Embed Size (px)
Citation preview

1
RESEARCH ARTICLE 1
2
Interacting genomic landscapes of REC8-cohesin, chromatin 3
and meiotic recombination in Arabidopsis thaliana 4 5
Christophe Lambinga, Andrew J. Tocka, Stephanie D. Toppa, Kyuha Choia,d, Pallas 6 C. Kuoa, Xiaohui Zhaoa, Kim Osmanb, James D. Higginsc, F. Chris H. Franklinb7 and Ian R. Hendersona,e8
9 a Department of Plant Sciences, University of Cambridge, Cambridge, CB2 3EA, United 10 Kingdom 11 b School of Biosciences, University of Birmingham, Birmingham, B15 2TT, United 12 Kingdom 13 c Department of Genetics and Genome Biology, University of Leicester, Leicester, LE1 14 7RH, United Kingdom 15 d Present Address: Department of Life Sciences, Pohang University of Science and 16 Technology, Pohang, Gyeongbuk, Republic of Korea. 17 e Corresponding Author: [email protected] 18
19 Short title: Genome-wide landscape of meiotic cohesin REC8. 20
21 Keywords: Meiosis, cohesin, REC8, chromatin, recombination, crossover, H3K9me2, 22 DNA methylation, Arabidopsis. 23
24 One-sentence summary: REC8 association with the genome correlates with multiple 25 chromatin states and is required to organize meiotic chromosome architecture and 26 interhomolog recombination. 27
28 The author responsible for distribution of materials integral to the findings presented in 29 this article in accordance with the policy described in the Instructions for Authors 30 (www.plantcell.org) is: Ian R. Henderson ([email protected]). 31
32 ABSTRACT 33 Meiosis recombines genetic variation and influences eukaryote genome evolution. 34 During meiosis, DNA double-strand breaks (DSBs) enter interhomolog repair to yield 35 crossovers and non-crossovers. DSB repair occurs as replicated sister chromatids are 36 connected to a polymerized axis. Cohesin rings containing the REC8 kleisin subunit 37 bind sister chromatids and anchor chromosomes to the axis. Here we report the 38 genomic landscape of REC8 using ChIP-seq in Arabidopsis thaliana. REC8 associates 39 with regions of high nucleosome occupancy in multiple chromatin states, including 40 histone methylation at H3K4 (expressed genes), H3K27 (silent genes) and H3K9 41 (silent transposons). REC8 enrichment is associated with suppression of meiotic DSBs 42 and crossovers at the chromosome and fine scales. As REC8 enrichment is greatest in 43 transposon-dense heterochromatin, we repeated ChIP-seq in kyp suvh5 suvh6 44 H3K9me2 mutants. Surprisingly, REC8 enrichment is maintained in kyp suvh5 suvh6 45 heterochromatin and no defects in centromeric cohesion were observed. REC8 46 occupancy within genes anti-correlates with transcription and is reduced in COPIA 47 transposons that reactivate expression in kyp suvh5 suvh6. Abnormal axis structures 48 form in rec8 that recruit DSB-associated protein foci and undergo synapsis, which is 49 followed by chromosome fragmentation. Therefore, REC8 occupancy correlates with 50 multiple chromatin states and is required to organize meiotic chromosome architecture 51 and interhomolog recombination. 52
Plant Cell Advance Publication. Published on February 4, 2020, doi:10.1105/tpc.19.00866
©2020 The author(s).

2
INTRODUCTION 53 54 The majority of eukaryotes reproduce via meiosis, which is a specialized cell division 55 that produces four haploid cells from a single diploid parent cell (Villeneuve and Hillers, 56 2001; Mercier et al., 2015). During meiosis, a single round of DNA replication is 57 followed by two rounds of chromosome segregation, which recombines the 58 chromosome complement (Villeneuve and Hillers, 2001; Mercier et al., 2015). In 59 addition, during meiotic prophase I, homologous chromosomes undergo programmed 60 DNA double-strand breaks (DSBs) generated via SPO11 complexes (Lam and Keeney, 61 2014; de Massy, 2013). Meiotic DSBs are resected to form 3′-single-stranded DNA 62 (ssDNA), which may invade the homologous chromosome, allowing repair as 63 crossovers or non-crossovers (Villeneuve and Hillers, 2001; Mercier et al., 2015). In 64 Arabidopsis thaliana approximately 200 meiotic DSBs are formed, of which around 10 65 are repaired as crossovers (Mercier et al., 2015). The majority of crossovers in plants 66 exhibit interference, which is the phenomenon that prevents multiple crossovers 67 occurring in close proximity along each chromosome pair (Villeneuve and Hillers, 2001; 68 Mercier et al., 2015). The combined effects of meiotic recombination and chromosome 69 segregation strongly influence genetic variation, genome evolution and adaptation 70 (Barton and Charlesworth, 1998; Kauppi et al., 2004). 71 72 Prior to the formation of meiotic DSBs, homologous chromosomes become associated 73 with a polymerized axis structure (Zickler and Kleckner, 1999). In Arabidopsis this 74 includes the ASY1 HORMA domain protein and the coiled-coil proteins ASY3 and 75 ASY4 (West et al., 2019; Armstrong et al., 2002; Chambon et al., 2018; Ferdous et al., 76 2012; Osman et al., 2018). During association with the axis, replicated sister 77 chromatids are held together by cohesin complexes containing the meiosis-specific 78 REC8 α-kleisin, which is also required to connect chromatin to the axis (Klein et al., 79 1999; Kim et al., 2010; Chelysheva et al., 2005; Cai et al., 2003). Meiotic DSBs are 80 proposed to form predominantly within the chromatin loops that become tethered to the 81 axis during interhomolog repair (Zickler and Kleckner, 1999). As meiotic prophase 82 proceeds, the axis is remodeled to form the synaptonemal complex (SC), coincident 83 with the progression of crossover repair (Zickler and Kleckner, 1999; Page and Hawley, 84 2003). The higher-order organization of meiotic chromosomes into a tethered-loop/axis 85 configuration is required for efficient and high-fidelity interhomolog recombination 86 (Zickler and Kleckner, 1999; Page and Hawley, 2003). 87 88 Epigenetic information on the chromosome loops, including nucleosome positions, 89 histone modifications, histone variants and DNA methylation, are also known to 90 influence meiotic recombination (Choi and Henderson, 2015; Choi et al., 2018; Pan et 91 al., 2011; Sommermeyer et al., 2013; Acquaviva et al., 2013; Choi et al., 2013; Yelina 92 et al., 2015). For example, DSBs and crossovers typically concentrate in narrow ~1 kb 93 hotspots, which are determined to varying degrees by genetic and epigenetic 94 information (Choi and Henderson, 2015; Lam and Keeney, 2014; Baudat et al., 2013). 95 During DSB formation, SPO11 enzymes become covalently bound to target site 96 oligonucleotides, which can be sequenced to identify sites of recombination initiation 97 (Pan et al., 2011; Fowler et al., 2014; Neale et al., 2005; Lange et al., 2016; Choi et al., 98 2018). SPO11-oligo sequencing in Arabidopsis and budding yeast has revealed 99 meiotic DSB hotspots located in nucleosome-free regions associated with gene 100 regulatory sequences (Choi et al., 2018; Pan et al., 2011). Crossovers in plants also 101 positively associate with H3K4me3 and histone variant H2A.Z, which show 5′ 102 enrichment at transcribed genes (Choi et al., 2013; Liu et al., 2009; Wijnker et al., 103 2013; He et al., 2017; Kianian et al., 2018). In budding yeast, H3K4me3 tethers DSB 104 hotspots to the Mer2 axis protein via the COMPASS complex Spp1 subunit, showing 105

3
how chromatin and axis organization can interact to promote meiotic recombination 106 (Sommermeyer et al., 2013; Acquaviva et al., 2013). 107 108 Meiotic recombination is also suppressed in broad regions of the chromosomes, 109 including heterochromatin, centromeres, ribosomal gene arrays, sex chromosomes 110 and mating-type loci (Underwood et al., 2018; Choi et al., 2018; Ellermeier et al., 2010; 111 Yelina et al., 2015; Vincenten et al., 2015). Crossover suppression in these regions 112 may be due to both structural rearrangements and epigenetic marks. Arabidopsis 113 centromeres are organized as megabase arrays of satellite repeats that are 114 surrounded by pericentromeric heterochromatin, which is densely modified with DNA 115 methylation in CG, CHG and CHH sequence contexts, H3K9me2, H3K27me1 and 116 histone variant H2A.W (Topp and Dawe, 2006; Stroud et al., 2013; Yelagandula et al., 117 2014; Jacob et al., 2009; Stroud et al., 2014). Arabidopsis pericentromeric 118 heterochromatin is suppressed for both meiotic DSBs and crossovers, and loss of 119 H3K9me2 and non-CG DNA methylation increases meiotic recombination in these 120 regions (Choi et al., 2018; Underwood et al., 2018). Crossover hotspots located in 121 Arabidopsis euchromatin have also been directly silenced via RNA-directed DNA 122 methylation and acquisition of H3K9me2 (Yelina et al., 2015). Although the presence of 123 specific heterochromatic marks varies between eukaryotic genomes, suppression of 124 meiotic recombination in centromeric and pericentromeric chromatin is widely observed 125 (Underwood et al., 2018; Vincenten et al., 2015; Pan et al., 2011; Fowler et al., 2014; 126 Ellermeier et al., 2010). 127 128 Despite the importance of the tethered-loop/axis architecture and chromatin for meiotic 129 recombination, how they interact to shape DSB and crossover frequency is 130 incompletely understood. REC8-cohesin occupancy has been analyzed via ChIP-seq 131 in budding and fission yeast, and found to accumulate most strongly in the centromeric 132 regions (Folco et al., 2017; Sun et al., 2015; Ito et al., 2014; Kugou et al., 2009). In 133 fission yeast, REC8 loading in heterochromatin requires H3K9 methylation, further 134 demonstrating the importance of chromatin for meiotic chromosome architecture 135 (Bernard et al., 2001; Nonaka et al., 2002; Mizuguchi et al., 2014; Folco et al., 2017). 136 Within the chromosome arms, transcription exerts an influence on mitotic and meiotic 137 cohesin occupancy, which accumulates at sites of convergent transcription in budding 138 and fission yeast (Sun et al., 2015; Lengronne et al., 2004; Mizuguchi et al., 2014). In 139 this study we explore REC8 localization in the Arabidopsis genome and its relationship 140 to recombination and chromatin state. The Arabidopsis genome is physically larger 141 than budding yeast and fission yeast, with greater gene and transposon numbers and 142 complex megabase-scale centromeres (Table 1). Heterochromatic regulation in 143 Arabidopsis also involves additional chromatin modifications, including DNA 144 methylation and H2A.W (Table 1) (Stroud et al., 2013; Yelagandula et al., 2014). 145 Notably, genome-wide maps of meiotic DSBs, crossovers and chromatin exist for each 146 of these species, making cross-species comparisons of interest (Tables 1 and 2) 147 (Fowler et al., 2014; Pan et al., 2011; Choi et al., 2018; Mancera et al., 2008). 148 149 In this work we map the landscape of Arabidopsis REC8 via ChIP-seq and observe a 150 positive correlation with nucleosome occupancy in multiple euchromatic and 151 heterochromatic chromatin states, including with histone H3K4, H3K9 and H3K27 152 methylation. At the chromosome scale, REC8 enrichment is greatest in pericentromeric 153 heterochromatin and associates with suppression of meiotic DSBs and crossovers. To 154 test the role of heterochromatin in REC8 enrichment we repeated ChIP-seq in kyp 155 suvh5 suvh6 mutants, which lose H3K9me2 and non-CG DNA methylation (Stroud et 156 al., 2014). Surprisingly, REC8 enrichment is maintained in kyp suvh5 suvh6 157 heterochromatin and no defects in centromeric cohesion occurred. We observe that 158

4
patterns of transcription influence REC8 occupancy within genes and transposons. For 159 example, REC8 decreases within COPIA transposable elements that become 160 transcriptionally up-regulated in kyp suvh5 suvh6. Together our work provides a 161 genome-wide map of meiotic cohesin in plants and insights into how chromatin and the 162 axis interact to shape meiotic chromosome architecture and recombination in the 163 Arabidopsis genome. 164 165 RESULTS 166 167 REC8-cohesin and chromatin organization during Arabidopsis meiosis 168 169 To detect REC8 during meiosis, we inserted 3×HA or 5×Myc epitopes into a REC8 170 genomic clone that uses the endogenous promoter, and transformed rec8-3 171 heterozygotes (Cai et al., 2003; Chelysheva et al., 2005). We assessed 172 complementation of rec8 by cytological analysis of meiosis in the resulting 173 transformants (Fig. 1A). rec8 shows severe meiotic defects in chromatin compaction, 174 axis formation and the presence of chromosome fragmentation, which cause complete 175 sterility (Fig. 1A) (Cai et al., 2003; Chelysheva et al., 2005; Bhatt et al., 1999). We 176 observed that transformation with the HA and Myc epitope-tagged REC8 constructs 177 fully complemented axis formation during prophase I, the presence of five bivalents at 178 metaphase I and chiasmata formation in rec8-3 (Fig. 1A and Supplemental Table 1). 179 180 To analyze REC8 accumulation on meiotic chromosomes, we immunostained male 181 meiocytes using α-HA or α-Myc antibodies and stained chromatin with DAPI (Fig. 1B 182 and Supplemental Figure 1A). We observed co-localization of REC8 with chromatin 183 from leptotene onwards, including within heterochromatin (Fig. 1B and Supplemental 184 Figure 1A). REC8 and chromatin co-localized as pairing and synapsis occured, with 185 strong co-staining of the paired axes at pachytene (Fig. 1B and Supplemental Figure 186 1A). The tagged REC8 proteins persisted on bivalents through diakinesis and 187 metaphase I (Fig. 1B and S1A), as reported (Chelysheva et al., 2005; Cai et al., 2003). 188 No signal was detected in non-transgenic wild type meiocytes, or somatic cells of 189 REC8 epitope-tagged lines (Fig. 1B and Supplemental Figure 1A). This demonstrates 190 specific detection of REC8-HA and REC8-Myc proteins with the expected localization 191 on meiotic chromosomes (Chelysheva et al., 2005; Cai et al., 2003). 192 193 To visualize chromatin organization during meiosis, we immunostained for euchromatic 194 (H3K4me3) and heterochromatic (H3K9me2 and H3K27me1) histone modifications, 195 together with the SC protein ZYP1 (Fig. 1C) (Higgins et al., 2005). During late 196 prophase I, when ZYP1 polymerization was complete, we observed clear differentiation 197 of euchromatin and heterochromatin, with ZYP1 signal continuous through both 198 chromatin types (Fig. 1C). To assess axis and SC signal through the centromeres, we 199 performed fluorescence in situ hybridization (FISH) for the satellite repeat CEN180 and 200 co-immunostained for ZYP1 or the SMC3 cohesin subunit (Fig. 1D). At the zygotene-201 pachytene transition, we observed CEN180-positive regions, through which the SMC3 202 and ZYP1 signals were continuous (Fig. 1D). Together these data show that the SC 203 and cohesin form a continuous axis through Arabidopsis euchromatin, heterochromatin 204 and the centromeres during meiosis. 205 206 Profiling the genomic landscape of REC8 enrichment by ChIP-seq 207 208 To generate a genome-wide map of REC8 enrichment we sought to use the HA and 209 Myc epitope-tagged lines to perform ChIP-seq. As REC8 is specifically expressed 210 during meiosis (Fig. 1B and Supplemental Figure 1A) (Cai et al., 2003; Chelysheva et 211

5
al., 2005), we collected unopened flower buds for ChIP. These flowers contain all 212 stages of meiosis, although prophase I has the longest duration (~31 of ~33 hours) 213 (Armstrong, 2013), during which REC8 associates with chromosomes (Fig. 1B and 214 Supplemental Figure 1A). We performed immunoblotting on meiotic-stage flowers 215 before and after α-HA or α-Myc immunoprecipitation, which revealed specific bands of 216 the expected size (77.2 kDa REC8-HA and 84.2 kDa REC8-Myc), in addition to bands 217 with an apparent ~20 kDa higher molecular mass, likely representing phosphorylated 218 forms of REC8 (Fig. 2A and Supplemental Figure 1B). For example, slow migrating 219 forms of REC8 have also been observed in yeast and mice, which are caused by 220 phosphorylation (Watanabe and Nurse, 1999; Kitajima et al., 2003). A non-specific α-221 HA band was detected in input samples from REC8-HA rec8 and wild type (Col), but 222 this band was greatly depleted after immunopurification (Fig. 2A). 223 224 Approximately 10 grams of unopened flowers collected from pools of plants were used 225 for ChIP from REC8-HA rec8 and the resulting DNA was used to construct sequencing 226 libraries (Supplemental Table 2). We analyzed genome coverage values from 227 biological replicate REC8-HA ChIP-seq libraries, which were highly correlated when 228 compared at varying physical scales (e.g., 10 kb windows rs=0.92) (Supplemental 229 Table 3). To test the specificity of REC8 enrichment we repeated α-HA ChIP from 230 untagged wild type (Col) flowers, which was used to generate a sequencing library. 231 The Col library yielded 3,681,603 read pairs of which only 0.75% mapped to the 232 Arabidopsis genome, compared to mapping rates of 91.34-93.22% of reads from the 233 REC8-HA libraries (Supplemental Table 2). This demonstrates the low background 234 obtained by our ChIP-seq protocol. 235 236 To further analyze the specificity of α-HA ChIP enrichment, three regions were 237 analyzed by qPCR from REC8-HA rec8 versus untagged Col (Fig. 2B and 238 Supplemental Table 4). The regions analyzed were heterochromatic ATHILA and 239 ATHILA6B transposable elements, and the 5′ end of a euchromatic gene (At2g02480), 240 which were predicted to show high REC8 occupancy (Fig. 2B and Supplemental Table 241 4). For each region, significantly greater enrichment was measured from ChIP relative 242 to input for REC8-HA rec8 compared to untagged wild-type Col, with highest REC8 243 enrichment within the transposons (Fig. 2B and Supplemental Table 4), further 244 confirming the specificity of our protocol. Finally, we analyzed α-Myc ChIP-seq data 245 from REC8-Myc rec8 plants, which showed a positive correlation with REC8-HA data 246 (e.g., 10 kb rs=0.83-0.86) (Fig. 3A and Supplemental Table 3). This demonstrates that 247 we obtain reproducible signal when performing REC8 chromatin immunoprecipitation 248 using two independent epitopes. Due to the high correlation of REC8-HA and REC8-249 Myc data, for subsequent analysis we focused on REC8-HA ChIP-seq data. 250 251 To assess REC8 enrichment at the fine scale we analyzed ChIP-seq values (per-base 252 coverage values normalized by total library coverage) and compared with chromatin 253 and recombination data (Fig. 2C and Table 2). These data included: (i) nucleosome 254 occupancy (MNase-seq) (Choi et al., 2016), (ii) markers of heterochromatin H3K9me2 255 (ChIP-seq), DNA methylation (BS-seq) and short interfering RNAs (siRNA-seq) (Lee et 256 al., 2012; Stroud et al., 2013), (iii) histone marks enriched at euchromatic genes 257 H3K4me3 (Choi et al., 2018), H3K4me2, H3K4me1 (ChIP-seq) and mRNA from 258 flowers or male meiocytes (RNA-seq) (Walker et al., 2018), (iv) the Polycomb histone 259 mark H3K27me3 (ChIP-seq) (Zhu et al., 2015), and (v) SPO11-1-oligos that mark 260 meiotic DSBs (Choi et al., 2018) and crossovers mapped by sequencing Col×Ler F2 261 plants (Serra et al., 2018; Rowan et al., 2019). 262 263 With reference to a representative 65 kb region of chromosome 3 we observed a 264

6
positive correlation between REC8 and nucleosome occupancy, both of which 265 negatively correlated with SPO11-1-oligos (Fig. 2C). High REC8 enrichment was 266 evident within different types of sequence element and chromatin state; for example, (i) 267 a transcriptionally silent LINE1 transposable element (AT3TE37690) that showed high 268 nucleosome density, DNA methylation, H3K9me2, and siRNAs, and suppressed 269 SPO11-1-oligos, (ii) expressed genes (e.g., TMKL1, PLL22, HCR1 and F-box) that 270 showed high nucleosome occupancy with H3K4me1, H3K4me2 and H3K4me3 271 modifications, and (iii) a H3K27me3-silenced gene (ABI3) (Fig. 2C). Therefore we next 272 sought to explore REC8 ChIP-seq enrichment at multiple scales and within different 273 sequence contexts in relation to chromatin and recombination datasets. 274 275 REC8 enrichment in euchromatin and heterochromatin at the chromosome scale 276 277 We compared REC8-HA ChIP-seq enrichment with markers of euchromatin and 278 heterochromatin and meiotic recombination along the Arabidopsis chromosomes (Fig. 279 3). For genome-wide analysis REC8-HA ChIP and input data were used to calculate 280 windowed log2(ChIP/input) enrichment values. At the chromosome scale, REC8-HA 281 enrichment was greatest in the centromeres and in proximal heterochromatin, which 282 positively correlated with nucleosome occupancy (rs=0.68) (Fig. 3A). Due to 283 pericentromeric REC8-HA enrichment, we also observed positive correlations with 284 transposable element density (rs=0.74), DNA methylation in CG (rs=0.79), CHG 285 (rs=0.72) and CHH (rs=0.72) sequence contexts, and the heterochromatic histone 286 modifications H3K9me2 (rs=0.78), H3K27me1 (rs=0.75) and histone variant H2A.W 287 (rs=0.83) (Fig. 3A) (Stroud et al., 2013; Yelagandula et al., 2014). These relationships 288 were further strengthened if the pericentromeres were considered separately from the 289 chromosome arms (Fig. 3B). Hence, as in budding and fission yeasts (Folco et al., 290 2017; Sun et al., 2015; Ito et al., 2014; Kugou et al., 2009), the strongest REC8 ChIP-291 seq enrichment in Arabidopsis is observed in proximity to the centromeres. 292 293 To provide cytological support for the trends observed via ChIP-seq, we analyzed 294 spread nuclei at pachytene stage that were immunostained for REC8-HA and 295 chromatin stained with DAPI (Fig. 3C). We tracked and quantified axial REC8-HA 296 signal as it transversed heterochromatin (Fig. 3C). We centered analysis on 297 heterochromatin over a distance of 100 pixels (equivalent to ~6.4 μM) in 20 sections 298 from a total of 10 meiocytes (Fig. 3C). We observed that mean REC8-HA and DAPI 299 signal intensity were significantly correlated over the tracked regions (Spearman’s 300 correlation coefficient rs=0.94 P=4.07×10-47). These cytological data are consistent with 301 REC8-HA ChIP-seq enrichment correlating with nucleosome-dense heterochromatin 302 (Fig. 3A-3B). 303 304 Due to REC8-HA enrichment in pericentromeric heterochromatin, genome-wide 305 negative correlations between REC8-HA and gene density (rs=-0.70), and the gene-306 associated chromatin modifications H3K4me1 (rs=-0.63), H3K4me2 (rs=-0.74), 307 H3K4me3 (rs=-0.75), H2A.Z (rs=-0.62) and H3K27me3 (rs=-0.26) were observed (Fig. 308 3A-3B) (Yelagandula et al., 2014). When the chromosome arms were considered 309 separately, a weaker positive correlation between REC8-HA and nucleosome 310 occupancy was observed (r=0.40) (Fig. 3B). The chromosome arms undergo the 311 greatest levels of meiotic DSBs (measured via SPO11-1-oligos) and crossovers (Fig. 312 3B). However, as noted previously (Choi et al., 2018; Underwood et al., 2018), while 313 crossovers and SPO11-1-oligos are positively correlated (genome-wide rs=0.63; 314 chromosome arms rs=0.46; pericentromeric rs=0.85), they show considerable variation 315 in their relative frequencies within the chromosome arms (Fig. 3A). These observations 316 prompted us to investigate the relationship between recombination, chromatin and 317

7
REC8-HA ChIP enrichment at the fine scale. 318 319 REC8 enrichment associates with suppression of meiotic DSBs and crossovers 320 321 REC8-HA enrichment peaks in ChIP-seq data were identified using the ranger tool 322 within the PeakRanger suite, providing input library reads as a control for background 323 (P ≤ 0.001 and FDR ≤ 0.01) (Feng et al. 2011). This approach identified 86,386 REC8-324 HA peaks, which had a mean width of 444 bp (Supplemental Figure 2 and 325 Supplemental Tables 5-6). The average REC8-HA coverage profile between the peak 326 start and end coordinates is positively correlated with nucleosome occupancy (rs=0.80) 327 and anti-correlated with SPO11-1-oligos (rs=-0.83), which was similar to the pattern 328 observed when well-positioned nucleosomes (n=57,734, mean width=145 bp) were 329 analyzed (Fig. 4A). We analyzed REC8-HA peaks separately according to their 330 location in the chromosome arms or pericentromeres and observed the same trends of 331 nucleosome enrichment and SPO11-1-oligo depletion (Fig. 4B). We next analyzed sets 332 of 3,320 crossovers (mean resolved width=976 kb) and 5,914 SPO11-1-oligo hotspots 333 and observed opposite trends to the REC8-HA peaks, with depleted nucleosomes and 334 REC8-HA, compared to the same number of random windows with the same widths 335 (Fig. 4A) (Choi et al., 2018; Serra et al., 2018). Hence, at the local scale, higher 336 nucleosome and REC8 occupancy associate with suppression of both meiotic DSBs 337 and crossovers. 338 339 REC8-HA peaks and SPO11-1-oligo hotspots were evaluated for overlap with other 340 genomic features by performing permutation tests using the R package regioneR (Gel 341 et al. 2016). For each test, 10,000 sets of randomly positioned loci with the same width 342 distribution as the REC8-HA peaks or SPO11-1-oligo hotspots were defined. For each 343 set, the number of random loci overlapping features of interest was then compared with 344 the observed number of REC8-HA peaks or SPO11-1-oligo hotspots overlapping the 345 features. This provides the basis for calculating an empirical P value denoting the 346 significance of the observed overlap (Supplemental Figure 2 and Supplemental Tables 347 5-6). REC8-HA peaks overlapped with well-positioned nucleosomes and H3K9me2 348 peaks significantly more than expected by chance, whereas they overlapped SPO11-1-349 oligo hotspots and crossovers significantly less than expected (Supplemental Figure 2 350 and Supplemental Tables 5-6). This provides statistical support for nucleosomes and 351 REC8 co-localizing and exerting a suppressive effect on meiotic recombination. 352 353 DNA base composition is known to influence nucleosome positioning, meiotic 354 recombination and cohesin occupancy in multiple systems (Glynn et al., 2004; Blat et 355 al., 2002; Laloraya et al., 2000; Choi et al., 2018). Therefore, we analyzed AT:GC 356 content around REC8-HA peaks, well-positioned nucleosomes, SPO11-1-oligo 357 hotspots and crossovers (Fig. 4C). Consistent with the known properties of 358 nucleosomes (Segal and Widom, 2009), we observed pronounced depletion of AT and 359 enrichment of GC bases around well-positioned nucleosomes (Fig. 4C). Similarly, 360 REC8-HA peaks show GC enrichment and AT depletion (Fig. 4C), supporting the 361 correlations between REC8-HA and nucleosome occupancy (Fig. 4A-4B). By contrast, 362 both SPO11-1-oligo hotspots and crossovers show the opposite trends of AT 363 enrichment and GC depletion (Fig. 4C), as reported (Choi et al., 2018). This reveals 364 that variation in DNA sequence frequency, chromatin structure, cohesin occupancy and 365 recombination are intimately connected in Arabidopsis, with GC-richness underlying 366 local nucleosome and REC8 enrichment and suppression of meiotic recombination. 367 368 REC8, chromatin and recombination within transposable elements 369 370

8
We previously observed that Arabidopsis transposable element families are 371 differentiated by levels of nucleosome occupancy and SPO11-1-oligos (Choi et al., 372 2018). For example, Gypsy, Copia and LINE1 RNA transposons are relatively 373 nucleosome-dense and suppressed for SPO11-1-oligos, compared with DNA elements 374 including MuDR and Helitron transposons, which are nucleosome-depleted, DSB 375 active and enriched in proximity to genes (Choi et al., 2018). We therefore compared 376 REC8-HA levels in DNA versus RNA transposable elements and at the same number 377 of randomly positioned loci of the same widths (Fig. 5A-5B). We observed that DNA 378 elements were relatively depleted for REC8-HA and nucleosomes, whereas higher 379 levels occurred in RNA elements (Fig. 5A-5B). This correlates with DNA elements 380 showing lower levels of the heterochromatic marks H3K9me2 and CG, CHG and CHH 381 DNA methylation, compared with RNA transposons (Fig. 5A-5B). 382 383 We next evaluated overlap of REC8 peaks, well-positioned nucleosomes and SPO11-384 1-oligo hotspots with different transposable element families (Fig. 5C). Gypsy, LINE1, 385 Copia RNA and EnSpm/CACTA DNA transposons showed significant positive overlap 386 with REC8 peaks and nucleosomes and negative overlap with SPO11-1-oligo hotspots 387 (Fig. 5C). By contrast, MuDR, HELITRON and Pogo/Tc1/Mariner DNA transposons 388 overlap REC8-HA and nucleosome peaks significantly less than expected by chance, 389 and SPO11-1-oligo hotspots significantly more than expected (Fig. 5C). To illustrate 390 differences in transposons we show a representative region on chromosome 3 391 containing a REC8-enriched COPIA81 RNA element, in the vicinity of a REC8-depleted 392 ATREP3 Helitron DNA element (Fig. 5D). Hence, Arabidopsis transposon families are 393 highly differentiated for REC8-cohesin enrichment, chromatin and recombination. 394 395 REC8 recruitment to heterochromatin is unimpaired in kyp suvh5 suvh6 396 H3K9me2 and non-CG DNA methylation mutants 397 398 Due to the enrichment of REC8 observed in the pericentromeres (Fig. 2A), we sought 399 to investigate whether loss of heterochromatic marks would change cohesin occupancy, 400 as observed in fission yeast H3K9 methylation mutants (Bernard et al., 2001; Nonaka 401 et al., 2002; Mizuguchi et al., 2014; Folco et al., 2017). Previously, we observed that 402 kyp suvh5 suvh6 triple mutants, which lose H3K9me2 and non-CG methylation, gain 403 SPO11-1-oligos within heterochromatic sequences (Underwood et al., 2018; Stroud et 404 al., 2014). Therefore, we repeated REC8-HA ChIP-seq in kyp suvh5 suvh6 mutants. 405 Floral buds from pooled F3 plants were used for this experiment, derived from the same 406 REC8-HA line crossed to kyp suvh5 suvh6. We also performed H3K9me2 ChIP-seq in 407 kyp suvh5 suvh6 to map the change of this chromatin modification. H3K9me2 and non-408 CG methylation are tightly coupled in Arabidopsis and both marks are lost in kyp suvh5 409 suvh6 mutants (Fig. 6A) (Stroud et al., 2014). 410 411 In contrast to fission yeast, we observed that the pattern of REC8-HA enrichment along 412 the chromosomes in kyp suvh5 suvh6 was highly similar to wild type, with strong 413 enrichment observed in proximity to the centromeres (Fig. 6A). We immunostained for 414 REC8-HA in wild-type and kyp suvh5 suvh6 male meiocytes and observed that normal 415 signal intensity with the heterochromatic chromocenters was maintained, using the F3 416 lines generated for ChIP analysis (Fig. 6B). Finally, we analyzed male meiocytes in kyp 417 suvh5 suvh6 cytologically for phenotypes associated with rec8 mutants (Fig. 6C) (Cai 418 et al., 2003; Chelysheva et al., 2005; Bhatt et al., 1999). At pachytene stage, normal 419 paired axes are observed in kyp suvh5 suvh6 and sister chromatid cohesion is 420 maintained at metaphase I and metaphase II, without obvious defects in chromosome 421 segregation (Fig. 6C). Hence, REC8 cohesin is recruited to levels sufficient to maintain 422 sister-chromatid cohesion and bivalent formation during meiosis in kyp suvh5 suvh6. 423

9
Transcription influences REC8 occupancy within genes and transposons 424 425 Transcription has been shown to shape mitotic and meiotic cohesin occupancy (Sun et 426 al., 2015; Lengronne et al., 2004; Mizuguchi et al., 2014). Therefore, we analyzed 427 REC8-HA enrichment in relation to gene bodies (TSS-TTS) and 2 kb of flanking 428 sequences, versus the same number of randomly positioned loci of the same widths 429 (Fig. 7A). As reported, SPO11-1-oligos are depleted within gene bodies and show 430 greatest enrichment in nucleosome-free regions in promoters and terminators (Fig. 7A) 431 (Choi et al., 2018). By contrast, REC8-HA, MNase-seq (nucleosomes) and RNA-seq 432 signal are enriched intragenically, and their average profiles show positive correlations 433 with one another (rs=0.66-0.96), with highest signal towards gene 3′ ends (Fig. 7A). For 434 example, REC8-HA is relatively depleted from the 5′ regions of genes that show 435 greatest enrichment of H2A.Z and H3K4me3 (Fig. 7A). These correlations indicate that 436 transcription may influence both nucleosome and REC8 occupancy within gene bodies, 437 or vice versa. 438 439 To explore the relationship with transcription, we ordered genes by increasing 440 expression measured from floral RNA-seq data and compared this with REC8-HA 441 ChIP-seq enrichment within genes (Fig. 7B). This showed a negative correlation 442 between REC8-HA occupancy within genes and expression level (rs=-0.40) (Fig. 7B). 443 As gene expression increases, REC8-HA and nucleosomes occur at lower levels 444 intragenically and show an increased bias towards the gene 3′ ends (Fig. 7B). This is 445 consistent with higher RNA polymerase activity causing migration of cohesin in the 446 direction of transcription. Genes with higher expression also show greater enrichment 447 of H3K4me3 and H2A.Z at their 5′ end, coincident with lower local REC8-HA levels (Fig. 448 7B). Interestingly, SPO11-1-oligos in gene promoters and terminators did not 449 significantly correlate with gene expression or intragenic levels of REC8-HA (Fig. 7B). 450 451 We performed RNA-seq from wild type and kyp suvh5 suvh6 meiotic-stage flowers and 452 identified 179 transposable elements that showed significant upregulation of 453 expression in the triple mutant. Upregulated transposons were enriched for elements 454 belonging to the EnSpm (hypergeometric test P=1.77×10-25), Copia (P=1.69×10-17), 455 Harbinger (P=0.0004), and LINE1 (P=0.0125) superfamilies (Supplemental Table 7). 456 Upregulated Copia transposons (n=45) show loss of H3K9me2 and non-CG DNA 457 methylation in kyp suvh5 suvh6 (Fig. 7C). As a control, we compared upregulated 458 Copia elements with the same number of randomly selected Copia elements that did 459 not show a significant change of expression in kyp suvh5 suvh6 (non-DE), or to the 460 same number of randomly positioned loci of the same widths (Fig. 7C). The 461 upregulated Copia transposons showed regions with significant gain of SPO11-1-oligos 462 in kyp suvh5 suvh6 compared to wild type, which overlapped with regions of significant 463 REC8 depletion, close to the transposon promoters (Benjamini-Hochberg adjusted 464 Mann-Whitney-Wilcoxon P<0.1) (Fig. 7C). No significant differences in SPO11-1-oligos 465 or REC8 were detected in the Copia transposons that were not upregulated (Fig. 7C). 466 This is consistent with changes to chromatin and transcription at these transposons 467 associating with modifications to REC8 occupancy and DSB levels. 468 469 Abnormal chromosome axis structures form in rec8 that recruit the homologous 470 recombination machinery 471 472 Finally, we sought to explore the consequences of rec8 mutation for meiotic 473 chromosome architecture and recombination at the cytological level. We used 474 epifluorescence immunocytology and structured illumination microscopy (SIM) to 475 analyze the axis proteins ASY1 and ASY3 in rec8. In wild type, ASY1 and ASY3 co-476

10
localize along linear axes from leptotene until zygotene (Fig. 8A–8D) (Ferdous et al., 477 2012). In rec8, ASY1 and ASY3 occurred in abnormal structures that persisted 478 throughout mid-prophase I (Fig. 8A–8D), as reported (Chelysheva et al., 2005). The 479 length chromosome axis structures stained by ASY1 in rec8 was significantly shorter 480 compared to the wild type at leptotene (mean=30 μm vs. 220 μm, MWW test 481 P=3.02×10-11) (Fig. 8D and Supplemental Table 8). 482 483 We analyzed the SC component ZYP1 in wild type and rec8 (Fig. 8A-8B). 484 Chromosome synapsis initiates at zygotene in wild type, with the formation of ZYP1 485 stretches that become depleted of ASY1 (Fig. 8A), until full synapsis is achieved at 486 pachytene (Lambing et al., 2015). We observed short stretches of ZYP1 polymerization 487 between the abnormal ASY1 axis structures in rec8 (Fig. 8A-8B). In wild type, ZYP1 488 polymerizes between axes separated by a mean distance of 109 nm (Fig. 8B and 489 Supplemental Table 9). In rec8, ZYP1 was detected between ASY1 axis structures with 490 a mean distance not significantly different from wild type (119 nm, MWW test P=0.22) 491 (Fig. 8B, 8D and Supplemental Table 9). PCH2 is a conserved meiotic AAA+ATPase 492 required to remodel the axis during synapsis, which forms a linear signal with ZYP1 at 493 pachytene (Fig. 8E) (Lambing et al., 2015). The rec8 axis structures co-stained for both 494 PCH2 and ZYP1 (Fig. 8E). Interestingly, the SMC3 cohesin subunit was also recruited 495 to the ASY1 axis structures, despite the absence of REC8 (Fig. 8F). Therefore, rec8 496 abnormal axial structures include ASY1, ASY3 and SMC3 and can recruit PCH2 and 497 ZYP1 to produce synapsed structures with a similar inter-axis width to wild type. 498 499 To visualize meiotic DSBs we immunostained for γH2A.X and ASY1 and observed a 500 mean of 202 axis-associated foci in wild type (Fig. 9A, 9C and Supplemental Table 10). 501 In rec8, γH2A.X foci were significantly reduced (mean=53, MWW test P=3.37×10-6), 502 although they remained associated with the ASY1 axis structures (Fig. 9A and 9C). We 503 also immunostained for the ssDNA binding proteins RAD51, RPA1a and DMC1, which 504 show mean axis-associated foci numbers of 181, 174 and 172 respectively at mid-505 prophase I in wild type, which were significantly reduced in rec8 to 39, 36 and 14 506 (MWW tests, P=3.37×10-6, P=3.33×10-6, P=1.08×10-5) (Fig. 9A, 9C and Supplemental 507 Tables 10-11). Positive correlations exist between axis length and γH2A.X and RAD51 508 foci per nuclei, in both wild type and rec8 (wild type rs=0.83 and 0.69, rec8 rs=0.92 and 509 0.77) (Fig. 9D and Supplemental Table 12), which is consistent with a requirement of 510 the axis for DSB formation. This provides cytological evidence that DSB formation and 511 interhomolog strand invasion are associated with abnormal axial structures in rec8 512 mutants. 513 514 Finally, we immunostained for DNA repair factors required for formation of interfering 515 crossovers. In wild type, the MutS homolog MSH4 forms a mean of 179 axis-516 associated foci at leptotene, which were significantly reduced in rec8 (mean=13 foci, 517 MWW test P=1.10×10-5), and to a greater extent than observed for DSB foci (Fig. 9A, 518 9C and Supplemental Tables 10-11). The MutL homolog MLH1 acts in late prophase I 519 and shows a mean of 10.4 chiasma-associated foci at diakinesis, which were 520 significantly reduced in rec8, yet remained associated with chromatin (mean=4.9 foci, 521 MWW test P=2.59×10-10) (Fig. 9C and Supplemental Table 13). Therefore, although 522 rec8 abnormal axis structures recruit recombination foci, they are reduced in number 523 relative to wild type. Together these cytological data support a role for REC8-cohesin in 524 organizing correct polymerization of the axis and SC to promote high-fidelity 525 interhomolog recombination. 526 527 DISCUSSION 528 529

11
At the chromosome scale, we observe greatest REC8 ChIP-seq enrichment in 530 proximity to the centromeres and within pericentromeric heterochromatin. This parallels 531 elevated REC8 enrichment observed in budding and fission yeast centromeres (Folco 532 et al., 2017; Sun et al., 2015; Ito et al., 2014; Kugou et al., 2009). In contrast to these 533 species, Arabidopsis possesses megabase-scale centromeres, consisting of CEN180 534 satellite repeats, a subset of which bind to the centromeric CENH3 histone variant 535 (Table 1) (Maheshwari et al., 2017; Zhang et al., 2008). The CEN180 repeats are 536 surrounded by repetitive heterochromatin enriched in Gypsy, Copia and EnSpm 537 transposable elements, which are epigenetically modified by DNA methylation, 538 H3K9me2 and H2A.W (Stroud et al., 2014; Yelagandula et al., 2014). We previously 539 observed that SPO11-1-oligos increase within heterochromatin in kyp suvh5 suvh6 540 H3K9me2/non-CG DNA methylation mutants (Underwood et al., 2018). Here we show 541 that REC8 is still effectively loaded in the centromeric regions in kyp suvh5 suvh6. In 542 fission yeast, which possesses complex regional centromeres, loss of centromeric 543 H3K9me2 in clr4 mutants greatly decreases REC8 loading and causes defects in 544 chromosome segregation (Bernard et al., 2001; Nonaka et al., 2002; Folco et al., 2017; 545 Ellermeier et al., 2010). Hence, in this respect Arabidopsis more closely resembles 546 mouse suv39h1 suv39h2 H3K9me2 mutants, where mitotic cohesin is recruited to 547 heterochromatin, but with remodeling on major versus minor satellite repeats (Koch et 548 al., 2008; Guenatri et al., 2004). Therefore, it is possible that other heterochromatic 549 marks, including CG DNA methylation, H3K27me1 or H2A.W (Stroud et al., 2013; 550 Yelagandula et al., 2014; Jacob et al., 2009; Stroud et al., 2014), or other features of 551 the centromere including the kinetochore (Hinshaw et al., 2017), could play important 552 roles in REC8 loading in the Arabidopsis centromeres and pericentromeric 553 heterochromatin. 554 555 We observe a strong correlation between REC8 enrichment and nucleosome 556 occupancy (measured via MNase-seq) across the genome. In this respect, it is notable 557 that in vitro binding experiments testing the mobility of a fission yeast cohesin complex 558 (Psm1, Psm3, Psc3, Rad21) along a single tethered DNA molecule showed hindrance 559 caused by the presence of nucleosomes (Stigler et al., 2016). Hence, it is possible that 560 Arabidopsis nucleosomes restrict REC8-cohesin mobility, contributing to the observed 561 correlations. In budding yeast, the Scc2/Scc4 complex functions to load cohesin within 562 nucleosome free gene promoter regions, with cohesin then migrating in the direction of 563 transcription towards gene terminator regions (Lengronne et al., 2004), which is also 564 evident for REC8-cohesin during meiosis (Sun et al., 2015), and during fission yeast 565 mitosis (Schmidt et al., 2009). In mouse cells the absence of CTCF leads to 566 accumulation of cohesin at the 3′ ends of transcribed genes (Busslinger et al., 2017). In 567 Arabidopsis we observe a negative relationship with transcription level and REC8 568 occupancy, with higher levels of transcription associating with a 3′ bias in REC8 569 enrichment, although we do not observe REC8 accumulation in gene terminators. In 570 contrast, Arabidopsis terminators are nucleosome-depleted and enriched for meiotic 571 DSBs (Choi et al., 2018), unlike in budding yeast where SPO11-1-oligo hotspots occur 572 at promoters and not at terminators (Pan et al., 2011). Hence, although a conserved 573 role for transcription patterning cohesin occupancy is apparent, the relative locations of 574 REC8 enrichment and DSBs with respect to gene architecture vary between genomes. 575 It is also important to consider that plant heterochromatin is actively transcribed by Pol 576 IV and Pol V RNA polymerases, which produce short transcripts required for RNA-577 directed DNA methylation (Law and Jacobsen, 2010). Hence, it will also be interesting 578 to explore the effects of heterochromatic transcription on cohesin occupancy in plants 579 in the centromere proximal regions. 580 581 We show that REC8 occupancy is associated with multiple chromatin states within the 582

12
Arabidopsis genome in both gene and repeat rich regions. We show significant 583 differentiation of DNA versus RNA transposon occupancy by REC8, which correlates 584 with chromatin and meiotic DSB levels. At both the chromosome and fine-scales, 585 REC8 enrichment associates with suppression of meiotic DSBs and crossovers. This is 586 consistent with anchoring of chromatin loops to the axis via REC8-cohesin causing 587 local exclusion of the recombination machinery. Cohesin has a role in DSB repair and 588 can antagonize interhomolog recombination by promoting intersister repair of DSBs 589 (Yoon et al., 2016; Hong et al., 2013; da Costa-Nunes et al., 2014; Bolaños-Villegas et 590 al., 2017). Hence, in Arabidopsis it is also possible that high levels of REC8 associate 591 with promotion of inter-sister repair, which could contribute to the low crossover 592 frequency observed in the heterochromatic pericentromeres. Despite the local 593 suppressive effect on recombination, REC8 is also required to establish an organized 594 meiotic chromosome architecture, and in its absence catastrophic recombination 595 occurs leading to chromosome fragmentation (Cai et al., 2003; Chelysheva et al., 596 2005; Bhatt et al., 1999). We show that in the absence of REC8, axis proteins undergo 597 polymerization into disorganised structures, which nevertheless recruit foci associated 598 with DSB formation and repair and form regions of synatonemal complex. We propose 599 that a subset of these repair foci represent illegitimate interhomolog recombination 600 events that cause chromosome fragmentation at metaphase I. In conclusion, we show 601 that chromatin, the axis and REC8-cohesin play tightly integrated roles organizing 602 meiotic chromosome architecture and recombination during Arabidopsis meiosis. 603 604 METHODS 605 606 Plant materials 607 608 Arabidopsis thaliana plants were grown on commercial F2 compost in controlled 609 environment chambers under the following conditions: 60% humidity, long day 610 conditions (16 hours light/8 hours dark) with 150 mol light intensity using LED bulbs 611 with standard NS1 spectrum and at 20C. The following mutant alleles in the Columbia 612 (Col) accession were used: rec8-1 (Salk_091193) was used for cytological 613 experiments (Ferdous et al., 2012), and rec8-3 (SAIL_807_B08) was used for 614 transformation with epitope-tagged REC8-HA and REC8-Myc transgenes (d’Erfurth et 615 al., 2009). The kyp suvh5 suvh6 triple mutant (SALK_041474, GK-263C05, 616 SAIL_1244_F04) was obtained from Steven Jacobsen (UCLA). 617 618 Cytological analysis of meiosis 619 620 Fixation of Arabidopsis inflorescences from pools of Col, rec8-1 or REC8-HA/Myc 621 plants and chromosome spreads of pollen mother cells (PMCs) were performed, as 622 described (Ross et al., 1996). Immunostaining of MLH1, REC8-HA and REC8-Myc was 623 performed on acetic acid chromosome spreads using fixed inflorescences, as 624 described (Chelysheva et al., 2010). Chromosome spreads of Arabidopsis PMCs and 625 immunostaining of ASY1, γH2A.X, RAD51, RPA1a, DMC1, MSH4, ASY3, SMC3, 626 PCH2 and ZYP1 were prepared using fresh buds from pools of Col or rec8-1 plants, as 627 described (Armstrong et al., 2002). The following antibodies were used: α-ASY1 628 (rat/rabbit, 1:500 dilution) (Armstrong et al., 2002), α-ZYP1 (rat/rabbit, 1:500 dilution) 629 (Higgins et al., 2005), α-γH2A.X (Ser 139, Merck Catalog No.07-164, 1:100 dilution) 630 (Ferdous et al., 2012), α-RAD51 (rabbit, 1:500 dilution) (Sanchez-Moran et al., 2007), 631 α-DMC1 (rabbit, 1:500 dilution) (Sanchez-Moran et al., 2007), α-MSH4 (rabbit, 1:500 632 dilution) (Higgins et al., 2004), α-RPA1a (rabbit, 1:500 dilution) (Osman et al., 2009), α-633 ASY3 (rabbit, 1:500 dilution) (Ferdous et al., 2012), α-MLH1 (rabbit, 1:200 dilution) 634 (Chelysheva et al., 2010), α-SMC3 (rabbit, 1:500) (Ferdous et al., 2012), α-PCH2 635

13
(rabbit, 1:500 dilution) (Lambing et al., 2015), α-Myc (mouse, 9E10 sc-40, Santa Cruz 636 Biotechnology, 1:50 dilution) and α-HA (rabbit, ab9110 Abcam, 1:250 dilution). 637 Fluorescence in situ hybridization was carried out on chromosome spreads of 638 metaphase nuclei using CEN180 probes, as described (Armstrong et al., 2001). 639 Microscopy was conducted using a DeltaVision Personal DV microscope (Applied 640 precision/GE Healthcare) equipped with a CDD Coolsnap HQ2 camera (Photometrics). 641 Image capture was performed using SoftWoRx software version 5.5 (Applied 642 precision/GE Healthcare). Image analysis and processing were performed using 643 ImageJ. 644 645 Generation of REC8-3×HA and REC8-5×Myc transgenic lines 646 647 The REC8 locus was PCR amplified from Columbia genomic DNA and 3×HA or 648 5×MYC epitopes introduced at the stop codon. For each of the constructs, a genomic 649 fragment containing 2.6 kb upstream of the REC8 5′-UTR was amplified using the 650 primers REC8-tag_F and REC8-tag_R (Supplemental Table 14). A linker coding for the 651 amino acids (GGGS)×4 was introduced between the last codon of the genomic 652 sequence and the HA and Myc tags. A Nos terminator was introduced at the end of the 653 HA and Myc tags. Tagged REC8 genomic fragments were cloned into the pPZP211 654 binary vector using HindIII and SalI restriction sites and used to transform rec8-3 655 heterozygous plants using Agrobacterium tumefaciens strain GV3101. Primary 656 transformants were selected on kanamycin supplemented MS medium. The presence 657 of transgenes in rec8-3 homozygous backgrounds was confirmed by PCR genotyping. 658 659 Immunoprecipitation and immunoblotting of epitope-tagged REC8 660 661 Unopened flower buds were collected from pools of Col, REC8-HA rec8 or REC8-Myc 662 rec8 F3 plants. For each genotype, one gram of unopened flower buds was ground in 663 liquid nitrogen and transferred to lysis buffer (25 mM Hepes-NaOH pH 7.9, 5 mM EDTA, 664 2% SDS, 1 mM PMSF, 2 mM DTT, 1×protein cocktail inhibitor (Roche, 11873580001)) 665 and incubated for 20 minutes at 95C. The solution was centrifuged at 17,000 × g for 666 20 minutes at 4C. The supernatant was diluted with 3 volumes of dilution buffer (14 667 mM Tris pH 8.0, 1% Triton X-100, 150 mM NaCl). For immunoprecipitation 4 μg of α-668 HA (ab9110, Abcam) or α-Myc (ab9132, Abcam) antibodies were used per 0.25 gram 669 of buds, which were bound to Protein G Dynabeads (Invitrogen, 10004D). The reaction 670 was incubated for 16 hours at 4C and then washed 4 times with wash buffer (14 mM 671 Tris pH 8.0, 1% Triton X-100, 150 mM NaCl). Immunoprecipitated proteins were eluted 672 from the beads by incubation with 1×SDS loading buffer for 5 minutes at 75C. The 673 eluted solution was used for immunoblotting. 674 675 Protein extracts and eluted proteins were loaded with 1×SDS loading buffer on a 676 NuPage 3–8% Tris acetate gel (Invitrogen, EA0375BOX). After proteins were 677 separated through the gel using electrophoresis, proteins were transferred to a PVDF 678 membrane (Bio-Rad, 162-0177) in transfer solution (25 mM Tris base, 192 mM glycine, 679 20% v/v methanol, pH=8.3) for 1 hour at 100 mV at 4C. After transfer, the membrane 680 was rinsed with TBST (20 mM Tris base, 150 mM NaCl, pH 7.6) and incubated in 681 blocking buffer (5% w/v non-fat dried milk in TBST) overnight at 4C with shaking. The 682 membrane was then washed for 1×15 minutes and 2×5 minutes with TBST at room 683 temperature. A solution of primary antibody diluted in blocking buffer was added (α-HA: 684 (Abcam ab9110 lot GR3231414) 1/15,000 dilution; α-Myc: (Abcam ab9132, lot 685 GR254156) 1/5,000 dilution) and the membrane was incubated for 2 hours at room 686 temperature. The membrane was washed 1×15 minutes and then 2×5 minutes with 687 TBST. A solution of secondary antibody diluted in blocking buffer (α-HA: α-rabbit IgG-688

14
HRP sc-2054 (Santa Cruz Biotechnology), 1/20,000 dilution; α-Myc: α-goat IgG-HRP 689 sc-2768 (Santa Cruz Biotechnology), 1/5,000 dilution) was added, and the membrane 690 was incubated for 1 hour at room temperature. The membrane was washed 1×15 691 minutes and 2×5 minutes with TBST at room temperature. The signal was detected on 692 film with ECL Prime Western Blotting detection reagents (GE Healthcare, RPN2232) 693 using a Xograph compact X4. 694 695 Chromatin immunoprecipitation sequencing of histone modifications 696 697 Unopened flower buds were collected from pools of Col or kyp suvh5 suvh6 plants. 698 Two grams of unopened floral buds were used to perform ChIP-seq for H3K4me1, 699 H3K4me2, H3K9me2, H3K27me1 and H3K9me2 in wild type (Col) or kyp suvh5 suvh6, 700 as described (Choi et al., 2018). 5 μg of antibodies were used per gram of tissue. The 701 following antibodies were used: α-H3K4me1 (Abcam, cat: ab8895, lot GR312093), α-702 H3K4me2 (Abcam, cat: ab7766, lot GR289627), α-H3K9me2 (Abcam, cat: ab1220, lot 703 GR244373) and α-H3K27me1 (Merck, cat: 07-448, lot 2869720). 704 705 Chromatin immunoprecipitation and sequencing of REC8 706 707 Unopened flower buds were collected from pools of Col, kyp suvh5 suvh6, REC8-HA 708 rec8 or REC8-Myc rec8 F3 plants. Ten grams of unopened floral buds were ground in 709 liquid nitrogen and used to perform ChIP-seq for REC8-HA in wild type (two biological 710 replicates representing two independent collections of flower buds from pools plants) 711 and kyp suvh5 suvh6 (two biological replicates representing two independent 712 collections of flower buds from pools plants), and for REC8-Myc in wild type (one 713 biological replicate), as described (Lambing et al., 2020). In vitro crosslinking and 714 nuclei isolation were performed in nuclei isolation crosslinking buffer (60 mM Hepes pH 715 8.0, 1 M sucrose, 5 mM KCl, 5 mM MgCl2, 5 mM EDTA, 0.6% Triton X-100, 0.4 mM 716 PMSF, 1 mM pepstatin, 1×protein cocktail inhibitor, 1% formaldehyde) at room 717 temperature for 25 minutes. Glycine was added to a final concentration of 125 mM and 718 the nuclei solution was incubated at room temperature for an additional 25 minutes. 719 The nuclei were purified from cellular debris by filtering the nuclei solution twice 720 through 1 layer of Miracloth and centrifugation at 12,000g for 10 minutes at 4C. The 721 pellet was resuspended in EB2 buffer (0.25 M sucrose, 10 mM Tris-HCl pH 8.0, 10 mM 722 MgCl2, 1% Triton X-100, 1 mM EDTA, 5 mM β-mercaptoethanol, 0.1 mM PMSF, 1 mM 723 pepstatin, 1×protein cocktail inhibitor) followed by a centrifugation at 12,000g for 10 724 minutes at 4C. The supernatant was discarded and the pellet was resuspended in 725 nuclei lysis buffer (50 mM Tris-HCl, pH 8.0, 10 mM EDTA, 1% SDS, 0.1 mM PMSF, 1 726 mM pepstatin, 1× protein cocktail inhibitor). DNA was sonicated using a Bioruptor for 727 15 minutes (High intensity 320W, 30 seconds ON – 30 seconds OFF). The nucleus 728 lysis solution was then diluted (3 volumes:2 volumes) with ChIP dilution buffer (1.1% 729 Triton X-100, 1.1 mM EDTA, 20 mM Tris-HCl pH 8.0, 167 mM NaCl, 1 mM pepstatin, 730 1×protein cocktail inhibitor) and a second cycle of DNA sonication was performed (25 731 minutes at high intensity 320W with 30 seconds ON – 1 minute OFF). The solution was 732 then centrifuged at 5,000g for 10 minutes at 4C. 5% of the supernatant was kept as 733 DNA input. The remaining supernatant was incubated with 7 μg of α-Myc (ab9132 734 Abcam) or 7 μg α-HA (ab9110 Abcam) antibodies that had been prebound to 50 μl of 735 Dynabead Protein G (Invitrogen, 10004D) per gram of floral buds and was then 736 incubated for 16 hours at 4C. The immunoprecipitated solution was washed 2 times 737 for 5 minutes with low salt buffer (150 mM NaCl, 0.1% SDS, 1% Triton X-100, 2 mM 738 EDTA, 20 mM Tris-HCl pH 8.0) and 2 times for 5 minutes with high salt buffer (500 mM 739 NaCl, 0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris-HCl pH 8.0). The proteins 740 bound to the antibody/beads were treated with elution buffer (1% SDS, 0.1 M NaHCO3) 741

15
at 65C for 10 minutes. DNA–protein complexes were reverse crosslinked by adding 742 NaCl to a final concentration of 240 mM and the solution was incubated at 65C 743 overnight. ChIP-DNA was purified with phenol/chloroform and 10 ng of ChIP-purified 744 DNA was used to generate libraries using the TruSeq DNA Sample Preparation Kit v2 745 (Illumina), as described (Choi et al., 2018). 746 747 ChIP-qPCR analysis 748 749 Enrichment of DNA following α-HA ChIP was estimated by quantitative PCR using a 750 QuantiNova SYBR Green PCR kit (Qiagen, 208054) and a CFX96 Bio-Rad qPCR 751 instrument. Briefly, 0.25 ng of DNA were used with 0.7 M of primers for qPCR 752 amplifications. The PCR program used was 40 cycles of; 95C for 2 minutes, 95C for 753 5 seconds and 60C for 10 seconds. To calculate α-HA ChIP enrichment of DNA, 754 amplifications were compared to input DNA, between REC8-HA rec8 transgenic plants 755 and non-transgenic wild-type (Col) plants. 756 757 RNA sequencing 758 759 Unopened flower buds were collected from pools of Col or kyp suvh5 suvh6 plants to 760 provide two biological replicates for each genotype. For each replicate, 100 mg of 761 unopened flower buds were ground in liquid nitrogen and RNA was extracted using 762 Trizol reagent. 6 μg of total RNA per sample were treated with DNaseI using TURBO 763 DNase reagents (ThermoFisher, AM2238). 4 μg of total DNaseI-treated total RNA was 764 then treated with Ribo-Zero rRNA Removal Kit (Epicentre, MRZPL116) to deplete 765 rRNA from the RNA sample. 50 ng of rRNA-depleted DNaseI-treated RNA were used 766 to prepare RNA-seq libraries using the ScriptSeq v2 RNA-seq Library Preparation Kit 767 (Epicentre, SSV21124), as described (Choi et al., 2018). 768 769 ChIP-seq data analysis 770 771 Deduplicated paired-end (2×76 bp) ChIP-seq reads were aligned to the TAIR10 772 reference genome using Bowtie2 (Version 2.2.9) (Langmead and Salzberg, 2012), with 773 the following settings: --very sensitive --no-discordant --no-mixed -p 4 -k 10. Up to 10 774 valid alignments were reported for each read. Aligned reads with more than 2 775 mismatches were discarded using the SAM optional field “XM:i”. Uniquely aligning 776 reads were extracted by removing alignments with the SAM optional field “XS:i” and 777 with Bowtie2-assigned MAPQ scores lower than 42. Alignments consisting of reads 778 that mapped to multiple loci were filtered such that only those with MAPQ scores 779 higher than or equal to 10 remained, from which the alignment with the highest MAPQ 780 score was retained. Where MAPQ scores for multiple valid alignments were equal, one 781 alignment was randomly selected. Alignments consisting of only one read in a pair 782 were discarded. The genome-wide average depth of coverage obtained for each library 783 is provided in Supplemental Table 2. 784 785 Unique and multiple alignments in BAM format were combined and coverage was 786 calculated for each coordinate in the genome using Rsamtools (Version 1.26.1). 787 Coverage was normalized by the sum of coverage for each library. The log2 ratio of 788 ChIP:input coverage was calculated to control for background and variation in 789 mappability across genomic loci. These values were used to generate chromosome-790 scale coverage profiles and fine-scale coverage profiles around genome features 791 within specific annotation categories, including REC8-HA peaks (defined as described 792 below), well-positioned nucleosomes (Choi et al. 2018), SPO11-1-oligo hotspots (Choi 793 et al. 2018), TAIR10 representative gene transcription start and termination sites (TSSs 794

16
and TTSs), transposable elements (Buisine et al., 2008) and a set of 3,320 crossover 795 intervals mapped using genotyping-by-sequencing of Col×Ler F2 plants (Choi et al., 796 2018; Serra et al., 2018). A coverage profile was generated for each feature using the 797 normalizeToMatrix function from the Bioconductor package EnrichedHeatmap (Version 798 1.11.1) (Gu et al., 2018). Due to varying feature lengths, each feature was divided into 799 proportionally scaled windows between start and end coordinates, and 2 kb flanking 800 regions were divided into 20 bp windows or 5 kb regions flanking crossovers were 801 divided into 50 bp windows. For each window along each feature and its flanking 802 regions, an average value was calculated using the “w0” method for ChIP-seq data or 803 the “absolute” method for bisulfite-seq data. The default profile-smoothing method 804 implemented in the normalizeToMatrix function was applied. The resulting matrix of 805 windowed coverage or DNA methylation proportion values was used to generate a 806 mean profile (averaged over all features), or a heat map in which each row represents 807 a single feature. Mean profiles and heat maps were plotted such that the distance 808 between feature start and end coordinates along the x-axis represents the mean 809 feature length. Previously published paired-end H3K4me3 ChIP-seq and MNase-seq 810 reads in wild type (Choi et al., 2018), and single-end SPO11-1-oligonucleotide reads in 811 wild type and kyp suvh5 suvh6 (Choi et al., 2018; Underwood et al., 2018) were 812 aligned to the TAIR10 reference genome and processed for downstream analysis, as 813 described (Choi et al., 2018). 814 815 REC8, H3K4me3, H3K9me2 and SPO11-1-oligo peaks were identified using the 816 ranger tool within the PeakRanger suite (Feng et al., 2011) with the --pad option 817 specified, providing ChIP input or gDNA reads as a control for background in each 818 case. H3K4me3 and H3K9me2 peaks were defined by applying P-value and false 819 discovery rate (FDR) thresholds of ≤0.05, with read extension lengths (--ext_length 820 option) of 125 and 150 nucleotides specified, respectively, based on mean estimated 821 fragment lengths derived from BAM alignment files. REC8 ChIP-seq peaks were 822 detected using more stringent significance thresholds (P ≤ 0.001 and FDR ≤ 0.01) and 823 with a read extension length of 200 nucleotides specified. SPO11-1-oligo hotspots 824 were identified as previously described (Choi et al., 2018). Any overlapping peaks were 825 merged. 826 827 REC8-HA peaks and SPO11-1-oligo hotspots were evaluated for overlap with genomic 828 features within other annotation categories by performing permutation tests using the 829 Bioconductor package regioneR (Version 1.6.2) (Gel et al., 2016). Overlaps were 830 defined as the number of REC8-HA peaks or SPO11-1-oligo hotspots that overlap one 831 or more features within a given annotation category. For each test, 10,000 sets of 832 randomly positioned loci with the same width distribution as the REC8-HA peaks or 833 SPO11-1-oligo hotspots were defined. For each set, the number of random loci 834 overlapping features within a given annotation category was compared with the 835 observed number of overlapping REC8-HA peaks or SPO11-1-oligo hotspots. This 836 provided the basis for calculating an empirical P value (≥1×10-4, with the minimum 837 obtainable P value determined by the number of permutations) denoting the 838 significance of the observed overlap. 839 840 Well-positioned nucleosomes were identified in mapped MNase-seq data (Choi et al., 841 2018), using the R package nucleR (Flores and Orozco, 2011). Well-positioned 842 nucleosomes are defined narrow peaks in nucleosomal signal corresponding to the 843 size of a single nucleosome core (~147 bp) and characterized by troughs in signal in 844 flanking regions. As nucleR identifies nucleosome positions from paired-end 100-bp 845 MNase-seq reads these were trimmed to the central 40 bp to improve detection. 846 Coverage at each position in the genome was calculated and normalized by library size. 847

17
Paired-end reads from a Col gDNA library were processed in the same way to provide 848 a control. These data were used to calculate log2(MNase-seq/gDNA) coverage ratios at 849 each genomic coordinate. To remove noise from the log2(MNase-seq/gDNA) profile, 850 the fast Fourier transform was applied using the filterFFT function within nucleR, 851 retaining 2% of the components of the original profile (pcKeepComp = 0.02). Peaks in 852 the signal were identified using the peakDetection function, specifying a peak width of 853 140 bp centered on the local maximum. Peaks with nucleR-assigned height scores 854 greater than 0.99 were retained. Any overlapping peaks were merged. Additionally, 855 DNA methylation proportions derived from published bisulfite sequencing reads in wild 856 type and kyp suvh5 suvh6 were used to profile DNA methylation levels at chromosome 857 and fine scales (Stroud et al., 2013). 858
859 RNA-seq data analysis 860
861 For analysis of chromosome-scale and fine-scale expression profiles, paired-end (2×76 862 bp) RNA-seq reads were aligned to the TAIR10 reference genome using STAR 863 (Version 2.5.3a) (Dobin et al., 2013), with the following settings: --runThreadN 24 --864 outFilterMultimapNmax 10 --outFilterMismatchNmax 2 --outMultimapperOrder Random 865 --outSAMattributes All --twopassMode Basic --twopass1readsN -1. Read pairs with up 866 to 10 valid alignments and up to 2 mismatches were reported as mapped. Uniquely 867 aligned reads were extracted by selecting alignments with STAR-assigned MAPQ 868 scores of 255. Alignments consisting of reads that mapped to multiple loci were 869 extracted using the SAM field “NH:i”. Multiple alignments with MAPQ scores lower than 870 3 were discarded. As the maximum STAR-assigned MAPQ score for multiple 871 alignments is 3, a primary multiple alignment was randomly selected. Alignments 872 consisting of only one read in a pair were discarded. Unique and multiple alignments 873 were combined and coverage was calculated for each coordinate in the genome using 874 Rsamtools (Version 1.26.1). Coverage was normalized by the total coverage for each 875 library. Analysis of differential expression between wild type and kyp suvh5 suvh6 876 mutant samples was performed using the Bioconductor package DESeq2 version 877 1.16.1 (Love et al., 2014), as described (Lawrence et al., 2019). TEs that were 878 upregulated in kyp suvh5 suvh6 relative to wild type were evaluated for over-879 representation of elements within each TE superfamily using the hypergeometric 880 distribution. 881
882 Accession numbers 883
884 The gene accession numbers for REC8, KYP, SUVH5 and SUVH6 are AT5G05490, 885 AT5G13960, AT2G35160, AT2G22740, respectively. ChIP-seq and RNA-seq data 886 have been deposited in the ArrayExpress database at EMBL-EBI 887 (www.ebi.ac.uk/arrayexpress) under accession numbers E-MTAB-7370 (ChIP-seq) and 888 E-MTAB-7371 (RNA-seq), respectively.889 Supplemental Data890
891 Supplemental Figure 1. Detection of REC8-Myc fusion proteins via immunoblotting 892 and immunostaining during meiosis. 893
894 Supplemental Figure 2. Overlap of REC8-HA peaks with other genome features. 895
896 Supplemental Table 1. Chiasma counts from wild-type (Col) and REC8-HA rec8 male 897 meiocytes at metaphase I. 898
899

18
Supplemental Table 2. Aligned reads from chromatin immunoprecipitation sequencing900 and RNA sequencing libraries. 901
902 Supplemental Table 3. Correlations between REC8-HA and REC8-Myc ChIP libraries 903 at varying physical scales. 904
905 Supplemental Table 4. ChIP-qPCR analysis of peak enrichment in REC8-HA rec8 and 906 wild-type (Col) flowers. 907
908 Supplemental Table 5. Overlap of REC8-HA peaks with other genome features in 909 chromosome arms. 910
911 Supplemental Table 6. Overlap of REC8-HA peaks with other genome features in the 912 pericentromeres. 913
914 Supplemental Table 7. Overrepresentation analysis of transposable element (TE) 915 superfamilies that are transcriptionally upregulated in kyp suvh5 ssuvh6. 916
917 Supplemental Table 8. ASY1 axis length measurements in wild type (Col) and rec8. 918
919 Supplemental Table 9. Inter-axis width measurements between synapsed ZYP1 920 structures in wild type (Col) and rec8. 921
922 Supplemental Table 10. Recombination foci counts of γH2A.X, RAD51 and RPA1a in 923 wild type (Col) and rec8 mid-prophase I nuclei. 924
925 Supplemental Table 11. DMC1 and MSH4 recombination foci counted in wild type 926 (Col) and rec8 during mid-prophase I. 927
928 Supplemental Table 12. Correlations between axis length and recombination foci in 929 wild type (Col) and rec8. 930
931 Supplemental Table 13. MLH1 foci counts in wild type (Col) and rec8 nuclei at late 932 prophase I. 933
934 Supplemental Table 14. Oligonucleotide sequences. 935
936 Acknowledgements 937
938 We thank Mathilde Grelon for the MLH1 antibody and the Gurdon Institute for access 939 to microscopes. Research was supported by grants from the ERC (SynthHotSpot) and 940 BBSRC (BB/L006847/1, BB/N007557/1 and BB/N002628/1), and an EMBO long term 941 postdoctoral fellowship (ALTF 807-2009). The authors declare no competing interests. 942
943 Author Contributions 944
945 CL, AJT, KC, PCK, KO, JDH, FCHF and IRH designed the research; CL, SDT, PCK 946 and JDH performed research; CL, AJT, XZ, KO, JDH, FCHF and IRH analyzed data; 947 CL, AJT, KO, JDH, FCHF and IRH wrote the paper. 948
949 950

19
Table 1. Genome architecture, chromatin and meiotic recombination in Arabidopsis, 951 budding yeast and fission yeast. 952
953
Species Genome size (Mb)
Chromosome number Genes
Transposable Elements
DNA transposable
elements
RNA transposable
elements
Schizosaccharomyces pombe 12.6 3 5,130 187 None. TF2 LTR
Saccharomyces cerevisiae 12.1 16 6,002 50 None. Ty1-Ty5 LTR
Arabidopsis thaliana 119.2 5 27,655 3,901
Ac/Ds, CACTA/En-SPM, Mu, Helitron,
Pogo/Tc1/Mariner
Gypsy, Copia, SINE, LINE1
Species Heterochromatin Euchromatin Centromere Polycomb/ H3K27me3
Schizosaccharomyces pombe H3K9me3
H3K4me1, H3K4me2, H3K4me3, H3K36me3, H2A.Z, H3/H4-Acetyl
Complex (35, 65 & 110
kb) No
Saccharomyces cerevisiae Sir2-mediated deacetylation
H3K4me1, H3K4me2, H3K4me3, H3K36me3, H2A.Z, H3/H4-Acetyl
Point (120 bp) No
Arabidopsis thaliana H3K9me2, H3K27me1, DNA
CG CHG CHH, H2A.W
H3K4me1, H3K4me2, H3K4me3, H3K36me3, H2A.Z, H3/H4-Acetyl
Complex (~1-5 Mb) Yes
Species cM cM/Mb ZMM Interference
distance Pachytene axis
μM
Schizosaccharomyces pombe 2,200 174.2 - - -
Saccharomyces cerevisiae 4,420 366.2
ZIP2, ZIP3, ZIP4, SPO16, MER3, MSH4, MSH5 Megabases 34
Arabidopsis thaliana 398 3.3 HEI10, SHOC1, PTD1, MER3, MSH4, MSH5 Megabases 179
954 Table 2. Genomic maps of chromatin, transcription and meiotic recombination in 955 Arabidopsis thaliana. The tissue collected for data generation is listed. 956
957 Dataset Reference Tissue
REC8 ChIP-seq This study Floral buds
H3K9me2 ChIP-seq This study Floral buds
SPO11-1-oligos Choi et al., 2018 Floral buds
Nucleosome MNase-seq Choi et al., 2018 Floral buds
H3K4me1 ChIP-seq This study Floral buds
H3K4me2 ChIP-seq This study Floral buds
H3K4me3 ChIP-seq Choi et al., 2018 Floral buds
DNA methylation BS-seq Stroud et al., 2013 Seedlings
siRNA-seq Lee et al., 2012 Immature inflorescences
Flower mRNA RNA-seq Choi et al., 2018 Floral buds
Meiocyte mRNA RNA-seq Walker et al., 2018 Floral buds
H3K27me3 ChIP-seq Zhu et al., 2015 Floral buds
Crossovers Rowan et al., 2019 Seedlings
958 959 960 961
REFERENCES: 962 963
Acquaviva, L., Székvölgyi, L., Dichtl, B., Dichtl, B.S., de La Roche Saint André, C., 964 Nicolas, A., and Géli, V. (2013). The COMPASS subunit Spp1 links histone 965 methylation to initiation of meiotic recombination. Science 339: 215–8. 966

20
Armstrong, S. (2013). A Time Course for the Analysis of Meiotic Progression in 967 Arabidopsis thaliana. Methods Mol. Biol. 990: 119–123. 968
Armstrong, S.J., Caryl, A.P., Jones, G.H., and Franklin, F.C.H. (2002). Asy1, a 969 protein required for meiotic chromosome synapsis, localizes to axis-associated 970 chromatin in Arabidopsis and Brassica. J. Cell Sci. 115: 3645–55. 971
Armstrong, S.J., Franklin, F.C., and Jones, G.H. (2001). Nucleolus-associated 972 telomere clustering and pairing precede meiotic chromosome synapsis in 973 Arabidopsis thaliana. J. Cell Sci. 114: 4207–17. 974
Barton, N.H. and Charlesworth, B. (1998). Why sex and recombination? Cold Spring 975 Harb. Symp. Quant. Biol. 281: 187–95. 976
Baudat, F., Imai, Y., and de Massy, B. (2013). Meiotic recombination in mammals: 977 localization and regulation. Nat. Rev. Genet. 14: 794–806. 978
Bernard, P., Maure, J.F., Partridge, J.F., Genier, S., Javerzat, J.P., and Allshire, 979 R.C. (2001). Requirement of Heterochromatin for Cohesion at Centromeres.980 Science (80-. ). 294: 2539–2542.981
Bhatt, A.M., Lister, C., Page, T., Fransz, P., Findlay, K., Jones, G.H., Dickinson, 982 H.G., and Dean, C. (1999). The DIF1 gene of Arabidopsis is required for meiotic983 chromosome segregation and belongs to the REC8/RAD21 cohesin gene family.984 Plant J. 19: 463–72.985
Blat, Y., Protacio, R.U., Hunter, N., and Kleckner, N. (2002). Physical and functional 986 interactions among basic chromosome organizational features govern early steps 987 of meiotic chiasma formation. Cell 111: 791–802. 988
Bolaños-Villegas, P., De, K., Pradillo, M., Liu, D., and Makaroff, C.A. (2017). In 989 Favor of Establishment: Regulation of Chromatid Cohesion in Plants. Front. Plant 990 Sci. 8: 846. 991
Buisine, N., Quesneville, H., and Colot, V. (2008). Improved detection and 992 annotation of transposable elements in sequenced genomes using multiple 993 reference sequence sets. Genomics 91: 467–75. 994
Busslinger, G.A., Stocsits, R.R., van der Lelij, P., Axelsson, E., Tedeschi, A., 995 Galjart, N., and Peters, J.-M. (2017). Cohesin is positioned in mammalian 996 genomes by transcription, CTCF and Wapl. Nature 544: 503–507. 997
Cai, X., Dong, F., Edelmann, R.E., and Makaroff, C.A. (2003). The Arabidopsis SYN1 998 cohesin protein is required for sister chromatid arm cohesion and homologous 999 chromosome pairing. J. Cell Sci. 116: 2999–3007. 1000
Chambon, A., West, A., Vezon, D., Horlow, C., De Muyt, A., Chelysheva, L., 1001 Ronceret, A., Darbyshire, A.R., Osman, K., Heckmann, S., Franklin, F.C.H., 1002 and Grelon, M. (2018). Identification of ASYNAPTIC4, a component of the 1003 meiotic chromosome axis. Plant Physiol. 178: 233–246. 1004
Chelysheva, L. et al. (2005). AtREC8 and AtSCC3 are essential to the monopolar 1005 orientation of the kinetochores during meiosis. J. Cell Sci. 118: 4621–32. 1006
Chelysheva, L., Grandont, L., Vrielynck, N., le Guin, S., Mercier, R., and Grelon, M. 1007 (2010). An easy protocol for studying chromatin and recombination protein 1008 dynamics during Arabidopsis thaliana meiosis: immunodetection of cohesins, 1009 histones and MLH1. Cytogenet. Genome Res. 129: 143–53. 1010
Choi, K. et al. (2018). Nucleosomes and DNA methylation shape meiotic DSB 1011 frequency in Arabidopsis thaliana transposons and gene regulatory regions. 1012 Genome Res. 28: 532–546. 1013
Choi, K. et al. (2016). Recombination Rate Heterogeneity within Arabidopsis Disease 1014 Resistance Genes. PLoS Genet. 12: e1006179. 1015
Choi, K. and Henderson, I.R. (2015). Meiotic recombination hotspots - a comparative 1016 view. Plant J. 83: 52–61. 1017
Choi, K., Zhao, X., Kelly, K.A., Venn, O., Higgins, J.D., Yelina, N.E., Hardcastle, 1018 T.J., Ziolkowski, P.A., Copenhaver, G.P., Franklin, F.C.H., Mcvean, G., and1019

21
Henderson, I.R. (2013). Arabidopsis meiotic crossover hot spots overlap with 1020 H2A.Z nucleosomes at gene promoters. Nat. Genet. 45: 1327–36. 1021
da Costa-Nunes, J.A., Capitão, C., Kozak, J., Costa-Nunes, P., Ducasa, G.M., 1022 Pontes, O., and Angelis, K.J. (2014). The AtRAD21.1 and AtRAD21.3 1023 Arabidopsis cohesins play a synergistic role in somatic DNA double strand break 1024 damage repair. BMC Plant Biol. 14: 353. 1025
d’Erfurth, I., Jolivet, S., Froger, N., Catrice, O., Novatchkova, M., and Mercier, R. 1026 (2009). Turning Meiosis into Mitosis. PLoS Biol. 7: e1000124. 1027
Dobin, A., Davis, C.A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., Batut, P., 1028 Chaisson, M., and Gingeras, T.R. (2013). STAR: ultrafast universal RNA-seq 1029 aligner. Bioinformatics 29: 15–21. 1030
Ellermeier, C., Higuchi, E.C., Phadnis, N., Holm, L., Geelhood, J.L., Thon, G., and 1031 Smith, G.R. (2010). RNAi and heterochromatin repress centromeric meiotic 1032 recombination. Proc. Natl. Acad. Sci. U. S. A. 107: 8701–5. 1033
Feng, X., Grossman, R., and Stein, L. (2011). PeakRanger: a cloud-enabled peak 1034 caller for ChIP-seq data. BMC Bioinformatics 12: 139. 1035
Ferdous, M., Higgins, J.D., Osman, K., Lambing, C., Roitinger, E., Mechtler, K., 1036 Armstrong, S.J., Perry, R., Pradillo, M., Cuñado, N., and Franklin, F.C.H. 1037 (2012). Inter-homolog crossing-over and synapsis in Arabidopsis meiosis are 1038 dependent on the chromosome axis protein AtASY3. PLoS Genet. 8: e1002507. 1039
Flores, O. and Orozco, M. (2011). nucleR: a package for non-parametric nucleosome 1040 positioning. Bioinformatics 27: 2149–50. 1041
Folco, H.D., Chalamcharla, V.R., Sugiyama, T., Thillainadesan, G., Zofall, M., 1042 Balachandran, V., Dhakshnamoorthy, J., Mizuguchi, T., and Grewal, S.I.S. 1043 (2017). Untimely expression of gametogenic genes in vegetative cells causes 1044 uniparental disomy. Nature 543: 126–130. 1045
Fowler, K.R., Sasaki, M., Milman, N., Keeney, S., and Smith, G.R. (2014). 1046 Evolutionarily diverse determinants of meiotic DNA break and recombination 1047 landscapes across the genome. Genome Res. 24: 1650–64. 1048
Gel, B., Díez-Villanueva, A., Serra, E., Buschbeck, M., Peinado, M.A., and 1049 Malinverni, R. (2016). regioneR: an R/Bioconductor package for the association 1050 analysis of genomic regions based on permutation tests. Bioinformatics 32: 289–1051 91. 1052
Glynn, E.F., Megee, P.C., Yu, H.-G., Mistrot, C., Unal, E., Koshland, D.E., DeRisi, 1053 J.L., and Gerton, J.L. (2004). Genome-wide mapping of the cohesin complex in1054 the yeast Saccharomyces cerevisiae. PLoS Biol. 2: E259.1055
Gu, Z., Eils, R., Schlesner, M., and Ishaque, N. (2018). EnrichedHeatmap: an 1056 R/Bioconductor package for comprehensive visualization of genomic signal 1057 associations. BMC Genomics 19: 234. 1058
Guenatri, M., Bailly, D., Maison, C., and Almouzni, G. (2004). Mouse centric and 1059 pericentric satellite repeats form distinct functional heterochromatin. J. Cell Biol. 1060 166: 493–505. 1061
He, Y. et al. (2017). Genomic features shaping the landscape of meiotic double-strand-1062 break hotspots in maize. Proc. Natl. Acad. Sci. U. S. A. 114: 12231–12236. 1063
Higgins, J.D., Armstrong, S.J., Franklin, F.C.H., and Jones, G.H. (2004). The 1064 Arabidopsis MutS homolog AtMSH4 functions at an early step in recombination: 1065 evidence for two classes of recombination in Arabidopsis. Genes Dev. 18: 2557–1066 70. 1067
Higgins, J.D., Sanchez-Moran, E., Armstrong, S.J., Jones, G.H., and Franklin, 1068 F.C.H. (2005). The Arabidopsis synaptonemal complex protein ZYP1 is required1069 for chromosome synapsis and normal fidelity of crossing over. Genes Dev. 19:1070 2488–2500.1071
Hinshaw, S.M., Makrantoni, V., Harrison, S.C., and Marston, A.L. (2017). The 1072

22
Kinetochore Receptor for the Cohesin Loading Complex. Cell 171: 72–84. 1073 Hong, S., Sung, Y., Yu, M., Lee, M., Kleckner, N., and Kim, K.P.P. (2013). The logic 1074
and mechanism of homologous recombination partner choice. Mol. Cell 51: 440–1075 453. 1076
Ito, M., Kugou, K., Fawcett, J.A., Mura, S., Ikeda, S., Innan, H., and Ohta, K. (2014). 1077 Meiotic recombination cold spots in chromosomal cohesion sites. Genes to Cells 1078 19: 359–373. 1079
Jacob, Y., Feng, S., LeBlanc, C.A., Bernatavichute, Y. V, Stroud, H., Cokus, S., 1080 Johnson, L.M., Pellegrini, M., Jacobsen, S.E., and Michaels, S.D. (2009). 1081 ATXR5 and ATXR6 are H3K27 monomethyltransferases required for chromatin 1082 structure and gene silencing. Nat. Struct. Mol. Biol. 16: 763–8. 1083
Kauppi, L., Jeffreys, A.J., and Keeney, S. (2004). Where the crossovers are: 1084 recombination distributions in mammals. Nat. Rev. Genet. 5: 413–24. 1085
Kianian, P.M.A. et al. (2018). High-resolution crossover mapping reveals similarities 1086 and differences of male and female recombination in maize. Nat. Commun. 9: 1087 2370. 1088
Kim, K.P., Weiner, B.M., Zhang, L., Jordan, A., Dekker, J., and Kleckner, N. (2010). 1089 Sister cohesion and structural axis components mediate homolog bias of meiotic 1090 recombination. Cell 143: 924–37. 1091
Kitajima, T.S., Miyazaki, Y., Yamamoto, M., and Watanabe, Y. (2003). Rec8 1092 cleavage by separase is required for meiotic nuclear divisions in fission yeast. 1093 EMBO J. 22: 5643–53. 1094
Klein, F., Mahr, P., Galova, M., Buonomo, S.B., Michaelis, C., Nairz, K., and 1095 Nasmyth, K. (1999). A central role for cohesins in sister chromatid cohesion, 1096 formation of axial elements, and recombination during yeast meiosis. Cell 98: 91–1097 103. 1098
Koch, B., Kueng, S., Ruckenbauer, C., Wendt, K.S., and Peters, J.-M. (2008). The 1099 Suv39h–HP1 histone methylation pathway is dispensable for enrichment and 1100 protection of cohesin at centromeres in mammalian cells. Chromosoma 117: 199–1101 210. 1102
Kugou, K., Fukuda, T., Yamada, S., Ito, M., Sasanuma, H., Mori, S., Katou, Y., Itoh, 1103 T., Matsumoto, K., Shibata, T., Shirahige, K., and Ohta, K. (2009). Rec8 guides 1104 canonical Spo11 distribution along yeast meiotic chromosomes. Mol. Biol. Cell 20: 1105 3064–76. 1106
Laloraya, S., Guacci, V., and Koshland, D. (2000). Chromosomal addresses of the 1107 cohesin component Mcd1p. J. Cell Biol. 151: 1047–56. 1108
Lam, I. and Keeney, S. (2014). Mechanism and Regulation of Meiotic Recombination 1109 Initiation. Cold Spring Harb. Perspect. Biol. 7: a016634. 1110
Lambing, C., Choi, K., Blackwell, A.R., and Henderson, I.R. (2020). Chromatin 1111 Immunoprecipitation of Meiotically Expressed Proteins from Arabidopsis thaliana 1112 Flowers. Methods Mol. Biol. 2061: 219–236. 1113
Lambing, C., Osman, K., Nuntasoontorn, K., West, A., Higgins, J.D., Copenhaver, 1114 G.P., Yang, J., Armstrong, S.J., Mechtler, K., Roitinger, E., and Franklin,1115 F.C.H. (2015). Arabidopsis PCH2 Mediates Meiotic Chromosome Remodeling and1116 Maturation of Crossovers. PLoS Genet. 11: e1005372.1117
Lange, J., Yamada, S., Tischfield, S.E., Pan, J., Kim, S., Zhu, X., Socci, N.D., Jasin, 1118 M., and Keeney, S. (2016). The Landscape of Mouse Meiotic Double-Strand 1119 Break Formation, Processing, and Repair. Cell 167: 695-708.e16. 1120
Langmead, B. and Salzberg, S.L. (2012). Fast gapped-read alignment with Bowtie 2. 1121 Nat. Methods 9: 357–9. 1122
Law, J.A. and Jacobsen, S.E. (2010). Establishing, maintaining and modifying DNA 1123 methylation patterns in plants and animals. Nat. Rev. Genet. 11: 204–220. 1124
Lawrence, E.J., Gao, H., Tock, A.J., Lambing, C., Blackwell, A.R., Feng, X., and 1125

23
Henderson, I.R. (2019). Natural Variation in TBP-ASSOCIATED FACTOR 4b 1126 Controls Meiotic Crossover and Germline Transcription in Arabidopsis. Curr. Biol. 1127 29: 2676–2686. 1128
Lee, T., Gurazada, S.G.R., Zhai, J., Li, S., Simon, S.A., Matzke, M.A., Chen, X., and 1129 Meyers, B.C. (2012). RNA polymerase V-dependent small RNAs in Arabidopsis 1130 originate from small, intergenic loci including most SINE repeats. Epigenetics 7: 1131 781–795. 1132
Lengronne, A., Katou, Y., Mori, S., Yokobayashi, S., Kelly, G.P., Itoh, T., Watanabe, 1133 Y., Shirahige, K., and Uhlmann, F. (2004). Cohesin relocation from sites of 1134 chromosomal loading to places of convergent transcription. Nature 430: 573–578. 1135
Liu, S., Yeh, C.-T., Ji, T., Ying, K., Wu, H., Tang, H.M., Fu, Y., Nettleton, D., and 1136 Schnable, P.S. (2009). Mu Transposon Insertion Sites and Meiotic 1137 Recombination Events Co-Localize with Epigenetic Marks for Open Chromatin 1138 across the Maize Genome. PLoS Genet. 5: e1000733. 1139
Love, M.I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change 1140 and dispersion for RNA-seq data with DESeq2. Genome Biol. 15: 550. 1141
Maheshwari, S., Ishii, T., Brown, C.T., Houben, A., and Comai, L. (2017). 1142 Centromere location in Arabidopsis is unaltered by extreme divergence in CENH3 1143 protein sequence. Genome Res. 27: 471–478. 1144
Mancera, E., Bourgon, R., Brozzi, A., Huber, W., and Steinmetz, L.M. (2008). High-1145 resolution mapping of meiotic crossovers and non-crossovers in yeast. Nature 1146 454: 479–85. 1147
de Massy, B. (2013). Initiation of meiotic recombination: how and where? 1148 Conservation and specificities among eukaryotes. Annu. Rev. Genet. 47: 563–99. 1149
Mercier, R., Mézard, C., Jenczewski, E., Macaisne, N., and Grelon, M. (2015). The 1150 molecular biology of meiosis in plants. Annu. Rev. Plant Biol. 66: 297–327. 1151
Mizuguchi, T., Fudenberg, G., Mehta, S., Belton, J.-M., Taneja, N., Folco, H.D., 1152 FitzGerald, P., Dekker, J., Mirny, L., Barrowman, J., and Grewal, S.I.S. (2014). 1153 Cohesin-dependent globules and heterochromatin shape 3D genome architecture 1154 in S. pombe. Nature 516: 432–435. 1155
Neale, M.J., Pan, J., and Keeney, S. (2005). Endonucleolytic processing of covalent 1156 protein-linked DNA double-strand breaks. Nature 436: 1053–7. 1157
Nonaka, N., Kitajima, T., Yokobayashi, S., Xiao, G., Yamamoto, M., Grewal, S.I.S., 1158 and Watanabe, Y. (2002). Recruitment of cohesin to heterochromatic regions by 1159 Swi6/HP1 in fission yeast. Nat. Cell Biol. 4: 89–93. 1160
Osman, K., Sanchez-Moran, E., Mann, S.C., Jones, G.H., and Franklin, F.C.H. 1161 (2009). Replication protein A (AtRPA1a) is required for class I crossover formation 1162 but is dispensable for meiotic DNA break repair. EMBO J. 28: 394–404. 1163
Osman, K., Yang, J., Roitinger, E., Lambing, C., Heckmann, S., Howell, E., Cuacos, 1164 M., Imre, R., Dürnberger, G., Mechtler, K., Armstrong, S., and Franklin, F.C.H. 1165 (2018). Affinity proteomics reveals extensive phosphorylation of the Brassica 1166 chromosome axis protein ASY1 and a network of associated proteins at prophase 1167 I of meiosis. Plant J. 93: 17–33. 1168
Page, S.L. and Hawley, R.S. (2003). Chromosome choreography: the meiotic ballet. 1169 Science 301: 785–9. 1170
Pan, J., Sasaki, M., Kniewel, R., Murakami, H., Blitzblau, H.G., Tischfield, S.E., 1171 Zhu, X., Neale, M.J., Jasin, M., Socci, N.D., Hochwagen, A., and Keeney, S. 1172 (2011). A hierarchical combination of factors shapes the genome-wide topography 1173 of yeast meiotic recombination initiation. Cell 144: 719–31. 1174
Ross, K.J., Fransz, P., and Jones, G.H. (1996). A light microscopic atlas of meiosis in 1175 Arabidopsis thaliana. Chromosome Res. 4: 507–16. 1176
Rowan, B.A., Heavens, D., Feuerborn, T.R., Tock, A.J., Henderson, I.R., and 1177 Weigel, D. (2019). An Ultra High-Density Arabidopsis thaliana Crossover Map 1178

24
That Refines the Influences of Structural Variation and Epigenetic Features. 1179 Genetics: doi: 10.1534/genetics.119.302406. 1180
Sanchez-Moran, E., Santos, J.-L., Jones, G.H., and Franklin, F.C.H. (2007). ASY1 1181 mediates AtDMC1-dependent interhomolog recombination during meiosis in 1182 Arabidopsis. Genes Dev. 21: 2220–33. 1183
Schmidt, C.K., Brookes, N., and Uhlmann, F. (2009). Conserved features of cohesin 1184 binding along fission yeast chromosomes. Genome Biol. 10: R52. 1185
Segal, E. and Widom, J. (2009). Poly(dA:dT) tracts: major determinants of 1186 nucleosome organization. Curr. Opin. Struct. Biol. 19: 65–71. 1187
Serra, H., Lambing, C., Griffin, C.H., Topp, S.D., Seguela-Arnaud, M., Fernandes, 1188 J., Mercier, R., and Henderson, I.R. (2018). Massive crossover elevation via 1189 combination of HEI10 and recq4a recq4b during Arabidopsis meiosis. Proc. Natl. 1190 Acad. Sci. U. S. A. 115: 2437–2442. 1191
Sommermeyer, V., Béneut, C., Chaplais, E., Serrentino, M.E., and Borde, V. (2013). 1192 Spp1, a member of the Set1 Complex, promotes meiotic DSB formation in 1193 promoters by tethering histone H3K4 methylation sites to chromosome axes. Mol. 1194 Cell 49: 43–54. 1195
Stigler, J., Çamdere, G.Ö., Koshland, D.E., and Greene, E.C. (2016). Single-1196 Molecule Imaging Reveals a Collapsed Conformational State for DNA-Bound 1197 Cohesin. Cell Rep. 15: 988–998. 1198
Stroud, H., Do, T., Du, J., Zhong, X., Feng, S., Johnson, L., Patel, D.J., and 1199 Jacobsen, S.E. (2014). Non-CG methylation patterns shape the epigenetic 1200 landscape in Arabidopsis. Nat. Struct. Mol. Biol. 21: 64–72. 1201
Stroud, H., Greenberg, M.V.C., Feng, S., Bernatavichute, Y. V, and Jacobsen, S.E. 1202 (2013). Comprehensive analysis of silencing mutants reveals complex regulation 1203 of the Arabidopsis methylome. Cell 152: 352–64. 1204
Sun, X., Huang, L., Markowitz, T.E., Blitzblau, H.G., Chen, D., Klein, F., and 1205 Hochwagen, A. (2015). Transcription dynamically patterns the meiotic 1206 chromosome-axis interface. Elife 4: doi: 10.7554/eLife.07424. 1207
Topp, C.N. and Dawe, R.K. (2006). Reinterpreting pericentromeric heterochromatin. 1208 Curr. Opin. Plant Biol. 9: 647–653. 1209
Underwood, C.J., Choi, K., Lambing, C., Zhao, X., Serra, H., Borges, F., 1210 Simorowski, J., Ernst, E., Jacob, Y., Henderson, I.R., and Martienssen, R.A. 1211 (2018). Epigenetic activation of meiotic recombination near Arabidopsis thaliana 1212 centromeres via loss of H3K9me2 and non-CG DNA methylation. Genome Res. 1213 28: 519–531. 1214
Villeneuve, A.M. and Hillers, K.J. (2001). Whence meiosis? Cell 106: 647–50. 1215 Vincenten, N., Kuhl, L.-M., Lam, I., Oke, A., Kerr, A.R., Hochwagen, A., Fung, J., 1216
Keeney, S., Vader, G., and Marston, A.L. (2015). The kinetochore prevents 1217 centromere-proximal crossover recombination during meiosis. Elife 4: doi: 1218 10.7554/eLife.10850. 1219
Walker, J., Gao, H., Zhang, J., Aldridge, B., Vickers, M., Higgins, J.D., and Feng, X. 1220 (2018). Sexual-lineage-specific DNA methylation regulates meiosis in Arabidopsis. 1221 Nat. Genet. 50: 130–137. 1222
Watanabe, Y. and Nurse, P. (1999). Cohesin Rec8 is required for reductional 1223 chromosome segregation at meiosis. Nature 400: 461–4. 1224
West, A.M., Rosenberg, S.C., Ur, S.N., Lehmer, M.K., Ye, Q., Hagemann, G., 1225 Caballero, I., Usón, I., MacQueen, A.J., Herzog, F., and Corbett, K.D. (2019). A 1226 conserved filamentous assembly underlies the structure of the meiotic 1227 chromosome axis. Elife 8: doi: 10.7554/eLife.40372. 1228
Wijnker, E. et al. (2013). The genomic landscape of meiotic crossovers and gene 1229 conversions in Arabidopsis thaliana. Elife 2: e01426. 1230
Yelagandula, R. et al. (2014). The histone variant H2A.W defines heterochromatin 1231

25
and promotes chromatin condensation in Arabidopsis. Cell 158: 98–109. 1232 Yelina, N.E., Lambing, C., Hardcastle, T.J., Zhao, X., Santos, B., and Henderson, 1233
I.R. (2015). DNA methylation epigenetically silences crossover hot spots and1234 controls chromosomal domains of meiotic recombination in Arabidopsis. Genes1235 Dev. 29: 2183–202.1236
Yoon, S.-W., Lee, M.-S., Xaver, M., Zhang, L., Hong, S.-G., Kong, Y.-J., Cho, H.-R., 1237 Kleckner, N., and Kim, K.P. (2016). Meiotic prophase roles of Rec8 in crossover 1238 recombination and chromosome structure. Nucleic Acids Res. 44: 9296–9314. 1239
Zhang, W., Lee, H.-R., Koo, D.-H., and Jiang, J. (2008). Epigenetic modification of 1240 centromeric chromatin: hypomethylation of DNA sequences in the CENH3-1241 associated chromatin in Arabidopsis thaliana and maize. Plant Cell 20: 25–34. 1242
Zhu, B., Zhang, W., Zhang, T., Liu, B., and Jiang, J. (2015). Genome-Wide 1243 Prediction and Validation of Intergenic Enhancers in Arabidopsis Using Open 1244 Chromatin Signatures. Plant Cell 27: 2415–2426. 1245
Zickler, D. and Kleckner, N. (1999). Meiotic chromosomes: integrating structure and 1246 function. Annu. Rev. Genet. 33: 603–754. 1247
1248 FIGURE LEGENDS 1249
1250 Figure 1. REC8-cohesin, chromatin and the synaptonemal complex during 1251 Arabidopsis meiosis. A. DAPI-stained spreads of wild-type (Col), rec8, REC8-HA 1252 rec8 and REC8-Myc rec8 male meiocytes, at the labeled stages of meiosis. B. Male 1253 meiocytes from wild type or REC8-HA rec8 immunostained for REC8-HA (red) and 1254 stained for DNA (DAPI, blue), at the labeled stages. C. Male meiocytes immunostained 1255 for ZYP1 (green), histone methylation (red, H3K4me3, H3K9me2 or H3K27me1) and 1256 stained for DNA (DAPI, blue) in wild type (Col), during late prophase I. D. Male 1257 meiocytes at pachytene stage immunostained for SMC3 (red) or ZYP1 (red) and 1258 stained for DNA (DAPI, blue). Fluorescence in situ hybridization (FISH) was performed 1259 against the CEN180 centromeric satellite sequence (green). Inset images show 1260 magnifications of the CEN180-positive regions. All scale bars=10 μm. 1261
1262 Figure 2. Profiling the genomic landscape of Arabidopsis REC8 by ChIP-seq. A. 1263 Immunoblot of wild-type (Col) and REC8-HA rec8 floral extracts before (input) and after 1264 α-HA immunoprecipitation (IP). Ponceau staining of the input lane is shown. The 1265 predicted molecular weight of REC8-HA is 77.2 kDa (*). Higher molecular weight 1266 REC8-HA bands are indicated (**). B. ChIP enrichment (% input) from REC8-HA rec8 1267 and untagged wild-type (Col) flowers measured via qPCR at the ATHILA, ATHILA6B 1268 and At2g02480 loci. Error bars show the standard deviation. C. A representative region 1269 from chromosome 3 showing chromatin and recombination data (Table 2). Transposon 1270 (red) and gene (green) annotation are shown, with elements of interest highlighted by 1271 gray shading and labeled beneath. 1272
1273 Figure 3. Genome-wide landscapes of REC8, heterochromatin, euchromatin and 1274 meiotic recombination. A. Genome-wide profiles of REC8-HA (red, log2(ChIP/input) 1275 red) and nucleosomes (purple, log2(MNase/gDNA), purple). Vertical solid lines indicate 1276 telomeres and dashed lines indicate centromeres, with the pericentromeres shaded in 1277 blue. Plots beneath compare REC8-HA (red) with transposable elements (dark green, 1278 TE/10kb), H3K9me2 (green, log2(ChIP/input)), DNA methylation (blue, proportion 1279 methylated in CG, CHG and CHH contexts/200 kb) (Stroud et al., 2013), genes (brown, 1280 genes/10kb), H3K4me3 (yellow, log2(ChIP/input)), SPO11-1-oligos (light blue, 1281 log2(oligos/gDNA)), crossovers (dark blue, crossovers/10 kb) and REC8-Myc (blue, 1282 log2(ChIP/input)). B. Correlation matrices showing genome-wide Spearman’s rank-1283 order correlation coefficients (rs) for the indicated parameter pairs, with cells color-1284

26
coded according to the correlation scale shown to the right. C. REC8-HA (red) and 1285 chromatin (blue/white, DAPI) were co-stained on pachytene stage chromosomes and 1286 20 axis sections of 100 pixels centered on heterochromatin were used to quantify 1287 signal intensity (arbitrary units). A representative full cell image is shown beneath 1288 (scale bar=10 μM). Arrows indicate the area of the cell used for the close-up image 1289 beneath, dashed line indicate the section of chromatin used for analysis (scale bar=2.5 1290 μM). 1291
1292 Figure 4. REC8-HA and nucleosome enrichment correlate with suppression of 1293 meiotic recombination at the fine scale. A. Mean coverage profiles for REC8-HA 1294 (red, log2(ChIP/input)), SPO11-1-oligos (blue, log2(oligos/gDNA)) and nucleosomes 1295 (purple, log2(MNase/gDNA)) within REC8-HA peaks and 2-kb flanking regions, or the 1296 same number of random positions of the same widths. Plots are repeated for well-1297 positioned nucleosomes, SPO11-1-oligo hotspots and crossovers (5-kb flanking 1298 regions). Plot ribbons denote 95% confidence intervals for windowed values. B. Heat 1299 maps of REC8-HA (log2(ChIP/input)), SPO11-1-oligos (log2(oligos/gDNA)) and 1300 nucleosomes (log2(MNase/gDNA)) within REC8-HA peaks and 2-kb flanking regions, 1301 with peaks (rows) ordered by descending REC8-HA values. Analysis was repeated 1302 according to peak locations in the chromosome arms (left) or pericentromeres (right). C. 1303 Plots as for A., but analyzing DNA base frequencies (AT=green; GC=pink) across the 1304 same regions in 4 kb or 10 kb regions around feature midpoints. 1305
1306 Figure 5. REC8, chromatin and recombination within Arabidopsis transposable 1307 element families. A. REC8-HA (red, log2(ChIP/input)), SPO11-1-oligos (blue, 1308 log2(oligos/gDNA)), nucleosomes (purple log2(MNase/gDNA)) and H3K9me2 (green, 1309 log2(ChIP/input)) within DNA transposons and 2 kb flanking regions, or the same 1310 number of random positions of the same widths. The mean width of the elements is 1311 indicated by vertical dashed lines. Beneath are plots analyzing the same transposons 1312 for mean DNA methylation proportion in CG (red), CHG (purple) and CHH (blue) 1313 sequence contexts. Plot ribbons denote 95% confidence intervals for windowed mean 1314 coverage or DNA methylation proportion. B. As for A, but analyzing RNA transposons. 1315 C. Bar graphs showing permutation-test derived log2(observed/expected) overlap of1316 REC8-HA peaks (red), SPO11-1-oligo hotspots (blue) or nucleosomes (purple) with the1317 indicated transposon superfamilies. Vertical gray lines mark significance thresholds1318 (α=0.05). D. A representative region from chromosome 3 showing chromatin,1319 recombination and genome annotation (Table 2). Transposon (red) and gene (green)1320 annotation are shown, with transposons of interest labeled and highlighted by gray1321 shading.1322
1323 Figure 6. REC8 occupancy in heterochromatin is maintained in kyp suvh5 suvh6. 1324 A. Genome-wide profiles of REC8-HA (log2(ChIP/input)) from wild type (red) and kyp1325 suvh5 suvh6 (blue). CHG context DNA methylation proportion from wild type (red) and1326 kyp suvh5 suvh6 (blue) are plotted beneath. Vertical solid lines indicate telomeres and1327 dotted lines indicate centromeres. B. Male meiocytes of wild type (Col) and kyp suvh51328 suvh6 were immunostained for REC8-HA (red) and stained for DNA (DAPI, blue) at1329 pachytene stage. White arrows indicate DAPI-dense chromocenters. All scale bars=101330 μm. C. DAPI-stained spreads of wild type (Col) and kyp suvh5 suvh6 male meiocytes1331 at the labeled stages of meiosis.1332
1333 Figure 7. Transcription shapes REC8 occupancy. A. Mean coverage profiles for 1334 REC8-HA (red, log2(ChIP/input)) between gene transcriptional start sites (TSS) and 1335 termination sites (TTS) and 2-kb flanking regions, or the same number of random 1336 regions of the same widths. Also plotted are SPO11-1-oligos (blue, log2(oligos/gDNA)), 1337

27
nucleosomes (purple, log2(MNase/gDNA)), RNA expression (pink, RNA-seq 1338 normalized by total coverage), H2A.Z (turquoise, log2(ChIP/input)) and H3K4me3 1339 (yellow, log2(ChIP/input)). B. Heat maps of REC8-HA, nucleosomes, SPO11-1-oligos, 1340 RNA-seq, H3K4me3 and H2A.Z within genes and 2-kb flanking regions. Rows have 1341 been ordered by decreasing RNA-seq within genes. C. Mean profiles in wild type and 1342 kyp suvh5 suvh6 for RNA-seq (purple), H3K9me2 (green, ChIP), CHG DNA 1343 methylation (yellow), REC8-HA (red) and SPO11-1-oligos (blue) within Copia 1344 transposons that are upregulated in kyp suvh5 suvh6, or a random subset of those that 1345 are not, or the same number of random positions of the same widths. Plot ribbons 1346 denote 95% confidence intervals for the windowed mean coverage or DNA methylation 1347 proportion values. Windows showing significant changes in kyp suvh5 suvh6 (with 1348 Benjamini-Hochberg-adjusted Mann-Whitney-Wilcoxon P<0.1) are indicated by colored 1349 ticks along the x-axis. 1350 1351 Figure 8. Abnormal axis structures in rec8 undergo synapsis. A. SIM images of 1352 wild-type (Col) and rec8 male meiocytes at zygotene stage, stained for ASY1 (red), 1353 ZYP1 (green) and chromatin (DAPI, blue). Scale bars=10 μm. Close-ups of synapsed 1354 regions are shown for each genotype where scale bars=2.5 μm. B. As for A., but 1355 showing synapsed regions stained with ASY1 (red) and ZYP1 (green) in wild type (Col) 1356 and rec8, with interaxis distance indicated in white. Scale bars=200 nm. C. As for A., 1357 but at leptotene stage and staining for ASY1 (green), ASY3 (red) and chromatin (DAPI, 1358 blue). D. Quantification of ASY1 axis length (μm) and inter-axis width (nm) in wild type 1359 (black) and rec8 (red). E. As for A., but pachytene stage cells stained for PCH2 (red), 1360 ZYP1 (green) and chromatin (DAPI, blue). F. As for A., but at leptotene stage and 1361 stained for ASY1 (green), SMC3 (red) and chromatin (DAPI, blue). Scale bars=10 μm. 1362 1363 Figure 9. Abnormal axis structures in rec8 recruit DSB and crossover foci. A. 1364 Wild-type (Col) and rec8 male meiocytes at leptotene stage were stained for ASY1 1365 (green), γH2A.X (red) and DNA (DAPI, blue). Also shown are male meiocytes stained 1366 for RPA1a, RAD51, DMC1 and MSH4 (all red) and ASY1 (green). B. Diakinesis-stage 1367 meiocytes from wild type and rec8 stained for MLH1 (red) and DNA (DAPI, blue). All 1368 scale bars=10 μm. C. γH2A.X, RPA1a, RAD51, DMC1, MSH4 and MLH1 foci number 1369 measured in wild type (black) and rec8 (red). D. Plot of ASY1 axis length (μm) against 1370 foci number of γH2A.X (black) and RAD51 (red) in wild type and rec8 meiocytes. 1371 1372

Figure 1. REC8-cohesin, chromatin and the synaptonemal complex during Arabidopsis meiosis. A. DAPI-stained spreads of wild type (Col), rec8, REC8-HA rec8 and REC8-Myc rec8 male meiocytes, at the labeled stages of meiosis. B. Male meiocytes from wild type or REC8-HA rec8 immunostained for REC8-HA (red) and stained for DNA (DAPI, blue), at the labeled stages. C. Male meiocytes immunostained for ZYP1 (green), histone methylation (red, H3K4me3, H3K9me2 or H3K27me1) and stained for DNA (DAPI, blue) in wild type (Col), during late prophase I. D. Male meiocytes at pachytene stage immunostained for SMC3 (red) or ZYP1 (red) and stained for DNA (DAPI, blue). Fluorescence in situ hybridization (FISH) was performed against the CEN180 centromeric satellite sequence (green). Inset images show zooms of the CEN180-positive regions. All scale bars=10 μm.
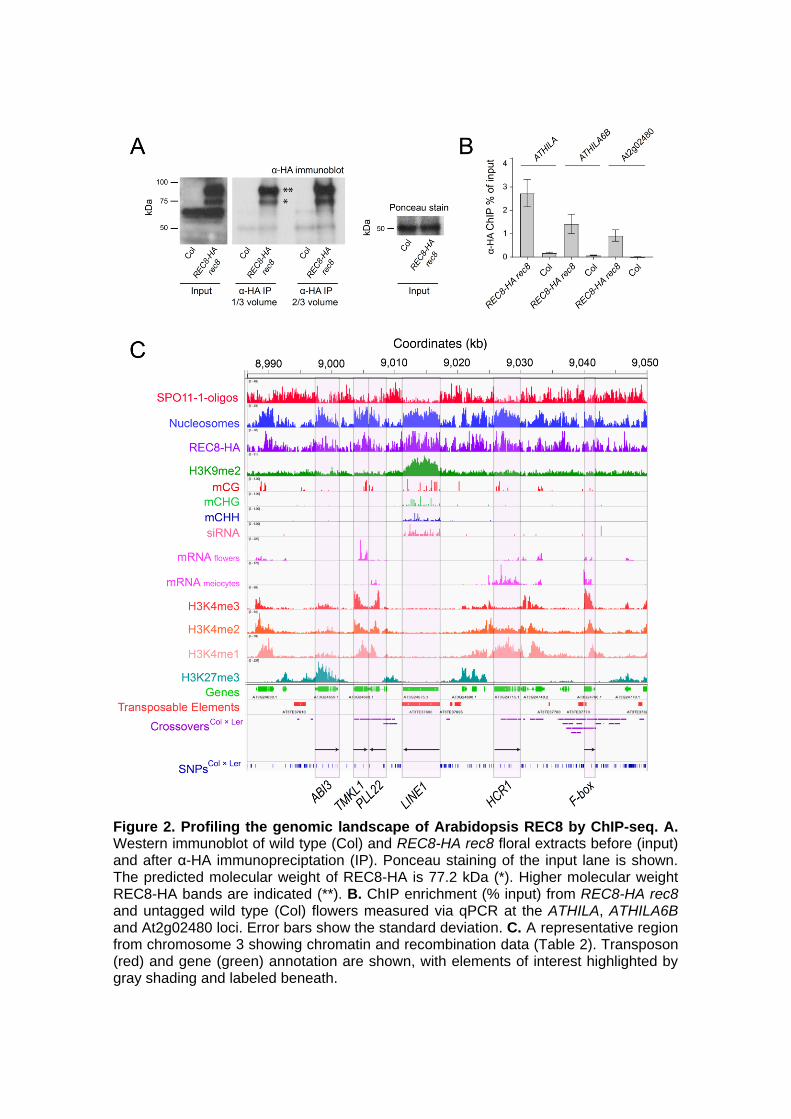
Figure 2. Profiling the genomic landscape of Arabidopsis REC8 by ChIP-seq. A. Western immunoblot of wild type (Col) and REC8-HA rec8 floral extracts before (input) and after α-HA immunopreciptation (IP). Ponceau staining of the input lane is shown. The predicted molecular weight of REC8-HA is 77.2 kDa (*). Higher molecular weight REC8-HA bands are indicated (**). B. ChIP enrichment (% input) from REC8-HA rec8 and untagged wild type (Col) flowers measured via qPCR at the ATHILA, ATHILA6B and At2g02480 loci. Error bars show the standard deviation. C. A representative region from chromosome 3 showing chromatin and recombination data (Table 2). Transposon (red) and gene (green) annotation are shown, with elements of interest highlighted by gray shading and labeled beneath.

Figure 3. Genome-wide landscapes of REC8, heterochromatin, euchromatin and meiotic recombination. A. Genome-wide profiles of REC8-HA (red, log2(ChIP/input) red) and nucleosomes (purple, log2(MNase/gDNA), purple). Vertical solid lines indicate telomeres and dashed lines indicate centromeres, with the pericentromeres shaded in blue. Plots beneath compare REC8-HA (red) with transposable elements (dark green, TE/10kb), H3K9me2 (green, log2(ChIP/input)), DNA methylation (blue, proportion methylated in CG, CHG and CHH contexts/200 kb) (Stroud et al., 2013), genes (brown, genes/10kb), H3K4me3 (yellow, log2(ChIP/input)), SPO11-1-oligos (light blue, log2(oligos/gDNA)), crossovers (dark blue, crossovers/10 kb) and REC8-Myc (blue, log2(ChIP/input)). B. Correlation matrices showing genome-wide Spearman’s rank-order correlation coefficients (rs) for the indicated parameter pairs, with cells color-coded according to the correlation scale shown to the right. C. REC8-HA (red) and

2
chromatin (blue/white, DAPI) were co-stained on pachytene stage chromosomes and 20 axis sections of 100 pixels centered on heterochromatin were used to quantify signal intensity (arbitrary units). A representative full cell image is shown beneath (scale bar=10 μM). Arrows indicate the area of the cell used for the close-up image beneath, showing a section of chromatin used for analysis (scale bar=2.5 μM).

Figure 4. REC8-HA, nucleosomes, meiotic recombination and DNA sequence at the fine scale. A. Mean coverage profiles for REC8-HA (red, log2(ChIP/input)), SPO11-1-oligos (blue, log2(oligos/gDNA)) and nucleosomes (purple, log2(MNase/gDNA)) within REC8-HA peaks and 2 kb flanking regions, or the same number of random positions of the same widths. Plots are repeated for well-positioned nucleosomes, SPO11-1-oligo hotspots and crossovers (5 kb flanking regions). Plot ribbons denote 95% confidence intervals for windowed values. B. Heat maps of REC8-HA (log2(ChIP/input)), SPO11-1-oligos (log2(oligos/gDNA)) and nucleosomes (log2(MNase/gDNA)) within REC8-HA peaks and 2 kb flanking regions, with peaks (rows) ordered by descending REC8-HA values. Analysis was repeated according to peak locations in the chromosome arms (left) or pericentromeres (right). C. Plots as for A., but analyzing DNA base frequencies (AT=green; GC=pink) across the same regions in 4 kb or 10 kb regions around feature midpoints.

Figure 5. REC8, chromatin and recombination within Arabidopsis transposable elements families. A. REC8-HA (red, log2(ChIP/input)), SPO11-1-oligos (blue, log2(oligos/gDNA)), nucleosomes (purple log2(MNase/gDNA)) and H3K9me2 (green, log2(ChIP/input)) within DNA transposons and 2 kb flanking regions, or the same number of random positions of the same widths. The mean width of the elements is indicated by vertical dashed lines. Beneath are plots analyzing the same transposons for mean DNA methylation proportion in CG (red), CHG (purple) and CHH (blue) sequence contexts. Plot ribbons denote 95% confidence intervals for windowed mean coverage or DNA methylation proportion. B. As for A, but analyzing RNA transposons. C. Bar graphs showing permutation-test derived log2(observed/expected) overlap of REC8-HA peaks (red), SPO11-1-oligo hotspots (blue) or nucleosomes (purple) with the indicated transposon superfamilies. Vertical gray lines mark significance thresholds (α=0.05). D. A representative region from chromosome 3 showing chromatin, recombination and genome annotation (Table 2). Transposon (red) and gene (green) annotation are shown, with transposons of interest labeled and highlighted by gray shading.

Figure 6. REC8 occupancy in heterochromatin is maintained in kyp suvh5 suvh6. A. Genome-wide profiles of REC8-HA (log2(ChIP/input)) from wild type (red) and kypsuvh5 suvh6 (blue). CHG context DNA methylation proportion from wild type (red) andkyp suvh5 suvh6 (blue) are plotted beneath. Vertical solid lines indicate telomeres anddotted lines indicate centromeres. B. Male meiocytes of wild type (Col) and kyp suvh5suvh6 were immunostained for REC8-HA (red) and stained for DNA (DAPI, blue) atpachytene stage. White arrows indicate DAPI-dense chromocenters. All scale bars=10μm. C. DAPI-stained spreads of wild type (Col) and kyp suvh5 suvh6 male meiocytesat the labeled stages of meiosis.

Figure 7. Transcription shapes REC8 occupancy. A. Mean coverage profiles for REC8-HA (red, log2(ChIP/input)) between gene transcriptional start sites (TSS) and termination sites (TTS) and 2 kb flanking regions, or the same number of random regions of the same widths. Also plotted are SPO11-1-oligos (blue, log2(oligos/gDNA)), nucleosomes (purple, log2(MNase/gDNA)), RNA expression (pink, RNA-seq normalized by total coverage), H2A.Z (turquoise, log2(ChIP/input)) and H3K4me3 (yellow, log2(ChIP/input)). B. Heat maps of REC8-HA, nucleosomes, SPO11-1-oligos, RNA-seq, H3K4me3 and H2A.Z within genes and 2 kb flanking regions. Rows have

1
been ordered by decreasing RNA-seq within genes. C. Mean profiles in wild type and kyp suvh5 suvh6 for RNA-seq (purple), H3K9me2 (green, ChIP), CHG DNA methylation (yellow), REC8-HA (red) and SPO11-1-oligos (blue) within Copia transposons that are upregulated in kyp suvh5 suvh6, or a random subset of those that are not, or the same number of random positions of the same widths. Plot ribbons denote 95% confidence intervals for the windowed mean coverage or DNA methylation proportion values. Windows showing significant changes in kyp suvh5 suvh6 (with Benjamini-Hochberg-adjusted Mann-Whitney-Wilcoxon P<0.1) are indicated by colored ticks along the x-axis.

Figure 8. Abnormal axis structures in rec8 undergo synapsis. A. SIM images of wild type (Col) and rec8 male meiocytes at zygotene stage, stained for ASY1 (red), ZYP1 (green) and chromatin (DAPI, blue). Scale bars=10 μm. Close-ups of synapsed regions are shown for each genotype where scale bars=2.5 μm. B. As for A., but showing synapsed regions stained with ASY1 (red) and ZYP1 (green) in wild type (Col) and rec8, with interaxis distance indicated in white. Scale bars=200 nm. C. As for A., but at leptotene stage and staining for ASY1 (green), ASY3 (red) and chromatin (DAPI, blue). D. Quantification of ASY1 axis length (μm) and inter-axis width (nm) in wild type (black) and rec8 (red). E. As for A., but pachytene stage cells stained for PCH2 (red), ZYP1 (green) and chromatin (DAPI, blue). F. As for A., but at leptotene stage and stained for ASY1 (green), SMC3 (red) and chromatin (DAPI, blue). Scale bars=10 μm.

Figure 9. Abnormal axis structures in rec8 recruit DSB and crossover foci. A. Wild type (Col) and rec8 male meiocytes at leptotene stage were stained for ASY1 (green), γH2A.X (red) and DNA (DAPI, blue). Also shown are male meiocytes stained for RPA1a, RAD51, DMC1 and MSH4 (all red) and ASY1 (green). B. Diakinesis-stage meiocytes from wild type and rec8 stained for MLH1 (red) and DNA (DAPI, blue). All scale bars=10 μm. C. γH2A.X, RPA1a, RAD51, DMC1, MSH4 and MLH1 foci number measured in wild type (black) and rec8 (red). D. Plot of ASY1 axis length (μm) against foci number of γH2A.X (black) and RAD51 (red) in wild type and rec8 meiocytes.

Parsed CitationsAcquaviva, L., Székvölgyi, L., Dichtl, B., Dichtl, B.S., de La Roche Saint André, C., Nicolas, A., and Géli, V. (2013). The COMPASSsubunit Spp1 links histone methylation to initiation of meiotic recombination. Science 339: 215–8.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Armstrong, S. (2013). A Time Course for the Analysis of Meiotic Progression in Arabidopsis thaliana. Methods Mol. Biol. 990: 119–123.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Armstrong, S.J., Caryl, A.P., Jones, G.H., and Franklin, F.C.H. (2002). Asy1, a protein required for meiotic chromosome synapsis,localizes to axis-associated chromatin in Arabidopsis and Brassica. J. Cell Sci. 115: 3645–55.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Armstrong, S.J., Franklin, F.C., and Jones, G.H. (2001). Nucleolus-associated telomere clustering and pairing precede meioticchromosome synapsis in Arabidopsis thaliana. J. Cell Sci. 114: 4207–17.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Barton, N.H. and Charlesworth, B. (1998). Why sex and recombination? Cold Spring Harb. Symp. Quant. Biol. 281: 187–95.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Baudat, F., Imai, Y., and de Massy, B. (2013). Meiotic recombination in mammals: localization and regulation. Nat. Rev. Genet. 14: 794–806.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bernard, P., Maure, J.F., Partridge, J.F., Genier, S., Javerzat, J.P., and Allshire, R.C. (2001). Requirement of Heterochromatin forCohesion at Centromeres. Science (80-. ). 294: 2539–2542.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bhatt, A.M., Lister, C., Page, T., Fransz, P., Findlay, K., Jones, G.H., Dickinson, H.G., and Dean, C. (1999). The DIF1 gene of Arabidopsisis required for meiotic chromosome segregation and belongs to the REC8/RAD21 cohesin gene family. Plant J. 19: 463–72.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Blat, Y., Protacio, R.U., Hunter, N., and Kleckner, N. (2002). Physical and functional interactions among basic chromosomeorganizational features govern early steps of meiotic chiasma formation. Cell 111: 791–802.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bolaños-Villegas, P., De, K., Pradillo, M., Liu, D., and Makaroff, C.A. (2017). In Favor of Establishment: Regulation of ChromatidCohesion in Plants. Front. Plant Sci. 8: 846.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Buisine, N., Quesneville, H., and Colot, V. (2008). Improved detection and annotation of transposable elements in sequenced genomesusing multiple reference sequence sets. Genomics 91: 467–75.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Busslinger, G.A., Stocsits, R.R., van der Lelij, P., Axelsson, E., Tedeschi, A., Galjart, N., and Peters, J.-M. (2017). Cohesin is positionedin mammalian genomes by transcription, CTCF and Wapl. Nature 544: 503–507.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Cai, X., Dong, F., Edelmann, R.E., and Makaroff, C.A. (2003). The Arabidopsis SYN1 cohesin protein is required for sister chromatid armcohesion and homologous chromosome pairing. J. Cell Sci. 116: 2999–3007.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chambon, A., West, A., Vezon, D., Horlow, C., De Muyt, A., Chelysheva, L., Ronceret, A., Darbyshire, A.R., Osman, K., Heckmann, S.,Franklin, F.C.H., and Grelon, M. (2018). Identification of ASYNAPTIC4, a component of the meiotic chromosome axis. Plant Physiol. 178:233–246.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chelysheva, L. et al. (2005). AtREC8 and AtSCC3 are essential to the monopolar orientation of the kinetochores during meiosis. J. CellSci. 118: 4621–32.
Pubmed: Author and Title

Google Scholar: Author Only Title Only Author and Title
Chelysheva, L., Grandont, L., Vrielynck, N., le Guin, S., Mercier, R., and Grelon, M. (2010). An easy protocol for studying chromatin andrecombination protein dynamics during Arabidopsis thaliana meiosis: immunodetection of cohesins, histones and MLH1. Cytogenet.Genome Res. 129: 143–53.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Choi, K. et al. (2018). Nucleosomes and DNA methylation shape meiotic DSB frequency in Arabidopsis thaliana transposons and generegulatory regions. Genome Res. 28: 532–546.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Choi, K. et al. (2016). Recombination Rate Heterogeneity within Arabidopsis Disease Resistance Genes. PLoS Genet. 12: e1006179.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Choi, K. and Henderson, I.R. (2015). Meiotic recombination hotspots - a comparative view. Plant J. 83: 52–61.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Choi, K., Zhao, X., Kelly, K.A., Venn, O., Higgins, J.D., Yelina, N.E., Hardcastle, T.J., Ziolkowski, P.A., Copenhaver, G.P., Franklin, F.C.H.,Mcvean, G., and Henderson, I.R. (2013). Arabidopsis meiotic crossover hot spots overlap with H2A.Z nucleosomes at gene promoters.Nat. Genet. 45: 1327–36.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
da Costa-Nunes, J.A., Capitão, C., Kozak, J., Costa-Nunes, P., Ducasa, G.M., Pontes, O., and Angelis, K.J. (2014). The AtRAD21.1 andAtRAD21.3 Arabidopsis cohesins play a synergistic role in somatic DNA double strand break damage repair. BMC Plant Biol. 14: 353.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
d'Erfurth, I., Jolivet, S., Froger, N., Catrice, O., Novatchkova, M., and Mercier, R. (2009). Turning Meiosis into Mitosis. PLoS Biol. 7:e1000124.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dobin, A., Davis, C.A., Schlesinger, F., Drenkow, J., Zaleski, C., Jha, S., Batut, P., Chaisson, M., and Gingeras, T.R. (2013). STAR:ultrafast universal RNA-seq aligner. Bioinformatics 29: 15–21.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ellermeier, C., Higuchi, E.C., Phadnis, N., Holm, L., Geelhood, J.L., Thon, G., and Smith, G.R. (2010). RNAi and heterochromatin represscentromeric meiotic recombination. Proc. Natl. Acad. Sci. U. S. A. 107: 8701–5.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Feng, X., Grossman, R., and Stein, L. (2011). PeakRanger: a cloud-enabled peak caller for ChIP-seq data. BMC Bioinformatics 12: 139.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ferdous, M., Higgins, J.D., Osman, K., Lambing, C., Roitinger, E., Mechtler, K., Armstrong, S.J., Perry, R., Pradillo, M., Cuñado, N., andFranklin, F.C.H. (2012). Inter-homolog crossing-over and synapsis in Arabidopsis meiosis are dependent on the chromosome axisprotein AtASY3. PLoS Genet. 8: e1002507.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Flores, O. and Orozco, M. (2011). nucleR: a package for non-parametric nucleosome positioning. Bioinformatics 27: 2149–50.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Folco, H.D., Chalamcharla, V.R., Sugiyama, T., Thillainadesan, G., Zofall, M., Balachandran, V., Dhakshnamoorthy, J., Mizuguchi, T., andGrewal, S.I.S. (2017). Untimely expression of gametogenic genes in vegetative cells causes uniparental disomy. Nature 543: 126–130.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fowler, K.R., Sasaki, M., Milman, N., Keeney, S., and Smith, G.R. (2014). Evolutionarily diverse determinants of meiotic DNA break andrecombination landscapes across the genome. Genome Res. 24: 1650–64.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gel, B., Díez-Villanueva, A., Serra, E., Buschbeck, M., Peinado, M.A., and Malinverni, R. (2016). regioneR: an R/Bioconductor packagefor the association analysis of genomic regions based on permutation tests. Bioinformatics 32: 289–91.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

Glynn, E.F., Megee, P.C., Yu, H.-G., Mistrot, C., Unal, E., Koshland, D.E., DeRisi, J.L., and Gerton, J.L. (2004). Genome-wide mapping ofthe cohesin complex in the yeast Saccharomyces cerevisiae. PLoS Biol. 2: E259.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Gu, Z., Eils, R., Schlesner, M., and Ishaque, N. (2018). EnrichedHeatmap: an R/Bioconductor package for comprehensive visualizationof genomic signal associations. BMC Genomics 19: 234.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Guenatri, M., Bailly, D., Maison, C., and Almouzni, G. (2004). Mouse centric and pericentric satellite repeats form distinct functionalheterochromatin. J. Cell Biol. 166: 493–505.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
He, Y. et al. (2017). Genomic features shaping the landscape of meiotic double-strand-break hotspots in maize. Proc. Natl. Acad. Sci. U.S. A. 114: 12231–12236.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Higgins, J.D., Armstrong, S.J., Franklin, F.C.H., and Jones, G.H. (2004). The Arabidopsis MutS homolog AtMSH4 functions at an earlystep in recombination: evidence for two classes of recombination in Arabidopsis. Genes Dev. 18: 2557–70.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Higgins, J.D., Sanchez-Moran, E., Armstrong, S.J., Jones, G.H., and Franklin, F.C.H. (2005). The Arabidopsis synaptonemal complexprotein ZYP1 is required for chromosome synapsis and normal fidelity of crossing over. Genes Dev. 19: 2488–2500.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hinshaw, S.M., Makrantoni, V., Harrison, S.C., and Marston, A.L. (2017). The Kinetochore Receptor for the Cohesin Loading Complex.Cell 171: 72–84.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hong, S., Sung, Y., Yu, M., Lee, M., Kleckner, N., and Kim, K.P.P. (2013). The logic and mechanism of homologous recombinationpartner choice. Mol. Cell 51: 440–453.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ito, M., Kugou, K., Fawcett, J.A., Mura, S., Ikeda, S., Innan, H., and Ohta, K. (2014). Meiotic recombination cold spots in chromosomalcohesion sites. Genes to Cells 19: 359–373.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jacob, Y., Feng, S., LeBlanc, C.A., Bernatavichute, Y. V, Stroud, H., Cokus, S., Johnson, L.M., Pellegrini, M., Jacobsen, S.E., andMichaels, S.D. (2009). ATXR5 and ATXR6 are H3K27 monomethyltransferases required for chromatin structure and gene silencing. Nat.Struct. Mol. Biol. 16: 763–8.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kauppi, L., Jeffreys, A.J., and Keeney, S. (2004). Where the crossovers are: recombination distributions in mammals. Nat. Rev. Genet.5: 413–24.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kianian, P.M.A. et al. (2018). High-resolution crossover mapping reveals similarities and differences of male and female recombinationin maize. Nat. Commun. 9: 2370.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kim, K.P., Weiner, B.M., Zhang, L., Jordan, A., Dekker, J., and Kleckner, N. (2010). Sister cohesion and structural axis componentsmediate homolog bias of meiotic recombination. Cell 143: 924–37.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kitajima, T.S., Miyazaki, Y., Yamamoto, M., and Watanabe, Y. (2003). Rec8 cleavage by separase is required for meiotic nucleardivisions in fission yeast. EMBO J. 22: 5643–53.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Klein, F., Mahr, P., Galova, M., Buonomo, S.B., Michaelis, C., Nairz, K., and Nasmyth, K. (1999). A central role for cohesins in sisterchromatid cohesion, formation of axial elements, and recombination during yeast meiosis. Cell 98: 91–103.
Pubmed: Author and Title

Google Scholar: Author Only Title Only Author and Title
Koch, B., Kueng, S., Ruckenbauer, C., Wendt, K.S., and Peters, J.-M. (2008). The Suv39h–HP1 histone methylation pathway isdispensable for enrichment and protection of cohesin at centromeres in mammalian cells. Chromosoma 117: 199–210.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Kugou, K., Fukuda, T., Yamada, S., Ito, M., Sasanuma, H., Mori, S., Katou, Y., Itoh, T., Matsumoto, K., Shibata, T., Shirahige, K., andOhta, K. (2009). Rec8 guides canonical Spo11 distribution along yeast meiotic chromosomes. Mol. Biol. Cell 20: 3064–76.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Laloraya, S., Guacci, V., and Koshland, D. (2000). Chromosomal addresses of the cohesin component Mcd1p. J. Cell Biol. 151: 1047–56.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lam, I. and Keeney, S. (2014). Mechanism and Regulation of Meiotic Recombination Initiation. Cold Spring Harb. Perspect. Biol. 7:a016634.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lambing, C., Choi, K., Blackwell, A.R., and Henderson, I.R. (2020). Chromatin Immunoprecipitation of Meiotically Expressed Proteinsfrom Arabidopsis thaliana Flowers. Methods Mol. Biol. 2061: 219–236.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lambing, C., Osman, K., Nuntasoontorn, K., West, A., Higgins, J.D., Copenhaver, G.P., Yang, J., Armstrong, S.J., Mechtler, K., Roitinger,E., and Franklin, F.C.H. (2015). Arabidopsis PCH2 Mediates Meiotic Chromosome Remodeling and Maturation of Crossovers. PLoSGenet. 11: e1005372.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lange, J., Yamada, S., Tischfield, S.E., Pan, J., Kim, S., Zhu, X., Socci, N.D., Jasin, M., and Keeney, S. (2016). The Landscape of MouseMeiotic Double-Strand Break Formation, Processing, and Repair. Cell 167: 695-708.e16.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Langmead, B. and Salzberg, S.L. (2012). Fast gapped-read alignment with Bowtie 2. Nat. Methods 9: 357–9.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Law, J.A. and Jacobsen, S.E. (2010). Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev.Genet. 11: 204–220.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lawrence, E.J., Gao, H., Tock, A.J., Lambing, C., Blackwell, A.R., Feng, X., and Henderson, I.R. (2019). Natural Variation in TBP-ASSOCIATED FACTOR 4b Controls Meiotic Crossover and Germline Transcription in Arabidopsis. Curr. Biol. 29: 2676–2686.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lee, T., Gurazada, S.G.R., Zhai, J., Li, S., Simon, S.A., Matzke, M.A., Chen, X., and Meyers, B.C. (2012). RNA polymerase V-dependentsmall RNAs in Arabidopsis originate from small, intergenic loci including most SINE repeats. Epigenetics 7: 781–795.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lengronne, A., Katou, Y., Mori, S., Yokobayashi, S., Kelly, G.P., Itoh, T., Watanabe, Y., Shirahige, K., and Uhlmann, F. (2004). Cohesinrelocation from sites of chromosomal loading to places of convergent transcription. Nature 430: 573–578.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Liu, S., Yeh, C.-T., Ji, T., Ying, K., Wu, H., Tang, H.M., Fu, Y., Nettleton, D., and Schnable, P.S. (2009). Mu Transposon Insertion Sitesand Meiotic Recombination Events Co-Localize with Epigenetic Marks for Open Chromatin across the Maize Genome. PLoS Genet. 5:e1000733.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Love, M.I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2.Genome Biol. 15: 550.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Maheshwari, S., Ishii, T., Brown, C.T., Houben, A., and Comai, L. (2017). Centromere location in Arabidopsis is unaltered by extremedivergence in CENH3 protein sequence. Genome Res. 27: 471–478.
Pubmed: Author and Title

Google Scholar: Author Only Title Only Author and Title
Mancera, E., Bourgon, R., Brozzi, A., Huber, W., and Steinmetz, L.M. (2008). High-resolution mapping of meiotic crossovers and non-crossovers in yeast. Nature 454: 479–85.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
de Massy, B. (2013). Initiation of meiotic recombination: how and where? Conservation and specificities among eukaryotes. Annu. Rev.Genet. 47: 563–99.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mercier, R., Mézard, C., Jenczewski, E., Macaisne, N., and Grelon, M. (2015). The molecular biology of meiosis in plants. Annu. Rev.Plant Biol. 66: 297–327.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Mizuguchi, T., Fudenberg, G., Mehta, S., Belton, J.-M., Taneja, N., Folco, H.D., FitzGerald, P., Dekker, J., Mirny, L., Barrowman, J., andGrewal, S.I.S. (2014). Cohesin-dependent globules and heterochromatin shape 3D genome architecture in S. pombe. Nature 516: 432–435.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Neale, M.J., Pan, J., and Keeney, S. (2005). Endonucleolytic processing of covalent protein-linked DNA double-strand breaks. Nature436: 1053–7.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nonaka, N., Kitajima, T., Yokobayashi, S., Xiao, G., Yamamoto, M., Grewal, S.I.S., and Watanabe, Y. (2002). Recruitment of cohesin toheterochromatic regions by Swi6/HP1 in fission yeast. Nat. Cell Biol. 4: 89–93.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Osman, K., Sanchez-Moran, E., Mann, S.C., Jones, G.H., and Franklin, F.C.H. (2009). Replication protein A (AtRPA1a) is required forclass I crossover formation but is dispensable for meiotic DNA break repair. EMBO J. 28: 394–404.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Osman, K., Yang, J., Roitinger, E., Lambing, C., Heckmann, S., Howell, E., Cuacos, M., Imre, R., Dürnberger, G., Mechtler, K., Armstrong,S., and Franklin, F.C.H. (2018). Affinity proteomics reveals extensive phosphorylation of the Brassica chromosome axis protein ASY1and a network of associated proteins at prophase I of meiosis. Plant J. 93: 17–33.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Page, S.L. and Hawley, R.S. (2003). Chromosome choreography: the meiotic ballet. Science 301: 785–9.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Pan, J., Sasaki, M., Kniewel, R., Murakami, H., Blitzblau, H.G., Tischfield, S.E., Zhu, X., Neale, M.J., Jasin, M., Socci, N.D., Hochwagen,A., and Keeney, S. (2011). A hierarchical combination of factors shapes the genome-wide topography of yeast meiotic recombinationinitiation. Cell 144: 719–31.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ross, K.J., Fransz, P., and Jones, G.H. (1996). A light microscopic atlas of meiosis in Arabidopsis thaliana. Chromosome Res. 4: 507–16.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rowan, B.A., Heavens, D., Feuerborn, T.R., Tock, A.J., Henderson, I.R., and Weigel, D. (2019). An Ultra High-Density Arabidopsisthaliana Crossover Map That Refines the Influences of Structural Variation and Epigenetic Features. Genetics: doi:10.1534/genetics.119.302406.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sanchez-Moran, E., Santos, J.-L., Jones, G.H., and Franklin, F.C.H. (2007). ASY1 mediates AtDMC1-dependent interhomologrecombination during meiosis in Arabidopsis. Genes Dev. 21: 2220–33.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Schmidt, C.K., Brookes, N., and Uhlmann, F. (2009). Conserved features of cohesin binding along fission yeast chromosomes. GenomeBiol. 10: R52.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Segal, E. and Widom, J. (2009). Poly(dA:dT) tracts: major determinants of nucleosome organization. Curr. Opin. Struct. Biol. 19: 65–71.

Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Serra, H., Lambing, C., Griffin, C.H., Topp, S.D., Seguela-Arnaud, M., Fernandes, J., Mercier, R., and Henderson, I.R. (2018). Massivecrossover elevation via combination of HEI10 and recq4a recq4b during Arabidopsis meiosis. Proc. Natl. Acad. Sci. U. S. A. 115: 2437–2442.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sommermeyer, V., Béneut, C., Chaplais, E., Serrentino, M.E., and Borde, V. (2013). Spp1, a member of the Set1 Complex, promotesmeiotic DSB formation in promoters by tethering histone H3K4 methylation sites to chromosome axes. Mol. Cell 49: 43–54.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Stigler, J., Çamdere, G.Ö., Koshland, D.E., and Greene, E.C. (2016). Single-Molecule Imaging Reveals a Collapsed ConformationalState for DNA-Bound Cohesin. Cell Rep. 15: 988–998.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Stroud, H., Do, T., Du, J., Zhong, X., Feng, S., Johnson, L., Patel, D.J., and Jacobsen, S.E. (2014). Non-CG methylation patterns shapethe epigenetic landscape in Arabidopsis. Nat. Struct. Mol. Biol. 21: 64–72.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Stroud, H., Greenberg, M.V.C., Feng, S., Bernatavichute, Y. V, and Jacobsen, S.E. (2013). Comprehensive analysis of silencing mutantsreveals complex regulation of the Arabidopsis methylome. Cell 152: 352–64.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Sun, X., Huang, L., Markowitz, T.E., Blitzblau, H.G., Chen, D., Klein, F., and Hochwagen, A. (2015). Transcription dynamically patternsthe meiotic chromosome-axis interface. Elife 4: doi: 10.7554/eLife.07424.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Topp, C.N. and Dawe, R.K. (2006). Reinterpreting pericentromeric heterochromatin. Curr. Opin. Plant Biol. 9: 647–653.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Underwood, C.J., Choi, K., Lambing, C., Zhao, X., Serra, H., Borges, F., Simorowski, J., Ernst, E., Jacob, Y., Henderson, I.R., andMartienssen, R.A. (2018). Epigenetic activation of meiotic recombination near Arabidopsis thaliana centromeres via loss of H3K9me2and non-CG DNA methylation. Genome Res. 28: 519–531.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Villeneuve, A.M. and Hillers, K.J. (2001). Whence meiosis? Cell 106: 647–50.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Vincenten, N., Kuhl, L.-M., Lam, I., Oke, A., Kerr, A.R., Hochwagen, A., Fung, J., Keeney, S., Vader, G., and Marston, A.L. (2015). Thekinetochore prevents centromere-proximal crossover recombination during meiosis. Elife 4: doi: 10.7554/eLife.10850.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Walker, J., Gao, H., Zhang, J., Aldridge, B., Vickers, M., Higgins, J.D., and Feng, X. (2018). Sexual-lineage-specific DNA methylationregulates meiosis in Arabidopsis. Nat. Genet. 50: 130–137.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Watanabe, Y. and Nurse, P. (1999). Cohesin Rec8 is required for reductional chromosome segregation at meiosis. Nature 400: 461–4.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
West, A.M., Rosenberg, S.C., Ur, S.N., Lehmer, M.K., Ye, Q., Hagemann, G., Caballero, I., Usón, I., MacQueen, A.J., Herzog, F., andCorbett, K.D. (2019). A conserved filamentous assembly underlies the structure of the meiotic chromosome axis. Elife 8: doi:10.7554/eLife.40372.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wijnker, E. et al. (2013). The genomic landscape of meiotic crossovers and gene conversions in Arabidopsis thaliana. Elife 2: e01426.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yelagandula, R. et al. (2014). The histone variant H2A.W defines heterochromatin and promotes chromatin condensation inArabidopsis. Cell 158: 98–109.
Pubmed: Author and Title

Google Scholar: Author Only Title Only Author and Title
Yelina, N.E., Lambing, C., Hardcastle, T.J., Zhao, X., Santos, B., and Henderson, I.R. (2015). DNA methylation epigenetically silencescrossover hot spots and controls chromosomal domains of meiotic recombination in Arabidopsis. Genes Dev. 29: 2183–202.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yoon, S.-W., Lee, M.-S., Xaver, M., Zhang, L., Hong, S.-G., Kong, Y.-J., Cho, H.-R., Kleckner, N., and Kim, K.P. (2016). Meiotic prophaseroles of Rec8 in crossover recombination and chromosome structure. Nucleic Acids Res. 44: 9296–9314.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang, W., Lee, H.-R., Koo, D.-H., and Jiang, J. (2008). Epigenetic modification of centromeric chromatin: hypomethylation of DNAsequences in the CENH3-associated chromatin in Arabidopsis thaliana and maize. Plant Cell 20: 25–34.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhu, B., Zhang, W., Zhang, T., Liu, B., and Jiang, J. (2015). Genome-Wide Prediction and Validation of Intergenic Enhancers inArabidopsis Using Open Chromatin Signatures. Plant Cell 27: 2415–2426.
Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zickler, D. and Kleckner, N. (1999). Meiotic chromosomes: integrating structure and function. Annu. Rev. Genet. 33: 603–754.Pubmed: Author and TitleGoogle Scholar: Author Only Title Only Author and Title

DOI 10.1105/tpc.19.00866; originally published online February 5, 2020;Plant Cell
Kim Osman, James Higgins, F. Christopher H. Franklin and Ian R. HendersonChristophe Lambing, Andrew J. Tock, Stephanie D. Topp, Kyuha Choi, Pallas C. Kuo, Xiaohui Zhao,
Arabidopsis thalianaInteracting genomic landscapes of REC8-cohesin, chromatin and meiotic recombination in
This information is current as of March 18, 2020
Supplemental Data /content/suppl/2020/02/05/tpc.19.00866.DC1.html
Permissions X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists





























