Embed Size (px)
Citation preview

BIOTECHNOLOGICALLY RELEVANT ENZYMES AND PROTEINS
Phosphorylation of pyruvate kinase A by protein kinase J leadsto the altered growth and differential rate of intracellular survivalof mycobacteria
Diwakar K. Singh & Pramod K. Singh & Sameer Tiwari &Susmita K. Singh & Ruma Kumari & Dinesh K. Tripathi &Kishore K. Srivastava
Received: 10 April 2014 /Revised: 26 May 2014 /Accepted: 27 May 2014# Springer-Verlag Berlin Heidelberg 2014
Abstract PknJ (Rv2088) is a serine/threonine protein kinaseof mycobacteria which is present in Mycobacteriumtuberculosis (MTB), but its gene is absent in Mycobacteriumsmegmatis (MS); a fast grower and nonpathogenic species ofmycobacteria. The heterologous expression of MTB-specificPknJ in MS altered the growth of recombinant mycobacteriahighlighting one of the characteristics of this protein. Thisnature of the protein was further confirmed whenMycobacte-rium bovis BCG (BCG) containing antisense copy of pknJresulted in the increased growth of BCG. The real-time RNAquantification analysis pointed out toward increased expres-sion of this protein during infection of THP-1 macrophagecells which further emphasized that the protein is essential forthe intracellular survival of mycobacteria. The differential ingel electrophoresis (DIGE) data followed by mass spectros-copy suggested that PknJ is involved in regulation of pyruvatekinase A (Rv1617). Since pyruvate kinase (PK) A is one ofthe key enzymes which controls glycolytic cycle inmycobacteria, we looked for its interaction with PknJ duringextracellular and intracellular growth of mycobacteria. In or-der to identify the specific residue(s) involved in post-translational modification, the phospho-null mutants of PKwere generated, and their substrate specificities in response toPknJ were assessed through kinase assay. The findings thusunderlined that the PK activity is predominantly dependent on
the threonine residue at the 94th position and further suggestedthat this site may be plausible in intracellular survival ofmycobacteria upon phosphorylation with PknJ.
Keywords Mycobacterium tuberculosis . STPK . PknJ .
Pyruvate kinase . Pathogenesis
Introduction
Protein phosphorylation and dephosphorylation are the prin-cipal mechanisms for translation of signals into cellular re-sponses. In mycobacteria, post-translational modification is aproven mechanism by which extracellular signals are trans-duced into various biological functions (Av-Gay and Everett2000). The regulation by kinases is frequently associated withthe disease, and thus, they are proposed in drug discoveryprograms as targets to modulate and control the cellular func-tions (Schreiber et al. 2009). Signalling pathways in Myco-bacterium tuberculosis (MTB) are controlled by 11 two-component systems, 11 serine threonine protein kinases(STPKs), one Ser/Thr phosphatase, one tyrosine kinase andtwo tyrosine phosphatases (Cole et al. 1998; Bach et al. 2009).Genomic analysis of MTB revealed 11 predicted eukaryotic-like STPKs; nine of them are transmembrane (Bach et al.2009; Av-Gay and Everett 2000). Although initial studieshave established roles for STPKs in prokaryotic development,stress responses and host–pathogen interactions (Av-Gay andEverett 2000), the upstream activators, downstream sub-strates, effectors and other binding partners need to be identi-fied. In order to adapt its physiology to the changing hostenvironmental conditions, MTB would need to sense theextracellular milieu to modulate the corresponding adaptivegene expression. The endogenous substrates for
Electronic supplementary material The online version of this article(doi:10.1007/s00253-014-5859-4) contains supplementary material,which is available to authorized users.
D. K. Singh : P. K. Singh : S. Tiwari : S. K. Singh : R. Kumari :D. K. Tripathi :K. K. Srivastava (*)Division of Microbiology and Academy of Scientific and InnovativeResearch, CSIR, Central Drug Research Institute, Lucknow 226031,Indiae-mail: [email protected]
Appl Microbiol BiotechnolDOI 10.1007/s00253-014-5859-4

mycobacterial STPKs are predominant to effect cellular ma-chinery of mycobacteria. Earlier findings that mycobacterialprotein Rv1747 is a predicted ABC transporter, which also isencoded in the same operon as its substrate, is phosphorylatedin vitro by PknF (Molle et al. 2004). Similarly, the pknK islocated in the vicinity of virS gene, which is known to regulatethe expression of the mycobacterial monooxygenase (mymA)operon (Kumar et al. 2009) while GarA as a substrate isidentified by in vitro kinase reaction (Villarino et al. 2005),and both of these substrates are known to control the myco-bacterial persistence after post-phosphorylation event. Thedetailed studies with protein kinases G and F have shown thatthese are key molecules which change the phosphorylationpattern of the host proteins upon infection, thereby promotingbacterial survival (Koul et al. 2001). The interdependent ac-tivities of two enzymes have been found to control the growthof mycobacteria, where the moderate downregulation of ppkgene by antisense was reported to inhibit the growth of MTB(H37Ra). It is concluded that the poly (P) and ATP glucoki-nase activities are catalysed by the same enzyme but that thetwo substrates may have different binding sites (Hsieh et al.1993). One of the key reasons for the impairment of growthhas been the kinase-mediated effect on the fatty acid synthesispathway. The mycobacterial FAS-II multienzyme complexhas been identified to be a target of MTB STPKs, withsubstrates, including the malonyl-CoA:acyl carrier protein(ACP) transacylase, FabD and the β-ketoacyl-ACP synthasesKasA and KasB. These proteins are phosphorylated by vari-ous kinases in vitro; however, the phosphorylation is found tobe the most efficient with PknA. Interestingly, though phos-phorylation has a negative impact on the activity of KasA, theKasB activity is enhanced upon phosphorylation (Khan et al.2010). Hence, mycobacterial STPKs influence diverse path-ways and play considerable roles in metabolism and in sur-vival of mycobacteria. Although the PknJ-dependent phos-phorylation of targets (EmbR, MmaA4, PepE and pyruvatekinase (PK)) has been reported recently, biological signifi-cance is still undefined (Jang et al. 2010; Arora et al. 2010).PknJ has no detectable orthologue in mycobacterial speciesoutside the MTB complex (Narayan et al. 2007). Thephosphorylation mediated effect on the growth has also beendemonstrated outside mycobacterial system. The 2-oxoglutarate dehydrogenase inhibitor protein (OdhI) of Cory-nebacterium glutamicum is phosphorylated by PknG. BesidesOdhI, the cell division protein FtsZ was identified as anin vitro substrate for PknA, PknB and PknL and as anin vivo substrate for Ser/Thr protein phosphatase inC. glutamicum (Schultz et al. 2009).
In the present study, we have established through a series ofexperiments that PknJ has an important role in the growth ofmycobacteria and an increased survival in THP-1 human cellline. Further, we have elaborated that this response is regulat-ed by phosphorylation of PK. The PK expression and the
activity of enzyme were altered during the expression of PknJ.PK is a rate-limiting enzyme in glycolysis which also plays animportant role in cell metabolism, and its phosphorylationfurther implicates a regulatory role played in mycobacterialphysiology.
Materials and methods
Cell line, bacterial cultures and animals
The macrophage cell line THP-1 was obtained from theAmerican Type Culture Collection (ATCC), USA, andcultured in RPMI-1640 medium (2 mML-glutamine,10 mM HEPES, 1 mM sodium pyruvate, 4.5 g/L glucoseand 1.5 g/L sodium bicarbonate), supplemented with 10 %heat-inactivated fetal calf serum (FCS) at 37 °C and 5 %CO2. Escherichia coli (E. coli) strains DH5α and BL21(DE3) were procured from NEB and were cultured inLur i a–Ber t an i med ium. Mycobac t e r i a l s t r a in sMycobacterium smegmatis mc2155 (MS, ATCC#700084),Mycobacterium bovis BCG laboratory strain (ATCC#35734,TMC 1011 BCG Pasteur) and M. tuberculosis H37Rv(ATCC#25618) were procured through ATCC (USA). Thestrains were maintained according to the protocol as reportedearlier (Singh et al. 2013a). Mice (BALB/c, 20–25 g) andalbino rabbits (1.5 kg) were obtained from the animal colonyof the institute. The institutional ethics committee for use oflaboratory animals has approved the study. The animals weremaintained in National Accreditation Board for Testing andCalibration Laboratories, accredited animal house and givenproper pellet diet, water ad libitum and environment (temper-ature 22±2 °C; humidity 45–55 %; light intensity 300–400 lx).
Cloning, expression and purification of PknJ and PK
The genes encoding Rv2088 of 1,770 bp and Rv1617 of1,419 bp were amplified by PCR from the genomic DNAof MTB using gene-specific primers having HindIII re-striction site at 5′ and 3′ ends of the primers (Table S1).PCR-amplified products were cloned in TA cloning vector(pTZ57R/T) and further sub-cloned at the same site intopTriEx4 E. coli expression vector, in mycobacterial inte-grative and in episomal vectors. The recombinant expres-sion vector plasmid was transferred into E. coli BL21 (DE3)cells, induced at 27 °C for 5 h. The harvested cells wereresuspended and sonicated in 20 mM Tris-HCl, pH 7.5,200 mM NaCl and protease inhibitor (Sigma). The expressedproteins were purified by nickel nitrilotriacetic acid columnusing 250 mM imidazole.
Appl Microbiol Biotechnol

Generation of polyclonal antisera and immunoblot analysesagainst PknJ and PK
Purified PknJ and PK proteins (200 μg) were mixed withFreund’s incomplete adjuvant (FIA) and injected subcutaneous-ly into rabbits. The injections of 50 μg of each proteins in FIAwere given at 15 days interval, and the sera were collected after10 days of the second boost. Equal amount of MTB cell lysateswere prepared and resolved on a 10 % sodium dodecyl sulfatepolyacrylamide gel electrophoresis (SDS-PAGE), transferredonto PVDF membrane and immunoblotted with anti-PknJand anti-PK antibodies (1:3,000). ECL detection was done byusing anti-rabbit IgG-HRP conjugated antibody (1:20,000) andanalysed on Chemi-Doc XRS system (Bio-Rad Laboratories,Hercules, CA) using Quantity One program.
PknJ-mediated growth of mycobacteria
The growth was monitored in mycobacterial strains of BCGandMS. Starter cultures were prepared by inoculatingMS andMS-J in LBGT and in Sauton media. Cultures were grown at37 °C in a shaking incubator (180 rpm) up to the OD of 0.05,and colony-forming unit (CFU) were normalized to 1×106/ml.These starter cultures were subjected to measure the growthkinetics. Twenty millilitre of medium was inoculated withstarter culture, and at different time intervals, plating for CFUwas done by passing the cultures five times through a 26-gaugeneedle and vortexed. The serially diluted cultures were platedon MB7H10 agar plates having 25 μg/ml of kanamycin todetermine CFU after 3 days incubation at 37 °C.
Starter cultures of BCG and BCG-AsJ having antisensecopy of pknJ gene were inoculated in MB7H9 medium sup-plemented with 10%ADC. At different time points, 100 μl ofeach cultures diluted in MB7H9 medium were plated onMB7H10 agar plates containing 10 % OADC supplementedwith 25 μg/ml kanamycin to determine CFU after 4 weeks.The fold difference was enumerated from the initial CFU atthe 0 time point. The progressive growth from one time pointto the next one was also considered for the logarithmic mul-tiplication of the mycobacteria by calculating Tx (nR)/T0(nR)=CFUn+STD [where T=time, x=experimental point,n=number and R=replicates].
RNA isolation and quantitative RT-PCR (qRT-PCR)of mycobacteria grown in acid stress and in hypoxiaconditions
Log and stationary phase cultures of BCG and MS weregrown, and the total RNA extraction was done using RNeasyMini kit (Qiagen) according to the manufacturer’s instruc-tions. The exponential growing cultures of MTB were sub-jected for in in vitro acid stress and hypoxia (Singh et al.2013a). DNase-treated RNA was used for making
complementary DNA (cDNA) using first-strand cDNA syn-thesis kit (Fermentas) and random hexamer primers. Quanti-tative real-time PCR was performed in 96 well plate on LightCycler 480 system (Roche) using QuantiTect SYBR greenPCR mix (Roche) with 5 μM primers (Table S1). The calcu-lated cycle threshold (Ct) of the test gene was normalized tothe Ct of the internal control (16S RNA) gene before calcu-lating the fold change of the test samples. Assay was per-formed as reported previously (Singh et al. 2013a, b).
The exponential growing cultures of BCG and MS havebeen used to infect THP-1 cells, and RNAwas extracted fromintracellular mycobacteria at 2, 24, 48 and 72-h post-infectionfor cDNA synthesis. The expression analysis was done byreal-time PCR.
In vitro kinase assay using MBP and PK as substrates
In vitro kinase assays were performed by ADP-glo kinaseassay according to manufacturer’s instruction. The assay wasperformed by incubating PknJ (25 ng) either with MBP(80 ng) or with PK (80 ng) in separate tubes. The reactionmixtures containing 40 mM Tris (pH 7.5), 2 mM MnCl2,20 mM MgCl2, 2 mM DTT, 0.5 mg/ml BSA and 0.1 mMATP were incubated at 24 °C for 1 h. The assay was alsostandardized using different concentrations of substrate (100,50, 25 ng) and monitored by luminometre (Berthold).
Infection of THP-1 macrophage cells with recombinant MS
The THP-1 human monocytic cell line was stimulated todifferentiate into macrophage-like cells by treatment withphorbol myristate acetate (PMA) as described earlier(Chaurasiya and Srivastava 2009; Kumari et al. 2012). THP-1 cells having density 5×105 cells/ml were plated in tissueculture plates. The single-cell suspension of log-phasemycobacteria was prepared by vortexing with the presenceof sterile glass beads and passing through 26-gauge needleand used for macrophage infection (MOI=1:10). The infec-tion was allowed to proceed for 4 h in case of BCG and 3 h incase of MS at 37 °C in 5 % CO2. The cells were washed withincomplete RPMI-1640 medium thrice, and extracellular bac-teria were removed by adding 100 μg/ml amikacin. Themacrophages were lysed with 0.05 % SDS at different timepoints to recover the bacteria and enumerated by platingappropriate dilutions on MB7H10 plates containing 10 %OADC and kanamycin (25 μg/ml). BCG and BCG-AsJ re-covered from large cultures of THP-1 cells were used for PKenzymatic activity at different time points (0, 48 and 96 h).
PK assay
Log-phase bacterial cultures were used for PK assay by mea-suring absorbance in spectrophotometer by determining the
Appl Microbiol Biotechnol

oxidation of NADH at 340 nm. Wild-type MS and MS-J wereharvested, lysed and cell debris was pelleted by centrifugationat 20,000×g for 30 min. Soluble cell extracts were furtherfiltered through a 0.8 μm filter. Forty to sixty micrograms ofproteins were used to determine PK enzymatic activity indifferent recombinants and at different stages of mycobacteria.The enzymatic reaction contained 50 mM Tris, 50 mM KCl,1 mM MgCl2, 1 mM PEP, 1.5 mM ADP, 2 U lactate dehy-drogenase (LDH) from rabbit muscle, 0.05 mM NADH and40–60 μg PK. PK enzymatic activity was determined bycontinuous assay coupled to LDH in which the change inabsorbance at 340 nm due to oxidation of NADH was mea-sured. The rate of change in absorbance at 340 nmwas used tocalculate PK activity.
Protein isolation and labelling with Cy dyes
Protein samples were prepared from log-phase cultures ofBCG and BCG-AsJ. The quality of the samples for 2D-differential in gel electrophoresis (DIGE) was evaluated bymini 2D-PAGE (7 cm IPG strips). The protein lysates werelabelled with Cy dyes according to the manufacturer’s proto-col for minimal labelling (Cy Dye DIGE Fluor minimal dyes,GE Healthcare). Protein amounts were quantified by theBradford method, and 35 μg of each protein was labelled with280 pmol of each Cy dyes in the dark for 30 min. In order tominimize dye-specific labelling artefacts, Cy3 and Cy5 label-ling patterns were swapped among the same group of samples.An internal standard pool with equal amounts of each proteinsample (17.5 μg) was used to reduce intergel variation. Thepooled internal standards were labelled with Cy2. Reactionwas quenched with 10 mML-lysine for 15 min under the sameconditions. Final volume was made up to 350 μl bydestreak solution to rehydrate the IPG strips. The isoelec-tric focusing (IEF) was performed on 13 cm immobilinedry strip pH 4–7 using Ettan IPG phore 3 isoelectricfocusing system (GE Healthcare). The strips were furtherrun on 12 % SDS-PAGE for second dimensional separa-tion of different proteins.
2DE and immunoblotting of BCG proteome
The log-phase cultures of BCG and BCG-AsJ were harvested,and samples for IEF were prepared using 200 μg of theproteins in 250 μl of rehydration buffer (9 M urea, 2 Mthiourea, 4 % CHAPS, 65 mM DTT, 0.5 % IPG buffer and0.002 % BPB) and were run on 11-cm IPG strip as describedearlier.
Electron microscopy
The log-phase cultures of MS, MS-J, BCG and BCG-AsJwere harvested, washed three times with phosphate buffer
(pH 7.2) and then fixed for 8 h in 2.5 % glutaraldehyde and2 % paraformaldehyde in 100 mM phosphate buffer (pH 7.2).The cells were dehydrated using 30, 50, 70, and then 100 %acetone (critical drying), and dried cells were mounted onmicroscope stubs and sputter coated on the coater. The sam-ples were then examined on a LEO 430 scanning electronmicroscope.
Generation of PK mutants
The potential serine and threonine residues were replaced withalanine and valine respectively. The parental recombinantpTriEx4/PK plasmid was used to generate mutations usingQuik change XL site directed mutagenesis kit, according tomanufacturer’s instructions (Stratagene). Table S1c providesthe detailed information about primers used for each muta-genesis. The integrity of all constructs was confirmed byDNAsequencing.
Results
PknJ-mediated growth and survival of mycobacteria
Wild-type and recombinant MS and BCG were cultured inSauton medium. Since MS does not express PknJ as con-firmed by genome wide search and by immunoblotting usingantiserum against PknJ (Fig. 1a), we used this bacteria as asurrogate model to define the function of this kinase. MScontaining a copy of MTB specific pknJ slowed down thegrowth by 1.5 to 10-fold at 24 and 48 h, respectively (Fig. 1b).To confirm whether this protein has any impact in the growthof BCG, we electroporated mycobacteria with the gene clonedin pMV361 vector in the wrong orientation, in order to ex-press antisense transcript (Kumari et al. 2012). The knock-down of the gene reduced the level of the protein considerably(Fig. 1c). The temporal growth of these bacteria was moni-tored. The BCG containing antisense transcript of the genegrew comparatively faster than the wild-type BCG, and thispattern of growth remained until day 10 (Fig. 1d). Thesefindings showed a clear evidence that PknJ is responsible forthe control of growth in mycobacteria. We elaborated thefindings to dissect in vivo and in vivo responses of PknJ ingrowth and in survival. THP-1 cell line was infected with wildtype (MS-V) and recombinant (MS-J) mycobacteria, and theintracellular survival was observed until 48 h. The resultsshowed that mycobacteria containing PknJ sustained the sur-vival in the macrophages and was found to be 1.3-fold (sig-nificant for MS) higher over the wild type infection (Fig. 1e).Since MS does not multiply in macrophage as sustainably asits pathogenic counterpart, we could not observe the prolongeffect of PknJ using MS. To validate whether this
Appl Microbiol Biotechnol
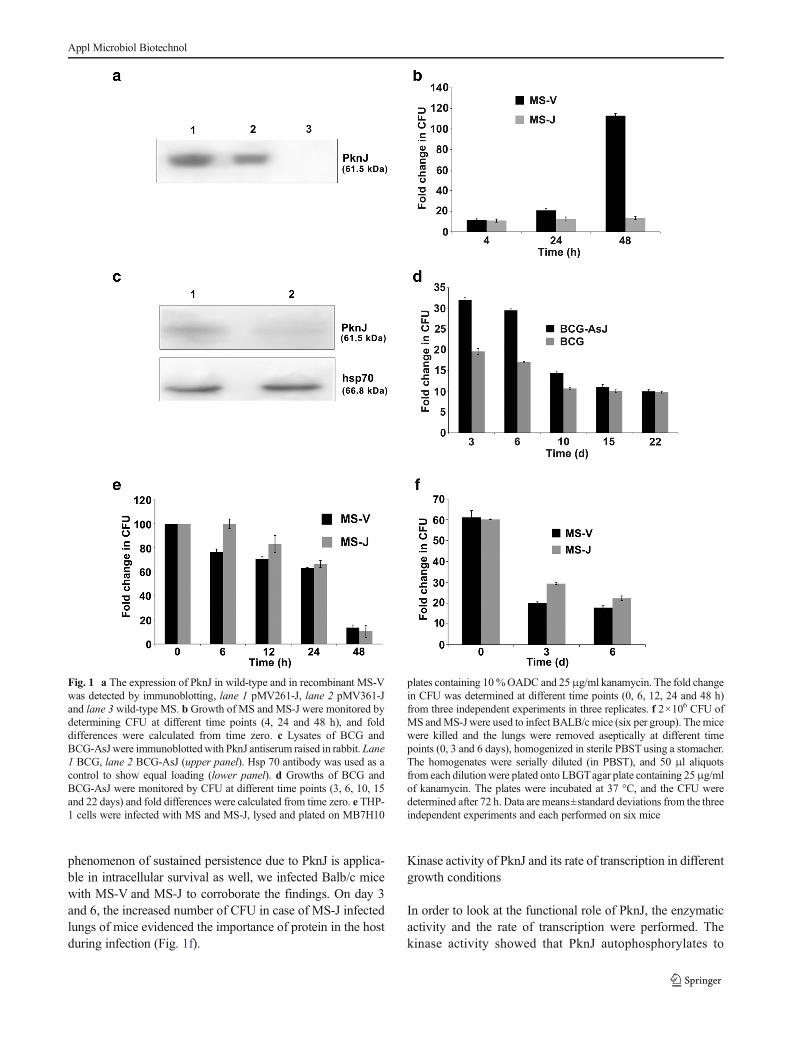
phenomenon of sustained persistence due to PknJ is applica-ble in intracellular survival as well, we infected Balb/c micewith MS-V and MS-J to corroborate the findings. On day 3and 6, the increased number of CFU in case of MS-J infectedlungs of mice evidenced the importance of protein in the hostduring infection (Fig. 1f).
Kinase activity of PknJ and its rate of transcription in differentgrowth conditions
In order to look at the functional role of PknJ, the enzymaticactivity and the rate of transcription were performed. Thekinase activity showed that PknJ autophosphorylates to
Fig. 1 a The expression of PknJ in wild-type and in recombinant MS-Vwas detected by immunoblotting, lane 1 pMV261-J, lane 2 pMV361-Jand lane 3 wild-type MS. b Growth of MS and MS-J were monitored bydetermining CFU at different time points (4, 24 and 48 h), and folddifferences were calculated from time zero. c Lysates of BCG andBCG-AsJwere immunoblottedwith PknJ antiserum raised in rabbit. Lane1 BCG, lane 2 BCG-AsJ (upper panel). Hsp 70 antibody was used as acontrol to show equal loading (lower panel). d Growths of BCG andBCG-AsJ were monitored by CFU at different time points (3, 6, 10, 15and 22 days) and fold differences were calculated from time zero. e THP-1 cells were infected with MS and MS-J, lysed and plated on MB7H10
plates containing 10%OADC and 25μg/ml kanamycin. The fold changein CFU was determined at different time points (0, 6, 12, 24 and 48 h)from three independent experiments in three replicates. f 2×106 CFU ofMS andMS-J were used to infect BALB/c mice (six per group). Themicewere killed and the lungs were removed aseptically at different timepoints (0, 3 and 6 days), homogenized in sterile PBST using a stomacher.The homogenates were serially diluted (in PBST), and 50 μl aliquotsfrom each dilution were plated onto LBGTagar plate containing 25μg/mlof kanamycin. The plates were incubated at 37 °C, and the CFU weredetermined after 72 h. Data are means±standard deviations from the threeindependent experiments and each performed on six mice
Appl Microbiol Biotechnol

exhibit its function in mycobacteria. Using myelin basic pro-tein as a universal kinase substrate, we observed the phosphor-ylation potential of PknJ (Fig. 2a) which led us to look for thespecific substrate(s) of this kinase in mycobacteria. The proteinis highly expressed during stationary stage of mycobacterialgrowth and is poorly expressed during early and logarithmicphases (Fig. 2b), indicating its role in slow growth ofmycobacteria. Since it is important to understand the physiol-ogy of mycobacteria in terms of acidic and basic conditions,
we grew the mycobacteria at different pH and quantified thetranscripts. The data showed that themaximum transcription ofthe gene was achieved at the neutral pH. The expression wasreduced to half when pH was dropped to 5.5. Further loweringof pH (4.5) drastically reduced the levels of transcripts(Fig. 2c). Although, PknJ is highly expressed in stationaryphase, nevertheless subjecting the mycobacteria with low ox-ygen concentration (Singh et al. 2013a and b), the rate oftranscription was found to be reduced by 60 % (Fig. 2d). Thus,
Fig. 2 In vitro kinase assay ofPknJ (a) in presence of MBP (1)and autophosphorylation of PknJ(2). The transcript level of PknJ indifferent growth conditions(in vitro and ex vivo). The BCGcultures were grown at log andstationary phases (b) at differentpH (c) in reduced oxygen (d) andlevel of pknJ transcripts afterinfection in THP-1 cells (e). THP-1 cells were infected with BCGand were osmotically lysed;bacteria were recovered aftercentrifugation at different timepoints. cDNAs were preparedfrom total bacterial RNA asmentioned above. The cDNA ofdifferent time points were used astemplates for SYBR green real-time PCR using pknJ primers(values presented are normalizedagainst 16 s rRNA). Data aremeans±standard deviations fromthree independent experimentseach performed at least six times
Appl Microbiol Biotechnol

the amount of RNA suggested that the expression of PknJprotein peaked at the stationary condition of growth, at neutralpH and at optimum oxygen concentration. The fate of thisprotein during intracellular survival showed that the increase intemporal expression of PknJ could be one of the possibilites forthe persistence of mycobacteria in THP-1 cells (Fig. 2e).
PknJ-mediated morphological changes in mycobacteria
A very clear and detailed resolution of ultrastructures wasachieved by electron microscopy. Samples for electron micro-scopic examinations were prepared from variants (wild-typeMS,MS-J, BCG and BCG-AsJ) and examined through SEM. Com-parisons of the shape and surface morphology revealed thatmycobacteria showed polymorphic characters of variable shapesand sizes. The mycobacteria containing PknJ (wild-type BCGand MS-J) were bulgy and morphologically bigger in the sizesand apparent with the definitive cell wall structures (Fig. 3b, cand Fig. S1b, c) as compared to mycobacteria without PknJ(BCG-AsJ and wild-type MS) as shown (Fig. 3a, d andFig. S1a, d). Thus, it became evident that PknJ is cell wallassociated and its downregulation may have a direct/indirectimpact on the cell wall morphology. To further confirm thelocalization of this protein, we separated the cytosolic and cellwall fractions ofmycobacteria and immunoblottedwith antibodyagainst PknJ. The result confirmed that majority of the protein ispresent in cell wall fraction (Fig. S2).
PK expression and activity in response to PknJ
In order to validate the role of PK in relation to mycobacterialgrowth, we measured its activity in slow (BCG) and in fast(MS) growing mycobacteria. As shown in Fig. 4a, the lysatesof MS exhibited 3-fold higher PK activity over BCG. Thisindicated that PK may be important in the metabolic regula-tion of the mycobacteria. To prove this phenomenon, wecompared the enzymatic activity in logarithmic and stationaryphases. The results showed that the activity of enzyme hasdirect relation to the growth pattern of BCG and is higherwhen the bacteria are in the dividing stage (Fig. 4b). Since MSdoes not contain PknJ, we made efforts to express this proteinin MS through episomal and integrative vectors. Upon ex-pression, the PK activity in MS enhanced as compared to thewild-type. This could further be validated by looking at thecomparative level of expressions by the low copy (integrativepMV361)) and high copy (episomal pMV261) of genes(Fig. 4c).
To re-confirm the fact that PknJ regulates PK activity, werepeated the experiment with BCG by knocking down pknJ.The data showed that PK activity and expression are signifi-cantly reduced in BCG-AsJ (Fig. 4d, e) as well. Furtherelaborations were done at the transcription level of PK inreaction to PknJ. Since MS has two copies of PK, we firstlooked at the differential regulation of these two genes. Onegene has a homologue of BCG and the other was found to be
Fig. 3 SEM analyses ofmycobacteria. The morphologicalchanges were observed. a wild-typeMS, b overexpressed pknJ inMS, c wild-type BCG and dBCG-AsJ having diminishedexpression of PknJ
Appl Microbiol Biotechnol

specific to MS. The BCG homologue in MS was found to betranscribed with much higher efficiency in log and in station-ary phases compared to the MS-specific PK (Fig. 4f).
Interaction of PknJ and PK in mycobacteria
2D-DIGE was applied to assess differential expression ofproteins in BCG and in BCG-AsJ. The labelled samples wereseparated, and the fluorescent images were obtained (Fig. S3).The lysates extracted from the BCG and BCG-AsJ werelabelled with either Cy5 or Cy3 to enable all comparisonsand eliminate any dye-labelling bias, while equal amounts ofprotein from the sample pairs were pooled together as aninternal standard and labelled with Cy2. The normalized ratioof the protein expression level for each spot was calculatedrelative to the internal standard, and intergel spot matchingwas performed using the DeCyder software. The incor-poration of this internal standard on each gel has
been reported to improve the accuracy of spot matching andthe reliability of the quantitative comparison of the spots be-tween the gels. Therefore, this method was sufficient to enableus running two DIGE-gels at the same time. Bioinformaticanalyses of the images detected approximately 1,200 signifi-cantly altered spots in GEL1; 1,207 spots in GEL2 and 1,150spots in both the gels. The matching ratio of the protein spotsbetween the two gels was approximately 95.3%, indicating thatDIGE-based proteomic analysis is an appropriate method tostudy the differentially regulated spots. The summary of thedata as shown in Table S2, indicating that two proteins inthe range of molecular mass 50 to 70 kDa were highlyupregulated. Since there were few proteins which showedup and down regulations in their apparent expressions, weexercised with the bioinformatic tool to narrow down theselection. Since PK has a relative molecular mass of50.6 kDa and pI of 5.37 and also there were reports forits interactions with PknJ, we chose to extend and elaborate
Fig. 4 Enzymatic activity of PKin MS and in BCG at log-phase.a, exponential growing culture ofboth the strains were harvested,and equal amount of proteinswere used for PK enzymaticactivity (μmole/min) as describedin “Materials and methods”. b,The BCG cultures were allowedto grow up to stationary phase,and equal amount of proteinswere used for PK activity. c, PKenzymatic activity (μmole/min)of recombinant MS havingoverexpression of PknJ throughintegrative and episomal vectors.d, PK enzymatic activity in BCGand BCG-AsJ. e, PK expressionwas detected by immunoblottingin BCG and AsJ-BCG. f, PKexpression was analysed bySYBR green real-time PCR in logand stationary phase cultures ofMS. Light bar represents MSspecific PK and dark barrepresents BCG homolog of PKin MS
Appl Microbiol Biotechnol

these findings by dissecting this association in the biologyof infection.
BCG and BCG-AsJ proteomes were further separated by2DE and stained with Coomassie blue (Fig. S4a, b) in order toselect the spots for mass spectrometry. The spots in the rangeof pI 5.0–5.6 were identified and were also confirmed byimmunoblotting using PK antiserum (Fig. S4c). The PK pro-tein was also confirmed for its phosphorylation by PknJ on2DE blot by using phosphorylated Ser/Thr antibody (Cellsignalling). A streaking could be observed showing the phos-phorylation of PK in BCG (Fig. S4d). The detected spot wasfocussed near pI 5.37 and showed difference in expression ofPK between BCG and BCG-AsJ.
Identification of phosphorylation sites in PK
PK was immunoprecipitated by the log-phase culture of BCGusing PK antiserum, and corresponding band was excised andused for LC/MS/MS analysis (Bencos). A total of 12 peptidefragments were generated which covered 52.97 % of the totalpeptides generated in PK (Fig. 5). Among these peptides, twoof them showed phosphorylation at Thr94 and Ser 337 positions(Fig. S5).
Mutant analysis and validation of phosphorylation sites in PK
To validate these phosphorylation sites at Ser337, Thr94 (pres-ent work) and Ser37 (earlier reported by other group, (Aroraet al. 2010), residues were replaced to Ala337, Val94 and Ala37,respectively, by site-directed mutagenesis to generatephospho-deficient mutants of PK (Fig. 6a). The mutant pro-teins were overexpressed in E. coli BL21 (DE3) cells andpurified by nickel nitrilotriacetic acid column (Fig. 6b). Theequal amount of purified proteins was used to detect theactivities of PK and PknJ. The data showed that Thr94 position
is important to confer the PK activity, and the mutation at thisposition abrogated the enzymatic activity (Fig. 6c). To lookwhether this position is also the site for making an impact onthe mycobacterial growth in response to PknJ, we used the wtand mutant proteins as substrates for PknJ activity. The resultsfurther confirmed that PknJ significantly phophorylates thre-onine at 94 position (Fig. 6d).
Discussion
PknJ is a transmembrane serine/threonine protein kinaseshowing sequence homology to the catalytic domain of eu-karyotic serine/threonine kinases. There are two lobes thatform a deep cleft in which ATP binds. The smaller lobe isreferred to as the N-terminal lobe that enhances the interactionbetween the lobes while the larger lobe is identified as C-terminal lobe. The key roles of the C-terminal lobe are to bindprotein substrate and initiate transfers of phosphoryl group.The importance of PknJ in the extracellular growth and inintracellular survival was the indication that this kinase maybe pertinent to decide the fate of slow and fast growth ofmycobacteria. Complementation of this kinase in MS throughintegrative vector showed the retardation in the growth of MS(Fig. 1b) and was confirmed when PknJ was depleted in BCG,resulting in the relatively better growth than the wild type(Fig. 1d). This phenomenon was also observed when therecombinant MS was used to infect either human macrophagecell line (Fig. 1e) or the Balb/c mice (Fig. 1f). Since MS doesnot persist longer in the macrophages and in the animals, thesignificance of discrepancy was established by the repetitionof experiments, by using number of technical replicates and atseveral occasions.
The cellular fractionation data along with SEM analysissubstantiated that majority of PknJ protein is associated with
Fig. 5 The selected spots from2D gels were used for LC/MS/MS analysis. The peptidesequence data matched with thewhole protein of PK. The peptideshowed the phosphorylationevents at 94 and 337 residues
Appl Microbiol Biotechnol

cell wall (Fig. S2), contemplating that the mycobacterialgrowth and survival are compromised upon knocking downof the expression (Fig. 1) (Hsieh et al. 1993; Peirs et al. 2000;Molle et al. 2003a, b). Since PknJ is a serine/threonine proteinkinase, it became obvious that the intracellular effect is due tophosphorylation of substrate(s) (Fig. 2a) (Molle et al. 2003a,b; Av-Gay et al. 1999). Because of its involvement in thecontrol of growth, we observed that this protein is expressed inhigher amounts during stationary phase as compared to loga-rithmic phase at an optimum pH and oxygen, as shown byquantitative RNA analysis (Fig. 2b-d). The preliminary datawere generated by SEM, which showed that the bacilli-expressing PknJ are more bulbous and have definitive cellwall boundaries as compared to the bacilli lacking PknJ. Thiswas confirmed by using recombinant MS and BCG-AsJ(Fig. 3). Earlier reports with some other transmembrane ki-nases like PknF, PknA and PknJ showed similar kind ofarchitecture (Gopalaswamy et al. 2004; Deol et al. 2005;Kang et al. 2005) of cell wall during overexpression of theproteins. Prior to these findings, little has been known aboutthe function of PknJ in physiology and virulence. In presentstudy, we report the functional importance of PknJ in growthand in the pathogenesis which we predict to be mediated byinteracting with the endogenous substrate either through
phosphorylation or by changing its expression level. In rela-tion to the last irreversible step in glycolysis, there has been noreport of PK being essential to in vivo growth of MTB,although Rubin and colleagues reported that for the optimalgrowth of MTB, PK is required to be supplemented in aglucose-containing medium (Chaba et al. 2002).
The unique property of PknJ is to co-express along with PKand affect many physiological properties of mycobacteriabecause it is an enzyme of glycolytic pathway. The PK isknown to catalyse the last steps of glycolysis usingphosphoenol pyruvate (PEP) and ADP as substrates to pro-duce pyruvate and ATP. PK expression and enzymatic activityincrease during exponential phase of Staphylococcus aureusgrowth, and the complete depletion of glucose occur duringthis phase of growth. PK gene disruption has demonstratedthat it is essential for S. aureus growth and survival, suggest-ing that this protein may be a potential drug target (Sassettiand Rubin 2003).
To address the interaction of PknJ with PK, we choose MSas a model and constructed different types of recombinant forMS and for BCG (MS-J and BCG-AsJ). Surprisingly, wefound alteration in PknJ expression which affected the PKexpression and its enzymatic activity in recombinantmycobacteria. PknJ is known to phosphorylate PK
Fig. 6 PKmutant architecture: purification of full length proteins and itsfunctional analysis. a, A schematic representation of PK and its generatedmutants. b, Full-length PK was purified, and equal amount of proteinswas run on 10 % SDS-PAGE and stained with Coomassie Brilliant Blue.
c, PK enzymatic activity of different mutant analysed by coupling reac-tion as stated in “Materials and methods”. d, In vitro autophosphorylationof PknJ and phosphorylation of wild-type and mutant PK as substrates forPknJ
Appl Microbiol Biotechnol

(Christofk et al. 2008b) and several other mycobacterial pro-teins (Arora et al. 2010). The specificity of phosphorylationwas confirmed by the linearity in kinase activity when usingdifferent doses of the substrates. (Fig. S6). The PK enzymaticactivity was considerably different in slow and in fast growingmycobacteria (BCG and MS). The MS has 3.25-fold higherPK enzymatic activity over BCG (Fig. 4a) because MS con-tains an additional copy of PK; however, the PK homologueof BCG is mainly responsible for this activity. This specificitywas re-confirmed by comparative real time PCR analysisusing BCG and MS-specific primers and demonstrated thatthe PK homologue of BCG in MS is mainly transcribedthroughout the life cycle (Fig. 4f). Unlike MTB andM. bovis,MS encodes an additional α-ketoic acid: ferredoxin oxidore-ductase homologue also, which is likely responsible for MVoxidoreductase activity. The α-ketoglutarate decarboxylase(KGD) also plays an important role in the growth ofmycobacteria using carbohydrates as the sole carbon andenergy source (Jang et al. 2010). PK activity was not detectedinM. bovis or inM. microti, strains having Glu220Asp muta-tion. However, MTB and BCG possess the Glu220 residueand have PK activity. To verify that the loss of activity was notattributable to lack of expression, real-time (RT)-PCR wasused and found that the level of PK mRNA expression inM. bovis is 2.46 times that expressed by M. bovis BCG(Baughn et al. 2009). Quantitative estimation of transcriptsshowed that PknJ and PK are expressed mostly in stationaryphase culture in BCG, which means that it is essential forsurvival in stress condition and protect the bacteria frommetabolite produced during growth. The induced levels ofPknK in stationary phase activate cellular targets or pathwaysthat slow down the growth of mycobacteria (Keating et al.2005). Since earlier studies with the interaction of PknJ andPK did not show significant differences in activity of phos-phorylated (WT-PykA) and unphosphorylated (PykA-S37A)forms of PK (Arora et al. 2010), we believed that event couldbe led by phosphorylation(s) at other site(s). Moreover, theearlier study was speculated on the basis of motif in PKreported in a eukaryotic system (Arora et al. 2010). In orderto precise the PknJ-mediated phosphorylation site(s) in PK inmycobacteria, we compared antisense and wt mycobacteriaand analysed with 2DE-DIGE (Fig. S3 and Table S2). Thedata showed 5–10-fold increase of PK in the recombinantmycobacteria and upon mass spectroscopic analysis showedtwo new phosphorylation sites at Thr94 and at Ser337. Surpris-ingly, PknJ did not phosphorylate PK at serine 37 (Fig. 5 andFig. S5) as reported earlier (Arora et al. 2010). This data led usto further analyse and validate these phosphorylation sites. Weconstructed the mutants by replacing serine and threonineresidues. The results were found to be very encouraging,showing that replacement of threonine at 94 position withvaline disrupts PK activity (Fig. 6c), and using thisphosphor-null protein as substrate considerably reduced
kinase activity by PknJ (Fig. 6d). Conclusively, the findingsdelineated that PknJ interacts with PK and regulates the my-cobacterial growth and the physiology.
The high level sequence and structural divergence betweenbacterial and mammalian PKs indicate that PK is a potentialnovel antibacterial target. It is widely suggested that potentialtarget genes should be essential for bacterial viability andpathogenesis (Sassetti and Rubin 2003; Zoraghi et al. 2010,2011; Christofk et al. 2008a) and hence the interaction of thesetwo kinases may draw the attention towards designing theinhibitors. Moreover, PK has recently been portraying a lotof attention as a novel target for antitrypanosomal,antileishmanial, antimalarial and anticancer drugs as well.
Acknowledgments We thank Director of Council of Scientific andIndustrial Research—Central Drug Research Institute (CSIR-CDRI) forhis encouragement and support. Financial supports by CSIR-UNDO andCSIR-SPLENDID are acknowledged. We thank Dr. J. K. Saxena forallowing us to use his facility for biochemical assay. The CSIR-CDRIcommunication allotted to this manuscript is 8705.
References
Arora G, Sajid A, Gupta M, Bhaduri A, Kumar P, Basu-Modak S, SinghY (2010) Understanding the role of PknJ in Mycobacteriumtuberculosis: biochemical characterization and identification of nov-el substrate pyruvate kinase A. PLoS One 5:e10772. doi:10.1371/journal.pone.0010772
Av-Gay Y, Everett M (2000) The eukaryotic-like Ser/Thr protein kinasesofMycobacterium tuberculosis. Trends Microbiol 8:238–244
Av-Gay Y, Jamil S, Drews SJ (1999) Expression and characterization ofthe Mycobacterium tuberculosis serine/threonine protein kinasePknB. Infect Immun 67:5676–5682
Bach H, Wong D, Av-Gay Y (2009)Mycobacterium tuberculosis PtkA isa novel protein tyrosine kinase whose substrate is PtpA. Biochem J420:155–160. doi:10.1042/BJ20090478
Baughn AD, Garforth SJ, Vilchèze C, Jacobs WR (2009) Ananaerobic-type alpha-ketoglutarate ferredoxin oxidoreductasecompletes the oxidative tricarboxylic acid cycle of Mycobacteriumtuberculosis. PLoS Pathog 5:e1000662. doi:10.1371/journal.ppat.1000662
Chaba R, RajeM, Chakraborti PK (2002) Evidence that a eukaryotic-typeserine/threonine protein kinase from Mycobacterium tuberculosisregulates morphological changes associated with cell division. EurJ Biochem/FEBS 269:1078–1085
Chaurasiya SK, Srivastava KK (2009) Downregulation of protein kinaseC-alpha enhances intracellular survival of mycobacteria: role ofPknG. BMC Microbiol 9:271. doi:10.1186/1471-2180-9-271
Christofk HR, Vander Heiden MG, Harris MH, Ramanathan A, GersztenRE, Wei R, Fleming MD, Schreiber SL, Cantley LC (2008a) TheM2 splice isoform of pyruvate kinase is important for cancer me-tabolism and tumour growth. Nature 452:230–233. doi:10.1038/nature06734
Christofk HR, Vander HeidenMG,WuN,Asara JM, Cantley LC (2008b)Pyruvate kinase M2 is a phosphotyrosine-binding protein. Nature452:181–186. doi:10.1038/nature06667
Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D, GordonSV, Eiglmeier K, Gas S, Barry CE, Tekaia F, Badcock K, BashamD,Brown D, Chillingworth T, Connor R, Davies R, Devlin K, FeltwellT, Gentles S, Hamlin N, Holroyd S, Hornsby T, Jagels K, Krogh A,
Appl Microbiol Biotechnol

McLean J, Moule S, Murphy L, Oliver K, Osborne J, Quail MA,Rajandream MA, Rogers J, Rutter S, Seeger K, Skelton J, SquaresR, Squares S, Sulston JE, Taylor K,Whitehead S, Barrell BG (1998)Deciphering the biology of Mycobacterium tuberculosis from thecomplete genome sequence. Nature 393:537–544. doi:10.1038/31159
Deol P, Vohra R, Saini AK, Singh A, Chandra H, Chopra P, Das TK,Tyagi AK, Singh Y (2005) Role ofMycobacterium tuberculosis Ser/Thr kinase PknF: implications in glucose transport and cell division.J Bacteriol 187:3415–3420. doi:10.1128/JB.187.10.3415
Gopalaswamy R, Narayanan PR, Narayanan S (2004) Cloning, overex-pression, and characterization of a serine/threonine protein kinasepknI from Mycobacterium tuberculosis H37Rv. Protein Expr Purif36:82–89. doi:10.1016/j.pep.2004.03.011
Hsieh PC, Shenoy BC, Jentoft JE, Phillips NF (1993) Purification ofpolyphosphate and ATP glucose phosphotransferase fromMycobacterium tuberculosis H37Ra: evidence that poly (P) andATP glucokinase activities are catalyzed by the same enzyme.Protein Expr Purif 4:76–84
Jang J, Stella A, Boudou F, Levillain F, Darthuy E, Vaubourgeix J, WangC, Bardou F, Puzo G, Gilleron M, Burlet-Schiltz O, Monsarrat B,Brodin P, Gicquel B, Neyrolles O (2010) Functional characterizationof the Mycobacterium tuberculosis serine/threonine kinase PknJ.Microbiology 156:1619–1631. doi:10.1099/mic.0.038133-0
Kang C-M, Abbott DW, Park ST, Dascher CC, Cantley LC, Husson RN(2005) The Mycobacterium tuberculosis serine/threonine kinasesPknA and PknB: substrate identification and regulation of cellshape. Genes Dev 19:1692–1704. doi:10.1101/gad.1311105
Keating LA, Wheeler PR, Mansoor H, Inwald JK, Dale J, Hewinson RG,Gordon SV (2005) The pyruvate requirement of some members ofthe Mycobacterium tuberculosis complex is due to an inactivepyruvate kinase: implications for in vivo growth. Mol Microbiol56:163–174. doi:10.1111/j.1365-2958.2005.04524.x
Khan S, Nagarajan SN, Parikh A, Samantaray S, Singh A, Kumar D, RoyRP, Bhatt A, Nandicoori VK (2010) Phosphorylation of enoyl-acylcarrier protein reductase InhA impacts mycobacterial growth andsurvival. J Biol Chem 285:37860–37871. doi:10.1074/jbc.M110.143131
Koul A, Choidas A, Tyagi AK, Drlica K, Singh Y (2001) Serine/threonine protein kinases PknF and PknG of Mycobacteriumtuberculosis: characterization and localization. Microbiology 147:2307–2314
Kumar P, Kumar D, Parikh A, Rananaware D, Gupta M, Singh Y,Nandicoori VK (2009) The Mycobacterium tuberculosis proteinkinase K modulates activation of transcription from the promoterof mycobacterial monooxygenase operon through phosphorylationof the transcriptional regulator VirS. J Biol Chem 284:11090–11099. doi:10.1074/jbc.M808705200
Kumari R, Singh SK, Singh DK, Singh PK, Chaurasiya SK, SrivastavaKK (2012) Functional characterization delineates that aMycobacterium tuberculosis specific protein kinase (Rv3080c) isresponsible for the growth, phagocytosis and intracellular survivalof avirulent mycobacteria. Mol Cell Biochem 369:67–74. doi:10.1007/s11010-012-1369-9
Molle V, Girard-blanc C, Kremer L, Doublet P, Cozzone AJ, Lyon D,National C, Recherche D (2003a) Protein PknE, a novel transmem-brane eukaryotic-like serine/threonine kinase from Mycobacterium
tuberculosis. Biochem Biophys Res Commun 308:820–825. doi:10.1016/S0006-291X(03)01476-1
Molle V, Kremer L, Girard-Blanc C, Besra GS, Cozzone AJ, Prost J-F(2003b) An FHA phosphoprotein recognition domain mediatesprotein EmbR phosphorylation by PknH, a Ser/Thr protein kinasefrom Mycobacterium tuberculosis. Biochemistry 42:15300–15309.doi:10.1021/bi035150b
Molle V, Soulat D, Jault JM, Grangeasse C, Cozzone AJPJ (2004) TwoFHA domains on an ABC transporter, Rv1747, mediate its phos-phorylation by PknF, a Ser/Thr protein kinase fromMycobacteriumtuberculosis. FEMS Microbiol Lett 234:215–223
Narayan A, Sachdeva P, Sharma K, Saini AK, Tyagi AK, Singh Y (2007)Serine threonine protein kinases of mycobacterial genus : phylogenyto function. Physiol Genomics 29:66–75. doi:10.1152/physiolgenomics.00221.2006
Peirs P, Parmentier B, De Wit L, Content J (2000) The Mycobacteriumbovis homologous protein of the Mycobacterium tuberculosisserine/threonine protein kinase Mbk (PknD) is truncated. FEMSMicrobiol Lett 188:135–139
Sassetti CM, Rubin EJ (2003) Genetic requirements for mycobacterialsurvival during infection. Proc Natl Acad Sci U S A 100:12989–12994. doi:10.1073/pnas.2134250100
SchreiberM, Res I,Matter A (2009) Protein kinases as antibacterial targets.Curr Opin Cell Biol 21:325–330. doi:10.1016/j.ceb.2009.01.026
Schultz C, Niebisch A, Schwaiger A, Viets U, Metzger S, BramkampM,Bott M (2009) Genetic and biochemical analysis of the serine/threonine protein kinases PknA, PknB, PknG and PknL ofCorynebacterium glutamicum: evidence for non-essentiality andfor phosphorylation of OdhI and FtsZ by multiple kinases. MolMicrobiol 74:724–741. doi:10.1111/j.1365-2958.2009.06897.x
Singh SK, Kumari R, Singh DK, Tiwari S, Singh PK, Sharma S,Srivastava KK (2013a) Putative roles of a proline-glutamic acid-rich protein (PE3) in intracellular survival and as a candidate forsubunit vaccine against Mycobacterium tuberculosis. MedMicrobiol Immunol 202:365–377. doi:10.1007/s00430-013-0299-9
Singh SK, Tripathi DK, Singh PK, Sharma S, Srivastava KK (2013b)Protective and survival efficacies of Rv0160c protein in murinemodel of Mycobacterium tuberculosis. Appl Microbiol Biotechnol97:5825–5837. doi:10.1007/s00253-012-4493-2
Villarino A, Duran R, Wehenkel A, Fernandez P, England P, Alzari PM(2005) Proteomic identification of M. tuberculosis protein kinasesubstrates: PknB recruits GarA, a FHA domain-containing protein,through activation loop-mediated interactions. J Mol Biol 350:953–963. doi:10.1016/j.jmb.2005.05.049
Zoraghi R, See RH, Gong H, Lian T, Swayze R, Finlay BB, BrunhamRC, McMaster WR, Reiner NE (2010) Functional analysis, overex-pression, and kinetic characterization of pyruvate kinase frommethicillin-resistant Staphylococcus aureus. Biochemistry 49:7733–7747. doi:10.1021/bi100780t
Zoraghi R, See RH, Axerio-Cilies P, Kumar NS, Gong H, Moreau A,Hsing M, Kaur S, Swayze RD, Worrall L, Amandoron E, Lian T,Jackson L, Jiang J, Thorson L, Labriere C, Foster L, Brunham RC,McMaster WR, Finlay BB, Strynadka NC, Cherkasov A, YoungRN, Reiner NE (2011) Identification of pyruvate kinase inmethicillin-resistant Staphylococcus aureus as a novel antimicrobialdrug target. Antimicrob Agents Chemother 55:2042–2053. doi:10.1128/AAC.01250-10
Appl Microbiol Biotechnol





























