Embed Size (px)
Citation preview

PLATE TETONICS AND SHALLOW MARINEDIVERSITY AND ENDEMISM, AN ACTUALISTIC MODEL
JAMES W. VALENTINE
AbstractValentine, James W. (Dept. Geology, Univ. Calif., Davis 95616) 1971. Plate tectonics
and shallow marine diversity and endemism, an actualistic model. Syst. Tool., 20:253-264.—The present shallow marine biogeographic pattern is geologically transitory and reflectsin large part the present state of the geography of continents and of lithospheric platemargins. Provincial boundaries are formed by thermal barriers and by land and deep-seabarriers, all of which are related to or controlled by tectonic patterns. Continental patterns,which greatly affect climate, reflect the patterns of plate tectonic processes. Deep-seabarriers are formed by ocean ridges and transform fault zones, while major oceanic dispersalroutes are formed by island arcs associated with subduction zones.
Patterns of species diversity can be related to tectonic as well as climatic patterns,presumably because the stability of shallow marine environments varies with the influenceof continentality, and this is under tectonic control. In particular it is the stability patternsof light and of nutrients (which directly affect the stability of trophic resource supplies)that correlate best with diversity patterns, and it is suggested that trophic resource stabilityis the fundamental regulator of species diversity within communities. Past changes incontinental relations and sizes, in oceanic sizes, and in the patterns and extent of island-arcchains have resulted from plate tectonic processes, and as these relations affect the trophicresource regime there have been corresponding alterations in patterns of both provincialityand diversity. [Species diversity, provinciality, biogeography.]
INTRODUCTION
The establishment of the mechanism ofsea-floor spreading (Vine and Mathews,1963; Vine, 1966) has led to the develop-ment of the theory of plate tectonics (Mc-Kenzie and Parker, 1967; Morgan, 1968;Le Pichon, 1968) and the demonstrationthat the geography of tectonic elements,including the pattern of continental dis-persion, has been radically different in thepast, and has probably been changing moreor less steadily throughout the Phanerozoic(Dewey and Horsfield, 1970; Valentine andMoores, 1970). Clearly, the climatic pat-terns associated with the continents havebeen different also. Indeed, world climatedepends to an extent upon continentalgeography and would vary with patternsof continental dispersion. Therefore thetwo most fundamental physical aspects ofbiogeography, climate and land-sea rela-tions, must have undergone significantchanges throughout much of geologic time,and these changes can be related to platetectonic processes. The geographic con-sequences of plate movements and associ-
ated processes, interacting with biologicalprocesses, must have led to the presentbiogeographic patterns.
This paper examines the present geog-raphy of continents, ocean ridges, subduc-tion zones and island arcs, fracture zones,and abyssal barriers, and relates them tomarine biogeographic patterns, especiallyto patterns of species diversity and pro-vinciality. Throughout this paper, the term'diversity' refers to the number of speciespresent in the region under discussion, anddoes not at all involve the size-frequencydistribution of their populations. Theserelationships form an actualistic model ofmarine biogeography which may serve asa tool in interpreting past patterns of bio-geography as well as past patterns of thegeography of tectonic elements.
PATTERNS OF ENDEMISM AND DISPERSAL:MARINE PROVINCES
At present, temperature is clearly thechief barrier to the latitudinal dispersal ofmarine shelf ectotherms (Bullock, 1955;Kinne, 1963). Probably this is chiefly due
253
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from

254 SYSTEMATIC ZOOLOGY
FIG. 1.—The major lithospheric plates of the earth and the character of their margins. Heavy lines areocean ridges which form spreading loci for the sea floor; most of them are active. Thin lines are trans-form faults which accommodate differential movement of plate segments or of entire plates (such asthe San Andreas fault system in California). Intermediate-weight lines are subduction zones, chieflydeep-sea trenches where crust is consumed; active zones are marked by triangles. After Le Pichon(1968) and Vine (1969).
to the temperature dependence of metabolicactivity rates, although in some cases itmay be due to the stability range of certaincompounds (such as lipids) or to othercauses. Furthermore, owing to the controlby water temperature of other water prop-erties, a great number of physiologicaleffects are correlated with temperature.Relatively few benthonic shelf speciesrange widely latitudinally, except in regionswhere climatic zones happen to be broadthemselves. Wherever the marine tempera-ture regime over the shelf changes mark-edly, it concentrates the range end-pointsof a relatively large number of species andforms a provincial boundary (Valentine,1966). Thus the primary latitudinal bio ticbarriers, which form provincial boundaries,are thermal.
Classically, the chief barriers to the longi-tudinal dispersal of marine shelf organismsare lands, as between the Caribbean andthe Gulf of Panama, and ocean deeps, as
between the eastern and western tropicalAtlantic (which might be called the mid-Atlantic Barrier) or between the rich Indo-Pacific biota and the American mainland—the East Pacific Barrier (Ekman, 1953).Both of these deep water barriers containa few isolated islands. Now, the EastPacific Barrier lies precisely over the EastPacific Rise, while the mid-Atlantic Barrieroverlies the mid-Atlantic Ridge. The richIndo-Pacific biota is developed principallyon islands and continental margins that areassociated with deep-sea trenches. Thusthere appears to be a tectonic basis for thebiogeographic patterns in these cases, whichprompts a closer look at the tectonic-bio-geographic relationship.
THE TECTONICS OF OCEAN BASINS ANDMARINE BIOGEOGRAPHY
In Fig. 1 some of the main tectonic fea-tures of the world's oceans are portrayed.The figure is generalized and simplified,
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from

TECTONICS: MARINE DIVERSITY AND ENDEMISM 255
but features that are important to bio-geography have been stressed. Accordingto the theory of plate tectonics, the plane-tary lithosphere is divided into a few largeplates, which are bounded at their marginsby either ridges (where crust is created),subduction zones (where crust is con-sumed), or fracture zones. Many platesbear one or more continents. Ocean ridgesare therefore a primary tectonic feature.Ridge crests, though rising well above thegeneral abyssal ocean floor, do not usuallyreach the ocean surface, although there areisolated island peaks such as the Azoresscattered along the mid-Atlantic and otherridges. In no case, however, are extensivechains of islands associated with ridges atpresent, but there is one large island—Iceland—that aids significantly in the dis-persal of marine biota. In general the ap-pearance and spreading of new oceaniccrust from ridge crests creates broad topo-graphic aprons gently inclined away fromthe crests and forming deep ocean floor.
The ridges and their spreading flanks arecut by numerous transverse fracture zones,both within plates and between them. Thesezones are transform faults, created by dif-ferential spreading of oceanic crust fromdifferent segments of the ridges and accom-modating the differential movement. Vol-canoes are generally not present along thefracture zones. As a result, the oceanridges together with their flanking crustalaprons generally form barriers, such as theEast Pacific Barrier, that block the dispersalof shallow-water organisms.
Subduction zones, which topographicallyare deep-sea trenches, also form primarytectonic features. If continents lie close tothem but opposite their consuming margins,such as where South America lies close tothe Peru-Chile Trench, mountain systemssuch as the Andes are formed by vulcanismrelated to crustal consumption. Whenthere are no continents closely associatedwith the plate margin formed by subductionzones, however, island arcs are formedopposite the consuming margin of the sub-duction zone.
The east-west chain of trenches, moun-tains and island arcs, which lies betweenBurma and the New Hebrides, borderscontinental plates to the northwest (Burmaand India) and in their south-central part(Australia-New Guinea). The subductionassociated with this chain chiefly accom-modates northward sea-floor spreading fromthe east-west trending Indian-Antarcticridge (Fig. 1; Le Pichon, 1968; Vine, 1969).These tropical islands and shelves supportthe richest of shallow marine biotas today.
Actually, the Indo-Pacific province ex-tends well beyond the range of the islandarcs, westward around the Indian shelf toeast Africa, eastward to the Tuamotus, andnorthward to the Gilbert, Marshall, andHawaiian Islands, and to numerous otherisland outliers. Presumably it extends west-ward simply because it abuts shelves therewhich provide a clear pathway for dispersalwithin a tropical climatic regime. Extensionof the Indo-Pacific province eastward andnorthward onto the island systems requiresspecial notice, for these islands cannot asyet be accounted for in terms of platetheory. Many of them are mid-plate islands,volcanic chains that rise from deep-oceanfloors but are not associated with platemargins nor with plate tectonics in anyobvious way, though they must move withthe crust in which they lie (see Wilson,1963, and Menard, 1969, for discussions).Others may have been generated at ridgesand have migrated laterally with spreadingcrust. If many islands of this sort happenedto occur east and north of the Tuamotus,forming easy dispersal routes leading to thetropical west American shelf, the EastPacific Rise would not form a barrier. Inany event, until their place in the schemeof global tectonics is more closely defined,the potentials of volcanoes as trans-ridgedispersal routes cannot be dismissed com-pletely.
LATITUDINAL DIVERSITY PATTERNS OFSHALLOW MARINE BIOTAS
The present configuration of the con-tinents creates long north-south trending
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from

256 SYSTEMATIC ZOOLOGY
stretches of essentially contiguous conti-nental shelves. The shelf biotas display awell-known latitudinal gradient in diversityfrom greater numbers of species at lowlatitudes to fewer at high latitudes (Fig.1). A number of explanations for thisgradient have been proposed (e.g., Fisher,1960; Pianka, 1966; Valentine, 1967; Sand-ers, 1968; Slobodkin and Sanders, 1969).A leading hypothesis is that the greater theenvironmental stability, the greater thenumber of species that may be accommo-dated, all other factors being equal (Fisher,1960; Hessler and Sanders, 1967; Sanders,1969).
The gradient in environmental stabilitythat most closely parallels the latitudinaldiversity gradient is in solar energy supply.As solar radiation is greatest in low latitudesand furthermore is received most evenlythroughout the year in low latitudes, andas seasonal differences increase polewarduntil day and night are seasonal, there is aresulting latitudinal gradient in the regimesof two major environmental factors, 1)temperature and 2) trophic resources.
Temperature has often been implicatedas a major cause of biotic diversity gradi-ents, but it now appears to be of secondaryimportance. Temperatures certainly varylatitudinally on the shelves and thereforecan be roughly correlated with diversitygradients. However, the discovery of highdiversity in the deep sea (Hessler andSanders, 1967) has demonstrated that lowtemperatures are not incompatible withhigh diversities, and in fact there aretheoretical reasons for supposing that lowtemperatures favor high diversity (Mar-galef, 1968). Another aspect of temperaturebesides its level is its variability or rangeand this has been suspected of influencingspecies diversity (Fisher, 1960). As sea-sonality is low in low latitudes, temperaturesthere have small annual ranges, whilemoving northward into mid-latitudes anincreasing temperature range is encount-ered. However in high latitudes the sea-sonal temperature range decreases, aswinter temperatures cannot fall below
freezing but summer temperatures are de-pressed. Thus, in general, there is a cor-relation between decreasing temperaturestability and decreasing diversity betweenlow- and mid-latitude shelves, but a cor-relation between increasing temperaturestability and decreasing diversity betweenmid- and high-latitude shelves. The cor-relation of thermal stability and diversityis thus seriously violated.
Turning to marine trophic resources, theyvary primarily with the availability of lightfor photosynthesis and with the nutrientsupply. In the deep-sea, low resource sup-ply is correlated with high diversity, andon the shelves there appears to be a generallatitudinal trend towards high annual pro-duction with increasing latitude and de-creasing diversity except at the highestlatitudes. Furthermore, upwelling createslocally high productivities by introducingnutrients into the euphotic zone, and insuch localities diversity is lower than incomparable regions that lack upwelling(Ryther, 1969). In addition to very highlatitudes, an exception to the correlation ofhigh productivity with low shallow-waterdiversity is posed by tropical reefs, whichare highly diverse and are reported to behighly productive (Sargent and Austin,1949; Odum and Odum, 1955; Kohn andHelfrich, 1957).
A strong correlation, which appears to bethe most significant of all, is between thestability of trophic resource supply anddiversity. The relatively constant supplyof solar energy to low latitudes results ina relatively constant productivity, while thehighly seasonal solar energy flux in highlatitudes results in high seasonal fluctua-tions in productivity. This correlates wellwith the latitudinal diversity gradient, fromequator to pole. Furthermore, the low di-versity of regions of seasonal upwelling canbe explained by the relative instability oftheir nutrient supply. And, finally, thealternation of cool northern waters andwarm southern waters that produces thehigh seasonal temperature variations withinmany mid-latitude shelf provinces must also
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from

TECTONICS: MARINE DIVERSITY AND ENDEMISM 257
cause a variation in productivity owing tothe associated changes in nutrient supply.Thus there appears to be a strong correla-tion, although to be sure the pattern ofproductivity fluctuation is not well knownand has not been compared to diversity inany detail. As investigations of marineproductivity patterns progress, the hy-pothesis of diversity regulation by resourcestability will be amply tested.
The focus here upon resource stabilityas the primary control of latitudinal di-versity is not intended to imply that otherfactors do not play important and locallysignificant roles. Spatial heterogeneity ofthe environment both on the scale of specieshabitats and of community biotopes clearlycontributes to diversity levels. Thereforeit is necessary to compare two communities(or provinces) from environments withsimilar spatial heterogeneities in order toreduce this effect. This was done for ex-ample by Hessler and Sanders (1957) whocompared the mud-bottom deep-sea ben-thos with the mud-bottom benthos ofshallow-water regions and not with theentire, heterogeneous shelf environment.Moreover, it has been recognized for manydecades that the isolation of islands frombiotic source regions lowers diversity inmany cases (see MacArthur and Wilson,1967). Other factors, such as the controlof population size or habitat by predators(Paine, 1966), competitors (Connell, 1970)or by density-independent factors surelyoperate as well and can be of considerablesignificance. It is noteworthy that whenthese factors enhance diversity it is usuallybecause they prevent populations fromutilizing all the resource supplies that theywould otherwise exploit.
LONGITUDINAL DIVERSITY PATTERNS OFSHALLOW MARINE BIOTAS
In addition to the latitudinal diversitygradients, there are large longitudinal dif-ferences in marine shelf diversities in dif-ferent oceans and on different sides of thesame ocean, even when similar latitudesor similar climatic provinces are compared.
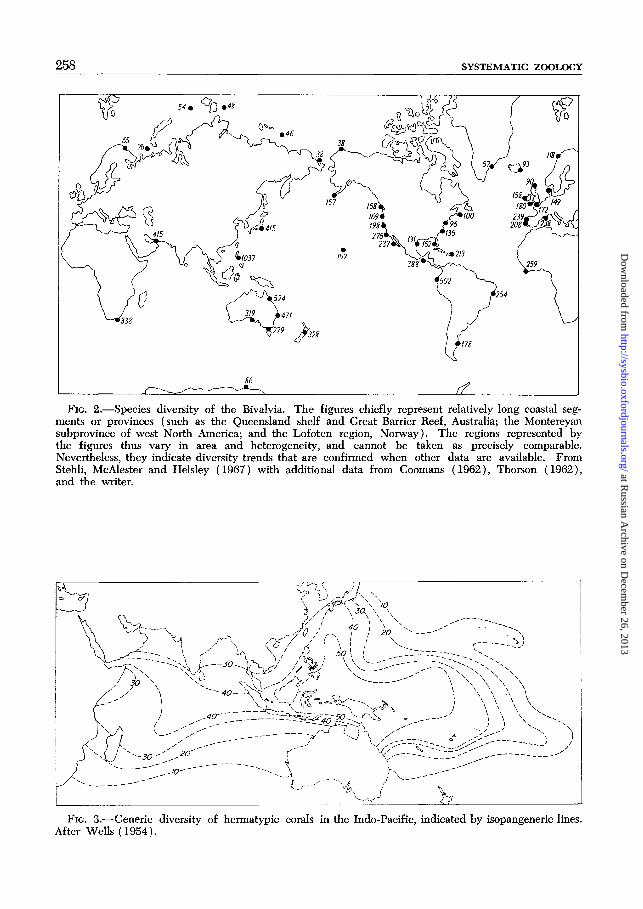
They are especially well shown by the bi-valve data presented by Stehli, McAlesterand Helsley (1967), which are plotted onFig. 2 with added data. If the westernPacific is taken as a starting point, diversitytends to fall in either direction, latitude forlatitude or climatic zone for climatic zone,reaching a low point in the western Atlantic.
There are risks associated with relyingupon the diversity pattern of only a singleclass such as the Bivalvia, for special adap-tive features perhaps related to the classground-plan might cause it to be unrepre-sentative of the entire biota at all localities.In fact, the Bivalvia are probably somewhatbiased in this way. For example, they seemto form a larger proportion of the westernAtlantic than of the eastern Pacific biotas.Nevertheless, wherever data are availablefor other large groups they support therelative trends displayed by the bivalves,and this is sufficient for present purposes.In Fig. 3, the Indo-Pacific region is con-toured according to the diversity of herma-typic coral genera (Wells, 1954). Thehighest diversity is centered on the broadshelves from Borneo eastward to the Gil-berts in low latitudes, with the well-knownnorthward extension along the KuroshioCurrent. In the east, generic diversity re-mains high along the chain of island arcsassociated with subduction zones, but dropsoff east of Samoa, so that the more isolatedand less continuous shallow-water regionsassociated with the eastern mid-plate vol-canic chains support fewer genera. Quanti-fication of total biotic diversity patternsand quantitative analysis of various effectsare much-desired goals, but data are in-sufficient at this time.
It seems possible to interpret these trendson the basis of the same principles that areused to explain latitudinal diversity, that is,with stability (especially of energy flow)as the chief environmental factor. Sanders(1969) has pointed out that there are longi-tudinal variations in environmental fluctua-tions—he was concerned principally withtemperature fluctuations—and that they ap-pear to correlate with the diversity trends
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from
258 SYSTEMATIC ZOOLOGY
FIG. 2.—Species diversity of the Bivalvia. The figures chiefly represent relatively long coastal seg-ments or provinces (such as the Queensland shelf and Great Barrier Reef, Australia; the Montereyansubprovince of west North America; and the Lofoten region, Norway). The regions represented bythe figures thus vary in area and heterogeneity, and cannot be taken as precisely comparable.Nevertheless, they indicate diversity trends that are confirmed when other data are available. FromStehli, McAlester and Helsley (1967) with additional data from Coomans (1962), Thorson (1962),and the writer.
FIG. 3.—Generic diversity of hermatypic corals in the Indo-Pacific, indicated by isopangeneric lines.After Wells (1954).
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from

TECTONICS: MARINE DIVERSITY AND ENDEMISM 259
where he has studied them. He recognizedtwo climatic situations in the boreal climaticzone, a continental and a maritime, whichare characterized by relatively large andsmall temperature variations, respectively.On the basis of the evidence available, con-tinental areas, as represented by the borealnorthwestern Atlantic, have less diversitythan maritime areas as represented by theboreal northeastern Pacific. Except in thehighest latitudes, seasonal variation in watertemperature usually signifies that summerand winter waters are derived from separatesources, with cool waters derived eitherfrom higher latitudes or from upwelling,and warm waters either from offshore orfrom lower latitudes. Owing to the patternsof water conditioning, circulation and mix-ing, and nutrient uptake and regeneration,nutrient concentrations are nearly alwayshigher in these cool waters (Redfield,Ketchum and Richards, 1963; Ryther, 1963).Thus the differences in temperature fluc-tuations in these different climatic situationsare probably associated with differences inproductivity regimes, with the greatestfluctuations in productivity where the tem-perature fluctuations are greatest. I there-fore suggest that the productivity factor ismore fundamental in regulating longi-tudinal diversity than is temperature.
There are some general considerationsthat could be expected to apply to the longi-tudinal distribution of environmental sta-bility. First, instability should tend to bemost pronounced on shelves that borderlands with the most continental climates(Sanders, 1969), which usually means thelargest land mass for any given latitude.Secondly, instability should be great alongthe smallest, most land-locked oceans,which would be expected to have the leastameliorating "maritime" influence on theclimate of bounding lands and to be mostunder the influence of continentality(Briggs, 1970). And third, instability shouldbe lowest around small islands in largeoceans, making due allowance for theirdegree of isolation and of their spatialheterogeneity. This pattern seems to be
clearly represented in the present oceans.For example, in very low latitudes thegreatest diversity is associated with thestring of small continents, island arcs andother volcanic chains in the Indo-Pacific,
'and diversity decreases with increasing iso-lation or latitude. Somewhat less diversityis associated with the tropical coastalmargin facing the largest ocean, on the eastPacific shelf. And the least diversity isassociated with the tropics of the narrowAtlantic Ocean. In very high latitudes, thehighest diversity (such as it is) is foundon the Antarctic shelf which chiefly faceslarge oceanic expanses, and the least di-versity is found around the small andrelatively landlocked Arctic Ocean (Fig. 2).
ADAPTIVE BASIS OF STABILITY REGULATION
OF DIVERSITY
It is appropriate to review briefly thereasons that resource stability is believedto be so important in regulating speciesdiversity. In environments with stable re-source supplies, the argument goes, popula-tions need not have much trophic breadthand indeed may become highly specializedon one or a few prey species or upon feed-ing in a particular habitat. The fact thatsuch specialization lowers the resourcesupply level and the population size pre-sents no special problem, for the stabilityof the resource guarantees adequate sup-port. When new species appear they canbe accommodated within a community insuch stable environments simply by a par-titioning of the resources at their trophiclevel. Again, the consequent reduction inaverage population size does not threatenthe ability of populations to endure.
In environments wherein trophic re-sources fluctuate, however, a flexible lifestyle is favored. When resources fall tolow levels, the populations that live in thegreatest variety of habitats and that eatthe largest range of food items will havethe best chance of surviving. And whenresource levels rise, the population thatresponds most rapidly and extensively byincreasing its population size is apt to ob-
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from

260 SYSTEMATIC ZOOLOGY
tain a larger share of the resources and tobe in a better position to endure the nextunfavorable period. When new speciesappear they are not easily accommodated,for the accent on selection for high re-productive potential results in a strongcompetition for resources. Furthermore, re-sources cannot easily be partitioned, for theresulting reduction in the sizes of the popu-lations involved would greatly lower theirchances of survival during unfavorableperiods.
Therefore in energetically stable environ-ments, numerous specialized, efficient pop-ulations can be accommodated within anecosystem, together with generalists as well,and population sizes may be small on theaverage. In energetically unstable environ-ments, by contrast, relatively few popula-tions can coexist, for they are chiefly ofbroadly-adapted, inefficient species that re-quire large populations during favorableperiods. Trophic resources are thus espe-cially important because they form a vari-able, density-dependent limiting factor. Theincrease in the numbers of species withinecosystems at low latitudes can thus in thisview be attributed chiefly to the increasingstability of trophic energy. For differentstatements of the adaptive strategies ofpopulations in stable versus unstable en-vironments, see MacArthur, 1955; Klopfer,1962; MacArthur and Wilson, 1967; Mar-galef, 1968; Levins, 1968; and Bretsky andLorenz, in press. To the extent that thesedifferent adaptive strategies are promotedby environmental regimes that result, atleast in part, from tectonic patterns, tec-tonics exercises an influence on evolutionarypathways.
SUMMARY OF TECTONIC-CLIMATIC MODEL OF
DIVERSITY AND PROVINCIALITY
The present geography of continents,ocean basins, ridges, trenches and lessertectonic features is transitory, and is basedupon the present state of plate tectonicprocesses. The interaction of this geo-graphic pattern with patterns of solarenergy reception and distribution that are
related to planetary shape and motion pro-duces a characteristic climatic pattern. Atpresent in the oceans there are many north-south trending continental shelves and ahigh latitudinal temperature gradient, creat-ing chains of latitudinal provinces. East-west trending shelves tend to support singleprovincial biotas so long as they are notinterrupted by irregularities that extendlatitudinally into different climatic regimes.The same principles apply to chains ofislands; those that trend north-south acrossclimatic regimes support latitudinally dis-tinctive provinces (as Japan) while thosethat trend east-west support single pro-vincial biotas, so long as they are not inter-rupted by barriers.
Diversity is highest in low latitudes, andhighest on island chains or highly maritimeshelves; it is lower in mid-latitudes andalong shelves that face large oceans; andit is least in high latitudes and on shelvesof large continents that face small oceans.This pattern appears to support the hy-pothesis that high diversity is permitted byand correlated with high trophic resourcestability. Local secondary effects, such asthe presence of a large seasonal runoff,certainly cause local deviations from thesetrends, which nevertheless hold on a globalscale.
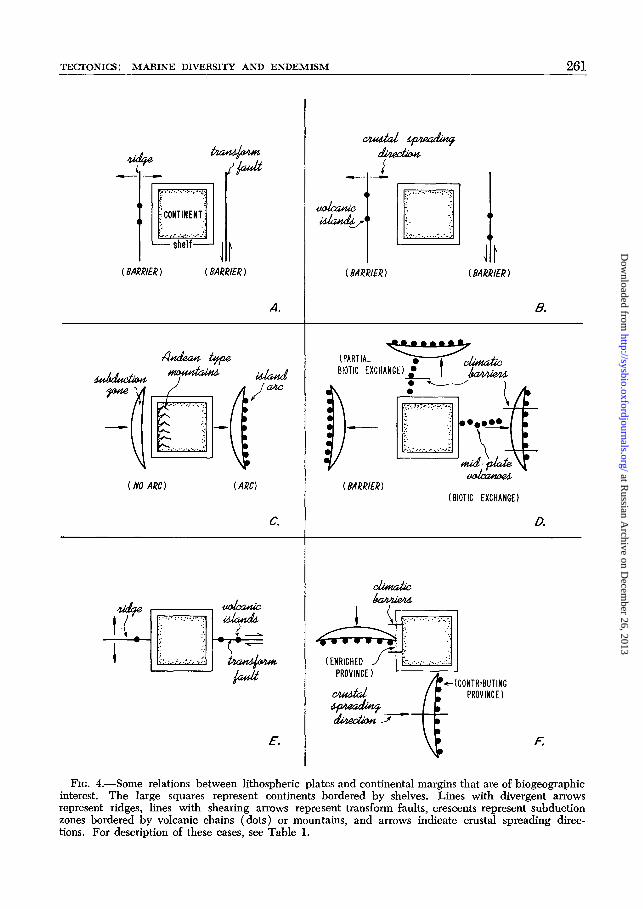
Thus from a biogeographic standpoint,oceanic plate margins fall into two classes.One class is formed by ridges and trans-form fault zones, which are particularlyeffective barriers, although they may beassociated with scattered islands that bearimproverished biotas. The other class isformed by subduction zones, which areaccompanied by island arcs and thus pro-vide island chains to support highly diversebiotas for their latitudes. In Table 1 andFig. 4 the possible relations of continentalshelves to plate margins are summarizedfrom a biogeographic standpoint. Trans-oceanic dispersals of major proportions arechiefly restricted to times when island arcsare near continents or when any interveningareas are occupied by mid-plate volcanoes
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from
TECTONICS: MARINE DIVERSITY AND ENDEMISM 261
lidqe.
CONTINENT
1 — shelf-
(BARRIER) (BARRIER)
cnuital
diaecti&n
——
<volcanic
1 \
i
J(BARRIER) (BARRIER)
B.
Andean.
Uiltductio+i aland(PARTIALBIOTIC EXCHANGE) •
one
(NO ARC) (ARC) (BARRIER)
mid-platevolcanoes
(BIOTIC EXCHANGE)
c. D.
climatic
lidae volcanicidiands
(ENRICHED JPROVINCE)
cnuital6-p/ieadinfdirection-
E.
-(CONTRIBUTINGPROVINCE)
F.
FIG. 4.—Some relations between lithospheric plates and continental margins that are of biogeographicinterest. The large squares represent continents bordered by shelves. Lines with divergent arrowsrepresent ridges, lines with shearing arrows represent transform faults, crescents represent subductionzones bordered by volcanic chains (dots) or mountains, and arrows indicate crustal spreading direc-tions. For description of these cases, see Table 1.
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from

262
Geometryrelation
TABLE 1.
of Charactermargin
POSSIBLE
of
RELATIONS
Distance
OF CONTINENTAL SHELVES
Biogeographic implications
SYSTEMATIC
TO PLATE MARGINS.
for Continental Shelf
ZOOLOGY
Fig.
PAR
AL
]
Ridgeor
Transform
Subduction
Zone
Near
Far
Near
Far
Barrier but with depauperate provincial out-liers on isolated islands 3A
Barrier 3B
Barrier if truly marginal with no island arc;Source of rich biota and dispersal route 3Cif arc present
No effect unless intervening region bridgedby mid-plate volcanoes, then source of 3Drich biota and dispersal route if no cli-matic barriers intervene
W
o<Xu
RidgeorTransform
Subduction
Zone
Near Little effect, with depauperate provincialoutliers on isolated islands 3E
Far Not a case
Near N-S shelf, E-W arc: arc system a source ofrich biota for local province. E-W shelf,N-S arc: proximal province of arc system 3Fa source of rich biota for entire shelf
Far Not a case
but not by ridges (and possibly not bytransform fault zones).
Many of the cases depicted in Fig. 4occur today. Fig. 4A illustrates the caseof a ridge near and parallel to a continentalmargin, which is represented by the EastPacific Rise off southern Mexico, and thecase of a transform fault near and parallelto a continental margin, such as representedby the San Andreas and other faults alongthe northeastern Pacific margin. Distantridges that are parallel to continental mar-gins (Fig. 4B) are represented by the EastPacific Rise off Central America, the mid-Atlantic Ridge, and the Antarctic-Pacificand Antarctic-Indian Ridges, and distanttransform faults that are parallel to con-tinental margins occur between the Ant-arctic and African Plates. In all of thesecases, barriers are formed by the transformfaults and ridges, although if the ridgeseventually abut continental shelves the shelfbiota may circumvent the barrier simplyby dispersing along the continental margin.
Subduction zones parallel to and close tocontinental margins may be of the Peru-
Chile Trench type, associated with terres-trial mountains (Fig. 4C), and in this eventthey form barriers. However, most sub-duction zones lie along island arcs and thechains of shoals on the island flanks areusually linked closely since active subduc-tion zones are continuous. Thus they canbe superhighways of dispersal (Fig. 4C);a modern example is formed by island arcsparallel with and just north of the Australia-New Guinea continent. If a subductionzone is distant from a continental margin,the sea floor intervening between island arcand continent forms a barrier. However,should mid-plate volcanoes rise in this inter-vening region the barrier might be broachedby "island stepping stones," which permitmore or less dispersal according to localconditions. In Fig. 4D, cases of north-southmid-plate volcanic chains and east-westones are depicted. Climatic barriers as-sociated with north-south chains will restrictabundant faunal deployment to the prov-ince nearest the shelf, a province whichmay not include the island arcs at thesubduction zone. Climatic barriers as-
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from

TECTONICS: MARINE DIVERSITY AND ENDEMISM 263
sociated with north-south island arcs willlimit dispersal and interchange to the prov-ince along the island arc which connectswith any mid-plate volcanic chain. Noclear examples of the linking of island arcsto continents are present today. When sub-duction zones abut continental margins athigh angles, two main cases are found (Fig.4F). If the arcs are chiefly east-west, theyserve as dispersal routes to and from thecontinental shelf province with which theyconnect, as for example, the Aleutian Is-lands with North America, and the Indo-nesian-Malaysian, archipelago and arcs withthe Indian continent. If on the other hand,the arcs trend principally north and south,then they are likely to be partitioned bioti-cally into latitudinal provinces, and onlythe province nearest the shelf will representan open biotic source and disperal routethereto. Of course, climatically tolerantspecies may penetrate provincial barriersinto more distant provinces in these cases.Intercontinental dispersal occurs when con-tinents themselves are in proximity. Innature the irregularities in coastlines andplate margins permit mixtures of thesecases to occur.
It is well known that continental posi-tions have been vastly different in the past,and it is certain that the relations oftrenches, ridges, and continents have alsochanged. Clearly there have been corre-sponding differences in past climatic re-gimes, in the sizes of continents and oceans,and in dispersal routes and barriers, changesthat are fundamental to the evolution of theecological structure of the biosphere (Val-entine, 1969, Valentine and Moores, 1970).This article has dealt chiefly with somebiogeographic consequences of present platetectonic patterns; a future contribution willexamine Phanerozoic variations in continent-continent and continent-plate margin rela-tions and their biogeographic results. It isappropriate to note here, however, that pastpatterns of marine faunal affinities do notnecessarily define relative continental po-sitions, for the relations of plate marginsto the continents must also be taken into
account. The "land bridges" and "islandstepping stones" of biogeography have atlast been given an effective tectonic for-mulation in trans-oceanic chains of islandarcs associated with plate margins.
ACKNOWLEDGMENTS
I have benefitted greatly throughout thepreparation of this paper from discussionswith Professor E. M. Moores, Departmentof Geology, University of California, Davis.The manuscript has been critically evalu-ated by Professor Moores and by ProfessorW. M. Hamner, Department of Zoology,University of California, Davis. Drafting isby Mrs. Ruth Darden.
REFERENCES
BRETSKY, P. W., AND D. M. LORENZ. In press.Adaptive response to environmental stability: aunifying concept in paleoecology. Proc. N.Amer. Paleontol. Conv.
BRIGGS, J. C. 1970. A faunal history of the NorthAtlantic Ocean. Syst. Zool., 19:19-34.
BULLOCK, T. H. 1955. Compensation for tem-perature in the metabolism and activity ofpoikilotherms. Biol. Rev., 30:311-342.
CONNELL, J. H. 1970. A predator-prey system inthe marine intertidal region. 1. Balanus glandulaand several predatory species of Thais. Ecol.Monogr., 40:49-78.
COOMANS, H. E. 1962. The marine molluskfauna of the Virginia area as a basis for definingzoogeographical provinces. Beaufortia, 9:83-104.
DEWEY, J. F., AND B. HORSFIELD. 1970. Platetectonics, orogeny and continental growth.Nature, 225:521-525.
EKMAN, S. 1953. Zoogeography of the sea. Sidg-wick and Jackson, London, 417 p.
FISHER, A. G. 1960. Latitudinal variations inorganic diversity. Evolution, 14:64—81.
HESSLER, R. R., AND H. L. SANDERS. 1967.Faunal diversity in the deep-sea. Deep-Sea Res.,14:65-79.
KINNE, O. 1963. The effects of temperature andsalinity on marine and brackish water animals. I.Temperature. Oceanogr. Marine Biol., 1:301—340.
KLOPFER, P. H. 1962. Behavioral aspects ofecology. Prentice-Hall, Englewood Cliffs, N. J.,173 p.
KOHN, A. J., AND P. HELFRICH. 1957. Primaryorganic productivity of a Hawaiian coral reef.Limnol. Oceanogr., 2:241-251.
LE PICHON, X. 1968. Sea-floor spreading and
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from

264 SYSTEMATIC ZOOLOGY
continental drift. Jour. Geophys. Res., 73:3661-3697.
MACARTHUR, R. H., AND E. O. WILSON. 1967.The Theory of island biogeography. PrincetonUniv. Press, Princeton, N. J., 203 p.
MARGALEF, R. 1968. Perspectives in ecologicaltheory. Chicago Univ. Press, Chicago, 111 p.
MCKENZIE, D. P., AND R. L. PARKER. 1967. TheNorth Pacific: An example of tectonics on asphere. Nature, 216:1276-1280.
MENARD, H. W. 1969. Growth of drifting vol-canoes. Jour. Geophys. Res., 74:4827-4837.
MORGAN, W. J. 1968. Rises, trenches, greatfaults, and crustal blocks. Jour. Geophys. Res.,73:1959-1982.
ODUM, H. T., AND E. P. ODUM. 1955. Trophicstructure and productivity of a windward coralreef community on Eniwetok Atoll. Ecol.Monogr., 25:291-320.
PAINE, R. T. 1966. Food web complexity andspecies diversity. Amer. Nat., 100:65-75.
PAINKA, E. R. 1966. Latitudinal gradients inspecies diversity: A review of concepts. Am.Naturalist, 100:33-46.
REDFBELD, A. C , B. H. KETCHUM, AND F. A. RICH-ARDS. 1963. The influence of organism on thecomposition of sea-water. In M. N. Hill, ed.,The Sea, 2:26-77. N. Y., Interscience.
RYTHER, J. H. 1963. Geographic variations inproductivity. In M. N. Hill, ed., The Sea, 2:347-380. N. Y., Interscience.
RYTHER, J. H. 1969. Photosynthesis and fishproduction in the sea. Science, 166:72-76.
SANDERS, H. C. 1968. Marine benthic diversity:a comparative study. Amer. Nat., 102:243-282.
SANDERS, H. C. 1969. Benthic marine diversityand the stability-time hypothesis. BrookhavenSymp. in Biol., 22:71-81.
SARGENT, M. C , AND T. S. AUSTIN. 1949. Or-ganic productivity of an atoll. Trans. Amer.Geophys. Union, 30:245-249.
SLOBODKIN, C. B., AND H. L. SANDERS. 1969.On the contribution of environmental predict-ability to species diversity. Brookhaven Symp.in Bio., 22:82-95.
STEHLI, F. G., A. L. MCALESTER, AND C. E. HELS-LEY. 1967. Taxonomic diversity of recent bi-valves and some implications for geology. Geol.Soc. Amer. Bull., 78:455-466.
THORSON, G. 1962. The distribution of benthicmarine mollusca along the N.E. Atlantic shelffrom Gibralter to Murmansk. Proc. First Europ.Malac. Congr., 5-23.
VALENTINE, J. W. 1966. Numerical analysis ofmarine molluscan ranges on the extratropicalnortheastern Pacific shelf. Limnol. and Oceangr.,11:198-211.
VALENTINE, J. W. 1967. Influence of climaticfluctuations on species diversity within theTethyan Provincial System. Syst. Assn. Pub. 7:153-166.
VALENTINE, J. W. 1969. Patterns of taxonomicand ecological structure of the shelf benthosduring Phanerozoic time. Palaeontology, 12:684-709.
VALENTINE, J. W., AND E. M. MOORES. 1970.Plate-tectonic regulation of biotic diversity andsea level: a model. Nature.
VINE, F. J. 1966. Spreading of the ocean floor:new evidence. Science, 154:1405-1415.
VINE, F. J. 1969. Sea-floor spreading—new evi-dence. Jour. Geol. Educa., 17:6-16.
VINE, F. J., AND D. H. MATHEWS. 1963. Mag-netic anomalies over oceanic ridges. Nature,199:947-949.
WELLS, J. W. 1954. Recent corals of the Mar-shall Islands. U. S. Geol. Surv., Prof. Paper260-1: 385-486, pis. 94-187.
WILSON, J. T. 1963. Evidence from islands onthe spreading of ocean floors. Nature, 197:536-538.
Manuscript received July, 1970
at Russian A
rchive on Decem
ber 26, 2013http://sysbio.oxfordjournals.org/
Dow
nloaded from





























