Embed Size (px)
Citation preview

Plant Physiol. (1984) 75, 387-3920032-0889/84/75/0387/06/$01.00/0
Pyruvate Orthophosphate Dikinase of C3 Seeds and Leaves asCompared to the Enzyme from Maize'
Received for publication October 26, 1983 and in revised form January 24, 1984
KAZUKO AOYAGI AND JAMES A. BASSHAM*Laboratory ofChemical Biodynamics, Lawrence Berkeley Laboratory, University ofCalifornia,Berkeley, California 94720
ABSTRACT
Pyruvate orthophosphate dikinase (PPDK) was found in varis im-mature seeds of C3 planRts (wheat, pea, green bean, plm, ad castorbean) in some C3 eaves (tobacco, spinach, sunflower, an wheat and inC (maize) kernels. The enzyme in the C3 plants crss-eacts with rabbitantserum against mae PPDK. Based on protein blot analysis, theapparent subuit size of PPDK from wheat seeds and leaves and fromsunflower leaves is about 94 kdaltos, the same as that of the enzymefrom iz but is slightly less (about 90 kdaltons) for the enzyme fromspiach and tobacco leaves. Te amount of this enzyme per mg of solubleprotein in C3 seeds and kaves is much less than in C4 eaves. PPDK bpresent in kernels of the C4 plant, Zea my in amounts comparable tothose in C4 leaves.
Regulatory properties of the enzyme from C3 tissues (wheat) aresimlr to those of the enzyme from C4 leaves with respect to i vwio lightactivation and dark inativatio (in leaves) and in vith cold lability (seedsand leaves).
Foilowing inoportion of 14 2 by illumiated wheat pericarp andadjoining tise for a few seconds, the labeled metabolies were predom-inantly products ret from carboxylatio of phosphoenolpyruvate,with lesser labeling of compounds formed by carboxylation of ribulose1,5-bisphosphate and operation of the reductive pentose phosphate cycleof photosynthesis. PPDK may be involved in of amino acidintercoversios dwing seed development.
Pyruvate orthophosphate dikinase (EC 2.7.9.1) is an essentialenzyme for photosynthetic carbon dioxide fixation in C4 plants(12), and in some CAM plants (14). PPDK2 activity is presentalso in seeds of barley (9) and wheat (18, 19), a C3 plant. Therehave been conflicting reports about the presence of PPDK insome C3 leaves (for references, see 1). Recently, the presence andamount ofPPDK activity in wheat seeds and leaves were reportedand compared with the amount ofPPDK activity in maize, andthe amounts ofPPDK protein in these tissues were estimated byimmunoprecipitation in the protein blot method (1).The role ofPPDK in the metabolism ofC3 plant tissues is not
established, although a role in guard cell stomatal opening hasbeen suggested (8, 25). The presence of PPDK in several addi-tional (3 seed and leaf tissues now has been seen. The character-
' Supported by the Office of Energy Research, Office of Basic EnergySciences, Biological Energy Research Division of the United StatesDepartment of Energy under Contract No. DE-AC03-76SF00098.
2Abbreviations: PPDK, pyruvate orthophosphate dikinase; PEPC,phosphoenolpyruvatecarboxylase; RuBPC, ribulose 1,5-biphosphate car-boxylase; MDH, malic dehydrogenase.
istics of the enzyme from wheat tissues are compared with thosefrom C4 tissues. The pattern ofincorporation of 14C into metab-olites during short periods ofphotosynthesis with 4C02 by wheatpericarp and associated tissue has been examined for evidenceof possible roles ofPPDK in the metabolism of these tissues.
MATERIAIS AND METHODSPlants. Wheat (Triticum aestivum L. cv Cheyenne, CI 8885),
kidney beans (Phaseolus vulgaris L. cv late red kidney), pea(Pisum sativum L.), spinach (Spinacia oleracea L. cv Highpackor cv 5286 America), Zea mays L., and tobacco (Nicotianatabacum L. cv Wisconsin) were grown in a greenhouse. Castorbean (Ricinus communis L.) and sunflower (Helianthus annuusL.) plants were grown outside in Berkeley. Immature fruit ofplum (Prunus tomentosa L.) was obtained from a tree in theBotanical Garden at University of California, Berkeley.
Reagents. Radiochemicals were purchased from New EnglandNuclear, malic dehydrogenase (MDH) was purchased from Boer-inger Mannheim Biochemicals; chemical reagents from SigmaChemical Co. and from Biorad Laboratories.
Protein Blot. PPDK was isolated from maize and purified aspreviously described (29). Antiserum to PPDK was prepared byinjecting New eland white rabbits with 150 pg of the purifiedenzyme and Freund's complete adjuvant, followed after 26 d bya booster containing the same amount of enzyme and Freund'sincomplete adjuvant and a second such booster at 40 d. Therabbits were bled at 41 d. The crude antiserum was used for theprotein blot. Nonimmunized serum showed no cross-reactionwith PPDK, and PPDK antiserum cross-reacted only with PPDK(1). This antiserum was used as a probe for the presence ofPPDKin.the various tissues studied.Each plant tissue studied (3 g) was ground in 6 ml of 0.1 M
Tris buffer, pH 7.4, containing 10 mM MgC2, 18% w/v sucrose,and 1% P-mercaptoethanol. The homogenate was filtered andthen centrifuged at 15,000g and OC for 20 min. Total solubleprotein was determined by the method of Bradford (5). Cross-reacting protein subunits similar in size to that of maize PPDKwere detected by the protein blot method (6, 21).SDS-PAGE of each plant extract was carried out in a gradient
gel (6.4 to 12.8%). Protein was transferred to cyanogen bromidepaper prepared by the method of Clarke et al. (6). The tansferpaper was probed with anti-PPDK serum and then with J125protein A. An autoradiograph was prepared using Kodak AR5x-ray film with an intensifying screen at -70°C overnight.
Densitometry. Relative levels of PPDK in preparations fromdifferent plant samples were estimated by densitometry of the x-ray film by comparison of the peak areas. Three or four knownamounts of purified PPDK were loaded on the SDS gel adjacentto the plant tissue samples each time to calibrate the amount.Under the conditions chosen there was a linear relationshipbetween the amount ofprotein loaded on the gel and the area of
387
AOYAGI AND BASSHAM
the densitometry peak.Ouchterlony Two-Dimensional Diffusion (22). Small wells
were cut in agar plates. One was filled with 20 Al of antiserum, asecond with solution containing 4 ,ug of purified PPDK frommaize, and the others with 20 !d of plant extract. The precipitinband was observed after 1 to 4 d of incubation at room temper-ature. The reaction was stopped by addition of0.9% (w/v) NaClsolution, and the bands were stained with Coomassie BriliantBlue. RuBPC antibody was kindly supplied by Dr. Mark Harps-ter, who prepared it by raising rabbit antibody to purified maizeleaf RuBPC.PPDK Activity Assay. PPDK activity was assayed as previ-
ously described (29). Dependence of the activity on ATP and onPi were demonstrated by separate controls in which each wasomitted from the reaction mixture. Protein content of tissueextracts was determined by the method of Bradford (5) and Chlcontent was measured according to Arnon's method (3).Cold Lability. The crude extract was prepared under sunlight
as described previously (1). The enzyme was then stored 30 and60 min at OC, then transferred back to room temperature andassayed after 30, 60, and 90 min.PEPC Activity Assay. PEPC was assayed by the reduction of
oxaloacetate with MDH and NADH to malate and spectropho-tometric observation ofthe change in absorption at 340 nm (17).Following 3 min preincubation of the PEPC at 22C in 0.9 mlof assay mixture containing 0.1 M Tris Ha (pH 8), 10 mMMg9l2, 2 mm DTT, 0.2 mm NADH, 10 mm NaHCO3, and 4units ofMDH, the reaction was initiated by the addition ofPEP(to give 2.5 mM).
Incorporation of Labeled Carbon Dioxide. Immature wheatseeds (25 d after flowering) were dissected. After removal ofendosperm tissue, the pericarp with adjoining tissue was cut intosmall pieces (about 2 x 2 x 2 mm). For each sample, 80 mggreen tissue was placed in 1 ml buffer (3 mm NaHCO3, 20 mMHepes, pH 7.8) in a glass homogenizing tube and preilluminatedat about 650 ,uE m-2sg' for 2 min. Then 0.2 ml NaH'4CO3solution was added to give a final concentration of 8.4 mm (25,uCi Amol-'), and illumination was continued for 5, 10, 15, 30,and 60 s for the five samples. Liquid N2 was poured into eachtube at the end of this time to stop the reactions. Methanol wasadded to give 50% methanol and the mixture was ground as it
A
thawed. An aliquot portion of this mixture was analyzed by two-dimensional paper chromatography and radioautography, andincorporation of "`C into individual compounds was measuredby liquid scintillation counting (23).
RESULTSAs previously reported, PPDK is present in wheat leaves and
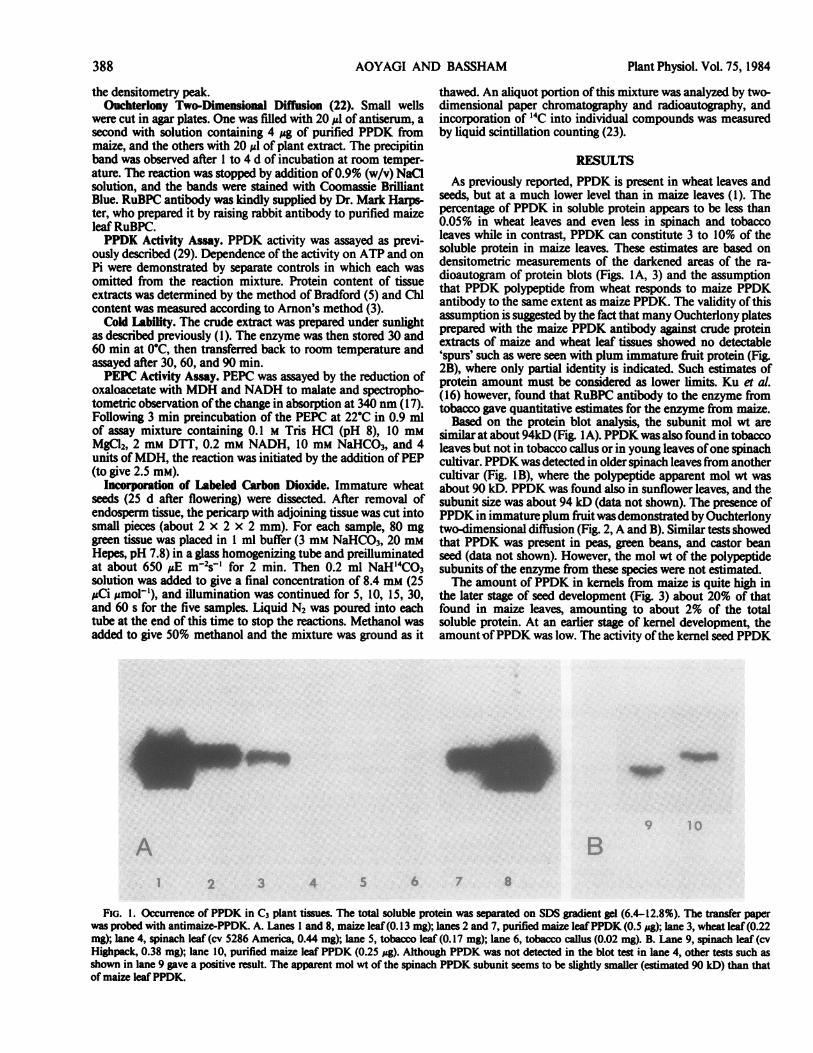
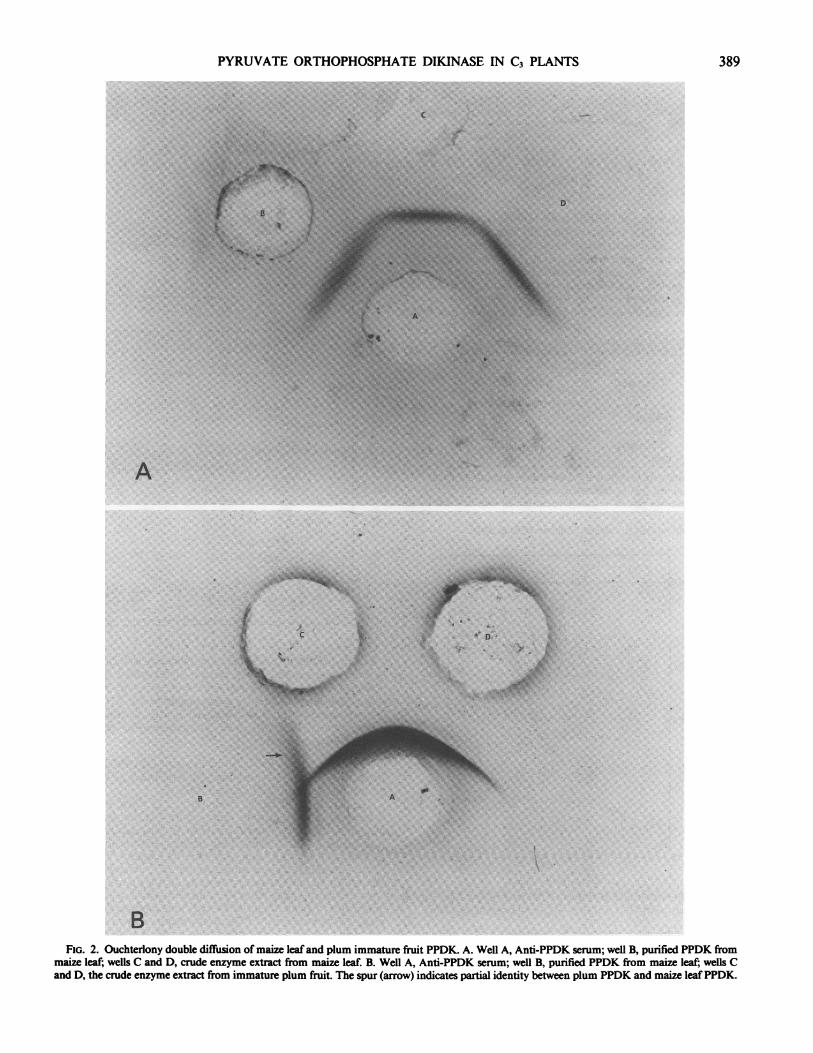
seeds, but at a much lower level than in maize leaves (1). Thepercentage of PPDK in soluble protein appears to be less than0.05% in wheat leaves and even less in spinach and tobaccoleaves while in contrast, PPDK can constitute 3 to 10% of thesoluble protein in maize leaves. These estimates are based ondensitometric measurements of the darkened areas of the ra-dioautogram of protein blots (Figs. IA, 3) and the assumptionthat PPDK polypeptide from wheat responds to maize PPDKantibody to the same extent as maize PPDK. The validity of thisassumption is suggested by the fact that many Ouchterlony platesprepared with the maize PPDK antibody against crude proteinextracts of maize and wheat leaf tissues showed no detectable'spurs' such as were seen with plum immature fruit protein (Fig.2B), where only partial identity is indicated. Such estimates ofprotein amount must be considered as lower limits. Ku et al.(16) however, found that RuBPC antibody to the enzyme fromtobacco gave quantitative estimates for the enzyme from maize.
Based on the protein blot analysis, the subunit mol wt aresimilarat about 94kD (Fig. lA). PPDKwas also found in tobaccoleaves but not in tobacco callus or in young leaves ofone spinachcultivar. PPDK was detected in older spinach leaves from anothercultivar (Fig. IB), where the polypeptide apparent mol wt wasabout 90 kD. PPDK was found also in sunflower leaves, and thesubunit size was about 94 kD (data not shown). The presence ofPPDK in immature plum fruitwasdemonstrated by Ouchterlonytwo-dimensional diffusion (Fig. 2, A and B). Similar tests showedthat PPDK was present in peas, green beans, and castor beanseed (data not shown). However, the mol wt of the polypeptidesubunits of the enzyme from these species were not estimated.The amount of PPDK in kernels from maize is quite high in
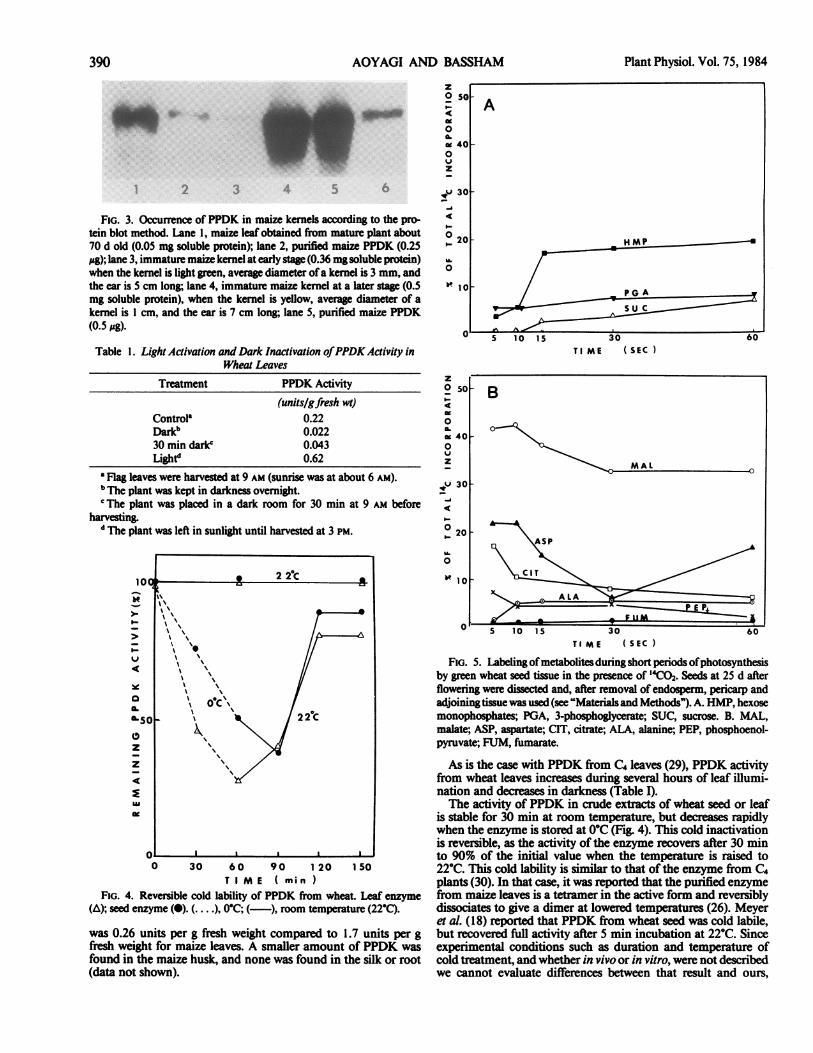
the later stage of seed development (Fig. 3) about 20% of thatfound in maize leaves, amounting to about 2% of the totalsoluble protein. At an earlier stage of kernel development, theamount ofPPDK was low. The activity ofthe kernel seed PPDK
BB..2 3 4 5. 6 7i, 8
FiG. 1. Occurrence of PPDK in C3 plant tissues. The total soluble protein was separated on SDS gradient gel (6.4-12.8%). The transfer paperwas probed with antimaize-PPDK. A. Lanes I and 8, maize leaf(0.13 mg); lanes 2 and 7, purified maize leafPPDK (0.5 gg); lane 3, wheat leaf(0.22mg); lane 4, spinach leaf (cv 5286 America, 0.44 mg); lane 5, tobacco leaf (0.17 mg); lane 6, tobacco callus (0.02 mg). B. Lane 9, spinach leaf (cvHighpack, 0.38 mg); lane 10, purified maize leaf PPDK (0.25 #tg). Although PPDK was not detected in the blot test in lane 4, other tests such asshown in lane 9 gave a positive result. The apparent mol wt of the spinach PPDK subunit seems to be slightly smaller (estimated 90 kD) than thatof maize leaf PPDK.
388 Plant Physiol. Vol. 75, 1984
PYRUVATE ORTHOPHOSPHATE DIKINASE IN C3 PLANTS
c
D
A
C } 4 CoD
B
FIo. 2. Ouchterlony double diffusion of maize leaf and plum immature fruit PPDK. A. Well A, Anti-PPDK serum; well B, purified PPDK frommaize leaf; wells C and D, crude enzyme extract from maize leaf. B. Well A, Anti-PPDK serum; well B, purified PPDK from maize leaf; wells Cand D, the crude enzyme extract from immature plum fruit. The spur (arrow) indicates partial identity between plum PPDK and maize leafPPDK.
389
...... ..
W.-o
AOYAGI AND BASSHAM
l2 3 4 5 6
FIG. 3. Occurrence of PPDK in maize kernels according to the pro-tein blot method. Lane 1, maize leaf obtained from mature plant about70 d old (0.05 mg soluble protein); lane 2, purified maize PPDK (0.25pg); lane 3, immature maize kernel at early stage (0.36 mg soluble protein)when the kernel is light green, average diameter ofa kernel is 3 mm, andthe ear is 5 cm long; lane 4, immature maize kernel at a later stage (0.5mg soluble protein), when the kernel is yellow, average diameter of akernel is I cm, and the ear is 7 cm long; lane 5, purified maize PPDK(0.5 sg).
Table 1. Light Activation and Dark Inactivation ofPPDKActivity inWheat Leaves
Treatment PPDK Activity(units/gfresh wt)
Control' 0.22Darkb 0.02230 min darkC 0.043Lightd 0.62
*Flag leaves were harvested at 9 AM (sunrise was at about 6 AM).b The plant was kept in darkness overnight.I The plant was placed in a dark room for 30 min at 9 AM before
harvesting.d The plant was left in sunlight until harvested at 3 PM.
0 30 60 90 120 150T I ME ( min )
FIG. 4. Reversible cold lability of PPDK fr-om wheat. Leaf enzyme
(A); seed enzyme (0). (.... ), 0iC; (-), room temperature (22"C).
was 0.26 units per g fresh weight compared to 1.7 units per gfresh weight for maize leaves. A smaller amount of PPDK wasfound in the maize husk, and none was found in the silk or root(data not shown).
z
oo soA0 40ag 400Uz
30-u 304
-20 _ HMP
U./0
10 PGA
Su C
0 5 10 is 30 60TI ME ( SEC
Z2 50 B
0ILat40-0UZ AL
30
220 ~~ASP0
P E P,
0 5 10 15 30 60T I M E (SEC
FIG. 5. Labeling ofmetabolitesduring short periods ofphotosynthesisby green wheat seed tissue in the presence of '4CO2. Seeds at 25 d afterflowering were dissected and, after removal of endosperm, pericarp andadjoiningtissue was used (see "Materials and Methods"). A. HMP, hexosemonophosphates; PGA, 3-phosphoglycerate; SUC, sucrose. B. MAL,malate; ASP, aspartate; CIT, citrate; ALA, alanine; PEP, phosphoenol-pyruvate; FUM, fumarate.
As is the case with PPDK from C4 leaves (29), PPDK activityfrom wheat leaves increases during several hours of leaf illumi-nation and decreases in darkness (Table 1).The activity of PPDK in crude extracts of wheat seed or leaf
is stable for 30 min at room temperature, but decreases rapidlywhen the enzyme is stored at 0C (Fig. 4). This cold inactivationis reversible, as the activity of the enzyme recovers after 30 minto 90% of the initial value when the temperature is raised to22C. This cold lability is similar to that of the enzyme from C4plants (30). In that case, it was reported that the purified enzymefrom maize leaves is a tetramer in the active form and reversiblydissociates to give a dimer at lowered temperatures (26). Meyeret a. (18) reported that PPDK from wheat seed was cold labile,but recovered full activity after 5 min incubation at 22°C. Sinceexperimental conditions such as duration and temperature ofcold treatment, and whether in vivo or in vitro, were not describedwe cannot evaluate differences between that result and ours,
390 Plant Physiol. Vol. 75, 1984

PYRUVATE ORTHOPHOSPHATE DIKINASE IN C3 PLANTS
although we note that a different cultivar was used. In a laterreport (19) the same authors failed to observe cold inactivationand light-mediated activation of wheat seed PPDK in prelimi-nary experiments.The pH optimum of the enzyme from wheat tissues is about
7.9. In wheat seeds 25 d after anthesis, when PPDK protein is atits highest level, its activity is about 0.1 unit per g fresh weight,while at that time the activity ofPEPC is about ten times greater.
Analysis of photosynthetic fixation of '4C-labeled bicarbonateby wheat seed green tissue (25 d after flowering) showed that thefixation at the shortest times is predominantly into compoundsderived from oxaloacetate (malate, aspartate, fumarate, and cit-rate (Fig. 5). After 5 s labeling, malate accounted for 42% ofradiocarbon found in compounds on the paper chromatogram.After longer periods of photosynthesis with labeled carbon, thenormal pattern of C3 assimilation (sugar phosphates, phospho-glycerate) becomes more prominent. Experiments with greenseed tissue 18 d after flowering gave similar results (not shown),although labeling of C4 compounds was slightly less dominant.Fixation of 14CO0 was not linear, being at a somewhat greaterrate during the first few seconds.
DISCUSSIONAlthough a limited number of C3 species so far have been
tested, PPDK appears to be more widely distributed among C3plants than had been generally realized. Negative results frompast efforts to detect enzyme activity may be due to a variety ofcauses including variability in amount due to age of leaves andother physiological conditions, inactivation during extraction,and lack of purification (since there is so little activity, it oftencannot be detected in crude extracts). The amount of PPDK isin fact very much less in C3 tissues as compared with C4 tissues.Moreover, PPDK thus far is found in C3 plants only in someseeds at certain stages ofdevelopment and in leaves. This limitedoccurrence, together with the regulatory properties ofthe enzymewhich mandate special conditions for obtaining the active form,may explain the scarcity of previous reports (9, 19, 25) of itspresence in such tissues.There are many similarities between the C3 PPDK (in wheat
seeds and leaves) and the enzyme from C4 plants. Mol wt (94kD) of the subunit, in vivo light activation (in leaves; seeds nottested), cold instability, and pH profile of the wheat enzyme areall similar to these properties in the enzyme from maize, a C4plant. The polypeptide subunit from leaves of spinach and to-bacco were slightly smaller (90 kD), while sunflower leaf PPDKsubunit had an apparent mol wt of about 94 kD.
Based on the Ouchterlony double diffusion technique (22),wheat seed or leaf PPDK appeared identical with maize leafPPDK, when tested with maize leafPPDK antibody. Wheat leafRuBPC, on the other hand, did not appear completely identicalwith maize leaf RuBPC, when tested with maize leaf RuBPCantibody (data not shown).The role of PPDK in the metabolism of C3 leaves is not well
established. It has been proposed that there may be an involve-ment of PPDK in control of stomatal opening through a partic-ipation in exchange in ion transport into guard cells (8, 25). Inseeds, PPDK may supply PEP for photosynthetic and dark CO2fixation. Photosynthesis by developing wheat grains can accountfor as much as 34% of the weight gain of the grains (10).Considerable additional photosynthesis by the grains may beinvolved in the refixation of respiratory CO2 (15). Stomata wereobserved on wheat and barley grains toward the brush end onthe flanks of the crease by Cochrane and Duffus (7) who exam-ined the ultrastructure of immature grains and discussed it inrelation to transport of gasses and metabolites and the relativelylow rate of photosynthetic 14CO2 fixation of intact seeds com-pared to the much larger rate seen upon removal of the outer
pericarp (24).The pattern of '4C-labeled compounds formed during short
periods of photosynthesis with '4C-labeled bicarbonate in wheatseeds 25 d after flowering indicates a very high PEPC activity(Fig. 5). Seeds at this late development stage were chosen for theexperiment since the highest level of PPDK is seen in such seeds(2). There is a preponderance of labeled products ofPEP carbox-ylation such as aspartate, malate, citrate, and glutamate. Thecontinued high percentage of labeling of these compounds to Imin seems to suggest a relatively high rate of net synthesis of C4and Cs compounds. The higher rate of fixation at the shortesttime may indicate that accumulated PEP was rapidly used upwhen bicarbonate was administered to the exposed tissue.Nutbeam and Duffus (20) reported predominant labeling of
malic acid at 1 min of photosynthetic 14C02 fixation by barleypericarp tissue, with labeling moving to sucrose and hexosephosphates at longer times, and they considered this to beevidence for C4 photosynthesis in that tissue. Similar results weresaid to have been obtained for wheat pericarp, but data were notpresented. In any event, the time scale for the kinetics of thoseresults with barley pericarp are quite different from those of thepresent study in which the labeling of malate and aspartate,although declining after the first 15 s, continued to be prominent.Wirth et al. (31) reported '4C labeling in wheat pericarp at 10 sonly, and found a predominance of labeled products character-istic ofC3 photosynthesis. Such differences in results from differ-ent studies may well be related to physiological differences in theseed tissue used, especially in stage of development and variety.In fact, in a preliminary experiment (data not shown) withanother variety (Anza) 18 d after flowering we obtained a pre-dominantly C3 fixation pattern.Given the greater activity of PEPC than of PPDK, some PEP
for carboxylation may come from glycolysis. However, conver-sion of pyruvate to PEP, mediated by PPDK, may be an impor-tant source of the PEP substrate for carboxylation. In particular,PEP carboxylation and a preceding conversion of pyruvate toPEP mediated by PPDK may be required for the net conversionof the C3 amino acid carbon skeleton of alanine to C4 and C5amino acids in wheat grains. Amino acids present in majoramounts in flag-leaf phloem are glutamate, aspartate, serine,alanine, and glycine (27). Conversion of this mixture of aminoacids to that required for seed protein synthesis must occur inthe seeds ( 13).Wheat storage protein such as gliadin is very rich in glutamate
and proline (11) which is made from glutamate. During thesynthesis of such proteins, the utilization of other amino acidsincluding alanine might lag, allowing them to accumulate in theendosperm during the first 3 weeks of seed development. Alanineis a major soluble amino acid of endosperm ofwheat grains, andwhen the plants are not irrigated, alanine can constitute as muchas 47% of total soluble amino acids (28). In this case, the peakin quantity of alanine occurs at about 25 d after flowering, atabout the same time as the maximum in PPDK protein seen inother studies (2). Perhaps during the last 2 weeks, conversion ofexcess accumulated alanine to glutamate is needed for efficientutilization of this stored alanine. Another possibility, the conver-sion of alanine to aspartate (19) seems less important since amuch smaller amount of C4 amino acids than of C5 amino acidsare required in the storage protein (1 1). For similar reasons, otherpossibilities such as conversion of malate to pyruvate followedby reduction to sugars seem less likely.
In maize, alanine is the amino acid occurring in highestamount among the free amino acids in the developing kernel(4). There may, therefore, be a necessity in corn kernels for theconversion of alanine to C4 and C5 amino acids during the latterstages of kernel development. Both PPDK protein and activitywere found in immature maize kernels. A previous study (19)
391

Plant Physiol. Vol. 75, 1984
based only on activity assay did not find PPDK in maize kernels,despite finding PPDK activity in wheat grains, where we foundlessPPDK activity than in maize kernels. Possibly in the previousstudy the kernels were at an earlier stage of development whenthe level of PPDK is much lower (Fig. 3), or the enzyme mayhave been inactivated during harvesting and extraction.The peak in PPDK activity in wheat seeds and in maize kernels
occurs well after the maximum in Chl content and RuBPCcontent of the green seed tissue or ear (2). This timing alsosuggests that the role ofPPDK in seed tissue is not primarily inphotosynthetic CO2 incorporation as it is in C4 leaf tissue. It isinteresting that nevertheless much more PPDK is expressed inmaize kernels than in wheat seed. Thus the greater expression ofthis enzyme seen in C4 leaves seems to be carried over into theseed, at least when maize and wheat are compared.
LITERATURE CITED
1. AOYAGI K, JA BASHAM 1983 Pyruvate orthophosphate dikinase in wheatleaves. Plant Physiol 73: 853-854
2. AOYAGI K, JA BASSHAM, PC GREENE 1983 Pyruvate orthophosphate dikinasegene expression in developing wheat seeds. Plant Physiol 75: 393-396
3. ARNON DJ 1949 Copper enzymes in isolated chloroplasts. Polyphenoloxidasein Beta vulgaris. Plant Physiol 24: 1-15
4. ARRUDA P, JW SILVA 1979 Amino acid composition of vascular sap of maizeear pduncle. Phytochemistry 18: 409-410
5. BRADFORD M 1976 A rapid and sensitive method for the quantitation ofmicrogram quantities of protein using the principle of protein dye binding.Anal Biochem 72: 248-254
6. CLARKE L, R HrTzMAN, J CARBON 1979 Selection of specific clones fromcolony banks screening with radioactive antibody. Methods Enzymol 68:436-442
7. COCHRANE MP,CM DUFFUS 1979 Morphology and ultrastructure ofimmaturecereal grains in relation to transport. Ann Bot 44: 67-72
8. DAs VSR, AS RAGHAVENDRA 1974 Control of stomatal opening by pyruvatemetabolism in light. Indian J Exp Bot 12: 425-428
9. DUFFUS CM, R ROsIE 1973 Some enzyme activities associated with thechlorophyll containing layers ofimmature barley pericarp. Planta 1 14: 219-226
10. EVANS LT, HM RAWSON 1970 Photosynthesis and respiration by the flag leafand components of the ear during grain development in wheat. Aust J BiolSC; 23: 245-254
11. JENNINGs AC, RK MORTON 1963 Changes in carbohydrate, protein, andnonprotein nitrogenous compounds of developing wheat grain. Aust J Biol
SCi 16: 318-33112. HATCH MD,CR SLAcK 1968 A newenzyme for the interconversion ofpyruvate
and phosphopyruvate and its role in the C. dicarboxylic acid pathway ofphotosynthesis. Biochem J 106: 141-146
13. KIRKMAN MA, BJ MIFLIN 1979 The nitrate content and amino acid compo-sition of the xylem fluid of spring wheat throughout the growing season. JSci Food Agric 30: 653-660
14. KLUGE M, CB OSMOND 1971 Pyruvate Pi dikdnase in crassulacean acidmetabolism. Naturwissenschaften 58: 414-415
15. KRIEDMANN P 1966 The photosynthetic activity ofthe wheat ear. Ann Bot 30:349-363
16. Ku MSB, MR SCHMIrr, GE EDWARDS 1979 Quantitative determination ofRuBPcarboxylase-oxygenase protein in leaves of several C3 and C4 plants. JExp Bot 30: 89-98
17. LANE MD, H MARUYAMA, RL EAStDAY 1969 Phosphoenolpyruvate carbox-ylase from peanut cotyledons. Methods Enzymol 13: 277-283
18. MEYER AO, GJ KELLY, E LArTxo 1978 Pyruvate orthophosphate dikinasefrom immature wheat grains. Plant Sci Lctt 12: 35-40
19. MER AO, GJ KELLY, E LATzKO 1982 Pyruvate orthophosphate dikinasefrom immature grains ofcereal graes. Plant Physiol 69: 7-10
20. NumEMm AR, CM DuFFs 1976 Evidence for C4 photosynthesis in barleypericarp tissue. Biochem Biophys Res Commun 70: 1198-1201
21. NYARI UJ, YH TAN, HA ERLuCH 1981 Production and characterization ofmonoclonal antibodies to human fibrobls -interferon. In E De Maeyer,G Galaso, H. Schelekens, eds, The Biology of the Interferon System.Elsevier North Holland, New York, pp 67-71
22. OUCHTERLONY 0 1968 Diffusion-in-gel methods for immunological analysis.Prog Allergy 5: 78
23. PEDERSEN TA, M KIRK, JA BAsSHAM 1966 Light-dark transients in levels ofintermediate compounds during photosynthesis in air-adapted Chlorella.Physiol Plant 19: 219-231
24. RADLEY M 1976 The development of wheat grain in relation to endogenousgrowth substances. J Exp Bot 27: 1009-1021
25. SCHNABLH 1981 The comparmentation ofcarboxylatingand decarboxylatingenzyme in guard cell protoplast Planta 152: 307-313
26. SHIRAHASHI K, S HAYAKAWA, T SUGIYAMA 1978 Cold lability of pyruvateorthophosphate dikinase in the maize leaf. Plant Cell Physiol 62: 826-830
27. SIMPSON RJ, MJ DALLING 1981 Nitrogen redistribution during grain growthin wheat (Triticum aestivum L). Planta 151: 447-456
28. SINGH U 1981 The influence ofirrigtion on the composition ofsoluble aminoacids in developing wheat grain. J Sci Food Agric 32: 203-207
29. SUGIYAMA T 1973 Purification, molecular, and catalytic properties ofpyruvatephosphate didnase from maize leaf. Biochemistry 12: 2862-2868
30. SUGIYAMA T, MR ScHMIDT, SB Ku, GE EDWARDS 1979 Difference in coldlability of pyruvate, Pi dikinase among C4 species. Plant Cell Physiol 20:965-97 1
3 1. WIRTH E, GJ KELLY, G FISCHER, E LATLKO 1976 Enzyme activities andproducts ofC02 fixation in various photosynthetic orans ofwheat and oat.Z Pflanzenphysiol 82: 78-87
392 AOYAGI AND BASSHAM






![Glucan, Water Dikinase Exerts Little Control over …Glucan, Water Dikinase Exerts Little Control over Starch Degradation in Arabidopsis Leaves at Night1[W][OPEN] Alastair W. Skeffington2,](https://img.pdfslide.us/doc/110x75/5e6be6838773a760e0501b6e/glucan-water-dikinase-exerts-little-control-over-glucan-water-dikinase-exerts.jpg)