Embed Size (px)
Citation preview

Polyamine Oxidase Is One of the Key Elements forOxidative Burst to Induce Programmed Cell Deathin Tobacco Cultured Cells1
Hiroshi Yoda*, Yoshinobu Hiroi, and Hiroshi Sano
Research and Education Center for Genetic Information, Nara Institute of Science and Technology,Nara 630–0192, Japan
Programmed cell death plays a critical role during the hypersensitive response in the plant defense system. One of componentsthat triggers it is hydrogen peroxide, which is generated through multiple pathways. One example is proposed to be polyamineoxidation, but direct evidence for this has been limited. In this article, we investigated relationships among polyamine oxidase,hydrogen peroxide, and programmed cell death using a model system constituted of tobacco (Nicotiana tabacum) cultured cell andits elicitor, cryptogein. When cultured cells were treated with cryptogein, programmed cell death occurred with a distinct patternof DNA degradation. The level of hydrogen peroxide was simultaneously increased, along with polyamine oxidase activity inapoplast. With the same treatment in the presence ofa-difluoromethyl-Orn, an inhibitor of polyamine biosynthesis, production ofhydrogen peroxide was suppressed and programmed cell death did not occur. A gene encoding a tobacco polyamine oxidase thatresides in the apoplast was isolated and used to construct RNAi transgenic cell lines. When these lines were treated withcryptogein, polyamines were not degraded but secreted into culture medium and hydrogen peroxide was scarcely produced, witha concomitant suppression of cell death. Activities of mitogen-activated protein kinases (wound- and salicylic acid-inducedprotein kinases) were also suppressed, indicating that phosphorylation cascade is involved in polyamine oxidation-derived celldeath. These results suggest that polyamine oxidase is a key element for the oxidative burst, which is essential for induction ofprogrammed cell death, and that mitogen-activated protein kinase is one of the factors that mediate this pathway.
Plant disease resistance is initiated by specific rec-ognition of pathogen-derived molecules by plant cells(Yang et al., 1997). Upon pathogen attack, necroticlesions are formed at the site of pathogen entry, re-sulting in prevention of further spread of disease, thisphenomenon being called the hypersensitive response(HR). One of the earliest events that occurs duringHR is the production of reactive oxygen intermediates(ROI), this is known as the oxidative burst. ROI mainlyinclude superoxide, hydroxyl radicals, and hydrogenperoxide, and play a critical role in triggering andmaintaining the HR by directly attacking pathogens,serving as substrates for lignification and cross-linkingof extensin, mediating signal transduction pathways,and inducing programmed cell death (Levine et al.,1994; Bestwick et al., 1997; Thordal et al., 1997). Assources of ROI, pathways catalyzed by several en-zymes have been proposed, including NADPH oxi-
dase (Keller et al., 1998), peroxidase (Bolwell andWojitaszek, 1997), oxalate oxidase (Hu et al., 2003),copper-containing amine oxidases (Allan and Fluhr,1997), and polyamine oxidases (Yoda et al., 2003). Whichof them constitutes the main source of ROI in partic-ular cases remains to be determined.
It is well documented that generation of ROI ex-hibits two phases during HR, the rapid and transientphase I and the late and persistent phase II (Grant,1997; Lamb and Dixon, 1997; Grant and Loake, 2000).This suggests that multiple sources of ROI might bepresent. NADPH oxidase located in plasma mem-branes may be the most significant source of ROI,being activated immediately after pathogen recogni-tion (Keller et al., 1998). In apoplasts, oxygen isconverted by NADPH oxidase into superoxide, whichspontaneously dismutates to give another active oxy-gen intermediate, hydrogen peroxide (Bestwick et al.,1997). Polyamines have also been proposed as impor-tant substrates for hydrogen peroxide production, be-ing degraded by copper-containing amine oxidasesor polyamine oxidases (Sebela et al., 2001; Yoda et al.,2003). In fact, enzymatic activities of polyamine me-tabolism are reported to increase in barley (Hordeumvulgare) seedlings during HR in response to powderymildew (Cowley and Walters, 2002). In animal cells,polyamines can be oxidized by polyamine oxidasesto yield hydrogen peroxide, which eventually inducesprogrammed cell death (Ha et al., 1997; Lindsay andWallace, 1999; Facchiano et al., 2001). It is therefore highlyprobable that enzymatic degradation of polyamines
1 This work was supported by a grant from the Research for theFuture Program of the Japan Society for the Promotion of Scienceand by a Grant-in-Aid for the 21st Century Center of ExcellenceResearch from the Ministry of Education, Culture, Sports, Scienceand Technology.
* Corresponding author; e-mail [email protected]; fax 81–743–72–5659.
The author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Hiroshi Yoda ([email protected]).
www.plantphysiol.org/cgi/doi/10.1104/pp.106.080515
Plant Physiology, September 2006, Vol. 142, pp. 193–206, www.plantphysiol.org � 2006 American Society of Plant Biologists 193
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021

also directly contributes to ROI production and pro-grammed cell death in plants.
Using Tobacco (Nicotiana tabacum) mosaic virus (TMV)and intact tobacco cultivars carrying the resistant (N)gene, we previously demonstrated that polyaminesare indeed one of the sources of hydrogen peroxideduring HR (Yoda et al., 2003). We found that, upon HRinduction, polyamines accumulate in apoplasts andare degraded, resulting in the production of hydrogenperoxide. Since polyamine oxidase appeared to play akey role in this system, we here isolated a gene en-coding a polyamine oxidase, and characterized its func-tion using tobacco cultured cells (Bright-Yellow 2 [BY2])and a non-race-specific proteinaceous elicitor, crypto-gein. Cryptogein is originally derived from a fungalpathogen, Phytophthora cryptogea, which induces anHR-like response in tobacco, including the oxidativeburst, nitric oxide production, and hypersensitive celldeath (Ponchet et al., 1999; Lamotte et al., 2004). Theadvantage of this system is not only that HR-like re-sponse can be synchronously induced, but also thatproduction of ROI clearly precedes HR-like cell death(Kadota et al., 2004).
During activation of plant defense responses, mitogen-activated protein kinases (MAPKs) play crucial rolesin signaling pathways. In tobacco plants carrying theN gene, TMV infection leads to activation of twoMAPKs, salicylic acid (SA)-induced protein kinase(SIPK), and wound-induced protein kinase (WIPK;Zhang and Klessig, 1998). The activation is also ob-served when tobacco plants or tobacco cultured cellsare treated with non-race-specific elicitors, includingfungal cell wall-derived elicitors, INF1, harpin, andcryptogein (Suzuki et al., 1999; Zhang et al., 2000;Sharma et al., 2003; Samuel et al., 2005). Their ortho-logs in other plant species have also been implicated indefense signaling (Zhang and Klessig, 2001). In allcases, the activation of MAPKs precedes and correlateswith the onset of HR-like cell death.
Here we provide evidence that polyamine oxidase isresponsible for generation of the oxidative burst, re-sulting in programmed cell death, and that MAPK isone of the signaling components in the polyamineoxidase-dependent defense response.
RESULTS
Programmed Cell Death Induced by Cryptogein
To determine whether cryptogein-induced cell deathis programmed, patterns of DNA cleavage were ana-lyzed. BY2 cells were treated with cryptogein for 24 hand DNA cleavage was examined with gel electro-phoresis. The results showed a specific DNA ladder-ing pattern, consisting of multiples of about 180 bp(Fig. 1A), which was not evident with DNA fromuntreated cells (Fig. 1A). In situ DNA cleavage wasthen examined by subjecting cryptogein-treated cellsto 3#-end DNA labeling with terminal deoxynucleo-
tide transferase-mediated dUTP nick-end labeling(TUNEL assay). Bright fluorescent signals were evi-dent in nuclei, indicating DNA to be fragmented (Fig.1B). Untreated cells were not labeled (Fig. 1B). Theresults are consistent with the case of animal cells un-dergoing apoptosis, showing a characteristic ladder-ing pattern with nucleosome-sized DNA fragmentsand positive TUNEL staining, and strongly suggestingthat programmed cell death indeed takes place in BY2cells upon cryptogein treatment.
Suppression of Programmed Cell Death
To establish an experimental system in which pro-grammed cell death is controlled, we first examinedthe effect of an elicitor, cryptogein, and inhibitors ofpolyamine synthetic pathways. a-Difluoromethyl-Arg,an irreversible inhibitor of Arg decarboxylase, whichcatalyzes conversion of Arg into agmatine, did not
Figure 1. Programmed cell death induced by cryptogein. A, DNAladdering pattern. A 5-mg aliquot of DNA extracted from BY2 cells treatedwith 250 nM cryptogein for 0 or 24 hwas fractionated on a 2% agarose gelandvisualizedwith ethidium bromide staining. Sizemarker DNA (M)wasrun in parallel. Arrowheads indicate DNA fragments of 180, 360, and 720bp. B, In situ DNA fragmentation. BY2 cells treated with 250 nMcryptogein for 24 h were fixed, subjected to TUNEL assay, and visualizedunder a fluorescent microscope (bottom section). Untreated cells werealso subjected to TUNEL assay as a control (top section).
Yoda et al.
194 Plant Physiol. Vol. 142, 2006
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021
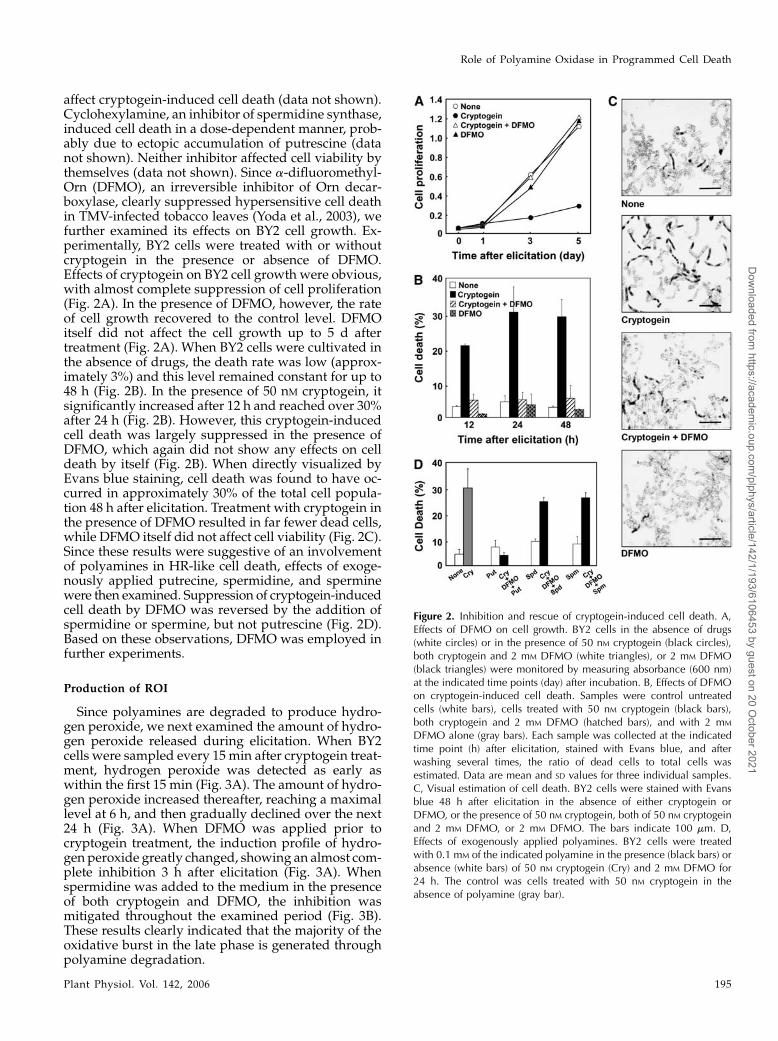
affect cryptogein-induced cell death (data not shown).Cyclohexylamine, an inhibitor of spermidine synthase,induced cell death in a dose-dependent manner, prob-ably due to ectopic accumulation of putrescine (datanot shown). Neither inhibitor affected cell viability bythemselves (data not shown). Since a-difluoromethyl-Orn (DFMO), an irreversible inhibitor of Orn decar-boxylase, clearly suppressed hypersensitive cell deathin TMV-infected tobacco leaves (Yoda et al., 2003), wefurther examined its effects on BY2 cell growth. Ex-perimentally, BY2 cells were treated with or withoutcryptogein in the presence or absence of DFMO.Effects of cryptogein on BY2 cell growth were obvious,with almost complete suppression of cell proliferation(Fig. 2A). In the presence of DFMO, however, the rateof cell growth recovered to the control level. DFMOitself did not affect the cell growth up to 5 d aftertreatment (Fig. 2A). When BY2 cells were cultivated inthe absence of drugs, the death rate was low (approx-imately 3%) and this level remained constant for up to48 h (Fig. 2B). In the presence of 50 nM cryptogein, itsignificantly increased after 12 h and reached over 30%after 24 h (Fig. 2B). However, this cryptogein-inducedcell death was largely suppressed in the presence ofDFMO, which again did not show any effects on celldeath by itself (Fig. 2B). When directly visualized byEvans blue staining, cell death was found to have oc-curred in approximately 30% of the total cell popula-tion 48 h after elicitation. Treatment with cryptogein inthe presence of DFMO resulted in far fewer dead cells,while DFMO itself did not affect cell viability (Fig. 2C).Since these results were suggestive of an involvementof polyamines in HR-like cell death, effects of exoge-nously applied putrecine, spermidine, and sperminewere then examined. Suppression of cryptogein-inducedcell death by DFMO was reversed by the addition ofspermidine or spermine, but not putrescine (Fig. 2D).Based on these observations, DFMO was employed infurther experiments.
Production of ROI
Since polyamines are degraded to produce hydro-gen peroxide, we next examined the amount of hydro-gen peroxide released during elicitation. When BY2cells were sampled every 15 min after cryptogein treat-ment, hydrogen peroxide was detected as early aswithin the first 15 min (Fig. 3A). The amount of hydro-gen peroxide increased thereafter, reaching a maximallevel at 6 h, and then gradually declined over the next24 h (Fig. 3A). When DFMO was applied prior tocryptogein treatment, the induction profile of hydro-gen peroxide greatly changed, showing an almost com-plete inhibition 3 h after elicitation (Fig. 3A). Whenspermidine was added to the medium in the presenceof both cryptogein and DFMO, the inhibition wasmitigated throughout the examined period (Fig. 3B).These results clearly indicated that the majority of theoxidative burst in the late phase is generated throughpolyamine degradation.
Figure 2. Inhibition and rescue of cryptogein-induced cell death. A,Effects of DFMO on cell growth. BY2 cells in the absence of drugs(white circles) or in the presence of 50 nM cryptogein (black circles),both cryptogein and 2 mM DFMO (white triangles), or 2 mM DFMO(black triangles) were monitored by measuring absorbance (600 nm)at the indicated time points (day) after incubation. B, Effects of DFMOon cryptogein-induced cell death. Samples were control untreatedcells (white bars), cells treated with 50 nM cryptogein (black bars),both cryptogein and 2 mM DFMO (hatched bars), and with 2 mM
DFMO alone (gray bars). Each sample was collected at the indicatedtime point (h) after elicitation, stained with Evans blue, and afterwashing several times, the ratio of dead cells to total cells wasestimated. Data are mean and SD values for three individual samples.C, Visual estimation of cell death. BY2 cells were stained with Evansblue 48 h after elicitation in the absence of either cryptogein orDFMO, or the presence of 50 nM cryptogein, both of 50 nM cryptogeinand 2 mM DFMO, or 2 mM DFMO. The bars indicate 100 mm. D,Effects of exogenously applied polyamines. BY2 cells were treatedwith 0.1 mM of the indicated polyamine in the presence (black bars) orabsence (white bars) of 50 nM cryptogein (Cry) and 2 mM DFMO for24 h. The control was cells treated with 50 nM cryptogein in theabsence of polyamine (gray bar).
Role of Polyamine Oxidase in Programmed Cell Death
Plant Physiol. Vol. 142, 2006 195
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021
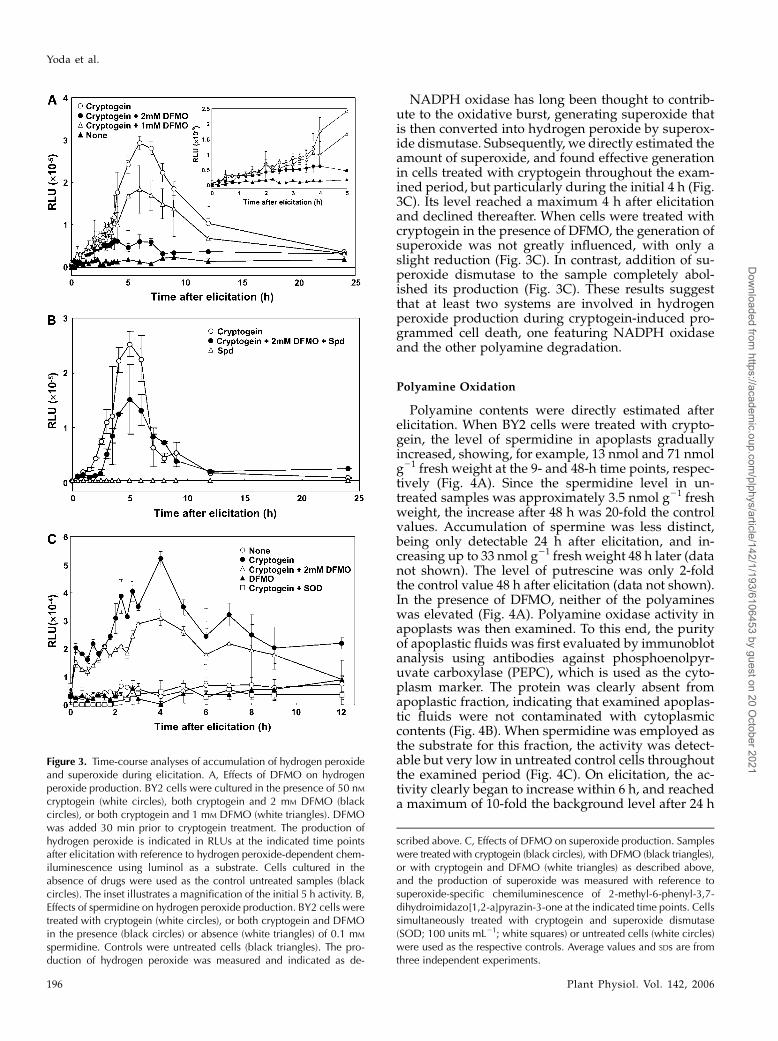
NADPH oxidase has long been thought to contrib-ute to the oxidative burst, generating superoxide thatis then converted into hydrogen peroxide by superox-ide dismutase. Subsequently, we directly estimated theamount of superoxide, and found effective generationin cells treated with cryptogein throughout the exam-ined period, but particularly during the initial 4 h (Fig.3C). Its level reached a maximum 4 h after elicitationand declined thereafter. When cells were treated withcryptogein in the presence of DFMO, the generation ofsuperoxide was not greatly influenced, with only aslight reduction (Fig. 3C). In contrast, addition of su-peroxide dismutase to the sample completely abol-ished its production (Fig. 3C). These results suggestthat at least two systems are involved in hydrogenperoxide production during cryptogein-induced pro-grammed cell death, one featuring NADPH oxidaseand the other polyamine degradation.
Polyamine Oxidation
Polyamine contents were directly estimated afterelicitation. When BY2 cells were treated with crypto-gein, the level of spermidine in apoplasts graduallyincreased, showing, for example, 13 nmol and 71 nmolg21 fresh weight at the 9- and 48-h time points, respec-tively (Fig. 4A). Since the spermidine level in un-treated samples was approximately 3.5 nmol g21 freshweight, the increase after 48 h was 20-fold the controlvalues. Accumulation of spermine was less distinct,being only detectable 24 h after elicitation, and in-creasing up to 33 nmol g21 fresh weight 48 h later (datanot shown). The level of putrescine was only 2-foldthe control value 48 h after elicitation (data not shown).In the presence of DFMO, neither of the polyamineswas elevated (Fig. 4A). Polyamine oxidase activity inapoplasts was then examined. To this end, the purityof apoplastic fluids was first evaluated by immunoblotanalysis using antibodies against phosphoenolpyr-uvate carboxylase (PEPC), which is used as the cyto-plasm marker. The protein was clearly absent fromapoplastic fraction, indicating that examined apoplas-tic fluids were not contaminated with cytoplasmiccontents (Fig. 4B). When spermidine was employed asthe substrate for this fraction, the activity was detect-able but very low in untreated control cells throughoutthe examined period (Fig. 4C). On elicitation, the ac-tivity clearly began to increase within 6 h, and reacheda maximum of 10-fold the background level after 24 h
Figure 3. Time-course analyses of accumulation of hydrogen peroxideand superoxide during elicitation. A, Effects of DFMO on hydrogenperoxide production. BY2 cells were cultured in the presence of 50 nMcryptogein (white circles), both cryptogein and 2 mM DFMO (blackcircles), or both cryptogein and 1 mM DFMO (white triangles). DFMOwas added 30 min prior to cryptogein treatment. The production ofhydrogen peroxide is indicated in RLUs at the indicated time pointsafter elicitation with reference to hydrogen peroxide-dependent chem-iluminescence using luminol as a substrate. Cells cultured in theabsence of drugs were used as the control untreated samples (blackcircles). The inset illustrates a magnification of the initial 5 h activity. B,Effects of spermidine on hydrogen peroxide production. BY2 cells weretreated with cryptogein (white circles), or both cryptogein and DFMOin the presence (black circles) or absence (white triangles) of 0.1 mM
spermidine. Controls were untreated cells (black triangles). The pro-duction of hydrogen peroxide was measured and indicated as de-
scribed above. C, Effects of DFMO on superoxide production. Sampleswere treated with cryptogein (black circles), with DFMO (black triangles),or with cryptogein and DFMO (white triangles) as described above,and the production of superoxide was measured with reference tosuperoxide-specific chemiluminescence of 2-methyl-6-phenyl-3,7-dihydroimidazo[1,2-a]pyrazin-3-one at the indicated time points. Cellssimultaneously treated with cryptogein and superoxide dismutase(SOD; 100 units mL21; white squares) or untreated cells (white circles)were used as the respective controls. Average values and SDs are fromthree independent experiments.
Yoda et al.
196 Plant Physiol. Vol. 142, 2006
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021
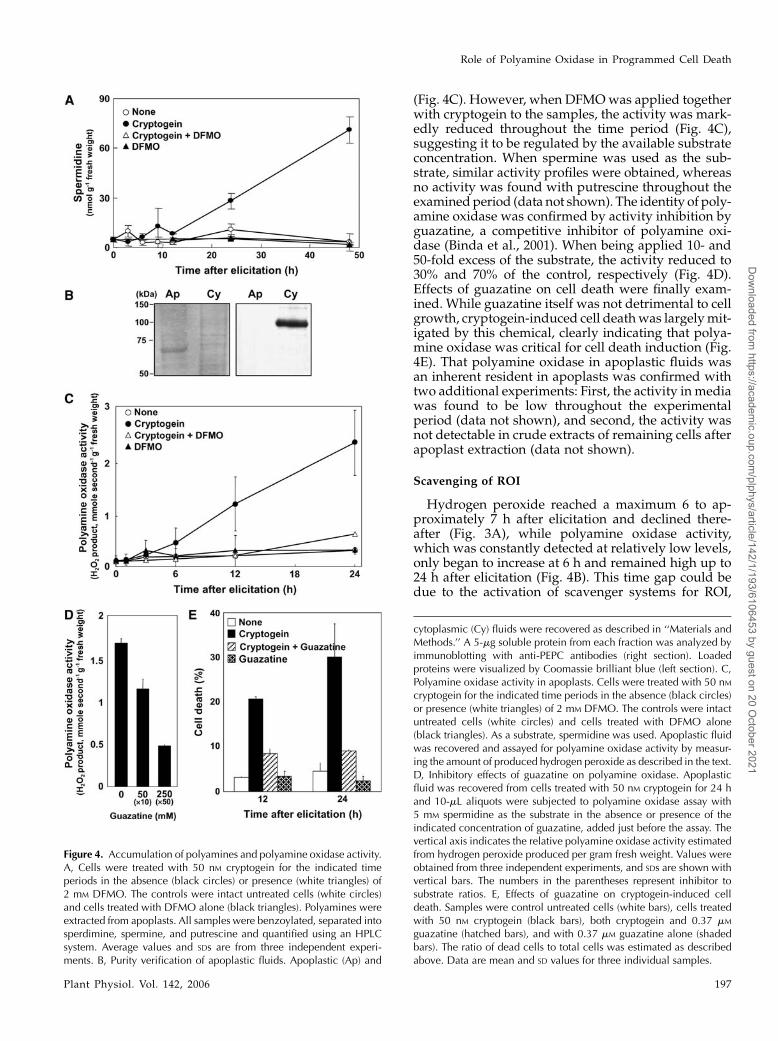
(Fig. 4C). However, when DFMO was applied togetherwith cryptogein to the samples, the activity was mark-edly reduced throughout the time period (Fig. 4C),suggesting it to be regulated by the available substrateconcentration. When spermine was used as the sub-strate, similar activity profiles were obtained, whereasno activity was found with putrescine throughout theexamined period (data not shown). The identity of poly-amine oxidase was confirmed by activity inhibition byguazatine, a competitive inhibitor of polyamine oxi-dase (Binda et al., 2001). When being applied 10- and50-fold excess of the substrate, the activity reduced to30% and 70% of the control, respectively (Fig. 4D).Effects of guazatine on cell death were finally exam-ined. While guazatine itself was not detrimental to cellgrowth, cryptogein-induced cell death was largely mit-igated by this chemical, clearly indicating that polya-mine oxidase was critical for cell death induction (Fig.4E). That polyamine oxidase in apoplastic fluids wasan inherent resident in apoplasts was confirmed withtwo additional experiments: First, the activity in mediawas found to be low throughout the experimentalperiod (data not shown), and second, the activity wasnot detectable in crude extracts of remaining cells afterapoplast extraction (data not shown).
Scavenging of ROI
Hydrogen peroxide reached a maximum 6 to ap-proximately 7 h after elicitation and declined there-after (Fig. 3A), while polyamine oxidase activity,which was constantly detected at relatively low levels,only began to increase at 6 h and remained high up to24 h after elicitation (Fig. 4B). This time gap could bedue to the activation of scavenger systems for ROI,
Figure 4. Accumulation of polyamines and polyamine oxidase activity.A, Cells were treated with 50 nM cryptogein for the indicated timeperiods in the absence (black circles) or presence (white triangles) of2 mM DFMO. The controls were intact untreated cells (white circles)and cells treated with DFMO alone (black triangles). Polyamines wereextracted from apoplasts. All samples were benzoylated, separated intosperdimine, spermine, and putrescine and quantified using an HPLCsystem. Average values and SDs are from three independent experi-ments. B, Purity verification of apoplastic fluids. Apoplastic (Ap) and
cytoplasmic (Cy) fluids were recovered as described in ‘‘Materials andMethods.’’ A 5-mg soluble protein from each fraction was analyzed byimmunoblotting with anti-PEPC antibodies (right section). Loadedproteins were visualized by Coomassie brilliant blue (left section). C,Polyamine oxidase activity in apoplasts. Cells were treated with 50 nMcryptogein for the indicated time periods in the absence (black circles)or presence (white triangles) of 2 mM DFMO. The controls were intactuntreated cells (white circles) and cells treated with DFMO alone(black triangles). As a substrate, spermidine was used. Apoplastic fluidwas recovered and assayed for polyamine oxidase activity by measur-ing the amount of produced hydrogen peroxide as described in the text.D, Inhibitory effects of guazatine on polyamine oxidase. Apoplasticfluid was recovered from cells treated with 50 nM cryptogein for 24 hand 10-mL aliquots were subjected to polyamine oxidase assay with5 mM spermidine as the substrate in the absence or presence of theindicated concentration of guazatine, added just before the assay. Thevertical axis indicates the relative polyamine oxidase activity estimatedfrom hydrogen peroxide produced per gram fresh weight. Values wereobtained from three independent experiments, and SDs are shown withvertical bars. The numbers in the parentheses represent inhibitor tosubstrate ratios. E, Effects of guazatine on cryptogein-induced celldeath. Samples were control untreated cells (white bars), cells treatedwith 50 nM cryptogein (black bars), both cryptogein and 0.37 mM
guazatine (hatched bars), and with 0.37 mM guazatine alone (shadedbars). The ratio of dead cells to total cells was estimated as describedabove. Data are mean and SD values for three individual samples.
Role of Polyamine Oxidase in Programmed Cell Death
Plant Physiol. Vol. 142, 2006 197
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021
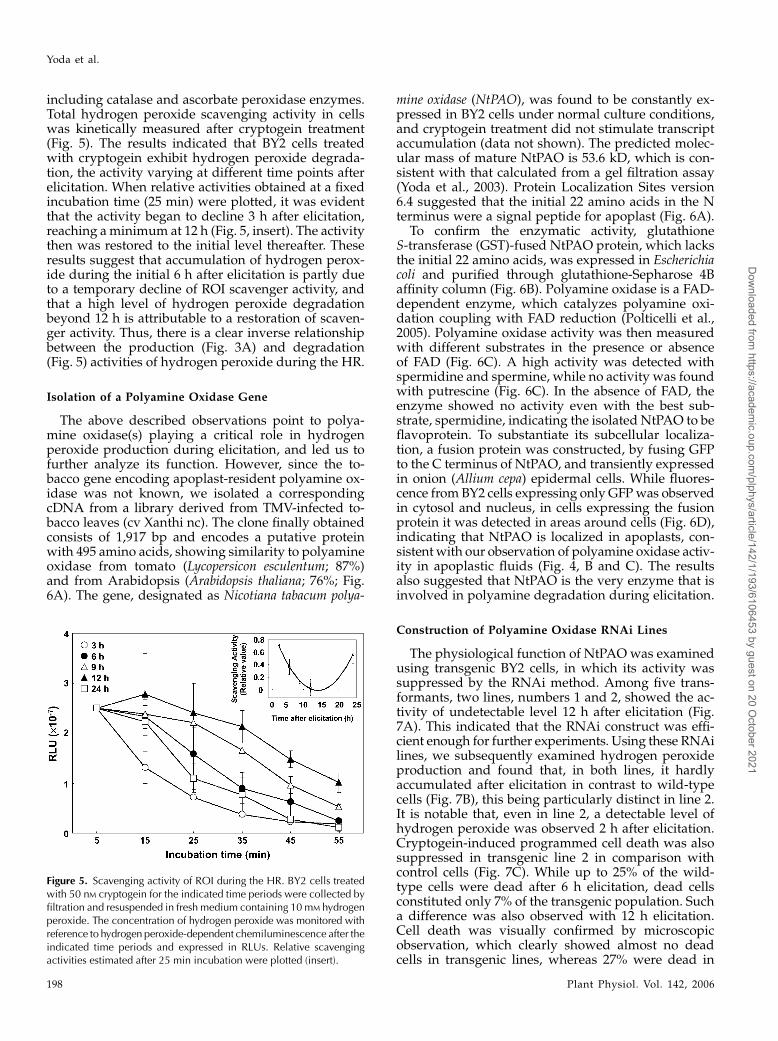
including catalase and ascorbate peroxidase enzymes.Total hydrogen peroxide scavenging activity in cellswas kinetically measured after cryptogein treatment(Fig. 5). The results indicated that BY2 cells treatedwith cryptogein exhibit hydrogen peroxide degrada-tion, the activity varying at different time points afterelicitation. When relative activities obtained at a fixedincubation time (25 min) were plotted, it was evidentthat the activity began to decline 3 h after elicitation,reaching a minimum at 12 h (Fig. 5, insert). The activitythen was restored to the initial level thereafter. Theseresults suggest that accumulation of hydrogen perox-ide during the initial 6 h after elicitation is partly dueto a temporary decline of ROI scavenger activity, andthat a high level of hydrogen peroxide degradationbeyond 12 h is attributable to a restoration of scaven-ger activity. Thus, there is a clear inverse relationshipbetween the production (Fig. 3A) and degradation(Fig. 5) activities of hydrogen peroxide during the HR.
Isolation of a Polyamine Oxidase Gene
The above described observations point to polya-mine oxidase(s) playing a critical role in hydrogenperoxide production during elicitation, and led us tofurther analyze its function. However, since the to-bacco gene encoding apoplast-resident polyamine ox-idase was not known, we isolated a correspondingcDNA from a library derived from TMV-infected to-bacco leaves (cv Xanthi nc). The clone finally obtainedconsists of 1,917 bp and encodes a putative proteinwith 495 amino acids, showing similarity to polyamineoxidase from tomato (Lycopersicon esculentum; 87%)and from Arabidopsis (Arabidopsis thaliana; 76%; Fig.6A). The gene, designated as Nicotiana tabacum polya-
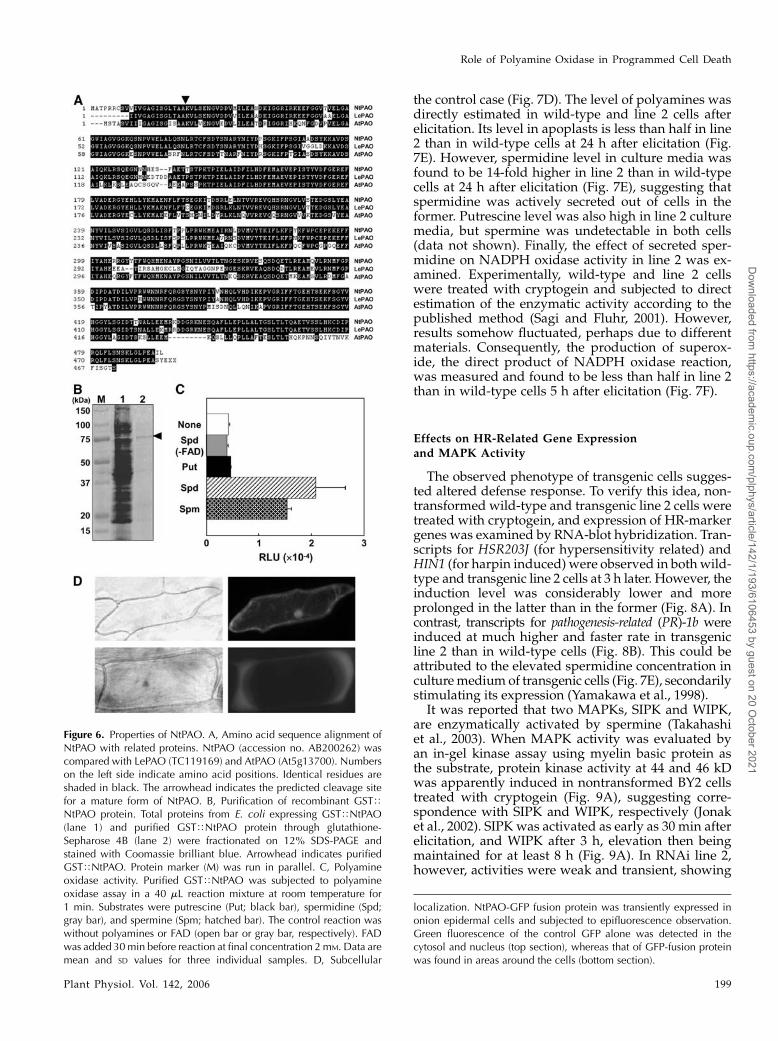
mine oxidase (NtPAO), was found to be constantly ex-pressed in BY2 cells under normal culture conditions,and cryptogein treatment did not stimulate transcriptaccumulation (data not shown). The predicted molec-ular mass of mature NtPAO is 53.6 kD, which is con-sistent with that calculated from a gel filtration assay(Yoda et al., 2003). Protein Localization Sites version6.4 suggested that the initial 22 amino acids in the Nterminus were a signal peptide for apoplast (Fig. 6A).
To confirm the enzymatic activity, glutathioneS-transferase (GST)-fused NtPAO protein, which lacksthe initial 22 amino acids, was expressed in Escherichiacoli and purified through glutathione-Sepharose 4Baffinity column (Fig. 6B). Polyamine oxidase is a FAD-dependent enzyme, which catalyzes polyamine oxi-dation coupling with FAD reduction (Polticelli et al.,2005). Polyamine oxidase activity was then measuredwith different substrates in the presence or absenceof FAD (Fig. 6C). A high activity was detected withspermidine and spermine, while no activity was foundwith putrescine (Fig. 6C). In the absence of FAD, theenzyme showed no activity even with the best sub-strate, spermidine, indicating the isolated NtPAO to beflavoprotein. To substantiate its subcellular localiza-tion, a fusion protein was constructed, by fusing GFPto the C terminus of NtPAO, and transiently expressedin onion (Allium cepa) epidermal cells. While fluores-cence from BY2 cells expressing only GFP was observedin cytosol and nucleus, in cells expressing the fusionprotein it was detected in areas around cells (Fig. 6D),indicating that NtPAO is localized in apoplasts, con-sistent with our observation of polyamine oxidase activ-ity in apoplastic fluids (Fig. 4, B and C). The resultsalso suggested that NtPAO is the very enzyme that isinvolved in polyamine degradation during elicitation.
Construction of Polyamine Oxidase RNAi Lines
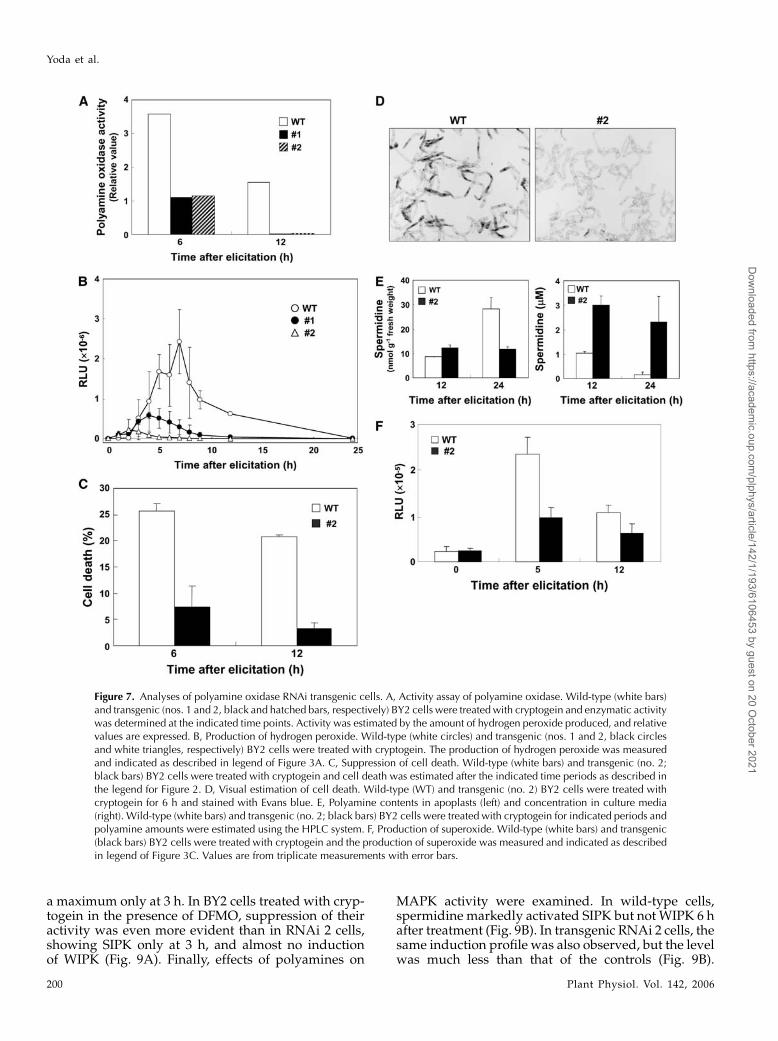
The physiological function of NtPAO was examinedusing transgenic BY2 cells, in which its activity wassuppressed by the RNAi method. Among five trans-formants, two lines, numbers 1 and 2, showed the ac-tivity of undetectable level 12 h after elicitation (Fig.7A). This indicated that the RNAi construct was effi-cient enough for further experiments. Using these RNAilines, we subsequently examined hydrogen peroxideproduction and found that, in both lines, it hardlyaccumulated after elicitation in contrast to wild-typecells (Fig. 7B), this being particularly distinct in line 2.It is notable that, even in line 2, a detectable level ofhydrogen peroxide was observed 2 h after elicitation.Cryptogein-induced programmed cell death was alsosuppressed in transgenic line 2 in comparison withcontrol cells (Fig. 7C). While up to 25% of the wild-type cells were dead after 6 h elicitation, dead cellsconstituted only 7% of the transgenic population. Sucha difference was also observed with 12 h elicitation.Cell death was visually confirmed by microscopicobservation, which clearly showed almost no deadcells in transgenic lines, whereas 27% were dead in
Figure 5. Scavenging activity of ROI during the HR. BY2 cells treatedwith 50 nM cryptogein for the indicated time periods were collected byfiltration and resuspended in fresh medium containing 10 mM hydrogenperoxide. The concentration of hydrogen peroxide was monitored withreference to hydrogen peroxide-dependent chemiluminescence after theindicated time periods and expressed in RLUs. Relative scavengingactivities estimated after 25 min incubation were plotted (insert).
Yoda et al.
198 Plant Physiol. Vol. 142, 2006
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021
the control case (Fig. 7D). The level of polyamines wasdirectly estimated in wild-type and line 2 cells afterelicitation. Its level in apoplasts is less than half in line2 than in wild-type cells at 24 h after elicitation (Fig.7E). However, spermidine level in culture media wasfound to be 14-fold higher in line 2 than in wild-typecells at 24 h after elicitation (Fig. 7E), suggesting thatspermidine was actively secreted out of cells in theformer. Putrescine level was also high in line 2 culturemedia, but spermine was undetectable in both cells(data not shown). Finally, the effect of secreted sper-midine on NADPH oxidase activity in line 2 was ex-amined. Experimentally, wild-type and line 2 cellswere treated with cryptogein and subjected to directestimation of the enzymatic activity according to thepublished method (Sagi and Fluhr, 2001). However,results somehow fluctuated, perhaps due to differentmaterials. Consequently, the production of superox-ide, the direct product of NADPH oxidase reaction,was measured and found to be less than half in line 2than in wild-type cells 5 h after elicitation (Fig. 7F).
Effects on HR-Related Gene Expression
and MAPK Activity
The observed phenotype of transgenic cells sugges-ted altered defense response. To verify this idea, non-transformed wild-type and transgenic line 2 cells weretreated with cryptogein, and expression of HR-markergenes was examined by RNA-blot hybridization. Tran-scripts for HSR203J (for hypersensitivity related) andHIN1 (for harpin induced) were observed in both wild-type and transgenic line 2 cells at 3 h later. However, theinduction level was considerably lower and moreprolonged in the latter than in the former (Fig. 8A). Incontrast, transcripts for pathogenesis-related (PR)-1b wereinduced at much higher and faster rate in transgenicline 2 than in wild-type cells (Fig. 8B). This could beattributed to the elevated spermidine concentration inculture medium of transgenic cells (Fig. 7E), secondarilystimulating its expression (Yamakawa et al., 1998).
It was reported that two MAPKs, SIPK and WIPK,are enzymatically activated by spermine (Takahashiet al., 2003). When MAPK activity was evaluated byan in-gel kinase assay using myelin basic protein asthe substrate, protein kinase activity at 44 and 46 kDwas apparently induced in nontransformed BY2 cellstreated with cryptogein (Fig. 9A), suggesting corre-spondence with SIPK and WIPK, respectively (Jonaket al., 2002). SIPK was activated as early as 30 min afterelicitation, and WIPK after 3 h, elevation then beingmaintained for at least 8 h (Fig. 9A). In RNAi line 2,however, activities were weak and transient, showing
Figure 6. Properties of NtPAO. A, Amino acid sequence alignment ofNtPAO with related proteins. NtPAO (accession no. AB200262) wascompared with LePAO (TC119169) and AtPAO (At5g13700). Numberson the left side indicate amino acid positions. Identical residues areshaded in black. The arrowhead indicates the predicted cleavage sitefor a mature form of NtPAO. B, Purification of recombinant GSTTNtPAO protein. Total proteins from E. coli expressing GSTTNtPAO(lane 1) and purified GSTTNtPAO protein through glutathione-Sepharose 4B (lane 2) were fractionated on 12% SDS-PAGE andstained with Coomassie brilliant blue. Arrowhead indicates purifiedGSTTNtPAO. Protein marker (M) was run in parallel. C, Polyamineoxidase activity. Purified GSTTNtPAO was subjected to polyamineoxidase assay in a 40 mL reaction mixture at room temperature for1 min. Substrates were putrescine (Put; black bar), spermidine (Spd;gray bar), and spermine (Spm; hatched bar). The control reaction waswithout polyamines or FAD (open bar or gray bar, respectively). FADwas added 30min before reaction at final concentration 2mM. Data aremean and SD values for three individual samples. D, Subcellular
localization. NtPAO-GFP fusion protein was transiently expressed inonion epidermal cells and subjected to epifluorescence observation.Green fluorescence of the control GFP alone was detected in thecytosol and nucleus (top section), whereas that of GFP-fusion proteinwas found in areas around the cells (bottom section).
Role of Polyamine Oxidase in Programmed Cell Death
Plant Physiol. Vol. 142, 2006 199
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021
a maximum only at 3 h. In BY2 cells treated with cryp-togein in the presence of DFMO, suppression of theiractivity was even more evident than in RNAi 2 cells,showing SIPK only at 3 h, and almost no inductionof WIPK (Fig. 9A). Finally, effects of polyamines on
MAPK activity were examined. In wild-type cells,spermidine markedly activated SIPK but not WIPK 6 hafter treatment (Fig. 9B). In transgenic RNAi 2 cells, thesame induction profile was also observed, but the levelwas much less than that of the controls (Fig. 9B).
Figure 7. Analyses of polyamine oxidase RNAi transgenic cells. A, Activity assay of polyamine oxidase. Wild-type (white bars)and transgenic (nos. 1 and 2, black and hatched bars, respectively) BY2 cells were treatedwith cryptogein and enzymatic activitywas determined at the indicated time points. Activity was estimated by the amount of hydrogen peroxide produced, and relativevalues are expressed. B, Production of hydrogen peroxide. Wild-type (white circles) and transgenic (nos. 1 and 2, black circlesand white triangles, respectively) BY2 cells were treated with cryptogein. The production of hydrogen peroxide was measuredand indicated as described in legend of Figure 3A. C, Suppression of cell death. Wild-type (white bars) and transgenic (no. 2;black bars) BY2 cells were treated with cryptogein and cell death was estimated after the indicated time periods as described inthe legend for Figure 2. D, Visual estimation of cell death. Wild-type (WT) and transgenic (no. 2) BY2 cells were treated withcryptogein for 6 h and stained with Evans blue. E, Polyamine contents in apoplasts (left) and concentration in culture media(right). Wild-type (white bars) and transgenic (no. 2; black bars) BY2 cells were treated with cryptogein for indicated periods andpolyamine amounts were estimated using the HPLC system. F, Production of superoxide. Wild-type (white bars) and transgenic(black bars) BY2 cells were treated with cryptogein and the production of superoxide was measured and indicated as describedin legend of Figure 3C. Values are from triplicate measurements with error bars.
Yoda et al.
200 Plant Physiol. Vol. 142, 2006
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021

Neither spermine nor putrescine was effective underour experimental conditions (data not shown).
DISCUSSION
ROI is a crucial element for induction of pro-grammed cell death during HR and HR-like responses(Hammond-Kosack and Jones, 1996; Lamb and Dixon,1997). Since ROI can be detected as early as within min-utes upon the onset of HR, its production is thought tobe one key factor in the defense response. The source,however, may vary, although NADPH oxidase is con-sidered to be a major contributor because of its sim-ilarity with neutrophil enzymes in mammals (Groomet al., 1996; Keller et al., 1998). In a previous investi-gation with intact tobacco leaves and TMV, we dem-onstrated that genes encoding enzymes involved inpolyamine biosynthesis are markedly up-regulatedupon the onset of HR (Yoda et al., 2003). Subsequently,polyamines were found to accumulate in apoplasts ofinfected leaves and to be degraded by polyamine oxi-dase, releasing hydrogen peroxide that causes hyper-sensitive cell death (Yoda et al., 2003). This raisedquestions as to the extent of the overall contribution ofpolyamine-derived hydrogen peroxide to hypersensi-tive cell death.
Temporal Features
To address this question, we developed the presentsystem of tobacco cultured cells exposed to crypto-gein. Trials established that BY2 cells, like tobacco cul-tured cells derived from tobacco cv Xanthi nc (Binetet al., 2001), can be efficiently stimulated to undergoprogrammed cell death by cryptogein. Time-courseanalyses using this system indeed showed clear spatialand temporal accumulation and degradation of com-
ponents involved in hydrogen peroxide metabolism(Fig. 10). Accumulation of polyamines proved varia-ble, with a gradual increase of spermidine but almostnone of spermine before 24 h. Polyamine oxidaseactivity was constitutively detectable at a low level,but markedly increased after elicitation. Hydrogenperoxide production occurred as early as 15 min afterelicitation, reaching a maximal level at 6 h and thendeclined. This feature was inversely well correlatedwith scavenger activity of hydrogen peroxide. The sig-nal transducers, SIPK and WIPK, were activated within30 min and 3 h, respectively. Hypersensitive cell deathwas apparent 6 h later. Based on these findings wepropose that upon elicitation, polyamines are simul-taneously produced and degraded, releasing hydro-gen peroxide, which activates signal transductionpathways including MAPKs to induce programmedcell death. Although the overall feature of polyamine-dependent programmed cell death can be explainedby this idea, some comments are needed as to rela-tionships among the factors involved.
Accumulation of polyamines began after the onsetof oxidative burst. This can be explained by their si-multaneous production and degradation, as seen froma vast accumulation of spermidine at relatively earlystage in culture fluid of the RNAi line, in which poly-amine oxidase activity is suppressed. Suppression ofoxidative burst in both DFMO-treated cells and poly-amine oxidase RNAi cell lines also supports this idea.In wild-type plants, perhaps polyamine synthesis iswell concerted with oxidation, resulting in apparently
Figure 9. Suppression of MAPK activity. A, In-gel kinase assays forMAPK activity. Wild-type BY2 cells were treated with cryptogein in theabsence (WT) or presence (DFMO) of DFMO for the indicated timeperiods. Transgenic line number 2 cells (#2) were also treated withcryptogein. Protein extracts were prepared from each sample, fraction-ated by 10% PAGE, and kinase activity was determined by the in-gelkinase assay method with myelin basic protein as the substrate. B, SIPKactivation by spermidine. Wild-type (WT) and transgenic line number 2(#2) cells were treated with 0.3 mM spermidine for the indicated timeperiods and proteins were extracted and subjected to in-gel kinaseassay as described above.
Figure 8. Change of HR-related gene expression in RNAi transgeniccells. Total RNA was isolated from wild-type (WT) and transgenic (no.2) cells, which were treated with cryptogein for indicated time periods.RNA gel-blot analysis was conducted with indicated probes that wereradioactively labeled with 32P. Experiments were repeated twice toconfirm similar results. rRNA was used as the reference to show equalloading of samples.
Role of Polyamine Oxidase in Programmed Cell Death
Plant Physiol. Vol. 142, 2006 201
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021

no accumulation during the oxidative burst occurringat the first 12 h. Polyamines are gradually accumulatedthereafter despite the increase of polyamine oxidase.We have no clear explanation for this, but several casesare conceivable; for example, differential rate betweenproduction and degradation, masking polyamine ox-idase activity in vivo, and protection of polyamines byother components.
Hydrogen peroxide level declines 6 h after elicita-tion, while polyamines and polyamine oxidase activityconstantly increase. This can be partly due to scav-enging activity of ROI, which is transiently decreasedduring the first 12 h after elicitation and increased tothe basal level thereafter. Hydrogen peroxide pro-duced later than 12 h might be erased by this recov-ered scavenging activity. Another possibility of itsapparent decline at the late stage is consumption ofhydrogen peroxide for repair process. For example, dur-ing defense response, ROI is consumed to strengthencell wall for physiological barrier against pathogensvia oxidative cross-linking of glycoproteins (Bradleyet al., 1992) and lignification from oxidative polymer-ization of monolignols (Kawasaki et al., 2006).
Spermidine was significantly secreted into mediumand not retained in apoplasts of RNAi cells. Since theactivity of hydrogen peroxide production is sup-pressed in RNAi transgenic lines, cell wall restorationmight be low as discussed above. This may cause a
loose structure of cell wall through which polyaminescould leak into the medium as observed.
Exogenously applied putrescine, a direct reactionproduct of Orn decarboxylase that is inhibited byDFMO, does not rescue DFMO-dependent suppres-sion of cell death. This suggests that putrescine isnot directly involved in polyamine-dependent celldeath. Since its accumulation was less evident incomparison with other polyamines, putrescine mayserve as the intermediate for polyamine synthesis inthe present system. Alternatively, since putrescine pre-vents programmed cell death syndrome (Papadakisand Roubelakis-Angelakis, 2005), it may also functionas a negative regulator to limit the propagation ofprogrammed cell death.
As spermine is known as an inducer for PR geneexpression (Yamakawa et al., 1998) and as an activatorfor MAPKs in tobacco plants (Takahashi et al., 2003),its role in our system should be considered. In the pres-ent study, spermine began to accumulate at least 24 hafter elicitation, when expression of PR-1b was fullydetectable. However, MAPKs are activated within30 min with a gradual increase during hours afterelicitation, when spermine accumulation is not ob-served. This suggests that spermine is not involved inmodulating the MAPK activities by itself. It is con-ceivable that spermine serves as the substrate for poly-amine oxidase at the early stage, and as the activatorfor such as PR genes at the late stage of the HR, al-though the possibility is not excluded that elicitationrenders BY2 cells more sensitive to spermine, resultingin activation of MAPKs before apparent accumulationof spermine takes place.
Source for Oxidative Burst
Regarding the two phases of the oxidative burst(Grant, 1997; Lamb and Dixon, 1997; Grant and Loake,2000), the amount of ROI in the prolonged phase II ismarkedly more abundant than in phase I (Draper,1997). At present, the biological function of ROI in eachphase is not completely understood. However, in phaseI it may not always correlate with plant disease resis-tance, whereas the link with the defense response ap-pears more absolute in phase II (Baker and Orlandi,1995). Analyses have suggested that ROI in phase I isdependent on NADPH oxidase based on the follow-ing observations: First, suppression of the gene encod-ing gp91phox homologs (rboh), one component of theNADPH oxidase complex, resulted in a disruption ofboth phases in Arabidopsis (Torres et al., 2002) and to-bacco (Simon-Plas et al., 2002); second, oxidative burstis sensitive to diphenylene iodonium, an inhibitor offlavin-containing oxidase including NADPH oxidase(Rusterucci et al., 1996; Simon-Plas et al., 1997); third, atobacco NtrbohD was rapidly and transiently expressedin cryptogein-treated cultured cells (Simon-Plas et al.,2002), this being consistent with our present observa-tion of rapid and transient production of superoxide.Further analysis recently indicated that spermine and
Figure 10. Kinetic outline of factors involved in polyamine-dependentcell death. The vertical axis indicates relative values for the indicatedevent, and the horizontal axis the time elapsed after onset of the HR.Spd and Spm stand for spermidine and spermine, respectively.
Yoda et al.
202 Plant Physiol. Vol. 142, 2006
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021

spermidine, but not putrescine, inhibit NADPH oxi-dase activity in a concentration-dependent manner(Papadakis and Roubelakis-Angelakis, 2005). Our pre-sent results agree with these observations and suggestthat observed inability of hydrogen peroxide produc-tion in RNAi cells was partly caused by the inhibitionof NADPH oxidase with leaked polyamines. However,the direct inhibition of polyamine oxidation was highlyconceived to be the main cause for it. Based on theavailable information, including our present findings,we speculate that ROI in phase I, which is the prereq-uisite to the next phase II oxidative burst, is producedby NADPH oxidase immediately after elicitation,whereas ROI in phase II is produced mainly by poly-amine degradation. We further speculate that oxida-tive burst in phase I may occur mainly in elicited cellsand serve as one of the triggers for subsequent HRevents, including phase II oxidative burst.
Signaling Pathways
MAPKs have long been considered to play a criticalrole in defense signal transduction and the defensegene activation (Tena et al., 2001; Zhang and Klessig,2001; Jonak et al., 2002; MAPK Group, 2002). For ex-ample, SIPK and WIPK are well-known tobacco MAPKsthat are activated together during defense responsesagainst TMV infection, elicitors, and wounding (Zhangand Klessig, 1998; Romeis et al., 1999; Liu et al., 2003).However, the immediate triggers for activation ofWIPK and SIPK themselves remain to be determined,although SIPK has been shown to respond to SA(Zhang and Klessig, 1997) and to ROI elements in-cluding hydrogen peroxide, superoxide, and ozone(Samuel et al., 2000). Activation of SIPK occurs withinminutes, suggesting that the responsible stimulant isROI rather than SA. This speculation is consistent witha report that the level of SIPK activity induced bycryptogein and harpin, a bacterial elicitor from Pseu-domonas syringae pv syringae, did not change in NahGtobacco plants in which SA does not accumulate dueto conversion into catechol (Lebrun-Garcia et al., 2002;Samuel et al., 2005). Our present finding that SIPKactivity was drastically suppressed in BY2 cells, inwhich hydrogen peroxide production was suppressed,further supports this idea. In wild-type BY2 cells, SIPKwas activated within 30 min after elicitation, whenhydrogen peroxide production in phase II had not yettaken place. This implies that the activation of SIPKprecedes polyamine oxidation, possibly being medi-ated by ROI generated in phase I through NADPHoxidase. However, activation of SIPK in phase I alonewas not sufficient to induce programmed cell death,hydrogen peroxide in phase II appeared to be a pre-requisite. This is in accordance with the finding thatthe phase I response does not necessarily correlatewith disease resistance (Baker and Orlandi, 1995).
Overall, this study with transgenic lines and phar-macological experiments provided strong evidencethat the majority of ROI in the late phase of oxidative
burst is derived from polyamine oxidation, and thatsuch ROI directly induces programmed cell death byactivating the MAPK cascade.
MATERIALS AND METHODS
Plant Materials and Chemical Treatments
Tobacco (Nicotiana tabacum) cultured cells and BY2 cells were grown in
Murashige and Skoog medium on a rotary shaker (115 rpm, 25�C) in the dark.
Cells in stationary phase at day 7 after subculture were used after dilution to a
1:50 ratio in fresh medium. A 5-mL aliquot of cells at day 7 was added to
50 mL of fresh medium containing aphidicolin (10 mg mL21) and cultured for
24 h for synchronization. For chemical treatments, synchronized cells were
collected by filtration, washed with fresh medium, diluted into 50 mL fresh
medium, and cultured for 1 h for acclimation. Cryptogein (Ponchet et al., 1999)
was added at an appropriate concentration as indicated for each experiment.
DFMO and guazatine were introduced into the medium 30 min before
cryptogein treatment at the concentrations indicated. Polyamines or hydrogen
peroxide were also added to the medium at the concentrations indicated in
other groups. For polyamine oxidase assays, guazatine and FAD were added
at appropriate concentrations as indicated in each experiment. Purified
guazatine triacetate (Mr 535.73) was commercially obtained (Wako) and
used at the concentration of 0.37 mM (0.2 mg mL21).
DNA Laddering and Fragmentation
DNA was extracted by the cetyltrimethyl ammonium bromide method
(Murray and Thompson, 1980), fractionated on agarose gel, and stained with
ethidium bromide. DNA fragmentation was examined using the TUNEL
method (in situ apoptosis detection kit, Takara; Tornusciolo et al., 1995)
according to the manufacturer’s protocol.
Quantitation of Cell Death
A 300 mL-aliquot of BY2 cells was incubated with 0.05% Evans blue for
15 min, washed several times to remove excess unbound dye, transferred
into 70 mL fresh medium, and examined for dead cells by light microscopy.
Data given are means from three replicate experiments, each with counts of
approximately 300 cells.
Chemiluminescence Assay
Production of hydrogen peroxide and superoxide was determined by
chemiluminescence using luminol and 2-methyl-6-phenyl-3, 7-dihydroimidazo-
[1,2-a]pyrazin-3-one as the respective reaction reagents. BY2 cells were
collected by filtration and washed several times with 5 mM MES buffer (pH
5.6) containing 175 mM mannitol, 0.5 mM CaCl2, and 0.5 mM K2SO4 (assay
buffer). After resuspension in assay buffer at the concentration of 50 mg fresh
weight per mL, samples were equilibrated by shaking at 115 rpm for 1 h at
25�C in the dark before cryptogein treatment. DFMO was added to the
medium 30 min before cryptogein treatment. After the indicated time period,
25-mL aliquots of cell suspension from each sample were subjected to chemi-
luminescence assay. Measurement was performed as previously described
(Yoda et al., 2003). Monitoring of hydrogen peroxide and potassium super-
oxide was performed with reference to results of adding known amounts of
the two products.
Quantitation of Polyamines
BY2 cells were collected by filtration and washed several times with
medium without Suc and 2,4-didhlorophenoxy-acetic acid. Proliferation was
monitored by directly measuring fresh weights of cells. Polyamines from
apoplasts were extracted with 50 mM MgCl2 (2.0 mL) in vacuo for 10 min.
A 500-mL aliquot of medium supernatant was collected by centrifugation and
the contained polyamines were derivatized with benzoyl chloride and sepa-
rated by HPLC as described (Yoda et al., 2003). Standard curves for estimation
were obtained by measuring a series of known amounts of each polyamine.
Role of Polyamine Oxidase in Programmed Cell Death
Plant Physiol. Vol. 142, 2006 203
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021

Assay for Polyamine Oxidase
BY2 cells were filtered through nylon mesh (20 mm) after an appropriate
time period and washed with fresh medium. Apoplastic proteins were
recovered by soaking cells in 10 mM Tris-HCl (pH 8.0; 2.5 mL) for 5 min and
further in vacuo for 10 min. A 10-mL aliquot of recovered fluid was subjected to
reaction with 5 mM of an appropriate polyamine substrate for 1 min, and
hydrogen peroxide was measured as described (Yoda et al., 2003).
Scavenging Activity Assay
BY2 cells were synchronized and cultured in fresh medium for 1 h, when
cryptogein was added to a final concentration of 50 nM. After incubation for
an appropriate time period, cells were collected and resuspended at 50 mg
fresh weight per mL of fresh medium containing 10 mM hydrogen peroxide.
Scavenging activity was estimated by measuring decrease of relative light
units (RLUs) in a chemiluminescence assay as described above after incuba-
tion at room temperature for the indicated time period.
Isolation of a cDNA for Polyamine Oxidase
Total RNA was prepared from TMV-infected tobacco leaves as described
above and poly (A)1 RNA was purified with a mRNA purification kit
(Pharmacia). A cDNA library was constructed using a l-ZAP cDNA synthesis
kit (Stratagene) with modification. The cDNAs, synthesized by reverse tran-
scriptase (Super Script II, Gibco BRL) from 5 mg of poly (A)1 RNA with linker
primers (anchored primers), were blunted and ligated to an EcoRI adaptor and
after phosphorylation of adaptor ends, digestion with XhoI was performed,
followed by fractionation by size with a SizeSep 400 Spun Column (Pharmacia).
The fractionated cDNA was ligated to Uni-ZAP XR Vector Arms and packaged
in lphage with MaxPlaxTM Lambda Packaging extract (Epicentre Technol-
ogy). The resulting fragment, designated as NtPAO, was used as a probe to
screen 2 3 105 plaques of the TMV-induced tobacco cDNA library. Positive
plaques were purified through three successive plaque hybridizations and
cDNA inserts were rescued in pBluescript SK(1) plasmids by ExAssist helper
phage-mediated in vivo excision and sequenced.
Bacterial Expression and Activity Assay
The cDNA of NtPAO was amplified by PCR with a set of synthetic
oligonucleotide primers containing a BamHI site (5#-GGATCCAAGGTGTTA-
TCGGAGAATGG-3#) and a SalI site (5#-GTCGACTCATAAGATAGCCTCTG-
GAAGTC-3#), respectively, using cloned pBluscript II SK(-)-NtPAO as the
template. The resulting fragment was ligated to the corresponding site of
pGEX-4T vector, which was transformed into Escherichia coli BL21. Bacterial
cells were grown at 37�C for 3 h and further incubated at 18�C for 12 h after
addition of isopropyl b-D-thiogalactopyranoside to a final concentration of
0.5 mM. Recombinant protein was purified with glutathione-Sepharose 4B
(Amersham) and subjected to polyamine oxidase assay as described above.
Approximately 4 mg recombinant protein was used for one reaction.
Transient Expression and Subcellular Localization
A fragment of NtPAO was amplified by PCR with a set of synthetic
oligonucleotide primers containing a SalI site (5#-GTCGACATGGCAACTCC-
CCG-3#) and a NcoI site (5#-CCATGGATAAGATAGCCTCTGGAAGTCC-3#),using cloned pBluscript II SK(-)-NtPAO as a template. This fragment was
introduced into the SalI/NcoI site of the CaMV35S-sGFP(S65T)-NOS3# vector,
creating an in-frame fusion product between the coding region and GFP. The
expression construct was delivered into onion (Allium cepa) epidermal cells by
particle bombardment as previously described (Yoda et al., 2002) and after
incubation of transformed cells for 8 h at 25�C in the dark, GFP was detected
by fluorescence microscopy.
Immunoblot Analysis
Apoplastic fluids were recovered as described above. Cytoplasmic con-
tents from remaining cells were extracted in 10 mM Tris-HCl (pH 6.8) with
ultrasonic. For immunoblot analysis, 5 mg each of soluble protein from
apoplasts and cytoplasm was separated on a 10% SDS-polyacrylamide gel and
transferred to a polyvinylidine fluoride membrane (Millipore). After blocking
overnight in 137 mM NaCl, 2.7 mM KCl, 1.5 mM KH2PO4, and 8.1 mM Na2hPO4
(PBS) containing 5% skim milk at 4�C, the membrane was incubated with
antibodies against PEPC diluted with PBS for 1 h at room temperature.
After several washings with PBS containing 0.1% Tween 20, the membrane
was incubated with horseradish peroxidase-conjugated anti-goat IgG anti-
bodies (Bio-Rad). Antibody-antigen complex was detected using ECL system
(Amersham).
Transformation of BY2 Cells
To generate transgenic BY2 cells with suppressed polyamine oxidase
activity, the pKANNIBAL (Wesley et al., 2001) vector designed for producing
hairpin RNA with a loop was employed. For sense orientation, the coding
region of NtPAO cDNA was amplified by PCR with a set of synthetic ol-
igonucleotide primers containing a XhoI site (5#-AGCTCGAGGTAATCGGTT-
TCAGC-3#) and a KpnI site (5#-GTGGTACCACTAAGGAAGAGTTGTCG-3#).For antisense orientation, the same region was amplified with a set of
synthetic oligonucleotide primers containing a BamHI site (5#-ACGGATCCG-
GAATAATCGGTTTCAGC-3#) and a ClaI site (5#-GTATCGATACTAAGGAA-
GAGTTGTCG-3#). The obtained fragments were introduced into the pGEM-T
Easy vector following digestion with designed restriction enzymes. Digested
fragments were ligated to the corresponding sites of the pKANNIBAL vector
and the resulting plasmid was digested with NotI and ligated to the cor-
responding pART27 vector, which was introduced into Agrobacterium tume-
faciens strain EHA105 cells. The final construct was introduced into tobacco
BY2 cells as previously described (Yamaguchi et al., 2003).
RNA Isolation and Gel-Blot Analysis
Total RNA was isolated by the acid guanidine thiocyanate-phenol/chloroform
method (Chomczynski and Sacchi, 1987) and gel-blot analyses were performed
as detailed earlier (Yoda et al., 2002) with probes produced as previously
described (Yoda et al., 2003). Probes for HSR203J and HIN1were synthesized
with a pair of specific primers: forward (5#-ATGGTTCATGAAAAGCAAGT-
GATAGAGG-3#) and reverse (5#-GCTTGTTGATGAACTCTGCAACGGCTTC-3#)for HSR203J, and forward (5#-CCCTTCCATTCCGCCACCAGCAAAATCC-3#)and reverse (5#-CTACCAATCAAGATGGCATCTGGTTTCC-3#) for HIN1.
In-Gel Kinase Activity Assay
In-gel kinase assays were performed as described previously (Zhang and
Klessig, 1997). Extracts containing 20 mg of protein were subjected to electro-
phoresis on 10% SDS-polyacrylamide gels embedded with 0.25 mg/mL of
myelin basic protein as a substrate for kinase. After electrophoresis, SDS was
removed by washing the gels with washing buffer (25 mM Tris-HCl, pH 7.5,
0.5 mM dithiothreitol [DTT], 0.1 mM Na3V04, 5 mM NaF, 0.5 mg mL21 bovine
serum albumin, and 0.1% Triton X-100 [v/v]) three times, each for 30 min at
room temperature. Protein kinase was allowed to renature in 25 mM Tris-HCl,
pH 7.5, 1 mM DTT, 0.1 mM Na3V04, and 5 mM NaF at 4�C for 16 h with three
changes of the buffer, and the gels were then incubated at room temperature in
a 30 mL reaction buffer (25 Tris-HCl, pH 7.5, 2 mM EGTA, 12 mM MgCl2, 1 mM
DTT, and 0.1 mM Na3V04) with 200 nM ATP plus 50 mCi [g-32P]ATP (3,000
Ci/mmol) for 60 min. The reaction was stopped by transferring the gel into 5%
trichloroacetic acid (w/v)/1% NaPPi (w/v). The unincorporated [g-32P]ATP
was removed by washing in the same solution for at least 6 h with five changes
and gels were then finally dried on Whatman 3MM paper for exposure of
x-ray film.
Sequence data from this article can be found in the GenBank/EMBL data
libraries under accession number AB200262.
ACKNOWLEDGMENTS
The authors thank Drs. Michel Ponchet and Jean-Claude Pernollet (Institut
National de la Recherche Agronomique, France), and Toyoki Amano
(Shizuoka University) for generous provision of cryptogein; Takashi Hashimoto
(Nara Institute of Science and Technology) and Kenzo Nakamura (Nagoya
University) for cDNA clones of spermidine synthase and S-adenosyl-
methionine decarboxylase, respectively; and Dr. Malcolm Moore (Intermal,
Nagoya) for critical reading of the manuscript.
Yoda et al.
204 Plant Physiol. Vol. 142, 2006
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021

Received March 15, 2006; accepted July 12, 2006; published July 14, 2006.
LITERATURE CITED
Allan AC, Fluhr R (1997) Two distinct sources of elicited reactive oxygen
species in tobacco epidermal cells. Plant Cell 9: 1559–1572
Baker CJ, Orlandi EW (1995) Active oxygen in plant pathogenesis. Annu
Rev Phytopathol 33: 299–321
Bestwick CS, Brown IR, Bennett MH, Mansfield JW (1997) Localization of
hydrogen peroxide accumulation during the hypersensitive reaction of
lettuce cells to Pseudomonas syringae pv phaseolicola. Plant Cell 9: 209–221
Binda C, Angelini R, Federico R, Ascenzi P, Mattevi A (2001) Structural
bases for inhibitor binding and catalysis in polyamine oxidase.
Biochemistry 40: 2766–2776
Binet M, Humbert C, Lecourieux D, VantardM, Pugin A (2001) Disruption
of microtubular cytoskeleton induced by cryptogein, an elicitor of
hypersensitive response in tobacco cells. Plant Physiol 125: 564–572
Bolwell GP, Wojitaszek P (1997) Mechanisms for the generation of reactive
oxygen species in plant defense: a broad perspective. Physiol Mol Plant
Pathol 51: 21–30
Bradley D, Kjellbom P, Lamb C (1992) Elicitor- and wound-induced
oxidative cross-linking of a proline-rich plant cell wall protein: a novel,
rapid defense response. Cell 7: 21–30
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation
by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal
Biochem 162: 156–159
Cowley T, Walters DR (2002) Polyamine metabolism in barley reacting
hypersensitively to the powdery mildew fungus Blumeria graminis f. sp.
hordei. Plant Cell Environ 25: 461–468
Draper J (1997) Salicylate, superoxide synthesis and cell suicide in plant
defence. Trends Plant Sci 2: 162–165
Facchiano F, D’Arcangelo D, Riccomi A, Lentini A, Beninati S, Capogrossi
MC (2001) Transglutaminase activity is involved in polyamine-induced
programmed cell death. Exp Cell Res 271: 118–129
Grant JJ (1997) Salicilate, superoxide synthesis and cell suicide in plant
defense. Trends Plant Sci 2: 162–165
Grant JJ, Loake GJ (2000) Role of reactive oxygen intermediates
and cognate redox signaling in disease resistance. Plant Physiol 124:
21–29
Groom QJ, Torres MA, Fordham-Skelton AP, Hammond-Kosack KE,
Robinson NJ, Jones JD (1996) rbohA, a rice homologue of the mam-
malian gp91phox respiratory burst oxidase gene. Plant J 10: 515–522
Ha HC, Woster PM, Yager JD, Casero RA (1997) The role of polyamine
catabolism in polyamine analogue-induced programmed cell death.
Proc Natl Acad Sci USA 94: 11557–11562
Hammond-Kosack KE, Jones JDG (1996) Resistance gene-dependent plant
defense responses. Plant Cell 8: 1773–1791
Hu X, Bidney DL, Yalpani N, Duvick JP, Crasta O, Folkerts O, Lu G (2003)
Overexpression of a gene encoding hydrogen peroxide-generating
oxalate oxidase evokes defense responses in sunflower. Plant Physiol
133: 170–181
Jonak C, Okresz L, Bogre L, Hirt H (2002) Complexity, crosstalk and
integration of plant MAP kinase signalling. Curr Opin Plant Biol 5:
415–424
Kadota Y, Goh T, Tomatsu H, Tamauchi R, Higashi K, Muto S, Kuchitsu K
(2004) Cryptogein-induced initial events in tobacco BY-2 cells: pharma-
cological characterization of molecular relationship among cytosolic
Ca21 transients, anion efflux and production of reactive oxygen species.
Plant Cell Physiol 45: 160–170
Kawasaki T, Koita H, Nakatsubo T, Hasegawa K, Wakabayashi K,
Takahashi H, Umemura K, Umezawa T, Shimamoto K (2006)
Cinnamoyl-CoA reductase, a key enzyme in lignin biosynthesis, is an
effector of small GTPase Rac in defense signaling in rice. Proc Natl Acad
Sci USA 103: 230–235
Keller T, Damude HG, Werner D, Doerner P, Dixon RA, Lamb C (1998) A
plant homolog of the neutrophil NADPH oxidase gp91phox subunit
gene encodes a plasma membrane protein with Ca21 binding motifs.
Plant Cell 10: 255–266
Lamb C, Dixon RA (1997) The oxidative burst in plant disease resistance.
Annu Rev Plant Physiol Plant Mol Biol 48: 251–275
Lamotte O, Gould K, Lecourieux D, Sequeira-Legrand A, Lebrun-Garcia
A, Durner J, Pugin A, Wendehenne D (2004) Analysis of nitric oxide
signaling functions in tobacco cells challenged by the elicitor crypto-
gein. Plant Physiol 135: 1–14
Lebrun-Garcia A, Chiltz A, Gout E, Bligny R, Pugin A (2002) Questioning
the role of salicylic acid and cytosolic acidification in mitogen-activated
protein kinase activation induced by cryptogein in tobacco cells. Planta
214: 792–797
Levine A, Tenhaken R, Dixon R, Lamb C (1994) H2O2 from the oxidative
burst orchestrates the plant hypersensitive disease resistance response.
Cell 79: 583–593
Lindsay GS, Wallace HM (1999) Changes in polyamine catabolism in
HL-60 human promyelogenous leukaemic cells in response to etoposide-
induced apoptosis. Biochem J 337: 83–87
Liu Y, Jin H, Yang K-Y, Kim CY, Baker B, Zhang S (2003) Interaction
between two mitogen-activated protein kinases during tobacco defense
signaling. Plant J 34: 149–160
MAPKGroup (2002) Mitogen-activated protein kinase cascades in plants: a
new nomenclature. Trends Plant Sci 7: 301–308
Murray MG, Thompson WF (1980) Rapid isolation of high molecular
weight plant DNA. Nucleic Acids Res 8: 4321–4325
Papadakis AK, Roubelakis-Angelakis KA (2005) Polyamines inhibit
NADPH oxidase-mediated superoxide generation and putrescine pre-
vents programmed cell death induced by polyamine oxidase-generated
hydrogen peroxide. Planta 220: 826–837
Polticelli F, Basran J, Faso C, Cona A, Minervini G, Angelini R, Federico R,
Scrutton NS, Tavladoraki P (2005) Lys300 plays a major role in the
catalytic mechanism of maize polyamine oxidase. Biochemistry 44:
16108–16120
Ponchet M, Panabieres F, Milat ML, Mikes V, Montillet JL, Suty L,
Triantaphylides C, Tirilly Y, Blein JP (1999) Are elicitins cryptograms
in plant-oomycete communications? Cell Mol Life Sci 56: 1020–1047
Romeis T, Piedras P, Zhang S, Klessig DF, Hirt H, Jones JDG (1999) Rapid
Avr 9- and Cf-9–dependent activation of MAP kinases in tobacco cell
cultures and leaves: convergence of resistance gene, elicitor, wound, and
salicylate responses. Plant Cell 11: 273–287
Rusterucci C, Stallaert V, Milat ML, Pugin A, Ricci P, Blein JP (1996)
Relationship between active oxygen species, lipid peroxidation, necro-
sis, and phytoalexin production induced by elicitins in Nicotiana. Plant
Physiol 111: 885–891
Sagi M, Fluhr R (2001) Superoxide production by plant homologues of the
gp91 phox NADPH oxidase: modulation of activity by calcium and by
tobacco mosaic virus infection. Plant Physiol 126: 1281–1290
Samuel MA, Hall H, Krzymowska M, Drzewiecka K, Hennig J, Ellis BE
(2005) SIPK signaling controls multiple components of harpin-induced
cell death in tobacco. Plant J 42: 406–416
Samuel MA, Miles GP, Ellis BE (2000) Ozone treatment rapidly activates
MAP kinase signaling in plants. Plant J 22: 367–376
Sharma PC, Ito A, Shimizu T, Terauchi R, Kamoun S, Saitoh H (2003)
Virus-induced silencing of WIPK and SIPK genes reduces resistance to a
bacterial pathogen, but has no effects on the INF-1-induced hypersen-
sitive response (HR) in Nicotiana benthamiana. Mol Genet Genom 269:
583–591
Simon-Plas F, Elmayan T, Blein JP (2002) The plasma membrane oxidase
NtrbohD is responsible for AOS production in elicited tobacco cells.
Plant J 31: 137–147
Simon-Plas F, Rusterucci C, Milat ML, Humbert C, Montillet JL, Blein JP
(1997) Active oxygen species production in tobacco cells elicited by
cryptogein. Plant Cell Environ 20: 1573–1579
Sebela M, Radova A, Angelini R, Tavladoraki P, Frebort I, Pec P (2001)
FAD-containing polyamine oxidases: a timely challenge for researchers
in biochemistry and physiology of plants. Plant Sci 160: 197–207
Suzuki K, Yano A, Shinshi H (1999) Slow and prolonged activation of the
p47 protein kinase during hypersensitive cell death in a culture of
tobacco cells. Plant Physiol 119: 1465–1472
Takahashi Y, Berberich T, Miyazaki A, Seo S, Ohashi Y, Kusano T (2003)
Spermine signalling in tobacco: activation of mitogen-activated protein
kinases by spermine is mediated through mitochondrial dysfunction.
Plant J 36: 820–829
Tena G, Asai T, Chiu W-L, Sheen J (2001) Plant mitogen-activated protein
kinase signaling cascades. Curr Opin Plant Biol 4: 392–400
Thordal CH, Zhang Z, Wei Y, Collinge DB (1997) Subcellular localization
of H2O2 in plants: H2O2 accumulation in papillae and hypersensitive
response during the barley-powdery mildew interaction. Plant J 11:
1187–1194
Role of Polyamine Oxidase in Programmed Cell Death
Plant Physiol. Vol. 142, 2006 205
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021

Tornusciolo DR, Schmidt RE, Roth KA (1995) Simultaneous detection of
TDT-mediated dUTP-biotin nick end-labeling (TUNEL)-positive cells
and multiple immunohistochemical markers in single tissue sections.
Biotechniques 19: 800–805
Torres MA, Dangl JL, Jones JDG (2002) Arabidopsis gp91 phox homologues
AtrbohD and AtrbohF are required for accumulation of reactive oxygen
intermediates in the plant defense response. Proc Natl Acad Sci USA 99:
517–522
Wesley SV, Halliwel CA, Smith NA, Wang M, Rouse DT, Liu O, Gooding
PS, Singh SP, Abbott D, Stoutjesdijk PA, et al (2001) Construct design
for efficient, effective and high-throughput gene silencing in plants.
Plant J 27: 581–590
Yamaguchi Y, Yoda H, Akiyama K, Sano H (2003) Induction of transcripts
encoding a novel seven-transmembrane protein during the hypersensi-
tive response to tobacco mosaic virus infection in tobacco plants. Planta
218: 249–253
Yamakawa H, Kamada H, Satoh M, Ohashi Y (1998) Spermine is a
salicylate-independent endogenous inducer for both tobacco acidic
pathogenesis-related proteins and resistance against tobacco mosaic
virus infection. Plant Physiol 118: 1213–1222
Yang Y, Shah J, Klessig DF (1997) Signal perception and transduction in
plant defense responses. Genes Dev 11: 1621–1639
Yoda H, Ogawa M, Yamaguchi Y, Koizumi N, Kusano T, Sano H (2002)
Identification of early-responsive genes associated with the hyper-
sensitive response to tobacco mosaic virus and characterization of a
WRKY-type transcription factor in tobacco plants. Mol Genet Genomics
267: 154–161
Yoda H, Yamaguchi Y, Sano H (2003) Induction of hypersensitive cell death
by hydrogen peroxide produced through polyamine degradation in
tobacco plants. Plant Physiol 132: 1973–1981
Zhang S, Klessig DF (1997) Salicylic acid activates a 48-kD MAP kinase in
tobacco. Plant Cell 9: 809–824
Zhang S, Klessig DF (1998) N resistance gene-mediated de novo synthesis
and activation of a tobacco MAP kinase by TMV infection. Proc Natl
Acad Sci USA 95: 7433–7438
Zhang S, Klessig DF (2001) MAPK cascades in plant defense signaling.
Trends Plant Sci 6: 520–527
Zhang S, Liu Y, Klessig DF (2000) Multiple levels of tobacco WIPK
activation during the induction of cell death by fungal elicitors. Plant J
23: 339–347
Yoda et al.
206 Plant Physiol. Vol. 142, 2006
Dow
nloaded from https://academ
ic.oup.com/plphys/article/142/1/193/6106453 by guest on 20 O
ctober 2021





























