Embed Size (px)
Citation preview

Plate tectonics inbiogeography
Steve TrewickMassey University, New Zealand
Plate tectonics deals with the motions of theEarth’s rocky shell, or lithosphere, which causethe gradual shift of continents and are alsoresponsible for mountain building, volcanicislands, and thermal vents. These geologicalprocesses have influenced the distribution ofbiodiversity. Prior to the identification of ageophysical mechanism that could drive move-ment at the Earth’s surface, its effects had beenpredicted. The concept of “continental drift”is usually attributed to Alfred Wegener, whopresented his ideas on the origin of continentsand oceans in 1915. However, in the 1500s theBelgian mapmaker Abraham Ortelius had notedthat the coincidence of the coastal outlines of theAmericas, Africa, and Europe implied formerjuxtaposition. In the 1800s, the “father of bio-geography,” Alfred Russell Wallace, developedhis ideas with the benefit of geographer CharlesLyell’s Principles of Geography. Lyell had notedthat “Continents … shift their positions entirelyin the course of ages.” Nevertheless, in the1900s continental drift was strongly opposed bymany, and the rejection of this hypothesis, whichwas built on extensive evidence, represents anotable failure of twentieth-century scientificmethod. The claim that consideration of thetheory required demonstration of a geophysicalmechanism was unjustified, but was neverthelessassuaged in the 1950s and 1960s when a tectonic
The International Encyclopedia of Geography.Edited by Douglas Richardson, Noel Castree, Michael F. Goodchild, Audrey Kobayashi, Weidong Liu, and Richard A. Marston.© 2017 John Wiley & Sons, Ltd. Published 2017 by John Wiley & Sons, Ltd.DOI: 10.1002/9781118786352.wbieg0638
model was presented. Today, paleomagnetism,radiometric dating, stratigraphy, and spatialinformation allow detailed interpretation andpaleogeographic reconstruction.
One type of evidence presented in supportof continental drift by Wegener and others was(paleo)biogeographic: the widespread distribu-tion of certain fossil animals and plants acrosslandscapes that are now disconnected (Figure 1).However, although widely reiterated even today,the occurrence of fossils did not in truth informon the process of continental isolation. An alter-native idea (land bridging) resolves the spatialdiscontinuity of biota problem equally well andmakes the same biological assumptions. As withcontinental drift, land bridging assumes that theorganisms of interest were unlikely to dispersebetween patches of suitable habitat. Thus bothhypotheses require habitat continuity if species(or their lineages) are to be shared among areas.In biogeography, distribution patterns are oftentaken as evidence of past processes but theyprovide only the basis for formulating alterna-tive hypotheses about the processes that mightexplain them. This is a subtle but vital distinctionthat underpins scientific method; a propositioncannot be simultaneously proposed and testedusing the same observations.
In later editions of his book, Wegener recog-nized that his continental drift hypothesis wasnot to be tested with fossils, or for that matter acoincidence of continental crust outline. Rather,the best approach to testing the hypothesis wasgeodetics (Earth measurement), because thehypothesis that continents had moved in the pastgenerated the prediction that continents wouldstill be moving. Wegener died in his tent duringa trip to Greenland to confirm longitudinal drift,

PLATE TECTONICS IN BIOGEOGRAPHY
Lystrosaurus
Cynognathus
Mesosaurus
Glossopteris
AfricaIndia
AustraliaAntartica
South America
Lystrosaurus
GlossopterisMesosaurus
Cynognathus
Figure 1 Distribution of four Permian and Triassic fossil groups used as biogeographic evidence for continentaldrift, and land bridging. Mesosaurus included a small number of meter-long marine reptile species recordedin early Permian time (299–280 Ma). It is said that Mesosaurus could not have crossed the Atlantic but this isspeculative, and at least one popular web resource incorrectly states that these were freshwater reptiles. Lystrosaurusincluded a debated number of species of pig-like land reptiles less than 1 m long. The genus is unusual in beingpresent on either side of the Permian-Triassic extinction boundary (255–241 Ma). Cynognathus (C. crateronotus)was a meter-long predatory mammal-like reptile usually treated as one species that existed in early to mid-Triassictime (247–237 Ma). Glossopteris was a group of woody gymnosperm plants with wide but mostly southern (i.e.,Gondwanan) distribution in the Permian (290–252 Ma). The identification of Glossopteris from northern areashas been questioned, and estimates of diversity vary.
Fossil sites were mapped using the database and tools of the Fossilworks Paleobiology Database, MacquarieUniversity ( John Alroy). Paleogeographic reconstruction is for Permian-Triassic time (250 mya). The inset on leftis a version of a popular iconographic but misleading depiction of the fossil distributions (The Snider-PellegriniWegener fossil map; WikiCommons). Permian Map by C.R. Scotese, PALEOMAP Project. Reproduced bypermission of C.R. Scotese.
but geodetic data have since demonstrated thefact that continents move. Plate tectonics explainshow. Geological (rather than biogeographic) datainforms paleogeographic reconstruction, and indoing so provides the context for biogeography.Plate tectonics is not, however, only aboutdrifting continents; it generates short- andlong-term environmental changes that influence
the distribution and evolution of biologicaldiversity, in many different ways.
Continental drift vicariance
Recognition of the potential for continental-scaleeffects of plate tectonics on biology developed
2

PLATE TECTONICS IN BIOGEOGRAPHY
side by side with plate tectonic theory. Thefounding concept of continental drift was thatmost of the Earth’s land had formerly been con-nected in a single supercontinent (Pangea). Fossilsof Permian age indicate that Pangea existed morethan 250 mya, and modern evidence supportsthis (see Figure 1). In fact the key fossils occurprimarily across the southern part of this conti-nent, a region usually referred to as Gondwana(or Gondwanaland), and it is noteworthy thatthis term was also used by land-bridgers who alsoenvisaged a former extensive continent, partsof which subsequently disappeared. Althoughthe geophysical evidence for a supercontinentis very clear, we now know that it did notinclude all continental areas at any one time.The idea that past breakup of a supercontinent(up to 200 million years later) could result inthe establishment of biotas that are visible todaymakes many assumptions, among them thata supercontinent would have a homogeneousenvironment with continuous distributions ofplants and animals, that other global events didnot cause local changes in biotic assemblages,and that dispersal between continents had littleinfluence on biological assemblages. In fact,paleoecological evidence indicates diverse andsometimes extreme environmental conditionsexisted across Pangea. Continental drift itselfresulted in changes in climatic conditions so thatnatural selection on the biota would change, anextreme case being the wholesale extinction ofAntarctic biota due to the drift of the continentover the South Pole. Mass extinctions acrossthe globe radically altered biotas and dispersalbetween continents influenced regional bioticcomposition through time.
Intriguingly, although vicariance biogeogra-phy associated with continental drift is a highlyattractive and popular idea, contrary evidenceof the power of biota to disperse and colonizeacross habitat discontinuities is abundant and well
understood (e.g., colonization and speciation onoceanic islands such as the Galapagos). In studiesof living biota, evidence for plate tectonic vicari-ance can come from dated phylogenetic analysis(see Phylogeography and landscape genetics).Appropriate sampling and fossil calibration (seeFigure 2) allow the timing of evolutionary eventsand continental drift to be compared. A matchbetween the estimated time of origins of regionalbiota and the time of continental separation isconsistent with, but not proof of, vicariance.
Land bridges and sea barriers
Plate tectonics can result in the connection offormerly disjunct land areas and simultaneouslythe sundering of marine environments, throughvolcanics, accretion, orogenics, and deformation.Although land bridging has been largely rejectedfrom consideration of ancient biological historyof Earth, younger examples show its influence.The Central American (or Panama) isthmusbetween North America and South America is anarrow strip of land (60–177 m wide) that finallyshut off the equatorial link between Atlantic andPacific oceans (the Central American seaway)in the late Pliocene (about 3 million years ago).Several tectonic plates intersect in the region(Figure 3) and their interaction appears to haveled to the formation of numerous volcanic islandsstarting at about 5.5 mya. Sediment accumu-lation between these islands is one mechanismproposed for the land bridge.
Closure forced the Gulf Stream to carry warmequatorial water into the northern AtlanticOcean, where it influences the compositionand ecology of regional marine biota. Separa-tion was accompanied by other environmentalchange; upwelling on the Pacific side see msto have increased while salinity has increasedin the Caribbean. Around Central America,
3
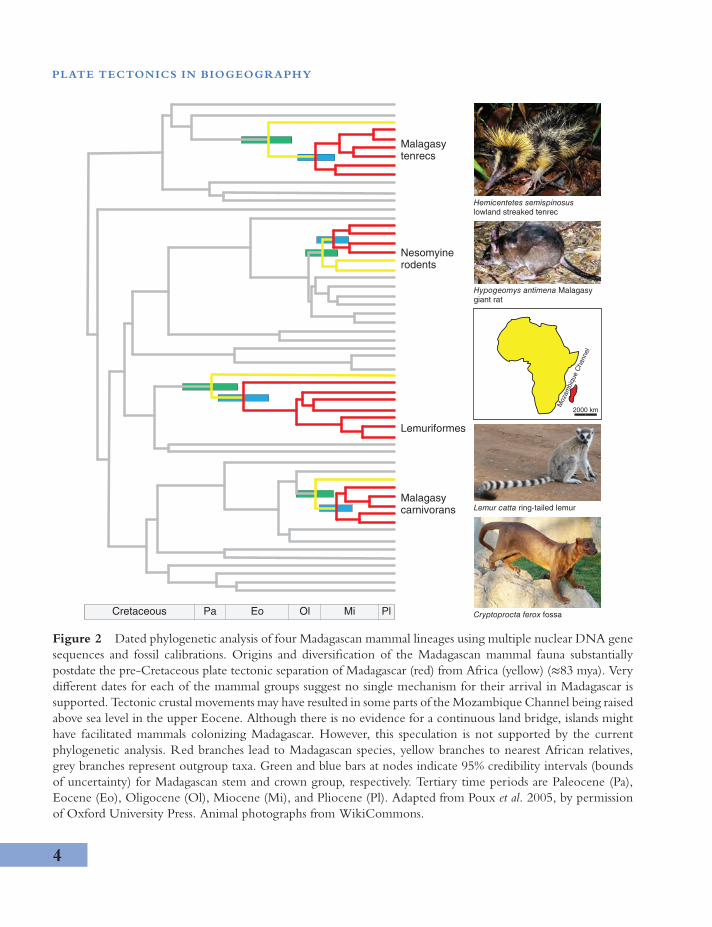
PLATE TECTONICS IN BIOGEOGRAPHY
Malagasytenrecs
Nesomyinerodents
Lemuriformes
Malagasycarnivorans
Cryptoprocta ferox fossa
Lemur catta ring-tailed lemur
Hypogeomys antimena Malagasygiant rat
Hemicentetes semispinosuslowland streaked tenrec
2000 km
Moza
mbiq
ue C
hannel
Cretaceous Pa Eo Ol Mi Pl
Figure 2 Dated phylogenetic analysis of four Madagascan mammal lineages using multiple nuclear DNA genesequences and fossil calibrations. Origins and diversification of the Madagascan mammal fauna substantiallypostdate the pre-Cretaceous plate tectonic separation of Madagascar (red) from Africa (yellow) (≈83 mya). Verydifferent dates for each of the mammal groups suggest no single mechanism for their arrival in Madagascar issupported. Tectonic crustal movements may have resulted in some parts of the Mozambique Channel being raisedabove sea level in the upper Eocene. Although there is no evidence for a continuous land bridge, islands mighthave facilitated mammals colonizing Madagascar. However, this speculation is not supported by the currentphylogenetic analysis. Red branches lead to Madagascan species, yellow branches to nearest African relatives,grey branches represent outgroup taxa. Green and blue bars at nodes indicate 95% credibility intervals (boundsof uncertainty) for Madagascan stem and crown group, respectively. Tertiary time periods are Paleocene (Pa),Eocene (Eo), Oligocene (Ol), Miocene (Mi), and Pliocene (Pl). Adapted from Poux et al. 2005, by permissionof Oxford University Press. Animal photographs from WikiCommons.
4

PLATE TECTONICS IN BIOGEOGRAPHY
populations of marine creatures (and evenseabirds that are reluctant to fly over land)were split between western Atlantic and easternPacific communities. On land, the isthmus ledto changes in rainfall and climate patterns andfacilitated the exchange of land creatures (a.k.a.the Great American Biotic Interchange). Manyanimals are inferred to have moved betweencontinents and contributed to biotic mixing andthus elevated biodiversity in Central America.
The late Pliocene formation of the PanamaIsthmus demonstrates a way that plate tectonicsinfluences biogeography, which is counter tothe effects of continental drift vicariance. Thisgeologically, ecologically, and phylogeneticallywell-studied phenomenon is precisely the typeof land bridge proposed by Charles Schuchertand others to explain exchange of biota betweencontinents in past times. Their ideas replaced ear-lier, simplistic land bridge models that requiredspecial, unknown, processes to destroy large partsof continents. Systems similar to the PanamaIsthmus might be geologically short-lived andthus hard to detect after the passage of time.Clearly, however, small and short-lived geophys-ical phenomena can have a profound influenceon biogeography.
Intriguingly, while many mammals appearto have walked between continents once thePanama Isthmus formed, molecular and fossildata indicate that habitat connectivity is notessential for exchange. For example, the best
explanation for the initial presence in SouthAmerica of several mammal lineages involvesoversea dispersal from Africa. This includessome that were subsequent North Americancolonists (e.g., porcupine). The marsupials, forinstance, are represented by just one species inNorth America, about 100 in South America,and about 230 in Australia. However, the oldestfossils (65 mya) are from North America. Expan-sion southwards through South America andAntarctica probably took marsupials to Australiaand eventually Borneo and Sulawesi, while thegroup disappeared from North America. Geneticanalysis suggests that all living marsupials havetheir ancestors in South America. This makessense because the only living marsupial in NorthAmerica (opossum) arrived over the PanamaIsthmus. Little is known about how these typesof animal interacted when they met, and conse-quently ad hoc interpretations are common. Forexample, the extinction of placental mammals inAustralia is often attributed to the arrival of mar-supials, yet the reverse (replacement of marsupialsby placentals) is suggested for North America.Very little is known about the marsupial faunathat must have existed in Antarctica, and none ofcourse are there today. Modern species distribu-tions are therefore revealed to be poor indicators,in many cases, of biogeographic history.
Other land bridges have resulted not fromtectonic activity but from sea level change. Mostrecently, during the Pleistocene (<2.6 mya),
−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−→Figure 3 The Panama Isthmus formed about 3 million years ago. Ocean currents (grey arrows) were altered.Range extension of various mammals is attributed to land bridging, from south to north (examples in blue) andnorth to south (examples in brown). Several of these have subsequently become extinct (†). Caribbean (pinkspots) and Pacific populations (yellow spots) of marine organisms were separated from one another. Red lines andletters refer to tectonic plates that intersect around the Panama Isthmus (NA, North American; CO, Cocoas; CA,Caribbean; NZ, Nazca; ND, North Andes; SA, South America). To the southwest is the Galapagos archipelago,with its famous biodiversity that stimulated Charles Darwin’s ideas about evolution. These volcanic islands grewfrom beneath the ocean as a result of plate tectonic activity, and the founding biota must have arrived by dispersalover the sea.
5
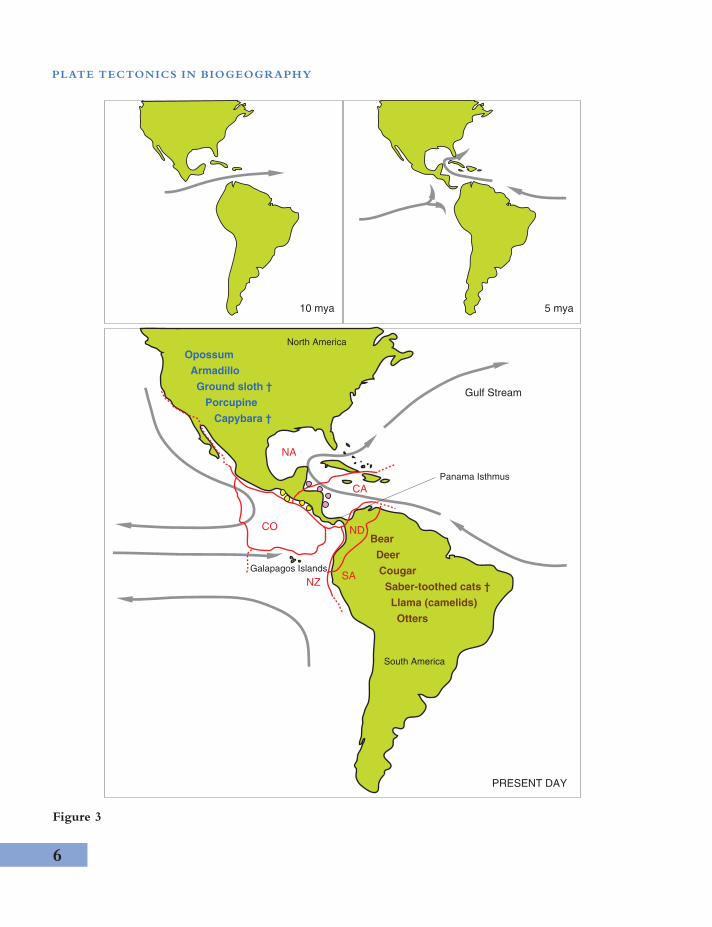
PLATE TECTONICS IN BIOGEOGRAPHY
Gulf Stream
PRESENT DAY
10 mya 5 mya
Galapagos Islands
North America
South America
Panama Isthmus
CO
NZ
CA
ND
NA
SA
Opossum
Armadillo
Ground sloth †
Porcupine
Capybara †
Bear
Deer
Cougar
Saber-toothed cats †
Llama (camelids)
Otters
Figure 3
6

PLATE TECTONICS IN BIOGEOGRAPHY
lowered sea level resulting from water beinglocked into polar ice caps created land con-nection in many parts of the world. Habitablecontinents were linked as recently as 13 000 yearsago through features such as the Bering landbridge between Siberia and Alaska that enabledthe migration of many plants and animals,including humans.
Mountains
Linear mountain ranges form when continentalplates collide, and it is routinely suggested that theformation of mountain ranges is a likely driverof allopatric speciation in terrestrial ecosystems.Populations sundered by geophysical barrierssuch as mountain ranges could evolve indepen-dently of one another if gene flow between themwas sufficiently impeded. Mountain formationprovides an appealing scenario for terrestrialvicariance because it generates abrupt environ-mental discontinuity (similar to land bridgesdividing marine environments). Abrupt changesin environmental conditions are expected toinfluence population density, range sizes, andnatural selection, but proven examples of theemergence of mountains causing speciationthrough vicariance are few. Cases where relatedspecies occupy habitat either side of moun-tains provide the expected distribution patternsbut might also have resulted from occupationof habitat patches after mountains formed.They might also be the remnants of speciesdistributions that predate mountain formation.
Mountain building contributes to biogeogra-phy in another way. The steep environmentalgradients and habitat mosaics that mountainranges produce appear to have stimulated adap-tive radiations in many taxa. Examples includealpine buttercups in New Zealand, paper daisiesin South Africa, and Andean hummingbirds inSouth America. Species that evolve to use alpine
conditions in one location may also be successfulon other ranges where similar ecological oppor-tunities exist even though these are island-like,and this can result in disjunct distributions thatare suggestive of vicariance.
Numerous studies in Europe and elsewhereshow that plants and animals responded to Pleis-tocene climate cycling by range shifting, and thisis contrary to the idea that mountains (and othergeophysical features) routinely make biologicalbarriers. The distribution across Europe ofgenetic variation within species and speciesgroups shows that only in some cases do somemountain ranges correspond with species limits.In these cases, of course, the mountains arealready present but the climate zones and speciesmove. Distinguishing between mountains (andsimilar features) as drivers of lineage splitting andtheir longer-term influence on environmentalgradients and ecosystem processes is challengingand exposes a general problem for biogeography.Mountains may come to be correlated with thelimits of species and assemblage ranges or theymight provide convenient landscape markersfor biogeographers, even if they were not themechanism of population sundering in the firstplace. Instead of looking for absolute interac-tions between geology and biology that focus onallopatric processes of lineage splitting, modernbiogeography now recognizes the ways geneflow, environmental gradients, environmentalfluctuations (e.g., climate change), extinction,sympatry, and uneven selection pressures acrossspecies ranges influence biotic composition.
Deep sea hydrothermal vents
Mineral-rich hot water emerges at temperaturesof 400 ○C or more in submarine conditionswhere tectonic plates meet. The heat andchemical resources fuel specialized faunas
7

PLATE TECTONICS IN BIOGEOGRAPHY
that constitute the only major biological sys-tems that do not draw their energy from thesun. Secondary production from Archaea andchemosynthetic bacteria supports faunas thatinclude giant worms, shrimps, clams, andlimpets, at depths of 1–4 km below the sea’ssurface. The habitat of hydrothermal vent faunasis dictated almost entirely by tectonic processesthat result in a patchy distribution around theglobe. These islands have histories tens of mil-lions of years long but their biological isolationappears to be influenced by their chemistryand the ecology of their fauna. Gene flowbetween invertebrate populations living aroundwidely spaced geothermal vents is mediatedby planktonic larvae. Plate tectonic activitymay be implicated in the origins of life in theancient seas of Earth as it has been proposedthat the thermal, chemical (e.g., acidity), andphysical (e.g., high pressure) conditions aroundhydrothermal vents could have supplied theenvironment suitable for initial emergence ofreplicating organic molecules and protocells.There is evidence for hydrothermal vents in theEarth’s oceans in the appropriate time framemore than 3 billion years ago.
Volcanoes
Oceanic volcanoes create virgin land that isphysically separate from existing, inhabited landareas. Although separated from other pieces ofland, islands are not isolated. Island biogeogra-phers recognize that the fauna and flora of islandsdevelops through the interaction of colonization,speciation, and extinction and these componentsare in some cases quantifiable. There is someevidence that the rate of colonization is influ-enced by the size and distance of islands frompotential biological sources, but increasingly itis apparent that establishment of migrant species
is strongly influenced by the composition of thebiota that has already arrived.
Classically, Charles Darwin’s observations oflife on the Galapagos archipelago were instru-mental in the founding of evolutionary theory(Figure 3). Volcanic island systems of this sortprovide natural laboratories where the effects ofcolonization, natural selection, and populationisolation can be observed. Because volcanic rocksare readily aged using radiometric analysis, max-imum ages for the local biota can be determined.
Volcanoes both destroy life and create opportu-nities, as has been demonstrated on a local, recentscale in systems such as Krakatau in Indonesia.On a much more profound scale, volcanic activ-ity is implicated in one or more ancient globalmass extinctions. Extreme volcanic activityassociated with the Central Atlantic Mag-matic Province is strongly indicated as drivingend-Triassic (201 mya) extinctions. Flood basalteruptions at the end of the Permian (252 mya)are likely to have contributed to extinction viaincreased atmospheric carbon dioxide, climatewarming, and associated ice melt methane pro-duction. The end of the Permian was markedby the extinction of about 70% of land animalspecies and 90% of marine animals. Amongthe casualties were the formerly widespreadGlossopertid plants, the fossils of which wereso influential in paleogeography. The Tri-assic saw the emergence of new ecologicalstrategies in mammal and dinosaur diversifi-cation. Ammonites emerged in the oceans,and crocodiles, pterosaurs, frogs, and sphen-odonts diversified on land.
Conclusion
On large and small scales, short and long timeframes, plate tectonics influences the formationand distribution of biological diversity; however,
8

PLATE TECTONICS IN BIOGEOGRAPHY
many of the details of how this happens remainto be thoroughly tested. A traditional focus onspatial effects (vicariance) is gradually beingreplaced by more in-depth analysis of the waygeophysical attributes of the planet influenceevolutionary ecology.
SEE ALSO: Biodiversity; Biogeography:history; Mountain biogeography; Oceanbiogeography; Zoogeography
Reference
Poux, C., O. Madsen, E. Marquard, et al. 2005.“Asynchronous Colonization of Madagascar by theFour Endemic Clades of Primates, Tenrecs, Car-
nivores, and Rodents as Inferred from NuclearGenes.” Systematic Biology, 54(5): 719–730.
Further reading
Lyell, C. 1872. Principles of Geology, 12th edn. London:John Murray.
Oreskes, N. 1999. The Rejection of Continental Drift:Theory and Method in American Earth Science. NewYork: Oxford University Press.
Schuchert, C. 1932. “Gondwana Land Bridges.” Bul-letin of the Geological Society of America, 43: 875–916.
Wallace, A.R. 1876. The Geographic Distribution of Ani-mals. London: MacMillan.
Wegener, A. 1968. The Origin of Continents and Ocean(trans. John Biram), 4th edn. London: Methuen.
9





























