Embed Size (px)
Citation preview

Plasminogen Activator and MatrixMetalloproteinase Production and Extracellular
Matrix Degradation by Rat Prostate CancerCells In Vitro: Correlation With Metastatic
Behavior In Vivo
P.H.A. Quax,1* A.C.W. de Bart,1 J.A. Schalken,2 and J.H. Verheijen1
1Gaubius Laboratory TNO-PG, Leiden, The Netherlands2Department of Urology, University Hospital, Nijmegen, The Netherlands
BACKGROUND. The plasminogen activation (PA) and metalloproteinase (MMP) system areinvolved in tumor cell migration and invasion.METHODS. The proteolytic activity of cell lines originating from the rat Dunning R-3327prostate tumor was analyzed by measuring in vitro extracellular matrix degradation, enzymeactivity, and mRNA levels of enzymes, inhibitors, and receptors, and compared with theirknown metastatic behavior in vivo.RESULTS. Only the highly metastatic sublines AT-3, MATLu, and MATLyLu showed a highextracellular matrix degradation mediated by urokinase-type plasminogen activator (u-PA).Relatively high levels of u-PA were present in the aggressive cell lines. u-PA receptor mRNAwas produced in all cells, and all but AT-1 produced LDL-receptor-related-protein (LRP)mRNA. t-PA mRNA was only found in HIF and MATLu. In gelatin, zymography lysis wasobserved at 72 kD and 74–76 kD in MATLu and MATLyLu cells, respectively. MMP-2 mRNAwas present in all cell lines except AT-1 and AT-2, and MMP-3 mRNA was present in AT-2,AT-3, and MATLu.CONCLUSIONS. These in vitro experiments show that in different rat prostate cancer sub-lines, proteolytic activity and u-PA-mediated extracellular matrix degradation correlate withtheir known metastatic behavior in vivo. Prostate 32:196–204, 1997. © 1997 Wiley-Liss, Inc.
KEY WORDS: prostate; cancer; urokinase-type plasminogen activator; matrix metallo-proteinase; extracellular matrix
INTRODUCTION
Both the plasminogen activator system, consistingof urokinase-type (u-PA) and tissue-type plasminogenactivator (t-PA), their inhibitors, and receptors, andthe matrix metalloproteinase (MMP) system, consist-ing of various MMPs and their inhibitors, have beenimplied in invasion and metastasis of various tumorcells [1,2] since malignant cells frequently produce el-evated levels of these proteolytic enzymes. In vivou-PA is found in malignant melanoma [3], just as hasbeen reported for breast and colon cancer in whichelevated levels of u-PA and plasminogen activator in-hibitor type 1 (PAI-1) appear to be associated with a
Abbreviations: t-PA, tissue-type plasminogen activator; u-PA, urokinase-type plasminogen activator; PAI-1, plasminogenactivator inhibitor type 1; PAI-2, plasminogen activator inhibitortype 2; u-PAR, urokinase-type plasminogen activator receptor;LRP, LDL-receptor related protein/a2-macroglobulin receptor;FCS, fetal calf serum; DME, Dulbecco’s modified Eagle’s me-dium; cDNA, complementary DNA; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; MMP, matrix metalloproteinase;TIMP, tissue inhibitor of metalloproteinase.*Correspondence to: Dr. P.H.A. Quax, Gaubius Laboratory TNO-PG, P.O. Box 2215, 2301 CE Leiden, The Netherlands.E-mail: [email protected] 1 April 1996; Accepted 28 August 1996
The Prostate 32:196–204 (1997)
© 1997 Wiley-Liss, Inc.

bad prognosis [4–6]. The involvement of u-PA in tu-mor development and metastasis has also been dem-onstrated in vitro [7,8]. Different members of the ma-trix metalloproteinase family are expressed in a largevariety of tumors including colon, prostate, and breastcancer [2,9–11] and tumor cell lines [12]. In malignantcells not only is the production of u-PA increased, butoften an increased production of the inhibitors PAI-1or PAI-2 and the receptor u-PAR can be observed[6,13,14].
In the normal prostate gland various proteases,such as plasminogen activators and matrix metal-loproteinases, are involved in the regulation of thedevelopment and normal cellular operations in theprostate gland [15]. The same enzymes appear to beinvolved in the development of prostate cancer. Ele-vated levels of activities of plasminogen activatorshave been found in prostatic tumors and tumor celllines both in rats and humans. For the human cell linesDu-145, PC-3, and 1-LN-PC-3-1A, a correlation is sug-gested between the levels of u-PA and the invasivebehavior of the cells [16,17]. Furthermore, it was sug-gested that certain MMPs are important in the regu-lation of prostate tumor progression since the moreaggressive cells show elevated levels of MMP expres-sion [17]. The presence of cell surface receptors forurokinase on PC-3 and Du-145 cells is also associatedwith invasive behavior [18,19]. The role of the variouscomponents of the plasminogen activator system andthe matrix metalloproteinase system in the stroma in-vasion and metastasis formation of prostate cancercells is not completely understood [20].
A well-defined model for prostate tumor is theDunning R-3327 rat prostate tumor [21] and the celllines derived from this tumor, with varying aggressiveand metastatic phenotypes [22,23]. Using these celllines it was demonstrated that various biochemicalmarkers for metastatic phenotypes, such as E-cadherin, differed in the sublines [23–25]. Elevatedlevels of plasminogen activator [26], and of MMP-2and MMP-9 [16], were found in the Dunning rat pros-tate tumor when compared with the dorsal prostate,which is considered the tissue source of the originaltumor [21]. Although Lowe and Isaacs [23] did notdetect any difference in plasminogen activator or ma-trix metalloproteinase activities in variants of the Dun-ning tumor cell lines, inhibition of u-PA activity wasshown to decrease tumor volume [26].
In malignant cells not only is the production ofu-PA increased, but often an increased production ofthe inhibitors PAI-1 or PAI-2 and the receptor u-PARcan be observed [6,13,14]. In this study we analyzedthe role of various components of the plasminogenactivation system and the metalloproteinase family inprostate tumor progression in a set of Dunning rat
prostate cancer cell sublines, and compared this withtheir known metastatic behavior in vivo in correlationwith the proteolytic activity in an in vitro matrix deg-radation assay.
MATERIALS AND METHODS
Dunning R-3327 Rat Prostatic Tumors
The parental tumor from which all rat prostatic tu-mor sublines have been derived is the original R-3327tumor initially discovered in 1963 by Dunning [21] ina 22-month-old inbred Copenhagen male rat, andoriginates from dorsal prostate tissue. From variousR-3327 tumors with different characteristics, cell lineswere established. Here we use the HIF, AT-1, AT-2,AT-3, MATLu, and MATLyLu cell lines. All cell linesexcept HIF have the same characteristics as the tumorsthey are derived from (HIF is not tumorigenic afterinoculation in animals), as confirmed by reinoculationexperiments [22]. The tumor sublines HIF and AT-1have a low metastatic ability, the AT-2 cell line has alow to moderate metastatic ability, and the cell linesAT-3, MAT-Lu, and MATLyLu are highly metastaticafter inoculation into animals, i.e., >75% of animalsinoculated with these tumor sublines develop distantmetastases. AT-3 and MATLyLu form metastasesmainly in the lymph nodes and the lung, whereasMATLu forms metastases only in the lung [22].
Tissue Extracts
Tissue extracts of tumors of AT-2, AT-3, MATLu,and MATLyLu grown in rats were prepared as de-scribed before [27]. Essentially, the samples wereweighed and homogenized in 1 ml 0.1% (v/v) Tween-80, 0.1 M Tris-HCl, pH 7.5, per 60 mg wet tissue at 0°C.The homogenate was centrifuged twice at 8,000g for2.5 min at 4°C, and the supernatant was stored at−20°C until analysis. Protein concentrations were de-termined using the BCA assay (Pierce, Rockford, IL).
Cell Culture
All cell lines were grown as monolayers in Dulbec-co’s modified Eagle’s (DME) medium supplementedwith 10% (v/v) fetal calf serum (FCS), 2 mM gluta-mine, 100 units/ml penicillin, and 100 mg/ml strepto-mycin. Culture media were changed every 2–3 days.For experiments, fresh medium without FCS, unlessindicated was added to the cells shortly before theyreached confluency. Twenty-four hours later condi-tioned medium was collected and cell extracts wereprepared for fibrin autography, MMP zymography,PA activity assay, and mRNA analysis.
PAs and MMPs in Rat Prostate Cancer Cell Lines 197

Fibrin Autography
Media, cell extracts, and tumor tissue extracts weresubjected to electrophoresis on 10% (w/v) sodium do-decyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels. After electrophoresis, the gels werewashed in 2.5% Triton X-100 (Sigma Chemical Co., St.Louis, MO), and plasminogen activator activities werevisualized essentially according to the method ofGranelli-Piperno and Reich [28].
Plasminogen Activator Activities
The plasminogen activator activities in the culturemedia, cell extracts, and tissue extracts were deter-mined using the indirect spectrophotometric assay asdescribed [29] in the presence and absence of antibod-ies against human t-PA [30] and u-PA [31], known toblock the activity of rat t-PA and u-PA, respectively.Activities were expressed in international units (IU).Purified u-PA or t-PA preparations standardizedagainst the respective International Standard prepara-tions on an activity basis, using the specific activitiesof 100,000 IU/mg u-PA (WHO preparation c66/46)[32] and 500,000 IU/mg t-PA (WHO preparation 83/517) [33], were used as standards.
Zymography for Metalloproteinases
Gelatinolytic activity of secreted MMPs was ana-lyzed by zymography on gelatin-containing poly-acrylamide gels as described previously [34]. Usingthis technique, both active and latent species of theMMP family can be visualized. Samples were sub-jected to electrophoresis on 10% (w/v) polyacryl-amide gels copolymerized with 0.2% (w/v) gelatin.After electrophoresis the gels were washed twice for15 min in 2.5% (w/v) Triton X-100 to remove the SDS,followed by two washes of 5 min in 50 mM Tris-HCl,pH 8.0, containing 5 mM CaCl2 and 1 mM ZnCl2 andincubated overnight in the same buffer at 37°C. Thegels were stained with Coomassie Brilliant Blue R-250(Sigma Chemical Co.).
mRNA Analysis
Cells were washed with phosphate-buffered saline(PBS) at 37°C and lysed in 4 M guanidnium thiocya-nate, 25 mM sodium citrate, pH 7.5, 0.5% (w/v) sar-kosyl, and 0.1 M 2-mercaptoethanol. RNA was iso-lated according to Chomczynski and Sacchi [35]. Thetotal amount of RNA isolated was determined bymeasuring the OD260, assuming that 1 OD260 unit isequivalent to 40 mg RNA.
RNA samples were electrophoresed on a 1.2% (w/
v) denaturating agarose gel containing 7.5% (v/v)formaldehyde and were transferred to a nylon mem-brane (Hybond N, Amersham, Buckinghamshire, UK)using a Vacugene system (Pharmacia, Uppsala, Swe-den). Membranes were hybridized with 32P-labeledcDNA fragments in 7% (w/v) SDS, 0.5 M NaHPO2,pH 7.2, and 10 mM EDTA at 65°C. Blots were rou-tinely washed with 2 × SSC, 1% w/v SDS for 1 hr at65°C (1 × SSC = 0.15 M NaCl, 0.015 M sodium citrate).cDNA fragments were labeled using the randomprimer method (Megaprime; Amersham), with 32P-dCTP (specific activity was approximately 109 cpm/mg DNA). Autoradiograms were prepared using Ko-dak XAR-5 films and intensifying screens at −70°C.
cDNA Probes
For hybridization experiments the following cDNAfragments were used as probes: a 0.3-kb EcoRI-AvaIfragment of the rat u-PA cDNA, a 0.3-kb PvuII frag-ment of rat t-PA cDNA, both kindly provided by Dr.J. Degen (Children’s Hospital, Cincinnati, OH), a 0.9-kb PvuII fragment of rat PAI-1 cDNA kindly providedby Dr. T. Gelehrter (University of Michigan, Ann Ar-bor, MI) [36], and a 0.9-kb PstI-PvuII fragment of mu-rine PAI-2 cDNA, kindly provided by Dr. D. Belin(University of Geneva, Geneva, Switzerland) [37]. ThecDNA probes used for human u-PAR, human LRP,MMP-1, MMP-2, MMP-3, MMP-7, MMP-9, TIMP1,and TIMP2, as well as rat glyceraldehyde-3-phosphatedehydrogenase (GADPH), were described elsewhere[38].
Extracellular Matrix Preparation and Degradation
Bovine aortic smooth muscle cells were grown toconfluency in DME medium supplemented with 10%(v/v) FCS, 2 mM glutamine, 100 units/ml penicillin,and 100 mg/ml streptomycin in 2 cm2 culture dishes.After the cultures had reached confluency and the for-mation of extracellular matrix was started, the cellswere incubated for 4 days with medium containing a3H-amino-acid mixture (1 mCi/ml, Amersham). Cellswere then lysed using 0.5% (v/v) Triton X-100 in PBS,and the cytoskeleton was removed by 25 mM ammo-niumhydroxide treatment. Unincorporated 3H-aminoacids were washed from the remaining extracellularmatrix using H2O (twice) and 75% (v/v) ethanol. Ma-trices were dried and stored at −20°C until use. Beforetumor cells were seeded onto the 3H labeled matrices,the latter were soaked with medium for 1 hr. For theextracellular matrix degradation assay, 105 cells/2 cm2
dish were seeded onto the matrix in 10% (v/v) FCScontaining DME medium or DME medium supple-mented with 100 U/ml aprotinin (as inhibitor for plas-
198 Quax et al.

min) or anti-catalytic antibodies against human u-PA[31] or t-PA [30]. Antibody concentrations used wereapproximately 250 mg/ml for anti-human-u-PA incase of the HIF, AT-1, AT-2, and AT-3 cell lines, andapproximately 500 mg/ml for anti-human-u-PA incase of the MATLu and MATLyLu cell lines. Antibod-ies against human t-PA were used in a concentrationof approximately 250 mg/ml. The concentrations usedwere calculated as to completely neutralize the u-PAor t-PA produced by the cell lines, as assayed in anactivity assay [29]. Human plasminogen (BIOFINE,Leiden, The Netherlands), was added to the mediumin all experiments to a final concentration of 0.14 mM.After 24 hr the conditioned medium was removed andthe remaining matrix was degraded with 0.25% (w/v)trypsin (1 hr at 37°C). 3H-amino-acid release was de-termined, and the 3H release by tumor cells was ex-pressed as percentage of the total amount of 3H re-leased (the sum of the release by the cells and by thetrypsin treatment).
Statistical Analysis
Statistical analysis was performed using ANOVAFisher’s least significant difference (LSD) analysis orStudent’s t-test. Significance was accepted for P < 0.05.
RESULTS
Plasminogen Activator Activities in Rat ProstateCancer Cells
Just before the cultures reached confluency theywere incubated with fresh serum-free medium. After24 hr the conditioned media were collected and cellextracts were prepared. Analysis of plasminogen acti-vator activity (Fig. 1) revealed that all the cells pro-duced both t-PA and u-PA, although the level of t-PAin MATLyLu and AT-2 was extremely low. Only 22–41% of the t-PA was detected in the medium, and therest was found in the cell extracts. u-PA was detectedfor >95% of cells associated (either intracellularly orbound to the cell surface) with the cell lines producingthe highest amounts of u-PA (AT-2, AT-3, MATLu,and MATLyLu), whereas in HIF and AT-1 only 54%and 24% of u-PA was found in the cell extracts, re-spectively.
A more qualitative analysis of these samples usingfibrin autography showed that AT-3, MATLyLu, andMATLu, the high producers for u-PA, produce an in-hibitor, possibly PAI-1, that forms complexes witht-PA and/or u-PA (results not shown). In the other celllines, HIF, AT-1, and AT-2, no t-PA, u-PA, or PA/PAIcomplexes could be observed.
Analysis of the mRNA levels of the various com-
ponents of the plasminogen activator system (Fig. 2)showed that t-PA mRNA can be clearly detected inHIF, while in the other cell lines hardly any (MATLU)or no t-PA mRNA could be detected. u-PA mRNA isfound in high levels in AT-3 and MATLyLu, and atlower levels in AT-1, AT-2, and MATLu. Concerningthe receptors u-PAR and LRP, it was found that allcells produced small amounts of u-PAR mRNA, andall but AT-1 produced LRP mRNA with strong signalsfor LRP mRNA in the AT-3 and MATLu cell lines. Thehighest levels of PAI-1 and PAI-2 mRNA could befound in HIF and AT-3.
Analysis of the plasminogen activator activities intissue extracts of tumors derived from the studied celllines reveals a different pattern of PA activities. Quan-titative analysis of the t-PA and u-PA activity showedthat in AT-3 and MATLyLu tumors, relatively hight-PA activity could be detected as well as high u-PAactivities, whereas AT-2 and MATLu had relativelylow t-PA and u-PA activities (Fig. 3). Fibrin autogra-phy revealed that in AT-2 and MATLyLu tumors largeamounts of PA/PAI complexes could be found (notshown), whereas in MATLu hardly any activity couldbe detected, neither free t-PA nor u-PA nor PA/PAIcomplexes.
Extracellular Matrix Degradation
The capacity of Dunning rat prostate cells to de-grade extracellular matrix was tested by culturingthem on 3H-labeled extracellular matrix produced bybovine smooth muscle cells. We have previously
Fig. 1. Levels of t-PA and u-PA activity, produced by Dunningrat prostate carcinoma cell lines, expressed in mIU per 106 cellsper 24 hr. Data represent the sum of the activities found in theconditioned medium and the cell extracts, each measured in du-plicate. Results shown are the mean ± SD. u-PA levels in AT-2,AT-3, MATLu, and MATLyLu are significantly higher (P < 0.05)than in HIF and AT-1. t-PA in HIF and MATLu are significantlyhigher than in other cell lines (P < 0.05), but the difference be-tween HIF and MATLu was not significant.
PAs and MMPs in Rat Prostate Cancer Cell Lines 199
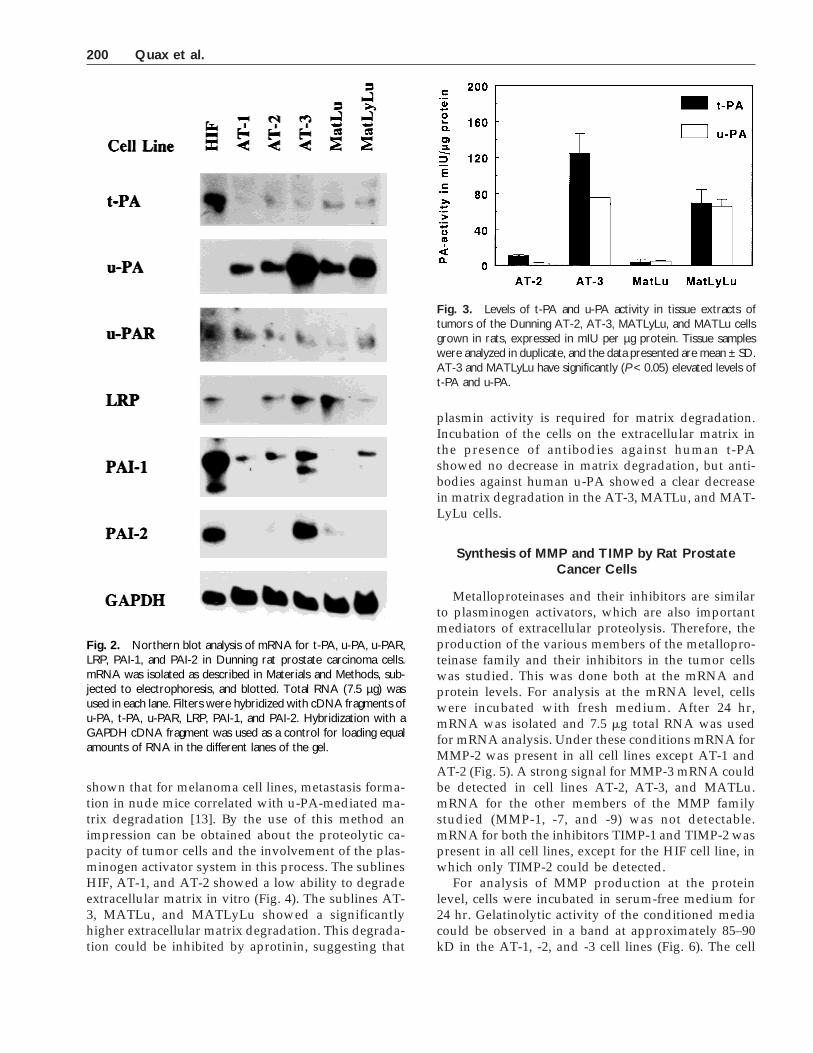
shown that for melanoma cell lines, metastasis forma-tion in nude mice correlated with u-PA-mediated ma-trix degradation [13]. By the use of this method animpression can be obtained about the proteolytic ca-pacity of tumor cells and the involvement of the plas-minogen activator system in this process. The sublinesHIF, AT-1, and AT-2 showed a low ability to degradeextracellular matrix in vitro (Fig. 4). The sublines AT-3, MATLu, and MATLyLu showed a significantlyhigher extracellular matrix degradation. This degrada-tion could be inhibited by aprotinin, suggesting that
plasmin activity is required for matrix degradation.Incubation of the cells on the extracellular matrix inthe presence of antibodies against human t-PAshowed no decrease in matrix degradation, but anti-bodies against human u-PA showed a clear decreasein matrix degradation in the AT-3, MATLu, and MAT-LyLu cells.
Synthesis of MMP and TIMP by Rat ProstateCancer Cells
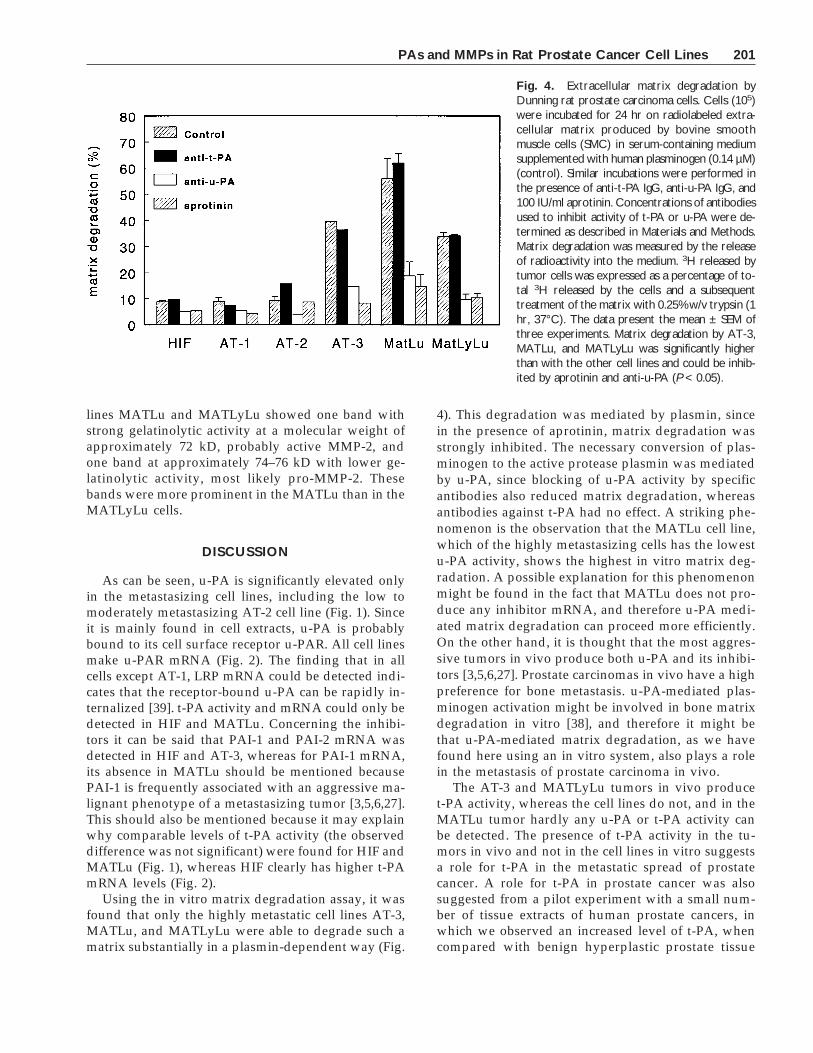
Metalloproteinases and their inhibitors are similarto plasminogen activators, which are also importantmediators of extracellular proteolysis. Therefore, theproduction of the various members of the metallopro-teinase family and their inhibitors in the tumor cellswas studied. This was done both at the mRNA andprotein levels. For analysis at the mRNA level, cellswere incubated with fresh medium. After 24 hr,mRNA was isolated and 7.5 mg total RNA was usedfor mRNA analysis. Under these conditions mRNA forMMP-2 was present in all cell lines except AT-1 andAT-2 (Fig. 5). A strong signal for MMP-3 mRNA couldbe detected in cell lines AT-2, AT-3, and MATLu.mRNA for the other members of the MMP familystudied (MMP-1, -7, and -9) was not detectable.mRNA for both the inhibitors TIMP-1 and TIMP-2 waspresent in all cell lines, except for the HIF cell line, inwhich only TIMP-2 could be detected.
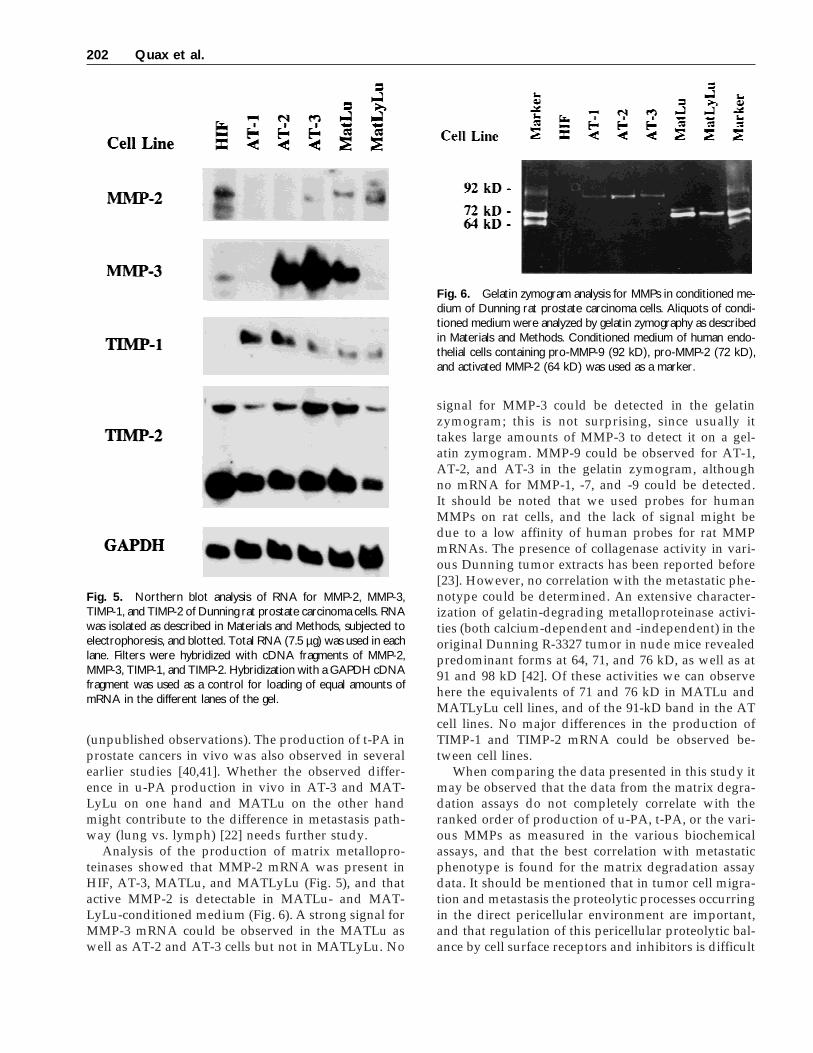
For analysis of MMP production at the proteinlevel, cells were incubated in serum-free medium for24 hr. Gelatinolytic activity of the conditioned mediacould be observed in a band at approximately 85–90kD in the AT-1, -2, and -3 cell lines (Fig. 6). The cell
Fig. 2. Northern blot analysis of mRNA for t-PA, u-PA, u-PAR,LRP, PAI-1, and PAI-2 in Dunning rat prostate carcinoma cells.mRNA was isolated as described in Materials and Methods, sub-jected to electrophoresis, and blotted. Total RNA (7.5 µg) wasused in each lane. Filters were hybridized with cDNA fragments ofu-PA, t-PA, u-PAR, LRP, PAI-1, and PAI-2. Hybridization with aGAPDH cDNA fragment was used as a control for loading equalamounts of RNA in the different lanes of the gel.
Fig. 3. Levels of t-PA and u-PA activity in tissue extracts oftumors of the Dunning AT-2, AT-3, MATLyLu, and MATLu cellsgrown in rats, expressed in mIU per µg protein. Tissue sampleswere analyzed in duplicate, and the data presented are mean ± SD.AT-3 and MATLyLu have significantly (P < 0.05) elevated levels oft-PA and u-PA.
200 Quax et al.
lines MATLu and MATLyLu showed one band withstrong gelatinolytic activity at a molecular weight ofapproximately 72 kD, probably active MMP-2, andone band at approximately 74–76 kD with lower ge-latinolytic activity, most likely pro-MMP-2. Thesebands were more prominent in the MATLu than in theMATLyLu cells.
DISCUSSION
As can be seen, u-PA is significantly elevated onlyin the metastasizing cell lines, including the low tomoderately metastasizing AT-2 cell line (Fig. 1). Sinceit is mainly found in cell extracts, u-PA is probablybound to its cell surface receptor u-PAR. All cell linesmake u-PAR mRNA (Fig. 2). The finding that in allcells except AT-1, LRP mRNA could be detected indi-cates that the receptor-bound u-PA can be rapidly in-ternalized [39]. t-PA activity and mRNA could only bedetected in HIF and MATLu. Concerning the inhibi-tors it can be said that PAI-1 and PAI-2 mRNA wasdetected in HIF and AT-3, whereas for PAI-1 mRNA,its absence in MATLu should be mentioned becausePAI-1 is frequently associated with an aggressive ma-lignant phenotype of a metastasizing tumor [3,5,6,27].This should also be mentioned because it may explainwhy comparable levels of t-PA activity (the observeddifference was not significant) were found for HIF andMATLu (Fig. 1), whereas HIF clearly has higher t-PAmRNA levels (Fig. 2).
Using the in vitro matrix degradation assay, it wasfound that only the highly metastatic cell lines AT-3,MATLu, and MATLyLu were able to degrade such amatrix substantially in a plasmin-dependent way (Fig.
4). This degradation was mediated by plasmin, sincein the presence of aprotinin, matrix degradation wasstrongly inhibited. The necessary conversion of plas-minogen to the active protease plasmin was mediatedby u-PA, since blocking of u-PA activity by specificantibodies also reduced matrix degradation, whereasantibodies against t-PA had no effect. A striking phe-nomenon is the observation that the MATLu cell line,which of the highly metastasizing cells has the lowestu-PA activity, shows the highest in vitro matrix deg-radation. A possible explanation for this phenomenonmight be found in the fact that MATLu does not pro-duce any inhibitor mRNA, and therefore u-PA medi-ated matrix degradation can proceed more efficiently.On the other hand, it is thought that the most aggres-sive tumors in vivo produce both u-PA and its inhibi-tors [3,5,6,27]. Prostate carcinomas in vivo have a highpreference for bone metastasis. u-PA-mediated plas-minogen activation might be involved in bone matrixdegradation in vitro [38], and therefore it might bethat u-PA-mediated matrix degradation, as we havefound here using an in vitro system, also plays a rolein the metastasis of prostate carcinoma in vivo.
The AT-3 and MATLyLu tumors in vivo producet-PA activity, whereas the cell lines do not, and in theMATLu tumor hardly any u-PA or t-PA activity canbe detected. The presence of t-PA activity in the tu-mors in vivo and not in the cell lines in vitro suggestsa role for t-PA in the metastatic spread of prostatecancer. A role for t-PA in prostate cancer was alsosuggested from a pilot experiment with a small num-ber of tissue extracts of human prostate cancers, inwhich we observed an increased level of t-PA, whencompared with benign hyperplastic prostate tissue
Fig. 4. Extracellular matrix degradation byDunning rat prostate carcinoma cells. Cells (105)were incubated for 24 hr on radiolabeled extra-cellular matrix produced by bovine smoothmuscle cells (SMC) in serum-containing mediumsupplemented with human plasminogen (0.14 µM)(control). Similar incubations were performed inthe presence of anti-t-PA IgG, anti-u-PA IgG, and100 IU/ml aprotinin. Concentrations of antibodiesused to inhibit activity of t-PA or u-PA were de-termined as described in Materials and Methods.Matrix degradation was measured by the releaseof radioactivity into the medium. 3H released bytumor cells was expressed as a percentage of to-tal 3H released by the cells and a subsequenttreatment of the matrix with 0.25% w/v trypsin (1hr, 37°C). The data present the mean ± SEM ofthree experiments. Matrix degradation by AT-3,MATLu, and MATLyLu was significantly higherthan with the other cell lines and could be inhib-ited by aprotinin and anti-u-PA (P < 0.05).
PAs and MMPs in Rat Prostate Cancer Cell Lines 201
(unpublished observations). The production of t-PA inprostate cancers in vivo was also observed in severalearlier studies [40,41]. Whether the observed differ-ence in u-PA production in vivo in AT-3 and MAT-LyLu on one hand and MATLu on the other handmight contribute to the difference in metastasis path-way (lung vs. lymph) [22] needs further study.
Analysis of the production of matrix metallopro-teinases showed that MMP-2 mRNA was present inHIF, AT-3, MATLu, and MATLyLu (Fig. 5), and thatactive MMP-2 is detectable in MATLu- and MAT-LyLu-conditioned medium (Fig. 6). A strong signal forMMP-3 mRNA could be observed in the MATLu aswell as AT-2 and AT-3 cells but not in MATLyLu. No
signal for MMP-3 could be detected in the gelatinzymogram; this is not surprising, since usually ittakes large amounts of MMP-3 to detect it on a gel-atin zymogram. MMP-9 could be observed for AT-1,AT-2, and AT-3 in the gelatin zymogram, althoughno mRNA for MMP-1, -7, and -9 could be detected.It should be noted that we used probes for humanMMPs on rat cells, and the lack of signal might bedue to a low affinity of human probes for rat MMPmRNAs. The presence of collagenase activity in vari-ous Dunning tumor extracts has been reported before[23]. However, no correlation with the metastatic phe-notype could be determined. An extensive character-ization of gelatin-degrading metalloproteinase activi-ties (both calcium-dependent and -independent) in theoriginal Dunning R-3327 tumor in nude mice revealedpredominant forms at 64, 71, and 76 kD, as well as at91 and 98 kD [42]. Of these activities we can observehere the equivalents of 71 and 76 kD in MATLu andMATLyLu cell lines, and of the 91-kD band in the ATcell lines. No major differences in the production ofTIMP-1 and TIMP-2 mRNA could be observed be-tween cell lines.
When comparing the data presented in this study itmay be observed that the data from the matrix degra-dation assays do not completely correlate with theranked order of production of u-PA, t-PA, or the vari-ous MMPs as measured in the various biochemicalassays, and that the best correlation with metastaticphenotype is found for the matrix degradation assaydata. It should be mentioned that in tumor cell migra-tion and metastasis the proteolytic processes occurringin the direct pericellular environment are important,and that regulation of this pericellular proteolytic bal-ance by cell surface receptors and inhibitors is difficult
Fig. 6. Gelatin zymogram analysis for MMPs in conditioned me-dium of Dunning rat prostate carcinoma cells. Aliquots of condi-tioned medium were analyzed by gelatin zymography as describedin Materials and Methods. Conditioned medium of human endo-thelial cells containing pro-MMP-9 (92 kD), pro-MMP-2 (72 kD),and activated MMP-2 (64 kD) was used as a marker.
Fig. 5. Northern blot analysis of RNA for MMP-2, MMP-3,TIMP-1, and TIMP-2 of Dunning rat prostate carcinoma cells. RNAwas isolated as described in Materials and Methods, subjected toelectrophoresis, and blotted. Total RNA (7.5 µg) was used in eachlane. Filters were hybridized with cDNA fragments of MMP-2,MMP-3, TIMP-1, and TIMP-2. Hybridization with a GAPDH cDNAfragment was used as a control for loading of equal amounts ofmRNA in the different lanes of the gel.
202 Quax et al.

to monitor using biochemical analysis of the proteo-lytic activity in cultured medium or cell extracts.
The data from these in vitro experiments show thatin different rat prostate cancer sublines, proteolyticactivity and u-PA-mediated extracellular matrix deg-radation correlate with their known metastatic behav-ior in vivo. The exact role and the interaction of pro-teases and their inhibitors in these cells in migration orinvasion processes remain to be resolved.
ACKNOWLEDGMENTS
This work was supported by the Dutch Cancer So-ciety and the Dutch Heart Foundation (grant M93001).
REFERENCES
1. Danø K, Andreasen PA, Grøndahl-Hansen J, Kristensen P,Nielsen LS, Skriver L: Plasminogen activators tissue degrada-tion and cancer. Adv Cancer Res 1985;44:139–264.
2. Stetler-Stevenson WG, Liotta LA, Kleiner DE: Role of matrixmetalloproteinases in tumor invasion and metastasis. FASEB J1993;7:1434–1441.
3. De Vries TJ, Quax PHA, Denijn M, Verrijp KN, Verheijen JH,Verspaget HW, Weidle UH, Ruiter DJ, van Muijen GNP: Plas-minogen activators, their inhibitors, and urokinase receptoremerge in late stages of melanocytic tumor progression. Am JPathol 1994;144:70–81.
4. Duffy MJ, Reilly D, O’Sullivan C, O’Higgins N, Jennelly JJ, An-dreasen P: Urokinase-plasminogen activator, a new and inde-pendent prognostic marker in breast cancer. Cancer Res 1990;50:6827–6829.
5. Sier CFM, Vloedgraven HJM, Ganesh S, Griffioen G, Quax PHA,Verheijen JH, Dooijewaard G, Welvaart K, Vandevelde CJH,Lamers CBHW, Verspaget HW: Inactive urokinase and in-creased levels of its inhibitor type 1 in colorectal cancer livermetastasis. Gastroenterology 1994;107:1449–1456.
6. Janicke F, Schmitt M, Pache L, Ulm K, Harbeck N, Hofler H,Graeff H: Urokinase (uPA) and its inhibitor PAI-1 are strongand independent prognostic factors in node-negative breastcancer. Breast Cancer Res Treat 1993;24:195–208.
7. Ossowski L, Reich E: Antibodies to plasminogen activator in-hibit human tumor metastasis. Cell 1983;35:611–619.
8. Mignatti P, Robbins E, Rifkin DB: Tumor invasion through thehuman amniotic membrane: Requirement for a proteinase cas-cade. Cell 1986;47:487–498.
9. Tryggvason K, Hoyhtya M, Salo T: Proteolytic degradation ofextracellular matrix in tumor invasion. Biochim Biophys Acta1987;907:191–217.
10. Cottam DW, Rees RC: Regulation of matrix metalloprotein-ases—Their role in tumor invasion and metastasis—Review. IntJ Oncol 1993;2:861–872.
11. McDonnell S, Wright JH, Gaire M, Matrisian LM: Expressionand regulation of stromelysin and matrilysin by growth factorsand oncogenes. Biochem Soc Trans 1994;22:58–63.
12. Sato H, Kida Y, Mai M, Endo Y, Sasaki T, Tanaka J, Seiki M:Expression of genes encoding type-IV collagen-degrading me-talloproteinases and tissue inhibitors of metalloproteinases invarious human tumor cells. Oncogene 1992;7:77–83.
13. Quax PHA, van Leeuwen RTJ, Verspaget HW, Verheijen JH:Protein and messenger RNA levels of plasminogen activators
and inhibitors analyzed in 22 human tumor cell lines. CancerRes 1990;50:1488–1494.
14. Quax PHA, van Muijen GNP, Weening-Verhoeff EJD, Lund LR,Danø K, Ruiter DJ, Verheijen JH: Metastatic behavior of humanmelanoma cell lines in nude mice correlates with urokinase-typeplasminogen activator, its type-1 inhibitor, and urokinase-mediated matrix degradation. J Cell Biol 1991;115:191–199.
15. Wilson MJ: Proteases in prostate development, function andpathology. Microsc Res Techn 1995;30:305–318.
16. Keer HN, Gaylis FD, Kozlowski JM, Kwaan HC, Bauer KD,Sinha AA, Wilson MJ: Heterogeneity in plasminogen activator(PA) levels in human prostate cancer cell lines: Increased PAactivity correlates with biologically aggressive behavior. Pros-tate 1991;18:201–214.
17. Wilson MJ, Sinha AA: Plasminogen activator and metalloprote-ase activities of Du-145, PC-3 and 1-LN-PC-3-1A human pros-tate tumors grown in nude mice—Correlation with tumor inva-sive behavior. Cell Mol Biol Res 1993;39:751–760.
18. Crowley CW, Cohen RL, Lucas BK, Liu GH, Shuman MA: Pre-vention of metastasis by inhibition of the urokinase receptor.Proc Natl Acad Sci USA 1993;90:5021–5025.
19. Hoosein NM, Boyd D, Hollas W, Mazar A, Henkin J, ChungLWK: Involvement of urokinase and its receptor in the inva-siveness of human prostatic carcinoma cell lines. Cancer Com1991;3:255–264.
20. Jarrard DF, Hansen NM, Patai B, Ruksalis DB: Urokinase plas-minogen activator is necessary but not sufficient for prostatecancer cell invasion. Invasion Metastasis 1995;15:34–45.
21. Dunning WF: Model systems for the study of prostatic cancer.NCI Monogr 1963;12:351–369.
22. Isaacs JT, Isaacs WB, Feitz WFJ, Scheres J: Establishment andcharacterization of seven Dunning rat prostatic cancer cell linesand their use in developing methods for predicting metastaticabilities of prostatic cancers. Prostate 1986;9:261–281.
23. Lowe FC, Isaacs JT: Biochemical methods for predicting meta-static ability of prostatic cancer utilizing the Dunning R-3327 ratprostatic adenocarcinoma system as a model. Cancer Res 1984;44:744–752.
24. Bussemakers MJG, van Moorselaar RJA, Giroldi LA, IchikawaT, Isaacs JT, Takeichi M, Debruyne FMJ, Schalken JA: Decreasedexpression of E-cadherin in the progression of rat prostatic can-cer. Cancer Res 1992;52:2916–2922.
25. Schalken JA, Ebeling SB, Isaacs JT, Treiger B, Bussemakers MJG,DeJong MEM, van de Ven WJM: Down modulation of fibronec-tin messenger RNA in metastasizing rat prostatic cancer cellsrevealed by differential hybridization analysis. Cancer Res 1988;48:2042–2046.
26. Hierowski MT, Schally AV: An inhibitor of urokinase and tissueplasminogen activators in Dunning R3327H prostate tumors ofrats treated with D-Trp6-LH-RH. Horm Res 1985;21:124–135.
27. Sier CFM, Verspaget HW, Griffioen G, Verheijen JH, Quax PHA,Dooijewaard G, De Bruin PAF, Lamers CBHW: Imbalance ofplasminogen activators and their inhibitors in human colorectalneoplasia: Implications of urokinase in colorectal carcinogene-sis. Gastroenterology 1991;101:1522–1528.
28. Granelli-Piperno A, Reich E: A study of proteases and protease-inhibitor complexes in biological fluids. J Exp Med 1978;148:223–234.
29. Verheijen JH, Mullaart E, Chang GTG, Kluft C, Wijngaards G: Asimple, sensitive spectrophotometric assay for extrinsic (tissue-type) plasminogen activator applicable to measurements inplasma. Thromb Haemost 1982;48:266–269.
30. Rijken DC, Wijngaards G, Welbergen J: Immunological charac-terization of plasminogen activator activities in human tissuesand body fluids. Lab Clin Med 1981;97:477–486.
PAs and MMPs in Rat Prostate Cancer Cell Lines 203

31. Binnema DJ, van Iersel JJL, Dooijewaard G: Quantitation of uro-kinase antigen in plasma and culture media by use of an ELISA.Thromb Res 1986;43:569–577.
32. Gaffney PJ, Heath AB: A collaborative study to establish a stan-dard for high molecular weight urinary-type plasminogen acti-vator (HMW/u-PA). Thromb Haemost 1990;64:398–401.
33. Gaffney PJ, Curtis AD: A collaborative study of a proposedinternational standard for tissue plasminogen activator (t-PA).Thromb Haemost 1985;53:134–136.
34. Hanemaaijer R, Koolwijk P, Leclercq L, Devree WJA, van Hins-bergh VWM: Regulation of matrix metalloproteinase expressionin human vein and microvascular endothelial cells—Effects oftumour necrosis factor-alpha, interleukin-1 and phorbol ester.Biochem J 1993;296:803–809.
35. Chomczynski P, Sacchi N: Single-step method of RNA isolationby acid guanidnium thiocyanate-phenol-chloroform extraction.Anal Biochem 1987;162:156–159.
36. Zeheb R, Gelehrter TD: Cloning and sequencing of cDNA forthe rat plasminogen activator inhibitor-1. Gene 1988;73:459–468.
37. Belin D, Wohlwend A, Schleuning WD, Kruithof EKO, VassalliJD: Facultative polypeptide translocation allows a single mes-senger RNA to encode the secreted and cytosolic forms of plas-minogen activators inhibitor-2. EMBO J 1989;8:3287–3294.
38. De Bart ACW, Quax PHA, Lowik CWGM, Verheijen JH: Regu-lation of plasminogen activation, matrix metalloproteinases andurokinase-type plasminogen activator-mediated extracellularmatrix degradation in human osteosarcoma cell line MG63 byinterleukin-1 alpha. J Bone Miner Res 1995;10:1374–1384.
39. Nykjaer A, Petersen CM, Moller B, Jensen PH, Moestrup SK,Holtet TL, Etzerodt M, Thogersen HC, Munch M, AndreasenPA, Gliemann J: Purified a2-macroglobulin receptor/LDL re-ceptor-related protein binds urokinase activator inhibitor type-1complex—Evidence that the a2-macroglobulin receptor medi-ates cellular degradation of urokinase receptor-bound com-plexes. J Biol Chem 1992;267:14543–14546.
40. Kirchheimer JC, Koller A, Binder BR: Isolation and character-ization of plasminogen activators from hyperplastic and malig-nant prostate tissue. Biochim Biophys Acta 1984;797:256–265.
41. Kirchheimer JC, Pfluger H, Ritschl P, Hienert G, Binder BR:Plasminogen activator activity in bone metastasis of prostaticcarcinomas as compared to primary tumors. Invasion Metasta-sis 1985;5:344–355.
42. Wilson MJ, Kapoor S, Vogel MM, Sinha AA: Characterization ofgelatin degrading metalloproteinase activities of the Dunningrat prostate tumor grown in nude mice. Prostate 1991;19:237–250.
204 Quax et al.





























