Embed Size (px)
Citation preview

of January 3, 2019.This information is current as
upon TLR4 StimulationBκNuclear Export and Retention of p65 NF-
Determining Factor in Crm1-Mediated Poly(ADP-Ribose) Polymerase-1 Is a
BoularesShahriar Koochekpour, Andrew Catling and A. Hamid Chetan P. Hans, Jong G. Kim, Zakaria Y. Abd Elmageed,Yasuhiro Suzuki, Hogyoung Kim, Jihang Ju, Tao Liu, Mourad Zerfaoui, Youssef Errami, Amarjit S. Naura,
ol.1000646http://www.jimmunol.org/content/early/2010/07/07/jimmun
published online 7 July 2010J Immunol
MaterialSupplementary
6.DC1http://www.jimmunol.org/content/suppl/2010/07/06/jimmunol.100064
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. All rights reserved.1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 3, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Poly(ADP-Ribose) Polymerase-1 Is a Determining Factor inCrm1-Mediated Nuclear Export and Retention of p65 NF-kBupon TLR4 Stimulation
Mourad Zerfaoui,* Youssef Errami,* Amarjit S. Naura,* Yasuhiro Suzuki,*
Hogyoung Kim,* Jihang Ju,* Tao Liu,† Chetan P. Hans,* Jong G. Kim,‡
Zakaria Y. Abd Elmageed,x Shahriar Koochekpour,† Andrew Catling,* and
A. Hamid Boulares*,x
The role of NF-kB in the expression of inflammatory genes and its participation in the overall inflammatory process of chronic
diseases and acute tissue injury are well established. We and others have demonstrated a critical involvement of poly(ADP-ribose)
polymerase (PARP)-1 during inflammation, in part, through its relationship with NF-kB. However, the mechanism by which
PARP-1 affects NF-kB activation has been elusive. In this study, we show that PARP-1 inhibition by gene knockout, knockdown,
or pharmacologic blockade prevented p65 NF-kB nuclear translocation in smooth muscle cells upon TLR4 stimulation, NF-kB
DNA-binding activity, and subsequent inducible NO synthase and ICAM-1 expression. Such defects were reversed by reconstitution
of PARP-1 expression. PARP-1 was dispensable for LPS-induced IkBa phosphorylation and subsequent degradation but was
required for p65 NF-kB phosphorylation. A perinuclear p65 NF-kB localization in LPS-treated PARP-12/2 cells was associated
with an export rather an import defect. Indeed, whereas PARP-1 deficiency did not alter expression of importin a3 and importin a4
and their cytosolic localization, the cytosolic levels of exportin (Crm)-1 were increased. Crm1 inhibition promoted p65 NF-kB
nuclear accumulation as well as reversed LPS-induced p65 NF-kB phosphorylation and inducible NO synthase and
ICAM-1 expression. Interestingly, p65 NF-kB poly(ADP-ribosyl)ation decreased its interaction with Crm1 in vitro. Pharmacologic
inhibition of PARP-1 increased p65 NF-kB–Crm1 interaction in LPS-treated smooth muscle cells. These results suggest that p65
NF-kB poly(ADP-ribosyl)ation may be a critical determinant for the interaction with Crm1 and its nuclear retention upon TLR4
stimulation. These results provide novel insights into the mechanism by which PARP-1 promotes NF-kB nuclear retention, which
ultimately can influence NF-kB–dependent gene regulation. The Journal of Immunology, 2010, 185: 000–000.
The role of poly(ADP-ribose) polymerase (PARP)-1 in in-flammation has been investigated intensely in the contextof its direct participation, by way of its catalytic activity, in
cellular responses to DNA-damaging agents, including oxidativestress (1). In a number of pathologic conditions that involve mas-sive DNA damage, the excessive activation of PARP-1 depletescellular stores of both NAD and its precursor ATP, leading toirreversible cytotoxicity and, potentially, cell death (2–4). We re-cently showed that PARP-1 plays important roles in allergic
asthma and atherosclerosis (5–7). An emerging role for this pro-tein, however, is the ability of PARP-1 to participate, directly orindirectly, in the regulation of a number of inflammatory genes,especially those mediated by NF-kB (reviewed in Ref. 8). NF-kBis a pleiotropic transcription factor that plays a critical role inthe regulation of the expression of multiple genes involved in in-flammatory responses, including inducible NO synthase (iNOS) andadhesion molecules (9). NF-kB binds to the promoter regions oftarget genes as a dimer of two Rel family proteins, most frequentlyp50 and p65 (9, 10). In quiescent cells, NF-kB is sequestered in thecytoplasm as a result of its interaction with members of the IkBfamily of proteins, which includes IkBa and IkBb. IkBa is phos-phorylated, polyubiquitinated, and degraded by the 26S proteasomein response to cell stimulation, resulting in the release of the nu-clear localization signal of NF-kB and its subsequent translocationto the nucleus (9).Interestingly, while p65 NF-kB nuclear translocation in TNF-
treated smooth muscle cells (SMCs) was sufficient for the expres-sion of VCAM-1, we recently demonstrated that PARP-1 is requiredfor expression of ICAM-1 (11). The expression of ICAM-1 wasassociated with a transient interaction between PARP-1 and p65NF-kB when examined in COS-7 cells and in the airway epithelialcell line A549. We (5, 7, 11, 12) and others (13, 14) have reportedthat NF-kB nuclear translocation requires PARP-1 expression asassessed by EMSA upon TLR4 stimulation by LPS treatment. Thedefect in the nuclear translocation of NF-kB culminated in thesevere reduction in the expression of NF-kB-targeted genes suchas iNOS, MCP-1, cyclooxygenase-2, and adhesion molecules.
*Department of Pharmacology and Experimental Therapeutics, †Department of Mi-crobiology and Immunology, ‡Department of Pathology, and xStanley Scott CancerCenter, Louisiana State University Health Sciences Center, New Orleans, LA 70112
Received for publication February 24, 2010. Accepted for publication May 29, 2010.
This work was supported in part by National Institutes of Health Grants HL072889 and1P20RR18766 (to D.K.), by American Cancer Society Grant RSG-116608, and byfunds from the Louisiana Cancer Research Consortium (New Orleans, LA) (to H.B.).This work was also supported by National Center for Research Resources Grant 1P20RR021970/COBRE (to S.K.; principal investigator, A. Ochoa).
Address correspondence and reprint requests to Dr. A. Hamid Boulares andDr. Mourad Zerfaoui, Department of Pharmacology and Experimental Therapeutics,Louisiana State University Health Sciences Center, 1901 Perdido Street, New Or-leans, LA 70112. E-mail addresses: [email protected] and [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this paper: Ad, adenoviral; Con, control; EYFP, enhanced yel-low fluorescent protein; iNOS, inducible NO synthase; LMB, leptomycin B; MEF,mouse embryonic fibroblast; PARG, poly(ADP-ribose) glycohydrolase; PARP, poly(ADP-ribose) polymerase; shRNA, short hairpin RNA; shRNA-Con, control shRNA;shRNA-PARP-1, shRNA targeting PARP-1; SMC, smooth muscle cell; WT, wild type.
Copyright� 2010 by The American Association of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1000646
Published July 7, 2010, doi:10.4049/jimmunol.1000646 by guest on January 3, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

The nuclear localization signal embedded within NF-kB binds toimportins and thus promotes nuclear translocation of thetranscription factor by allowing its passage through the nuclearpore complex (15). The import of p65 NF-kB to the nucleusupon stimulation has been attributed predominantly to importina3 and importin a4 (15). Additionally, NF-kB has been reportedto interact with exportins, which through a multifactor complex cantransport the transcription factor from the nucleus to the cytosol(16). Crm1, an important member of the nuclear export machinery,recognizes and exports proteins containing a leucine-rich nuclearexport signal (17). Crm1 binds cooperatively with RanGTP and thecargo to form a trimeric complex, nuclear export signal–RanGTP–Crm1, which then exports through the nuclear pore complexes.Themechanisms bywhich PARP-1, a nuclear enzyme, influences
cytosolic events leading to the release of NF-kB from the cytosoland translocation to the nucleus are not understood. Thus, the goalsof the present study were to determine the mechanisms by whichPARP-1 participates in the retention of NF-kB in the nucleus uponTLR4 stimulation by LPS.
Materials and MethodsAnimals, isolation of SMCs and fibroblasts, and treatmentprotocols
Mice were bred in a specific pathogen-free facility at the Louisiana StateUniversity Health Sciences Center (New Orleans, LA) and allowed unlim-ited access to sterilized chow and water. Maintenance, experimental proto-cols, and procedures were all approved by the Louisiana State UniversityHealth Sciences Center Animal Care and Use Committee. C57BL/6 wildtype (WT) mice were purchased from The Jackson Laboratory (Bar Harbor,ME). The generation of C57BL/6 PARP-12/2 mice was previouslydescribed (6). All animals were genotyped by PCR.
Isolation and assessment of purity of SMC preparations were conductedas described (11). SMCs were used between passages 4 and 7. Mouseembryonic fibroblasts (MEFs) were isolated as described (18). 293T cellsand SMCs were maintained in DMEM supplemented with 10% FBS,penicillin, and streptomycin. Prior to treatment, SMCs at 50–80% con-fluence were starved by incubation in DMEM/F12 with 0.5% cell culture-tested BSA (Sigma-Aldrich, St. Louis, MO) for 18 h. Cells were treatedwith 1 mg/ml LPS (Axxora, San Diego, CA) for the indicated times in theabsence or presence 50 mM of the PARP-1 inhibitor NU1025 (Santa CruzBiotechnology, Santa Cruz, CA) or 1 mM 3-aminobenzamide (Santa CruzBiotechnology). In some experiments, cells were treated with 1 mM H2O2
for the indicated intervals before processing for immunofluorescence. Formicroscopy, cells were incubated in CO2-independent medium.
Total, cytoplasmic, and nuclear extract preparations andimmunoblot analysis
After treatment with the indicated agents, cells were washed with ice-coldPBS followed by centrifugation. For total protein extracts, cell pellets wereincubated for 15 min on ice in lysis buffer supplemented with proteases andphosphatase inhibitors as previously described (19). The preparation ofcytoplasmic and nuclear extracts was performed using a commercial kit(Active Motif, Carlsbad, CA) according to the manufacturer’s instructions.A portion (15 mg of protein) of each lysate was then fractionated by SDS-PAGE on a 4–20% gradient gel, and the separated proteins were transferredto a nitrocellulose filter. The filter was stained with Ponceau S to confirmequal loading and transfer of samples, and was then probed with Abs toIkBa, ICAM-1, VCAM, iNOS, Crm1, p65 NF-kB, poly(ADP-ribose) gly-cohydrolase (PARG), or actin (Santa Cruz Biotechnology), PARP-1 (BDPharmingen, San Jose, CA), importin a3 (Sigma-Aldrich), importin a4(Imgenex, San Diego, CA), GST (Novus Biologicals, Littleton, CO), as wellas Abs to the phosphorylated form (serine residues 32 and 36) of IkBa (CellSignaling Technology, Beverly, MA), or of p65 NF-kB at Ser276, Ser529, orSer536. Immune complexes were detected with appropriate secondary Absand chemiluminescence reagents (PerkinElmer Life Science, Boston, MA).
Conventional RT-PCR
RNA was extracted from cells using standard methods, and cDNA wasgenerated using Reverse Transcriptase III (Invitrogen, Carlsbad, CA). Oli-gonucleotide primers to specifically amplify a fragment of ICAM-1, iNOS, or
b-actin were purchased from Integrated DNATechnologies (Coralville, IA).The specific primers were as follows: ICAM-1, forward primer, 59-TCCTAAAATGACCTGCAGACG-39, reverse primer, 59-AGTTTTATGGCCTCCTCC TGA-39; iNOS, forward primer, 59-CAG CTG GGC TGT ACA AACCTT-39, reverse primer, 59-CAT TGG AAG TGA AGC GTT TCG-39; andb-actin, forward primer, 59-ACC GTG AAA AGATGA CCC AGATC-39,reverse primer, 59-TAG TTT CAT GGA TGC CAC AGG-39. Theamplification program was as follows: 3 min at 95˚C, 30 s at 95˚C, 45 s at60˚C, and 45 s at 72˚C. The cycle numbers were optimized for each primerpair. The PCR products were then incubated for 15min at 72˚C. The resultingPCR products were subjected to electrophoresis in a 2% agarose gel andstained with ethidium bromide.
Construction of the GST–p65 NF-kB, p65 NF-kB–enhancedyellow fluorescent protein, and PARP-1–EYFP plasmids, aswell as adenoviral vector–expressing PARP-1–EYFP,transduction, and transfections
Construction of the p65NF-kB–enhanced yellow fluorescent protein (EYFP)expression vector was described elsewhere (11). For the construction of theGST–p65 NF-kB expression vector, the p65 NF-kB cDNAwas cloned intoa Gateway entry vector, pENTR/SD/D-TOPO (Invitrogen). The accuracy ofthe construct was verified by sequence analysis. The expression clone wasgenerated by performing an LR recombination reaction between the entryclone and the destination vector GST-pDEST27 (Invitrogen).
SMCs were transiently transfected with p65 NF-kB–EYFP using theMirus TransIT-LT1 transfection reagent (Mirus Bio, Madison, WI) accord-ing to the manufacturer’s instructions. After starvation, SMCs were culturedin CO2-independent medium (Invitrogen) and treated with LPS as describedearlier (11). Subcellular localization of p65 NF-kB was monitoredthroughout the treatment using a Zeiss inverted fluorescence microscope.The PARP-1–EYFP expression vector was described previously (20). Allsequences were confirmed by sequencing. Knockdown of PARP-1 (sc-29438-V; Santa Cruz Biotechnology) or PARG (sc-152026-V; Santa CruzBiotechnology) in WT SMCs was achieved using lentiviral vectors encod-ing the respective targeting short hairpin RNA (shRNA), according to themanufacturer’s specifications and instructions, before treatment with theindicated reagents.
Pulldown assay, EMSA, and luciferase assay
293T cells were transiently transfected with the GST–p65 NF-kB vectorusing Lipofectamine LTX (Invitrogen) and were treated with 1 mg/ml LPS(Axxora) for the indicated times. GST–p65 NF-kB was pulled down usingGlutathione Sepharose 4 Fast Flow (GE Healthcare, Piscataway, NJ).EMSA analysis of DNA-binding activity in prepared nuclear extractswas performed as described (21). MEFs were transiently transfected witha total of 5 mg of the RapidReporter pRR-High-NF-kB plasmid (ActiveMotif) using Lipofectamine 2000 (Invitrogen) according to themanufacturers’ instructions. After treatment, cells were lysed (20 ml oflysis buffer per well) and then incubated for 30 min at room temperature.Substrate was added to extracts according to the manufacturers’instructions and measured on a TriStar LB 941 luminometer (BertholdTechnologies, Oak Ridge, TN).
Poly(ADP-ribosyl)ation in vitro
Purified recombinant GST–p65 or untagged p65 (Active Motif) NF-kB andpurified PARP were incubated in a mixture (25 ml) containing reactionbuffer (100 mM Tris-HCl, 1 mM DTT, 10 mM MgCl2), sonicated salmonsperm DNA, and 2 mM NAD (Sigma-Aldrich) for 30 min at 37˚C. Thereaction was either terminated by the addition of an equal volume of SDSsample buffer and heating at 95˚C for 5 min or subjected to pulldown withGST beads after addition of recombinant Crm1 protein. Samples were thensubjected to immunoblot analysis with Abs to poly(ADP-ribose) (Axxora),p65 NF-kB, or GST.
Indirect immunofluorescence
Cells grown on chamber slides were washed in PBS three times after theindicated treatment and then fixed with 3.7% paraformaldehyde for 20 minat room temperature. After permeabilization in 0.05% Triton X-100 in PBSfor 5 min, cells were incubated with primary Abs for 1 h at room temper-ature. Ab–Ag complexes were detected with Alexa Fluor 594-conjugatedsecondary Ab for 1 h at room temperature. The cells were thoroughlywashed after each incubation and then counterstained with DAPI. Cellswere examined under a fluorescence microscope (Leica Microsystems,Deerfield, IL) using a 340 objective lens.
2 REGULATION OF LPS-INDUCED NF-kB NUCLEAR RETENTION BY PARP-1
by guest on January 3, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
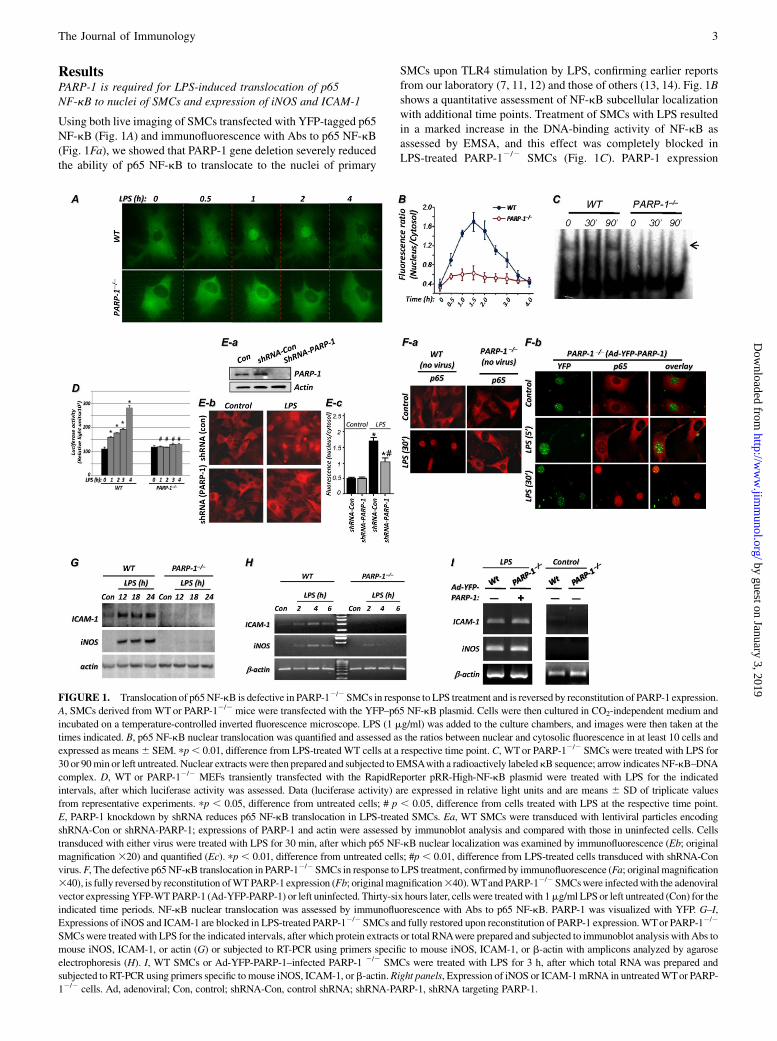
ResultsPARP-1 is required for LPS-induced translocation of p65NF-kB to nuclei of SMCs and expression of iNOS and ICAM-1
Using both live imaging of SMCs transfected with YFP-tagged p65NF-kB (Fig. 1A) and immunofluorescence with Abs to p65 NF-kB(Fig. 1Fa), we showed that PARP-1 gene deletion severely reducedthe ability of p65 NF-kB to translocate to the nuclei of primary
SMCs upon TLR4 stimulation by LPS, confirming earlier reportsfrom our laboratory (7, 11, 12) and those of others (13, 14). Fig. 1Bshows a quantitative assessment of NF-kB subcellular localizationwith additional time points. Treatment of SMCs with LPS resultedin a marked increase in the DNA-binding activity of NF-kB asassessed by EMSA, and this effect was completely blocked inLPS-treated PARP-12/2 SMCs (Fig. 1C). PARP-1 expression
FIGURE 1. Translocation of p65NF-kB is defective in PARP-12/2SMCs in response to LPS treatment and is reversed by reconstitution of PARP-1 expression.
A, SMCs derived from WTor PARP-12/2 mice were transfected with the YFP–p65 NF-kB plasmid. Cells were then cultured in CO2-independent medium and
incubated on a temperature-controlled inverted fluorescence microscope. LPS (1 mg/ml) was added to the culture chambers, and images were then taken at the
times indicated. B, p65 NF-kB nuclear translocation was quantified and assessed as the ratios between nuclear and cytosolic fluorescence in at least 10 cells and
expressed as means6 SEM. pp, 0.01, difference from LPS-treated WT cells at a respective time point. C, WTor PARP-12/2 SMCs were treated with LPS for
30 or 90min or left untreated. Nuclear extracts were then prepared and subjected to EMSAwith a radioactively labeledkB sequence; arrow indicates NF-kB–DNA
complex. D, WT or PARP-12/2 MEFs transiently transfected with the RapidReporter pRR-High-NF-kB plasmid were treated with LPS for the indicated
intervals, after which luciferase activity was assessed. Data (luciferase activity) are expressed in relative light units and are means 6 SD of triplicate values
from representative experiments. pp , 0.05, difference from untreated cells; # p , 0.05, difference from cells treated with LPS at the respective time point.
E, PARP-1 knockdown by shRNA reduces p65 NF-kB translocation in LPS-treated SMCs. Ea, WT SMCs were transduced with lentiviral particles encoding
shRNA-Con or shRNA-PARP-1; expressions of PARP-1 and actin were assessed by immunoblot analysis and compared with those in uninfected cells. Cells
transduced with either virus were treated with LPS for 30 min, after which p65 NF-kB nuclear localization was examined by immunofluorescence (Eb; original
magnification 320) and quantified (Ec). pp , 0.01, difference from untreated cells; #p , 0.01, difference from LPS-treated cells transduced with shRNA-Con
virus.F, The defective p65NF-kB translocation in PARP-12/2SMCs in response to LPS treatment, confirmed by immunofluorescence (Fa; originalmagnification
340), is fully reversed by reconstitution ofWTPARP-1 expression (Fb; originalmagnification340).WTand PARP-12/2SMCswere infectedwith the adenoviral
vector expressingYFP-WTPARP-1 (Ad-YFP-PARP-1) or left uninfected. Thirty-six hours later, cells were treatedwith 1mg/ml LPSor left untreated (Con) for the
indicated time periods. NF-kB nuclear translocation was assessed by immunofluorescence with Abs to p65 NF-kB. PARP-1 was visualized with YFP. G–I,
Expressions of iNOS and ICAM-1 are blocked in LPS-treated PARP-12/2 SMCs and fully restored upon reconstitution of PARP-1 expression. WTor PARP-12/2
SMCswere treated with LPS for the indicated intervals, after which protein extracts or total RNAwere prepared and subjected to immunoblot analysis with Abs to
mouse iNOS, ICAM-1, or actin (G) or subjected to RT-PCR using primers specific to mouse iNOS, ICAM-1, or b-actin with amplicons analyzed by agarose
electrophoresis (H). I, WT SMCs or Ad-YFP-PARP-1–infected PARP-1 2/2 SMCs were treated with LPS for 3 h, after which total RNA was prepared and
subjected to RT-PCR using primers specific tomouse iNOS, ICAM-1, orb-actin.Right panels, Expression of iNOS or ICAM-1mRNA in untreatedWTor PARP-
12/2 cells. Ad, adenoviral; Con, control; shRNA-Con, control shRNA; shRNA-PARP-1, shRNA targeting PARP-1.
The Journal of Immunology 3
by guest on January 3, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

appeared to be required for the expression of ICAM-1 and iNOS aswell as other NF-kB–dependent genes in cells other than SMCs.Indeed, expression of ICAM-1 (see Supplemental Fig. 1) and iNOS(7, 22) was blocked in LPS-treated PARP-12/2 macrophages.Furthermore, in MEFs transiently transfected with the RapidRe-porter pRR-High-NF-kB plasmid, treatment with LPS induceda pronounced increase in luciferase activity; such an increase wascompletely blocked in the PARP-12/2 counterparts (Fig. 1D),which is consistent with published reports (5, 14, 23). PARP-1knockdown with a lentiviral vector encoding a shRNA targetingmouse PARP-1 (Fig. 1Ea) significantly reduced nuclear localiza-tion of p65 NF-kB upon LPS treatment of WT SMCs (Fig. 1Eb,Ec). The impairment of p65 NF-kB nuclear translocation in LPS-treated PARP-12/2 cells (Fig. 1Fa) was fully reversed upon re-constitution of PARP-1 expression using adenovirus gene transfer(Ad–YFP–PARP-1) as assessed by immunofluorescence with Absto p65 NF-kB (Fig. 1Fb). PARP-1 gene deletion markedly reducedexpression of iNOS and ICAM-1 at the protein (Fig. 1G) andmRNA levels (Fig. 1H). The expression of iNOS and ICAM-1was fully restored by adenoviral-mediated reconstitution of PARP-1expression (Fig. 1I), confirming the specificity and validity of ourobservations.
PARP-1 expression is dispensable for IkBa phosphorylationand subsequent degradation but is required for p65 NF-kBphosphorylation in response to TLR4 stimulation
We next investigated whether PARP-1 gene deletion preventedNF-kB nuclear translocation in response to LPS by affecting thecritical events leading to its activation and consequent nucleartranslocation as well as its subsequent posttranslocation phos-phorylation. Surprisingly, IkBa phosphorylation and subsequentdegradation were almost identical in both LPS-treated WT andPARP-12/2 SMCs (Fig. 2A) with the clear preservation of thetransient nature of IkBa phosphorylation and degradation. Theseresults strongly suggest that the NF-kB signal transduction ma-chinery post-TLR4 stimulation is intact in PARP-12/2 cells andthat the defect may reside in the actual translocation of the tran-scription factor to the nucleus rather than its association with IkBa.
Accumulating evidence indicates that after the release of NF-kBfrom inhibition by IkBa, the phosphorylation of the p65 subunitat Ser276, Ser529, and Ser536 is critical for its interaction with transcrip-tional coactivators such as CBP/p300 and for binding to its target siteson DNA (24). We and others have reported that interaction betweenPARP-1, p65 NF-kB, CBP/p300 takes place upon stimulation withTNF or LPS (11, 25). Given that the TLR4-associated signal trans-duction appeared to be intact in PARP-12/2 cells, while expression ofNF-kB–dependent genes was compromised, it became important to
determine whether PARP-1 plays a role in events leading to p65NF-kB phosphorylation. Fig. 2B shows that phosphorylation of p65NF-kB at Ser276 and Ser529 was evident in WT SMCs but almostcompletely absent in PARP-12/2 cells upon TLR4 stimulation byLPS. These results clearly suggest that PARP-1 expression is neces-sary for events leading to the phosphorylation of p65 NF-kB. It isunclear how PARP-1 influences these phosphorylation events.
Expression of importin a3 and importin a4 and their cytosoliclocalization are not altered in PARP-12/2 cells
A closer examination of p65 NF-kB localization in LPS-treatedPARP-12/2 cells revealed that the protein appeared to accumulatein the perinuclear area of a subpopulation (30–40%) of theexamined cells (Fig. 3A), which suggested a potential defect inthe nuclear import system in these cells. We surmised that thedefect in p65 NF-kB nuclear translocation in response to LPSmay be coupled to a defective importin system. First, weexamined whether PARP-1 gene deletion exerted any effect onexpression of importin a3 and importin a4. Fig. 3B shows thatthe expression levels of importin a3 and importin a4 were notaffected by PARP-1 gene deletion in SMCs. Given that interactionbetween p65 NF-kB and importins takes place in the cytosol afterrelease from IkBa upon stimulation, we examined whether thelevels of cytosolic importins were different between WT andPARP-12/2 cells upon stimulation with LPS. The levels ofcytosolic importin a3 (Fig. 3C) and importin a4 (Fig. 3D) werenot different between WT and PARP-12/2 cells, suggesting thatthe defect in p65 NF-kB nuclear translocation in LPS-treatedPARP-12/2 cells resides upstream of importins.
Inhibition of Crm1 by leptomycin B promotes accumulation ofp65 NF-kB in nuclei of PARP-12/2 cells and reversesexpression of NF-kB–dependent genes upon TLR4 stimulation
Because the importin system was not affected by PARP-1 gene de-letion, we reasoned that elevated export activity resulted in NF-kBnuclear accumulation upon TLR4 stimulation in PARP-12/2 cells.Accordingly, we tested the hypothesis that NF-kB was notdetected in the nucleus because it was rapidly exported byCrm1-associated export machinery. PARP-1 gene deletion didnot alter total Crm1 expression (Fig. 4Aa) but was associated withan increase in cytosolic Crm1 in control cells (Fig. 4Ab, Ac). Thisincrease in cytosolic Crm1 was verified by cell fractionation fol-lowed by immunoblot analysis (Fig. 4B). LPS treatment promoteda decrease in cytosolic Crm1 in WT cells. In sharp contrast, thelevels of cytosolic Crm1 were relatively unchanged in LPS-treatedPARP-12/2 cells (Fig. 4B). Interestingly, no major changes innuclear Crm1 were observed in WT and PARP-12/2 cells,
FIGURE 2. Effects of PARP-1 gene deletion on
phosphorylation and degradation of IkBa and phos-
phorylation of p65 NF-kB in SMCs upon LPS
treatment. WT or PARP-12/2 SMCs were treated
with 1 mg/ml LPS for different intervals or left
untreated (Con). A, Proteins extracts were subjected
to immunoblot analysis with Abs to IkBa or
phospho-IkBa at Ser32/Ser36. B, The same extracts
were subjected to immunoblot analysis with Abs to
phospho-p65 NF-kB at Ser276 or Ser529, total p65
NF-kB, or actin. Con, control.
4 REGULATION OF LPS-INDUCED NF-kB NUCLEAR RETENTION BY PARP-1
by guest on January 3, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

suggesting that the level of the translocated Crm1 to the cytosolwas minor compared with that in the nucleus. A quantitativeassessment of the data is presented in Fig. 4Bb. The persistenceof Crm1 in the cytosol of control and LPS-treated PARP-12/2
SMCs was verified by immunofluorescence (Fig. 4C, 4D). Theseresults suggest that PARP-1 may play a role in the dynamics ofCrm1 trafficking.The coincidence of Crm1 and p65 NF-kB in the cytosolic
compartments of LPS-treated PARP-12/2 cells is suggestive ofa potential link between PARP-1 gene deletion and the defect inNF-kB nuclear translocation. To further investigate the relationshipbetween PARP-1, p65 NF-kB, and Crm1 and to establish that theexport system is affected by PARP-1 gene deletion, we hypothe-sized that inhibition of Crm1 would allow p65 NF-kB nuclearaccumulation in LPS-treated PARP-12/2 cells. Indeed, Crm1inhibition allowed p65 NF-kB nuclear accumulation in PARP-12/2
cells without LPS treatment, similar to that observed in WT cells(Fig. 4E). LPS treatment appeared to increase the nuclear retentionof p65 NF-kB with almost a complete absence of the protein in thecytosolic compartment in PARP-12/2 cells (Fig. 4E). These resultssuggest that nuclear accumulation of p65 NF-kB in PARP-12/2
cells in response to TLR4 stimulation results from enhancedCrm1-associated export of the transcription factor from the nucleus.The nuclear retention of p65 NF-kB in leptomycin B (LMB)-
treated PARP-12/2 cells coincided with a robust increase in itsphosphorylation state at Ser276, Ser529, and Ser536 but only uponLPS treatment, suggesting a potential transcriptional activity (Fig.4F). An important question to be addressed was whether suchnuclear accumulation of p65 NF-kB and concomitant phosphory-lation at Ser536 in LPS- and LMB-treated PARP-12/2 cellsculminated in expression of NF-kB-driven genes such as iNOSor ICAM-1. Fig. 4G shows that, indeed, inhibition of Crm1 re-stored LPS-stimulated iNOS or ICAM-1 expression in PARP-12/2
cells. Collectively, these data strongly suggest that PARP-1 pro-motes the nuclear function of NF-kB by preventing its exportfrom the nucleus by Crm1.
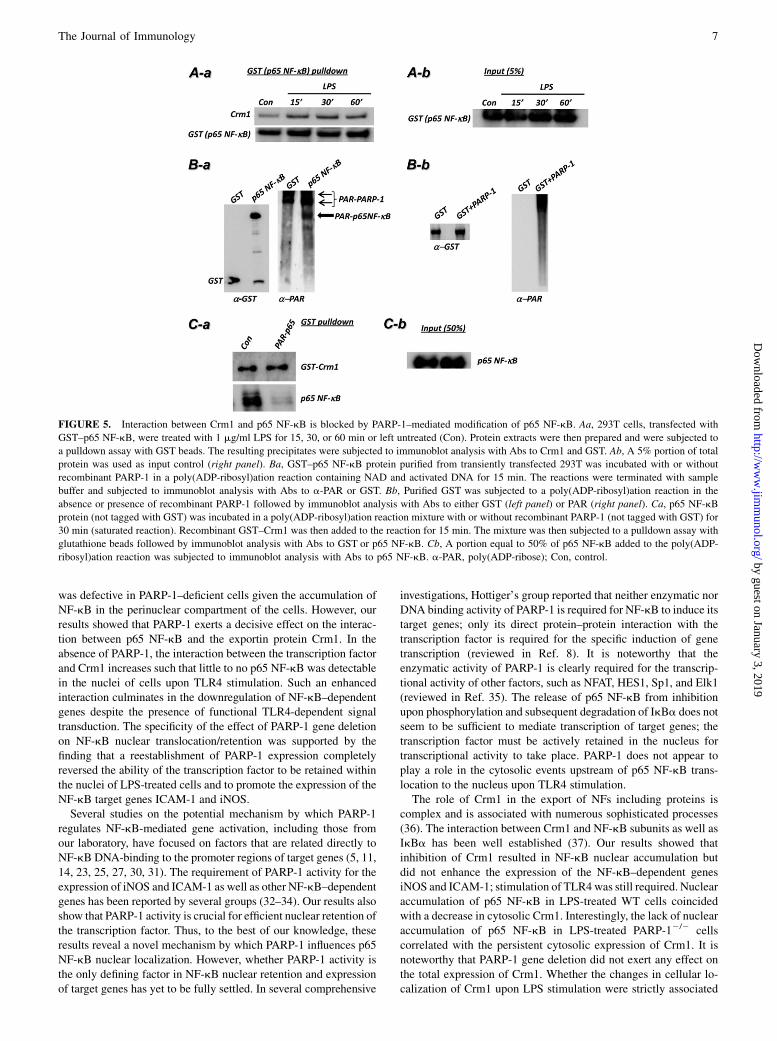
Poly(ADP-ribosyl)ation of p65 NF-kB by PARP-1 decreases itsinteraction with Crm1
We next investigated the potential mechanism by which PARP-1influences the relationship between Crm1 and p65 NF-kB. Fig.5A confirms the interaction between Crm1 and p65 NF-kB andthe dynamic nature of such an interaction upon LPS stimulationin 293T cells. Using an in vitro poly(ADP-ribosyl)ation system
(26), we showed that p65 NF-kB can be modified by PARP-1(Fig. 5B), confirming the report by Kameoka et al. (27). We nextexamined whether PARP-1–mediated modification of p65 NF-kBwould affect the interaction between the transcription factor andCrm1 in vitro. Fig. 5C shows that poly(ADP-ribosyl)ation of p65NF-kB almost completely abolished the interaction between thetwo proteins. These observations are consistent with the hypo-thesis that poly(ADP-ribosyl)ation of p65 NF-kB inhibits its as-sociation with Crm1 and hence attenuates nuclear export of thetranscription factor.
Pharmacologic inhibition of PARP-1 reduces p65 NF-kBnuclear retention and subsequent expression of ICAM-1 andiNOS in response to TLR4 stimulation, in part, by promotingpersistence of p65 NF-kB–Crm1 interaction
While the involvement of the PARP-1 enzymatic activity in the ex-pression of NF-kB–dependent genes and interaction with relatedtranscription factors remains controversial, its involvement inNF-kB nuclear translocation upon TLR4 stimulation is com-pletely unknown. Our data predict that inhibition of PARP1 enzy-matic activity should promote the p65 NF-kB–Crm1 interactionand thereby enhance Crm1-mediated nuclear export of the tran-scription factor. Fig. 6A shows that inhibition of PARP-1 by eithera noncompetitive (NU1025) or a competitive (3-aminobenzamide)inhibitor significantly reduced p65 NF-kB nuclear accumulationin LPS-treated SMCs. Such reduction in nuclear translocationcorrelated with a decrease in p65 NF-kB phosphorylation uponpharmacologic inhibition of PARP-1 (see Supplemental Fig. 2).In agreement with these findings, expression of iNOS and ICAM-1in LPS-stimulated SMCs was markedly reduced by the PARP-1inhibitors (Fig. 6B). Accordingly, these results demonstrate thatthe enzymatic activity of PARP-1 is required for a full nucleartranslocation of p65 NF-kB and subsequent expression of iNOSand ICAM-1. The requirement of PARP-1 enzymatic activity forNF-kB–dependent gene expression was verified by a luciferaseactivity assay using WT MEFs that were transiently transfectedwith the RapidReporter pRR-High-NF-kB plasmid (Fig. 6C).Whether the requirement for PARP-1 enzymatic activity appliesto all NF-kB stimuli and all cell types is not fully established.Inhibition of PARP-1 activity can also be achieved by reduc-
ing expression of PARG, the PAR-hydrolyzing enzyme, throughan increase in the population of automodified PARP-1 molecules(28, 29). Indeed, poly(ADP-ribosyl)ation activity was markedlyreduced in cells that were subjected to PARG knockdown with
FIGURE 3. Effect of PARP-1 gene deletion on expression and cytosolic localization of importins in SMCs upon LPS treatment. A, High magnification
image of nuclear and perinuclear localization of YFP–p65 NF-kB in LPS-treated WT and PARP-12/2 SMCs, respectively (original magnification 363).
B, Total protein extracts, prepared from WT or PARP-12/2 SMCs, were subjected to immunoblot analysis with Abs to importin a3, importin a4, or actin.
WT or PARP-12/2 SMCs were treated with 1 mg/ml LPS for 30 or 90 min or left untreated (Con), after which cytosolic fractions were prepared. The
resulting extracts were subjected to immunoblot analysis with Abs to importin a3 (C), importin a4 (D), or actin. Con, control.
The Journal of Immunology 5
by guest on January 3, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

a lentiviral vector encoding PARG-targeting shRNA (Supplementaldata 3A) upon treatment with the DNA damage-inducing agentH2O2 (Supplemental data 3B). The decreased in PARP-1 activityin PARG-depleted cell extracts was confirmed using an in vitropoly(ADP-ribosyl)ation assay (Supplemental data 3C). PARGknockdown significantly decreased p65 NF-kB retention in SMCsupon LPS treatment (Supplemental data 3D), which provides addi-tional support for the involvement of PARP-1 activity in the nuclearretention of the transcription factor.We next investigated the potential effect of pharmacologic in-
hibition of PARP-1 on the interaction between Crm1 and p65 NF-kB. Fig. 6D shows that the interaction of p65 NF-kB was persistentin cells treated with LPS in the presence of the PARP-1 inhibitorNU1025 compared with that observed in cells treated with LPS
alone. These results strongly suggest that PARP-1 enzymatic activ-ity is required for an efficient nuclear retention of p65 NF-kB uponTLR4 stimulation and confirm our in vitro data.Overall, these results strongly suggest that the potential key
regulatory mechanism of NF-kB nuclear retention may reside in themodification of p65 NF-kB by poly(ADP-ribosyl)ation, which pre-vents its interaction with Crm1, rendering the transcription factorresistant to export and promoting its retention in the nucleus.
DiscussionThemechanisms bywhich PARP-1, a nuclear protein, affects the ac-tivation of NF-kB in the cytosol and its nuclear translocation are notunderstood. The results of the present study shed light on thesemechanisms. Our initial prediction was that the importin system
FIGURE 4. Effect of PARP-1 gene deletion on expression and cytosolic localization of Crm1 and effects of Crm1 inhibition on p65 NF-kB nuclear
accumulation and expression of iNOS and ICAM-1 in LPS-treated SMCs. A, WT or PARP-12/2 SMCs were subjected to protein extracts followed by
immunoblot analysis with Abs to Crm1 or actin (Aa) or subjected to immunofluorescence with Abs to Crm1 (Ab). Ac represents ratios between nuclear and
cytosolic fluorescence of Crm1 in at least 10 cells and expressed as mean 6 SEM. *p , 0.01, difference from WT cells. B, The same cytosolic fractions
described in Fig. 3C were subjected to immunoblot analysis with Abs to Crm1 and actin (Ba) and quantified (Bb). C, WT or PARP-12/2 SMCs cultured on
chamber slides were treated with 1mg/ml LPS for 30 or 120min or left untreated (Con), after which cells were fixed and subjected to immunofluorescencewith
Abs to Crm1 (Alexa Fluor 488, green) and p65 NF-kB (Alexa Fluor 594, red) (original magnification 340). D, Crm1 cellular distribution was quantified as
described for p65 NF-kB in Fig. 1C. pp , 0.01, difference from respective untreated cells; #p , 0.01, difference from WT cells at the respective treatment
time. E, WT or PARP-12/2 SMCs were pretreated with LMB or left untreated, after which they were treated with 1 mg/ml LPS for the indicated intervals
(original magnification 340). Cells were then fixed and subjected to immunofluorescence with Abs to p65 NF-kB. Inhibition of Crm1 by LMB promotes
accumulation of p65 NF-kB in nuclei of PARP-12/2 cells and expression of iNOS and ICAM-1 in LPS-treated PARP-12/2 SMCs. WTor PARP-12/2 SMCs,
pretreated with LMB or left untreated, were exposed to 1 mg/ml LPS for different intervals or left untreated (Con). Proteins extracts were prepared and
subjected to immunoblot analysis with Abs to phospho-p65NF-kB at Ser276, Ser529, or Ser536, or actin (F).G, Cells were treated as in F, but for 3 h, after which
total RNAwas extracted and subjected to RT-PCR analysis with primers to iNOS, ICAM-1, and b-actin. Con, control.
6 REGULATION OF LPS-INDUCED NF-kB NUCLEAR RETENTION BY PARP-1
by guest on January 3, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
was defective in PARP-1–deficient cells given the accumulation ofNF-kB in the perinuclear compartment of the cells. However, ourresults showed that PARP-1 exerts a decisive effect on the interac-tion between p65 NF-kB and the exportin protein Crm1. In theabsence of PARP-1, the interaction between the transcription factorand Crm1 increases such that little to no p65 NF-kB was detectablein the nuclei of cells upon TLR4 stimulation. Such an enhancedinteraction culminates in the downregulation of NF-kB–dependentgenes despite the presence of functional TLR4-dependent signaltransduction. The specificity of the effect of PARP-1 gene deletionon NF-kB nuclear translocation/retention was supported by thefinding that a reestablishment of PARP-1 expression completelyreversed the ability of the transcription factor to be retained withinthe nuclei of LPS-treated cells and to promote the expression of theNF-kB target genes ICAM-1 and iNOS.Several studies on the potential mechanism by which PARP-1
regulates NF-kB-mediated gene activation, including those fromour laboratory, have focused on factors that are related directly toNF-kB DNA-binding to the promoter regions of target genes (5, 11,14, 23, 25, 27, 30, 31). The requirement of PARP-1 activity for theexpression of iNOS and ICAM-1 as well as other NF-kB–dependentgenes has been reported by several groups (32–34). Our results alsoshow that PARP-1 activity is crucial for efficient nuclear retention ofthe transcription factor. Thus, to the best of our knowledge, theseresults reveal a novel mechanism by which PARP-1 influences p65NF-kB nuclear localization. However, whether PARP-1 activity isthe only defining factor in NF-kB nuclear retention and expressionof target genes has yet to be fully settled. In several comprehensive
investigations, Hottiger’s group reported that neither enzymatic norDNA binding activity of PARP-1 is required for NF-kB to induce itstarget genes; only its direct protein–protein interaction with thetranscription factor is required for the specific induction of genetranscription (reviewed in Ref. 8). It is noteworthy that theenzymatic activity of PARP-1 is clearly required for the transcrip-tional activity of other factors, such as NFAT, HES1, Sp1, and Elk1(reviewed in Ref. 35). The release of p65 NF-kB from inhibitionupon phosphorylation and subsequent degradation of IkBa does notseem to be sufficient to mediate transcription of target genes; thetranscription factor must be actively retained in the nucleus fortranscriptional activity to take place. PARP-1 does not appear toplay a role in the cytosolic events upstream of p65 NF-kB trans-location to the nucleus upon TLR4 stimulation.The role of Crm1 in the export of NFs including proteins is
complex and is associated with numerous sophisticated processes(36). The interaction between Crm1 and NF-kB subunits as well asIkBa has been well established (37). Our results showed thatinhibition of Crm1 resulted in NF-kB nuclear accumulation butdid not enhance the expression of the NF-kB–dependent genesiNOS and ICAM-1; stimulation of TLR4was still required. Nuclearaccumulation of p65 NF-kB in LPS-treated WT cells coincidedwith a decrease in cytosolic Crm1. Interestingly, the lack of nuclearaccumulation of p65 NF-kB in LPS-treated PARP-12/2 cellscorrelated with the persistent cytosolic expression of Crm1. It isnoteworthy that PARP-1 gene deletion did not exert any effect onthe total expression of Crm1. Whether the changes in cellular lo-calization of Crm1 upon LPS stimulation were strictly associated
FIGURE 5. Interaction between Crm1 and p65 NF-kB is blocked by PARP-1–mediated modification of p65 NF-kB. Aa, 293T cells, transfected with
GST–p65 NF-kB, were treated with 1 mg/ml LPS for 15, 30, or 60 min or left untreated (Con). Protein extracts were then prepared and were subjected to
a pulldown assay with GST beads. The resulting precipitates were subjected to immunoblot analysis with Abs to Crm1 and GST. Ab, A 5% portion of total
protein was used as input control (right panel). Ba, GST–p65 NF-kB protein purified from transiently transfected 293T was incubated with or without
recombinant PARP-1 in a poly(ADP-ribosyl)ation reaction containing NAD and activated DNA for 15 min. The reactions were terminated with sample
buffer and subjected to immunoblot analysis with Abs to a-PAR or GST. Bb, Purified GST was subjected to a poly(ADP-ribosyl)ation reaction in the
absence or presence of recombinant PARP-1 followed by immunoblot analysis with Abs to either GST (left panel) or PAR (right panel). Ca, p65 NF-kB
protein (not tagged with GST) was incubated in a poly(ADP-ribosyl)ation reaction mixture with or without recombinant PARP-1 (not tagged with GST) for
30 min (saturated reaction). Recombinant GST–Crm1 was then added to the reaction for 15 min. The mixture was then subjected to a pulldown assay with
glutathione beads followed by immunoblot analysis with Abs to GST or p65 NF-kB. Cb, A portion equal to 50% of p65 NF-kB added to the poly(ADP-
ribosyl)ation reaction was subjected to immunoblot analysis with Abs to p65 NF-kB. a-PAR, poly(ADP-ribose); Con, control.
The Journal of Immunology 7
by guest on January 3, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

with p65 NF-kB translocation or were a general phenomenonrelated to the overall signal transduction associated with TLR4stimulation is not clear. However, it is plausible to predict that atleast a portion of the cytosolic Crm1 is responsible for the lack ofnuclear accumulation of p65 NF-kB in LPS-treated PARP-12/2
cells. This observation is supported by the finding that Crm1inhibition by LMB promoted the accumulation of nuclear p65NF-kB in LPS-treated PARP-12/2 cells. According to ourin vitro data, such a role involved PARP-1–mediated poly(ADP-ribosyl)ation of p65 NF-kB, which reduced the interaction of themodified protein with Crm1. This observation was confirmed inLPS-treated cells using NU1025, a potent inhibitor of PARP-1 en-zymatic activity. A recent report by Fukasawa’s group (38) inves-tigating Crm1 in the context of nuclear accumulation of p53supports the notion that this mechanism of regulation is not re-stricted to NF-kB alone.PARP-1appears toberequiredforthephosphorylationofp65NF-kB
at serine residues 276, 529, and 536. Although the mechanism bywhich PARP-1 influences p65 NF-kB phosphorylation is not clear,PARP-1 has been reported to interact with several kinases such asDNA-dependent protein kinase and ERK (39). Thus, it will be veryimportant to determine how p65NF-kBphosphorylation relates to therole of PARP-1–mediated regulation of this transcription factor andits interaction with Crm1. It is noteworthy that PARP-1 deficiency didnot seem to affect the phosphorylation of p65 NF-kB given the
observation that such event was reversed by LMB-mediated Crm1inhibition in LPS-treated PARP-12/2 cells. This may suggest thatthe phosphorylation was related to a defect in nuclear retentionrather thanPARP-1deficiency.What is clear and established, however,is that the phosphorylation of the transcription factor at these sites iscritical for the expression of NF-kB–dependent genes (40–43).Additionally, the expression levels of IkBa (balance betweenexpression and degradation) also may dictate the nature of theinteraction between PARP-1 and p65 NF-kB as well as with Crm1.Furthermore, the role of acetylation undoubtedly is very important andmay be a critical factor in the functionality of NF-kB and its ability todrive target gene expression. As stated above, we and others recentlyshowed an interaction between PARP-1 and p300/CBP that appearsto take place immediately after the interaction of PARP-1 with NF-kB (11, 25). The consequences of the sequence of these events are farfrom clear.An additional important aspect of NF-kB nuclear translocation is
the importin system (44). Although the data on the role ofimportins in p65 NF-kB shuttling between the cytosol and thenucleus presented in this study are not extensive, they do suggestthat there is no relationship between PARP-1 and these proteins asit relates to nuclear localization of p65 NF-kB, especially given thefact that PARP-1 gene deletion exerted no effect on total or cyto-solic levels of importin a3 and importin a4 upon LPS treatment.These observations are supported by the fact that Crm1 inhibition
FIGURE 6. Pharmacologic inhibition of PARP-1 reduces p65 NF-kB nuclear retention and subsequent expression of ICAM-1 and iNOS in response to
TLR4 stimulation, in part, by promoting persistence of p65 NF-kB–Crm1 interaction. A, PARP-1 inhibitors reduce LPS-induced p65 NF-kB nuclear
translocation. WT SMCs were treated with LPS in the presence or absence of the noncompetitive (NU1025) or the competitive (3-AB) PARP-1 inhibitor for
30 min, after which p65 NF-kB nuclear localization was examined by immunofluorescence (Aa; original magnification 320) and quantified (Ab) as
described in Fig. 1C. pp , 0.01, difference from untreated cells; #p , 0.01, difference from LPS-treated cells. B, WT SMCs were treated as in A, but for 2
or 4 h, after which total RNAwas prepared and subjected to RT-PCR using primers specific to mouse iNOS, ICAM-1, or b-actin with amplicons analyzed
by agarose electrophoresis. C, WT MEFs transiently transfected with the RapidReporter pRR-High-NF-kB plasmid were treated with LPS for 4 h in the
presence or absence of NU1025, after which luciferase activity was assessed. Data (luciferase activity) are expressed in relative light units and are means6SD of triplicate values from representative experiments. pp, 0.05, difference from untreated cells; #p, 0.05, difference from cells treated with LPS alone.
Da, 293T cells, transfected with GST–p65 NF-kB, were treated with LPS for 15 or 30 min or left untreated (Con) in the absence or presence of NU1025.
Protein extracts were then prepared and were subjected to a pulldown assay with GST beads. The resulting precipitates were subjected to immunoblot
analysis with Abs to Crm1 and GST. Db, A 5% portion of total protein was used as input control. Con, control.
8 REGULATION OF LPS-INDUCED NF-kB NUCLEAR RETENTION BY PARP-1
by guest on January 3, 2019http://w
ww
.jimm
unol.org/D
ownloaded from

by LMB leads to accumulation of p65 NF-kB within the nucleus,which again suggests that the importin system may be intact;however, this remains to be verified. Furthermore, it was recentlyshown that the nucleoporin Nup214 modulates NF-kB activation(in Drosophila) and influences the relative strength and duration ofNF-kB signaling responses (45). Again, whether Nup214 influ-ences the relationship between PARP-1, NF-kB, and Crm1remains to be studied. In conclusion, the results of the presentstudy provide novel insights into the mechanism by whichPARP-1 influences activation of TLR4-associated signal transduc-tion leading to NF-kB activation and subsequent nuclear retention.
AcknowledgmentsWe thank Ji-Won Park and Waleed Elsegeiny for technical assistance.
DisclosuresThe authors have no financial conflicts of interest.
References1. Burkle, A. 2001. Physiology and pathophysiology of poly(ADP-ribosyl)ation.
Bioessays 23: 795–806.2. Ha, H. C., and S. H. Snyder. 1999. Poly(ADP-ribose) polymerase is a mediator
of necrotic cell death by ATP depletion. Proc. Natl. Acad. Sci. USA 96: 13978–13982.
3. Chiarugi, A., E. Meli, M. Calvani, R. Picca, R. Baronti, E. Camaioni,G. Costantino, M. Marinozzi, D. E. Pellegrini-Giampietro, R. Pellicciari, et al.2003. Novel isoquinolinone-derived inhibitors of poly(ADP-ribose) polymerase-1:pharmacological characterization and neuroprotective effects in an in vitro modelof cerebral ischemia. J. Pharmacol. Exp. Ther. 305: 943–949.
4. Nicoletti, V. G., and A. M. Stella. 2003. Role of PARP under stress conditions:cell death or protection? Neurochem. Res. 28: 187–194.
5. Boulares, A. H., A. J. Zoltoski, Z. A. Sherif, P. Jolly, D. Massaro, andM. E. Smulson. 2003. Gene knockout or pharmacological inhibition of poly(ADP-ribose) polymerase-1 prevents lung inflammation in a murine model ofasthma. Am. J. Respir. Cell Mol. Biol. 28: 322–329.
6. Boulares, H., A. Zoltoski, S. Kandan, T. Akbulut, A. Yakovlev, andM. Oumouna. 2006. Correlation between decreased sensitivity of the Daudilymphoma cells to VP-16-induced apoptosis and deficiency in DNAS1L3 ex-pression. Biochem. Biophys. Res. Commun. 341: 653–662.
7. Oumouna-Benachour, K., C. P. Hans, Y. Suzuki, A. Naura, R. Datta,S. Belmadani, K. Fallon, C. Woods, and A. H. Boulares. 2007. Poly(ADP-ribose) polymerase inhibition reduces atherosclerotic plaque size and promotesfactors of plaque stability in apolipoprotein E-deficient mice: effects on macro-phage recruitment, nuclear factor-kB nuclear translocation, and foam cell death.Circulation 115: 2442–2450.
8. Hassa, P. O., and M. O. Hottiger. 2002. The functional role of poly(ADP-ribose)polymerase 1 as novel coactivator of NF-kB in inflammatory disorders. Cell.Mol. Life Sci. 59: 1534–1553.
9. Karin, M., Y. Cao, F. R. Greten, and Z. W. Li. 2002. NF-kB in cancer: frominnocent bystander to major culprit. Nat. Rev. Cancer 2: 301–310.
10. Yamamoto, Y., and R. B. Gaynor. 2001. Role of the NF-kB pathway in thepathogenesis of human disease states. Curr. Mol. Med. 1: 287–296.
11. Zerfaoui, M., Y. Suzuki, A. S. Naura, C. P. Hans, C. Nichols, and A. H. Boulares.2008. Nuclear translocation of p65 NF-kB is sufficient for VCAM-1, but notICAM-1, expression in TNF-stimulated smooth muscle cells: differential re-quirement for PARP-1 expression and interaction. Cell. Signal. 20: 186–194.
12. Hans, C. P., Y. Feng, A. S. Naura, M. Zerfaoui, B. M. Rezk, H. Xia, A. D. Kaye,K. Matrougui, E. Lazartigues, and A. H. Boulares. 2009. Protective effects ofPARP-1 knockout on dyslipidemia-induced autonomic and vascular dysfunctionin ApoE mice: effects on eNOS and oxidative stress. PLoS ONE 4: e7430.
13. Oliver, F. J., J. Menissier-de Murcia, C. Nacci, P. Decker, R. Andriantsitohaina,S. Muller, G. de la Rubia, J. C. Stoclet, and G. de Murcia. 1999. Resistance toendotoxic shock as a consequence of defective NF-kB activation in poly(ADP-ribose) polymerase-1 deficient mice. EMBO J. 18: 4446–4454.
14. Ha, H. C. 2004. Defective transcription factor activation for proinflammatorygene expression in poly(ADP-ribose) polymerase 1-deficient glia. Proc. Natl.Acad. Sci. USA 101: 5087–5092.
15. Fagerlund, R., K. Melen, X. Cao, and I. Julkunen. 2008. NF-kB p52, RelB and c-Rel are transported into the nucleus via a subset of importin a molecules. Cell.Signal. 20: 1442–1451.
16. Harhaj, E. W., and S. C. Sun. 1999. Regulation of RelA subcellular localizationby a putative nuclear export signal and p50. Mol. Cell. Biol. 19: 7088–7095.
17. Kutay, U., and S. Guttinger. 2005. Leucine-rich nuclear-export signals: born tobe weak. Trends Cell Biol. 15: 121–124.
18. Boulares, A. H., A. J. Zoltoski, A. Yakovlev, M. Xu, and M. E. Smulson. 2001.Roles of DNA fragmentation factor and poly(ADP-ribose) polymerase in anamplification phase of tumor necrosis factor-induced apoptosis. J. Biol. Chem.276: 38185–38192.
19. Boulares, A. H., A. G. Yakovlev, V. Ivanova, B. A. Stoica, G. Wang, S. Iyer, andM. Smulson. 1999. Role of poly(ADP-ribose) polymerase (PARP) cleavage in
apoptosis: caspase 3-resistant PARP mutant increases rates of apoptosis in trans-fected cells. J. Biol. Chem. 274: 22932–22940.
20. Belmadani, S., M. Zerfaoui, H. A. Boulares, D. I. Palen, and K. Matrougui. 2008.Microvessel vascular smooth muscle cells contribute to collagen type I de-position through ERK1/2 MAP kinase, avb3-integrin, and TGF-b1 in responseto ANG II and high glucose. Am. J. Physiol. 295: H69–H76.
21. Boulares, H. A., C. Giardina, C. L. Navarro, E. A. Khairallah, and S. D. Cohen.1999. Modulation of serum growth factor signal transduction in Hepa 1–6 cellsby acetaminophen: an inhibition of c-myc expression, NF-kB activation, andRaf-1 kinase activity. Toxicol. Sci. 48: 264–274.
22. Naura, A. S., R. Datta, C. P. Hans, M. Zerfaoui, B. M. Rezk, Y. Errami,M. Oumouna, K. Matrougui, and A. H. Boulares. 2009. Reciprocal regulation ofiNOS and PARP-1 during allergen-induced eosinophilia. Eur. Respir. J. 33: 252–262.
23. Hassa, P. O., and M. O. Hottiger. 1999. A role of poly(ADP-ribose) polymerasein NF-kB transcriptional activation. Biol. Chem. 380: 953–959.
24. Li, Q., and I. M. Verma. 2002. NF-kB regulation in the immune system. Nat.Rev. Immunol. 2: 725–734.
25. Hassa, P. O., S. S. Haenni, C. Buerki, N. I. Meier, W. S. Lane, H. Owen,M. Gersbach, R. Imhof, and M. O. Hottiger. 2005. Acetylation of poly(ADP-ribose) polymerase-1 by p300/CREB-binding protein regulates coactivation ofNF-kB-dependent transcription. J. Biol. Chem. 280: 40450–40464.
26. Boulares, A. H., A. J. Zoltoski, F. J. Contreras, A. G. Yakovlev, K. Yoshihara,and M. E. Smulson. 2002. Regulation of DNAS1L3 endonuclease activity bypoly(ADP-ribosyl)ation during etoposide-induced apoptosis: role of poly(ADP-ribose) polymerase-1 cleavage in endonuclease activation. J. Biol. Chem. 277:372–378.
27. Kameoka, M., K. Ota, T. Tetsuka, Y. Tanaka, A. Itaya, T. Okamoto, andK. Yoshihara. 2000. Evidence for regulation of NF-kB by poly(ADP-ribose)polymerase. Biochem. J. 346: 641–649.
28. Rapizzi, E., S. Fossati, F. Moroni, and A. Chiarugi. 2004. Inhibition of poly(ADP-ribose) glycohydrolase by gallotannin selectively up-regulates expressionof proinflammatory genes. Mol. Pharmacol. 66: 890–898.
29. Erdelyi, K., A. Kiss, E. Bakondi, P. Bai, C. Szabo, P. Gergely, F. Erdodi, andL. Virag. 2005. Gallotannin inhibits the expression of chemokines and in-flammatory cytokines in A549 cells. Mol. Pharmacol. 68: 895–904.
30. Hassa, P. O., M. Covic, S. Hasan, R. Imhof, and M. O. Hottiger. 2001. Theenzymatic and DNA binding activity of PARP-1 are not required for NF-kBcoactivator function. J. Biol. Chem. 276: 45588–45597.
31. Kraus, W. L., and J. T. Lis. 2003. PARP goes transcription. Cell 113: 677–683.32. Le Page, C., J. Sanceau, J. C. Drapier, and J. Wietzerbin. 1998. Inhibitors of
ADP-ribosylation impair inducible nitric oxide synthase gene transcriptionthrough inhibition of NF-kB activation. Biochem. Biophys. Res. Commun. 243:451–457.
33. Chiarugi, A. 2002. Poly(ADP-ribose) polymerase: killer or conspirator? The“suicide hypothesis” revisited. Trends Pharmacol. Sci. 23: 122–129.
34. Nakajima, H., H. Nagaso, N. Kakui, M. Ishikawa, T. Hiranuma, and S. Hoshiko.2004. Critical role of the automodification of poly(ADP-ribose) polymerase-1 innuclear factor-kB-dependent gene expression in primary cultured mouse glialcells. J. Biol. Chem. 279: 42774–42786.
35. Kraus, W. L. 2008. Transcriptional control by PARP-1: chromatin modulation,enhancer-binding, coregulation, and insulation. Curr. Opin. Cell Biol. 20: 294–302.
36. Sorokin, A. V., E. R. Kim, and L. P. Ovchinnikov. 2007. Nucleocytoplasmictransport of proteins. Biochemistry (Mosc.) 72: 1439–1457.
37. Huang, T. T., N. Kudo, M. Yoshida, and S. Miyamoto. 2000. A nuclear exportsignal in the N-terminal regulatory domain of IkBa controls cytoplasmic local-ization of inactive NF-kB/IkBa complexes. Proc. Natl. Acad. Sci. USA 97:1014–1019.
38. Kanai, M., K. Hanashiro, S. H. Kim, S. Hanai, A. H. Boulares, M. Miwa, andK. Fukasawa. 2007. Inhibition of Crm1-p53 interaction and nuclear export ofp53 by poly(ADP-ribosyl)ation. Nat. Cell Biol. 9: 1175–1183.
39. Cohen-Armon, M., L. Visochek, D. Rozensal, A. Kalal, I. Geistrikh, R. Klein,S. Bendetz-Nezer, Z. Yao, and R. Seger. 2007. DNA-independentPARP-1 activation by phosphorylated ERK2 increases Elk1 activity: a link tohistone acetylation. Mol. Cell 25: 297–308.
40. Zhong, H., R. E. Voll, and S. Ghosh. 1998. Phosphorylation of NF-kB p65 byPKA stimulates transcriptional activity by promoting a novel bivalent interactionwith the coactivator CBP/p300. Mol. Cell 1: 661–671.
41. Yang, F., E. Tang, K. Guan, and C. Y. Wang. 2003. IKKb plays an essential rolein the phosphorylation of RelA/p65 on serine 536 induced by lipopolysaccha-ride. J. Immunol. 170: 5630–5635.
42. Seldon, M. P., G. Silva, N. Pejanovic, R. Larsen, I. P. Gregoire, J. Filipe,J. Anrather, and M. P. Soares. 2007. Heme oxygenase-1 inhibits the expressionof adhesion molecules associated with endothelial cell activation via inhibitionof NF-kB RelA phosphorylation at serine 276. J. Immunol. 179: 7840–7851.
43. Chen, L. Y., W. W. Pan, M. Chen, J. D. Li, W. Liu, G. Chen, S. Huang,T. J. Papadimos, and Z. K. Pan. 2009. Synergistic induction of inflammation bybacterial products lipopolysaccharide and fMLP: an important microbial path-ogenic mechanism. J. Immunol. 182: 2518–2524.
44. Fagerlund, R., L. Kinnunen, M. Kohler, I. Julkunen, and K. Melen. 2005. NF-kBis transported into the nucleus by importin a3 and importin a4. J. Biol. Chem.280: 15942–15951.
45. Xylourgidis, N., P. Roth, N. Sabri, V. Tsarouhas, and C. Samakovlis. 2006. Thenucleoporin Nup214 sequesters CRM1 at the nuclear rim and modulates NFkBactivation in Drosophila. J. Cell Sci. 119: 4409–4419.
The Journal of Immunology 9
by guest on January 3, 2019http://w
ww
.jimm
unol.org/D
ownloaded from
![Untersuchungen zum Wirkmechanismus von 6-Amino-11,12 ... · PARP Poly [ADP-ribose] polymerase PBGD Porphobilinogen deaminase PBS Phosphate buffered saline PCR Polymerase chain reaction](https://img.pdfslide.us/doc/110x75/5d5cbcc088c9939b368b7c27/untersuchungen-zum-wirkmechanismus-von-6-amino-1112-parp-poly-adp-ribose.jpg)













![Novel therapies are changing treatment paradigms in ... · Polyadenosine diphosphate [ADP]-ribose polymerase (PARP) is a nuclear enzyme that aids the repair of single-strand DNA breaks](https://img.pdfslide.us/doc/110x75/60f47096160be920b7480ca6/novel-therapies-are-changing-treatment-paradigms-in-polyadenosine-diphosphate.jpg)














