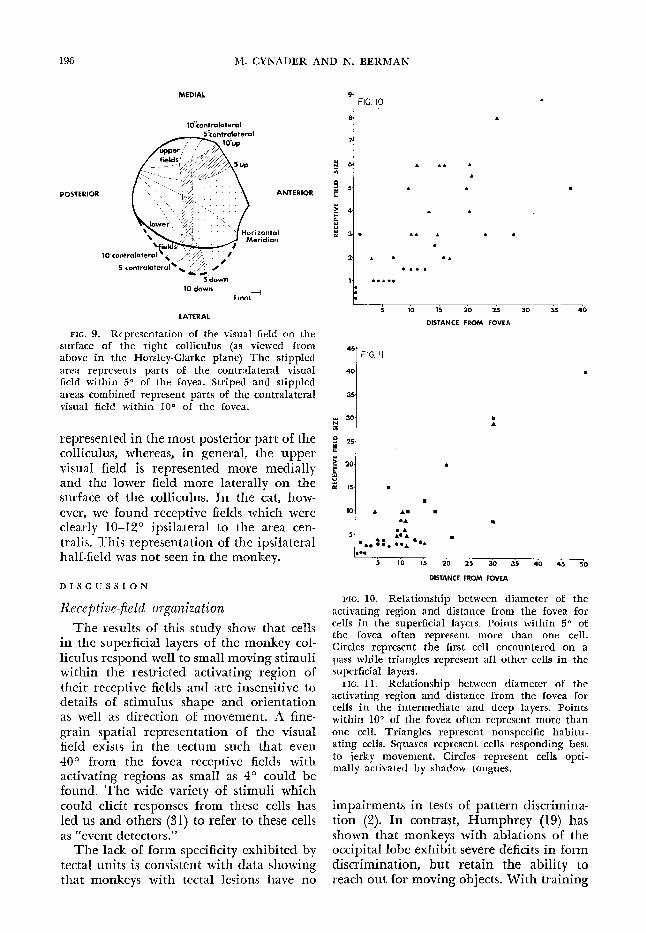
The size of receptive fields tended to in- crease with incre& ing distance from the fovea and also with depth in the colliculus. Figures 10 and II show the relationship between size of the activating region and distance from the fovea for cells in the superficial and deep layers of the monkey co113 culus.
The map of the retina onto the superior colliculus appears to be similar in the mon- key and the cat. In both instances, the periphery of the contralateral half-field is
POSTERtOR
. . ..>.1:: 30
FIG. 8. Relationship between receptive-field location, receptive-field size, and unit location on the collic- ulus surface. The left-hand side of the figure shows a reconstruction of the right superior colliculus as viewed from above in the Horsley-Clarke plane. The encircled numbers on the figure show our pene- tration relative to the collicuIus surface. Lines join these to corresponding encircled numbers in the visual field, The area enclosed by the dotted line, including points 8 and 14, represents the surface of the colliculus which lies on the lateral margin, The right-hand side of the figure shows the visual field with the point marked F representing the fovea, the ordinate representing the vertical meridian, and the abscissa the horizontal. meridian. The scale is in degrees along the horizontal and vertical meridians. The encircled numbers show the location and size of the receptive field of the first cell encountered in the penetration corresponding to this number. Points on the colliculus with their receptive fields shown on
the fovea actually had receptive fields which differed in position by about 14?.
196 M. CYNADER AND N. BERMAN
9 1 FIG. IO
POSTERIOR
IO’contralateral
5htmkteral
1 O’down - --. .+ Imm
LATERAL
,NTERIOR
FIG. 9. Representation of the visual field on the surface of the right colliculus (as viewed from above in the Horsley-Clarke plane) The stippled area represents parts of the contralateral visual field within 5” of the fovea. Striped and stippled areas combined represent parts of the contralateral visual field within HP of the fovea.
represented in the most posterior part of the colliculus, whereas, in general, the upper visual field is represented more medially and the lower field more laterally on the surface of the colliculus. In the cat, how- ever, we found receptive fields which were clearly 1612” ipsilateral to the area cen- tralis. This representation of the ipsilateral half-field was not seen in the monkey.
8 1
AA
A
A
AA A l
5 10 15 20 25 30 3s 40
DISTANCE FROM FOVEA
“‘1 FIG. I I
40
I 354
Am 5 IO I5 20 25 30 35 40 45 50
Receptive-field organization
The results of this study show that cells in the superficial layers of the monkey col- liculus respond well to small moving stimuli within the restricted activating region of their receptive fields and are irken&ive to details of stimulus shape and orientation as well as direction of Imovement. A fine- grain spatial representation of the visual field exists in the tectum such that even 40” from the fovea receptive fields with activating regions as small as 4’ could be found. The wide variety of stimuli which could elicit responses from these cells has led us and others (31) to refer to these cells as “event detectors.”
The lack of form specificity exhibited by tectal units is consistent with data showing that monkeys with tectal lesions have no
l
DISTANCE FROM FOVEA
DISCUSSION
FIG. IO. Relationship between diameter of the activating region and distance from the fovea for cells in the superficial layers. Points within 50 of the fovea often represent more than one cell, Circles represent the first cell encountered on a pass while triangles represent a11 other cells in the superficial layers.
FIG. 11. Relationship between diameter of the activating region and distance from the fovea for cells in the intermediate and deep layers. Points within 100 of the fovea often represent more than one cell. Triangles represent nonspecific habitu- ating cells. Squares represent cells responding best to jerky movement. Circles represent cells opti - mally activated by shadow tongues.
impairments in tests of pattern discrimina- tion (2). In contrast, Humphrey (19) has shown that monkeys with ablations of the occiGta1 lobe exhibit severe deficits in form dis&mination, but retain the ability to reach out for moving objects. With training

MONKEY SUPERIOR COLLICULUS 197
and practice the capacity to localize even stati onary objects improves, but there is no evidence of recognition of objects by sight. Here again, the distinction between localiz- ing and identifying systems can be made (33). It seems possible that the fine-grain representation of the retina on the tectal surface provides the substrate for object localization which survives occipital cortex ablation. Behavioral experiments in mon- keys involving serial cortical and tectal lesions might bear on this hypothesis.
In the deeper layers some of the units studied appeared to be true newness neu- rons, giving weaker responses to repeated presentation of any stimulus. Response decrement to repeated stimuli has been ob- served in tectal neurons in several species (14,23, 37). In cats and rabbits, neurons with these properties are reported to occur in the deeper layers of the colliculus, and thev tend to be associated with multimodal responsivity. Units related to eye or head movements have been found in these lavers (30, 34, 38,42) and it may be of interest to try to conceive of the newness units more in terms of their possible relation to the oculomotor system- than as purely sensory cells. As an example, we may consider the hypothetical results of an experiment in which one attempted to map receptive fields of single units in the oculomotor nuclei of a paralyzed animal. These units would presumably fire when he attempted to move his eyes. A description of their re- ceptive-field characteristics would probably inilude multimodal responsivi ty, strong habituation to repeated stimulus presenta- tion, and perhaps, in abducens, the oculo- motor nucleus associated with lateral eye movements, the units would exhibit direc- tional selectivity with the preferred direc- tion away from the midline.
In the monkey a clear laminar dissocia- tion between these nonspecific newness neu- rons and units with multimodal responsivity is evident. Weakened responses to repeated stimulus presentation can be seen in the intermediite layers, whereas multimodal. re- sponsivity is confined to the deep layers. We believe that our newness units are closely related to units found in the inter- mediate and deep layers of the colliculus by Schiller and Koerner (31). They found a
population of units which discharge before eye movements and have visual receptive fields. These units discharge when an eye movement occurs which results in brin e% a target placed in the receptive field of the unit to the fovea.
We have also found other units exhibit- ing more specific responses to visual stimuli in- the intermediate layers of the colliculus. Some cells responded well to jerky stimulus moveme nt while others responded well to shadow tongues entering an illumina ted area. We suggest that these units extract stimulus features which could be useful to a visual attention and eye-m ovement mech- anism. The presence of these units in the same collicular laminae as eye-movement- related units reinforces this possibility (3 1).
There are some interesting parallels be- tween our results and those of Lettvin and co-workers (23), who have studied the re- sponses to visual stimuli of single units in the frog optic tectum, These workers report that in the deep layers of the tectum, where the cell bodies- are located, there are new- ness neurons which resemble, in many ways, some of the cells which we encountered in the intermediate and deep collicular layers of the monkey. The other category of cells found by this group, the “sameness” neu- rons, exhibits the property of “noticing” a stimulus and then responding to every jerk or movement of the stimulus with a burst of impulses. These cells bear a resemblance to some of our jerky-movement units in the monkey, which also seemed to lock onto the jerky stimulus and follow its every move- ment for a period of time.
Comj!hrison of responses monkev colliculi
in cat and
Collicular cells in these two species ex- hibit many similarities. In both cases, the receptive fields consist of a homogeneous activating region flanked by a surround which can inhibit res ponses to the center, but from which no responses can be evoked (36). In both cases, units respond best to moving stimuli, are insensitive to details of stimulus shape or orientation, and often respond well to stimuli much smaller than the size of the activating region. In both species the deeper lavers contain cells which

198 M. CYNADER AND N, BERMAN
habituate to repeated stimulus presenta- tions (37).
The major differences between the re- sponse properties of superior colliculus units in the two species were the strong direc- tional specificity exhibited by about 67% of the cat colliculus cells versus 5% in the monkey and the absence of cells in the cat responding specifically to shadow tongues entering illuminated areas or to jerky stim- ulus movement. Some of these differences may be related to the ecological niches oc- cupied by these species. It seems reasonable that the arboreal diurnal primate might be more likely to possess specific mechanisms for the detection of shadow tongues than the more nocturnal carnivore.
Other differences may be related to ac- cumulating evidence which suggests that similar stimulus features are reflected in the firing patterns of cells in different brain structures in the two species. In the cat, units optimally activated by horizontally disparate stimulation of the two retinas oc- cur in area 17 (27). In the monkey such cells do not occur in area 17, but do occur in area 18 (17). In the cat, cells responding well to jerky stimulus movement over a large area of the visual field have been described. These cells are located not in the colliculus, but in the anterior middle suprasylvian gyrus of the cortex (7).
Comparison of geniculocortical and collicular systems
Recent anatomical evidence suggests that while the monkey superior colliculus re- ceives its major visual input from the retina and from the striate and peristriate cortex, the projection of the central visual field onto the colliculus seems to be exclusively of cortical origin (40). This region occupies the anterior one-third of the collicular sur- face. The units in the monkey colliculus with receptive fields within the central vi- sual field may thus be considered to repre- sent a further step in processing of visual information by a retina-geniculate-striate cortex-colliculus system. We may thus com- pare the receptive fields in this area with those of the previous stage and see what changes have occurred in the receptive-field properties at this level. Both cortical (16) and collicular cells are binocularly driven,
but in the colliculus, the percentage of cells driven equally by the two eyes is much higher. Collicular cells do resemble “hyper- complex” cortical cells in that their recep- tive fields are not separable into discrete on and off areas, the responses to stimuli are the same independent of their loci in the receptive field, and the optimal stimuli may be much smaller than the- receptive field.
Since cortical cells respond best to lines of a particular orientation, it is surprising that collicular units whose visual input comes exclusively from cells of this type show no preference for a particular stim- ulus orientation. Similarly, in the cortex a considerable percentage of cells are reported to show directional selectivity, but this feature is rarely seen in collicular cells. Thus, the superficial collicular cells have some properties which are similar to those of cortical units, but the specificity for par- ticular stimulus shape and direction pos- sessed by cortical units is not found in the colliculus, It is surprising in light of the anatomic evidence that we have found no systematic difference between collicular cells in the perifoveal and peripheral visual fields in terms of shape specificity or direc- tional selectivity. While receptive fields in the periphery are larger, stimuli much smaller than the activating region still produce good responses. Cells in the deeper layers of the colliculus exhibit habituation to repeated stimuli, and this has not been noted in striate cortex cells.
Retinot+c organization
The projection of the various parts of the visual field onto the surface of the tec- turn has been studied in many vertebrates (3, 5, 10, 11, 20). In many species all parts of the visual field are represented equally on the surface of the tectum, but in some cases relative magnification of parts of the visual field on the surface of the tectum has been noted (20, 22). In the monkey, as in the cat, the upper visual field is represented on the more medial parts of the superior colliculus while the lower field projects to more lateral portions of this structure. The contralateral periphery is represented on the posterior part of the colliculus whereas the central field is represented on the more anterior

MONKEY SUPERIOR COLLICULUS 199
portion. In the monkey the central field (within 10° of the fovea) occupies over one- third of the surface of the tectum while the peripheral parts of the field are crammed into the remaining portion. The expansion of fovea1 and perifoveal represen- tation on the tectal surface is seen in the severe distortion in the representation of the vertical meridian
These data are in general agreement with recent anatomical work on the projection of the retina and visual cortex to the supe- rior colliculus and with older electrophysio- logic observations of the representation of the visual field in the macaque colliculus (39, 40). Interestingly, the map shown in Fig. 9 is in approximate agreement with an eye-movement map produced by s timula- tion of the colliculus if one transforms the latter from polar to rectangular coordinates (29). It has been shown that small saccades are elicited by stimulation of more poste- ior parts of this structure, Upward-directed saccades follow stimulation of the medial colliculus while lateral stimulation results in downward eye movement. The relation- ship between the two maps is consistent with the idea that stimulation of a given point in the superior colliculus results in a saccade whose length and direction are equal to the distance and direction. of the receptive fields of the stimulated cells from the fovea.
SUMMARY
The responses of single cells in the superior colliculus to visual stimuli were studied in paralyzed, N,O-anesthetized monkeys. Units in the superficial layers were binocularly driven and had receptive fields which consisted of small circular or ellipsoid activating regions surrounded by suppressive areas which could inhibit re- sponses to central stimulation, but from which no response could be evoked. These units preferred moving to flashing or sta- tionary stimuli and showed no shape or directional specificity. Kesponses to stimuli extending outside the activating region were usually weaker, and many units pre- ferred stimuli much smaller than the acti- vating region.
In the intermediate and deep layers of the colliculus, cells responded to suboptimal stimuli with habituation. About 30% of these units could be best described as “newness neurons,” giving weakened re- sponses to repeated presentation of any stimulus, but other units responded to particular aspects of the visual stimulus. One class of cells responded best to short jerky movements of the visual stimulus while another class responded preferen- tially to shadow tongues entering illumi- nated areas. Other cells fell into transition groups between these types and the non- specific habituating cells. A few cells exhibiting multimodal responsivi ty were observed in the deep layers. Directional selectivity was observed in 5% of all tectal units studied, There is a topographic map of the contralateral half-field onto the colliculus with the upper field represented medial to the horizontal meridian and lower field lateral. There is an expansion of the representation of the perifoveal re- gions on the colliculus with over one-third of the colliculus surface devoted to the central field within X0* of the fovea. The more peripheral field is represented in the posterior part of the colliculus.
In the cat, 687$ of all units studied ex- hibited directional selectivity, but in other ways showed many resemblances to monkey colliculus cells, including binocular activa- tion and insensitivity to details of stimulus shape or orientation. In the deeper layers no cells giving selective responses to jerky stimulus movement or to shadow tongues were observed.
These data are consistent with recent electrophysiologic and behavioral work re- lating the superior colliculus to visually guided behavror and oculomotor function.
ACKNOWLEDGMENTS
The authors express their gratitude to Professor Peter H. Schiller for his valuable advice at all stages of this work.
This research was supported by National Science Foundation Grant GB-17047 to Professor Schiller, by a grant from the Alfred P. Sloan Foundation, and National Aeronautics and Space Adminis- tration Grant NGR-Z-009-038 to Professor Hans- Lukas Teuber,

200 M. CYNADER AND nT. BERMAnt
REFERENCES
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
39.
ADAMUK, E. uber die Innervation der Augen- bewegungen. Zenttr. Med. Wiss. 8: 65-67, 1870. ANDERSON, K. F’. AND SYMMES, D. The superior coIliculus and higher visual functions in the monkey. Bruin Res. 13: 37-52, 1969. APTER, J. T. Projection of the retina on supe- rior colliculus of cats. J. Neurophysiol. 8: 123- 134, 1945. APTER, J. T. Eye movements following strych- ninization of Ihe superior colliculus of cats. J. Neurophysiol. 9: 73-86, 1946. CHOW, K. E. Influences of residual eye movc- merits in single unit studies of the visual sys- tem. Bruin Res. 8: 385-388, 1968, DENNY-BROWN, D. The midbrain and motor integration. Proc. Roy. SW. Med. 55: 527-538, 1962. Dow, B. M. AND DUBNER, R. Single-unit responses to moving visual stimuli in middle suprasylvian gyrus of the cat. J, Neurophysiol, 34: 47-55, 1971. I~ELDON, S., FELDON, P., AND KRUGER, L. To- pography of the retinal projection upon the superior colliculus of the cat. Vision Res. 10: 135-143, 1970. GERSTEIN, G. L. AND KIANG, N. Y,-S, An ap- proach to the quantitative analysis of electro- physiological data from single neurons. Bio- phys. Je 1: 15-28, 1960. HAMDI, F. A. AND WHITTERIDGE, D. The reprc- sentation of the retina on the optic tectum of the pigeon. Quart. J. Exptl. Physiol. 39: lH-- 119, 1954. HERXC, 71’. M. AND KRUGER, L. Organization of the visual projection upon the optic tectum of a reptile (Alligator mississipensis). J. Camp. Neurol. 124: 101-111, 1965. HESS, W. R,, BUERCI, S., AND BIJCHER, V. Motorische Funktion des Tcctal und Teg- mentalgebietes. Mona tsschr. Psychia t. Neurol. 112: I-52, 1946. I-IxLL, R. M. AND GOODWIN, H. Visual receptive fields from cells of a marsupial (DideZphis virginiana) superior colliculus. Experentia 24: 559-560, 1968. I-IoRN, G. AND HILL, R. M. Responsiveness to sensory stimulation of units in the superior colliculus and subjacent tectotcgmental regions of the rabbit. ExptE. Neurol. 14: 199-223, 1966. HUBEL, D. H. AND WIESEL, T. N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J. Physiol., London 160: 106-154, 1962. HUBEL, D. H. AND WIESEL, T. N. Receptive fields and functional architecture of monkey striate ::ortex. J. Physiol., London 195: 215-243, 1968. HUBEL, D. H. AND WIESEL, T. N. Stereoscopic vision in macaque monkey. Nuttdre 225: 41-44, 1970, HUMPHREY, N. K. Responses to visual stimuli of units in the superior colliculus of rats and monkeys. Exptl. Neural. 20: 312-340, 1968. HUMPHREY, N. K. FZrhat the frog’s eye tells the
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36,
monkey’s brain. Brain Beha-u. Evol. 3: 324-337, 1970.
JACOBSON, M. The representation of the retina on the optic tectum of the frog. Correlation between retinotectal magnification factor and retinal ganglion cell count. Quart. J, Exptl. Physiol. 57: 170-178, 1962. KADOYA, S., WOLIN, L., AND MASSOPUST, L. Photically evoked unit activity in the tectum opticum of the squirrel monkey. J. Camp. New-ok 142: 495408, 1971. KRUGER, L. The topography of the visual projection to the mesencephalon: a compara- tive survey. Brain Behav. Evol. 3: 169-177, 1970. LETTVIN, J. Y., MATURANA, H. R., PITTS, W. H., AND MCCULLOCH, W. S. Two remarks on the visual system of the frog. In: Sensory Com- munication, edited by W, A. Rosenblith. New York: Wiley, 1961. MASLAND, R. H., CHOW, K. C., AND STEWART, D. C. Receptive-field characteristics of superior colliculus neurons in the rabbit. J. Neuro- physiol. 34: 148-156, 1971. MCILWAIN, J. F. AND BUSER, P. Receptive fields of single cells in the cat’s superior colliculus. Exptl. Brain Res. 5: 314-325, 1968. MICHAEL, C. R. Integration of retinal and cortical information in the superior colliculus of the ground squirrel. Brain Behav. Evol. 3: 205-209, 1970. NIKARA, T., BISHOP, P. O., AND PETTIGREW, J* D. Analysis of retinal correspondence by studying receptive fields of binocular singIe units in cat striate cortex. Exptl. Brain Rss. 6: 353-372, 1968. PASIK, T., PASXK, P., AND BENDER, M. B. The superior colliculi and eye movements. Arch l
Neurol. 15: 420-436, 1966. ROBINSON, D. A. Eye movements evoked by superior colliculus stimulation in the alert monkey. Meeting Assoc. Res. Vision Uph- thalmoI, Sarasota, Fla., 19X SCHAEFER, K. P. Unit analysis and electrical stimulation of the optic tectum of rabbits and cats. Bruin Behav, Evol. 3: 222-240, 1970. SCHILLER, P. H. AND KOERNER, F. Discharge characteristics of single units in superior COT- liculus of the alert rhesus monkey. J. Neuro- physiol. 34: 920-936, 1971, SCHNEIDER, G, IX:, Contrasting visuomotor func- tion of tectum and cortex in the golden hamster. Psychol. Forsch. 31: 52-62, 1967. SCHNEIDER, G. E. Two visual systems. Science 163: 895-902, 1969. SPRAGUE, J. M., MARCHIAFAVA, P. L., AND &ZZOLAT'rI, G. Unit responses to visual stimuli in the superior colliculus of the unanesthe- tized, mid-pontine cat, Arch. Ital. BioZ, 106: 169-193, 1968. SPRAGUE, J. M. AND MEiKLE, T, H., JR. The role of the superior colliculus in visually guided behavior. Exptl. Neurol. 11: 115-146, 1965. STERLING, P. AND WICKELGREN, B. G. Visual

MONKEY SUPERIOR COLLICULUS 201
receptive fieIds in the superior colliculus of physics, edited by R. Jung and H. Rornhuber. the cat. J. Neurophysiol. 32: 1-15, 1969, Berlin: Springer, 1961, p. 210-211~
37. STRASCHILL, M. AND HOFFMAN, K. P. Functional 40. WILSON, M. E, AND TOYNE, M. J. Retino-tectal
aspects of localization in the cat’s tectum opti- and cortico-tectal projections in Macaca mu-
cum. Brain Res. 13: 274-283, 1969. lutta. Bruin Res. 24: 395406, 1970.
38. STRASCHILL, M. AND HOFFMAN, K. P. Activity 41. WOLBARSHT, M. C., MACNXCHOL, E. F., AND
of movement sensitive neurons of the cat’s WAGNER, H. G. Glass insuIated platinum micro-
tectum opticum during spontaneous eye move- electrode. Science 132: 1309-1310, 1960.
ments. ExptZ. Brain Res. 11: 318-326, 1970. 42. WURTZ, R. H. AND GOLDBERG, M. E. Superior
39. 'WHITTERIDGE, D. Tectum opticum. In: The colliculus cell responses related to eye move- ments in awake monkeys. Science 171: 82-84,
Visual System: Neurophysiology and Psycho- 1971.





























