Embed Size (px)
Citation preview

Perilipin Family Members Preferentially Sequester to Either Triacylglycerol-
or Cholesteryl Ester-Specific Intracellular Lipid Storage Droplets
Kai Hsieh*, Yun Kyung Lee*, Constantine Londos*, Bruce M. Raaka+, Knut Tomas Dalen*#, and Alan R. Kimmel*^
* Laboratory of Cellular and Developmental Biology, + Clinical Endocrinology Branch,
National Institute of Diabetes and Digestive and Kidney Diseases The National Institutes of Health, Bethesda, MD 20892
and
# Department of Nutrition, Institute of Basic Medical Sciences, Medical Faculty,
University of Oslo, P.O. Box 1046 Blindern, N-0316 Oslo, Norway
^ Telephone: (301) 496-3016 Facsimile: (301) 496-5239 e-mail: [email protected]
Running Title: Perilipin Lipid Specificities
Key Words: PLIN, ADRP, TIP47, LSDP5, S3-12, triacylglyceride, cholesterol, fatty acids,
lipolysis
© 2012. Published by The Company of Biologists Ltd.Jo
urna
l of C
ell S
cien
ceA
ccep
ted
man
uscr
ipt
JCS online publication date 8 June 2012

2
SUMMARY
Perilipin family proteins (Plins) coat the surface of intracellular neutral lipid
storage droplets in various cell types. Studies across diverse species
demonstrate that Plins regulate lipid storage metabolism through recruitment of
lipases and other regulatory proteins to lipid droplet surfaces. Mammalian
genomes encode 5 distinct Plin gene members and additional protein forms
derive from specific mRNA splice variants. However, it is not known if the
different Plins have distinct functional properties. Using biochemical, cellular
imaging, and flow cytometric analyses, we now show that within individual cells
of various types, the different Plin proteins preferentially sequester to separate
pools of lipid storage droplets. By examining ectopically expressed GFP fusions
and all endogenous Plin protein forms, we demonstrate that different Plins
sequester to lipid droplets, comprised distinctly of either triacylcerides or of
cholesterol esters. Further, Plins with strong association preferences to TAG (or
CE) droplets can re-direct the relative intracellular TAG/CE balance toward the
targeted lipid. Our data suggest diversity of Plin function, alter previous
assumptions about shared collective actions of the Plins, and indicate that each
Plin can have separate and unique functions.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

3
INTRODUCTION
Intracellular neutral lipid storage droplets (LSDs) are unique organelles
that store metabolic precursors of cellular energy, membrane biosynthesis,
steroid hormone synthesis, and signaling (Farese and Walther, 2009; Kimmel et
al., 2010; Londos et al., 2005). LSDs contain different lipids (e.g. triacylglycerides
(TAG) or cholesteryl esters (CE) at their core, surrounded by a phospholipid
monolayer. LSD surfaces in organisms as diverged as mammals, Drosophila,
and Dictyostelium are targeted by an evolutionarily related family of proteins
(Kimmel et al., 2010; Lu et al., 2001; Miura et al., 2002), the Perilipins (Plins).
Mammalian genomes encode 5 distinct Plin gene members and additional
protein forms derived from specific mRNA splice variants (Kimmel et al., 2010).
Plin1 is the major LSD coat protein in adipocytes and steroidogenic cells
(Greenberg et al., 1993; Servetnick et al., 1995). Other Plins exhibit different
expression patterns. Plin2 is the predominant, but not exclusive, form in liver
(Dalen et al., 2006), whereas Plin5 is primarily expressed in oxidative tissues,
including heart, soleus muscle, and brown fat (Dalen et al., 2007; Wolins et al.,
2006; Yamaguchi et al., 2006). Based on Plin1 function (Martinez-Botas et al.,
2000; Sztalryd et al., 2003; Tansey et al., 2001; Wang et al., 2009), the Plins are
viewed as fundamental regulators of lipolytic activity. Loss of Plin1 (Martinez-
Botas et al., 2000; Tansey et al., 2001) or Plin2 (Chang et al., 2006) in mice
significantly reduces intracellular lipid levels in adipocytes and hepatocytes,
respectively. Furthermore, heterozygous loss-of-function mutations in human
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

4
PLIN1 leads to a familial partial lipodystrophy, supporting a required role for
Perilipin in TAG storage within human adipocyte LSDs (Gandotra et al., 2011).
Regardless, little is known of lipid interaction specificity of the various Plins. Here,
we show that distinct Plins differentially sequester to either TAG- or CE-specific
LSDs and can alter relative intracellular TAG or CE levels toward the
preferentially targeted lipid. These data demonstrate and emphasize diverse
functions for the different Plins.
RESULTS
Exogenous fatty acids and cholesterol differentially stabilize accumulation
of Plin protein family members.
Intracellular LSDs accumulate substantially when cells are cultured
overnight in the presence of high concentrations of various exogenous lipids (Xu
et al., 2005). Since Plins primarily sequester to LSD surfaces (Miura et al., 2002),
we determined if different Plins exhibited differential regulation in response to
either fatty acids or cholesterol, lipids that mobilize separate pathways.
Y1 mouse adrenocortical cells have robust capacity for steroid hormone
synthesis and accumulate TAG and CE LSDs as energy and metabolic precursor
stores. Further, steroidogenic cells are able to synthesize all 4 Plin1 mRNA splice
variants (Servetnick et al., 1995; Xu et al., 2005) and express all other Plin genes.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

5
Y1 cells were cultured under standard conditions or in medium
supplemented with oleic acid and/or cholesterol. Endogenous Plin proteins were
quantified in whole cell lysates by specific immunoblotting (Figure 1A). In general,
none of the Plins exhibited significant accumulation in unsupplemented medium.
However, significant Plin accumulation differences were observed in the
presence of oleic acid or cholesterol. The two major Plin1 variants of
steroidogenic cells, Plin1a and Plin1c, exhibited reciprocal patterns. Plin1a was
enhanced by oleic acid, but not by cholesterol, whereas the Plin1c response was
exactly opposite (Figure 1A). The effects were largely activating, since the
expressions of Plin1a and Plin1c were not diminished in cells cultured
simultaneously with oleic acid and cholesterol. Plin1b and Plin1d proteins are not
easily detected in Y1 cells (Servetnick et al., 1995), although Plin1b appears to
be regulated similarly to Plin1a (Figure 1A).
Plin2 and Plin3 accumulate similarly regardless of the exogenous lipid
moiety, although Plin2 may be slightly more responsive to oleic acid. Conversely,
Plin4 and Plin5 show extreme lipid specificity, largely mimicking that of Plin1c
and Plin1a, respectively (Figure 1A).
Since exogenous lipids may have differential regulatory effects on the
transcription or translation of endogenous Plin mRNAs and, thus, indirectly
impact Plin protein accumulation, we also examined the effects of oleic acid and
cholesterol using GFP-Plin protein fusions expressed from identical constitutively
active promoter vectors. McARH7777 rat hepatoma cells were transiently
transfected with vectors to separately express each GFP-Plin protein fusion and
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

6
cultured under standard conditions or in medium supplemented with oleic acid
and/or cholesterol.
The GFP-Plin proteins in McARH7777 cells showed identical responses to
oleic acid and cholesterol as their endogenous counterparts in Y1 cells (Figures
1A and 1B). GFP-Plin1a and GFP-Plin5 were specifically responsive to the
positive effects of oleic acid, whereas GFP-Plin1c and GFP-Plin4 were only
detected in the presence of cholesterol (Figure 1B). GFP-Plin2 and GFP-Plin3
did not show a significant preference to either lipid. The responses of Plin1b and
Plin1d were more clear in this heterologous expression system. Plin1b is
structurally most similar to Plin1a (Lu et al., 2001) and GFP-Plin1b behaves
identically to both Plin1a and GFP-Plin1a. Plin1d is the smallest variant (Lu et al.,
2001) and GFP-Plin1d shows limited lipid preference, acting more like Plin2 and
Plin3.
Differential sub-cellular localizations of FA- or Chl- tagged markers.
Data (Figure 1) suggest that the sequestration and stabilization of
individual Plins to different classes of LSDs may influence the specific
accumulation of particular Plin proteins. To examine this further, we established
conditions to preferentially tag and purify TAG- and CE-specific LSDs.
Y1, McARH7777, and AML12 cells were cultured overnight in the
presence of both oleic acid and cholesterol, plus BODIPY 558/568 C12 [as a
fluorescent fatty acid (FA) dye marker] and cholesteryl BODIPY 500/510 FL C12
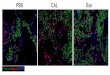
[as a fluorescent cholesteryl (Chl) dye marker] and imaged (Figure 2A). All three
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

7
types of cells showed definitive separation FA (red) and Chl (green) markers into
distinct LSDs clusters. Few sectors show any co-localization, a pattern similarly
observed in 4T1 mouse mammary tumor, primary mouse liver, C2C12 mouse
myoblast, 3T3-L1 mouse fibroblast, J774A.1 mouse monocyte-macrophage, and
CHOK1 Chinese hampster ovary cells (Supplemental Figure S1A). In general,
the different markers, though distinctly separate, were largely intermingled.
However, the McARH7777 cells (Figure 2A) were most distinctive. FA- and Chl-
tagged droplets segregated to entirely unique sub-cellular regions. FA-tagged
droplets were polarized to the cell periphery, while Chl-tagged droplets were
centrally localized. Less distinctive labeling and polarized separation of the
droplets is seen in the McARH7777 cells cultured for shorter incubation periods
(Supplemental Figure S1B).
The polarized localizations of FA- and Chl-tagged droplets suggested that
it might be possible to distinguish separate associations of these distinct droplets.
McARH7777 cells were cultured with oleic acid, cholesterol, and FA- or Chl-
tagged fluorescent markers, and co-stained using antibodies against several
organelle-specific proteins (Figure 2B) or organelle-specific dye trackers (Figure
2C). In McARH7777 cells, the FA-tagged droplets localized entirely separate
from lysosomes, mitochondria, and early endosomes (Figures 2B,C). The Chl-
tagged droplets and other organelles are more centrally localized and perinuclear.
Nonetheless, there was nominal overlap of Chl-tagged droplets with these other
structures (Figures 2B,C). Lysosomes and mitochondria remained largely
separate, and there was minimal intermingling with early endosomes. The data
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

8
indicate that the FA- or Chl- fluorescent tags do not broadly label these other
organelles.
FACS purification of TAG- and CE-LSDs.
To determine if FA- and Chl-tagged LSDs had distinct lipid compositions,
we first developed conditions for their separate purifications. Labeled Y1,
McARH7777, and AML12 cells were lysed, LSDs floated by gradient
centrifugation, and the FA- and Chl-fluorescently tagged LSDs separated by
FACS (Figure 3A and Supplemental Figures S2A,B,C). The FA- and Chl- specific
labelings were relatively similar and reproducible. Generally >70% of the particles
were dye-tagged in dual label experiments. FACS separations were distinct with
<5% of unsorted particles carrying both FA and Chl dye markers (Figures 3A,B).
The particles also showed reasonable size homogeneity; ~80% of all particles
had diameters of 2-6 μm. Mixing experiments of labeled and unlabeled lysates
and particles showed <2% dye marker transfer and/or non-specificity
(Supplemental Figures S2A,B,C).
Lipids were extracted from sorted FA- and Chl-tagged LSDs and analyzed
and quantified by thin layer chromatography (Figure 3C, Table I, and
Supplemental Figure S3A). Total LSD content was similar within and between
cell types. The Chl-tagged droplets were primarily comprised of cholesteryl ester
and a smaller amount of cholesterol; we were unable to detect triacylglycerols,
fatty acids, and related metabolites in the sorted Chl-tagged droplets. Conversely,
the isolated FA-tagged droplets were comprised primarily of triacylglycerol and
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

9
minor quantities of metabolites (di- and mono-acylglycerides and fatty acids);
small amounts of cholesterol were present, but cholesteryl ester was not
detected. Thus, we have established conditions to fluorescently tag, image, and
purify LSDs specific to either CE or TAG.
We also traced the cell fate of the Bodipy precursor markers
(Supplemental Figure S3B). The native Bodipy-Chl dye marker migrates at a
unique position that differs from both cholesterol and choleterol ester and retains
its identical motility even after integration within the CE-droplets Thus, Bodipy-
Chl appears to be incorporated directly into CE droplets and is not significantly
metabolized into other droplet-specific lipids during the time frame studied.
Bodipy-FA migrates less far than does untagged FA, but changes mobility when
integrated into TAG droplets. We suggest that this represents the incorporation of
the Bodipy-FA into TAG, which migrates less far than untagged TAG and
distinctly from phospholipids. Regardless, the FA- and Chl-markers represent
specific tracer tags that allow us to separately purify the distinct populations of
LSDs that are biochemically enriched in either triacylglcerol (TAG) or cholesterol
ester (CE), hereafter referred to as TAG- or CE-LSDs, respectively.
Plin associations with TAG- or CE- LSDs.
We next examined Plin protein associations with isolated TAG- or CE-
LSDs. Y1 cells were cultured with oleic acid and cholesterol, plus FA- and Chl-
fluorescent markers. LSDs were floated, TAG- or CE-LSDs purified by FACS,
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

10
and associations of Y1 endogenous Plins within unsorted LSDs or to TAG- or
CE-specific populations then assayed by specific immunoblotting.
All of the Plin proteins were detected in the unsorted LSD population
(Figure 3D), consistent with previous data (Figures 1A,B). The relative increase
in endogenous Plin1b and Plin1d signals probably reflects the enrichment of
these forms in floating droplets compared to whole cell lysates (see Figures 1A,
3D). In this assay, the individual Plins show distinct sequestration to TAG- or CE-
specific LSDs. Plin1a, Plin1b, and Plin5 are restricted to TAG-populations,
whereas Plin1c and Plin4 localize with the CE-LSDs (Figure 3D). Plin2, Plin3,
and Plin1d show limited association preference (Figure 3D).
Sub-cellular localization of GFP-Plins to TAG- or CE-LSDs.
The whole cell accumulations of specific Plin proteins in response to either
oleic acid or cholesterol correlate with the associations of the same Plins, to
either TAG- or CE-LSDs (Figures 1,3D). Still, we wished to investigate specific
Plin/droplet associations by alternative methods. Since McARH7777 cells
accumulate TAG- and CE-LSDs that are completely separated (Figure 2A), we
could visually observe if GFP-Plins exhibited parallel localization differences. In
addition we could quantify the relative distributions of GFP-Plins among CE- or
TAG-LSDs by FACS segregation.
McARH7777 cells were transiently transfected individually with vectors
that specifically encode different eGFP-Plin fusions and cultured with oleic acid
and cholesterol plus either BODIPY 558/568 C12 (TAG marker) or cholesteryl
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

11
BODIPY 576/589 C11 (CE marker). Both dye markers have excitation/emission
spectra that differ from eGFP (488/509). GFP/TAG and GFP/CE experiments
were always performed in pairs and data analyzed in parallel to assess reciprocal
responses (Figures 4, 5, and 6 and Supplemental Figure S4).
GFP-Plin1a localizes to the polarized periphery of McARH7777 cells and
primarily with TAG-LSDs (Figure 4A and 6). In contrast, while CE-LSDs are
sequestered to the cell interior, none of the GFP-Plin1a signal co-localizes with
the CE marker (Figure 4A). FACS profiles of cell populations support these
conclusions; ~95% of expressed GFP-Plin1a co-sorts with the TAG marker, while
only ~5% GFP-Plin1a co-sorts with CE-LSDs (Figures 4A and 6).
GFP-Plin1b also exhibits a localization preference for TAG-LSDs
compared to CE-LSDs, although the FACS profiles show slightly less specificity
(Figures 4B and 6).
GFP-Plin1c shows the expected reciprocal pattern to GFP-Plin1a (see
Figures 1 and 3D). GFP-Plin1c localizes to the cell interior, surrounding CE-LSDs,
and distinctly separate from the polarized TAG-LSDs (Figure 4C). The FACS
data are consistent, where >90% of GFP-Plin1c sorts with CE and separate from
TAG (Figures 4C and 6).
The GFP fusions of Plin1d, Plin2, and Plin3 show minimal preference for
TAG- or CE-LSDs and segregate with both (Figures 4D, 5A, and 5B). All 3
protein fusions are found in both interior and peripheral cell regions, co-localize
with both TAG- and CE-LSDs within the cell, and co-segregate with both by
FACS, but to varying degrees (Figures 4D, 5A, 5B, and 6).
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

12
GFP-Plin4 and GFP-Plin5 exhibit reciprocal LSD preferences and are,
thus, respectively, similar to Plin1c and Plin1a (see Figures 1 and 3). While GFP-
Plin4 sequesters with CE-LSDs in the cell interior (Figure 5C), GFP-Plin5
polarizes with TAG-LSDs to the cell periphery (Figure 5D). FACS data
substantiate these preferences; ~85% of GFP-Plin4 co-sorts with a CE marker,
while ~85% of GFP-Plin5 segregates with TAG (Figures 5C, 5D, and 6).
Plins with TAG- or CE-binding preferences can alter cellular TAG/CE
distributions.
Loss of Plin1 and Plin2 in mice alters the targeted accumulation of lipid
levels in defined cell types (Martinez-Botas et al., 2000; Tansey et al., 2001;
Chang et al., 2006). We were, thus, interested to determine if there were a
preferential relationship among individual Plin proteins, their lipid targeting
preference, and the cellular accumulation of either TAG or CE. We first
attempted siRNA approaches to deplete Plins targeted to either TAG- or CE-
LSDs. Data from other systems clearly show that depletion of one Plin protein
type results in compensation by other Plins (Martinez-Botas et al., 2000; Tansey
et al., 2001; Chang et al., 2006; Sztalryd et al., 2006). Further, Plin2 and Plin3
are expressed in most cells and exhibit no TAG or CE targeting preference. Thus,
experiments directed toward defining Plin effects on TAG or CE levels required
us to simultaneously target multiple Plins in any individual cell. Several cultured
cells lines were selected but we were unsuccessful in depleting any of the TAG-
or CE-LSD specific Plins in combination with Plin2 and Plin3. As we were unable
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

13
to deplete all Plins associated with either TAG- or CE-LSDs, we sought an
alternative approach.
When we compared FACS analyses of GFP-Plin1a- and GFP-Plin1c-
expressing cells (Figure 4), we noticed a conspicuous difference in the relative
distributions of TAG- and CE-LSDs. >60% of total LSD particles in Plin1a-
expressing cells were tagged by the TAG-dye marker (Figure 4A), whereas the
Plin1c-expressing cells were predominantly (>60%) populated with CE-containing
LSDs (Figure 4C). These data suggested that the ectopic expression of Plins
with specific lipid targeting preferences might quantitatively alter the balance of
TAG/CE levels in individual cells. We thus analyzed the relative TAG/CE
distributions in cells expressing various Plin proteins.
AML12 cells were transiently transfected individually with vectors that
specifically encode different Plin proteins and cultured with oleic acid and
cholesterol. Cells cultured without exogenous lipids accumulated only limited
levels of TAG or CE regardless of Plin expression (data not shown), whereas
untransfected control cells accumulate large quantities of both TAG and CE.
LSDs were isolated from the various Plin-expressing and control cells and their
TAG/CE levels quantified by TLC. Cells expressing Plin1a or Plin1c were
analyzed in pairs, and normalized in parallel experiments of lipid loaded and
unloaded control cells (Figure 7). Similar paired and normalized experiments
were used to analyze cells expressing Plin4 or Plin5.
Plin1a and Plin1c exhibited largely reciprocal effects on TAG/CE levels,
with an ~60% TAG lipid bias observed in Plin1a-expressing cells and an ~60%
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

14
CE bias observed for Plin1c-expressing cells (Figure 7). Plin5/Plin4 differences in
relative TAG/CE accumulations were similarly polarized toward either TAG or CE,
respectively. Thus, ectopic expression of Plins that exhibit lipid targeting
preferences can polarize relative cellular lipid-type distributions. These effects
are seen for both the TAG-specific (i.e. Plin1a and Plin5) and the CE-specific (i.e.
Plin1c and Plin4) Plin proteins, but not for the non-preferential Plins 2, 3, and 1d
(data not shown).
DISCUSSION
We have demonstrated that distinct Plins differentially sequester to either
TAG- or CE-specific LSDs, emphasizing diversity of function for the different
Plins. These significant functional differences towards TAG- or CE-LSDs impact
previous assumptions about commonality of Plin action. The large unilocular
TAG-LSDs that are targeted by Plin1a in adipocytes are proposed to derive from
nascent droplets marked by Plin2, Plin3, and Plin4 (Wolins et al., 2005). While
Plin2 and Plin3 interact with TAG-LSDs, this association, unlike that of Plin1a, is
not exclusive, as Plin2 and Plin3 also co-segregate with CE-LSDs. Since Plin4
primarily targets CE-LSDs, conclusions regarding linear development of Plin-
specific LSD populations, based upon static global imaging of cells cultured with
exogneous fatty acids (Wolins et al., 2005), must be tempered.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

15
Plin structural domains that direct LSD targeting are still poorly defined. All
Plins share an N-terminal, ~100 amino acid PAT domain and a distal 11-mer
amphipathic helical repeat (Bussell and Eliezer, 2003; Lu et al., 2001; Miura et al.,
2002). C-terminal to both, Plins are increasingly diverged. Several groups have
probed for LSD-interacting motifs through domain-specific expressions (Garcia et
al., 2003; Hickenbottom et al., 2004; McManaman et al., 2003; Nakamura and
Fujimoto, 2003; Ohsaki et al., 2006; Subramanian et al., 2004a; Subramanian et
al., 2004b; Yamaguchi et al., 2006), but these studies have focused on TAG-
enriched cells and ignored effects of CE-LSDs. In addition, some constructs have
exposed amphipathic helices that are usually masked in endogenous Plins and,
thus, may target inappropriately (Hickenbottom et al., 2004).
No simple and consistent structural model for preferential Plin
associations with TG- or CE-LSDs can be easily deduced from sequence or
structural motif scanning and interrogation. While the PAT and associated 11-
mer domains may be involved in LSD interaction, these regions cannot be
sufficient determinants for specific targeting. Although all Plin1 variants have
identical N-terminal 198 residues, which include the PAT and 11-mer regions
(Kimmel et al., 2009; Lu et al., 2001; Miura et al., 2002), they have dissimilar LSD
targeting. Further, the PAT and 11-mer domains differ greatly among the other
Plin proteins in both length and sequence. Lipid binding discrimination can also
not be simply deduced by analyses of C-termini, which differ highly among Plin4,
Plin5, and each of the unique Plin1 forms. Signaling or targeting motifs may
instead reside in non-common segments, as hydrophobic segments are
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

16
suggested to facilitate Plin1 targeting to LSDs (Garcia et al., 2003). While Plin-
specific interacting proteins possibly help direct TAG- or CE-LSD recognition,
one must consider that the extremely diverged single Plin species in
Dictyostelium specifically targets LSDs when expressed in mammalian CHO cells
(Lu et al., 2001; Miura et al., 2002). Furthermore, although Plin interactions that
are unique to TAG- or CE-specific LSDs may involve the surrounding
phospholipid monolayer, these would also require commonality in both diverse
tissues (e.g. adrenal and liver cells) and species.
Pathogenesis associated with abnormal lipid storage has serious health
consequences. Thus, understanding the mechanisms that direct lipid storage and
lipolytic breakdown is paramount. The Plins regulate access of lipases to lipids
stored within the LSD core (Granneman et al., 2009; Granneman et al., 2010;
Martinez-Botas et al., 2000; Miyoshi et al., 2007; Sztalryd et al., 2003; Tansey et
al., 2001; Wang et al., 2010; Wang et al., 2009; Yamaguchi et al., 2004), and the
cellular content of Plins and accumulated LSDs seem intimately coordinated. In
addition, various Plins may have significantly different regulatory effects on
cellular lipolytic activity depending upon tissue context (Dalen et al., 2007;
Tansey et al., 2003). Accordingly, recent attention has been directed towards
possible causal linkages of aberrant Plin function with human disease. In
particular, heterozygous loss-of-function mutations of PLIN1 causes a familial
partial lipodystrophy in humans (Gandotra et al., 2011) and polymorphisms in
human genes for Plin1 (Qi et al., 2004) and Plin4 (Richardson et al., 2011) have
been associated with obesity.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

17
Our data add novel conceptual parameters for LSD and Perilipin function
and analyses. The various Plins target different classes of LSDs even within a
single cell population and Plins with LSD lipid specificities can preferentially
affect the accumulation of the targeted LSD class. Intriguingly, tissues such as
adipose, heart, and oxidative muscle, that predominantly accumulate TAG, have
highest expression of Plin 1a, 1b, or 5, which specifically target TAG-LSDs.
Conversely, steroidogenic cells, which accumulate CE, express these Plins more
poorly in comparison to Plin1c. We suggest that Plin expression may not impact
all cellular lipids and LSDs equivalently, but that the actions of individual Plin-
types may be differentially targeted to distinct LSD classes within a given cell. It
will, thus, be of interest to evaluate the full spectrum of Plin variants in broad
tissue panels that exhibit differential TAG/CE biases. Our new findings
underscore the view that each Plin is likely to have very separate and perhaps
unique functions associated with their LSD-specific targeting.
MATERIALS AND METHODS
Materials.
Fatty acid free bovine serum albumin (BSA) was purchased from Fisher
Scientific (Pittsburgh, PA). Phosphate buffered saline (PBS), glutamine, fetal
bovine serum (FBS), horse serum, cholesteryl BODIPY® 500/510 FL C12
[cholesteryl 4,4-difluoro-5,7-dimethyl-4-bora-3a,4a-diaza-s-indacene-3-
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

18
dodecanoate; cholesteryl dye], BODIPY® 558/568 C12 [4,4-difluoro-5-(2-thienyl)-
4-bora-3a,4a-diaza-s-indacene-3-dodecanoic acid; fatty acid dye], cholesteryl
BODIPY® 576/589 C11 [cholesteryl 4,4-difluoro-5-(2-pyrrolyl)-4-bora-3a,4a-diaza-
s-indacene-3-undecanoate; cholesteryl dye], and BODIPY® 500/510 FL C12 [4,4-
difluoro-5-methyl-4-bora-3a,4a-diaza-s-indacene-3-dodecanoate; fatty acid dye]
were from Invitrogen (Carlsbad, CA). MitoTracker (579/599) and LysoTracker
(577/590) were also from Invitrogen (Carlsbad, CA). Antibodies to LAMP1 and
EEa1 were from Abcam (Cambridge, MA) and to TOM2 from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA). The 250 microns silica gel H TLC plates
were from Analtech (Newark, DE). Complete protease inhibitor cocktail tablets
were from Roche Diagnostics (Indianapolis, IN). Paraformaldehyde was from
Electron Microscopy Sciences (Hatfield, PA). Cell media and all other chemicals
were from Sigma-Aldrich (St. Louis, MO).
Cell culture.
Y1 mouse adrenalcortical, AML12 mouse liver, and McARH7777 rat
hepatoma cells were obtained from the American Tissue Culture Collection
(Manassas, VA). Cells were grown to subconfluence in medium supplemented
with 100 μg/mL penicillin and 100 μg/mL streptomycin and incubated in
humidified air containing 5% CO2 at 37°C. Y1 adrenal cells were grown in
Dulbecco's modified Eagle medium (DMEM)/Ham’s nutrient mixture F-12
supplemented with 15% horse serum, 2.5% FBS, and 2 mM glutamine.
McARH7777 cells were grown in DMEM supplemented with 10% FBS. AML12
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

19
cells were grown in DMEM/Ham’s nutrient mixture F-12 supplemented with 5
mg/ml insulin, 5 mg/ml tranferrin, 5 ng/ml selenium, 10% FBS, and 2 mM
glutamine
Plin-GFP fusion constructs and transfection.
The perilipin expression vectors were generated using Multi-Site Gate-
Way (Invitrogen, Carlsbad, California). Mouse Plins 1a, 2, 3, and 5 cDNAs were
amplified from previously described pSG5 vectors (Dalen et al., 2007). Plins 1b,
1c, 1d, and 4 cDNAs were amplified from Y-1 cells or adipose tissue mRNA
using PfuTurbo® DNA Polymerase (Stratagene). Primers used contained
overhangs for insertion into the pDONR-221 P4r-P3r vector, Kozak sequence
(ACCTAG) and stop codon (CTA), and were designed using Vector NTI 10.0
(Invitrogen) with Tm set to 65ºC.
PCR products were recombined into the pDONR-221 P4r-P3r vector using
BP Clonase II (Invitrogen) to produce pENTR-vectors. GFP was amplified from
pEGFP-C1 (Clontech) and cloned into the pDONR-221 P1-P4. The V5-6x-His-
Gly epitope was cloned into the pDONR-221 P3-P2 vector.
The destination vector was generated by replacing the multi-cloning site of
pcDNA3 (Invitrogen) with the attR1-ccdB-chloramphenicol-attR2-cassette (R1-R2)
from pLenti6/v5-DEST (Invitrogen). The R1-R2 cassette was amplified with
PfuTurbo, and digested with HindIII/ApaI prior to ligation into HindIII/ApaI
digested pcDNA3 vector. The ligation mixture was transformed into ccdB Survival
TR cells (Invitrogen) and clones selected on Ampicilin (100 µg/ml) and
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

20
Chloramphenicol (25 µg/ml) plates. The novel pcDNA3-R1- R2 vector (pcDNA3-
KTD2-DEST) was subsequently recombined with the above pENTR-vectors
using LR clonase II (Invitrogen) to generate the pKTD2-G-Perilipin-VH
expression vectors. Due to the stop codon inserted into the pENTR-perilipin
vectors, the 3-end V5-6xHis-G tag will not be translated. Correctly amplified
sequences were confirmed by sequencing (Macrogen, Korea).
Transient transfection was carried out following the manufacturer's
instructions (Invitrogen) using lipofectamine LTX reagent. McARH7777 cells (cell
density 6.25x104/cm2) were incubated in standard growth conditions in medium
lacking antibiotic for one day and then incubated with fresh media containing
plasmid DNA (250 ng/cm2), lipofectamine LTX reagent (1 µl/cm2), and Opti-MEM
(50 µl/cm2) for one day.
Immunoblot analyses.
Proteins were separated by electrophoresis in 10% NuPAGE gels
(Invitrogen, Carlsbad, CA) using MOPS running buffer and then subjected to
immunoblot analyses as described (Kim et al., 2002). For immunoblot analyses,
we used rabbit polyclonal antisera (1:3000) to mouse Plin1 (Servetnick et al.,
1995), Plin2 (Xu et al., 2005), or Plin3 (Sztalryd et al., 2006), and guinea pig
polyclonal antisera (1:3000) to human Plin4 (American Research Product,
03-GP31) and human Plin5 (Progen Biotechnik, GmbH GP34). Rabbit antibody
(1:5000) to β-actin and GFP were, respectively, from Abcam (Cambridge, MA)
and Invitrogen (Carlsbad, CA). Secondary antibodies to rabbit or guinea pig
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

21
IgG were from Jackson ImmunoResearch (West Grove, PA) and used
at 1:5000 dilution.
Lipid loading.
Cells were grown overnight to <50% confluence in media supplemented
with 100-200 μM oleic acid bound to fatty acid free BSA (2.5:1 mol oleic acid: mol
BSA) (Dalen et al., 2006) and/or 50 μM cholesterol complexed with β-methyl
cyclodextrin (8:1 mol β-methyl cyclodextrin: mol cholesterol) (Christian et al.,
1997). For fatty acid and cholesteryl dye labeling, cells were grown overnight to
<50% confluence in media supplemented with 100 μM oleic acid and 50 μM
cholesterol, plus 1.0 μM BODIPY® 558/568 C12 (fatty acid dye) and 0.5 μM
cholesteryl BODIPY® 500/510 FL C12 (cholesteryl dye). GFP-Plin expressing
cells were grown overnight to <50% confluence in media supplemented with 100
μM oleic acid and 50 μM cholesterol, plus either 1.0 μM BODIPY® 558/568 C12
(fatty acid dye) or 0.5 μM cholesteryl BODIPY® 576/589 C11 (cholesteryl dye).
Lipid droplet preparation.
Cells were grown overnight, washed with phosphate-buffered saline (PBS),
scraped into PBS, and pelleted by centrifugation (300xg for 5 min). The cell pellet
was resuspended in 4 ml hypotonic lysis solution (50 mM HEPES pH 7.3, 0.1 M
KCl, 2 mM MgCl2; 4 °C) containing protease inhibitors (20 μg/ml leupeptin, 1 mM
benzamidine and 100 μM 4-(2-aminoethyl)-benzenesulfonylfluoride) and lysed by
incubation on ice for 30 min. 1 ml of 50% (w/v) sucrose in lysis solution was
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

22
added to the cell lysate for a total volume of 5 ml. This 10% w/v sucrose solution
was layered at the bottom of an ultracentrifuge tube and a step gradient of 5%,
2.5%, and 0% sucrose was carefully layered above; centrifugation (Beckman
Coulter Optima XL100K Ultracentrifuge) was at 154,000xg for 1 h at 4 °C. The
floating lipid layer was collected using a Beckman tube slicer (Brea, CA) for
immunoblotting or flow cytometry (Brasaemle et al., 2004; Hsieh and Huang,
2005)
FACS separation of fluorescently labeled lipid droplets.
Lipid droplets isolated from cells labeled with fluorescent fatty acid dye,
fluorescent cholesteryl dye, and/or expressing GFP were sorted using a
FACSAria II cytometer (BD Biosciences). All fluorochromes were excited using a
488 nm laser. Fluorescent emissions from lipid droplets labeled with cholesteryl
BODIPY® 500/510 FL C12 or with GFP-tagged perilipin proteins were collected
through a 502 nm longpass filter followed by a 530/30 nm bandpass filter.
Emissions from lipid droplets labeled with BODIPY® 558/568 C12 or with
cholesteryl BODIPY® 576/589 C11 were collected through a 556 nm longpass
filter followed by a 575/26 bandpass filter. Compensation settings for
experiments with multiple fluorescent dyes were established using lipid droplets
isolated from cells labeled with a single fluorescent dye (see Figures S1 and S3).
The forward scatter threshold was set at the lowest possible linear signal height
of 200 (within an allowed range of 200 to 262,143), permitting detection of lipid
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

23
droplets with diameters >1 μm. For sorting, ~5,000 events/second were
processed and sort precision was set on the default purity mode.
Confocal laser scanning microscopy (CLSM).
CLSM was carried out using a Zeiss LSM 510 (Jena, Germany) inverted
confocal microscope with a 100x (Plan-Apochromat, NA1.40) oil objective lens.
GFP and cholesteryl BODIPY® FL 500/510 FL C12 were imaged using argon
488-nm laser and a 505-530-nm BP emission filter. BODIPY® 558/568 C12 and
cholesteryl BODIPY® 576/589 C11 were imaged using a He/Ne 543-nm laser
excitation and a 580-nm LP emission filter. The software for confocal microscopic
image generation was LSM510 software 3.2. The cells were seeded and
maniplulated in 35 mm glass bottom culture dishes (MatTek, Ashland, MA), fixed
with 4% paraformaldehyde in PBS for 30 minutes, and washed twice for 5
minutes before observation. To visualize isolated lipid droplets, the unsorted or
sorted lipid droplet populations were first mixed with an equal volume of glycerol
before observation.
Lipid extraction and analyses.
Lipid droplets were extracted twice with 2 volumes of
chloroform:heptane:methanol (4:3:2; v/v/v) (Hsieh and Huang, 2007). The lipids
were applied to TLC plates and separated in hexane:diethyl ether:acetic acid
(70:30:1; v/v/v); the plates were stained overnight in an iodine chamber to
visualize the lipids. Extracted lipids were separated in parallel to a dilution series
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

24
of lipid standards applied to the same plate. The standards were cholesteryl
oleate (cholesteryl ester; CE), glyceryl trioleate (triacylglycerol; TAG), oleic acid
(fatty acid; FA), cholesterol (Chol), dioleoylglycerol (diacyclglycerol; DAG), DL-α-
Monoolein (monoacyclglycerol; MAG), and L-α-phosphatidylcholine (phospholipid;
PL).
TIFF images of the iodine stained TLC plates were analyzed by
ImageQuant TL with module 1D gel analyses (GE Health Life Sciences,
Piscataway, NJ). Stored 8 bit grayscale images were created manually, since
single lane images had various shapes and spot distances. Spot intensities were
corrected for background and data exported to generate curve standards for the
various lipid marker controls. Masses of the unknown lipids were extrapolated in
comparison to standard curves (see Figure S3A).
Statistical analyses.
Confocal data (Figures 4,5,6) are from at least 3 independent, paired
experiments (see Figure 3), where the TAG/CE sum for each pair was 98-102%
(see Figure 3). McARH7777 cells were transiently transfected with the indicated
GFP-Plin expressing constructs and cultured overnight in the presence of 100
μM oleic acid and 50 μM cholesterol, plus either 1 μM BODIPY 558/568 C12 (FA)
or 0.5 μM cholesteryl BODIPY 576/589 C11 (Chl) dye markers. TAG or CE
distributions represent the % of the total GFP+ signal that co-sorts, respectively;
with either FA+ or Chl+ tags. The numbers are shown ± standard deviation
(Figure 6).
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

25
ACKNOWLEDGEMENTS
This research was supported by the Intramural Research Program of the
National Institutes of Health, the National Institute of Diabetes and Digestive and
Kidney Diseases. K.T.D. was supported by a travel grant from the Henning and
Johan Throne-Holst's Foundation. There are no conflicts or competing interests.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

26
REFERENCES
Brasaemle, D. L., Dolios, G., Shapiro, L. and Wang, R. (2004). Proteomic analysis of proteins associated with lipid droplets of basal and lipolytically stimulated 3T3-L1 adipocytes. J Biol Chem 279, 46835-42. Bussell, R., Jr. and Eliezer, D. (2003). A structural and functional role for 11-mer repeats in alpha-synuclein and other exchangeable lipid binding proteins. J Mol Biol 329, 763-78. Chang, B. H., Li, L., Paul, A., Taniguchi, S., Nannegari, V., Heird, W. C. and Chan, L. (2006). Protection against fatty liver but normal adipogenesis in mice lacking adipose differentiation-related protein. Mol Cell Biol 26, 1063-76. Christian, A. E., Haynes, M. P., Phillips, M. C. and Rothblat, G. H. (1997). Use of cyclodextrins for manipulating cellular cholesterol content. J Lipid Res 38, 2264-72. Dalen, K. T., Dahl, T., Holter, E., Arntsen, B., Londos, C., Sztalryd, C. and Nebb, H. I. (2007). LSDP5 is a PAT protein specifically expressed in fatty acid oxidizing tissues. Biochim Biophys Acta 1771, 210-27. Dalen, K. T., Ulven, S. M., Arntsen, B. M., Solaas, K. and Nebb, H. I. (2006). PPARalpha activators and fasting induce the expression of adipose differentiation-related protein in liver. J Lipid Res 47, 931-43. Farese, R. V., Jr. and Walther, T. C. (2009). Lipid droplets finally get a little R-E-S-P-E-C-T. Cell 139, 855-60. Gandotra, S., Le Dour, C., Bottomley, W., Cervera, P., Giral, P., Reznik, Y., Charpentier, G., Auclair, M., Delepine, M., Barroso, I. et al. (2011). Perilipin deficiency and autosomal dominant partial lipodystrophy. N Engl J Med 364, 740-8. Garcia, A., Sekowski, A., Subramanian, V. and Brasaemle, D. L. (2003). The central domain is required to target and anchor perilipin A to lipid droplets. J Biol Chem 278, 625-35. Granneman, J. G., Moore, H. P., Krishnamoorthy, R. and Rathod, M. (2009). Perilipin controls lipolysis by regulating the interactions of AB-hydrolase containing 5 (Abhd5) and adipose triglyceride lipase (Atgl). J Biol Chem 284, 34538-44. Granneman, J. G., Moore, H. P., Mottillo, E. P., Zhu, Z. and Zhou, L. (2010). Interactions of perilipin-5 (Plin5) with adipose triglyceride lipase. J Biol Chem 286, 5126-35. Greenberg, A. S., Egan, J. J., Wek, S. A., Moos, M. C., Jr., Londos, C. and Kimmel, A. R. (1993). Isolation of cDNAs for perilipins A and B: sequence and expression of lipid droplet-associated proteins of adipocytes. Proc Natl Acad Sci U S A 90, 12035-9. Hickenbottom, S. J., Kimmel, A. R., Londos, C. and Hurley, J. H. (2004). Structure of a lipid droplet protein; the PAT family member TIP47. Structure 12, 1199-207. Hsieh, K. and Huang, A. H. (2005). Lipid-rich tapetosomes in Brassica tapetum are composed of oleosin-coated oil droplets and vesicles, both
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

27
assembled in and then detached from the endoplasmic reticulum. Plant J 43, 889-99. Hsieh, K. and Huang, A. H. (2007). Tapetosomes in Brassica tapetum accumulate endoplasmic reticulum-derived flavonoids and alkanes for delivery to the pollen surface. Plant Cell 19, 582-96. Kim, H. U., Hsieh, K., Ratnayake, C. and Huang, A. H. (2002). A novel group of oleosins is present inside the pollen of Arabidopsis. J Biol Chem 277, 22677-84. Kimmel, A. R., Brasaemle, D. L., McAndrews-Hill, M., Sztalryd, C. and Londos, C. (2009). Adoption of PERILIPIN as a unifying nomenclature for the mammalian PAT-family of intracellular lipid storage droplet proteins. J Lipid Res 51, 468-71. Kimmel, A. R., Brasaemle, D. L., McAndrews-Hill, M., Sztalryd, C. and Londos, C. (2010). Adoption of PERILIPIN as a unifying nomenclature for the mammalian PAT-family of intracellular lipid storage droplet proteins. J Lipid Res 51, 468-71. Londos, C., Sztalryd, C., Tansey, J. T. and Kimmel, A. R. (2005). Role of PAT proteins in lipid metabolism. Biochimie 87, 45-9. Lu, X., Gruia-Gray, J., Copeland, N. G., Gilbert, D. J., Jenkins, N. A., Londos, C. and Kimmel, A. R. (2001). The murine perilipin gene: the lipid droplet-associated perilipins derive from tissue-specific, mRNA splice variants and define a gene family of ancient origin. Mamm Genome 12, 741-9. Martinez-Botas, J., Anderson, J. B., Tessier, D., Lapillonne, A., Chang, B. H., Quast, M. J., Gorenstein, D., Chen, K. H. and Chan, L. (2000). Absence of perilipin results in leanness and reverses obesity in Lepr(db/db) mice. Nat Genet 26, 474-9. McManaman, J. L., Zabaronick, W., Schaack, J. and Orlicky, D. J. (2003). Lipid droplet targeting domains of adipophilin. J Lipid Res 44, 668-73. Miura, S., Gan, J. W., Brzostowski, J., Parisi, M. J., Schultz, C. J., Londos, C., Oliver, B. and Kimmel, A. R. (2002). Functional conservation for lipid storage droplet association among Perilipin, ADRP, and TIP47 (PAT)-related proteins in mammals, Drosophila, and Dictyostelium. J Biol Chem 277, 32253-7. Miyoshi, H., Perfield, J. W., 2nd, Souza, S. C., Shen, W. J., Zhang, H. H., Stancheva, Z. S., Kraemer, F. B., Obin, M. S. and Greenberg, A. S. (2007). Control of adipose triglyceride lipase action by serine 517 of perilipin A globally regulates protein kinase A-stimulated lipolysis in adipocytes. J Biol Chem 282, 996-1002. Nakamura, N. and Fujimoto, T. (2003). Adipose differentiation-related protein has two independent domains for targeting to lipid droplets. Biochem Biophys Res Commun 306, 333-8. Ohsaki, Y., Maeda, T., Maeda, M., Tauchi-Sato, K. and Fujimoto, T. (2006). Recruitment of TIP47 to lipid droplets is controlled by the putative hydrophobic cleft. Biochem Biophys Res Commun 347, 279-87.
Qi, L., Shen, H., Larson, I., Schaefer, E.J., Greenberg, A.S., Tregouet, D.A., Corella, D. and Ordovas, J.M. (2004). Gender-specific association of a
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

28
perilipin gene haplotype with obesity risk in a white population. Obes Res 12, 1758-1765.\
Richardson. K., Louie-Gao, Q., Arnett, D.K., Parnell, L.D., Lai, C.Q., Davalos, A., Fox, C.S., Demissie, S., Cupples, L.A., Fernandez-Hernando, C. and Ordovas, J.M. (2011). The PLIN4 variant rs8887 modulates obesity related phenotypes in humans through creation of a novel miR-522 seed site. PLoS One 6, e17944. Servetnick, D. A., Brasaemle, D. L., Gruia-Gray, J., Kimmel, A. R., Wolff, J. and Londos, C. (1995). Perilipins are associated with cholesteryl ester droplets in steroidogenic adrenal cortical and Leydig cells. J Biol Chem 270, 16970-3. Subramanian, V., Garcia, A., Sekowski, A. and Brasaemle, D. L. (2004a). Hydrophobic sequences target and anchor perilipin A to lipid droplets. J Lipid Res 45, 1983-91. Subramanian, V., Rothenberg, A., Gomez, C., Cohen, A. W., Garcia, A., Bhattacharyya, S., Shapiro, L., Dolios, G., Wang, R., Lisanti, M. P. et al. (2004b). Perilipin A mediates the reversible binding of CGI-58 to lipid droplets in 3T3-L1 adipocytes. J Biol Chem 279, 42062-71. Sztalryd, C., Bell, M., Lu, X., Mertz, P., Hickenbottom, S., Chang, B. H., Chan, L., Kimmel, A. R. and Londos, C. (2006). Functional compensation for adipose differentiation-related protein (ADFP) by Tip47 in an ADFP null embryonic cell line. J Biol Chem 281, 34341-8. Sztalryd, C., Xu, G., Dorward, H., Tansey, J. T., Contreras, J. A., Kimmel, A. R. and Londos, C. (2003). Perilipin A is essential for the translocation of hormone-sensitive lipase during lipolytic activation. J Cell Biol 161, 1093-103. Tansey, J. T., Huml, A. M., Vogt, R., Davis, K. E., Jones, J. M., Fraser, K. A., Brasaemle, D. L., Kimmel, A. R. and Londos, C. (2003). Functional studies on native and mutated forms of perilipins. A role in protein kinase A-mediated lipolysis of triacylglycerols. J Biol Chem 278, 8401-6. Tansey, J. T., Sztalryd, C., Gruia-Gray, J., Roush, D. L., Zee, J. V., Gavrilova, O., Reitman, M. L., Deng, C. X., Li, C., Kimmel, A. R. et al. (2001). Perilipin ablation results in a lean mouse with aberrant adipocyte lipolysis, enhanced leptin production, and resistance to diet-induced obesity. Proc Natl Acad Sci U S A 98, 6494-9. Wang, H., Bell, M., Sreenevasan, U., Hu, H., Liu, J., Dalen, K., Londos, C., Yamaguchi, T., Rizzo, M. A., Coleman, R. et al. (2010). Unique Regulation of Adipose Triglyceride Lipase (ATGL) by Perilipin 5, a Lipid Droplet-associated Protein. J Biol Chem 286, 15707-15. Wang, H., Hu, L., Dalen, K., Dorward, H., Marcinkiewicz, A., Russell, D., Gong, D., Londos, C., Yamaguchi, T., Holm, C. et al. (2009). Activation of hormone-sensitive lipase requires two steps, protein phosphorylation and binding to the PAT-1 domain of lipid droplet coat proteins. J Biol Chem 284, 32116-25. Wolins, N. E., Quaynor, B. K., Skinner, J. R., Schoenfish, M. J., Tzekov, A. and Bickel, P. E. (2005). S3-12, Adipophilin, and TIP47 package lipid in adipocytes. J Biol Chem 280, 19146-55.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

29
Wolins, N. E., Quaynor, B. K., Skinner, J. R., Tzekov, A., Croce, M. A., Gropler, M. C., Varma, V., Yao-Borengasser, A., Rasouli, N., Kern, P. A. et al. (2006). OXPAT/PAT-1 is a PPAR-induced lipid droplet protein that promotes fatty acid utilization. Diabetes 55, 3418-28. Xu, G., Sztalryd, C., Lu, X., Tansey, J. T., Gan, J., Dorward, H., Kimmel, A. R. and Londos, C. (2005). Post-translational regulation of adipose differentiation-related protein by the ubiquitin/proteasome pathway. J Biol Chem 280, 42841-7. Yamaguchi, T., Matsushita, S., Motojima, K., Hirose, F. and Osumi, T. (2006). MLDP, a novel PAT family protein localized to lipid droplets and enriched in the heart, is regulated by peroxisome proliferator-activated receptor alpha. J Biol Chem 281, 14232-40. Yamaguchi, T., Omatsu, N., Matsushita, S. and Osumi, T. (2004). CGI-58 interacts with perilipin and is localized to lipid droplets. Possible involvement of CGI-58 mislocalization in Chanarin-Dorfman syndrome. J Biol Chem 279, 30490-7.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

30
FIGURE LEGENDS
Figure 1. Differential accumulation of Plins in cells cultured in the absence
or presence of fatty acid and/or cholesterol.
A. Y1 adrenal cells were cultured overnight in the absence or presence of oleic
acid and/or cholesterol. Whole cell lysates were prepared and endogenous Plins
assayed by immunoblot. Data are representative of 3 experiments.
B. McARH7777 rat liver cells were transiently transfected with the indicated GFP-
Plin expressing constructs and cultured overnight in the absence or presence of
oleic acid and/or cholesterol. Whole cell lysates were prepared and GFP-Plin
assayed by immunoblot. Data are representative of 3 experiments.
Figure 2. Spatially distinct intracellular accumulations of fatty acid- and
cholesteryl-dye markers.
A. Y1 adrenal, McARH7777 rat liver, and AML12 mouse liver cells were cultured
overnight in the presence of oleic acid and cholesterol, plus BODIPY 558/568 C12
[fatty acids (FA)] and cholesteryl BODIPY 500/510 FL C12 (Chl). Representative
confocal images are shown with red and green indicating the localization of FA
and Chl, respectively.
B. McARH7777 cells were cultured overnight in the presence of oleic acid and
cholesterol, plus BODIPY 558/568 C12 [fatty acids (FA)] and cholesteryl BODIPY
500/510 FL C12 (Chl) and probed for organelle localization by immunoflorescence
detection of LAMP1 (Lysosomal-associated membrane protein 1), TOM2
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

31
(translocase of outer mitochondrial membranes 20 kDa), and EEA1 (Early
Endosome Antigen 1.
C. McARH7777 cells were cultured overnight in the presence of oleic acid and
cholesterol, plus BODIPY 500/510 C12 [fatty acids (FA)] or cholesteryl BODIPY
500/510 FL C12 (Chl) and probed for organelle localization by staining with dye
specific markers.
Figure 3. Differential Localization of Plins to TAG or CE droplets.
Y1 adrenal, McARH7777 rat liver, and AML12 mouse liver cells were
cultured overnight in the presence of oleic acid and cholesterol, plus BODIPY
558/568 C12 [fatty acids (FA)] and cholesteryl BODIPY 500/510 FL C12 (Chl).
A. Representative FACS profiles of purified FA- and Chl-tagged lipid droplets are
shown, with relative distributions of total lipid droplet numbers indicated in the
different quadrants (FA-/Chl-, FA+/Chl-, FA-/Chl+, and FA+/Chl+); blue represents
dual signals of FA and Chl probes. Y1 cells (25,000 total droplets); McARH7777
cells (20,000 total droplets); AML12 cells (25,000 total droplets).
B. Confocal images of unsorted and FACS sorted FA- and Chl-tagged lipid
droplets from Y1, McARH7777, and AML12 cells.
C. Lipids isolated from FA- and Chl-sorted droplets were separated by TLC in
parallel with lipid markers and detected by staining with iodine vapor. 1x indicates
2x105 LSD particles; 5x indicates 1x106 LSD particles. Marker lanes have 50μg
of CE: Cholesteryl Ester* and 10μg each of TAG: Triacylglycerols; FA: Fatty
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

32
Acids; DAG: Diacylglycerides; Chol: Cholesterol; MAG: Monoacylglyceride; PL:
Phospholipids. PLs do not migrate from the origin in this system.
D. Proteins were prepared from unsorted and FACS sorted TAG- or CE-LSD
isolated from Y1 cells. Endogenous Plin proteins were assayed by immunoblot.
Identical lipid droplet particle numbers were loaded in each lane. Data are
representative of 3 experiments.
Figure 4. Differential localization of Plin1 proteins to TAG or CE lipid
droplets.
McARH7777 cells were transiently transfected with the indicated GFP-
Plin1 expressing constructs and cultured overnight in the presence of oleic acid
and cholesterol, plus either BODIPY 558/568 C12 (TAG) or cholesteryl BODIPY
576/589 C11 (CE).
Representative confocal images are shown for each with red indicating the
localization of either TAG or CE droplets, green indicating localization of GFP-
Plin1 proteins, and yellow (or rings) indicating co-localization.
Representative FACS profiles of TAG- and GFP-labeled lipid droplets or of
CE- and GFP-labeled lipid droplets are shown, with relative distributions of total
particle numbers indicated in the different quadrants, and the relative Plin1
variant localizations indicated as % of total GFP+ signal that co-sorts (blue) with
either TAG+ or CE+ tags. Each experiment was performed at least 3 times (see
Figure 6).
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

33
A. Plin1a (25,000 total droplets for each population); B. Plin1b (20,000
total droplets for each population); C. Plin1c (25,000 total droplets for each
population); D. Plin1d (10,000 total droplets for each population).
Figure 5. Differential localization of Plin2-5 proteins to TAG or CE lipid
droplets.
McARH7777 cells were transiently transfected with the indicated GFP-Plin
expressing constructs and cultured overnight in the presence of oleic acid and
cholesterol, plus either BODIPY 558/568 C12 (TAG) or cholesteryl BODIPY
576/589 C11 (CE).
Representative confocal images are shown for each with red indicating the
localization of either TAG or CE droplets, green indicating localization of GFP-
Plin proteins, and yellow (or rings) indicating co-localization.
Representative FACS profiles of TAG- and GFP-labeled lipid droplets or of
CE- and GFP-labeled lipid droplets are shown, with relative distributions of total
particle numbers indicated in the different quadrants, and the relative Plin2-5
variant localizations indicated as % of total GFP+ signal that co-sorts (blue) with
either TAG+ or CE+ tags. Each experiment was performed at least 3 times (see
Figure 6).
A. Plin2 (15,000 total droplets for each population); B. Plin3 (20,000 total
droplets for each population); C. Plin4 (25,000 total droplets for each population);
D. Plin5 (10,000 total droplets for each population).
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

34
Figure 6. Relative distributions of Plin family proteins to TAG- or CE-
specific intracellular lipid storage droplets.
McARH7777 cells were transiently transfected with the indicated GFP-Plin
expressing constructs and cultured overnight in the presence of oleic acid and
cholesterol, plus either BODIPY 558/568 C12 (FA) or cholesteryl BODIPY
576/589 C11 (Chl) dye markers. TAG or CE distributions represent the % of the
total GFP+ signal that co-sorts, respectively; with either FA+ or Chl+ tags (see
Figures 4,5). Data are from at least 3 independent, paired experiments, where
the TAG/CE sum for each pair was 98-102%. The numbers are shown as the
mean ± standard deviation.
Plin1a, Plin1b, and Plin5 show strong preference for localization to TAG-
LSDs, with Plin1a consistently exhibiting a stronger TAG signal than did Plin1b or
Plin5. Plin1c and Plin4 show strong preference for localization to CE-LSDs, with
Plin1c consistently exhibiting a stronger CE signal than did Plin4. Plin1d, Plin2,
and Plin3 exhibit minor localization preferences.
Figure 7. Relative change in TAG- or CE- specific intracellular lipid storage
upon expression of various Plin proteins.
AML12 cells were transiently transfected with the indicated GFP-Plin
expressing constructs and cultured overnight in the presence of oleic acid and
cholesterol. Transfection efficiencies were confirmed by visualizing GFP
fluorescence. Untransfected cells, cultured with/without exogenous oleic acid and
cholesterol, were grown in parallel. LSDs were isolated by centrifugation and
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

35
lipids were extracted, separated by TLC in parallel with lipid markers, and TAG
and CE detected after staining with iodine vapor. Relative TAG/CE ratios were
quantified in cells transfected with each specific Plin-expressing construct and
analyzed in parallel with identically grown untransfected cells for normalization
and TLC background correction.
The TAG/CE-ratio for untransfected lipid loaded cells (controls) was set to
0.5. The relative TAG/CE-ratio of Plin1a- and Plin1c- expressing cells were
always analyzed in parallel and normalized to those of control cells, and then
secondarily compared to results determined for its Plin-expressing counterpart.
Numbers >0.5 indicate a proportional increase in TAG lipid bias, whereas
numbers <0.5 indicate a proportional increase in CE lipid bias. Relative
distributions of TAG/CE levels are shown as the mean ± standard deviation for
each paired comparison. Data for each pair are based on three independent
experiments. Plin4 and Plin5 expressing cells were similarly analyzed as pairs
and internally compared as described for the Plin1a/Plin1c pair.
Plin1a and Plin5 show a relative bias for cellular TAG accumulation and a
strong preference for binding specificity to TAG-LSDs (see Figure 6). Conversely,
Plin1c and Plin4 show a relative increase in cellular CE levels and a strong
preference for binding specificity to CE-LSDs.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

36
SUPPLEMENTAL FIGURE LEGENDS
Figure S1. Spatially distinct intracellular accumulations of fatty acid- and
cholesteryl-dye markers.
A. 4T1 mouse mammary tumor, primary mouse liver, C2C12 mouse myoblast,
3T3-L1 mouse fibroblast, J774A.1 mouse monocyte-macrophage, and CHOK1
Chinese hampster ovary cells were cultured overnight in the presence of oleic
acid and cholesterol, plus BODIPY 558/568 C12 [fatty acids (FA)] and cholesteryl
BODIPY 500/510 FL C12 (Chl). Representative confocal images are shown with
red and green indicating the localization of FA and Chl, respectively.
B. McARH7777 cells were cultured for the times indicated in the presence of
oleic acid and cholesterol, plus BODIPY 558/568 C12 [fatty acids (FA)] and
cholesteryl BODIPY 500/510 FL C12 (Chl). Representative confocal images are
shown with red and green indicating the localization of FA and Chl, respectively.
Figure S2. Control FACS profiles of lipid dye-marked Y1 cells.
Cells were cultured overnight in the presence of 100 μM oleic acid and 50
μM cholesterol, and in the absence or presence of 1 μM BODIPY 558/568 C12
(FA) or 0.5 μM cholesteryl BODIPY 500/510 FL C12 (Chl). Cells were
hypotonically lysed and lipid droplets purified by sucrose gradient centrifugation.
Representative FACS profiles of unlabeled lipid droplets and of FA- or Chl-
labeled lipid droplets are shown, with relative distributions of total particle
numbers indicated in the different quadrants (FA-/Chl-, FA+/Chl-, FA-/Chl+, and
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

37
FA+/Chl+). In one FACS panel (FA lysate + Chl lysate), lysates were prepared
from cells that had been separately labeled for either FA or Chl and then mixed
and sorted simultaneously. A. Y1 cells; B. McARH7777 cells; C. AML12 cells.
Figure S3. Lipid quantification.
A. Lipid markers were separated by TLC and detected by staining with iodine
vapor (see Figure 3C). TIFF images of the iodine stained TLC plates were
analyzed to generate standard curves for each lipid. CE: Cholesteryl Ester; TAG:
Triacylglycerols; FA: Fatty Acids; DAG: Diacylglycerides; Chol: Cholesterol; MAG:
Monoacylglyceride.
B. Lipids isolated from FA- and Chl-sorted droplets were separated by TLC in
parallel with lipid markers and detected by fluorescence under UV or by staining
with iodine vapor. 1x indicates 2x105 LSD particles; 5x indicates 1x106 LSD
particles. Marker lanes had various lipid concentrations. Bodipy-FA is the input
BODIPY 558/568 C12 marker and Bodipy-Chl is the input cholesteryl BODIPY
500/510 marker. CE: Cholesteryl Ester; TAG: Triacylglycerols; FA: Fatty Acids;
DAG: Diacylglycerides; Chol: Cholesterol; MAG: Monoacylglyceride; PL:
Phospholipids. PLs do not migrate from the origin in this system.
Figure S4. Control FACS profiles of lipid dye-marked or GFP-Plin
expressing McARH7777 cells cells.
McARH7777 cells were cultured overnight in the presence of 100 μM oleic
acid and 50 μM cholesterol. In one cell population, 1 μM BODIPY 558/568 C12
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

38
(FA) was added to the culture media. Another population of McARH7777 cells
was transiently transfected with a GFP-Plin1a expressing construct before
culturing. Cells were then hypotonically lysed and lipid droplets purified by
flotation following centrifugation. Representative FACS profiles of unlabeled lipid
droplets and of FA- or GFP-labeled lipid droplets are shown, with relative
distributions of total particle numbers indicated in the different quadrants (FA-
/GFP-, FA+/GFP-, FA-/GFP+, and FA+/GFP+). In one FACS panel (FA lysate +
GFP lysate), lysates were prepared from cells that had been separately labeled
for either FA or GFP and then mixed and sorted simultaneously.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Table I
Lipid Specificities in FACS Purified Lipid Droplets
LIPID Chl-Tagged Droplets FA-Tagged Droplets s
...... Y1 McA AML12 Y1 McA AML12
CE 90μg 65μg 70μg ND (3<μg) ND (3<μg) ND (3<μg)
TAG ND (0.5<μg) ND (0.5<μg) ND (0.5<μg) 70μg 100μg 100μg
FA ND (0.5<μg) ND (0.5<μg) ND (0.5<μg 10μg 15μg 20μg
DAG ND (0.5<μg) ND (0.5<μg) ND (0.5<μg 20μg 10μg 10μg
Chol 50μg 15μg 15μg 3-5μg 3-5μg 3-5μg
MAG ND (2<μg) ND (2<μg) ND (2<μg) 15μg 10μg 10μg
Chl- and FA-tagged lipid droplets from Y1, McARH7777, and AML12 cells were separated by FACS
and lipid profiles analyzed by TLC (see Figure 3C). Values/2x106 droplets were extrapolated (± 30%)
from relative staining intensity data in comparison to standards (see Figure S3B) for CE: Cholesteryl
Ester, TAG: Triacylglycerol, FA: Fatty Acid, DAG: Diacylglycerol, Chol: Cholesterol, and MAG:
Monoacylglycerol. Phospholipid (PL) spot staining was similar for each, but did not resolve from the
origin. ND: not detected.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

- - + + Oleic Acid
- + - + Cholesterol
Plin1a
Plin1b
Plin1c
Plin1d
Plin2
Plin3
Plin4
Plin5
actin
Endogenous
Y1 Plins
Figure 1A
A.
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

- - + + Oleic Acid
- + - + Cholesterol
Plin1a
actin
Plin1b
actin
Plin1c
actin
Plin1d
actin
Plin2
actin
Plin3
actin
Plin4
actin
Plin5
actin
GFP-Plin B.
Figure 1B
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Y1 Mouse Adrenalcortical
FA Chl
A.
Figure 2A
McARH7777 Rat Liver
FA Chl
AML12 Mouse Liver
FA Chl
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

LAMP1 (lysosomes)
FA
Chl
EEA1 (early endsomes) Tom20 (mitochondria)
B.
Figure 2B
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

MitoTracker LysoTracker
C.
Figure 2C
FA
Chl
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Fatt
y A
cid
s
Cholesteryl
Figure 3A
A.
Y1
Cholesteryl Cholesteryl
McARH7777 AML12
0.381
0.194
0.400
0.025
0.356
0.042 0.330
0.272 0.515
0.294
0.149
0.042
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

McARH7777
Unsorted
Droplets
FA-
Sorted
Droplets
Chl-
Sorted
Droplets
Y1 AML12
10mm
B.
Figure 3B
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

AML12 Sorted
Lipid Droplets
DAG Chol
MAG
McA Sorted
Lipid Droplets Lipid Markers
(10 mg each)
4x105 Chl-Droplets
4x105 FA-Droplets
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
5x 5x
CE
TAG
FA
PL
DAG Chol
MAG
Y1 Sorted
Lipid Droplets
CE
FA
PL
Chol
MAG
TAG
DAG
+*
-
-
-
-
-
-
-
-
5x 1x 1x 1x
1x 1x 1x
* (50mg) CE
TAG
FA
(origin) PL
C.
Figure 3C
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

+ - + TAG Lipid Droplets
+ + - CE Lipid Droplets
Plin1a
Plin1b
Plin1c
Plin1d
Plin2
Plin3
Plin4
Plin5
Endogenous
Y1 Plins
FACS
Purified
D.
Figure 3D
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

TA
G
GFP-Plin1a GFP-Plin1a
CE
95% of
GFP+
5% of
GFP+
0.032
0.574
0.305
0.0
89
0.505
0.027
0.080
0.3
87
Plin1a
TAG
Plin1a
CE
A.
Figure 4A
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

GFP-Plin1b GFP-Plin1b
80% of
GFP+
0.092
0.358
0.361
0.1
89
19% of
GFP+
0.328
0.079
0.133
0.4
60
Plin1b
TAG Plin1b
CE
TA
G
CE
B.
Figure 4B
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

GFP-Plin1c GFP-Plin1c
8% of
GFP+
0.562
0.048
0.063
0.3
28
94% of
GFP+
0.041
0.639
0.274
0.0
46
Plin1c
TAG
Plin1c
CE
TA
G
CE
C.
Figure 4C
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

GFP-Plin1d GFP-Plin1d
47% of
GFP+
0.207
0.191
0.299
0.3
03
57% of
GFP+
0.173
0.225 0
.27
3
Plin1d
TAG
Plin1d
CE
0.329
TA
G
CE
D.
Figure 4D
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

GFP-Plin2 GFP-Plin2
65% of
GFP+
0.112
0.210
0.480
0.1
98
32% of
GFP+
0.225
0.108 0
.36
5
Plin2
TAG
Plin2
CE
0.302
TA
G
CE
A.
Figure 5A
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

GFP-Plin3 GFP-Plin3
55% of
GFP+
0.141
0.171
0.383
0.3
05
43% of
GFP+
0.154
0.118 0
.31
8
Plin3
TAG Plin3
CE
0.410
TA
G
CE
B.
Figure 5B
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

GFP-Plin4 GFP-Plin4
19% of
GFP+
0.301
0.072
0.4
39
84% of
GFP+
0.072
0.1
41
0.384
Plin4
TAG
Plin4
CE
0.403 0.189
TA
G
CE
C.
Figure 5C
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

GFP-Plin5 GFP-Plin5
88% of
GFP+
0.051
0.356
0.441
0.1
52
13% of
GFP+
0.339
0.050
0.205
0.4
06
Plin5
TAG Plin5
CE
TA
G
CE
D.
Figure 5D
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

Plin1a
Plin1b
Plin1c
Plin1d
Plin2
Plin3
Plin4
Plin5
TAG CE
Plin1a 93.0 2.4% 6.0 1.2%
Plin1b 82.6 6.7% 16.1 6.2%
Plin1c 9.5 2.3% 91.4 3.7%
Plin1d 46.2 3.9% 57.2 3.1%
Plin2 67.8 3.6% 34.5 3.0%
Plin3 57.2 3.3% 46.0 3.6%
Plin4 17.7 2.0% 81.9 2.4%
Plin5 87.9 1.5% 12.5 0.4%
GFP-Plin % TAG Distribution % CE Distribution
Figure 6
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t

No
rma
lize
d T
AG
/CE
0
0.5
1.0
1a 1c 5 4
Lipid Quantity
Figure 7
Plin
p<0.05 p<0.05
Jour
nal o
f Cel
l Sci
ence
Acc
epte
d m
anus
crip
t