Embed Size (px)
Citation preview

Proc. Indian Acad. Sci. (Anim. Sci.), Vol. 95, No.3, June 1986, pp. 303-324.© Printed in India.
Population dynamics of a tropical lepidopteran Catopsilia crocale(Pieridae)
M S M CHRISTOPHER· and S MATHAVANSchool of Biological Sciences, Madurai Kamaraj University, Madurai 625021, India
• Mis Ram Bahadur Thakur (P) Ltd, Travancore Tea Estates Group, Siddharth Building,Cochin 682003, India
MS received 21 May 1985; revised 16 August 1985
Abstract. Extensive fieldobservations were undertaken to study the population dynamics ofthe larvae of Catopsilia crocale in the Madurai Kamaraj University campus during the years1980-81 and 1981-82. Though oligophagous, C. crocaleadopted 'monophagic strategy' foroviposition and larval feeding. Seasonal changes in the nitrogen content of the food Cassiaalata leaf influenced the pupal/adult weight and oviposition. Commencement of rain triggeredthe reproductive activity and continued precipitation facilitated maximum egg deposition. Oftotal egg production in a year, about 80 % was oviposited during northeast monsoon(September-December) and the rest during the south-west monsoon (May-August). Egg, Iinstar, V instar and pupa suffered more mortality than the other lifestages. Ofthe total numberof eggs laid, about 1·01 o,~ emerged as adult and 98·99 ~.~ died during the process of larval andpupal development.
Keywords. Lepidopteran larvae; Catopsilia crocale; fecundity; life table; populationdynamics.
1. Introduction
The occurrence of fluctuations in tropical butterfly populations has been reported by anumber of workers (Ehrlich and Gilbert 1973; Watt et a11979; Vats and KaushaI1980).However, most of the workers have restricted their studies to the adult populations.Only a few publications report on fluctuations in the larval populations and thecausative factors of the fluctuations (e.g. Hayes 1981). Environmental factors such asrainfall, temperature and relative humidity have been shown to influence thepopulation either directly (Chew 1975; Watt et aI1977; Shapiro 1978a, b; Blau 1980) orthrough the host plants (Wolda 1978b; Mooney et al 1980, 1981; Lakhani andDempster 1981).
While reviewing butterfly ecology, Gilbert and Singer (1975) stated that a few lifetables have been compiled for butterflies, that too for a single generation (e.g. Pierisrapae; Harcourt 1966; Dempster 1967; Thomas 1974; Lycaena dispar; Duffey 1968).Recently, life table has been constructed for Papilio polyxenes (Blau 1980).Hayes (1981)made a detailed long term study on the population ecology and life table of the pieridbutterfly Calias alexandra.
In the present study it has been attempted to analyse the factors influencingoviposition. larval population dynamics and life table of the butterfly Catopsiliacrocale.
303

304 M S M Christopher and S Mathaoan
2. Materials and methods
2.1 The study site
The study site is situated within the Madurai Kamaraj University campus, Madurai(780 100Eand 90 58'N). Altitude is about 132·5m, Madurai climate is monsoonic with anaverage annual rainfall of 400-700 mm. During the study period the mean maximumand minimum temperature varied from 38°C (June) to 21°C (January). It is a semi aridregion consisting of a variety of complex diversified tropical fauna and flora. The areachosen for the study was about 7,5OOm2• The topography of the study site is given infigure 1.
22 The organism
Catopsilia crocale Cramer belongs to the Order Lepidoptera, Family Pieridae, subfamily Coliadinae. It is a tropical species, very commonly found in India, Ceylon andBurma (Wynter-Blyth 1935). The females are distinguished from the males by thepresence ofa black mark in the apex and the costa. In males, the apex and the costa arenarrowly black, whereas in females they are broadly black in colour. Adult C. crocaleoccurs concomitantly with other related species like C. pomona, C. pyranthae and C.florella (Beeson 1941).
Scale 1 em =15 m
-!....-...._II~E3 ~_
c:J,I II II II II I
L.--
XX Distribution of Ca. alata
123 4 5 Biology Dept. s6 instrumenta tion Centre
7 8 Chemistry
9 Physics
10 librar y
Figure 1. Topography of the study area.

Population dynamics of C. crocale 305
C. crocale is a multivoltine species, breeds continuously throughout the year except inthe dry season. However, adult population persists both in the dry and wet seasons ofthe year. C. crocale is oligophagous; it has been reported that the larvae utilize plantsbelonging to Cassia sp. such as Ca. alata, Ca. siamea, Ca. nodosa, Ca. javanica and Ca.grandis as food sources (Browne 1968). However, in the study area (habitat) though avariety of Cassia sp. were present, C. crocale females oviposited only on Ca. alata andlarvae utilized the same plant as the potential food. Adults visited a number of plantsbelonging to various families for nectar.
2.3 Food plant
In India. Ca. alata is called as dadmurdum, which may be translated as herps-killer. Ca.alata is a soft woody shrub, belonging to the sub-family Cecalpiniae; cosmopolitan indistribution in the tropics and semi arid regions. These plants usually grow upto 3 mheight. In the study area though plants ofdifferent height/age are distributed randomly,small plants (-- 1·5m) outnumbered the large plants. At the beginning of summer, i.e.during the months ofFebruary-March they shed the old leaves and new buds sprout inMay following the first precipitation. Maximum leaf production is synchronized withmaximum precipitation i.e. during the months of September-November. Floweringcommences during NovemberfDecember. Since most of the conserved energy isutilized for flowering and seed production in the tall plants, leaf production is reducedafter December.
2.4 Egg and larval counting
The study was conducted for two years during the period from January 1980 to January1982. Total number of eggs oviposited on all Ca. alata plants in the study area duringthe two seasons were determined by direct counting method. Field observations weremade on alternate days and each Ca. alata leaf was examined carefully. Eggs oncecounted were marked by a magic marker at the tip of the egg without causing anydamage and hence the error of repeated counting was avoided. Eggs were depositedexclusively on the tender leaves and hence it was easy to locate them for counting.
For accurate censusing oflarval population, the following method was adopted. Lifestagewise number oflarvae in each plant was counted on the day ofobservation. Since I,II, III and IV instars required only 2 days each to complete the feeding period, those atthe II instar on the day of observation would have moulted into III instar on thesubsequent observation, hence a particular stage larva was not counted twice for thesame age. However, since final instar required about 2'5-3,0 days, total number of finalinstar larvae in each plant was counted in each observation; the increase or decrease ofthis larval stage in the subsequent observation was estimated by counting the newlyformed pupa or newly moulted larvae. After a few weeks of careful regular fieldobservations it became easier for us to census accurately.
2.5 Life table
To obtain complete information of mortality at each life stage of C. crocale, a life tablewas constructed using data from regular field observations on the number of eggs,

306 M S M Christopher and S Mathacan
larvae, pupae and adults. Causes of mortality were also recorded from the observations.From this, age specific life table was constructed for each month of the seasons. Lifetable data were subjected to 'key-factor' analysis (Varley and Gradwell 1960). k orkilling factor was a concept originally by Haldane (1949)and applied by Morris (1959)and Varley and Gradwell (1960), as a statistical method for identifying the age specificcauses of population change. The k value for the mortality of each life stage is thedifference between the common logarithm of the number entering that stage and thesubsequent one. The total generation mortality (K) was calculated by adding all the kvalues. In order to know the monthly changes in the surviving individuals in each stage,survivorship curve was drawn by plotting the data obtained for monthly survivor ineach life stage (lx) against the age (x).
Data on daily temperature (maximum and minimum), humidity (maximum andminimum) and rainfall during the study period were collected from the minimeteorological station of the School of Biological Sciences (courtesy: Unit of AnimalBehaviour, School of Biological Sciences). From these data a monthly mean wascalculated.
3. Results
The following plants have been reported to serve as food for C. crocale larvae: Cassiaalata, Ca. fistula, Ca. simona and Ca. jauanica (Browne 1968).
However, from extensive regular field observations, it was found that C. crocaleoviposited on leaves ofCa. alata with nearly 100% accuracy. Other cassia species in thesame area did not attract the animal. Although the C. crocale larvae are considered to beoligophagous, the adults strictly follow the monographic type of oviposition. A similartype of oligophagous butterflies preferring monophagic type of oviposition has beenreported for Papilia machaon (Wiklund 1974) and Euphydryas editha (Holdren andEhrlich 1982).
C. crocale lays eggs singly on the leaves of Ca. alata. After landing on the plant, thefemale C. crocale requires about 4-6 seconds for the deposition of an egg. After eggdeposition, she usually moves to another plant or another leaf of the same plant todeposit the next egg.
A careful observation revealed that though the food plants were distributedrandomly, egg deposition was heavy on small plants and it was poor on large/tall plants.The height of the plant varied and a maximum height of 3·0m was observed. Of thefood plants present in the study area, about 20%were above 2·5m height. Surprisinglynot even a single egg was observed in these plants during the study period of25 months.
For plants of different heights, the number of eggs laid per plant were counted for aperiod of 5 days and are presented in table 1. The number of eggs laid per plantdecreased with increasing height of the plant. A significant negative correlation wasobtained and the slope of the regression, calculated are presented in figure 2 (r =-0'95; Y = 341+ 121·5X). With increasing nitrogen content of the leaf, total numberof eggs deposited per plant increased (Y = 237·5 - 98·7 X; r = -0'933). Even in thesmall plants, C. crocale preferred mostly tender leaves for egg deposition.
The pattern of egg distribution on the leaf was observed and presented in table 2. Ofthe total 259 eggs observed in a chosen period, 126, 30, 103 eggs were found in theupper, lower and margin of the leaves. respectively. The eggs were observed
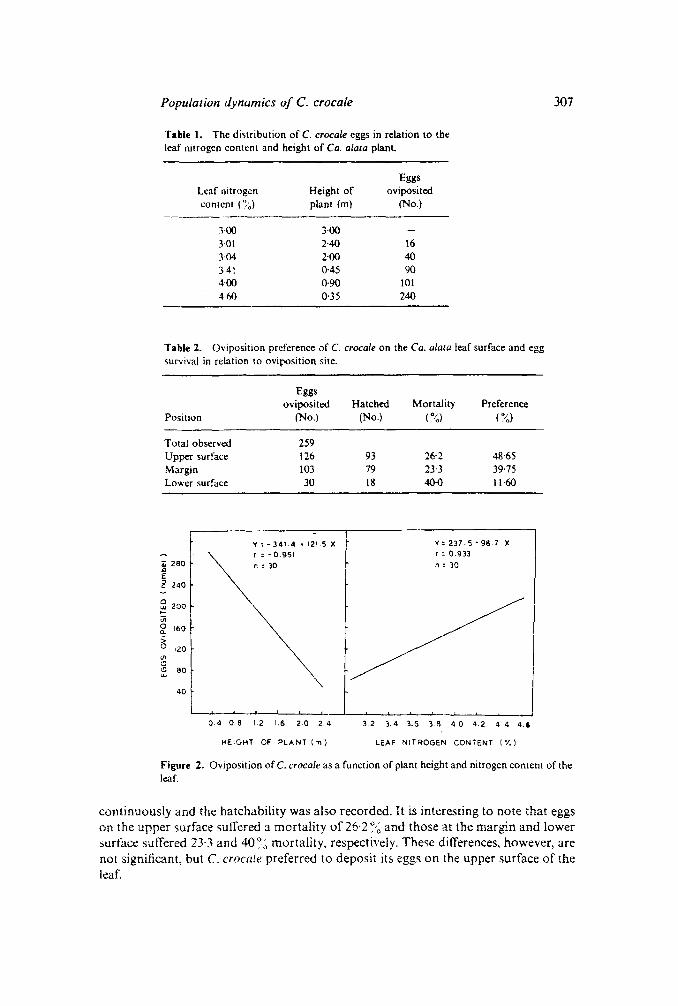
Population dynamics of C. crocale
Table 1. The distribution of C. crocale eggs in relation to theleaf nitrogen content and height of Ca. alala plant.
EggsLeaf nitrogen Height of ovipositedcontent (~~~) plant (m) (No.)
1·00 3·00301 NO 163·04 2·00 40HI 0-45 904·00 (}9O 10141iO 0·35 240
Table 2. Oviposition preference of C. crocale on the Ca. alma leaf surface and eggsurvival in relation to oviposition site.
Eggsoviposited Hatched Mortality Preference
Position (No.) (No.) ( .J,;) ( ~<,)
Total observed 259Upper surface 126 93 26·2 48·65Margin 103 79 23-3 39·75Lower surface 30 18 40·0 11·60
307
.. 280<:IE~ 240
l/lo" 80w
40
Y:-341.4· 121.5 X
~ r: -0.95\
~,."o
~o 4 0.8 1.2 1.5 2·0 2.4
HEIGHT OF PLANT (rn )
Y: 237.5 - 98.7 Xr: 0.933n: 30
3.2 3.4 3.5 3.8 40 4.2 4.4 4'LEAF NITROGEN CONTENT 1"1.)
Figure 2. Oviposition of C. crocale as a function of plant height and nitrogen content of theleaf.
continuously and the hatchability was also recorded. It is interesting to note that eggson the upper surface suffered a mortality of 26·2 ~,~ and those at the margin and lowersurface suffered 23·3 and 40~;) mortality, respectively. These differences, however, arenot significant, but C. crocale preferred to deposit its eggs on the upper surface of theleaf.

308 M S M Christopher and S Mathaoan
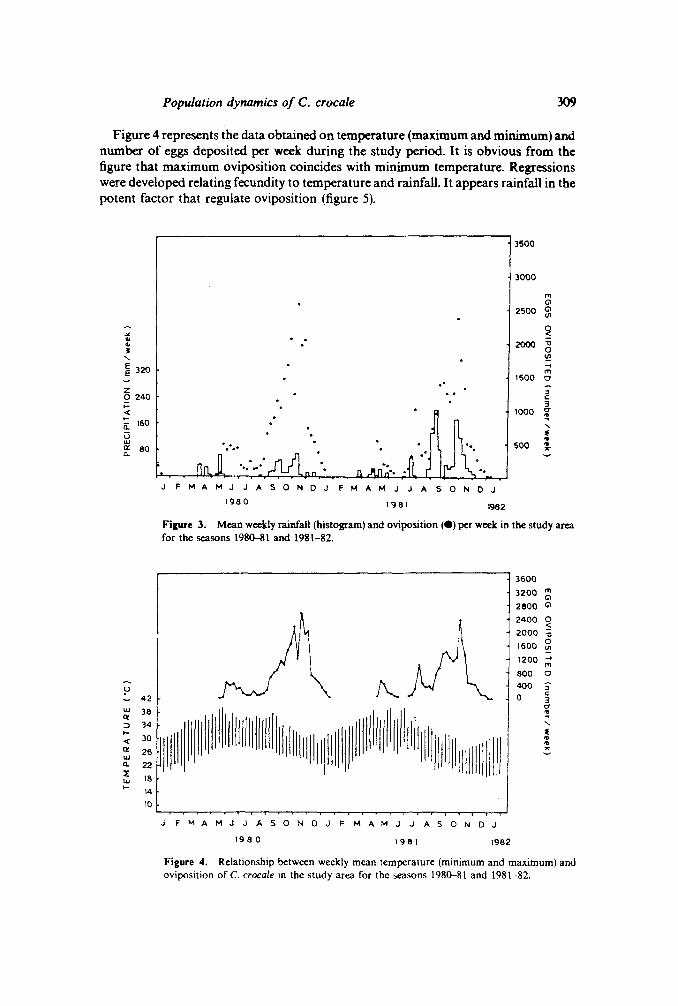
Mean monthly data on rainfall, temperature and humidity recorded in the study areaduring the 25 month period is presented in table 3. From regular field observationsnumber of eggs laid per week was calculated and related to rainfall for thecorresponding period (figure 3). Oviposition was observed from January 1980 toJanuary 1982.It isobvious from the figure that initiation of oviposition follows rains. C.crocale did not oviposit during the months of February, March, April 1980.First rainwas recorded in April 1980. Following that C. crocale initiated oviposition in May. Aprecipitation of about 74mm was recorded during the last week of May 1980 andconsequently the oviposition increased in the following week. Subsequent absence ofrain forced the C. crocale to reduce oviposition during June-August till thecommencement of the northeast monsoon in September. The maximum oviposition ofabout 7869eggswere recorded in the month of November 1980and isclearly related tothe maximum rainfall of 120mm during the same period. Subsequently, as theprecipitation decreased the oviposition also dwindled. Almost a similar pattern of egglaying was observed during 1981-82 season and the maximum was recorded in earlyNovember. From marking and recapture studies it was found that adults emergedduring the end of the season i.e. during January 1980,survived upto April/May withoutmuch reproductive activity during the interim period.
Table 3. Monthly rainfall, temperature and relative humidity in the study area fromJanuary 1980 to January 1982.
Temperature ("C) Humidity (~.-;;)
RainfallMonth (mm) Max Min Max Min
January 1980 0-00 30-88 20-83 75·63 31·70February 0-00 33·25 21·70 90-55 34·68March 0-00 36-00 23-93 84-45 25·95April 68·70 36·65 25·95 88·\0 33-18May 72-30 38·33 27·80 78·68 31·88June 0-00 36·50 27·78 66·13 35·53July 0-00 36·36 27·57 65-80 37'10August 4·00 36-35 26·78 67·28 34·00September 99'04 36-04 26·08 75-33 34·00October 102·00 33-00 24·85 86·48 40-73November 128·00 31'38 24-25 85·78 45·70December 27-40 30-53 22-68 88'13 45·85
January 1981 1'10 31·53 21-30 84-40 33·78February 2'50 33·55 21·33 83-85 28'25March 20-30 35·55 24·45 90-80 32-45April 37·50 37·70 26·98 86·63 32·70May 43·90 37·60 27·\3 79·50 32·65June 0-00 35'10 27·03 75·78 40-90July 110-80 34·87 25·95 78·19 41·38August 26·70 35·21 25-44 77-19 37·35September 424·00 32·95 23-63 92·00 47·\3October 271·90 31·29 23·13 96·55 53·84November 250-20 27-80 21·00 95·41 60·00December 10-40 28·20 20-60 91·85 47·54
January 1982 0-00 31·10 21-00 83·50 33·00
Population dynamics of C. crocale 309
Figure 4 represents the data obtained on temperature (maximum and minimum) andnumber of eggs deposited per week during the study period. It is obvious from thefigure that maximum oviposition coincides with minimum temperature. Regressionswere developed relating fecundity to temperature and rainfall. It appears rainfall in thepotent factor that regulate oviposition (figure 5).
3500
3000
'"Q2500 ~
500
""....J
<,
~320
zo 240>=-ci 160
UuJ
g: 80
..
..
~2000 ;g
(II
....'"1500 0
"c3
1000 g"•....,..
19821981
J FMAMJ JASONDJ FMAMJJASONDJ
1980
Figure 3. Mean weekly rainfall (histogram) and oviposition (e) per week in the study areafor the seasons 1980--81 and 1981-82.
P" ~p}l~ ~ 1I11I11111111111111111111111111111111111111111111111111111/111111111111111111111111111111111111
~ 18... 14
10L-..,--r~~~-..--,--,.---,....-.--r---'---'-~-"--,,--,--"--""""'r-r--.----.-r-~
3600
3200 ~
2800 Q
2400 ~
2000 "1600 ~1200 ....
m800 0
400 "o ~
0..<,
•....,..
J FMAMJ JASON DJ F MAMJ JASON DJ
1980 19111 \982
Figure 4. Relationship between weekly mean temperature (minimum and maximum) andoviposition of C. crocale in the study area for the seasons 1980--81 and 1981-82.
310 M S M Christopher and S Mathauan
1980 - 8\ 1981-82
/'
Y:5381.140Xr : 0.8590
Y : 530. 18 • 43.9 Xr:0.8373
8000 ,-----------.---------
t~ 5000::>c
~ 4000isz~
~ 2000lL
~I-~I
, • i r'
30 50 90 120 100 200 300 400
1980 - 81 RAINFALL (mm ) 1981- 82
Y :15243.1.(-420.31 X Y :13816.(-413 84 X)r:-0.3896 r:-0.5120t
D
3 6000c:
~ 8000
>~ 4000
1,=1L......,.....,..-....---.-...,........,..~_....-"'-..---.,.....,..-..,...-,-~:>~125 27 29 31 33 24 26 28
TEM PERA TURE I' C )
30 32
Figure 5. Regressions showing the relationship between rainfall fecundity and temperatureand fecundity.
Table 4. Correlation between leaf nitrogen content and egg production during theyear 1980.
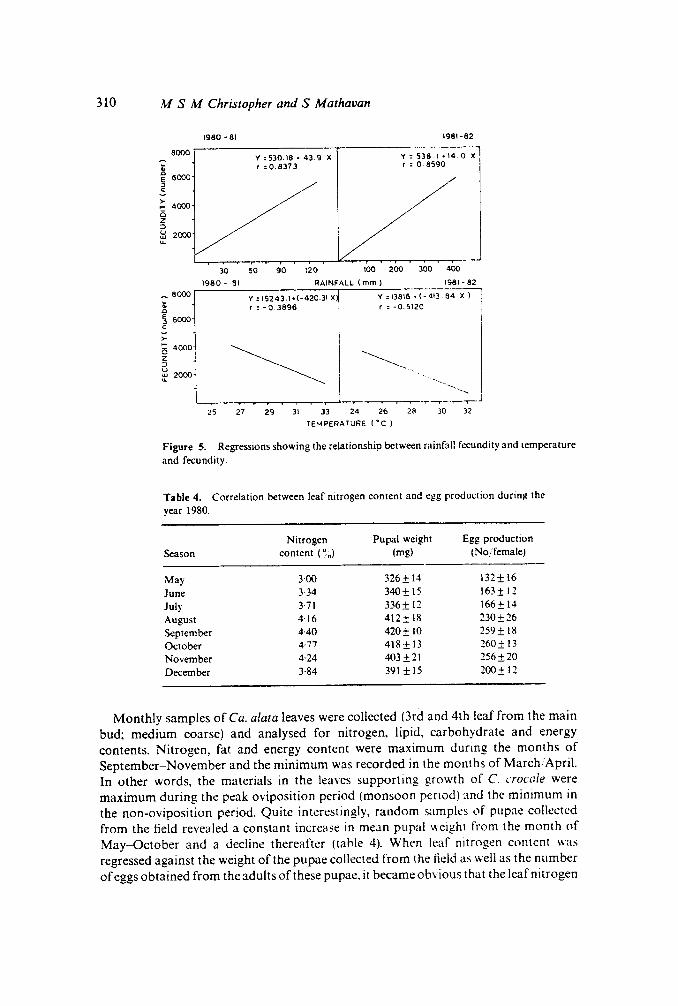
Nitrogen Pupal weight Egg productionSeason content (~o) (mg) (No/femalej
May 3-()() 326± 14 132±16June 3·34 340± 15 163± 12July 3·71 336± 12 166± 14August 4·16 412± 18 230±26September 4·40 420± 10 259 ± 18October 4·77 418 ± 13 260± 13November 4·24 403±21 256±20December 3·84 391 ±15 200± 12
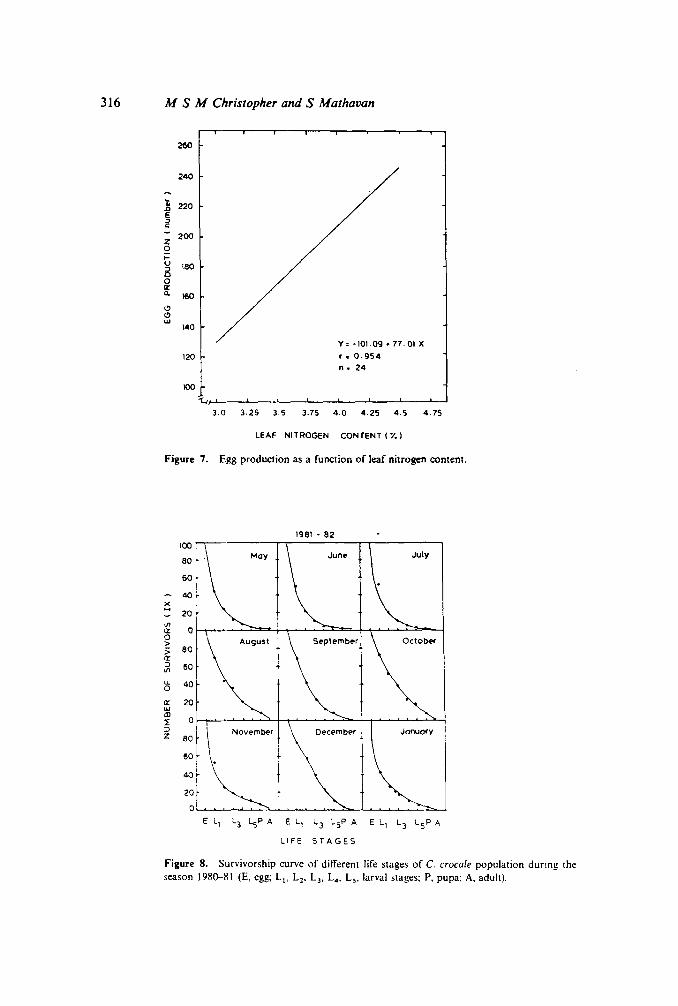
Monthly samples of Ca. alata leaves were collected Ord and 4th leaf from the mainbud; medium coarse) and analysed for nitrogen, lipid, carbohydrate and energycontents. Nitrogen, fat and energy content were maximum during the months ofSeptember-November and the minimum was recorded in the months of March/April.In other words, the materials in the leaves supporting growth of C. crocale weremaximum during the peak oviposition period (monsoon penod) and the minimum inthe non-oviposition period. Quite interestingly, random samples of pupae collectedfrom the field revealed a constant increase in mean pupal weight from the month ofMay-October and a decline thereafter (table 4). When leaf nitrogen content wasregressed against the weight of the pupae collected from the field as well as the numberofeggs obtained from the adults of these pupae. it became obvious that the leaf nitrogen

Population dynamics of C. crocale 311
content is highly correlated with the pupal and the adult weights and consequently withegg production (figures 6 and 7).
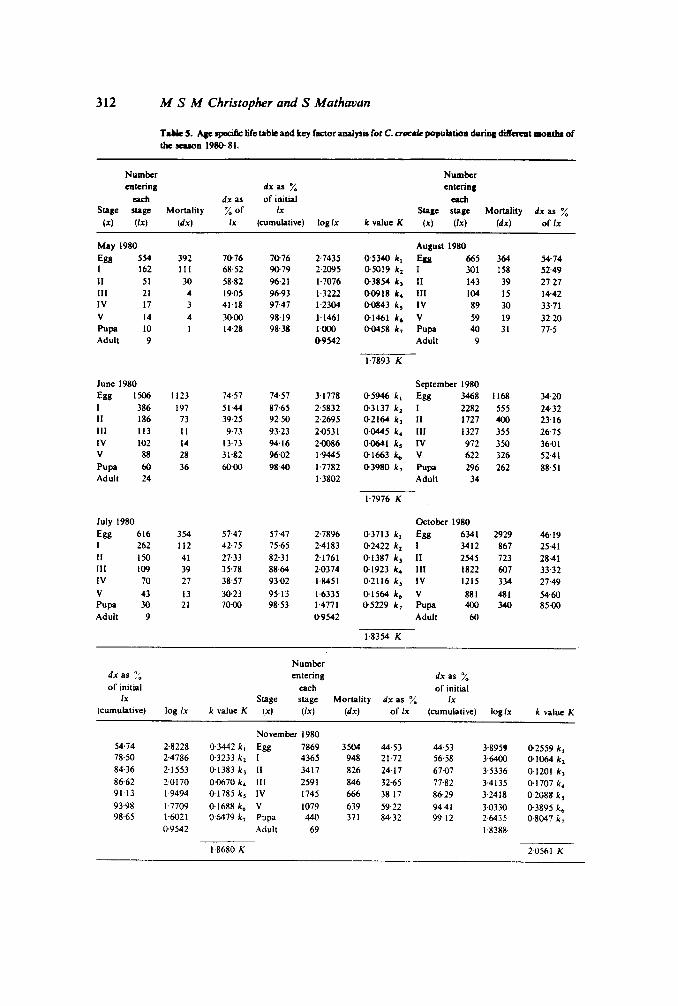
Using the basic data obtained every month on number ofeggs and larvae surviving ineach life stage of C. crocale, a life table was constructed for the seasons 1980-81 and1981-82. First oviposition was observed in the month of May during 1980-81 seasonand in April during 1981-82 season.
Life tables constructed for each month are presented in tables 5 and 6. Based on thelife tables, monthly survivorship curves of C. crocale were obtained for the seasons1980-81 and 1981--82 (figures 8 and 9). It may be seen from the figures that survival wasconstantly low in egg and the I instar larvae throughout the season. Survival of the pupaand final instar was better during the early season than their survival in the middle orlate seasons.
The survivorship curve obtained for C. crocale is a typical concave type; more than70~/~of the animals died before they reached the mid larval stage (Price 1975).From thesurvivorship curve represented for the month of May 1980 and 1981, it is observed thatover 96 /~ of the individuals died before reaching the mid larval stage. With advancingseason the mortality percentage decreased and during the month of December 1980,65 /~ of the individuals died before attaining middle larval stage.
Infertility of egg, feeding of the eggs by ants and the I instar larvae of C. crocale arethe major factors of egg mortality. In the case of I instar in addition to the naturalmortality, ant predators and rainfall are other factors that account for the loss of thelarvae. During the II. III and IV larval stages, the major causes of mortality werepredation by wasps, spiders, mantids. birds and ants. During the final larval and pupalstages the major loss was due to the heavy infection of bacterial and parasitichymenoptera. Birds also prefer to predate upon the fully grown larva. Mortality of
i I
~ 75
Y,133 05 .62.99 X l'r .0 913
n e 24 ~
__L.. L-._J-.._--'----'-._...L-__-L.Jj2'; s s j7C 4,0 425 4.5
.0<)0 ~~
L.t,._.L.3 ,}
320
420
400 .:coE,... sso:I: -:'2w
/~ 360....<{a.::. .140a.
/
.//
cEAF NITROGtN CONTENT ('I. \
Figure 6. Pur:>1 weight :>" ;l Iunct ion of leaf nitrogen content.
312 M S M Christopher and S Mathauan
T.We5. Aae specific life table and key factor analysis for C. CTtICIW population dlIriDa differeDt IBOBtIu of
tbe season 198()--81.
Number Number
entering dx as % enterinl
each dx as of initial eachStage stage Mortality %of lx Stale stage Mortality dx as %
(x) (Ix) (dx) Ix (cumulative) log tx k value K (x) (Ix) (dx) of lx
May 1980 August 1980
Egg 554 392 7Q-76 7Q-76 2-7435 Q-5340 k, Egg 665 364 54-74I 162 III 68·52 90-79 2·2095 Q-5019 k, I 301 158 52-49II 51 30 58·82 96·21 1·7076 Q-3854 k, II 143 39 27·27111 21 4 19·05 96'93 1'3222 0-0918 k. III 104 15 14·42IV 17 3 41·18 97-47 1'2304 0-0843 k, IV 89 30 33·7\V 14 4 30·00 98·19 1·1461 Q-1461 k. V 59 19 32·20Pupa 10 1 14·28 98·38 1·000 0-0458 k7 Pupa 40 31 77-5Adult 9 Qo9542 Adult 9
1·7893 K
June 1980 September 1980Egg 1506 1123 74·57 74·57 3'1778 Q-5946 k, Egg 3468 1168 34·20I 386 197 51·44 87·65 2·5832 0-3137 k, 1 2282 555 24·32II 186 73 39·25 92-50 2·2695 0-2164 k, II 1727 400 23-16III 113 II 9·73 93·23 2·0531 0·0445 k. III 1327 355 26-75IV 102 14 13·73 94·16 2·0086 0·0641 k, IV 972 350 36·01V 88 28 31·82 96·02 1·9445 0-1663 ko V 622 326 52·41Pupa 60 36 60·00 9840 1·7782 0-3980 k 7 Pupa 296 262 88·51Adult 24 1'3802 Adult 34
1·7976 K
July 1980 October 1980Egg 616 354 57-47 57-47 2·7896 0-3713 k, Egg 6341 2929 46·19I 262 112 42·75 75·65 204183 0-2422 k, 1 3412 867 25-41II 150 41 27·33 82-31 2·1761 0-1387 k, II 2545 723 28'41III 109 39 35·78 88·64 2·0374 0-1923 k. III 1822 607 33·32IV 70 27 38·57 93-02 1·8451 0-2116 k, IV 1215 334 27·49
V 43 13 3Q-23 95·13 1·6335 0-1564 k. V 881 481 54·60Pupa 30 21 70·00 98·53 1·4771 0-5229 k ; Pupa 400 340 85-00Adult 9 0-9542 Adult 60
1·8354 K
Numberdx as ~~ entering dx as %of initial each of initial
Ix Stage stage Mortality dx as % lx(cumulative) log lx k value K (x) (Ix) (dx) of lx (cumulative) log lx k value K
November 198054·74 2-8228 0·3442 k, Egg 7869 3504 44·53 44'53 3-895" 0-2559 k,78·50 204786 0-3233 k, I 4365 948 21·72 56'58 3·6400 Q-1064 k,84·36 2·1553 0-1383 k, II 3417 826 24'17 67·07 3·5336 0·1201 k,86·62 2·0170 0·0670 k. III 2591 846 32-65 77-82 N135 0-1707 k.91·13 1·9494 0·1785 k, IV 1745 666 38·17 86·29 3·2418 0·2088 k,93·98 1·7709 0'1688 k. V 1079 639 59·22 94·41 3·0330 0-3895 k.98·65 1-6021 0·6479 k7 Pupa 440 371 84·32 99·12 2·6435 0·8047 k7
0-9542 Adult 69 1·8388-
1'8680 K 20561 K
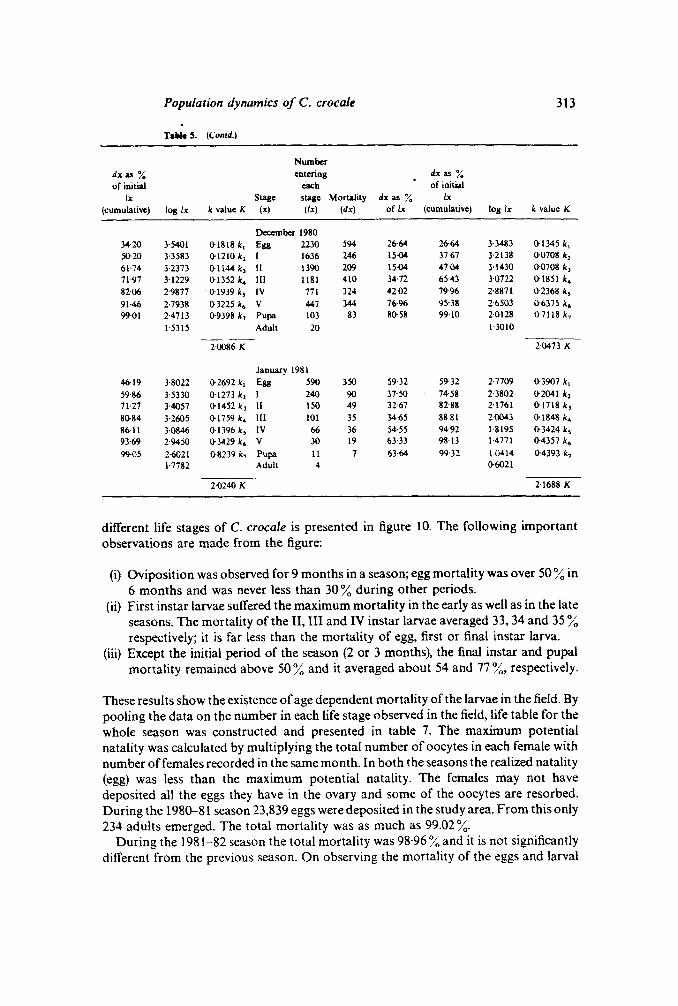
Population dynamics of C. crocale 313
Tallie 5. (Con/d.)
Numberdxas % entering dx as %of initial each of initial
Ix Stage stage MonaJity dx as % Ix(cumulative) log lx k value K (x) (Ix) (dx) of lx (cumulative) log Ix k value K
December 198034·20 3·5401 0-1818 k, Egg 2230 594 26-64 26-64 3·3483 0-1345 k,50-20 3·3583 0-1210 k, 1 1636 246 15·04 3767 3·2138 (}()708 k,61-74 3-2373 0-1l44 k, 11 1390 209 15·04 47·04 3·1430 0·0708 k,71·97 3-1229 0-1352 k. III 1181 410 34·72 65-43 3·0722 0-1851 k.82·06 2·9877 0-1939 k, IV 771 324 42·02 79·96 2·8871 0-2368 k,91·46 2·7938 0-3225 k. V 447 344 76·96 95'38 2'6503 0·6375 k.99·01 H713 0-9398 k, Pupa 103 83 so58 99-10 2·0128 0'7118k,
1·5315 Adult 20 1·3010
2·0086 K 20473 K
January 198146·19 3·8022 0-2692 k, Egg 590 350 59·32 59·32 2·7709 ll-3907 k,59·86 3·5330 ll-1273 k, I 240 90 37'50 74'58 2-3802 0-2041 k,71·27 3·4057 ll-1452 k, 11 150 49 32-67 82-88 2-1761 0-1718k,8ll-84 3-2605 0-1759k. III 101 35 3465 88-81 HlO43 0'1848 k.86·11 3·0846 0·1396 k, IV 66 36 54·55 94'92 1'8195 0-3424 k,93·69 2·9450 ll-3429 k. V 30 19 63-33 98·13 1·4771 0-4357 k.99·05 2-6021 0·8239 k, Pupa II 7 63·64 99'32 1·0414 ll-4393 k,
1·7782 Adult 4 0-6021
2·0240 K 2'1688 K
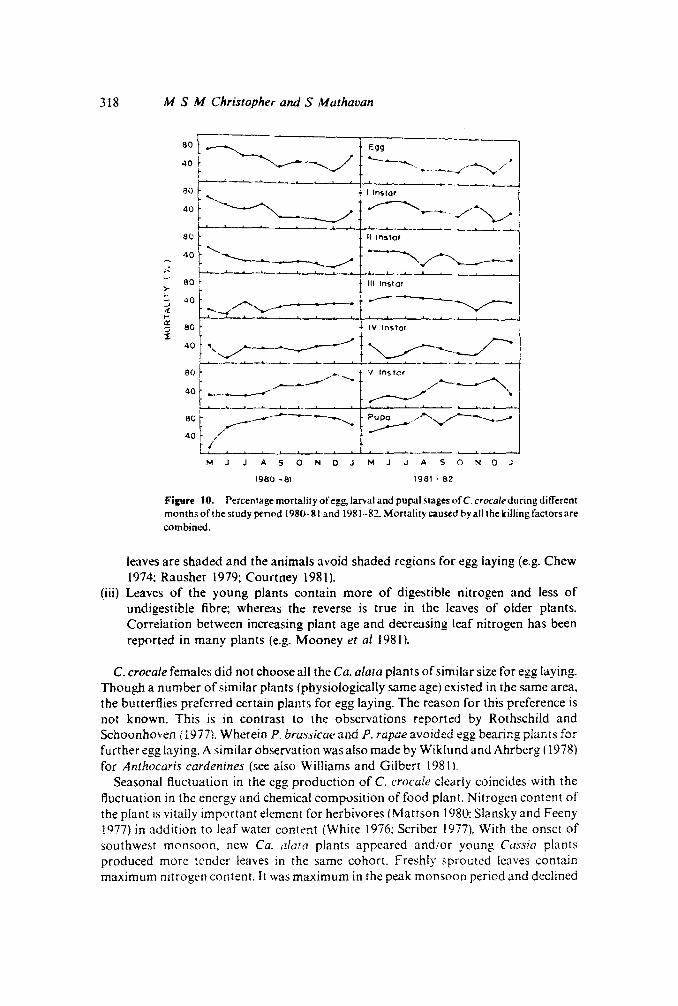
different life stages of C. crocale is presented in figure 10. The following importantobservations are made from the figure:
(i) Oviposition was observed for 9 months in a season; egg mortality was over 50%in6 months and was never less than 30% during other periods.
(ii) First instar larvae suffered the maximum mortality in the early as well as in the lateseasons. The mortality of the II, III and IV instar larvae averaged 33, 34 and 35%respectively; it is far less than the mortality of egg, first or final instar larva.
(iii) Except the initial period of the season (2 or 3 months), the final instar and pupalmortality remained above 50% and it averaged about S4 and 77%, respectively.
These results show the existence ofage dependent mortality of the larvae in the field. Bypooling the data on the number in each life stage observed in the field, life table for thewhole season was constructed and presented in table 7. The maximum potentialnatality was calculated by multiplying the total number ofoocytes in each female withnumber of females recorded in the same month. In both the seasons the realized natality(egg) was less than the maximum potential natality. The females may not havedeposited all the eggs they have in the ovary and some of the oocytes are resorbed.During the 1980-81 season 23,839eggs were deposited in the study area. From this only234 adults emerged. The total mortality was as much as 99.02%.
During the 1981-82 season the total mortality was 98·96%and it is not significantlydifferent from the previous season. On observing the mortality of the eggs and larval
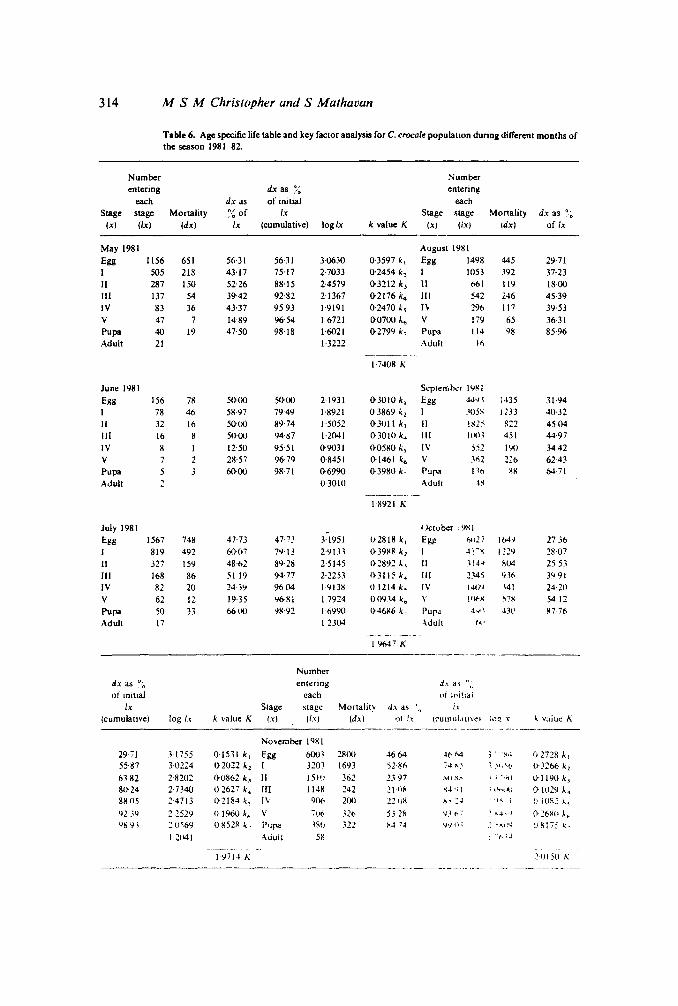
314 M S M Christopher and S Mathavan
T.ble 6. Age specific life table and key factor analysis for C. crocale populauon during different months ofthe season J981-82
Number Numberentering dx as I~ entering
each dx as of initial eachStage stage Mortality ~,~ of lx Stage stage Mortality dx as ~,~
(x) (Ix) (dx) lx (cumulative) log lx k value K (x) (ix) Idx) of lx
May 1981 August 1981
Egg 1156 651 56·31 56·31 3·0630 D-3597 k l Egg 1498 445 29·71
I 505 218 43-17 75·17 2·7033 D-2454 k, I 1053 392 37·23
II 287 150 52-26 88·15 2'4579 D-3212 k, II 661 119 18·00
III 137 54 39·42 92-82 2·1367 0·2176 k. III 542 246 45·39IV 83 36 43·37 9593 1·9191 0·2470 k, 1\1 296 117 39·53y 47 7 14·89 96·54 16721 D-070() i; V 179 65 36·31Pupa 40 19 47·50 98'18 1·6021 D-2799 k, Pupa 114 98 85·96Adult 21 1-3222 Adult 16
----,_._-.1·7408 K
June 1981 September 19~ I
Egg 156 78 50·00 50'00 2·1931 0'3010 k, Egg 44'11 ,·135 31-94
1 78 46 58·97 79·49 1·8921 03869 k, 1 -'05S 12D 40·32
II 32 16 50-00 89·74 1-5052 0·3011 k, 11 !~2:' S22 45-04
III 16 8 50-(J() 94'87 1-2041 D- 3010 k. III IW.l 451 44-97
IV 8 I 12·50 95-51 0'9031 0-0580 k, IV ;~2 190 34-42y 7 2 28-57 96·79 0-8451 0-1461 k. V Yi2 ~2b 6243Pupa 3 60-(J() 98·71 0-6990 D-3980 k· Pupa I1n 88 64-71
Adult 2 0·3010 Adull ~s
---"--_. --18921 K
July 1981 October :9Xl
Egg 1567 748 47-73 4P3 3-1951 0·2818 k, Egg ~!27 164'} 27 36
I 819 492 60·07 79-13 2'91." 0·3988 k, 1 ..n-~ 11:!9 2807
II 327 159 48-02 8928 25145 1}2892 k, II .'! 4\.1 S04 2553
III 168 86 5119 94·77 2·2253 ().J115 k: III 2345 931- 3'191
IV 82 20 14-39 9604 1-9138 01214 k. IV J-lI.N \41 2420
V 62 12 19·35 96·81 17924 0·0934 k. V 1'11-8 ~7x 5412
Pupa 50 33 66(J() 98·92 1-6990 1)'4686 k- PUpJ .1-/,) ..IW 8776
Adult 17 12304 -\dull I.'
19647 K-~------
Numberdx as (J,~ entering d.r. as ~:.
of mitial each of witiai
lx Stage stage Mortality dx as '1" lx
(cumulative) log lx k value K (x) (Ix) (dxl 01 tx {cumuta nvet !nQ T ~. value K-----._.----_.-- ..• _---_.._- _._._-------
November 1981
2971 31755 0·1531 k, Egg 6003 28(J() 4664 ..II> 1>4 " .'::-;., () 2728 i,
55·87 3-02:4 O'2022 k, [ 3203 1693 52-81\ i4 "-.' ;.".'-.t> 0-3266 k,
63-82 28202 0'0862 k.\ II 1510 362 23 97 ;.oIl~k ;.! ',itl U-llIJO k.,81}24 2·7340 02627 k. 1lI J 1..18 242 21"~ 1'\4'~ t .. \~'t~j 01029 k•8805 24713 0·218..1 k. IV 9()(' 2(~) n'i8 ~."I :.J ')0\ i o iO~'; ;..~
92.19 22529 o 1960 k. V 7\.to 321> 5;28 1.1.1,...: :-" ......J (1-:1'80 k.9S9} 20569 IJ8528 k, PIJpa iso 322 84 74 'I'Jtl' .~ "Xl ;I,i u·8175 ,,-
121141 A<iult 58 : .'t-.;.1.
[y7 It K ,O! 50 K--------"-------.._--- . --------

Population dynamics of C. crocale 315
Table 6. (Coned_)
Numberdx as ?Ict entering d."( as ~~
of initial each of initiallx Stage stage Mortality dx as ~~ lx
(cumulative) log tx k value K (x) (Ix) (.Ix) of Ix (cumulative) log x k value K
December 1981
31-94 3-6525 0-1671 k, Egg 1903 504 26-48 26-48 3-2794 0-13.16 k,59-38 H854 0-2241 k, 1 l.199 291 20-28 4178 ll458 O-HJl3 k,77-68 3-2613 Q-2600 k, II 1108 368 33-21 61-11 1-0445 o 1753 k,87-71 3-0013 0-2594 k. III 740 310 41-89 7NO 2-81>92 0-2357 k.91-94 2-7419 0-1832 k, IV 430 240 5581 9O-\l! 26335 0-3547 k,96-97 2-5587 04252 k. V 190 126 6632 96-64 227MM 0-4726 k.98-93 2-1335 0-4523 k, Pupa 64 47 73-44 99-10 18062 0-5758 k,
1-6812 Adult 17 1-2304----_.-.-- ------ -
1-9713 K 20490 K
'anuary 1982
27-36 3-7801 0-1388k, Egg 504 281 55-75 55-75 2-7024 U-3541 k,4775 36413 0-1431 k, I 223 82 36-77 nU2 23483 0-1991 k,
6109 3-4982 0-1281 k I II 141 45 31-91 80-95 21492 U-1689k,7662 3-3701 U2212 k. 111 96 36 3750 8809 19823 02041 k.8228 3-1489 Q-12U3 k, IV 60 29 4833 93-85 17782 0-2868 k,9187 3-0286 0-JJ84 k. V 31 10 32.16 95K3 1-4914 0-1692 k.99-00 2-6902 U9120 k, Pupa 21 17 80-95 9921 I-un 0-7201 k,
1-7782 Adult 4 v-6021~- ~._----
-_._-_.2-0019 K 2-ltJ03 K
stages of 1980--81 season, It became apparent that about 40 ~/o of the eggs failed to hatch.The mortality in the first 4 larval stages averaged to about 30 ~{) while the V instar andpupae suffered 57 and 83 ~~. respectively. During 1981-82 season it ranged from 36 ~~ inthe egg to 30 ~<, in the IV instar. However, V instar (5(}l5 o/~) and pupae (81 0 0) showedthe maximum percentage of mortality. This clearly depicts the low survivorship of thefinal instar and pupae for the season 198Q..8 t and 1981-82. Analysis of k factor revealsthat the maximum k value of above 0·7 was observed for the pupal stage III both theseasons and the second maximum was observed for the V instar (about 0-3).
4. Discussion
Even though females of C. crocale were confronted with an array of host plants such asCa. fistula, Ca. siamea, Ca. jal'anica and Ca. grand is. they restricted oviposition to theleaves of Ca. alata which appears to be a case of'rnonophagic strategy'. Fernales clearlypreferred to oviposit on the leaves of Ca. ulaia and avoided plants belonging to otherCassia species. though these plants are present in a similar topography along with Ca.aluta. This type of rnonophagic strategy is found in many oligophagous butterflies suchas P machaon (Wiklund 1974.1975). E_ editha (Singer 1971; Holdren and Ehrhch 19~2)
and Heliconius meinom-m» (Smiley t 9781. Slobodk in and Sanders i 1Y(9) and Wiklund\1(74) stated that rnonophagic strategy would be favoured by species living In
316 M S M Christopher and S Mathavan
260
240
! 220E
""z 200
0...U 180a0II:Q. 160
""... 140
Y= -101.09.77.01 X
120 r.0.954n. 24
100't.,!-'--_-.L._---'-_--..L_--'__'--_.L-_-'--'
3.0 3.25 3.5 3.75 4.0 4.25 4.5 4.75
LEAF NITROGEN CONTENT (Y.)
Figure 7. Egg production as a function of leaf nitrogen content.
1981 - 82100
80July
60
40)(- 20inQ: 00>s 80ce:J !O<Jl
u,0
ce...III1::JZ 80
!O
4O~II
20 r0
E' L1 L3 LSP A E L1 L3 LSP A E II l3 LSP A
LI FE STAGES
Figure 8. Survivorship curve of different life stages of C. crocale population during theseason 1980--81 (E, egg; t., L2, LJ , L4 , t.; larval stages; P, pupa: A, adult).

Population dynamics of C. crocaJe
1980 - 81100r-r-----r------,-,-----...,80
60
;; 40
l/IIXo>>IX::ll/I
u.oII:....cD~::lZ
E LI L3 LSP A E L1 L3 Ls P A E LI L3 ls P A
LIFE STAGES
317
Figure 9. Survivorship curve of different life stages of C. crocale population during theseason 1981-82 (E, egg; L t , L z, L 3, L., L,. larval stages; P, pupa; A, adult).
predictable environments (habitats) which allow specialization to narrow niches.Moreover oligophagous specieshaving adopted the monophagic strategy will predominantly utilize perennial plant species or species which are abundant in the habitat(Wiklund 1974).
Similarly, Smiley (1978) and Gilbert (1978) suggested that monophagic strategyobserved in the butterfly H. melpomene might have originated, since food plants mightdiffer in their suitability for oviposition, larval growth and survival or for reasons otherthan biochemical or nutritional differences. In addition, ecological factors such aspredation might have affected the evolution of monophagy in these butterflies. Holdrenand Ehrlich (1982)concluded that the monophagic strategy found in E. editha towardsCastilleja linarifolis plant is not because of their biochemical make-up but because oftheir ecological characteristics, primarily their phenologies. Courtney (1983) hasfurther showed that evolution of monophagous relationship is possible even whenalternative host plants are present. The oviposition choice of e.crocale females seems toagree with the Hopkins host selection principle (Hopkins 1917); it states that a female'soviposition preference is biased in favour of the plant species which she fed upon aslarva.
e. crocale females identified young Ca. alata plants for potential egg laying andcarefully avoided the older (taller) plants. It may be due to the following reasons.
(i) The young plants have more number of tender leaves and thus supported thefreshly emerged larvae better than the older plants.
(ii) In small plants, the young tender leaves are exposed to sunlight for a maximumperiod of time in a day and attract ovipositing females. In older plants, the best
318 M S M Christopher and S Mathaoan
MJJASONDJMJJASONDJ
1980 - 81 1981 ·82
figure to. Percentage mortality of egg, lanai and pupal stages ofC.crocaleduring differentmonths of the study period 1980-81 and 1981--82. Mortality caused by all the killing factors arecombined.
leaves are shaded and the animals avoid shaded regions for egg laying (e.g, Chew1974; Raushcr 1979; Courtney 1981).
(iii) Leaves of the young plants contain more of digestible nitrogen and less ofundigestible fibre; whereas the reverse is true in the leaves of older plants.Correlation between increasing plant age and decreasing leaf nitrogen has beenreported in many plants (e.g. Mooney et al 1981).
C. crocale females did not choose all the Ca. alata plants of similar size for egg laying.Though a number of similar plants (physiologically same age) existed in the same area,the butterflies preferred certain plants for egg laying. The reason for this preference isnot known. This is in contrast to the observations reported by Rothschild andSchoonhoven (1977), Wherein P. brassicae and P. rapae avoided egg bearing plants forfurther egg laying. A similar observation was also made by Wiklund and Ahrberg (1978)for Anthocaris cardenines (see also Williams and Gilbert 1981.1-
Seasonal fluctuation in the egg production of C. crocale clearly coincides with thefluctuation in the energy and chemical composition of food plant. Nitrogen content ofthe plant is vitally important element for herbivores (Mattson 1980: Siansky and FeenyJ977) in addition to leaf water content (White 1976: Scriber 1977). With the onset ofsouthwest monsoon, new Ca. alma plants appeared and/or young Cassia plantsproduced more tender leaves in the same cohort. Freshly sprouted leaves containmaximum nitrogen content. It was maximum in the peak monsoon period and declined

Population dynamics of C. crocale 319
Table 7. Age specific life table and key factor analysis for C. croca/e during 1980-81 and1981 -82 seasons
dx as o_~ ofdx as o~ initial/x log
X tx dx of /x (cumulative) value k value k------_._,---------
1980--81PN 55534Egg 23839 10796 45-29 45·29 4-3773 (}2619 k,Instar I 13043 3284 25·18 59·06 4-1154 o1260 k2
lnstar II 975IJ 2390 24·49 69·09 3-9894 (}1220 k J
Instar III 7369 2322 31·51 78-83 3-8674 (}1644 k4
Instar IV 5047 1784 35·35 86·31 3-7030 0·1894 ksInstar V 3263 1873 57-40 94·17 3·5136 0·3706 k.PUl'a 1390 1156 83·16 99-02 3-1430 (}7738 k7
Adult 234 2-3692
2·0081 K
1981-82PN 566~O
Egg 23396 8649 36·97 36·97 4'3691 (}2004 k,Instar I 14747 5693 38·60 61-30 4'1687 (}21 (9 k2
Instar II 9054 2854 31·52 73-50 3·9568 (}1644 k]Instar III 620(l 2)71 38-24 83·63 3·7924 0·2093 k4
Instar IV 3829 1177 3(}74 88-66 3-5831 (}1595 ksInstar V 2652 1352 50·98 94-44 J4236 0·3097 k.Pupa 1300 1057 81·31 98·96 3·1139 (}7283 k,Adult 243 2·3856
1·9835 K
PN, Potential natality.
as the rainy season declined. As the plants started flowering, maximum nitrogen waschannelled for reproductive part of the plants. Mooney et al (1981) reported that leafnitrogen of the Californian shrub Deplacus aurantiacus declined as the floweringcommenced. A similar case is true for Ca. alata. It is assumed that leaf quality of Ca.alata determines the reproductive potential ofC. crocale. Wiklund and Person (1983)showed that females of the speckled wood butterfly, Pararge aegeria lay eggs thatdecrease in size over the oviposition period. However, we did not make suchobservation during the oviposition of C. crocale.
In southern India, the southwest monsoon starts from April and continues to Augustand the northeast monsoon commences in September extending to December. Duringthe 1980-81 season, the first rain (29 mm) was recorded on 5th April 1980. Followingthis precipitation, the first oviposition was observed on 4th May 1980. During thesouthwest monsoon total precipitation was 145mm. Temperature ranged between amaximum of 37' e and a minimum of noeand the humidity between 73 and 34 %rhoTotal number of eggs produced during the same period was 3341. It represents 14%ofthe total egg production or the 198(}-81 season. During the same year the northeastmonsoon brought about 356 mm rain and 83 ~o of the total egg production was
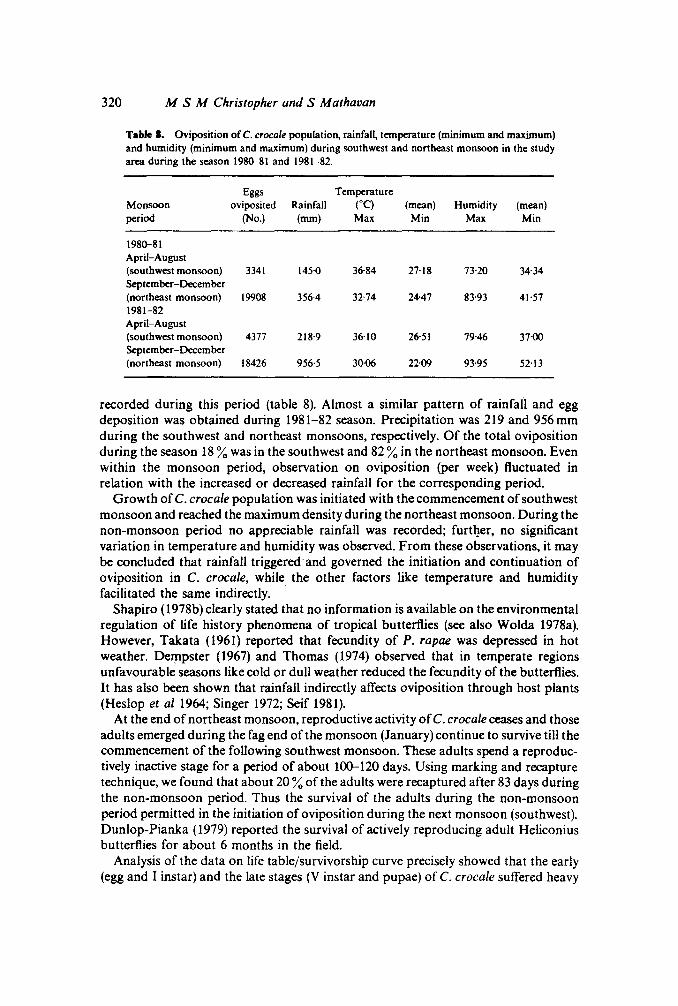
320 M S M Christopher and S Mathaoan
Table I. Oviposition of C. crocale population, rainfall, temperature (minimum and maximum)and humidity (minimum and maximum) during southwest and northeast monsoon in the studyarea during the season 1980--81 and 1981-82.
Eggs TemperatureMonsoon oviposited Rainfall (0C) (mean) Humidity (mean)period (No.) (mm) Max Min Max Min
1980--81April-August(southwest monsoon) 3341 145·0 36·84 27·18 73·20 34·34September-December(northeast monsoon) 19908 356'4 32·74 24-47 83-93 41'571981-82April-August(southwest monsoon) 4377 218·9 36·10 26·51 79·46 37·00September-December(northeast monsoon) 18426 956·5 30·06 22·09 93-95 52·13
recorded during this period (table 8). Almost a similar pattern of rainfall and eggdeposition was obtained during 1981-82 season. Precipitation was 219 and 956mmduring the southwest and northeast monsoons, respectively. Of the total ovipositionduring the season 18%was in the southwest and 82 %in the northeast monsoon. Evenwithin the monsoon period, observation on oviposition (per week) fluctuated inrelation with the increased or decreased rainfall for the corresponding period.
Growth ofC. crocale population was initiated with the commencement ofsouthwestmonsoon and reached the maximum density during the northeast monsoon. During thenon-monsoon period no appreciable rainfall was recorded; further, no significantvariation in temperature and humidity was observed. From these observations, it maybe concluded that rainfall triggered and governed the initiation and continuation ofoviposition in C. crocale, while the other factors like temperature and humidityfacilitated the same indirectly.
Shapiro (1978b) clearly stated that no information is available on the environmentalregulation of life history phenomena of tropical butterflies (see also Wolda 1978a).However, Takata (1961) reported that fecundity of P. rapae was depressed in hotweather. Dempster (1967) and Thomas (1974) observed that in temperate regionsunfavourable seasons like cold or dull weather reduced the fecundity of the butterflies.It has also been shown that rainfall indirectly affects oviposition through host plants(Heslop et al 1964; Singer 1972; Seif 1981).
At the end ofnortheast monsoon, reproductive activity ofC. crocale ceases and thoseadults emerged during the fag end of the monsoon (January) continue to survive till thecommencement of the following southwest monsoon. These adults spend a reproductively inactive stage for a period of about 100-120 days. Using marking and recapturetechnique, we found that about 20 %of the adults were recaptured after 83 days duringthe non-monsoon period. Thus the survival of the adults during the non-monsoonperiod permitted in the initiation of oviposition during the next monsoon (southwest).Dunlop-Pianka (1979) reported the survival of actively reproducing adult Heliconiusbutterflies for about 6 months in the field.
Analysis of the data on life table/survivorship curve precisely showed that the early(egg and I instar) and the late stages (V instar and pupae) of C. crocale suffered heavy

Population dynamics of C. crocale 321
mortality during the entire study period of the 1980-81 and 1981-82 seasons. Keyfactor analysis (k) was followed to estimate age specific mortalities (Morris 1959; Varleyand Gradewelll960; Hayes 1981).These age specific mortalities (k) were added togetherto give the total generation mortality (K). Such analysis revealed that k values alwaysremained high for egg and I instar as well as for V and pupal stages. Observation ofHayes (1981) for estimating the k values for the pierid butterfly Colias alexandra, wassimilar to the one observed in the present study. He reported that egg, diapausing larvae(final instar) and post diapausing larvae suffered the maximum mortality during theextended study period of 5 years. Working on the pierid butterfly Anthochariscardamines,Courtney (1981) showed that these polyphagous larvae fed and survived ona variety of cruciferous food plants; irrespective of larval host plant, maximummortality was obtained during egg, early and late larval instars. Egg mortality was dueto cannibalism; late larvae and pupae of A. cardamines suffered a maximum of about25-40 %due to bacterial attack and 40 % due to parasitism.
Wolda and Foster (1978) reported an outbreak of Zunacetha annulata in aneotropical forest on Barro Colorado Island during 1971-73 and showed that theoutbreak was very effectively ended by fungus and/or either bacterial or viral disease.They have found that 90 %of the pupae were killed by these factors and suggested theexistence of density dependent factor which brought the numbers down whenever thelarval population exceeded certain level.
Mathavan (1975) reported the incidence of a tachinid Strumia concerqens on apopulation of the monarch butterfly Danaus chrysippus during 1973-74 and showedthat about 15% of the pupae were killed by the parasite. Pandian and Delvi (1973 j
showed that about 11%of the grasshopper P. pictus population was destroyed by thesarcophagid fly Blaesoxipha kaestneri. Normally, both D. chrysippus and P. pictus aresubjected to limited predator/parasite mortality, since they feed on the Calotropis plantand accumulate the carotenoid poison in their body (Brower 1969). Hence thepercentage mortality due to parasite in P. pictus and D. chrysippus was minimal whencompared to the data available in the literature. In the tropical moth Achea janata,Muthukrishnan (1980) observed about 70-80 %of the population was reduced by thehymenopteran parasite Microplitis ophiusa (see also Nayar et al 1976). Presenting thelife tables of caterpillar Hemileuca oliviae, Henson et al (1982) concluded that withoutparasitism, caterpillar density remained constant after a 70 ~~ minimum loss in the earlystages. However, the occurrence of techinid parasite and predation decreased thedensity to an insignificant level.
C. crocale is a continuous breeder, and hence mortality per generation cannot befixed for the population in the field. However, percentage ofadults surviving in a seasoncan be calculated. The calculation thus made revealed that, of the total eggs laid in theseason about 1 o.~ or less than 1 'j~ successfully emerged as adults in the 1980- 81 and1981 -82 seasons. McClure (1981) found that in the homopteran populations ofFiorinia externa and Tsugaspidiotus tsuqae, the generation mortality ranged from93-99°~. Investigating the key mortality factors of Heliothis armigera. Bilapate (1981)reported a generation mortality of99·2 (~o and the major killing factors the populationare the parasites and predators. A similar generation mortality was also observed in thepierid butterfly C. alexandra. which ranges from 97·1 0'0 in the year 1975 to 99·5 0" in theyear 1978 (Hayes 1981). Latheef and Harcourt (1974) reported that the Coloradopotato beetle Leptinotarsa decemlineata population suffered 99 "" mortality in thepotato fields of eastern Ontario.

322 M S M Christopher and S Mathavan
Many studies have shown that insects in the tropics are seasonal (GoeI1976; Wold a1977, 1978b). Mostly, the fluctuation pattern is related 10 rainfall and wet and dryseasons. Working on the leaf hoppers in Panama, Wolda (1980) reported that in mostspecies of Homopterans the maximum abundance peak was observed in the rainyseason and the minimum in the dry season. This is very similar to the population ofC. crocale. Moreover, seasonality of insect species is synchronized with the seasonalpresence of its food (Wolda 1978b). Such a type of fluctuation related to rainfall is notuncommon in the tropics.
Wolda (l978a) clearly showed that such a type of fluctuation is found both in tropicaland temperate zones. C. crocale population fluctuation is clearly influenced by themagnitude ofrainfall during ditTerentmonths (dry and wet season) of the season. As theincidence ofhymenopteran entomophagous parasite and bacteria fluctuates along withthe C. crocale population it may be concluded that these factors influence C. crocalepopulation considerably. Similar controlling factors for the population regulation havebeen reported (Wolda and Foster 1978; Harcourt 1971). According to Varley andGradwell (1970) such fluctuation in the population should not be unexpected sincemortality factor may not remain constant throughout the whole range of populationdensity. Ehrlich (1965) also emphasized the fact that one factor which is extremelyimportant in the dynamics of E. editha population is parasitism in addition to themicroclimatic changes of the area.
The unfavourable environmental situations such as low rainfall, high temperatureand piling up of heavy predation pressure plus maximum parasitic load resulted in thedecline of C. crocale population to a minimum level. Decline in the density of the hostresulted in the disappearance of the controlling factors. Ultimately, C. crocale adultssurvived during the non-seasonal period without much predation pressure and beganto initiate the population build up with the commencement of the subsequentmonsoon. The foregoing discussion clearly shows that though the environmentcontrols the reproductive success of the butterfly, the survival and continuouspropagation of the population mainly depend upon the load of parasitism andpredation on the larval population.
References
Beeson C F C 1941 The ecology and control ofthe forest insects ofIndia and the neighbouring countries (Govtof India)
Bilapate G G 1981 Investigations on Heliothis armiqera (Hulirer) in Marathwada-XXIII. Key mortalityfactors on cotton, pigeon pea and chick pea; Proc. Indian Nat. Sci. Acad. 847 637-646
Blau W S 1980 The effect of environmental disturbance on a tropical butterfly population; Ecology 611005-1012
Browne F G 1968 Pests and diseases offorest plantation trees (Oxford: Clarendon Press)Brower L P 1969 Ecological chemistry; Sci. Am. 220 1-8Chew F S 1974 Strategies offood plant exploitation in a complex of oligophagous butterflies (Lepidoptera),
Ph.D. thesis, Yale University, New Haven, Connecticut, USAChew F S 1975 Coevolution of pierid butterflies and their cruciferous food plants. I. The relative quality of
available resources; Oecologia 20 117-127Courtney S P 1981 Coevolution of pierid butterflies and their cruciferous food plants. III Anthocharis
cardamines (L) survival, development and oviposition on different host plants; Oecoloqia 51 91-96Courtney S P 1983 Models of host plant location by butterflies; the effect of search images and search efficiency;
Oecologia 59317-321

Population dynamics of C. crocale 323
Dempster J P 1967 The control of Pieris rapae with DDT I. The natural mortality of the young stages ofPieris; J. Appl. Eco!. 4485-500
Duffey E 1968 Ecological studies on a large copper butterfly, Lycaena disper; Haw. batayus Obth. atWoowalton Fen National Nature Reserve, Huntingtonshire; J. App!. Ecol. 569-96
Dunlop-Pianka H L 1979Ovarian dynamics in Heliconius butterflies: Correlations among daily ovipositionrates, egg weights, and quantitative aspects of oogenesis; J. Insect Physiol. 25 741-749
Ehrlich P R 1965The population biology of the butterfly Euphydryas editha. The structure of the Jasper ridgecolony; Evolution 19 327-336
Ehrlich P Rand Gilben L E 1973 Population structure and dynamics of the tropical butterfly Heliconiusethilla; Biotropica 5 69-83
Gilbert L E 1978 Development of theory in the analysis of insect-plant interactions; in Analysis ofEcologicalsystems. (ed.) D Hom, R Michel1and G Stairs (Columbus: Ohio State University Press) 117-153
Gilbert L E and Singer M C 1975 Butterfly ecology; Annu. Rev. Eco!. Syst. 6 365-397Goel S C 1976 An ecological study of the Hemipteran catch on the light trap at MuzafTarnagar, India; Trap.
Ecol. 17 89-94Harcourt D G 1966 Major factors in survival of the immature stages of Pieris rapae (L.); Can. Eniomol. 98
653-662Harcourt D G 1971 Population dynamics of Leptinotarsa decemlinecta (say) in eastern Ontario III. Major
population process; Can. Entorool. 103 1049-1061Haldane J B 1949 Disease and evolution; Ric. Sci. 193-11Hayes J L 1981 The population ecology of a natural population of the pierid butterfly Colias alexandra;
Oecolagia 49 188-200Heslop I P R, Hyde D E and Stockley R E 1964 Notes and viewsof the purple emperor (Brighton: Southern)
248Holdren C E and Ehrlich P R 1982Ecological determinants of food plant choice in the checkerspot butterfly
Euphydryas editha; Oecologia 52 417-423Hopkins AD 1917 A discussion of CG Hewitt's paper on "Insect Behaviour"; J. Econ. Entorool. 1092-93Henson J D, Owens J C and Huddleston E W 1982 Life tables of the range caterpillar, Hemileuca oliviae
(Lepidoptera: Saturnidae); Environ. Entorool. 11 355-360Lakhani K H and Dempster J P 1981 Cinnabar moth and its food plant; Further analysis of a simple
interaction model; J. Anim. Ecol. 50 231-249Latheef M A and Harcourt D G 1974 The dynamics of Leptinotarsa decemlineata; Enromol. Exp. Appl. 17
67-76Mathavan S 1975 Observation on the mortality of the monarch butterfly Danaus chrysippus (Linnaeus)
infected by Sturmia convergens (Wiedemann) (Diptera: Tachinidae); Curro Sci. 44 554--555Mattson W J 1980 Herbivory in relation to plant nitrogen content; Annu. Rev. Ecol. Syst. 11 119-161McClure M S 1981 Effects of voltinism, interspecific competition and parasitism on the hemlock scales,
Fiorinia externa and Tsugaspidiotus tsuqae (Homoptera: Diaspididae); Ecol. Entomol. 647-54Mooney H A, Ehrlich P R, Lincoln D E and Williams K S 1980 Environmental controls of the seasonality of
the drought deciduous shrub, Diplacus aurantiacus; Oecologia 50 195-198Mooney H A. Williams K S, Lincoln D E and Ehrlich P R 1981 Temporal and spatial variability in the
interaction between the checkerspot butterfly, Euphydryas cholcedona and its principal food source, theCalifornia shrub, Diplacus auranriacus; Oecologia 50 195-198
Morris R F 1959 Single-factor analysis in population dynamics; Ecology 40 58(}-588Muthukrishnan J 1980 Physiological studies on chosen arthropods, Ph.D. thesis, Madurai Kamaraj
University, MaduraiNayar K K, Ananthakrishnan T N and David B V 1976 General and applied entomology; (New Delhi: Tata
McGraw-Hill Pub\. Co. Ltd)Pandian T J and Delvi M R 1973Observation on the mortality of population of the grasshopper Poecilocerus
pictus infected by sarcophagous parasite Blaesoxipha kaestneri; Indian J. Entomol. 35 50-52Price P W 1975 Insect Ecoloqy (New York: John Wiley)Rausher M D 1979 Larval habitat suitability and oviposition preference in three related butterflies; Ecology
66 503-511Rothchild M and Schoonhoven L M 1977 Assessment of egg load by Pieris brassicae (Lepidoptera: Pieridae);
Nature (London) 266 352-355Scriber J M 1977 Limiting effects of low leaf water content on the nitrogen utilization, energy budget and
larval growth of Hyalophora cecropia (Lepidoptera: Satyrinae); Oecoloqia 28 269-287

324 M S M Christopher and S Mathavan
Seif A A 1981Seasonal fluctuation ofadult population of the whitefly, Bemisia tabaci on cassava; Insect Sci.Appl. 1 363-364
Shapiro A M 1978a Developmental and Phenotypic responses to photoperiod and temperature in anequitorial montene butterfly Tatochila xanthodice (Lepidoptera: Pieridae); Biotropica 10297-301
Shapiro A M 1978b Weather and the lability of breeding populations of the chekersp.ot white butterfly, Pierisprotodice; J. Res. Lepid. 17 1-23
Singer M C 1971Evolution of food plant preference in the butterfly Euphydryas editha; Evolution 25 383-389Singer M C 1972 Complex components of habitat suitability within a butterfly colony; Science 173 75-77Slobodkin L B and Sanders H L 1969 On the contribution of environmental predictability to species
diversity. In Diversity and stability in ecological systems; Brookhaven Symp. Bioi. 22 82-95Smiley J 1978 Plant chemistry and the evolution of host specificity, New evidence from Heliconius and
Passifiora; Science 201 745-747Siansky F Jr and Feeny P 1977 Stabilization of the rate of nitrogen accumulation by larvae of the cabbage
butterfly on wild and cultivated food plants; Ecol. Monogr. 47 209-228Takata N 1961Studies on the host preference of common cabbage butterfly-XlI; Jpn. J. Ecol. 11 141-154Thomas J A 1974Autecoloqical studies ofhairstick butterflies, Ph.D. thesis, Univ Leicester,Leicester,EnglandVarley G C and Gradwell G R 1960 Key factors in population studies; J. Anim. Ecol. 29 399-401Varley G C and Gradwell G R 1970 Recent advances in insect population dynamics; Annu. Rev. Entomol. 15
1-24Vats L K and Kaushal B R 1980 Fluctuations of lepidopteran populations, secondary productivity and
energy flow through Belenois mesentina (Pieridae); Aqro-Ecosystems 6161-176Watt W B, Chew F S, Snyder L R G, Watt A G and Rothchild D E 1977 Population structure of pierid
butterflies. I. Densities and movementsof three montene Colias species; Oecoloqia 27 1-22Watl W B, Han D and Tabashrik B E 1979 Population structure of pierid butterflies. II. A "native"
population of Colias phi/odice eriphyle in Colorado; Oecoloqia 44 44-52White T C R 1976 Weather, food and plagues of locusts; Oecoloqia 22 119-134Wiklund C 1974The concept of oligophagy and the natural habitats and host plants of Papilio machaon in
Fennoscandia; Entomol. Scand. 5 151-160Wiklund C 1975 The evolutionary relationship between adult oviposition preference and larval host plant
range in Papilio machaon; Oecoloqia 18 185-197Wiklund C and Ahrberg C 1978 Host plants, nector source plants and habitat selection of malesand females
of Anthocharis cardamines (Lepidoptera); Oikos 31 169-183Wiklund C and Person A 1983Fecundity and the relation of egg weight variations to offspring fitness in the
speckled wood butterfly Pararge aeqeria; or why don't butterfly females lay more eggs;Oikos 40 5H3Williams K S and Gilbert L E 1981 Insects as selectiveagents on plant vegetative morphology; egg mimicry
reduces egg laying by butterflies; Science 212 467-469Wolda H 1971 Fluctuations in abundance of some Homoptera in a tropical forest; Geo-Eco-Trop. 3 229-257Wolda H 1918a Fluctuations in the abundance of tropical insects; Am. Nat. 112 1017-1045Wolda H 1978b Seasonal fluctuations in rainfall, food and abundance of tropical insects; J. Anim. Ecol.47
369-381Wolda H 1980Seasonality of tropical insects.1.Leafhoppers (Homoptera) in Les Cumbres, Panama; J. Anim.
Ecol. 49 277-290Wolda H and Foster R 1978 Zunacetha annulota (Lepidoptera: Dioptidae) an outbreak insect in a neo
tropical forest; Geo-Eco-Trop. 2 443-454Wynter-Blyth 1935 Butterflies of the Indian region; Bombay Nat. Hist. Soc., Bombay














![Tropical Wave Dynamics [Lectures 11,12]](https://img.pdfslide.us/doc/110x75/58a1a2b51a28ab32438b9281/tropical-wave-dynamics-lectures-1112.jpg)














