Embed Size (px)
Citation preview

Physical Properties of a Genomic CondensedChromatin Fragment
Rodolfo Ghirlando†, Michael D. Litt†, Marie-Noelle PrioleauFelix Recillas-Targa and Gary Felsenfeld*
Department of Health andHuman Services, Laboratory ofMolecular Biology, NationalInstitute of Diabetes, Digestiveand Kidney Diseases, NationalInstitutes of Health, BethesdaMD 20892, USA
We have studied the physical properties of a segment of condensed chro-matin that lies upstream of the chicken b-globin locus. This segment canbe excised from an avian erythroleukemia cell line by restriction enzymedigestion and released from the nucleus as an essentially homogeneousfragment about 15.5 kbp long. Because of this homogeneity we couldmeasure its sedimentation coefficient quite accurately by a combinationof sucrose gradient and analytical ultracentrifugation. By measuringadditionally the buoyant density of the cross-linked particle in CsCl wecould deduce the total mass of the particle, hence its frictional coefficient,f ; directly related to its shape. The measured value of f is consistent witha rod-like particle of the approximate length and diameter proposed ear-lier for the 30 nm chromatin fiber. The method is generally applicable tohomogeneous particles of unique sequence at genomic abundance.
Published by Elsevier Ltd.
Keywords: chromatin structure; condensed chromatin; heterochromatin;hydrodynamics; physical genomics*Corresponding author
Introduction
Within the eukaryotic nucleus, the structure oflocal chromatin domains that contain genes reflectsthe state of local transcriptional activity. Transcrip-tionally active chromatin, although largely stillpackaged in nucleosomes, has a more open, nucle-ase-accessible structure marked by numeroushistone modifications that may serve to recruit acti-vating complexes as well as to maintain the partialdisruption of the chromatin higher order folding.Except for housekeeping genes, this activity andthe associated chromatin modifications are specificto certain cell types or developmental stages. How-ever, in vertebrates a large part of the genome iscomposed of non-coding sequences, including
extended regions of repetitive DNA that have nolong open reading frames and are, so far as weknow, almost never transcribed. These are pack-aged in condensed structures in which the nucleo-somes assume some kind of more closely packedorganization, which has been given the descriptivename “constitutive heterochromatin”.
A possibly distinct kind of chromatin is foundnear centromeres, and has been particularly wellcharacterized in Drosophila. The properties ofthis pericentromeric heterochromatin include theability to silence genes placed nearby, presumablyby an extension of the condensed chromatin struc-ture. This frequently is associated with epigeneti-cally stable variegation of expression (positioneffect variegation). Pericentromeric heterochroma-tin has served as a model for all heterochromaticstructures, but its relationship to the constitutiveheterochromatin of vertebrates, scattered through-out the genome, is unclear.1 – 3
This issue is further complicated by uncertaintiesabout the nature of higher order chromatin struc-ture. DNA is packaged in chromatin as a poly-nucleosome string. Each nucleosome contains anoctamer of core histones about which are wrapped165 bp of DNA in two superhelical turns; thenucleosomes are connected by short stretches oflinker DNA. Linker histone (histone H1 or H5) is
0022-2836/$ - see front matter Published by Elsevier Ltd.
† R.G. and M.D.L. contributed equally to this work.Present addresses: M.-N. Priouleau, Laboratoire de
Genetique Moleculaire, CNRS URA 1302, 46 Rue d’Ulm,75230 Paris Cedex 05, France; F. Recillas-Targa, Institutode Fisiologıa Celular, Universidad Nacional Autonomade Mexico, Departamento de Genetica Molecular,Apartado Postal 70-242, Mexico, DF.
E-mail address of the corresponding author:[email protected]
Abbreviations used: bp, base pairs; MNase,micrococcal nuclease.
doi:10.1016/j.jmb.2003.12.049 J. Mol. Biol. (2004) 336, 597–605

bound to the outside of the nucleosome and to thelinker. Under physiological ionic conditions, acompact fiber structure is formed with a diameterof about 30 nm. The external appearance of thisfiber under many experimental conditions suggestsa superhelical array of nucleosome monomers,with a repeat of about six nucleosomes per turn,each turn occupying about 11 nm along the helicalaxis. The way in which this compact structure isformed, and in particular the path of the linkerDNA between successive nucleosomes within the30 nm fiber, has been debated.4 – 11 Furthermore,although idealized models and many electronmicrographs of short segments appear to depict aregular rod-like structure, there has been somequestion about the regularity and rigidity of suchstructures in solution and within the nucleus. Thisis a difficult question to address because mucheukaryotic chromatin is interspersed with regulat-ory sites that bind non-histone proteins and couldlead to breaks in an otherwise uniform structure.
We have shown earlier that the region 50 of thechicken b-globin locus contains an approximately16 kbp region of condensed chromatin, shownhere to be closer in average value to 15.5 kbp (thelatter is the length used throughout this paper).This region is marked by high levels of DNA meth-ylation, low levels of histone H3 and H4 acety-lation and H3 K4 methylation, and high levels ofH3 K9 methylation (Figure 1).12,13 These are allmarkers of condensed chromatin. The DNA itselfincludes multiple copies of the chicken CR1 repeat,and it has no long open reading frame. This regionoffered the opportunity to study an extendedregion of condensed chromatin structure, possiblyfree of interspersed non-histone binding sites. Wedescribe here the application of precise methodsof sedimentation and buoyant density measure-ment to obtain a detailed picture of the hydro-dynamic properties, and consequently the shape,of a chromatin fragment containing this region.
Results
Hydrodynamic analysis
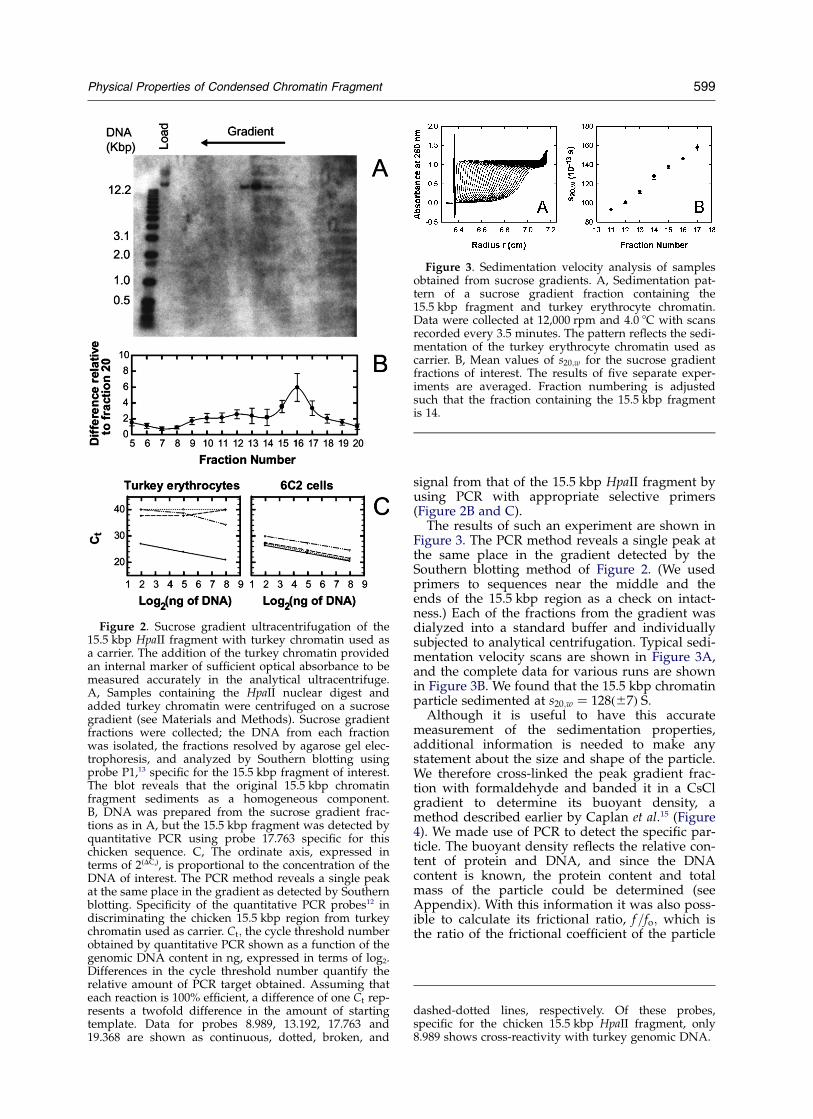
An earlier study of this 15.5 kbp region exam-ined the distribution of methylated CpG sites
resistant to cleavage by the restriction enzymeHpaII. All of these sites were highly resistant, indi-cating that they were also highly methylated. Atthe margins of the region, however, there were sev-eral sites that were unmethylated and could be cutby HpaII.13 We took advantage of this pattern ofmethylation to excise the 15.5 kbp element. Studiesin other laboratories as well as our own had shownthat chromatin fragments of moderate size couldbe liberated from nuclei by digestion withnucleases.14 – 17 We prepared nuclei from chicken6C2 cells, digested with HpaII, and dialyzed into abuffer that liberated digestion products from thenuclei. In the first experiments, the digest wasthen subjected to sucrose gradient sedimentation,and DNA was purified from the fractions collected.The individual fractions were electrophoresed onan agarose gel and blotted to a membrane, whichwas then hybridized to a radioactively labeledprobe targeted to a unique sequence within thecondensed chromatin region. As can be seen inFigure 2A, the fragment sediments largely in asingle peak with a large sedimentation coefficient.The size of the DNA was determined precisely bypulsed field agarose gel electrophoresis (data notshown), which revealed a nearly monodisperseDNA fragment, sometimes resolving into a closelyspaced pair, with mean value of approximately15.5 kbp, consistent with the known sequence inthis region. The slight polydispersity in DNA sizearises because there are clusters of unmethylatedHpaII sites at the 30 end of the fragment, and theremay be some variation in the accessibility of DNAin this region.
In order to draw conclusions about the physicalproperties of this particle it is necessary to makequantitative measurements of its sedimentationcoefficient. First, we established PCR methods forfollowing the particle on sucrose gradients, allow-ing smaller volumes of sample to be layered onthe gradient and to be used in detection (Figure2B). In principle it should be possible to transferthe individual sucrose gradient fractions to theanalytical ultracentrifuge and determine the sedi-mentation properties of the bulk chromatin con-tained in each fraction. This proved difficultbecause the yield of large (.10 kbp) and solublechromatin fragments from the HpaII digestionprocedure is low, preventing accurate opticalabsorbance measurements in the analytical ultra-centrifuge, unless large numbers of cells are used.We therefore prepared a larger quantity of “bulk”chromatin fragments by mild micrococcal nuclease(MNase) digestion of turkey adult red cell nuclei,and added this to the HpaII chicken 6C2 nucleardigest as carrier before loading the gradient. Wechose turkey erythrocyte nuclei as the source ofthis chromatin because the use of chicken erythro-cyte nuclei would have raised unacceptably thebackground level of randomly cut fragments fromthe 15.5 kbp region. We found that although thereappears to be cross-hybridizing material in the tur-key erythrocyte nucleus, we could distinguish this
Figure 1. Diagram of the region at the 50 end of thechicken b-globin locus, showing the 15.5 kbp condensedchromatin region that separates the upstream folatereceptor gene from the downstream b-globin locus con-trol elements and gene cluster. (Adapted from Prioleauet al.13.)
598 Physical Properties of Condensed Chromatin Fragment
signal from that of the 15.5 kbp HpaII fragment byusing PCR with appropriate selective primers(Figure 2B and C).
The results of such an experiment are shown inFigure 3. The PCR method reveals a single peak atthe same place in the gradient detected by theSouthern blotting method of Figure 2. (We usedprimers to sequences near the middle and theends of the 15.5 kbp region as a check on intact-ness.) Each of the fractions from the gradient wasdialyzed into a standard buffer and individuallysubjected to analytical centrifugation. Typical sedi-mentation velocity scans are shown in Figure 3A,and the complete data for various runs are shownin Figure 3B. We found that the 15.5 kbp chromatinparticle sedimented at s20;w ¼ 128ð^7Þ S:
Although it is useful to have this accuratemeasurement of the sedimentation properties,additional information is needed to make anystatement about the size and shape of the particle.We therefore cross-linked the peak gradient frac-tion with formaldehyde and banded it in a CsClgradient to determine its buoyant density, amethod described earlier by Caplan et al.15 (Figure4). We made use of PCR to detect the specific par-ticle. The buoyant density reflects the relative con-tent of protein and DNA, and since the DNAcontent is known, the protein content and totalmass of the particle could be determined (seeAppendix). With this information it was also poss-ible to calculate its frictional ratio, f=fo; which isthe ratio of the frictional coefficient of the particle
Figure 2. Sucrose gradient ultracentrifugation of the15.5 kbp HpaII fragment with turkey chromatin used asa carrier. The addition of the turkey chromatin providedan internal marker of sufficient optical absorbance to bemeasured accurately in the analytical ultracentrifuge.A, Samples containing the HpaII nuclear digest andadded turkey chromatin were centrifuged on a sucrosegradient (see Materials and Methods). Sucrose gradientfractions were collected; the DNA from each fractionwas isolated, the fractions resolved by agarose gel elec-trophoresis, and analyzed by Southern blotting usingprobe P1,13 specific for the 15.5 kbp fragment of interest.The blot reveals that the original 15.5 kbp chromatinfragment sediments as a homogeneous component.B, DNA was prepared from the sucrose gradient frac-tions as in A, but the 15.5 kbp fragment was detected byquantitative PCR using probe 17.763 specific for thischicken sequence. C, The ordinate axis, expressed interms of 2(DCt), is proportional to the concentration of theDNA of interest. The PCR method reveals a single peakat the same place in the gradient as detected by Southernblotting. Specificity of the quantitative PCR probes12 indiscriminating the chicken 15.5 kbp region from turkeychromatin used as carrier. Ct; the cycle threshold numberobtained by quantitative PCR shown as a function of thegenomic DNA content in ng, expressed in terms of log2.Differences in the cycle threshold number quantify therelative amount of PCR target obtained. Assuming thateach reaction is 100% efficient, a difference of one Ct rep-resents a twofold difference in the amount of startingtemplate. Data for probes 8.989, 13.192, 17.763 and19.368 are shown as continuous, dotted, broken, and
dashed-dotted lines, respectively. Of these probes,specific for the chicken 15.5 kbp HpaII fragment, only8.989 shows cross-reactivity with turkey genomic DNA.
Figure 3. Sedimentation velocity analysis of samplesobtained from sucrose gradients. A, Sedimentation pat-tern of a sucrose gradient fraction containing the15.5 kbp fragment and turkey erythrocyte chromatin.Data were collected at 12,000 rpm and 4.0 8C with scansrecorded every 3.5 minutes. The pattern reflects the sedi-mentation of the turkey erythrocyte chromatin used ascarrier. B, Mean values of s20;w for the sucrose gradientfractions of interest. The results of five separate exper-iments are averaged. Fraction numbering is adjustedsuch that the fraction containing the 15.5 kbp fragmentis 14.
Physical Properties of Condensed Chromatin Fragment 599
(determined from s20;w) to the frictional coefficientof a sphere of the same mass (Table 1).
Comparison with hydrodynamic theory
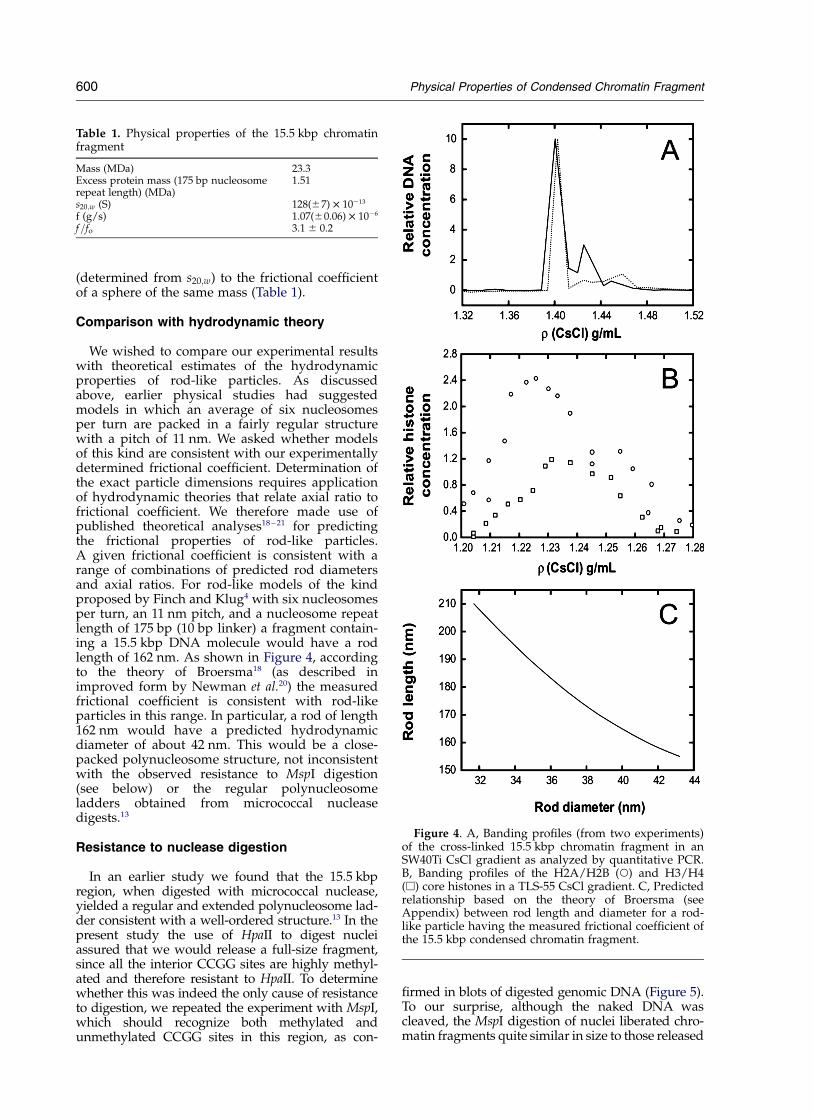
We wished to compare our experimental resultswith theoretical estimates of the hydrodynamicproperties of rod-like particles. As discussedabove, earlier physical studies had suggestedmodels in which an average of six nucleosomesper turn are packed in a fairly regular structurewith a pitch of 11 nm. We asked whether modelsof this kind are consistent with our experimentallydetermined frictional coefficient. Determination ofthe exact particle dimensions requires applicationof hydrodynamic theories that relate axial ratio tofrictional coefficient. We therefore made use ofpublished theoretical analyses18 – 21 for predictingthe frictional properties of rod-like particles.A given frictional coefficient is consistent with arange of combinations of predicted rod diametersand axial ratios. For rod-like models of the kindproposed by Finch and Klug4 with six nucleosomesper turn, an 11 nm pitch, and a nucleosome repeatlength of 175 bp (10 bp linker) a fragment contain-ing a 15.5 kbp DNA molecule would have a rodlength of 162 nm. As shown in Figure 4, accordingto the theory of Broersma18 (as described inimproved form by Newman et al.20) the measuredfrictional coefficient is consistent with rod-likeparticles in this range. In particular, a rod of length162 nm would have a predicted hydrodynamicdiameter of about 42 nm. This would be a close-packed polynucleosome structure, not inconsistentwith the observed resistance to MspI digestion(see below) or the regular polynucleosomeladders obtained from micrococcal nucleasedigests.13
Resistance to nuclease digestion
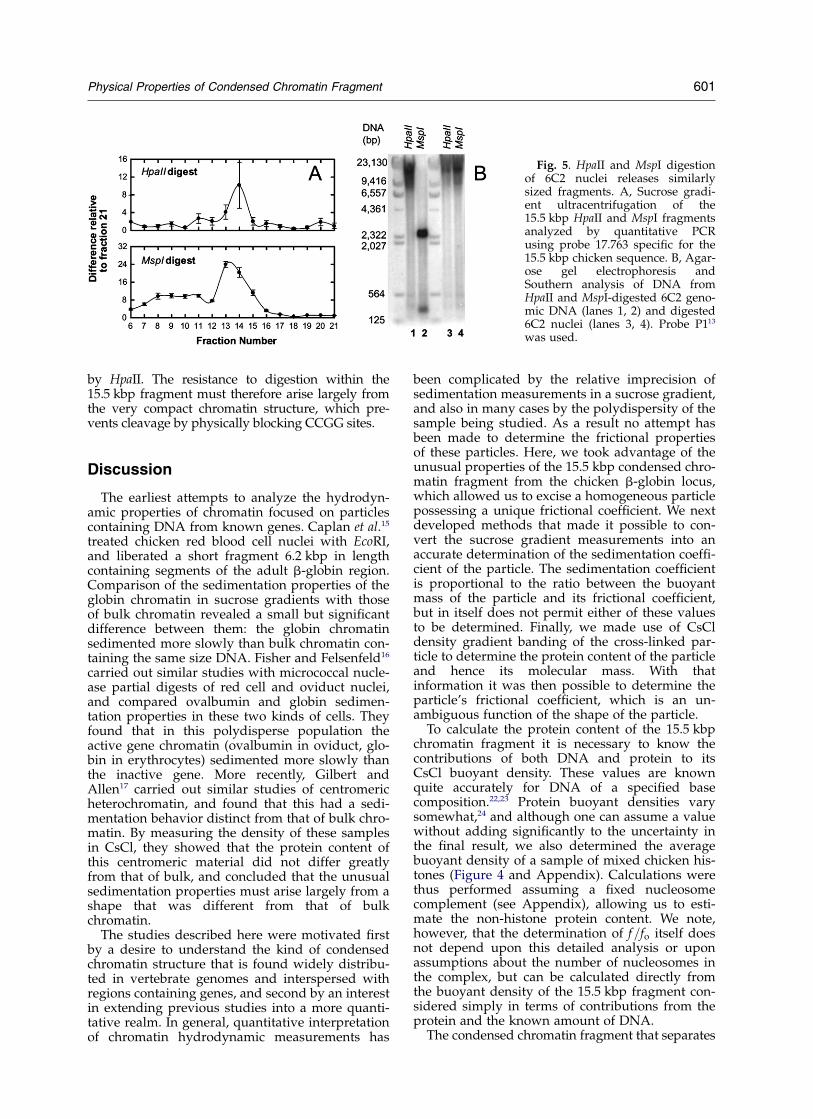
In an earlier study we found that the 15.5 kbpregion, when digested with micrococcal nuclease,yielded a regular and extended polynucleosome lad-der consistent with a well-ordered structure.13 In thepresent study the use of HpaII to digest nucleiassured that we would release a full-size fragment,since all the interior CCGG sites are highly methyl-ated and therefore resistant to HpaII. To determinewhether this was indeed the only cause of resistanceto digestion, we repeated the experiment with MspI,which should recognize both methylated andunmethylated CCGG sites in this region, as con-
firmed in blots of digested genomic DNA (Figure 5).To our surprise, although the naked DNA wascleaved, the MspI digestion of nuclei liberated chro-matin fragments quite similar in size to those released
Figure 4. A, Banding profiles (from two experiments)of the cross-linked 15.5 kbp chromatin fragment in anSW40Ti CsCl gradient as analyzed by quantitative PCR.B, Banding profiles of the H2A/H2B (W) and H3/H4(A) core histones in a TLS-55 CsCl gradient. C, Predictedrelationship based on the theory of Broersma (seeAppendix) between rod length and diameter for a rod-like particle having the measured frictional coefficient ofthe 15.5 kbp condensed chromatin fragment.
Table 1. Physical properties of the 15.5 kbp chromatinfragment
Mass (MDa) 23.3Excess protein mass (175 bp nucleosomerepeat length) (MDa)
1.51
s20;w (S) 128(^7) £ 10213
f (g/s) 1.07(^0.06) £ 1026
f=fo 3.1 ^ 0.2
600 Physical Properties of Condensed Chromatin Fragment
by HpaII. The resistance to digestion within the15.5 kbp fragment must therefore arise largely fromthe very compact chromatin structure, which pre-vents cleavage by physically blocking CCGG sites.
Discussion
The earliest attempts to analyze the hydrodyn-amic properties of chromatin focused on particlescontaining DNA from known genes. Caplan et al.15
treated chicken red blood cell nuclei with EcoRI,and liberated a short fragment 6.2 kbp in lengthcontaining segments of the adult b-globin region.Comparison of the sedimentation properties of theglobin chromatin in sucrose gradients with thoseof bulk chromatin revealed a small but significantdifference between them: the globin chromatinsedimented more slowly than bulk chromatin con-taining the same size DNA. Fisher and Felsenfeld16
carried out similar studies with micrococcal nucle-ase partial digests of red cell and oviduct nuclei,and compared ovalbumin and globin sedimen-tation properties in these two kinds of cells. Theyfound that in this polydisperse population theactive gene chromatin (ovalbumin in oviduct, glo-bin in erythrocytes) sedimented more slowly thanthe inactive gene. More recently, Gilbert andAllen17 carried out similar studies of centromericheterochromatin, and found that this had a sedi-mentation behavior distinct from that of bulk chro-matin. By measuring the density of these samplesin CsCl, they showed that the protein content ofthis centromeric material did not differ greatlyfrom that of bulk, and concluded that the unusualsedimentation properties must arise largely from ashape that was different from that of bulkchromatin.
The studies described here were motivated firstby a desire to understand the kind of condensedchromatin structure that is found widely distribu-ted in vertebrate genomes and interspersed withregions containing genes, and second by an interestin extending previous studies into a more quanti-tative realm. In general, quantitative interpretationof chromatin hydrodynamic measurements has
been complicated by the relative imprecision ofsedimentation measurements in a sucrose gradient,and also in many cases by the polydispersity of thesample being studied. As a result no attempt hasbeen made to determine the frictional propertiesof these particles. Here, we took advantage of theunusual properties of the 15.5 kbp condensed chro-matin fragment from the chicken b-globin locus,which allowed us to excise a homogeneous particlepossessing a unique frictional coefficient. We nextdeveloped methods that made it possible to con-vert the sucrose gradient measurements into anaccurate determination of the sedimentation coeffi-cient of the particle. The sedimentation coefficientis proportional to the ratio between the buoyantmass of the particle and its frictional coefficient,but in itself does not permit either of these valuesto be determined. Finally, we made use of CsCldensity gradient banding of the cross-linked par-ticle to determine the protein content of the particleand hence its molecular mass. With thatinformation it was then possible to determine theparticle’s frictional coefficient, which is an un-ambiguous function of the shape of the particle.
To calculate the protein content of the 15.5 kbpchromatin fragment it is necessary to know thecontributions of both DNA and protein to itsCsCl buoyant density. These values are knownquite accurately for DNA of a specified basecomposition.22,23 Protein buoyant densities varysomewhat,24 and although one can assume a valuewithout adding significantly to the uncertainty inthe final result, we also determined the averagebuoyant density of a sample of mixed chicken his-tones (Figure 4 and Appendix). Calculations werethus performed assuming a fixed nucleosomecomplement (see Appendix), allowing us to esti-mate the non-histone protein content. We note,however, that the determination of f=fo itself doesnot depend upon this detailed analysis or uponassumptions about the number of nucleosomes inthe complex, but can be calculated directly fromthe buoyant density of the 15.5 kbp fragment con-sidered simply in terms of contributions from theprotein and the known amount of DNA.
The condensed chromatin fragment that separates
Fig. 5. HpaII and MspI digestionof 6C2 nuclei releases similarlysized fragments. A, Sucrose gradi-ent ultracentrifugation of the15.5 kbp HpaII and MspI fragmentsanalyzed by quantitative PCRusing probe 17.763 specific for the15.5 kbp chicken sequence. B, Agar-ose gel electrophoresis andSouthern analysis of DNA fromHpaII and MspI-digested 6C2 geno-mic DNA (lanes 1, 2) and digested6C2 nuclei (lanes 3, 4). Probe P113
was used.
Physical Properties of Condensed Chromatin Fragment 601

the chicken folate receptor gene from the b-globinlocus has a measured frictional coefficient of1.07(^0.06) £ 1026 g/s. Its frictional ratio, the ratioof this coefficient to the coefficient calculated for aspherical particle of the same mass, is 3.1(^0.2).This frictional ratio corresponds to an elongated par-ticle with a very high axial ratio. Comparison with atheory of the hydrodynamics of rod-like moleculesshows that this ratio is consistent with a rod about170 nm long and with a diameter of about 40 nm.Although numbers obtained from this theory areonly approximate, they are clearly consistent withestimates that have been made by other methods.
The predicted dimensions of the particle alsoinclude a contribution from the non-histone pro-tein component. However, even if this proteinwere organized as a uniform layer coating thecylinder, it could contribute only about 1 nm tothe overall cylindrical diameter. The identity ofthis protein remains to be determined by othermethods. Although our results are consistent withthe model proposed by Finch and Klug4 andothers,5 – 10 namely a continuously wound poly-nucleosome superhelix, they would also fit othermodels with similar values of mass per unit lengthbut more complicated paths of the polynucleosomebackbone,11 or with a somewhat irregular numberof nucleosomes per turn. The data and the analysisdo however support the idea that the 15.5 kbp frag-ment of condensed chromatin is an extended rod-like structure.
The experimental approach described here dif-fers from earlier approaches in three respects.First, we used a chromatin particle of definedDNA size. The determination of the molecularmass and therefore the frictional coefficient of theparticle based on CsCl buoyant density (Figure 4)depends on this information, which is not availablewhen studying polydisperse samples.16,17 Second,we developed internal calibration methods thatallowed us to measure precisely the sedimentationcoefficient of the particle. Third, we studied a frag-ment of considerably greater length than that avail-able in one earlier study of transcriptionally activechromatin14,15 which did make use of a 6.2 kbprestriction fragment from the coding region of theb-globin locus. The latter study concluded thatthis fragment sedimented more slowly than bulk(mostly inactive) chromatin with the same sizeDNA, and attributed this to the presence of abreak, possibly at a hypersensitive site, in an other-wise condensed structure. A short particle withsuch a break is best approximated as a pair ofspheres connected by a small link,19 and its struc-ture is clearly different from that assumed by longstretches of condensed chromatin.
The 15.5 kbp condensed chromatin region thatwe describe here contains many repeat sequences(CR1 repeats) and no long open reading frames orhypersensitive sites. It therefore provided an excel-lent opportunity to determine whether such a“heterochromatic” sequence in fact exists as anextended chromatin fiber, rather than as a col-
lapsed random coil. Our results provide evidencethat it does assume the extended form, andstrongly suggest that other condensed chromatinregions dispersed throughout the genome willhave similar properties, affecting the overall rigidityand accessibility of the regions in which they areembedded. The methods described here are appli-cable to other domains, and should open the way todetailed studies of the physical properties of definedchromatin fragments and organized chromatin com-plexes that can be liberated from the nucleus.
Materials and Methods
Hpa II digestion of 6C2 nuclei
Nuclei were isolated from six T150 dishes of 6C2 cellsas described.13 The nuclei were washed at least twice inNEBuffer no. 2 (50 mM NaCl, 10 mM Tris–HCl, 10 mMMgCl2, 1 mM dithiothreitol (pH 7.9 at 25 8C)) and resus-pended in NEBuffer no. 2 to an A260 nm of 100. The nucleiwere then digested with five units/ml of HpaII for fourhours at 37 8C. Reactions were stopped by adding 1/3volume of 50 mM EDTA. In order to facilitate the releaseof large chromatin fragments, the digested nuclei werepelleted in a microcentrifuge for seven minutes at1200 g, resuspended in 0.5 ml of buffer A (80 mM KCl,10 mM NaCl, 0.5 mM EDTA, 0.1 mM EGTA, 0.5 mMspermidine, 0.15 mM spermine, 15 mM 2-mercaptoetha-nol, 15 mM Tris–HCl (pH 8.0), 0.5% Brijw 58) and incu-bated for ten minutes at 4 8C. A half volume of 2 M ureawas added and nuclei were incubated at 4 8C for twohours. The addition of urea removes the nuclear envel-ope while preserving the native chromatin morphology.25
The addition of urea did not alter the sedimentationproperties of the 15.5 kbp chromatin fragment in anyway (data not shown). Prior to fractionation on a sucrosegradient, nuclei were dialyzed overnight at 4 8C in DBbuffer (80 mM NaCl, 10 mM Tris–HCl (pH 7.4), 0.2 mMEDTA) and pelleted at 16,000 g for ten minutes at 4 8Cin a microcentrifuge. The supernatant containing the sol-uble chromatin was retained for further analysis. Notethat all solutions, except those used for sedimentationvelocity, contain the following protease inhibitors:0.75 mg/ml of pepstatin A, 0.50 mg/ml of leupeptin,1.0 mg/ml of aprotinin and 0.2 mM PMSF.
MNase digestion of turkey erythrocyte nuclei
Nuclei were isolated from 10 ml of adult turkey redblood cells as described.13 The nuclei were washedtwice in lysis buffer (10 mM Tris–HCl (pH 7.4), 50 mMNaCl, 3 mM MgCl2 0.4% NP-40) containing 1 mMCaCl2, and then resuspended to an A260 nm of 100 in lysisbuffer þ 1 mM CaCl2. Nuclei were digested with3.8 £ 1023 units of MNase/ml for ten minutes at 37 8C.Reactions were stopped by adding 1/3 volume of50 mM EDTA. Soluble chromatin was prepared asdescribed above.
Fractionation of the 15.5 kbp chromatin fragment
Gradients of 5%–30% (w/v) sucrose in DB bufferwere poured in SW40 14 mm £ 95 mm centrifuge tubeswith a BioComp Gradient Master 107 using the followingparameters: 35 s at an angle of 868 and 28,000 rpm
602 Physical Properties of Condensed Chromatin Fragment

(program long, sucrose, 5–30% (w/v), first). Then, 400–500 ml of digested chromatin were layered on the gradi-ent for separation. In experiments in which sampleswere analyzed on Southern blots or used for formal-dehyde cross-linking, 400–500 ml of 6C2 HpaII-digestedchromatin were used. In the case of experiments inwhich the sedimentation coefficient of fractions of inter-est were measured, 100 ml of 6C2 HpaII-digested chro-matin was mixed with 300 ml of MNase-digested turkeyerythrocyte chromatin and layered on top of the gradi-ent. Samples were centrifuged in an SW40Ti rotor at38,000 rpm and 4.0 8C for 1.5 hours. After centrifugation,500 ml fractionations were collected from the top usingthe BioComp Piston Gradient Fractionator (BioCompInstruments Inc., Fredericton, Canada). In order toidentify the fraction(s) containing the 15.5 kbp chroma-tin fragment, DNA obtained from 50 ml of each fractionwas purified by phenol/chloroform-extraction and etha-nol-precipitation. The purified DNA samples were ana-lyzed by real-time PCR on an ABI 7700 sequencedetector using DNA primers and Taqman probesspecific to the 15.5 kbp region (numbers 8.989, 17.763,19.368).12 The 15.5 kbp chromatin fragment was repro-ducibly found in fractions 14, 15 or 16.
Pulse-field gel electrophoresis and Southernblot analysis
Analysis of the chromatin fragments released from6C2 nuclei by HpaII digestion was performed by sizeseparation of the DNA on a BioRad CHEF-DRII. Bestseparation was achieved when samples were run at14 8C in 0.5 £ TBE at 200 V for 18 hours with an initialpulse time of 0.2 s and a final pulse time of 1 s. DNA ofinterest was sized by Southern blot analysis. DNA waspurified from 450 ml of the sucrose gradient fractions byphenol/chloroform-extraction and ethanol-precipitation,analyzed by pulse-field gel electrophoresis and trans-ferred to an Amersham Hybond N þ membrane asdescribed.26 Membranes were probed with probe P2, a566 bp fragment specific for the 15.5 kbp fragment,13
which also hybridizes to the Invitrogen 5 kbp ladderused as a marker. Blots were analyzed on an AmershamMolecular Dynamics Typhoon 8600 phosphorimager.
Sedimentation velocity experiments
Sedimentation velocity experiments were performedon selected sucrose gradient fractions containing both6C2 HpaII and turkey erythrocyte MNase-digested chro-matin. Fractions were dialyzed overnight at 4 8C againstDB buffer. Experiments were performed in a BeckmanOptima XL-A analytical ultracentrifuge at 4.0 8C. Thereference and sample sectors of Epon charcoal-filled,double-sector cells were loaded respectively with 350 mlof dialysis buffer and 300 ml of chromatin having anA260 nm of about 1.0. Sedimentation velocity runs wereperformed at speeds ranging from 10,000 rpm to16,000 rpm and monitored at 260 nm. Scans were takenat 3.25–4.00 minute intervals with a continuous0.003 cm radial step size. Sedimentation coefficientswere determined by fitting the data to a single species(Fujita) using the program Svedberg.27,28 Identical valuesof s (second moment) were obtained when data wereanalyzed using the program UltraScan II under Linux†.
Uncorrected sedimentation coefficients were convertedto s20;w values using measured solvent densities andviscosities.
Determination of the buoyant density of the15.5 kbp fragment
Sucrose gradient fractions containing the 15.5 kbpHpaII chromatin fragment from 6C2 nuclei were dialyzedinto 80 mM NaCl, 10 mM triethanolamine–NaOH (pH7.6), 0.2 mM EDTA and cross-linked with 0.5% formal-dehyde as described.17 Excess cross-linker was removedby dialysis into DB buffer. Samples were made up to1.0 ml and treated with an equal volume of cesium chlor-ide in dialysis buffer having a concentration of 1.06 g/ml, to a final density of approximately 1.40 g/ml. An ali-quot (2.0 ml) of this solution was centrifuged in a Beck-man TLS-55 rotor at 20.0 8C and 30,000 rpm for 96 hours.Experiments show that cesium chloride sedimentationequilibrium is achieved within 72 hours. Gradients werefractionated from the top into 20 £ 100 ml fractions. Analiquot (30 ml) of each fraction was used to determinethe refractive index, which was converted into a densitybased on an experimentally determined calibrationcurve using densities measured at 20.000 8C on anAnton Paar DMA 5000 densimeter. The remaining 70 mlwas treated as described15 to obtain free DNA and ana-lyzed by quantitative PCR using the 17.763 primer setand Taqman probe12 as described above. Note thatinitial buoyant density experiments were performed in14 mm £ 95 mm centrifuge tubes on an SW40Ti rotorspun at 30,000 rpm and 20.0 8C for 96 hours. Eventhough the cesium chloride had not yet reached sedi-mentation equilibrium, the 128 S 15.5 kbp chromatinfragment fractionated at its buoyant density due to itslarge size. In this manner, a density range of about 1.2–1.7 g/ml was analyzed. Furthermore, an analysis of thecross-linked chromatin fraction by 5–30% (w/v) sucrosegradients as described, demonstrates that materialretains its original sedimentation properties.
Determination of the buoyant density of chickencore histones
Histones H2A/H2B and H3/H4 were fractionated onhydroxylapatite using MNase-treated chicken erythro-cyte nuclei as described.29 Approximately 0.25 mg ofeach histone preparation was made up to 0.29 g/ml ofcesium chloride in DB buffer (final density, approxi-mately, 1.22 g/ml). Samples were centrifuged and col-lected as described above. Fractions were diluted sixtimes with water; histone proteins were precipitatedwith trichloroacetic acid and resuspended in 50 ml ofwater. Proteins were quantified using a Molecular ProbesCBQCA protein quantification kit following the manu-facturer’s recommendations, based on a histone proteincalibration curve.
Acknowledgement
This paper is dedicated to the memory of Pro-fessor Claude Helene, a friend and colleague whomade many important contributions to our under-standing of the molecular biology of nucleic acids.† http://www.ultrascan.uthscsa.edu/
Physical Properties of Condensed Chromatin Fragment 603

References
1. Elgin, S. C. (1996). Heterochromatin and gene regu-lation in Drosophila. Curr. Opin. Genet. Dev. 6, 193–202.
2. Grewal, S. I. S. & Elgin, S. C. R. (2002). Heterochroma-tin: new possibilities for the inheritance of structure.Curr. Opin. Genet. Dev. 12, 178–187.
3. Richards, E. J. & Elgin, S. C. R. (2002). Epigeneticcodes for heterochromatin formation and silencing:rounding up the usual suspects. Cell, 108, 489–500.
4. Finch, J. T. & Klug, A. (1976). Solenoidal model forsuperstructure in chromatin. Proc. Natl Acad. Sci.USA, 73, 1897–1901.
5. Lee, K. S., Mandelkern, M. & Crothers, D. M. (1981).Solution structural studies of chromatin fibers. Bio-chemistry, 20, 1438–1445.
6. McGhee, J. D., Nickol, J. M., Felsenfeld, G. & Rau,D. C. (1983). Higher order structure of chromatin:orientation of nucleosomes within the 30 nm chroma-tin solenoid is independent of species and spacerlength. Cell, 33, 831–841.
7. Widom, J. & Klug, A. (1985). Structure of the 300 Achromatin filament: X-ray diffraction from orientedsamples. Cell, 43, 207–213.
8. Widom, J., Finch, J. T. & Thomas, J. O. (1985). Higher-order structure of long repeat chromatin. EMBO J. 4,3189–3194.
9. McGhee, J. D. & Felsenfeld, G. (1986). Structure of the30 nm chromatin fiber. Cell, 44, 375–377.
10. Sen, D., Mitra, S. & Crothers, D. M. (1986). Higherorder structure of chromatin: evidence from photo-chemically detected linear dichroism. Biochemistry,25, 3441–3447.
11. Williams, S. P. & Langmore, J. P. (1991). Small angleX-ray scattering of chromatin. Radius and mass perunit length depend on linker length. Biophys. J. 59,606–618.
12. Litt, M. D., Simpson, M., Recillas-Targa, F., Prioleau,M. N. & Felsenfeld, G. (2001). Transitions in histoneacetylation reveal boundaries of three separatelyregulated neighboring loci. EMBO J. 20, 2224–2235.
13. Prioleau, M. N., Nony, P., Simpson, M. & Felsenfeld,G. (1999). An insulator element and condensed chro-matin region separate the chicken b-globin locusfrom an independently regulated erythroid-specificfolate receptor gene. EMBO J. 18, 4035–4048.
14. Kimura, T., Mills, F. C., Allan, J. & Gould, H. (1983).Selective unfolding of erythroid chromatin in theregion of the active b-globin gene. Nature, 306,709–712.
15. Caplan, A., Kimura, T., Gould, H. & Allan, J. (1987).Perturbation of chromatin structure in the region ofthe adult b-globin gene in chicken erythrocyte chro-matin. J. Mol. Biol. 193, 57–69.
16. Fisher, E. A. & Felsenfeld, G. (1986). Comparison ofthe folding of b-globin and ovalbumin gene contain-ing chromatin isolated from chicken oviduct anderythrocytes. Biochemistry, 25, 8010–8016.
17. Gilbert, N. & Allan, J. (2001). Distinctive higher-orderchromatin structure at mammalian centromeres.Proc. Natl Acad. Sci. USA, 98, 11949–11954.
18. Broersma, S. (1960). Viscous force constant for aclosed cylinder. J. Chem. Phys. 32, 1632–1635.
19. Garcia de la Torre, J. & Bloomfield, V. A. (1981).Hydrodynamic properties of complex, rigid, biologi-cal macromolecules: theory and applications. Quart.Rev. Biophys. 14, 81–139.
20. Newman, J., Swinney, H. L. & Day, L. A. (1977).
Hydrodynamic properties and structure of fd virus.J. Mol. Biol. 116, 593–606.
21. Tirado, M. M. & Garcia de la Torre, J. (1980). Transla-tional friction coefficients of rigid symmetric topmacromolecules. Application to circular cylinders.J. Chem. Phys. 71, 2581–2588.
22. Eisenberg, H. (1989). Solution properties of DNA:sedimentation, scattering of light, X-rays and neu-trons, and viscometry. Landolt-Bornstein New SeriesVII/1c, 257–276.
23. Sober, H. A. (1970). Editor of Handbook of Biochemis-try, 2nd edit., pp. H13–H17, CRC Press, Cleveland,OH.
24. Ifft, J. B. (1973). Proteins in density gradients at sedi-mentation equilibrium. In Methods in Enzymology,(Hirs, C. H. W. & Timasheff, S. N., eds), vol. 27, pp.128–140, Academic Press, New York.
25. Sedat, J. & Manuelidis, L. (1977). A direct approachto the structure of eukaryotic chromosomes. ColdSpring Harbor Symp. Quant. Biol. 42, 331–350.
26. Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). Mol-ecular Cloning. A Laboratory Manual, pp. 9.42–9.58,Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
27. Philo, J. S. (1994). Measuring sedimentation, diffu-sion and molecular weights of small molecules bydirect fitting of sedimentation velocity concentrationprofiles. In Modern Analytical Ultracentrifugation(Schuster, T. M. & Laue, T. M., eds), pp. 156–170, Bir-khauser, Boston.
28. Philo, J. S. (1997). An improved function for fittingsedimentation velocity data for low-molecular-weight solutes. Biophys. J. 72, 435–444.
29. Simon, R. H. & Felsenfeld, G. (1979). A new pro-cedure for purifying histone pairs H2A þ H2B andH3 þ H4 from chromatin using hydroxylapatite.Nucl. Acids Res. 6, 689–696.
Appendix
Determination of the 15.5 kbp chromatinfragment mass from the CsCl gradient
We observed that the 15.5 kbp HpaII chromatinfragment bands at r(CsCl) ¼ 1.404 g/ml.
At this concentration of CsCl, the specificvolume of DNA having a GC content similar tothat of chicken is nDNA ¼ 0.587 ml/g.A1 The buoy-ant densities of the chicken core histones in CsClwere measured (Figure 4) leading to specificvolumes of nH2A/H2B ¼ 0.8163 ml/g and nH3/
H4 ¼ 0.8097 ml/g. An average of these two valueswas used for the linker histones H1 and H5. Con-sider a chromatin complex consisting only of15.5 kbp DNA, histone octamers and histone lin-kers H1 and H5, with one nucleosome for every175 bp of DNA. Based on the specific volumes inCsCl, this chromatin complex is predicted to havea buoyant molecular mass, M(1 2 nr) of 186 kDaat r(CsCl) ¼ 1.404 g/ml. At this buoyant density,however, the measured M(1 2 nr) ¼ 0, indicatingthat excess protein is bound to the cross-linked15.5 kbp chromatin complex. Using a specificvolume of 0.80 ml/g for the excess protein,A2 theexcess protein mass is calculated at 1.51 MDa,
604 Physical Properties of Condensed Chromatin Fragment

equivalent to 17.0 kDa of bound protein pernucleosome. Based on this value and the expectedcontributions from the DNA and histone proteins,a mass of 23.28 MDa is determined for the15.5 kbp chromatin fragment.
Determination of the frictional coefficient fromthe sedimentation coefficient
Sedimentation velocity measurements in the ana-lytical ultracentrifuge yielded the values20;w ¼ 128ð^7Þ £ 10213 S for the sedimentation coef-ficient of the 15.5 kbp fragment. Based on the aminoacid sequence of the chicken histone proteins, andbased on the known stoichiometry of the core par-ticle, an effective n ¼ 0.7501 ml/g is calculated forthe histone core particle and H1/H5.A3 Assumingthat other proteins have a partial specific volumen ¼ 0.75 in waterA4 and that f0
DNA ¼ 0.52 ml/g,A1 wefind that M(1 2 nr) ¼ 8.21 MDa for the complex inwater. Since s ¼ M(1 2 nr)/Nf (where f is the fric-tional coefficient and N is Avogadro’s number), thevalue of f is (1.07 ^ 0.06) £ 1026 g/s. The value ofthe frictional coefficient for a sphere of equivalentmass, fo; is fo ¼ 6phð3Mn=4pNÞ1=3; which has thevalue 0.343 £ 1026 g/s. Therefore the frictional ratioof the 15.5 kbp chromatin particle is f=fo ¼3:1ð^0:2Þ; indicative of a highly asymmetric particle,such as a rod.
Dimension of a rod-like particle
The theory of BroersmaA5 (described inimproved form by Newman et al.6) allows calcu-lation of the linear diffusion coefficient D of a rodof length L and diameter d: If d ¼ lnð2L=dÞ and his the viscosity, then:
D ¼ ðkT=3phLÞ½dþ 3:7ð1=d2 0:34Þ2 þ 2:1ð1=d
2 0:39Þ2 2 0:73� ðA1Þ
The frictional coefficient is given by f ¼ kT=D:A given value of f is consistent with a range ofvalues of d; but for any value of d the value of L isdetermined. The solutions consistent with the
measured value of f for the 15.5 kbp condensedchromatin fiber are shown in Figure 4C of themain text. We have excluded solutions for particleswith diameters greater than 43 nm, but all of theshown combinations of values are in reasonableagreement with those expected for a chromatinfiber with a helix containing about six nucleo-somes/turn and with a pitch of about 11 nm.Application of another theoretical treatmentA7,A8
gives a dependence of L on d shifted slightly tothe right, so that for any assumed rod diameter,the predicted value of L is somewhat larger thanthat obtained from equation (A1) above.
References
A1. Eisenberg, H. (1989). Solution properties of DNA;sedimentation, scattering of light, X-rays and neu-trons, and viscometry. Landolt-Bornstein New SeriesVII/1c, 257–276.
A2. Ifft, J. B. (1973). Proteins in density gradients at sedi-mentation equilibrium. In Methods in Enzymology,(Hirs, C. H. W. & Timasheff, S. N., eds), vol. 27, pp.128–140, Academic Press, New York.
A3. Perkins, S. J. (1986). Protein volumes and hydrationeffects. The calculations of partial specific volumes,neutron scattering matchpoints and 280-nm absorp-tion coefficients for proteins and glycoproteins fromamino acid sequences. Eur. J. Biochem. 157, 169–180.
A4. Durchschlag, H. (1986). Specific volumes of biologi-cal macromolecules and some other molecules ofbiological interest. In Thermodynamic Data for Bio-chemistry and Biotechnology (Hinz, H.-J., ed.), pp.45–128, Springer, Berlin.
A5. Broersma, S. (1960). Viscous force constant for aclosed cylinder. J. Chem. Phys. 32, 1632–1635.
A6. Newman, J., Swinney, H. L. & Day, L. A. (1977).Hydrodynamic properties and structure of fd virus.J. Mol. Biol. 116, 593–606.
A7. Garcia de la Torre, J. & Bloomfield, V. A. (1981).Hydrodynamic properties of complex, rigid, biologi-cal macromolecules: theory and applications. Quart.Rev. Biophys. 14, 81–139.
A8. Tirado, M. M. & Garcia de la Torre, J. (1980). Transla-tional friction coefficients of rigid symmetric topmacromolecules. Application to circular cylinders.J. Chem. Phys. 71, 2581–2588.
Edited by A. Klug
(Received 29 September 2003; received in revised form 10 December 2003; accepted 12 December 2003)
Physical Properties of Condensed Chromatin Fragment 605








![Interacting Genomic Landscapes of REC8-Cohesin ...Interacting Genomic Landscapes of REC8-Cohesin, Chromatin, and Meiotic Recombination in Arabidopsis[CC-BY] Christophe Lambing,a Andrew](https://img.pdfslide.us/doc/110x75/60dbfd21ae2a635f0e07e13a/interacting-genomic-landscapes-of-rec8-cohesin-interacting-genomic-landscapes.jpg)




















