Embed Size (px)
Citation preview

PHYSICAL REVIEW E 83, 051909 (2011)
Phase description of spiking neuron networks with global electric and synaptic coupling
Dipanjan Roy,* Anandamohan Ghosh,† and Viktor K. Jirsa‡
Theoretical Neuroscience Group, Institut des Sciences du Mouvement, UMR6233 CNRS and Universite de la Mediterranee,163 Avenue de Luminy, F-13288 Marseille, France
(Received 24 July 2010; revised manuscript received 4 February 2011; published 11 May 2011)
Phase models are among the simplest neuron models reproducing spiking behavior, excitability, andbifurcations toward periodic firing. However, coupling among neurons has been considered only using genericarguments valid close to the bifurcation point, and the differentiation between electric and synaptic couplingremains an open question. In this work we aim to address this question and derive a mathematical formulationfor the various forms of coupling. We construct a mathematical model based on a planar simplification of theMorris-Lecar model. Based on geometric arguments we then derive a phase description of a network of theabove oscillators with biologically realistic electric coupling and subsequently with chemical coupling under fastsynapse approximation. We demonstrate analytically that electric and synaptic coupling are differently expressedon the level of the network’s phase description with qualitatively different dynamics. Our mathematical analysisshows that a breaking of the translational symmetry in the phase flows is responsible for the different bifurcationspaths of electric and synaptic coupling. Our numerical investigations confirm these findings and show excellentcorrespondence between the dynamics of the full network and the network’s phase description.
DOI: 10.1103/PhysRevE.83.051909 PACS number(s): 87.19.ll, 87.19.lj, 87.19.lm
I. INTRODUCTION
Cortical neurons display various dynamic behaviors in-cluding spike train generation, various bursting behaviors,sustained constant frequency firing and adaptive frequencyfiring [1–4]. Significant theoretical efforts have been devotedto characterizing the biophysical mechanisms that underliesuch repetitive activity [5]. In parallel mathematical effortshave been undertaken to capture the dynamical mechanismsunderlying this repertoire of neuronal behaviors [6–11]. Oneof the simplest mathematical representations is the canonicalphase model referred to as the theta neuron [12], which canbe derived from a saddle node on an invariant circle (SNIC)bifurcation and displays threshold properties and excitablebehavior. Beyond the bifurcation point, the membrane voltagedisplays repetitive firing [14]. SNIC bifurcations lead toClass I excitability observed in various neuron models[10,13,15]. When coupling neurons in a network, previousmathematical studies have used generic local bifurcationarguments to derive the corresponding representation ofcoupling in the phase description [13,16–21]. No distinctionof electric and synaptic coupling can be established throughthis approach due to the generic nature of the argument. Toaddress this issue of the correct differentiation of electric andsynaptic coupling in the phase description, we here take thefollowing approach: We formulate a planar neuron model thatallows for a convenient phase description via a coordinatetransformation. Under a circular approximation of the phasespace portrait, the electric and synaptic couplings assumevery distinct and simple characteristic forms, whereas themathematical expressions characterizing the intrinsic neuronaldynamics are more complicated (that is, they are not as simpleas in the theta neuron, for instance). Through a perturbation
*[email protected]†[email protected]‡[email protected]
analysis we demonstrate that deviations of the phase portraitfrom the unit circle do not alter the basic mathematical formof the electric and synaptic phase couplings in the lowest-order approximation. We then numerically confirm that thebifurcation structure of the full spiking neuron network model(using planar as well as Hodgkin-Huxley neuron models) iscorrectly captured by the reduced phase model.
II. NEURON MODEL
As a first step we formulate our neuron model, which shallbe based on the Morris-Lecar type biophysical model andrelated “planar” simplifications [24]. We introduce a cubicnullcline and piecewise (PWL) nullcline. The motivation forthis kind of model comes from the reasoning that it can inprinciple generate type I behavior that is often associated witheither a homoclinic or a SNIC bifurcation observed in detailedbiophysical models [20,25]. The introduction of the PWLnullcline simplifies the nonlinearity for the gating variablesand was previously introduced by Tonnelier et al. [25,26].Chik et al. have successfully studied clustering throughpostinhibitory rebound in synaptically coupled neurons usingthis type of PWL caricature [27]. The governing equations forthe single-neuron model are
dx
dt= −y − x3 + x + I (1)
anddy
dt= [−y + f (x)]/τ , (2)
where
f (x) ={
0, for x < x0,
c(x + x0) , for x � x0.(3)
Here, x represents the membrane potential that meets therequirement of a “fast” depolarizing Ca++ current, and y
represents the gating variables that capture the dynamics of
051909-11539-3755/2011/83(5)/051909(10) ©2011 American Physical Society

DIPANJAN ROY, ANANDAMOHAN GHOSH, AND VIKTOR K. JIRSA PHYSICAL REVIEW E 83, 051909 (2011)
a slow repolarizing K+ current. The above system, Eqs. (1)and (2), is a two-dimensional planar representation of thedynamics of the Morris-Lecar model [1] in which the fastcalcium channels instantaneously assume their steady-statevalues following a change in the membrane potential. Theinput current I acts as a bifurcation parameter in our modeland moves the nullcline in the vertical direction with noinfluence on its shape [24]. The parameter values for c =4I , x0 = −0.25 are chosen such that the nullclines alwaysintersect at the center of the homoclinic orbit as I > Ic.From initial simulations we find that the resulting homoclinicorbit is sufficiently circular. The intrinsic timescale of theslow dynamics is τ ; all numerical studies are carried outfor τ > 1. Specifically, here we seek to derive a model,which does not necessarily yield a simple mathematicallyconvenient form (such as the theta neuron model) but ratherallows for an explicit formulation of coupling terms. To
this end we make the following ansatz: [x + i(y − y0)] =r(θ ) exp(iθ ), where x = r(θ ) cos θ and y = y0 + r(θ ) sin θ ,where y0 = I . The amplitude correction is defined as r(θ ) =1 + εh(θ ), where ε is small for near-circular orbits andh(θ ) is the phase dependence of the homoclinic orbit.Substituting this in our governing equations (1)–(3) (wepresent the detailed calculus in Appendix A) we arriveat the following phase evolution equation for the singleneuron:
θ = 1
r ′2 + r2
{[r ′ cos(θ ) − r(θ ) sin(θ )][(I − y0)
− r(θ ) sin(θ ) − r3(θ ) cos3(θ ) + r(θ ) cos(θ )]
+ r ′ sin(θ ) + r(θ ) cos(θ )
τ[−y0 − r(θ ) sin(θ ) + f (θ )]
},
(4)
f (θ ) ={
0 , for r(θ ) cos θ < x0,
c[
r ′ sin θ+r(θ) cos θ
τ
][r(θ ) cos θ + x0] , for r(θ ) cos θ � x0,
(5)
where r ′ = drdθ
. For sufficiently small enough ε the dynamicsevolves on an invariant circular orbit of unit radius, r(θ ) = 1.Then the above phase equation simplifies to
θ = 0.5 +4∑
j=1
aj sin (jθ ) +4∑
j=1
bj cos (jθ ) + f (θ ) + O(ε)
= F (θ ) + O(ε), (6)
f (θ ) ={
0 for cos θ < x0c(1+cos 2θ )
2τ+ cx0(cos θ)
τfor cos θ � x0,
(7)
where the nonlinear flow of the phase is captured by F (θ ),and O(ε) contains all terms of first and higher order in ε. Theparameters are a1 = −(I − y0 ), a2 = −(0.25 + 1/2τ ), a3 =0, a4 = 0.425, b1 = −y0/τ , b2 = −0.5, b3 = 0, and b4 = 0.
Figure 1 shows the phase plane trajectories obtained for thefull model in Eqs. (1)–(3) and the phase model in Eqs. (4)–(5),as well as the circular approximation of the phase modelin Eqs. (6)–(7). We express h(θ ) = a0 + a cos (θ ) + b sin (θ )through a truncated Fourier expansion and determine the coef-ficients numerically as a0 = −1.5, a = −0.34, b = −0.64. Wechoose ε = 0.1, which results in a deviation from circularity oforbit. Simulations of Eqs. (1)–(3) and Eqs. (6)–(7) are shownin Fig. 2. When I < Ic the above system has a pair of fixedpoints; one is stable and the other is unstable [see Fig. 2(a)].These are also captured by the phase model as indicated bythe vector field in Fig. 2(c). When I ≈ Ic, the two fixed pointsapproach each other, collide, and disappear. For I slightlygreater than Ic the trajectory spends practically all its timemoving through the bottleneck. Beyond this, for I > Ic, asshown in Figs. 2(b) and 2(d), we are left with an inflectionpoint right at the center of the orbit. As can be appreciated fromFigs. 2(c) and 2(d), the nonlinear flow F (θ ) is actually quitesimple despite its complicated formulation in Eq. (6). Relevantfor a good qualitative correspondence between the dynamics
of two levels of description is a constant amplitude. This isjustified by the phase-plane structure of our model, which canbe divided into two main regions of interest. The first regionlies in quadrants 2 and 3, and the second in quadrants 1 and4. In quadrants 1 and 4 where x � x0, using two-time scaleanalysis, we show that the magnitude of amplitude obeys thefollowing equation: dr
dt= 0.5r[(1 − 1/τ ) − 3r2] [28]. Setting
the time dependence of r to zero and τ � 1 results inr ≈ 1/
√3. We verify numerically that this is true for τ � 1,
where our model shows a stable invariant cycle that is nearlycircular. In order to show the existence of a SNIC bifurcation,
− 1.5 − 0.5 0 0.5 1.0 1.5
y
x− 1.0
− 1.5
− 1.0
− 0.5
0.5
0
1.0
1.5
FIG. 1. Phase space dynamics for the planar neuron model andthe approximate phase models with and without amplitude correction.The planar model is shown in black dotted curves, the corrected phasemodel in thick gray dotted curves, and the circular phase model inlight gray dotted curves. The two intersecting curves with black squareand black circular points are the two nullclines.
051909-2

PHASE DESCRIPTION OF SPIKING NEURON NETWORKS . . . PHYSICAL REVIEW E 83, 051909 (2011)
FIG. 2. Phase space portrait, x, y time series, and nullclines forplanar model Eq. (1)–(3) in the excitable [left top panel in (a)] andin the oscillatory [left bottom panel in (b)] regimes. Both dynamicalstates are linked through a SNIC at Ic = 0.38 (see text for moredetails). (c) (right top panel) Stability of the fixed points (in black astable fixed point and in white an unstable fixed point) are depictedin a one-dimensional vector field flow on the real line for the phasemodel in the excitable regime and in (d) (right bottom panel) in theoscillatory regime. The arrow heads on the real line indicate the phaseflow.
we perform a linear stability analysis for the full systemEqs. (1)–(3) and find that xc = ±1/
√3 and Ic = ∓2/3
√3.
It is now straightforward to show that near the fixed point,the lowest-order contribution of the local dynamics reduces todxdt
= I − bx2, where I = (√
3I+1)√3
and b = 3√3.
III. NETWORK MODELS
Motivated by the correspondence between the single-neuron model and its approximate circular phase descriptionwe now analyze a globally coupled network. Our objective isto evaluate to what degree the here adapted phase descriptionof a network reproduces the qualitative dynamic features andbifurcations as observed in the full network. Kuramoto [22]was among the first to describe how to compute the interactionbetween the limit cycle oscillators in the limit of infinitesimallyweak coupling. By using the method of averaging, it is possibleto compute the interaction between the oscillators in the limitwhen the interactions are vanishingly small. However, thisdescription works well for the limit cycle regime, but not whenthe neurons are in a nonoscillatory regime. Similar constraintsapply to approaches using phase response curves outside ofthe regular firing regime [23]. Here we take the followingapproach: We introduce a biologically realistic electrical andsynaptic coupling in the neuron model, which is used in manystudies to understand clustering, synchronization, etc., in a net-work of globally coupled oscillators [16,26,29–31]. Then weperform the same transformation as in the previous section forthe uncoupled neuron with the goal to derive an expressionfor the coupling in the limit of circular orbit. The expression
of the couplings in the circular limit will be meaningful if theerrors introduced by the noncircular contributions are at leastof the order ε. In this case the zero-order term of coupling willbe the leading term.
A. Electric coupling
Following is the set of equations that describes a globallyelectrically coupled system of nonlinear oscillators:
dxi
dt= −yi − x3
i + xi + I + K(X − xi) , (8)
dyi
dt= [−yi + f (xi)]/τ , (9)
f (xi) ={
0 for xi < x0
c(xi + x0) for xi � x0,(10)
where i = 1 . . . N and where K is the coupling constant. Foran uncoupled network, K = 0. Each neuron receives inputfrom all other neurons, and the average activity of the networkis X = 1/N
∑Ni=1 xi . The generalized phase equation in the
limit of a circular orbit with electric coupling is as follows:
θi = F (θi) − K sin θi(cos � − cos θi) + O(ε), (11)
where all parameters in F (θ ) are the same as in (6), andcos � = 1/N
∑Nl=1 cos θl , i �= l. The noncircular contribu-
tions are absorbed in O(ε) (see Appendix A). Every neuronis connected to every other neuron in the network, and thecoupling strength between any two neurons is K
N. There is no
self-coupling between the neurons, and the electric couplingconstant usually assumes only positive values. However, inthis case we extended the parameter space search also tonegative (and hence nonbiological) values of K to allow adirect comparison with the effects of synaptic coupling.
Results of the simulations of Eqs. (8)–(11) are shownin Fig. 3, which displays four different dynamics in theparameter space as we vary the control parameters (K, I). Wecompare the four different dynamics obtained from the fullsystem with the phase description of the system for N = 100oscillators and find good correspondence in the qualitativedynamics (see Fig. 4). We employ the following scheme forthe identification of the parameter spaces: First, we determinethe saddle-node bifurcation curve by imposing the conditionsfor phase-coupled networks dθi/dt = 0, det J = 0, where J
is the Jacobian matrix; then we solve these simultaneouslyand sweep θi over the full range from [−π,π ]. Similarlywe determine the Hopf bifurcation curve by imposing theconditions dθi/dt = 0, T rJ = 0. For the planar and the phasemodel composed of identical elements we hold the couplingstrength K to be fixed and allow for adjusting I continuouslywhich results in a degenerate point in the (I, K) plane. We fixK to be zero and in the absence of the coupling determine thecritical I value for both the full and phase models. Subsequentlywe adjust K continuously by holding I fixed at its criticalvalue to compute the bifurcation lines limiting synchronyand incoherence. In the numerical bifurcation analysis thedegenerate codimension 2 point in the (I, K) plane is detected asa cusp bifurcation point in the parameter plane using forwardand backward bifurcation techniques. From simulations wefind for both these models starting with random initial phases
051909-3

DIPANJAN ROY, ANANDAMOHAN GHOSH, AND VIKTOR K. JIRSA PHYSICAL REVIEW E 83, 051909 (2011)
FIG. 3. Schematic of different scenarios for a population ofcoupled neurons in the phase space spanned by the variablesx and y. Each dot represents the state of a neuron. (a) Thepopulation remains in the vicinity of a fixed point attractor. (b)The population shows clustering in the phase space and bistability.(c) The population oscillates out of phase. (d) The population of anoscillator in synchronization. The time series on the right are froma population of 10 neurons for various (I–K) values for the reducedmodel.
for more negative values of K (linear coupling strength)population of neurons exhibit bistability and the existence ofa separatrix. The increase in coupling strength toward morepositive values results in a stable fixed point attractor forboth these systems. As we go above I > Ic for fixed couplingstrength the systems become oscillatory through a saddle-nodebifurcation. Now, depending on the sign of K the entirepopulation of neurons either synchronizes or shows incoherentbehavior about their center of mass. This is quite well capturedin the simulation as shown in Fig. 3. We numerically verifiedthe robustness of the parameter space structure up to N = 1000oscillators. The structure of the parameter space is similar toother oscillator types (typically globally coupled phase modelsclose to SNIC bifurcation) like the one proposed by Ref. [16].
To gain insight into the mechanisms underlying the emer-gence of the oscillatory behavior of the network, we pursuean analytical approach making use of the network’s phaseevolution equations
θi = F (θi) − K sin θi(cos � − cos θi), (12)
where all the parameters are the same as described in Eq. (11).In the parameter space in Fig. 4 the region for K > 0corresponds to mostly monostable and synchronized solutions,that is, phase-locked solutions where cos θi = cos �. Forphase-locked solutions the coupling term is essentially zero,and � must be a solution of � = F (�), and hence thenetwork’s monostable solution changes stability for increasingI at the same time as when the stability change occurs for thesingle-neuron solution. On the other hand, regions with K < 0show bistable and incoherent solutions and are characterizedby cos � = 0; for the bistable case, θi ∈ A, which is the firstfixed point θi = 0, and θi ∈ B, which is the second fixed pointθi = π ; for the incoherent oscillations, the θi are distributedacross the closed orbit. This leads to the following phaseevolution equation:
θi = F (θi) + K sin (θi) cos (θi). (13)
I II
Bis
tabl
eM
onos
tabl
e
Synchrony
Incoherence
Mon
osta
ble
Synchrony
Incoherence
Bis
tabl
e
Synchrony
Incoherence
Mon
osta
ble
Bis
tabl
e
0 0.2 0.4 0.6 0.8 1
0 0 0
1
-1 -1
1 1
-1
(a) (b) (c)
K
I0 0.2 0.4 0.6 0.8 1
I0 0.2 0.4 0.6 0.8 1
I
FIG. 4. Parameter space with global electric coupling for (a) phase model, (b) phase model with amplitude correction, and (c) fullmodel.
051909-4
PHASE DESCRIPTION OF SPIKING NEURON NETWORKS . . . PHYSICAL REVIEW E 83, 051909 (2011)
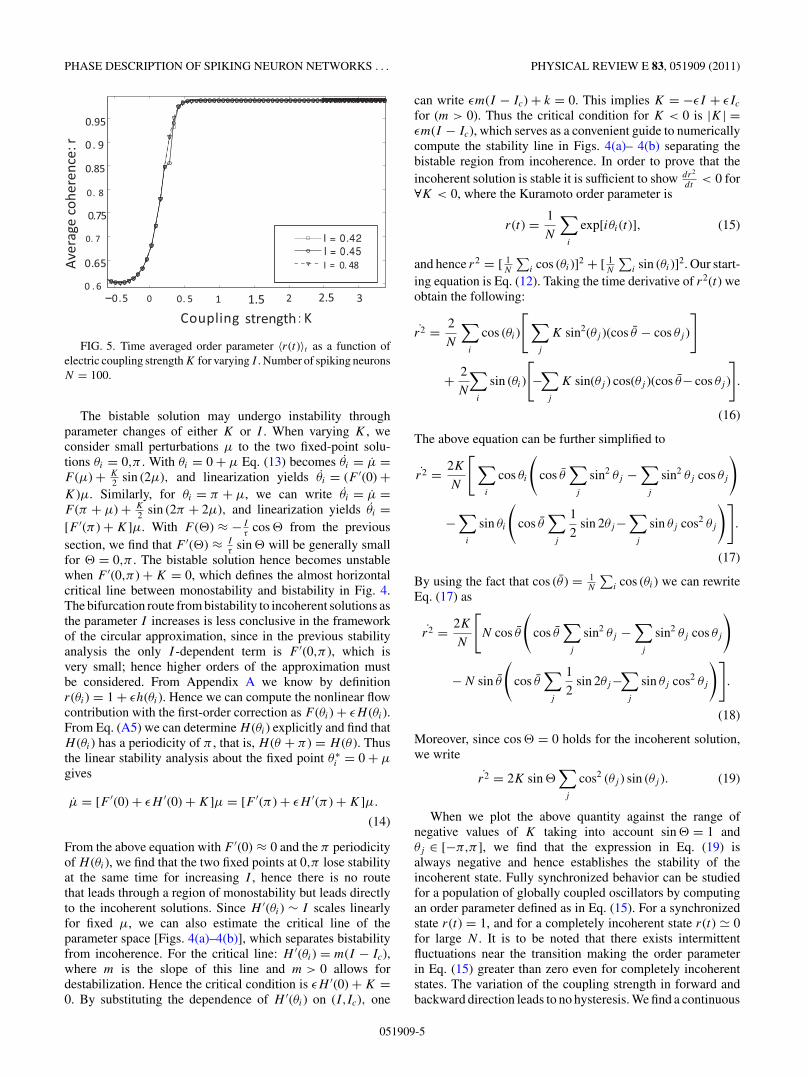
FIG. 5. Time averaged order parameter 〈r(t)〉t as a function ofelectric coupling strength K for varying I . Number of spiking neuronsN = 100.
The bistable solution may undergo instability throughparameter changes of either K or I . When varying K , weconsider small perturbations μ to the two fixed-point solu-tions θi = 0,π . With θi = 0 + μ Eq. (13) becomes θi = μ =F (μ) + K
2 sin (2μ), and linearization yields θi = (F ′(0) +K)μ. Similarly, for θi = π + μ, we can write θi = μ =F (π + μ) + K
2 sin (2π + 2μ), and linearization yields θi =[F ′(π ) + K]μ. With F (�) ≈ − I
τcos � from the previous
section, we find that F ′(�) ≈ Iτ
sin � will be generally smallfor � = 0,π . The bistable solution hence becomes unstablewhen F ′(0,π ) + K = 0, which defines the almost horizontalcritical line between monostability and bistability in Fig. 4.The bifurcation route from bistability to incoherent solutions asthe parameter I increases is less conclusive in the frameworkof the circular approximation, since in the previous stabilityanalysis the only I -dependent term is F ′(0,π ), which isvery small; hence higher orders of the approximation mustbe considered. From Appendix A we know by definitionr(θi) = 1 + εh(θi). Hence we can compute the nonlinear flowcontribution with the first-order correction as F (θi) + εH (θi).From Eq. (A5) we can determine H (θi) explicitly and find thatH (θi) has a periodicity of π , that is, H (θ + π ) = H (θ ). Thusthe linear stability analysis about the fixed point θ∗
i = 0 + μ
gives
μ = [F ′(0) + εH ′(0) + K]μ = [F ′(π ) + εH ′(π ) + K]μ.
(14)
From the above equation with F ′(0) ≈ 0 and the π periodicityof H (θi), we find that the two fixed points at 0,π lose stabilityat the same time for increasing I , hence there is no routethat leads through a region of monostability but leads directlyto the incoherent solutions. Since H ′(θi) ∼ I scales linearlyfor fixed μ, we can also estimate the critical line of theparameter space [Figs. 4(a)–4(b)], which separates bistabilityfrom incoherence. For the critical line: H ′(θi) = m(I − Ic),where m is the slope of this line and m > 0 allows fordestabilization. Hence the critical condition is εH ′(0) + K =0. By substituting the dependence of H ′(θi) on (I,Ic), one
can write εm(I − Ic) + k = 0. This implies K = −εI + εIc
for (m > 0). Thus the critical condition for K < 0 is |K| =εm(I − Ic), which serves as a convenient guide to numericallycompute the stability line in Figs. 4(a)– 4(b) separating thebistable region from incoherence. In order to prove that theincoherent solution is stable it is sufficient to show dr2
dt< 0 for
∀K < 0, where the Kuramoto order parameter is
r(t) = 1
N
∑i
exp[iθi(t)], (15)
and hence r2 = [ 1N
∑i cos (θi)]2 + [ 1
N
∑i sin (θi)]2. Our start-
ing equation is Eq. (12). Taking the time derivative of r2(t) weobtain the following:
r2 = 2
N
∑i
cos (θi)
[ ∑j
K sin2(θj )(cos θ − cos θj )
]
+ 2
N
∑i
sin (θi)
[−∑
j
K sin(θj ) cos(θj )(cos θ− cos θj )
].
(16)
The above equation can be further simplified to
r2 = 2K
N
[ ∑i
cos θi
(cos θ
∑j
sin2 θj −∑
j
sin2 θj cos θj
)
−∑
i
sin θi
(cos θ
∑j
1
2sin 2θj−
∑j
sin θj cos2 θj
)].
(17)
By using the fact that cos (θ) = 1N
∑i cos (θi) we can rewrite
Eq. (17) as
r2 = 2K
N
[N cos θ
(cos θ
∑j
sin2 θj −∑
j
sin2 θj cos θj
)
−N sin θ
(cos θ
∑j
1
2sin 2θj−
∑j
sin θj cos2 θj
)].
(18)
Moreover, since cos � = 0 holds for the incoherent solution,we write
r2 = 2K sin �∑
j
cos2 (θj ) sin (θj ). (19)
When we plot the above quantity against the range ofnegative values of K taking into account sin � = 1 andθj ∈ [−π,π ], we find that the expression in Eq. (19) isalways negative and hence establishes the stability of theincoherent state. Fully synchronized behavior can be studiedfor a population of globally coupled oscillators by computingan order parameter defined as in Eq. (15). For a synchronizedstate r(t) = 1, and for a completely incoherent state r(t) � 0for large N . It is to be noted that there exists intermittentfluctuations near the transition making the order parameterin Eq. (15) greater than zero even for completely incoherentstates. The variation of the coupling strength in forward andbackward direction leads to no hysteresis. We find a continuous
051909-5

DIPANJAN ROY, ANANDAMOHAN GHOSH, AND VIKTOR K. JIRSA PHYSICAL REVIEW E 83, 051909 (2011)
phase transition of the order parameter toward r(t) = 1, adistinct signature for supercritical Hopf transition near whichthe order parameter r(t) ≈ √
(K − Kc) as shown in Fig. 5.
B. Synaptic coupling
Here we use a globally coupled network model withchemical synapses such as the one proposed by Golomb andRinzel [32,33]. A single neuron in our model system of N
globally synaptically coupled (all-to-all) neurons is similarlydescribed by a set of three nonlinear differential equations:
xi = −yi − x3i + xi + I + gS(xi − vth) , (20)
yi = [−yi + f (xi)]/τ , (21)
si = asi(xi)(1 − si) − si/β, (22)
f (xi) ={
0 for xi < x0
c(xi + x0) for xi � x0.(23)
The collective synaptic action is given by S = 1N
∑Ni=1 si ,
where asi(xi) = 1
[1+exp(−xi/2)] . The synaptic constant g is thesame for all the neurons. In the case of a fast synapse(AMPA-type glutamate receptors), such as those found inthe auditory system, the rise time is instantaneous, and post-synaptic responses commence almost instantaneously afterthe start of presynaptic action potential [34,35]. This briskcommunication is a consequence of rapid calcium-channelkinetics, which allows significant calcium entry during theupstroke of the presynaptic action potential [36]. The timecourse of the postsynaptic conductivity caused by an activationof AMPA receptors can be captured by a rise time βrise =0.09 ms and decay time βdecay = 1.5 ms [37]. For our simulationwe use these values as guidelines. Under the fast synapseapproximation the variable si relaxes much more rapidly thanxi , in which case we may apply a quasistatic approximationto (22), si ≈ 0, allowing us to adiabatically eliminate thesynaptic variable via si = β
[1+β+exp(−xi/2)] . From numericswe find this approximation provides good results for τs in therange between 0.01 and 0.5 ms. Then the generalized phase
equation in the limit of a circular orbit with synaptic couplingis as follows:
θi = F (θi) − � sin θi(cos θi − vth) + O(ε), (24)
where the mean field for synaptic action for a given populationof neurons is expressed as
� = g
N
N∑l=1
β[1 + β + exp
( − cos θl
2
)] , i �= l, (25)
and higher orders of ε are absorbed in O(ε).Figure 6 shows the parameter space diagram for the full
and phase models presented in Eqs. (20)–(23) and Eq. (24).Over a wide range of I-g values the collective dynamics of thetwo systems primarily show four distinct regions of interest,which are close to each other in the parameter space. However,there are qualitative differences in the collective dynamicscompared to electric coupling in the previous section. Fromsimulation we find both the full model and the reduced modelexhibit bistability and the existence of a separatrix for negativeas well as positive values of g (inhibitory and excitatorycoupling, respectively). For decreasing coupling strength g,the network displays monostable behavior. Increasing I forfixed values of g results in a phase transition from bistablebehavior to monostable behavior. With further increase ofthe current I > Ic both networks become oscillatory througha saddle-node bifurcation. Now, depending on the couplingstrength, the entire population of neurons either synchronizesor shows incoherent behavior about their center of mass. Wehave obtained numerically the boundaries that separate thesefour regions in the parameter space. For the entire simulation,we fixed the reversal potential of potassium ions to vth ≈ −1.0.The parameter space structure for both planar and phasemodels are obtained numerically with standard bifurcationanalysis software [38,39]. We test our approximation for fastsynapses numerically and find excellent agreement betweenthe planar model and reduced phase model for different valuesof the time constant β.
Following the same lines of thought as we did for theelectric coupling, we consider the instability of the bistable
Synchrony
Incoherence
Synchrony
Incoherence
Synchrony
Incoherence
Bis
tabl
e
Bis
tabl
e
Bis
tabl
e
Mon
osta
ble
Mon
osta
ble
Mon
osta
ble
0 0.40.2 0.6 0.8 1I
0 0.40.2 0.6 0.8 1I
0 0.40.2 0.6 0.8 1I
g
-1 -1 -1
000
1 1 1
I II
(a) (b) (c)
FIG. 6. Parameter space with global synaptic coupling for the (a) phase model, (b) phase model with amplitude correction and (c) fullmodel.
051909-6

PHASE DESCRIPTION OF SPIKING NEURON NETWORKS . . . PHYSICAL REVIEW E 83, 051909 (2011)
dynamic region for increasing parameters I . In particular, wewish to understand why for synaptic coupling the bifurcationpath leads us always through the monostable region towardincoherence. The governing equation for the phase networkevolution with synaptic coupling reads
θi = [F (θi) − � sin θi(cos θi − Vth)], (26)
where � = g
N
∑Nj=1
β
[1+β+exp(− cos θj
2 )]. As observed numerically
the bistable solution has two fixed points θ∗ = 0,π , whereθ∗ = 0 implies �0 = g
β
[1+β+exp(− 12 )]
and subsequently for
θ∗ = π , �π = gβ
[1+β+exp( 12 )]
. Note that � �= �(�) and � =�0 + �π = gβ( 1
[1+β+exp(− 12 )]
+ 1[1+β+exp( 1
2 )]). In the following
we perform the linear stability analysis around these two fixedpoints.
(a) For θi = 0 + μ we write
θi = μ = F (μ) − � sin 2θi
2+ �Vth sin θi . (27)
Linearization of the above leads to μ = [F ′(0) − � cos(0) +�Vth cos(0)]μ, and thus μ = [F ′(0) − (1 − Vth)�]μ = λμ. Toallow for a direct comparison with the electric coupling,we consider also noncircular contributions, r(θ ) = 1 + εh(θ )and F (θ ) + εH (θ ). This additional contribution extends thestability analysis to
θi = μ = [F ′(0) + εH ′(0) − �(1 − Vth)]μ, (28)
where F ′(0) ≈ 0, H ′(0) = H ′(π ). Then the linear stability isgiven by λ0 = εH ′(0) − �(1 − Vth).
(b) For the second fixed point θi = π + μ in Eq. (26) wewrite after linearization
θi = μ = [F ′(π ) − � cos(2π ) + �Vth cos(π )]μ, (29)
which reduces to μ = (F ′(π ) − �(1 + Vth)μ. Again, whenextending toward noncircular contributions we obtain
θi = μ = [F ′(π ) + εH ′(π ) − �(1 + Vth)]μ, (30)
where F ′(π ) ≈ 0 and λπ = εH ′(π ) − �(1 + Vth).For Vth > 0 the following inequality is satisfied: 0 >
λ0 > λπ which demonstrates that θi = 0 is less stable anddestabilizes first for increasing I [note H (θ ) scales linearlyin I ]. In other words, the translational symmetry of the phaseflow under translation of π is broken for synaptic coupling.As a consequence, the bifurcation path from bistability toincoherence always leads through monostability.
IV. SUMMARY
Here we have developed a phase description of globallycoupled neurons with fast chemical synapses and biologicallyrealistic electric synapses, which is not constrained to the limitcycle regime but extends to the excitable regime displayingfeatures of multistability and threshold behavior. We demon-strated analytically that the electric and synaptic couplingsassume different mathematical forms in the phase descriptionand tested their validity computationally. Notably, the synapticcoupling is expressed in a different mathematical form in thereduced equations than so far assumed in the literature [25,27,40–43]. The previously assumed coupling expression correctly
reflects gap junction coupling, but does not reflect synapticcoupling. The latter assumes a form of multiplicative type. Thetwo different types of coupling are exact for circular orbits andare valid approximations of the network dynamics for smalldeviations from circular symmetry. This finding is meaningful,since the resulting qualitative network behaviors are different.The advantage of the phase description of the network dynam-ics is its simpler mathematical form. Key to the here appliedapproximation is the assumption of amplitude independence ofthe couplings. This assumption is trivially satisfied for circularorbits. For more realistic scenarios, there will be a smooth de-pendence of the amplitude on the phase angle, which is whereour approximations hold well. Nevertheless they will fail whenthe dependencies become too complex, such as in weaklycoupled chaotic oscillators. It is important to note that the cou-plings in the phase descriptions maintain their mathematicalexpression plus some linearly added correction terms, whichscale with the degree of order of deviation from the circle. As aconsequence of this additivity of couplings and deviations fromcircularity, the here presented networks of phase oscillatorsoffer a reasonable simple framework for the study of spikingneurons with mixed couplings and close to circular orbits.
ACKNOWLEDGMENTS
This research has been supported by the ATIP Plus Program(CNRS) and the James S. McDonnell Foundation.
APPENDIX A: DEVIATION FROM CIRCULARAPPROXIMATION OF THE HOMOCLINIC ORBIT
The planar network model under global synaptic couplingis governed by the following equations:
xi = −yi − x3i + xi + I + gS(t)(xi − vth) , (A1)
yi = [−yi + f (xi)]/τ , (A2)
si = asi(xi)(1 − si) − si/β, (A3)
f (xi) ={
0 for xi < x0
c(xi + x0) for xi � x0.(A4)
Using the approximation of adiabatic elimination (seemain text) Eq. (A3) assumes the following form: si =
β
[1+β+exp(−xi/2)] . Now we make the following ansatz: [xi +i(yi − y0)] = r(θi) exp(iθi), where x = r(θi) cos θi and y =y0 + r(θi) sin θi , where i = 1 . . . N . Substituting this in ourgoverning equations [Eqs. (A1)–(A3)] with the definitionr(θi) = 1 + εh(θi) allows us to write the phase evolutionequation for our network as follows:
θi = 1
r ′2+r2
{[r ′ cos(θi)−r(θi) sin(θi)][(I − y0)−r(θi) sin(θi)
− r3(θi) cos3(θi) + r(θi) cos(θi) + Isynapse]
+ r ′ sin(θi) + r(θi) cos(θi)
τ[−y0−r(θi) sin(θi) + f (θi)]
},
(A5)
051909-7

DIPANJAN ROY, ANANDAMOHAN GHOSH, AND VIKTOR K. JIRSA PHYSICAL REVIEW E 83, 051909 (2011)
f (θi) ={
0 for r(θi) cos θi < x0
c[
r ′ sin θi+r(θi ) cos θi
τ
][r(θi) cos θi + x0] for r(θi) cos θi � x0.
(A6)
In the above equations r ′ = dr(θ)dθ
. Hence, the denomi-nator in the above expression assumes the following form:r ′2 + r2 = [1 + εh(θi)]2 + ε2h′2. In Eq. (A5) an explicit formof Isynapse is
Isynapse = B
B1
g
N
N∑j=1
β
{1 + β + exp[−r cos(θj )/2]} , (A7)
where we have collected the various orders of ε in the termsof B and B1, in particular,
B = (Vth sin θi − 0.5 sin 2θi) + ε[h′ cos2 θi − h(θi) sin 2θi]
−Vth[h′ cos θi − h(θi) sin θi] + O(ε2), (A8)
and B1 is [1 + εh(θi)]2 + ε2h′2. When carrying out a Taylorexpansion of 1/B1 for small ε, we obtain
1
B1= [1 − 2εh(θi)] + O(ε2). (A9)
Also, from Eq. (A7) the mean field of synaptic action S is
S =N∑
j=1
β
{1 + β + exp[−r cos(θj )/2]} . (A10)
The above expression can be rearranged and written in thefollowing manner:
S =N∑
j=1
β{1 + β + exp[−0.5 cos(θj )] exp
[ − ε2h(θj ) cos θj
]} .
(A11)
A Taylor series expansion for small ε in the denominator ofEq. (A11) yields
S(θj ) =N∑
j=1
β
γ
1[1 − ε
exp(−0.5 cos θj )2γ
h(θj ) cos θj + O(ε2)] .
(A12)
We can further approximate Eq. (A12) in ε and obtain
S(θj ) =N∑
j=1
β
γ
[1+ε
exp(−0.5 cos θj )
2γh(θj ) cos θj+O(ε2)
],
(A13)
where γ = [1 + β + exp(−0.5 cos θj )]. Equations (A7) and(A8) together with Eqs. (A9) and (A13) can now be condensedinto a single equation that describes the total synaptic currentin the phase model
Isynapse = g/N
N∑j=1
β
γ
[1 + ε
exp(−0.5 cos θj )
2γh(θj ) cos θj
]
× [1 − 2εh(θi)](Vth sin θi − 0.5 sin 2θi)
+ ε[h′ cos2 θi − h(θi) sin 2θi]−Vth[h′ cos θi
−h(θi) sin θi] + O(ε2). (A14)
We rewrite the above equation by collecting O(ε) contribu-tions:
Isynapse = g/N
N∑j=1
β
γ
[(Vth sin θi − 0.5 sin 2θi)
+ ε
((Vth sin θi − 0.5 sin 2θi)
exp(−0.5 cos θj )
2γ
×h(θj ) cos(θj ) − 2h(θi)(Vth sin θi − 0.5 sin 2θi)
+{h′ cos2 θi − h(θi) sin 2θi − Vth[h′ cos θi
−h(θi) sin θi]})]
+ O(ε2). (A15)
Equation (A13) describes the modification of the mean fieldof synaptic action for the population of neurons under a smalldeviation of the amplitude from circularity, and Eq. (A15)accounts for the corresponding total synaptic current. Allderivations are carried out without assuming a specific form offunction h(θ ). In the main text we approximated numericallyan expression for h(θ ) including the first leading expressionsof a Fourier series. Assuming here a simpler specific form suchas h(θ ) = sin θ for the illustration of the correction term in thesynaptic current, Eq. (A15) can be expressed as
Isynapse = g/N
N∑j=1
β
γ
((Vth sin θi − 0.5 sin 2θi)
+ ε
{(Vth sin θi − 0.5 sin 2θi)
exp(−0.5 cos θj )
2γ
× sin θj cos θj − 2 sin θi(Vth sin θi − 0.5 sin 2θi)
+ [cos3 θi − sin θi sin 2θi − Vth(cos2 θi
− sin2 θi)]
})+ O(ε2). (A16)
As ε → 0 the total synaptic current in Eq. (A14) reduces tothe same expression as obtained in Eq. (24).
APPENDIX B: COMPARISON BETWEEN THECOLLECTIVE DYNAMICS OF A NETWORK OF NEURONS
AND A NETWORK OF A PHASE MODEL
In this section we test the generality of the couplingsat the network level that we derived so far in this work.We computationally test our proposition in this paper byconsidering a globally synaptically coupled network of Class Iexcitable neurons such as Hodgkin-Huxley (HH) neurons [44].
051909-8

PHASE DESCRIPTION OF SPIKING NEURON NETWORKS . . . PHYSICAL REVIEW E 83, 051909 (2011)
I I
Synchrony
Incoherence
Synchrony
Incoherence
Mon
osta
ble
Bis
tabl
e
Mon
osta
ble
0 0.2 0.4 0.6 0.8 1 0 0.2 0.4 0.6 0.8 1
g
I I
a
a
1 1
0 0
-1 -1
(a) (b)
FIG. 7. Parameter space with global synaptic coupling for(a) phase model and (b) HH model.
A single neuron is described by standard HH equations, andsynaptically coupled network equations are
Vi = −0.1(Vi + 65) − 35m3∞hi(Vi − 55) − 9n4
i (Vi
+ 90) + I + gS(xi − Vth), (B1)
ni = 5[ani
Vi(1 − ni) − bniVi
], (B2)
hi = 5[ahi
Vi(1 − hi) − bhiVi
], (B3)
si = asi(Vi)(1 − si) − si/β, (B4)
where all the gating functions used here are taken fromRef. [45]. As before the mean synaptic action is givenby S = 1
N
∑Ni=1 si , where asi
(Vi) = 1[1+exp(−Vi/2)] . Here we
simulate a network of 100 neurons of the above type withrandom initial conditions and compare the (I, g) parameterspace with a globally coupled network of phase model as inEq. (24). As appears in Fig. 7 from the partitioning of theparameter space (I, g) to a good degree, the dynamics of anensemble of biologically realistic neural oscillators resemblesthe dynamics obtained from phase-coupled neural oscillators.It is noteworthy there are some differences in the dynamicsof these two networks that arise as we take I close to 0.0. Incase of HH networks we do not obtain any further states inthe parameter space; however, for a network of phase-coupledneurons we obtain a bistable behavior in a narrow region ofparameter space.
[1] C. Morris and H. Lecar, Biophys. J. 35, 193 (1981).[2] D. A. McCormick, B. W. Connors, J. W. Lighthall, and D. A.
Prince, J. Neurophysiol. 54, 782 (1985).[3] A. Kaske and N. Bertschinger, Biol. Cybern. 92, 3 (2005).[4] B. S. Gutkin, G. B. Ermentrout, and A. D. Reyes,
J. Neurophysiol. 94, 1623 (2005).[5] B. Naundorf, F. Wolf, and M. Volgushev, Nature (London) 440,
7087 (2006).[6] E. M. Izhikevich, Dynamical Systems in Neuroscience: The
Geometry of Excitability and Bursting (Computational Neuro-science) (MIT Press, Cambridge, MA, 2006), p. 457.
[7] J. L. Hindmarsh and R. M. Rose, Proc. R. Soc. Lond. B 221, 87(1984).
[8] X. J. Wang, Neuroscience 59, 21 (1994).[9] P. F. Pinsky and J. Rinzel, J. Comput. Neurosci. 1, 39
(1994).[10] J. C. Brumberg and B. S. Gutkin, Brain Res. 1171, 122
(2007).[11] A. Ghosh, D. Roy, and V. K. Jirsa, Phys. Rev. E 80, 041930
(2009).[12] G. B. Ermentrout and N. Kopell, SIAM J. Appl. Math. 46, 359
(1986).[13] E. M. Izhikevich, Int. J. Bifurcation Chaos 10, 1171 (2000).[14] B. S. Gutkin and G. B. Ermentrout, Information Processing in
Cells and Tissues (Plenum Press, New York, 1998), p. 57.[15] B. S. Gutkin, J. Jost, and H. C. Tuckwell, Europhys. Lett. 81,
20005 (2008).[16] D. Pazo and E. Montbrio, Phys. Rev. E 73, 055202(R)
(2006).[17] G. B. Ermentrout, Neural Comput. 6, 679 (1994).[18] F. C. Hoppendsteadt and E. M. Izhikevich, Weakly Connected
Neural Networks (Springer-Verlag, New York, 1997).[19] E. S. Brown, J. Mohelis, and P. Holmes, Neural Comput. 16, 4
(2004).
[20] S. Coombes, Phys. Rev. E 67, 041910 (2003).[21] T. B. Kepler, L. F. Abbott, and E. Marder, Biol. Cybern. 66, 381
(1992).[22] Y. Kuramoto, Chemical Oscillations, Waves and Turbulence
(Springer-Verlag, New York, 1984).[23] R. M. Smeal, G. B. Ermentrout, and J. A. White, Phil. Trans. R.
Soc. B 365, 2407 (2010).[24] V. Jirsa and A. McIntosh, Handbook of Brain Connectivity
Understanding complex systems Springer complexity. NonlinearDynamics (Springer, Berlin, 2007).
[25] A. Tonnelier and W. Gerstner, Phys. Rev. E 67, 021908(2003).
[26] S. Coombs, SIAM J. Appl. Dyn. Syst. 7, 1101 (2008).[27] D. T. W. Chik, S. Coombes, and Z. D. Wang, Phys. Rev. E 70,
011908 (2004).[28] S. H. Strogatz, Nonlinear Dynamics and Chaos: With Applica-
tion to Physics, Biology, Chemistry, and Engineering (WestviewPress, 2000), p. 498.
[29] C. G. Assisi, V. K. Jirsa, and J. A. S. Kelso, Phys. Rev. Lett. 94,018106 (2005).
[30] M. Dhamala, V. K. Jirsa, and M. Ding, Phys. Rev. Lett. 92,028101 (2004).
[31] M. Dhamala, V. K. Jirsa, and M. Ding, Phys. Rev. Lett. 92,074104 (2004).
[32] X. J. Wang and J. Rinzel, Neural Comput. 4, 84 (1992).[33] D. Golomb and J. Rinzel, Phys. Rev. E 48, 4810 (1993).[34] E. C. Keen and A. J. Hudspeth, Proc. Natl. Acad. Sci. 103, 14
(2006).[35] H. Parnas and I. Parnas, J. Membr. Biol. 142, 267
(1994).[36] B. L. Sabatini and W. G. Regher, Nature (London) 384, 170
(1996).[37] F. Gabbiani, J. Midtgarred, and T. Knopfel, J. Neurophysiol. 72,
999 (1994).
051909-9

DIPANJAN ROY, ANANDAMOHAN GHOSH, AND VIKTOR K. JIRSA PHYSICAL REVIEW E 83, 051909 (2011)
[38] E. J. Doedl, [http://indy.cs.concordia.ca/auto/].[39] R. H. Clewley, W. E. Sherwood, M. D. LaMar, and J. M.
Guckenheimer, [http://pydstool.sourceforge.net].[40] M. Bazhenov, R. Huerta, M. I. Rabinovich, and T. Sejnowskic,
Physica D 116, 3 (1998).[41] M. Bazhenov, N. F. Rulkov, and I. Timofeev, J. Neurophysiol.
100, 1562 (2008).
[42] C. Koch, Biophysics of Computation (Oxford University Press,New York, 1999).
[43] W. Gerstner and W. Kistler, Spiking Neuron Models: AnIntroduction (Cambridge University Press, Cambridge, 2002).
[44] A. L. Hodgkin and A. F. Huxley, J. Physiol. 117, 500 (1952).[45] N. Kopell and G. B. Ermentrout, Proc. Natl. Acad. Sci. USA
101, 15482 (2004).
051909-10