Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 296, No. 1, July, pp. 302-307, 1992
Rat Lung Polychlorinated Biphenyl-Binding Protein: Effect of Glucocorticoids on the Expression of the Clara Cell-Specific Protein during Fetal Development’
Magnus Nerd,* Olof Andersson,*,T Mikael Brijnnegiird,*,$ and Johan Lund**’ *Department of Medical Nutrition, TDepartment of Lung Medicine, and *Department of Pediatrics, Karolinska Institutet, Huddinge University Hospital F60, Novum, S141 86 Huddinge, Sweden
Received January 21, 1992, and in revised form March 5, 1992
Certain metabolites of polychlorinated biphenyls (PCBs) are retained in the Clara cells and in the airway lumen of rodent lung due to their interaction with a se- cretory 13-kDa protein. The expression of this Clara cell- specific, PCB-binding protein (PCB-BP) during the fetal development of the rat lung was studied by means of li- gand binding and a monospecific antiserum. The PCB- BP and specific 4,4’-bis([SH]methylsulfonyl)-2,2’,5,6’- tetrachlorobiphenyl binding was first detected on ges- tational Day 19 and subsequently the levels of PCB-BP and specific ligand binding increased as a function of gestational age. The start site of transcription for the rat PCB-BP gene was determined by primer extension anal- ysis and the information thus obtained was used to de- velop a quantitative assay for the corresponding mRNA based on solution hybridization and Sl nuclease mapping. The appearance of PCB-BP mRNA during fetal lung de- velopment preceded the detection of immunoreactive protein and ligand binding by 1 day. By Day 2 1, the level of PCB-BP mRNA was 15 ng/lOO pg total lung RNA which is approximately 30-40% of adult levels. In utero exposure to the synthetic glucocorticoid betamethasone was shown to increase specific 4,4’-bis([3H]methylsul- fony1)2,2’,5,5’-tetrachlorobiphenyl binding, PCB-BP pro- tein, and PCB-BP mRNA if administered from gesta- tional Day 18 and onward. By Days 21-22, glucocorti- coid treatment resulted in a two- to threefold increase in the levels of specific ligand binding, immunoreactivity, and mRNA, i.e., to approximately adult levels. o 1992
Academic Press, Inc.
Previous studies indicate that xenobiotics, i.e., certain methylsulfonyl metabolites of polychlorinated biphenyls (PCBS),~ may selectively accumulate in Clara cells of the rodent lung due to the presence in these cells of a specific, secretory binding protein (l-4). Exposure of humans to PCBs is associated with inflammatory symptoms from the airways and an accumulation of methylsulfonyl-PCB in the lungs of exposed subjects is well documented (5, 6). A similar binding protein for PCBs has also been shown to be present in bronchoalveolar lavage fluid from healthy human volunteers (7). The PCB-binding protein (PCB-BP) has been purified from rat lung cytosol (2) as well as from human bronchoalveolar lavage fluid (8). Par- tial amino acid sequence analysis of the purified proteins as well as the recent isolation of the cDNA for the rat PCB-BP (9) suggests that the PCB-binding proteins con- stitute the rat and human homologues of rabbit uterog- lobin [reviewed in (10) and (ll)] and that they are iden- tical to the rat and human Clara cell-specific lo-kDa secretory proteins (CClO) (12-14).
Rabbit uteroglobin is a dimeric protein first identified in uterine secretions (15) but it is also produced by epi- thelial cells of the lung as shown by, e.g., in situ hybrid- ization (16). Uteroglobin binds progesterone and the expression of its corresponding gene is under hormonal control by progesterone in the uterus and by glucocorti- coids in the lung [reviewed in (10) and (ll)]. The three- dimensional structure of uteroglobin has been determined by X-ray crystallography (17-19). Particular residues in uteroglobin believed to participate in ligand binding have
3 Abbreviations used: PCB, polychlorinated biphenyl; (MeSO,),TCB, 4,4’-bis(methylsulfonyl)-2,2’,5,5’-tetrachlorobipheny~ PCB-BP, PCB- binding protein; CClO, Clara cell-specific lo-kDa secretory protein; GR,
r This work was supported by the Swedish Medical Research Council, glucocorticoid receptor; PEPCK, phosphoenolpyruvate carboxykinase; Magnus Bergvalls Stiftelse, Swedish Heart-Lung Foundation, Stiftelsen PAG, polyacrylamide gel; SDS-PAGE, sodium dodecyl sulfate-poly- Lars Hiertas Minne. acrylamide gel electrophoresis; SP-A, -B, and -D, surfactant apoproteins
’ To whom correspondence should be addressed. A, B, and D, respectively.
302 0003-9%x/92 $5.00 Copyright 0 1992 by Academic Press, Inc.
All rights of reproduction in any form reserved.

RAT FETAL POLYCHLORINATED BIPHENYL-BINDING PROTEIN 303
to a greater extent than other residues been conserved in the rat and human homologues, implying that ligand binding is linked to the function of the proteins. Subtle differences in ligand binding characteristics exist and a recent study by Peter et al. (20) shows that changing Thr60 of rabbit uteroglobin to the residue present at the corresponding position in the human homologue decreases the affinity of the expressed protein for progesterone. Thus, although the structural and functional similarities of the three proteins suggest a common ancestry, differ- ences exist and in particular the tissue-specific and hor- monal control of the three genes require further studies. Glucocorticoids induce the uteroglobin gene in fetal rabbit lung explants (21) and recently Hagen et al. (14) showed that the mRNA for the rat homologue can be increased by glucocorticoids in adult rats. The developmental and glucocorticoid regulation of rat PCB-BP in lung was in- vestigated given the suggested role of the uteroglobins in inflammation as inhibitors of phospholipase A2 (11, 13) and the important role of glucocorticoids in fetal lung development [reviewed in (22) and (23)] as well as their use in the treatment of inflammation.
MATERIALS AND METHODS
Chemicals. 4,4’-bis([3H]methylsulfonyl)-2,2’, 5,5’-tetrachlorobiphenyl [(3H-MeS02)2TCB, sp act 4 Ci/mmol, purity > 90%] and unlabeled (MeSO,),TCB were kind gifts from Drs. A. Bergman, E. Klasson-Wehler, and C.-A. Wachtmeister, Wallenberg Laboratory, University of Stock- holm, Stockholm, Sweden (24). ‘251-protein A was purchased from NEN (Boston, MA). Radioactive nucleotides, restriction enzymes, and mod- ifying enzymes were obtained from Amersham (Buckinghamshire, England). All other reagents were analytical grade chemicals from Sigma Chemical Co. (St. Louis, MO) or Merck A. G. (Darmstadt, Germany).
Animals. Pregnant Sprague-Dawley rats were injected i.m. with 0.2 mg/kg betamethasone (Celestona Soluspan, Schering Corp., NJ; con- taining equal amounts of betamethasone acetate and betamethasone phosphate) or saline (controls) 12 and 24 h prior to sacrifice. The time of conception was known within 8 b. Betamethasone was used because this glucocorticoid has a prolonged biological activity in target tissues and no mineralocorticoid effects. The rats were killed by cervical dis- location and the fetuses delivered by cesarean section. The number of fetuses was registered and the lungs were removed under the microscope. The lungs from each litter were pooled, weighed, and then further pro- cessed.
Preparation of cytosol and in vitro ligand binding assay. Pooled lungs from rat fetuses of different gestational age were homogenized in 20 mM potassium phosphate, 1 mM EDTA, pH 7.2, using a tight fitting Teflon/ glass Potter-Elvehjem homogenizer. The homogenate was centrifuged at 105,OOOg for 60 min. The supernatant was removed and stored at -70°C until further use. The in vitro ligand binding assay was performed on cytosolic preparations as previously described (1). The data from two saturation analysis experiments for each time-point were plotted according to Woolf (25) in order to obtain equilibrium binding param- eters. The validity of the Woolf transformation was determined by the r-value and only r-values > 0.9 were accepted.
SDS-PAGE and Western immunoblotting. Aliquots of cytosolic preparations (100 ag of protein/aliquot) were subjected to SDS-PAGE (15% PAG), transferred to nitrocellulose and the filters probed with antibodies to rat PCB-BP as previously described (2). Complexes of antigen, antibody, and 1251-protein A were visualized by autoradiography and then cut out of the nitrocellulose filters and the radioactivity was
determined by use of a LKB Wallac Minigamma counter. The Western immunoblots were run in duplicate. In parallel, partially purified PCB- BP was run as an internal control to allow comparisons between blots.
Primer extension and 5’1 nuclease analysis. Primer extension analysis was performed on poly(A)+mRNA from adult rat lung according to Domdey et al. (26) in order to determine the start-site of transcription of the rat PCB-BP gene. The primer (B-GGCTGAGCTGCAGCA- GATGGACAGCATGAG-3’) corresponds to the lower strand of the published cDNA between nucleotides +85 and +114 (9) and primer extension was performed using reverse transcriptase from either Moloney murine leukemia virus (37°C) or avian myeloblastosis virus (42°C). Parallel dideoxy-sequencing reactions were performed using the same oligonucleotide primer and a Sac1 fragment of a genomic clone isolated from a rat genomic EMBL SP6/T7 library (Clontech, CA) using the rat PCB-BP cDNA. This clone generated five fragments when digested with Sacl, one of which (approximately 4.5 kb in size) was positive when probed by Southern blotting with the 32P-labeled PCB-BP cDNA. This fragment was subcloned into pUCl9 and its sequence determined. The 5’-end of the Sac1 fragment corresponds to nucleotide -167 in the num- bering of the rat CC10 gene (14). This Sac1 fragment thus overlaps the Pstl-Sac1 fragment used by Hagen et al. (14) to map the start-site of transcription of the CC10 gene by Sl nuclease mapping. Primer-extended products and the dideoxy-sequencing products were analyzed by 6% denaturing PAG/urea electrophoresis.
A synthetic oligonucleotide for Sl analysis of correctly initiated PCB- BP mRNA transcripts was made based on the determined start-site of transcription of the PCB-BP gene and the assay was performed as pre- viously described (27). The oligonucleotide is 60 nucleotides in length (5’.GTCTGTAGATGTGGGCTGATGTTGTAATGTGAGGGTATG- TGTGGGTATGTGTGGGTATGC-3’) and correctly initiated tran- scripts give rise to protected fragments of 47 and 42 nucleotides in length as analyzed by 10% denaturing PAG/ urea electrophoresis. Protected fragments were cut out of the dried gels after, autoradiography and the radioactivity was determined by liquid scintillation counting. An internal standard of pooled RNA from adult rat lung were run in each assay to allow for a quantitation of specific PCB-BP mRNA. The pooled RNA had in turn been calibrated against in vitro-synthesized mRNA obtained by transcription of the cDNA in a pGEM vector using T7 polymerase. The Sl nuclease analysis was run in triplicate.
RESULTS
Pregnant Sprague-Dawley rats, where the time of con- ception was known, were injected i.m. with 0.2 mg beta- methasone/kg body wt or saline at 12 and 24 h prior to sacrifice. The fetuses were delivered by cesarean section and lungs were removed under the microscope and then weighed. The lungs from each litter were pooled and then processed for cytosolic preparations and isolation of RNA. The average number of live fetuses obtained upon cesar- ean section was 12.8 + 1.2 (mean f SD) and was not influenced by the betamethasone treatment.
In order to investigate the effect of in utero exposure to glucocorticoids on the expression of the rat lung PCB- BP we analyzed cytosolic preparations from pools of lungs from exposed and nonexposed rat fetuses for specific (MeSO&TCB-binding. As shown in Fig. 1, specific ligand binding was first observed on Day 19 in lung cytosol from both control and treated fetuses. However, the level of specific PCB-binding in cytosolic preparations from treated animals was approximately 200% compared to control preparations. The levels of specific PCB-binding increased as a function of gestational age in both control
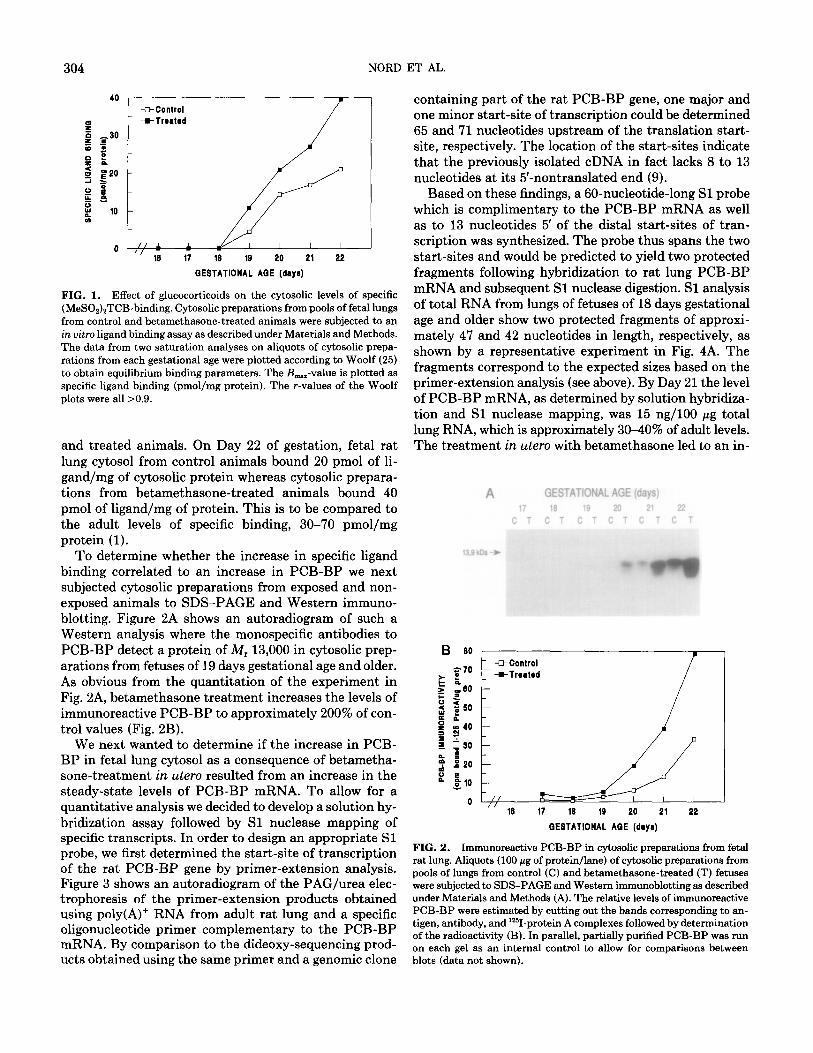
304 NORD ET AL.
/ & I
O I’/ 16 17 18 19 20 21 22
GESTATIONAL AGE (days)
FIG. 1. Effect of glucocorticoids on the cytosolic levels of specific (MeSO&TCB-binding. Cytosolic preparations from pools of fetal lungs from control and betamethasone-treated animals were subjected to an in vitro ligand binding assay as described under Materials and Methods. The data from two saturation analyses on aliquots of cytosolic prepa- rations from each gestational age were plotted according to Woolf (25) to obtain equilibrium binding parameters. The &,-value is plotted as specific ligand binding (pmol/mg protein). The r-values of the Woolf plots were all >0.9.
and treated animals. On Day 22 of gestation, fetal rat lung cytosol from control animals bound 20 pmol of li- gand/mg of cytosolic protein whereas cytosolic prepara- tions from betamethasone-treated animals bound 40 pmol of ligand/mg of protein. This is to be compared to the adult levels of specific binding, 30-70 pmol/mg protein (1).
To determine whether the increase in specific ligand binding correlated to an increase in PCB-BP we next subjected cytosolic preparations from exposed and non- exposed animals to SDS-PAGE and Western immuno- blotting. Figure 2A shows an autoradiogram of such a Western analysis where the monospecific antibodies to PCB-BP detect a protein of M, 13,000 in cytosolic prep- arations from fetuses of 19 days gestational age and older. As obvious from the quantitation of the experiment in Fig. 2A, betamethasone treatment increases the levels of immunoreactive PCB-BP to approximately 200% of con- trol values (Fig. 2B).
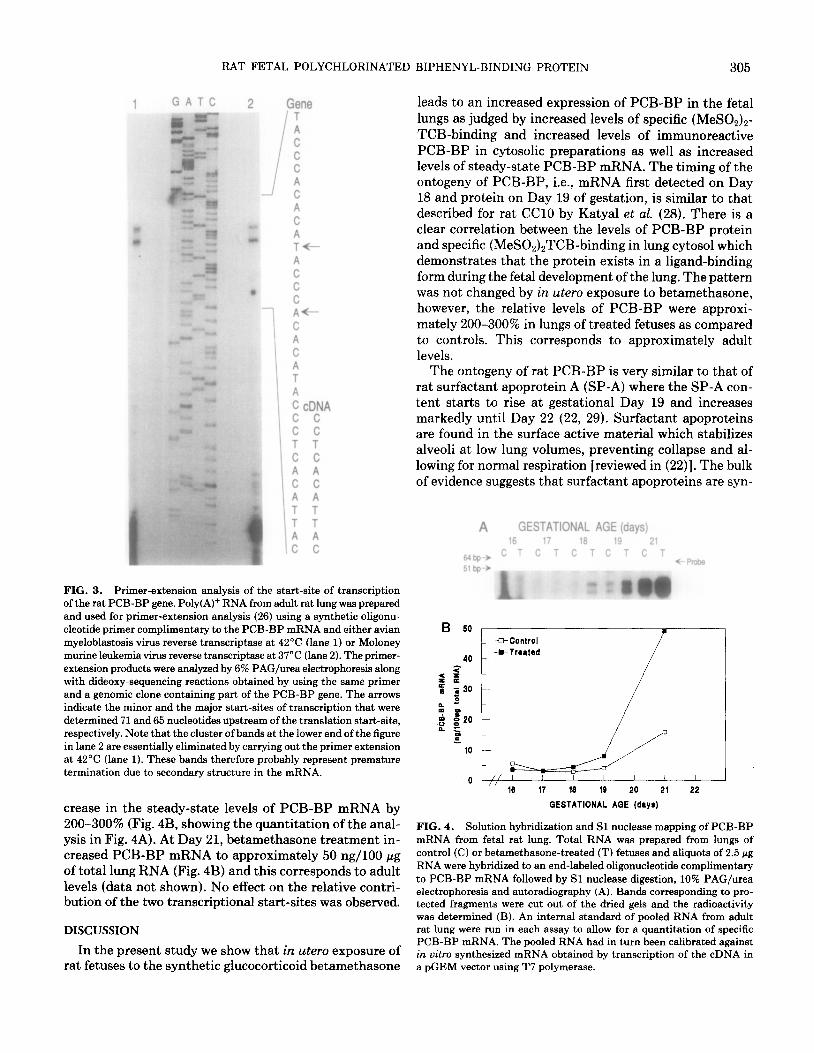
We next wanted to determine if the increase in PCB- BP in fetal lung cytosol as a consequence of betametha- sone-treatment in utero resulted from an increase in the steady-state levels of PCB-BP mRNA. To allow for a quantitative analysis we decided to develop a solution hy- bridization assay followed by Sl nuclease mapping of specific transcripts. In order to design an appropriate Sl probe, we first determined the start-site of transcription of the rat PCB-BP gene by primer-extension analysis. Figure 3 shows an autoradiogram of the PAG/urea elec- trophoresis of the primer-extension products obtained using poly(A)+ RNA from adult rat lung and a specific oligonucleotide primer complementary to the PCB-BP mRNA. By comparison to the dideoxy-sequencing prod- ucts obtained using the same primer and a genomic clone
containing part of the rat PCB-BP gene, one major and one minor start-site of transcription could be determined 65 and 71 nucleotides upstream of the translation start- site, respectively. The location of the start-sites indicate that the previously isolated cDNA in fact lacks 8 to 13 nucleotides at its 5’-nontranslated end (9).
Based on these findings, a 60-nucleotide-long Sl probe which is complimentary to the PCB-BP mRNA as well as to 13 nucleotides 5’ of the distal start-sites of tran- scription was synthesized. The probe thus spans the two start-sites and would be predicted to yield two protected fragments following hybridization to rat lung PCB-BP mRNA and subsequent Sl nuclease digestion. Sl analysis of total RNA from lungs of fetuses of 18 days gestational age and older show two protected fragments of approxi- mately 47 and 42 nucleotides in length, respectively, as shown by a representative experiment in Fig. 4A. The fragments correspond to the expected sizes based on the primer-extension analysis (see above). By Day 21 the level of PCB-BP mRNA, as determined by solution hybridiza- tion and Sl nuclease mapping, was 15 ng/lOO pg total lung RNA, which is approximately 30-40% of adult levels. The treatment in hero with betamethasone led to an in-
A GESTATIONAL AGE (days) 17 18 19 20 21 22
CTCTCTCTCTCT
.::“::
” 18 17 18 19 20 21 22
QESTATIONAL ABE (daya)
FIG. 2. Immunoreactive PCB-BP in cytosolic preparations from fetal rat lung. Aliquots (100 pg of protein/lane) of cytosolic preparations from pools of lungs from control (C) and betamethasone-treated (T) fetuses were subjected to SDS-PAGE and Western immunoblotting as described under Materials and Methods (A). The relative levels of immunoreactive PCB-BP were estimated by cutting out the hands corresponding to an- tigen, antibody, and “‘I-protein A complexes followed by determination of the radioactivity (B). In parallel, partially purified PCB-BP was run on each gel as an internal control to allow for comparisons between blots (data not shown).
RAT FETAL POLYCHLORINATED BIPHENYL-BINDING PROTEIN 305
1 GATC 2 Gene T A C
: A C A C A T A C C C A C A C A T
C C
i A C A T T A C
f
c
cD!?A C T C A C A T T A C
FIG. 3. Primer-extension analysis of the start-site of transcription of the rat PCB-BP gene. Poly(A)+ RNA from adult rat lung was prepared and used for primer-extension analysis (26) using a synthetic oligonu- cleotide primer complimentary to the PCB-BP mRNA and either avian myeloblastosis virus reverse transcriptase at 42°C (lane 1) or Moloney murine leukemia virus reverse transcriptase at 37°C (lane 2). The primer- extension products were analyzed by 6% PAG/urea electrophoresis along with dideoxy-sequencing reactions obtained by using the same primer and a genomic clone containing part of the PCB-BP gene. The arrows indicate the minor and the major start-sites of transcription that were determined 71 and 65 nucleotides upstream of the translation start-site, respectively. Note that the cluster of bands at the lower end of the figure in lane 2 are essentially eliminated by carrying out the primer extension at 42°C (lane 1). These bands therefore probably represent premature termination due to secondary structure in the mRNA.
crease in the steady-state levels of PCB-BP mRNA by 200-300% (Fig. 4B, showing the quantitation of the anal- ysis in Fig. 4A). At Day 21, betamethasone treatment in- creased PCB-BP mRNA to approximately 50 ng/lOO pg of total lung RNA (Fig. 4B) and this corresponds to adult levels (data not shown). No effect on the relative contri- bution of the two transcriptional start-sites was observed.
DISCUSSION
In the present study we show that in utero exposure of rat fetuses to the synthetic glucocorticoid betamethasone
leads to an increased expression of PCB-BP in the fetal lungs as judged by increased levels of specific (MeSO& TCB-binding and increased levels of immunoreactive PCB-BP in cytosolic preparations as well as increased levels of steady-state PCB-BP mRNA. The timing of the ontogeny of PCB-BP, i.e., mRNA first detected on Day 18 and protein on Day 19 of gestation, is similar to that described for rat CC10 by Katyal et al. (28). There is a clear correlation between the levels of PCB-BP protein and specific (MeSO&TCB-binding in lung cytosol which demonstrates that the protein exists in a ligand-binding form during the fetal development of the lung. The pattern was not changed by in utero exposure to betamethasone, however, the relative levels of PCB-BP were approxi- mately 200-300% in lungs of treated fetuses as compared to controls. This corresponds to approximately adult levels.
The ontogeny of rat PCB-BP is very similar to that of rat surfactant apoprotein A (SP-A) where the SP-A con- tent starts to rise at gestational Day 19 and increases markedly until Day 22 (22, 29). Surfactant apoproteins are found in the surface active material which stabilizes alveoli at low lung volumes, preventing collapse and al- lowing for normal respiration [reviewed in (22)]. The bulk of evidence suggests that surfactant apoproteins are syn-
A GESTATIONAL AGE (days) 16 17 18 19 21
!34bp+ c T c T c T c T c T fProbe
6 50 UControl
22
GESTATIONAL APE (days)
FIG. 4. Solution hybridization and Sl nuclease mapping of PCB-BP mRNA from fetal rat lung. Total RNA was prepared from lungs of control (C) or betamethasone-treated (T) fetuses and aliquots of 2.5 pg RNA were hybridized to an end-labeled oligonucleotide complimentary to PCB-BP mRNA followed by Sl nuclease digestion, 10% PAG/urea electrophoresis and autoradiography (A). Bands corresponding to pro- tected fragments were cut out of the dried gels and the radioactivity was determined (B). An internal standard of pooled RNA from adult rat lung were run in each assay to allow for a quantitation of specific PCB-BP mRNA. The pooled RNA had in turn been calibrated against in oitro synthesized mRNA obtained by transcription of the cDNA in a pGEM vector using T7 polymerase.

306 NORD ET AL.
thesized in alveolar type II cells, whereas the PCB-BP is a product of the nonciliated bronchiolar Clara cell (3). Results from studies of the localization of surfactant apo- protein mRNA have been conflicting in regard to Clara cells and whether the immunocytochemical findings of surfactant apoprotein in Clara cells represent phagocy- tosis or synthesis remain unclear [reviewed in (22); (29, 30)]. Also the prenatal developmental profile of gluco- corticoid responsiveness of SP-A shows striking similar- ities to that of PCB-BP with marked increases by glu- cocorticoid treatment of the SP-A content at gestational Days 19 and 21 (29). The ontogeny and developmental responsiveness to glucocorticoids of rat surfactant apo- protein B (SP-B) differ slightly from those of SP-A and PCB-BP (29) and show a response to glucocorticoids at Day 19 but not at Day 21. Recent results regarding the ontogeny of rat surfactant apoprotein D @P-D) (31) shows a pattern similar to those of SP-A and PCB-BP. The similarity of the prenatal developmental profile and responsiveness to glucocorticoids of PCB-BP to those of the surfactant apoproteins suggests an important role for PCB-BP in the maturation of the lung prior to birth.
The rat PCB-BP cDNA (9) was used to isolate a ge- nomic clone which contains part of the structural gene for rat PCB-BP, including the first exon. Partial analysis of the first intron (data not shown) as well as upstream sequences (Fig. 3) shows almost perfect homology to the rat CC10 gene (14), thus providing further evidence for their identity. We also determined the location of the cap site for the rat PCB-BP gene by primer extension analysis. One major and one minor start-site of transcription were determined at 65 and 71 nucleotides upstream of the translation start-site. The assignment of these start-sites is not identical to that of Hagen et al. (14). The Sl nu- clease assay used in that study, however, yielded a cluster of bands centered around 72 to 74 nucleotides upstream of the translation start-site. Given the fact that the ge- nomic CC10 sequence of Hagen et al. (14) contains an extra CCA-codon just upstream of the initiator ATG not present in the PCB-BP cDNA or genomic clone, the two studies are probably in good agreement also taking into account that two independent methods were used. No change in the relative contribution of the start-sites could be observed as a consequence of the exposure to gluco- corticoids.
An important question is whether glucocorticoids act directly on the Clara cells to stimulate PCB-BP expres- sion. To date no studies of the effect of glucocorticoids on rat Clara cells in culture are available but such studies are needed. Also the level at which the regulation occurs is not known. Clearly, the homology to uteroglobin sug- gests that the hormonal effect is primarily at the tran- scriptional level [reviewed in (lo)]. However, comparisons of the 5’-flanking region of the uteroglobin gene with that of the rat CC10 gene show some major differences (14). Two regions of 0.3 kb and 2.1 kb are absent in the rat
CC10 upstream gene region. The 2.1-kb region in the uteroglobin gene contains a cluster of hormone receptor binding sites, believed to be responsible for the differential regulation by glucocorticoids and progesterone. It will therefore be important to study in detail the regulatory region of the rat PCB-BP/CC10 gene since some aspects of regulation most likely will differ from that of utero- globin.
The effects of glucocorticoids during lung development are complex. Previous studies have shown the glucocor- ticoid receptor (GR) mRNA and protein to be present in fetal rat lungs of gestational age 16 to 21(32,33). In utero exposure to betamethasone resulted in a significant down- regulation of GR mRNA at all gestational ages studied, whereas the GR protein was not down-regulated by its ligand until after Day 19 (33). These studies demonstrate that the potential for a receptor-mediated glucocorticoid response in fetal rat lung exists prior to Days 18-19; i.e., the receptor is present but somehow the function of the receptor must be restricted. It further illustrates that the ontogenetic pattern of PCB-BP expression and its re- sponsiveness to glucocorticoids is not a general phenom- enon in fetal rat lung since the expression and regulation of GR is different. Another example is phosphoenolpyru- vate carboxykinase [PEPCK; (34)], a gene known to con- tain glucocorticoid responsive elements (35-37) but which does not become active in rodent lung until Day 7 post- partum.
The ontogenies of PCB-BP, GR, PEPCK, and the sur- factant apoproteins thus illustrate the complexity of glu- cocorticoid action in rodent lung development. An im- portant feature emerging from the studies of a number of glucocorticoid responsive cellular genes is the presence of &-acting elements that activate transcription via collab- oration between the GR and an alternative transcription factor, so called composite hormone response elements or glucocorticoid response units (37-40). In this way, glu- cocorticoid responsiveness of different genes can be con- trolled by the number and/or activity of the auxilliary factors. Importantly, such a hypothesis for a transition to induction competence of genes during lung develop- ment is becoming possible to address experimentally since the genes for GR, PCB-BP, surfactant apoproteins, and PEPCK have all been isolated.
The roles of PCB-BP and other Clara cell secretory proteins in the lung homeostasis are not known but clearly merit further attention based on the developmental pat- tern of PCB-BP expression, which suggests a role in the final airway maturation prior to birth [this study, (28)], and the suggested role of uteroglobin/PCB-BP/CC10 in the inflammatory process (8,11,41). In addition, the tox- icological consequences of the binding of PCBs to these homologous proteins should be investigated further.
ACKNOWLEDGMENTS We thank Lena Nordlund-Mijller and Pia L&strand for expert tech-
nical assistance..

RAT FETAL POLYCHLORINATED BIPHENYL-BINDING PROTEIN 307
REFERENCES 21.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15. 16.
17.
18.
19.
20.
Lund, J., Brandt, I., Poellinger, L., Bergman, A., Klasson-Wehler, E., and Gustafsson, J.-A. (1985) Mol. Pharmacol. 27, 314-323.
Lund, J., Nordlund, L., and Gustafsson, J.-A. (1988) Biochemistry 27, 7895-7901.
Lund, J., Devereux, T., Glaumann, H., and Gustafsson, J.-A. (1988) Drug Metab. Dispos. 16, 590499.
Brandt, I., Lund, J., Bergman, A., Klasson-Wehler, E., Poellinger, L., and Gustafsson, J.-A. (1985) Drug Metab. Dispos. 13, 490-496.
Shigematsu, N., Ishimaru, R., Saito, R., Ikedu, T., Matsuba, K., Suginama, K., and Masuda, Y. (1978) Enuiron. Res. 16,92-100. Haraguchi, K., Kuroki, H., and Masuda, Y. (1984) Food Ckm. Ton- icol. 22, 283-284.
Lund, J., Andersson, O., and Ripe, E. (1986) Toxicol. Appl. Phw- macol. 83,486-493.
Andersson, O., Nordlund-Moller, L., Bronnegird, M., Sirzea, F., Ripe, E., and Lund, J. (1991) Am. J. Respir. Cell Mol. Biol. 5, 6-12.
Nordlund-Moller, L., Andersson, O., Ahlgren, R., Schilling, J., Gill- ner, M., Gustafsson, J.-A., and Lund, J. (1990) J. Biol. Chem. 265, 12,690-12,693.
Savouret, J.-F., and Milgrom, E. (1983) DNA 2, 99-104.
Miele, L., Cordella-Miele, E., and Mukherjee, A. B. (1987) Endocrine Rev. 8,474-490.
Singh, G., Singh, J., Katyal, S. L., Brown, W. E., Kramps, J. A., Paradis, I. L., Dauber, J. H., Macpherson, T. A., and Squeglia, N. (1988) J. Histochem. Cytochem. 36, 73-80.
Singh, G., Katyal, S. L., Brown, W. E., Kennedy, A. L., Singh, U., and Wong-Chong, M.-L. (1990) Biochim. Biophys. Acta 1039,348- 355. Hagen, G., Wolf, M., Katyal, S. L., Singh, G., Beato, M., and Suske, G. (1990) Nucleic Acids Res. 18, 2939-2946.
Beier, H. M. (1968) Biochim. Biophys. Acta 160, 289-291.
Warembourg, M., Tranchant, O., Atger, M., and Milgrom, E. (1986) Endocrinology 119, 1632-1640.
Mornon, J. P., Fridlansky, F., Bally, R., and Milgrom, E. (1980) J. Mol. Biol. 137, 415-429.
Morize, I., Surcouf, E., Vaney, M. C., Epelboin, Y., Buehner, M., Fridlansky, F., Milgrom, E., and Mornon, J. P. (1987) J. Mol. Biol. 194,725-739. Bally, R., and Delettre, J. (1989) J. Mol. Biol. 206, 153-170.
Peter, W., Brtiller, H.-J., Vriend, G., Beato, M., and Suske, G. (1991) J. Steroid Biochem. Mol. Biol. 38, 27-33.
22. 23. 24.
25. 26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
Lopez de Haro, M. S., and Nieto, A. (1985) Biochem. J. 225, 255- 258.
Ballard, P. L. (1989) Endocrine Reu. 10, 165-181.
Gross, I. (1990) Am. J. Physiol. 259, L337-L334.
Klasson-Wehler, E., Bergman, A. L., and Wachtmeister, C. A. (1983) J. Label. Cmpd. 20, 1407-1412.
Haldane, J. B. (1957) Nature 179, 832.
Domdey, H., Apostol, B., Lin, R.-J., Necoman, A., Brody, E., and Abelson, J. (1984) Cell 39, 611-621.
Lund, J., Ahlgren, R., Wu, D., Kagimoto, M., Simpson, E. R., and Waterman, M. R. (1990) J. Biol. Chem. 265, 3304-3312.
Katyal, S. L., Singh, G., Brown, W. E., Kennedy, A. L., Squeglia, N., and Wong Chong, M.-L. (1990) Progr. Respir. Res. 25, 29-35.
Shimizu, H., Miyamura, K., and Kuroki, Y. (1991) Biochim. Biophys. Acta 1081,53-60. Crouch, E., Rust, K., Marienchek, W., Parghi, D., Chang, D., and Persson, A. (1991) Am. J. Respir. Cell Mol. Biol. 5, 13-18.
Ogasawara, Y., Kuroki, Y., Shiratori, M., Shimizu, H., Miyamura, K., and Akino, T. (1991) Biochim. Biophys. Acta 1083, 252-256. Bronnegard, M., and Okret, S. (1988) J. Steroid Biochem. 3 1,809- 817.
Bronnegird, M., and Okret, S. (1991) J. Steroid Biochem. Mol. Biol. 39,13-17. Zimmer, D. B., and Magnuson, M. A. (1990) J. Hi&o&em. Cytochem. 38, 171-178.
Sasaki, K., Cripe, T. P., Koch, S. R., Andreone, T. L., Petersen, D. D., Beale, E. G., and Granner, D. K. (1984) J. Biol. Chem. 259, 15,242-l&251.
Short, J. M., Wynshaw-Boris, A., Short, H. P., and Hanson, R. W. (1986) J. Biol. Chem. 261,9721-9726. Imai, E., Stromstedt, P.-E., Quinn, P. G., Carlstedt-Duke, J., Gus- tafsson, J.-A., and Granner, D. K. (1990) Mol. Cell. Biol. 10, 4712- 4719.
Danesch, U., Gloss, B., Schmid, W., Schlitz, G., Schiibe, R., and Reukawitz, R. (1987) EMBO J. 6, 625-630.
Diamond, M. I., Miner, J. N., Yoshinaga, S. K., and Yamamoto, K. R. (1990) Science 249, 1266-1271.
Zhang, H., and Young, A. P. (1991) J. Biol. Chem. 266, 24,332- 24,338. Mihal, K., and Riedel, N. (1991) Am. J. Respir. Cell Mol. Biol. 5, l-3.





























